jasmonic acid as modulator of lead toxicity in aquatic plant wolffia arrhiza (lemnaceae)
TRANSCRIPT

J(
AMa
b
a
ARRA
KAAGJLW
1
1laoaerfgtflm
om
Mr
0d
Environmental and Experimental Botany 66 (2009) 507–513
Contents lists available at ScienceDirect
Environmental and Experimental Botany
journa l homepage: www.e lsev ier .com/ locate /envexpbot
asmonic acid as modulator of lead toxicity in aquatic plant Wolffia arrhizaLemnaceae)
licja Piotrowskaa,∗, Andrzej Bajguza, Beata Godlewska-Zyłkiewiczb, Romuald Czerpaka,ałgorzata Kaminskaa
University of Bialystok, Institute of Biology, Department of Plant Biochemistry and Toxicology, Swierkowa 20 B, 15-950 Bialystok, PolandUniversity of Bialystok, Institute of Chemistry, Department of Analytical Chemistry, Hurtowa 1, 15-399 Bialystok, Poland
r t i c l e i n f o
rticle history:eceived 26 November 2008eceived in revised form 27 March 2009ccepted 31 March 2009
eywords:
a b s t r a c t
The present study was undertaken to test the influence of exogenously applied jasmonic acid (JA) uponthe growth and metabolism of Wolffia arrhiza (Lemnaceae), the smallest vessel aquatic plant exposedto lead (Pb) stress. It was found, that JA acted in a concentration-dependent manner. Treatment withJA at the highest concentration 100 �M resulted in the enhancement of heavy metal toxicity leadingto increase in metal biosorption and formation of lipid peroxides as well as decrease in fresh weight,
ntioxidantsntioxidant enzymesrowth
asmonic acideadolffia arrhiza
chlorophyll a, carotenoid, monosaccharide and soluble protein content. In contrast, this phytohormoneapplied at 0.1 �M protected W. arrhiza fronds against Pb stress inhibiting heavy metal accumulation,restoring plant growth and primary metabolite level. Moreover, JA at 0.1 �M activated enzymatic (catalase,ascorbate peroxidase, NADH peroxidase) and non-enzymatic antioxidant (ascorbate, glutathione) systemin W. arrhiza, and therefore, suppressed oxidative destruction of cellular components induced by heavymetal. The data suggest that JA plays an important role in the growth and metabolism of W. arrhiza exposed
high
to abiotic stressor and its. Introduction
Wolffia arrhiza (Lemnaceae) is the smallest vessel plant aboutmm in length, which neither has leaves nor a stem and even
acks roots. The whole plant called frond may set flowers and seeds,lthough rapid multiplication is achieved by budding. W. arrhiza isne of the important ecological species in freshwater ecosystemsccording to mixotrophic properties, high rate of absorption nutri-nts from polluted water, quick growth, asexual reproduction andesistance to numerous toxins (Fujita et al., 1999). Moreover, plantsrom Lemnaceae family have been identified as potentially usefulroup for accumulating and bioconcentrating heavy metals in phy-oremediation techniques (Hou et al., 2007). Therefore, this free-oating easy to culture in laboratory plant can be used as experi-
ental model system to investigate heavy metal induced responses.Heavy metals, such as Pb are strongly phytotoxic partly becausef the generation of reactive oxygen species (ROS) that damageembranes, lipids, nucleic acids, proteins and photosynthetic pig-
Abbreviations: DTT, dithiothreitol; JA, jasmonic acid; MDA, malondialdehyde;eJA, methyl jasmonate; NEM, N-ethylmaleimide; PVP, polyvinylpyrrolidone; ROS,
eactive oxygen species; TBA, thiobarbituric acid; TCA, trichloroacetic acid.∗ Corresponding author. Tel.: +48 857457292; fax: +48 857457302.
E-mail address: [email protected] (A. Piotrowska).
098-8472/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.envexpbot.2009.03.019
adaptation ability to metal contamination of aquatic environment.© 2009 Elsevier B.V. All rights reserved.
ments (Rucinska-Sobkowiak and Pukacki, 2006). However, plantshave evolved enzymatic (catalase, ascorbate peroxidase, NADH per-oxidase) and non-enzymatic (ascorbate, glutathione) antioxidantmechanisms to prevent the oxidative stress and lipid peroxidationgenerated by heavy metals (Blokhina et al., 2003). Under severestress conditions, however, the antioxidant capacity may not besufficient to minimize the harmful effect of heavy metals. Survivalunder stress conditions may also depend on the plants ability togenerate and transit signals that adjust the metabolism accordingly(Krantev et al., 2008). Therefore, the search for signal moleculesmediating stress tolerance is an important step towards the betterunderstanding how the plants respond to metal pollution. Somedata suggest that jasmonic acid (JA), naturally occurring phyto-hormone, may activate signaling pathways leading to increasedresistance to abiotic stress conditions. The cross-talk in action ofheavy metals and JA is discussed by Poschenrieder et al. (2006,2008). This phytohormone is formed from lipid peroxides as aresult of enhanced lipooxygenase activity, but it is quite possi-ble that the formation may be a prolonged consequence of heavymetal-induction of oxidative stress. Moreover, jasmonates showed
a protective effect against Cu and Cd ions on the growth, chlorophylland carotenoid content as well as photosynthesis in Arabidopsisthaliana (Maksymiec and Krupa, 2002). Despite presented literaturedata, the interaction between JA and heavy metal stress in aquaticplants is still unknown.
5 nd Exp
cetactfaw
2
2
scwciK00Cuwtoiice
2m
tuabmaae(bStweBs
2
(Hwt(a
08 A. Piotrowska et al. / Environmental a
For this reason, the effect of JA on the growth and biochemicalhanges in W. arrhiza exposed to phytotoxic concentration of Pb wasxamined. This study was also undertaken to determine whetherhis phytohormone is involved in induction of defense responsesnd to test the hypothesis that the ability of JA to protect cellularomponents against Pb bioaccumulation is mediated by its effect onhe antioxidant defense system. Obtained results may be importantor better understanding of the plant hormone role in biochemicaldaptation of W. arrhiza to stress conditions present in pollutedater ecosystems.
. Material and methods
.1. Plant material and growth conditions
W. arrhiza (L.) Hork. ex. Wimm. (Lemnaceae) was grown in smallterile, plastic vessels containing 100 mL of culture solution. Theultures were grown under controlled conditions at 25 ± 0.5 ◦C,ith a day/night cycle 16/8 h (photon flux of 50 �mol m−2 s−1). The
ulture medium used was Hutner’s medium (pH 7.0) in the follow-ng nutritive solution: 1.3 mM EDTA, 2 mM MgSO4·7H2O, 2.9 mMH2PO4, 1.5 mM Ca(NO3)2·4H2O, 3.6 mM KOH, 1.7 mM NH4VO3,.2 mM ZnSO4·7H2O, 0.1 mM Na2MoO4·2H2O, 0.1 mM FeSO4·7H2O,.1 mM MnCl2·4H2O, 0.2 mM H3BO3, 20 �M CuSO4·5H2O, 0.7 �Mo(NO3)2·6H2O (Hutner, 1953). 1/50 Dilution of the medium wassed in this study. 0.5 (±0.005) g of fresh weight was treatedith JA (Sigma Chemical Co., USA) at the range of concentra-
ions of 0.01–100 �M on the 1st day of culture. Sterile solutionsf Pb(NO3)2 were added to obtain required concentration of leadn the growing solutions (1–1000 �M). Cultures were conductedn four replications and samples for determination of the bio-hemical parameters were collected on the 7th and 14th day ofxposure.
.2. Determination of growth, photosynthetic pigments,onosaccharides and proteins
For fresh weight determination, the plants were filtered, washedhree times with distilled water, kept on filter paper for a few min-tes to remove of excess liquid and weighed. For chlorophyll and carotenoids determination, the cultures were first collectedy filtration and then the pellets (0.1 g) were homogenized inethanol. The absorbance of the extract was measured at 652.4
nd 665.2 nm for chlorophyll a and 470.0 nm for carotenoids. Themounts of photosynthetic pigments present in the methanolxtract were calculated according to the equations of Wellburn1994). For sugar determination, the cultures were first collectedy filtration and then the pellets (0.1 g) were assessed using theomogyi (1954) method. The measurement of the protein con-ent was done by the homogenization of biomass. The homogenateas centrifuged for 10 min at 12,000 × g and an aliquot of the
xtract was used to determine protein content following theradford (1976) method, using bovine serum albumin as thetandard.
.3. Heavy metal determination
For the determination of total Pb level in W. arrhiza, the biomass1 g) was harvested by filtration and plant material was digested in a
2SO4:HNO3 mixture (1:5, v/v) for 24 h, after which it was treatedith HNO3:HClO4 mixture (5:1, v/v). The concentration of Pb inhe digest was measured using an atomic absorption spectrometerCarl Zeiss Jena, Germany) with a background compensator and anir–acetylene flame.
erimental Botany 66 (2009) 507–513
2.4. Determination of antioxidant level
Extraction and determination of total ascorbate from W. arrhizabiomass was carried out following the method of Kampfenkel et al.(1995). Plant material (1 g) was harvested by filtration and quicklyhomogenized in liquid N2 and thereafter extracted with 5% (w/v)TCA. The homogenate was centrifuged for 5 min at 15,600 × g (4 ◦C).Then the supernatant was transferred to a new reaction vessel andimmediately assayed for the ascorbate content in a reaction mixturecontaining supernatant, 10 mM DTT, 0.2 M phosphate buffer (pH7.4), 0.5% NEM, 10% TCA, 42% H3PO4, 4% 2,2′-dipyridyl and 3% FeCl3.
Determination of glutathione level was essentially as described(De Kok et al., 1986). Briefly, glutathione was extracted from filteredfresh weight in 2 volumes of extracting buffer (2% sulfosalicylicacid, 1 mM Na2EDTA and 0.15% ascorbate) and homogenized. Thehomogenate was centrifuged at 12,000 × g for 5 min. An aliquot ofsupernatant was then used for the measurement of the glutathionecontent in W. arrhiza by glutathione assay kit (Sigma Chemical Co.,USA).
2.5. Determination of the antioxidant enzymes activities
Enzymatic extracts were obtained from 1 g of fresh weight ofW. arrhiza. The biomass has been filtered and homogenized inliquid N2. Next, the samples were homogenized with 0.05 M phos-phate buffer (pH 7.0) containing 0.1 M EDTA and 1% PVP at 4 ◦C.The plant biomass:extraction buffer (w/v) proportion was 1:2. Thehomogenate was centrifuged for 10 min at 15,000 × g (4 ◦C) andthe supernatant was dialyzed overnight in phosphate buffer. Esti-mation of the activity of the selected enzymes was performed asfollows.
Catalase (EC 1.11.1.6) activity was determined following Aeby(1984). The rate of H2O2 decomposition at 240 nm was measuredspectrophotometrically and calculated using a molar extensioncoefficient ε = 45.2 mM−1 cm−1. The reaction mixture consistedof phosphate buffer, 0.1 mM H2O2 and supernatant. One unit ofcatalase activity was assumed as the amount of enzyme thatdecomposed 1 �mol of H2O2 per mg of soluble protein per minuteat 30 ◦C.
Total ascorbate peroxidase (EC 1.11.1.11) was determined accord-ing to the method described by Nakano and Asada (1981). Thereaction mixture consisted of phosphate buffer, 5 mM sodiumascorbate, 0.1 mM H2O2 and supernatant. Total ascorbate perox-idase activity was determined as the decrease in absorbance ofascorbate at 290 nm and calculated using a molar extension coeffi-cient ε = 2.8 mM−1 cm−1. The enzyme activity was calculated as theamount of the enzyme that oxidizes 1 �mol of ascorbate consumedper mg of soluble protein per min at 30 ◦C.
The NADH peroxidase (EC 1.11.1.1) activity was determinedaccording to Ishida et al. (1987). The reaction mixture consisted of50 mM pH 6.0 sodium acetate buffer and 0.2 mM NADH. The reac-tion was initiated by adding the enzymatic extract and lasted for upto 5 min. The peroxidase activators p-cumaric acid and 5 nM MnCl2were used. The activity was calculated with a molar extinctioncoefficient ε = 6.300 mM−1 cm−1 for NADH. One unit of NADH per-oxidase activity was assumed as the amount of the enzyme whichoxidizes 1 �mol of NADH per mg of soluble protein per min at 30 ◦C.
2.6. Determination of lipid peroxidation
The lipid peroxidation in the W. arrhiza biomass was determined
as MDA content by the TBA reaction as described by Heath andPacker (1968). Filtered biomass (1 g) was homogenized in 0.1% TCA.The homogenate was centrifuged at 10,000 × g for 5 min. Reactionmixture containing supernatant, 20% TCA and 0.5% TBA was heatedat 95 ◦C for 30 min and then quickly cooled on ice. After centrifuga-
nd Exp
tt
2
wpto
3
3
nFcca1dP0
i
Fad±
A. Piotrowska et al. / Environmental a
ion at 10,000 × g for 10 min, the absorbance at 532 nm was read andhe value for the non-specific absorption at 600 nm was subtracted.
.7. Replication and statistical analysis
Each treatment consisted of four replicates and each experimentas carried out at least twice at different times. A minitab statisticalackage was used to carry out a one-way ANOVA. The Student’s t-est was used to estimate the difference between means at 5% levelf significance.
. Results
.1. Growth of W. arrhiza
Preliminary experiments showed that presence of Pb in theutrient solution inhibited the growth of W. arrhiza culture (Fig. 1A).resh weight decreased proportionally with increasing Pb con-entration. The most phytotoxic effect of Pb was observed atoncentration 1000 �M resulting in cellular fragmentation, lysisnd finally in the death of the plant culture. Pb at lower dose00 �M caused significant 62 and 80% decrease in biomass pro-
uction on the 7th and 14th day of culture, respectively. Therefore,b at 100 �M combined with JA at the range of concentrations.01–100 �M were used in the following experiments.JA at the highest dose 100 �M strongly enhanced the inhibitorynfluence of the heavy metal on plant growth leading to 73 and 89%
ig. 1. Growth of Wolffia arrhiza culture under the influence of 1–1000 �M Pb (A)nd 0.1–100 �M jasmonic acid during 100 �M Pb stress (B). Fresh weight on the 1stay of culture 0.50 (±0.05) g. Data are the means of four independent experimentsS.D.
erimental Botany 66 (2009) 507–513 509
decrease in fresh weigh on the 7th and 14th day of experiment,respectively (Fig. 1B). On the other hand, JA at 0.1 �M showed theopposite effect on W. arrhiza exposed to 100 �M Pb inducing 8–25%increase in the fresh weight.
3.2. Photosynthetic pigment, monosaccharide and protein content
W. arrhiza fronds treated with 100 �M Pb displayed chlorosisbecause 58% decline in chlorophyll a and 35% decrease in carotenoidcontent was found during the whole 14-day period of the culture(Fig. 2A and B). These symptoms were enhanced after JA applica-tion at extremely inhibitory dose (100 �M) on plant growth leadingto 70 and 51% reduction in chlorophyll a and carotenoids contents,respectively on the 14th day of culture. However, treatment plantswith 0.1 �M JA reduced the phytotoxic effect of Pb on the pho-tosynthetic pigments and led to restoration of chlorophyll a andcarotenoid level.
The destruction of photosynthetic pigments in plants exposed to100 �M Pb was accompanied by significant 42–65% on the averagedecrease in the monosaccharide content (Fig. 2C). When W. arrhizawas treated with both 100 �M Pb and 100 �M JA this biochemicalparameter reached the minimum value during 2 weeks of the cul-ture. By contrast, JA at lower dose (0.1 �M) showed additive effecton the monosaccharides in fronds exposed to heavy metal resultingin twofold higher increase in their content.
The application of 100 �M Pb resulted in 56% reduction in theprotein content after 14 days exposure (Fig. 2D). This phytotoxiceffect of heavy metal was enhanced in the presence of 100 �M JAbecause protein level decreased by 44 and 66% on the 7th and 14thday, respectively. On the other hand W. arrhiza plants treated withboth 100 �M Pb and JA at 0.1 �M were characterized by 46–50%restoration of protein content.
3.3. Pb bioaccumulation
JA influenced Pb biosorption by W. arrhiza plants in aconcentration-dependent trend (Fig. 3). The highest Pb contentwas observed in the culture after exogenous application of both100 �M Pb and 100 �M JA, where it reached 0.49 and 0.90 mg g−1
fresh wt. on the 7th and 14th day of cultivation, respectively. Bycomparison, JA at lower concentrations inhibited the Pb biosorp-tion from the nutrient solution. For example, JA at 0.1 �M reducedthe accumulation of Pb in biomass to 0.06 mg g−1 fresh wt. andto 0.09 mg g−1 fresh wt. on the 7th and 14th day of the culture,respectively.
3.4. Lipid peroxidation
Because Pb is known to induce an oxidative stress, damage to themembranes was investigated by monitoring the content of MDA,the toxic product of lipid peroxidation. Relative to the control Pb-treated plants exhibited higher by 92–106% rate of lipid peroxidesformation (Fig. 4). The stimulation of lipid peroxidation by 108and 140% was observed under the influence of exogenous 100 �MPb and 100 �M JA on the 7th and 14th day of culture, respec-tively. However, the application of JA at 0.1 �M inhibited membranedestruction by heavy metal, because the significant decrease inMDA level was observed.
3.5. Levels of ascorbate and glutathione
The presence of exogenous Pb resulted in non-enzymatic antiox-idant response in W. arrhiza fronds expressed as the increase inascorbate and glutathione level in relation to the control culture(Fig. 5). Application of both 100 �M Pb and JA at the most stimulat-ing concentration 0.1 �M showed an additive effect on the content

510 A. Piotrowska et al. / Environmental and Experimental Botany 66 (2009) 507–513
F hyll aT 4.36)( s of fo
oa0
3
s
Fmf
ig. 2. The combined effect of 0.1–100 �M jasmonic acid and 100 �M Pb on chlorophe biochemical parameter level on the 1st day of culture: chlorophyll a 205.04 (±±0.35) mg g−1 fresh wt., proteins 24.77 (±0.93) mg g−1 fresh wt. Data are the mean
f antioxidants in fronds. The threefold higher increase in ascorbatend glutathione accumulation was observed after 100 �M Pb and.1 �M JA supply during 2 weeks of cultivation.
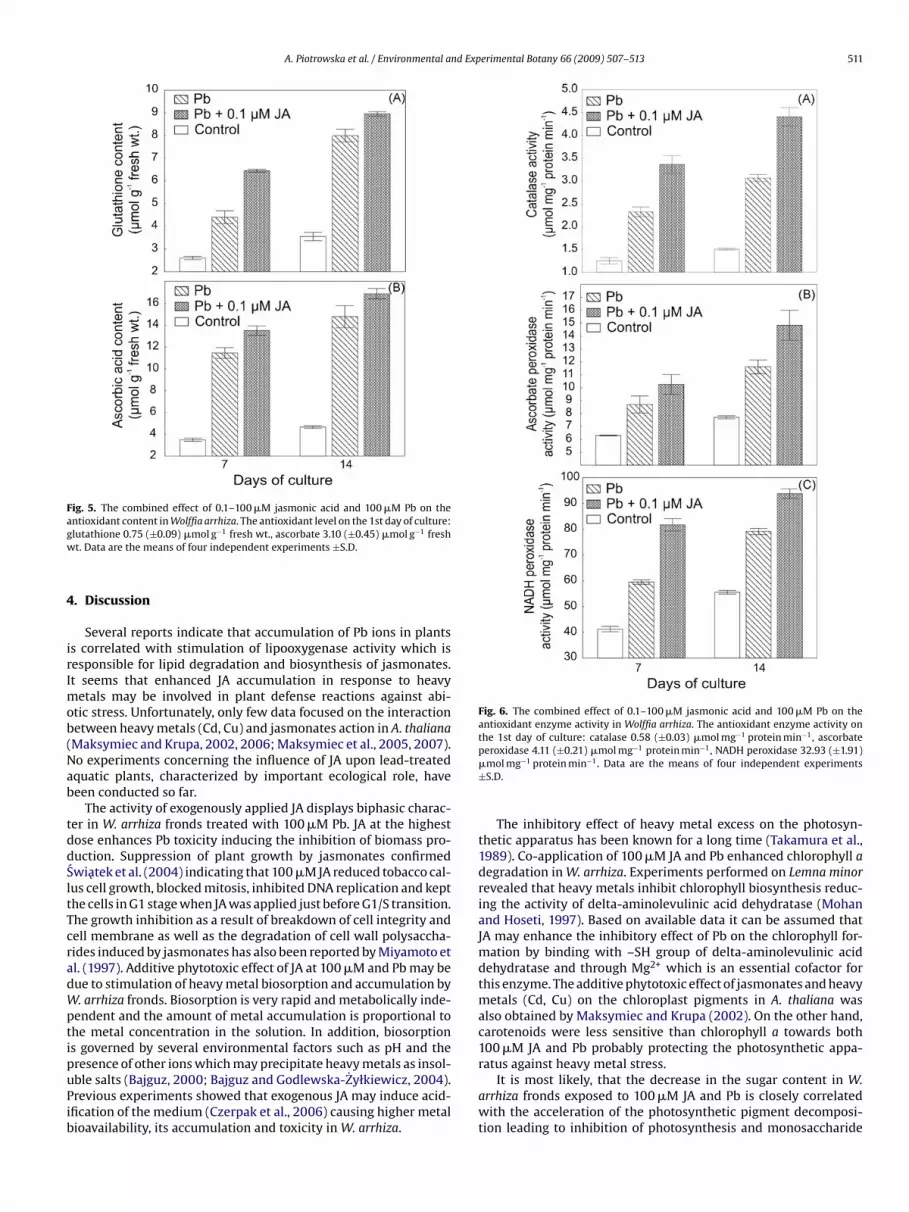
.6. Activities of antioxidant enzymes
The presence of 100 �M Pb in the nutrient solution led to thetimulation of antioxidant enzymes in W. arrhiza fronds (Fig. 6).
ig. 3. The combined effect of 0.1–100 �M jasmonic acid and 100 �M Pb on heavyetal biosorption in Wolffia arrhiza. The Pb level on the 1st day of culture 0 mg g−1
resh wt. Data are the means of four independent experiments ±S.D.
(A), carotenoids (B), monosaccharide (C), and protein (D) content in Wolffia arrhiza.�g g−1 fresh wt., carotenoids 65.41 (±2.35) �g g−1 fresh wt., monosaccharides 3.11ur independent experiments ±S.D.
Moreover, the significant enhancement of the enzymes activitiesinvolved in scavenging of ROS appeared as a consequence of plantexposure to 0.1 �M JA plus 100 �M Pb. Exogenously applied JA at0.1 �M and Pb at 100 �M possessed additive effect on ascorbateperoxidase which activity was stimulated by 63 and 93% on the
7th and 14th day. The significant increase by 67–98% in NADH per-oxidase activity was also observed when plants were treated withJA at 0.1 �M and Pb at 100 �M. Analogically catalase activity waselevated by 164 and 190% on the 7th and 14th day, respectively.Fig. 4. The combined effect of 0.1–100 �M jasmonic acid and 100 �M Pb on the lipidperoxidation in Wolffia arrhiza. The MDA level on the 1st day of culture 15.09 (±0.9)nmol g−1 fresh wt. Data are the means of four independent experiments ±S.D.
A. Piotrowska et al. / Environmental and Experimental Botany 66 (2009) 507–513 511
Fig. 5. The combined effect of 0.1–100 �M jasmonic acid and 100 �M Pb on theagw
4
irImob(Nab
tddSltTcradWptipuPib
Fig. 6. The combined effect of 0.1–100 �M jasmonic acid and 100 �M Pb on theantioxidant enzyme activity in Wolffia arrhiza. The antioxidant enzyme activity onthe 1st day of culture: catalase 0.58 (±0.03) �mol mg−1 protein min−1, ascorbate
ntioxidant content in Wolffia arrhiza. The antioxidant level on the 1st day of culture:lutathione 0.75 (±0.09) �mol g−1 fresh wt., ascorbate 3.10 (±0.45) �mol g−1 fresht. Data are the means of four independent experiments ±S.D.
. Discussion
Several reports indicate that accumulation of Pb ions in plantss correlated with stimulation of lipooxygenase activity which isesponsible for lipid degradation and biosynthesis of jasmonates.t seems that enhanced JA accumulation in response to heavy
etals may be involved in plant defense reactions against abi-tic stress. Unfortunately, only few data focused on the interactionetween heavy metals (Cd, Cu) and jasmonates action in A. thalianaMaksymiec and Krupa, 2002, 2006; Maksymiec et al., 2005, 2007).o experiments concerning the influence of JA upon lead-treatedquatic plants, characterized by important ecological role, haveeen conducted so far.
The activity of exogenously applied JA displays biphasic charac-er in W. arrhiza fronds treated with 100 �M Pb. JA at the highestose enhances Pb toxicity inducing the inhibition of biomass pro-uction. Suppression of plant growth by jasmonates confirmed
´wiatek et al. (2004) indicating that 100 �M JA reduced tobacco cal-us cell growth, blocked mitosis, inhibited DNA replication and kepthe cells in G1 stage when JA was applied just before G1/S transition.he growth inhibition as a result of breakdown of cell integrity andell membrane as well as the degradation of cell wall polysaccha-ides induced by jasmonates has also been reported by Miyamoto etl. (1997). Additive phytotoxic effect of JA at 100 �M and Pb may beue to stimulation of heavy metal biosorption and accumulation by. arrhiza fronds. Biosorption is very rapid and metabolically inde-
endent and the amount of metal accumulation is proportional tohe metal concentration in the solution. In addition, biosorptions governed by several environmental factors such as pH and theresence of other ions which may precipitate heavy metals as insol-
ble salts (Bajguz, 2000; Bajguz and Godlewska-Zyłkiewicz, 2004).revious experiments showed that exogenous JA may induce acid-fication of the medium (Czerpak et al., 2006) causing higher metalioavailability, its accumulation and toxicity in W. arrhiza.peroxidase 4.11 (±0.21) �mol mg−1 protein min−1, NADH peroxidase 32.93 (±1.91)�mol mg−1 protein min−1. Data are the means of four independent experiments±S.D.
The inhibitory effect of heavy metal excess on the photosyn-thetic apparatus has been known for a long time (Takamura et al.,1989). Co-application of 100 �M JA and Pb enhanced chlorophyll adegradation in W. arrhiza. Experiments performed on Lemna minorrevealed that heavy metals inhibit chlorophyll biosynthesis reduc-ing the activity of delta-aminolevulinic acid dehydratase (Mohanand Hoseti, 1997). Based on available data it can be assumed thatJA may enhance the inhibitory effect of Pb on the chlorophyll for-mation by binding with –SH group of delta-aminolevulinic aciddehydratase and through Mg2+ which is an essential cofactor forthis enzyme. The additive phytotoxic effect of jasmonates and heavymetals (Cd, Cu) on the chloroplast pigments in A. thaliana wasalso obtained by Maksymiec and Krupa (2002). On the other hand,carotenoids were less sensitive than chlorophyll a towards both100 �M JA and Pb probably protecting the photosynthetic appa-ratus against heavy metal stress.
It is most likely, that the decrease in the sugar content in W.arrhiza fronds exposed to 100 �M JA and Pb is closely correlatedwith the acceleration of the photosynthetic pigment decomposi-tion leading to inhibition of photosynthesis and monosaccharide

5 nd Exp
botfoppClelepcatsmae
dtirpidaKatscg
malSdibeC
cmrapspttiiboe
iEpsp
12 A. Piotrowska et al. / Environmental a
iosynthesis. JA at the highest dose might enhance the sensitivityf photosynthetic apparatus of W. arrhiza to Pb stress. This observa-ion is in good agreement with the increased rate of lipid peroxidesormation expressed as MDA content in fronds under the influencef both 100 �M JA and Pb. MDA is the decomposition product ofolysaturated fatty acids of biomembranes and its increase showslants are under high-level oxidative stress (Singh et al., 2006).hloroplasts are unique organelles that contain a relatively high
evel of unsaturated fatty acids in its membranes, therefore arespecially sensitive to stress factors generating oxidative burst andipid peroxidation (Apel and Hirt, 2004). As suspected, stimulatingffect of 100 �M JA plus Pb on MDA level in W. arrhiza indicatedlants encountered enhanced lipid peroxidation causing oxidativeellular damage, inhibition of the growth and development as wells the loss in photosynthetic pigment and monosaccharide con-ent. Our finding also corresponds well with another literature datahowing that high amounts of JA are accumulated during heavyetal stress in A. thaliana and Phaseolus coccineus (Maksymiec et
l., 2005) accelerating the senescence program and plant death (Het al., 2002).
Cultivation of W. arrhiza with 100 �M JA and Pb resulted in theegradation of proteins. Soluble protein content in plants, an impor-ant indicator of reversible and irreversible changes in metabolism,s known to respond to a wide variety of stressors such as natu-al and xenobiotics. Probably, 100 �M JA enhances the damage ofroteins caused by acute oxidative stress induced by heavy metals
n plant cells. Moreover, heavy metals could induce DNA break-own that may significantly affect RNA and protein synthesis. Theccumulation of Pb in plant cells blocks also the entry of Mg andon which protein synthesis system relied (Gardea-Torresdey et
l., 2004). The mechanism of Pb-induced protein degradation inhe presence of high dose of JA is complex and needs furthertudy. Summarizing, JA at higher doses accelerates Pb biosorptionontributing to oxidative degradation of primary metabolites androwth inhibition.
However, the response of W. arrhiza to both Pb and JA isore multifaceted. JA applied at lower concentrations, especially
t 0.1 �M, displays protective function against heavy metal stresseading to restoration of fresh weight. It is not surprising becausecenedesmus incrassulatus (Chlorophyceae) showed enhanced cellivision and algal viability in response to exogenous jasmonate
n temperature stress (Christov et al., 2001). Similar data haveeen also reported by Maksymiec and Krupa (2002) indicating thatxogenous MeJA at 1 �M did not enhance the inhibitory action ofu and Cd on A. thaliana growth.
The protective role of 0.1 �M JA against Pb toxicity in W. arrhizaulture may be explained by JA-induced inhibition of Pb bioaccu-ulation which allow plant cells to maintain precise homeostatic
egulation of intracellular heavy metal level. The removal of met-ls from highly polluted waters may depend on the presence oflant growth regulators and physiology of plants. For example,alicylic acid pretreatment alleviated the negative effect of Cd onlant growth parameters reducing the accumulation this metal inhe maize roots (Krantev et al., 2008). The results exclude alsohe possibility that JA is an effective signaling molecule upregulat-ng the expression of genes for phytochelatin synthesis extensivelynvolved in heavy metal detoxification. Our hypothesis is supportedy observation that MeJA stimulated significantly the accumulationf phytochelatins in A. thaliana in response to Cu and Cd leading tonhanced plant tolerance to stress (Maksymiec et al., 2007).
In addition to above presented facts, JA at 0.1 �M is character-
zed by protective role on proteins in W. arrhiza exposed to Pb.xplanation for JA-induced increase in protein level despite theresence of toxic concentration of heavy metal may be referred totimulation of the expression of genes resulting in the synthesis ofroteins with the protective function. Many studies have describederimental Botany 66 (2009) 507–513
JA-regulated genes expressed under stress. For example, the biosyn-thesis of 3-subtilisin-like serine proteases (At-SLP2) is induced byJA and Cd in A. thaliana (Golldack et al., 2003). Furthermore, theexpression of jasmonic acid-inducible rice MAP kinase gene (OsB-WMK1) is induced by diverse abiotic stresses such as Cu, Cd andHg (Agrawal et al., 2003) suggesting that JA and heavy metals mayactivate similar defense responses.
The results suggest that the application of 0.1 �M JA has a benefi-cial effect on photosynthetic apparatus in W. arrhiza treated with Pb.Owing to photosynthetic pigments’ protection function from unfa-vorable influence UV irradiation, oxidative stress and heavy metals,the increase in their content has a positive effect on the adaptationability of W. arrhiza plants to environment pollution. Carotenoids,which play the part of guard of chlorophyll, serve also as antioxi-dants to quench or scavenge the free radicals and reduce the damageof cell, cell membrane and its main genetic composition induced byheavy metal (Czerpak et al., 2006). The stimulation of photosynthe-sis by 0.1 �M JA applied together with Pb contributes significantly tothe production of monosaccharides which are building substancesfor plant and play a key role as source of energy. This finding is alsocomparable with data indicating JA affects the mobilization of stor-age sugars by stimulation of polysaccharide hydrolysis (Bogatek etal., 2002).
In the light of presented facts, we can summarize that JA at0.1 �M is characterized by unusual biological and antistress activityin W. arrhiza culture. However, an important question in this studywas how JA may help plants to avoid cellular damage upon exposureto Pb which is extensively involved in oxidative stress generation.For this reason, the present study was designed to test the hypoth-esis that JA at optimal stimulating concentration 0.1 �M possessessignificant role in activating antioxidant system in W. arrhiza treatedwith Pb.
Our results revealed that JA at 0.1 �M may be involved in inhi-bition of lipid peroxides formation which was obvious from lowerMDA content in W. arrhiza exposed to Pb. JA diminishes lipid per-oxidation through the stimulation of non-enzymatic (ascorbate,glutathione) antioxidant machinery responsible for a tight regu-lation of ROS homeostasis during heavy metal stress. Ascorbateis known to operate as an antioxidant either in direct chemicalinteraction with ROS, or during the reaction catalyzed by ascorbateperoxidase (Kampfenkel et al., 1995; Singh et al., 2006). Ascorbatederives its role from its sensitivity to ROS and from the fact thatits oxidation affects the redox balance of other metabolites, suchas glutathione which being themselves involved in the perceptionof the cellular redox unbalance contributed to signal amplification(Apel and Hirt, 2004). Glutathione is an important water-phaseantioxidant with proposed roles in the storage and transport ofreduced sulfur, the synthesis of proteins and nucleic acids and asmodulator of enzyme activity (De Kok et al., 1986). The glutathionelevel has been shown to correlate with the plant adaptation toextreme heavy metal stress and reduced glutathione pool showsmarked alterations in response to heavy metal stress (Singh et al.,2006; Jin et al., 2008). Moreover, glutathione is also a precursor ofphytochelatins, low molecular mass peptides produced by plantsto immobilize heavy metals. Xiang and Oliver (1998) indicatedthat JA stimulated the expression of enzymes of the glutathionebiosynthetic pathways leading to an increase in phytochelatinbiosynthesis and heavy metal tolerance in A. thaliana. Therefore,our results ruled out the possibility that JA-induced increase inascorbate and glutathione content plays a key role in W. arrhizatolerance to Pb providing the protection against oxidative stress
and that glutathione accumulation may precede the phytochelatinsynthesis.Apart from the additive effect of 0.1 �M JA and Pb on the non-enzymatic antioxidants, jasmonate and heavy metal affected theenzymatic antioxidants. The majority of ROS-scavenging pathways

nd Exp
oiattcw(potsgasi(as
iaaPdatdctmntotn
R
AA
A
B
B
B
B
B
C
C
C
A. Piotrowska et al. / Environmental a
f plants include ascorbate peroxidase, catalase and NADH perox-dase which can prevent H2O2 accumulation. Enhanced activity ofscorbate peroxidase is generally associated with an acclimationo elevated amounts of active oxygen species and plant adapta-ion to environmental changes. Moreover, ascorbate peroxidase canooperate with NADH peroxidase stimulating lignin biosynthesishich may build up a physical barrier against toxic heavy metal
Chaoui et al., 2004; De Pinto and De Gara, 2004). Catalase partici-ates in the main defense system against accumulation and toxicityf active oxygen species, such as hydrogen peroxide and can con-rol H2O2 level in plant cells (Blokhina et al., 2003). As suspected,timulation of antioxidant enzymes by both 0.1 �M JA and Pb is inood agreement with decreased rate of lipid peroxides productionnd increased plant tolerance to the presence of heavy metal. Aimilar effect of both heavy metals and jasmonates on the antiox-dant enzymes activities was reported by Maksymiec and Krupa2002) for A. thaliana. Summarizing, JA at low dose may protect W.rrhiza against Pb toxicity through the activation of antioxidativeystem.
The physiological role of JA is usually considered to be signal-ng molecule extensively involved in plant responses to biotic andbiotic stress (Wasternack, 2007). Obtained data suggest that JA isn important plant hormone modifying resistance of W. arrhiza tob and increases the stress or/and enhances the plant resistanceepending on its concentration. JA at higher dose acts as stressornd stimulates Pb biosorption as well as lipid peroxides forma-ion leading therefore to inhibition of plant growth and metaboliteegradation. On the other hand, JA at lower concentrations, espe-ially at 0.1 �M displays the beneficial effect which may help planto avoid cumulative damage upon exposure to Pb. Probably, the
echanism of plant stress tolerance induced by JA is closely con-ected with the blockade of heavy metal entry to the cell andhe activation of antioxidant defense responses to minimize thexidative damage induced by Pb presence. However, the interac-ive action between heavy metal and jasmonates is not clear andeeds further investigation.
eferences
eby, H., 1984. Catalase in vitro. Methods Enzymol. 105, 125–212.pel, K., Hirt, H., 2004. Reactive oxygen species: metabolism, oxidative stress and
signal transduction. Annu. Rev. Plant Biol. 55, 373–399.grawal, G.K., Tamogami, S., Iwahashi, H., Agrawal, V.P., Rakwal, R., 2003. Tran-
sient regulation of jasmonic acid-inducible rice MAP kinase gene (OsBWMK1)by diverse biotic and abiotic stresses. Plant Physiol. Biochem. 41, 355–361.
ajguz, A., 2000. Blockade of heavy metals accumulation in Chlorella vulgaris cellsby 24-epibrassinolide. Plant Physiol. Biochem. 38, 797–801.
ajguz, A., Godlewska-Zyłkiewicz, B., 2004. Protective role of 20-hydroxyecdysoneagainst lead stress in Chlorella vulgaris cultures. Phytochemistry 65,711–720.
lokhina, O., Virolainen, E., Fagerstedt, K.V., 2003. Antioxidants, oxidative damageand oxygen deprivation stress: a review. Ann. Bot. 91, 179–194.
ogatek, R., Côme, D., Corbineau, F., Ranjan, R., Lewak, S., 2002. Jasmonic acid affectsdormancy and sugar catabolism in germinating apple embryos. Plant Physiol.Biochem. 40, 167–173.
radford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein–dye binding. Anal.Biochem. 72, 248–254.
haoui, A., Jarrar, B., El Ferjani, E., 2004. Effects of cadmium and copper on peroxidase,NADH oxidase and IAA oxidase activities in cell wall, soluble and microsomalmembrane fractions of pea roots. J. Plant Physiol. 161, 1225–1234.
hristov, C., Pounieva, M., Bozhkova, M., Toncheva, S., Fournadzieva, S., Zafirova, T.,2001. Influence of temperature and methyl jasmonate on Scenedesmus incrassu-latus. Biol. Plant. 44, 367–371.
zerpak, R., Piotrowska, A., Szulecka, K., 2006. Jasmonic acid affects changes in thegrowth and some components content in alga Chlorella vulgaris. Acta Physiol.Plant. 28, 195–203.
erimental Botany 66 (2009) 507–513 513
De Kok, L.J., Maas, F.M., Godeke, J., Haaksma, A.B., Kuiper, P.J.C., 1986. Glutathione, atripeptide which may function as a temporary storage compound of excessivereduced sulphur in H2S fumigated spinach plants. Plant Soil 91, 349–352.
De Pinto, M.C., De Gara, L., 2004. Changes in the ascorbate metabolism of apoplasticand symplastic spaces are associated with cell differentiation. J. Exp. Bot. 55,2559–2569.
Fujita, M., Mori, K., Kodera, T., 1999. Nutrient removal and starch production throughcultivation of Wolffia arrhiza. J. Biosci. Bioeng. 87, 194–198.
Gardea-Torresdey, J.L., Peralta-Videa, J.R., Montes, M., de la Rosa, G., Corral-Diaz, B.,2004. Bioaccumulation of cadmium, chromium and copper by Convolvulus arven-sis L.: impact on plant growth and uptake of nutritional elements. Bioresour.Technol. 92, 229–235.
Golldack, D., Vera, P., Dietz, K.J., 2003. Expression of subtilisin-like serine proteasesin Arabidopsis thaliana is cell-specific and responds to jasmonic acid and heavymetals with developmental differences. Physiol. Plant. 118, 64–73.
He, Y., Fukushige, H., Hildebrand, D.F., Gan, S., 2002. Evidence supporting a role ofjasmonic acid in Arabidopsis leaf senescence. Plant Physiol. 128, 876–884.
Heath, R.L., Packer, L., 1968. Photoperoxidation in isolated chloroplasts. I. Kinetics andstoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125, 189–198.
Hou, W., Chen, X., Song, G., Wang, Q., Chang, C.C., 2007. Effects of copper and cadmiumon heavy metal polluted waterbody restoration by duckweed (Lemna minor).Plant Physiol. Biochem. 45, 62–69.
Hutner, S.H., 1953. Comparative physiology of heterotrophic growth in plants. In:Loomis, W.E. (Ed.), Growth and Differentiation in Plants. Iowa State College Press,Ames, pp. 417–446.
Ishida, A., Ookubu, K., Ono, K., 1987. Formation of hydrogen peroxide by NAD(P)Hoxidation with isolated cell wall-associated peroxidase from cultured liverwortcells, Marchantia polymorpha L. Plant Cell Physiol. 28, 723–726.
Jin, X.F., Yang, X.E., Islam, E., Liu, D., Mahmood, Q., Li, H., Li, J., 2008. Ultrastruc-tural changes, zinc hyperaccumulation and its relation with antioxidants in twoecotypes of Sedum alfredii Hance. Plant Physiol. Biochem. 46, 997–1006.
Kampfenkel, K., Van Montagu, M., Inzé, D., 1995. Extraction and determination ofascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 225, 165–167.
Krantev, A., Yordanova, R., Janda, T., Szalai, G., Popova, L., 2008. Treatment with sali-cylic acid decreases the effect of cadmium on photosynthesis in maize plants. J.Plant Physiol. 165, 920–931.
Maksymiec, W., Krupa, Z., 2002. Jasmonic acid and heavy metals in Arabidopsisthaliana—a similar physiological responses to both stressors? J. Plant Physiol.159, 509–515.
Maksymiec, W., Krupa, Z., 2006. The effects of short-term exposition to Cd, excessCu ions and jasmonate on oxidative stress appearing in Arabidopsis thaliana.Environ. Exp. Bot. 57, 187–194.
Maksymiec, W., Wianowska, D., Dawidowicz, A.L., Radkiewicz, S., Mardarowicz, M.,Krupa, Z., 2005. The level of jasmonic acid in Arabidopsis thaliana and Phaseoluscoccineus plants under heavy metal stress. J. Plant Physiol. 162, 1338–1346.
Maksymiec, W., Wójcik, M., Krupa, Z., 2007. Variation in oxidative stress and photo-chemical activity in Arabidopsis thaliana leaves subjected to cadmium and excesscopper in the presence or absence of jasmonate and ascorbate. Chemosphere 66,421–427.
Miyamoto, K., Oka, M., Ueda, J., 1997. Update in the possible mode of action of jas-monates: focus on the metabolism of cell wall polysaccharides in relation togrowth and development. Physiol. Plant. 100, 631–638.
Mohan, B.S., Hoseti, B.B., 1997. Potential phytotoxicity of lead and cadmium to Lemnaminor grown in sewage stabilization ponds. Environ. Pollut. 98, 233–238.
Nakano, Y., Asada, K., 1981. Hydrogen peroxidase is scavenged by ascorbate-specificperoxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880.
Poschenrieder, C., Gunsé, B., Corrales, I., Barceló, J., 2008. A glance into aluminumtoxicity and resistance in plants. Sci. Total Environ. 400, 356–368.
Poschenrieder, C., Tolrà, R., Barceló, J., 2006. Can metals defend plants against bioticstress? Trends Plant Sci. 11, 288–295.
Rucinska-Sobkowiak, R., Pukacki, P.M., 2006. Antioxidative defense system in lupinroots expose to increasing concentrations of lead. Acta Physiol. Plant. 28,357–364.
Singh, S., Eapen, S., D’Souza, S.F., 2006. Cadmium accumulation and its influence onlipid peroxidation and antioxidative system in an aquatic plant Bacopa monnieriL. Chemosphere 62, 233–246.
Somogyi, M., 1954. Notes on sugar determination. J. Biol. Chem. 195, 19–23.Swiatek, A., van Dongen, W., Esmans, E.L., van Onckelen, H.A., 2004. Metabolic fate
of jasmonates in tobacco Bright Yellow-2 cells. Plant Physiol. 135, 161–172.Takamura, N., Kusai, F., Watanabe, M., 1989. Effects of Cu, Cd, Zn on photosynthesis
of freshwater benthic algae. J. Appl. Phycol. 1, 39–52.Wasternack, C., 2007. Jasmonates: an update on biosynthesis, signal transduction
and action in plant stress response, growth and development. Ann. Bot. 100,
681–697.Wellburn, A.R., 1994. The spectral determination of chlorophylls a and b, as well astotal carotenoids, using various solvents with spectrophotometers of differentresolution. J. Plant Physiol. 144, 307–313.
Xiang, C., Oliver, D.J., 1998. Glutathione metabolic genes coordinately respond toheavy metals and jasmonic acid in Arabidopsis. Plant Cell 10, 1530–1550.