resveratrol production as a part of the hypersensitive-like response of grapevine cells to an...
TRANSCRIPT

New PhytoL (1993), 124, 455-463
Resveratrol production as a part of thehypersensitive-like response of grapevinecells to an elicitor from Trichoderma viride
BY ANTONIO A. CALDERON, JOSE M. ZAPATA,ROMUALDO MUNOZ, MARIA A. PEDRENO AND A. ROS BARCELO*
Department of Plant Biology {Plant Physiology), University of Murcia,E-30100 Murcia, Spain
{Received 13 December 1992; accepted 18 March 1993)
SUMMARY
Suspension cell cultures of grapevine {Vitis vinifera, cv. Monastrell) treated with an elicitor (cellulase, OnozukaR-10) from Trichoderma viride showed a hypersensitive-like response. This was characterized by cell plasmolysisand was accompanied by localized cell death, which was concomitant with cell culture browning, itself probablydue to an activation of oxidative phenolic metabolism driven by a large increase in endogenous levels of H. O2. Inaddition to these responses, the treatment of cell cultures with the elicitor produced an increase in amounts ofbenzoic acid and of resveratrol, the latter a potent phytoalexin of grapevines. This hypersensitive-like response wasspecific since none of the above responses was obtained with other cell wall-degrading enzymes from severalsources, or with inocula of either mycelial extracts or culture filtrates of Botrytis cinerea. These results arediscussed in the light of a disease-resistance reaction induced in grapevine cells by a product of T. viride, a fungalagent characterized by its effective biocontrol of Botrytis cinerea, the causal agent of grey mould in grapevines.
Key words: Benzoic acid, elicitor (cellulase Onozuka R-10), Vitis vinifera (grapevine), suspension cell cultures,induced resistance, resveratrol, Trichoderma viride.
INTRODUCTION
Trichoderma and Gliocladium spp. are thought tohave potential for the biological control of plant-pathogenic fungi (Papavizas, 1985). This direct effecton plant protection can also be brought about bybacteria and other fungi (Adams, 1990; Lugtenberg,de Weger & Bennett, 1991). Interest in biocontrolhas gained momentum, due in part to the in-creasingly negative attitude towards chemical pesti-cides (Lugtenberg et al., 1991).
Four mechanisms have been proposed to explainthe effect of fungal antagonists: concurrence, anti-biosis, mycoparasitism and induced resistance(Lugtenberg et al, 1991). Induced resistance hasbeen defined as a dynamic resistance based onphysical and chemical barriers induced by pre-liminary or concomitant inoculation with an in-compatible pathogen or non-pathogen, or by treat-ment with their products (Ouchi, 1983).
Resistance can be induced by biocontrol agentsthrough the production of extracellular enzymes and
* To whom all correspondence should be addressed.
metabolites that trigger the hypersensitive responsein the host (Lugtenberg et al., 1991). The hyper-sensitive response (HR), characterized by a rapidcollapse and desiccation of the host tissue in the areaimmediately adjacent to the site of attemptedpenetration (Ouchi, 1983), is often accompanied by arapid production of phytoalexins (Darvill & Alber-sheim, 1984).
Experiments with Trichoderma spp. have revealedthat this antagonist may be used as an effectivebiocontrol agent against Botrytis cinerea in grape-vines (Dubos, Jailloux & Bulit, 1982). We havestudied the ability of an elicitor obtained fromTrichoderma viride Pers. ex fr. (cellulase Onozuka R-10) to induce HR in suspension-cultured cells ofMonastrell {Vitis vinifera), a grapevine cultivarsusceptible to B. cinerea.
M.ATERIALS AND METHODSPlant material and culture
Suspension cell cultures from the pericarp ofimmature grapes {Vitis vinifera, cv. Monastrell), of
31 ANP 124

456 A. A. Calderdn and others
approx. 5 mm diameter, were grown in Gamborg'sB5 medium at 25 °C in the dark (Zapata et al., 1991).
Flicitor
The elicitor (cellulase Onozuka R-10, ex. Tricho-derma viride, c. 1-34 U mg">) was purchased fromServa Feinbiochemica GmbH (Heidelberg, FRG).Examination of this crude enzyme fraction by themanufacturer shows the presence of pectinase(0-53 U mg^') and hemicellulase activities(M Umg-^).
Grapevine cell cultures were treated with differentconcentrations of the T. viride elicitor (in H . ^ )during the log phase of the growth cycle. The elicitorwas sterilized by filtration through a 0 2 fim pore-size filter with luer lock fitting (Sigma Chemical Co.,Madrid, Spain). Controls were carried out with0-1 mg m r ' autoclaved elicitor, and in the absence ofelicitor.
Other fungal preparations
Botrytis cinerea Pers. CECT 2100 (Coleccion Espan-ola de Cultivos Tipo, Department of Microbiology,University of Valencia, Spain) was grown at 25 °C instationary culture in 250 ml conical fiasks containing100 ml of 2"o (w/v) bacteriological malt extract(Biokar S.A., France) for 28 d.
After sterilization by filtration, elicitation ofsuspension-cultured grapevine cells by the culturefiltrate or mycelial extract of Botrytis cinerea wasassayed by dilution 1 :10 (v/v) of the inocula in theplant cell culture medium. For this, the culturefiltrate was used directly. A mycelial extract wasobtained by grinding the frozen mycelium (1 vol.) ina Sorvall Omni-mixer (Sorvall Inc., Newton, Con-necticut, USA), this being extracted with 5 vol. ofredistilled and deionized water.
Other fungal preparations used as elicitors werepectinase from Rhizopus (Sigma), pectinase fromAspergillus niger (Serva) and pectolyase from As-pergillus faponicus (Sigma).
Cell wall preparations and hydrolysates
Cell wall fractions were obtained from suspension-cultured cells by resuspension of 40 g of packed cellsin 200 ml H.O. Cells were killed by immersion in aboiling water bath for 10 min and homogenized in amechanical blender in the presence of 0-1 M sodiumdodecylsulphate, 50 mM sodium ethylenediamine-tetraacetate, 20 mM sodium tetraborate buffer,pH 7-0, containing 1 "^ (v/v) methylglycol. Thehomogenate was incubated for 16 h at 25 °C in thebuffer with agitation. Cell wall debris was recoveredby filtration, washed extensively with hot water andsubsequently with ethanol. Finally, it was dried at50 °C and stored at - 2 0 °C until use.
The purity of cell wall fractions was tested with aDNA staining reagent, consisting of 0-2 "„ orcein in10 mM HCl containing 60% (w/v) sucrose, andexamination under a light microscope.
To obtain cell wall hydrolysates, 180 mg of cellwalls were resuspended in H^O and dialyzed over-night against H2O at 4 °C. Suspensions were incu-bated in the presence of 0-1 mg ml"' elicitor (cellulaseOnozuka R-10) for 24 h at 25 °C. The hydrolyticreaction was stopped by autoclaving. Hydrolysateswere further filtered through a 0 2/^m pore-sizefilter.
Packed cell volume, conductivity and pH
Packed cell volume was determined by centri-fugation of suspension-cultured cells at 260^ for5 min. After the removal of cell material by centri-fugation, the conductivity of the spent medium wasdetermined at 25 °C using a Crison 525 conductivitymeter (Crison Instruments, Barcelona, Spain) and a109803000 standard conductivity cell. The samefractions were used for the measurement of the pHof the spent medium.
Cell viability assay
The viability of the control and elicited cells wastested using the 2,3,5-triphenyltetrazolium chloridemethod (Zapata et al., 1991).
H.fi., formation and lipid peroxidation assays
Production of HgO^ in control and elicited cells wastested by chemiluminescence reaction with luminol(Shimada et al,, 1991). The chemiluminescence ofcontrol and elicited suspension-cultured cells wasdetermined as follows: 100 //I of suspension-culturedcells were mixed with 10/il saturated luminol inGamborg's B5 medium. The chemiluminescencewas examined under a fiuorescence microscopeequipped with an epi-illumination system (NikonDiaphot TMD) . The excitation light was obtainedby using a u.v. filter (excitor 330-380 nm, barrier420 nm, dichroic mirror 400 nm). Chemilumines-cence caused by H.3O2 was observed before and afteraddition of 40 mg ml*^ catalase.
Lipid peroxidation in the control and elicited cellswas measured using the thiobarbituric acid test formalondialdehyde (MDA) (Placer, Cushman &Johnson, 1966). A \00/i\ sample was mixed with1400//I of 20 "o trichloroacetic acid (TCA) con-taining 0 - 5 % thiobarbituric acid (TBA) in 15 mlplastic centrifuge tubes and incubated at 95 °C for30 min. The tubes were cooled immediately in an icebath and centrifuged at 10000 rpm in a Biofuge A(Heraeus, Madrid, Spain) for 5 min. Followingcentrifugation, the absorbance at 532 nm was read

Resveratrol in grape 457
using 20% TCA-+0-5 % TBA as the reference. TheMDA standard was prepared from 1,1,3,3-tetra-methoxypropane as described by Yoden & Iio (1989).
Identification and quantification of resveratrol
Phenolics were extracted by a modification of themethod of Langcake & Pryce (1976). For this,control and elicited suspension-cultured cells (25 ml)were extracted with 70 " o (v/v) methanol for 24 h at4 °C. The mixture was filtered through filter paper,and the eluate was evaporated to dryness at 40 °C inthe dark. The residue was extracted by phasepartitioning between ethyl acetate and 3 'o (w/v)NaHCOg in water. The organic phase was washedwith water and the ethyl acetate removed at 40 °C invacuo. Phenolic extracts were purified by solid-phaseextraction as described by Jeandet, Bessis &Gautheron (1991) with minor modifications asfollows: samples were dissolved in 20 % (v/v)methanol and applied to a Sep-Pak C j, column(Waters Assoc, Millipore®, Milford, MA, USA)preconditioned with 20% (v/v) methanol. Theloaded cartridge was washed with 20 °o (v/v)methanol and dried in a stream of nitrogen. Afterelution with acetone, the eluate was evaporated todryness under a nitrogen stream, and then redis-solved in methanol for HPLC analysis. For TLC,samples dissolved in acetone were used directly.
Resveratrol was identified by its R^ in severalsolvent systems during T L C analysis (Pool, Creasy& Frackelton, 1981), by its u.v. spectrum, and by theblue fluorescence of the bands when inspected underu.v. light (Aggj) (Ingham, 1976). /?,s shown byresveratrol at 25 °C were 0-37 (toluene:ethylformate: formic acid, 5 :4 :1 , silicagel), 0-82
4 : 1 , silicagel), and 0-402 1:1, cellulose), which agree with
values given by Pool et al. (1981). After elution of theblue fiuorescent spots from the TLC plates, thecompound showed A^ ,, (FtOH) = 306 nm and322 nm [sh], which agrees with the values given byJeandet et al. (1991).
Resveratrol production was quantified by HPLC(Langcake, Cornford & Pryce, 1979). Reverse-phaseHPLC was carried out on a 15 cm x 04 cm i.d.Spherisorb ODS-2 (5-0 /tm) column (Tracer anali-tica S.A., Barcelona, Spain) using a Waters (WatersAssoc, Millipore*, Milford, MA, USA) systemcomprising a Model 510 pump. Model U6K sampleinjector. Model 730 system controller and Model481 u.v. detector operated at 300 nm (0-2 a.u.f.s.).Acetonitrile-water (35:65, v/v) was used for iso-cratic chromatography at a flow rate of 025 ml min .Runs were performed at room temperature.
Identification of resveratrol in HPLC and quan-titative assessments were made by comparisons ofthe retention time and peak areas with those of anexternal standard of resveratrol purified by pre-
parative TLC (MeOH:H2O, 1:1, cellulose), whichgives a homogeneous peak during HPLC. For thisstandard, resveratrol concentration was calculatedfrom the absorbance at 306 nm using e.,,, =26800 M-^cm"^ (Langcake & Pryce, 1977). Reten-tion volume {R^) for resveratrol under theseconditions (/?,. = 5-6 ml) was close to the value(i?,, = 5-1 ml) given by Langcake et al. (1979).
Gas chromatography-mass spectrometry of phenolicfractions
Phenolics, recovered in the ethyl acetate fraction (seeabove) and purified by absorption on Sep-Pak Cjgcartridges, were analyzed by GC-MS (Greenawayet al., 1991). For this, the acetone solution wastransferred to a screw-top conical glass tube and theacetone evaporated to dryness under a stream of N2.After addition of 50//I pyridine and 100//I bis(tri-methylsilyl)trifluoroacetamide (BSTFA) containing1 % trimethylchlorosilane (TMCS), the tube wassealed and heated for 30 min at 100 °C to producetrimethylsilyl (TMS) derivatives for GC.
GC-MS was performed on a 5993 HewlettPackard model coupled to a 5995 gas chromato-graph/mass spectrometer and to a 2648A Graphicsterminal, using an OW Chrompak 50 m x 0-20 mmi.d. column. He pressure of 0-4 MPa and a GCtemperature programme of 90-280 °C at 5 °C min ,with a 5 min hold at 280 °C.
Browning determination
Browning was determined quantitatively from theabsorbance at 390 nm of 70 °o (v/v) methanolicextracts of control and elicited cells (see above), andexpressed as the equivalent concentration of p-benzoquinone, which was used as calibration stan-dard.
RESULTS
Cell responses to elicitor
Addition of the elicitor at 0-1-1-0 mg mL^ produceda strong reduction in the packed cell volume (Fig.1 a) and a net increase in the conductivity of spentmedium (Fig. 1 b) after 6 h of elicitation. Over 48 h,no significant change in the pH of the spent mediumwas observed for the elicited cells w ith respect to thatof controls (Fig. 1 c).
Elicitor concentrations in the order of, or greaterthan, 0-1 mg ml"' invariably produced cellular lysis,which was responsible for the decrease in the packedcell volume (Fig. 1 a) and probably, in part, for theincrease in the spent medium conductivity (Fig. 1 b).This was apparently due to the capacity of thecellulase to digest plant cell walls. Digestion wasprobably accompanied by some protoplast rupture.Under these circumstances, ionic fluxes through the
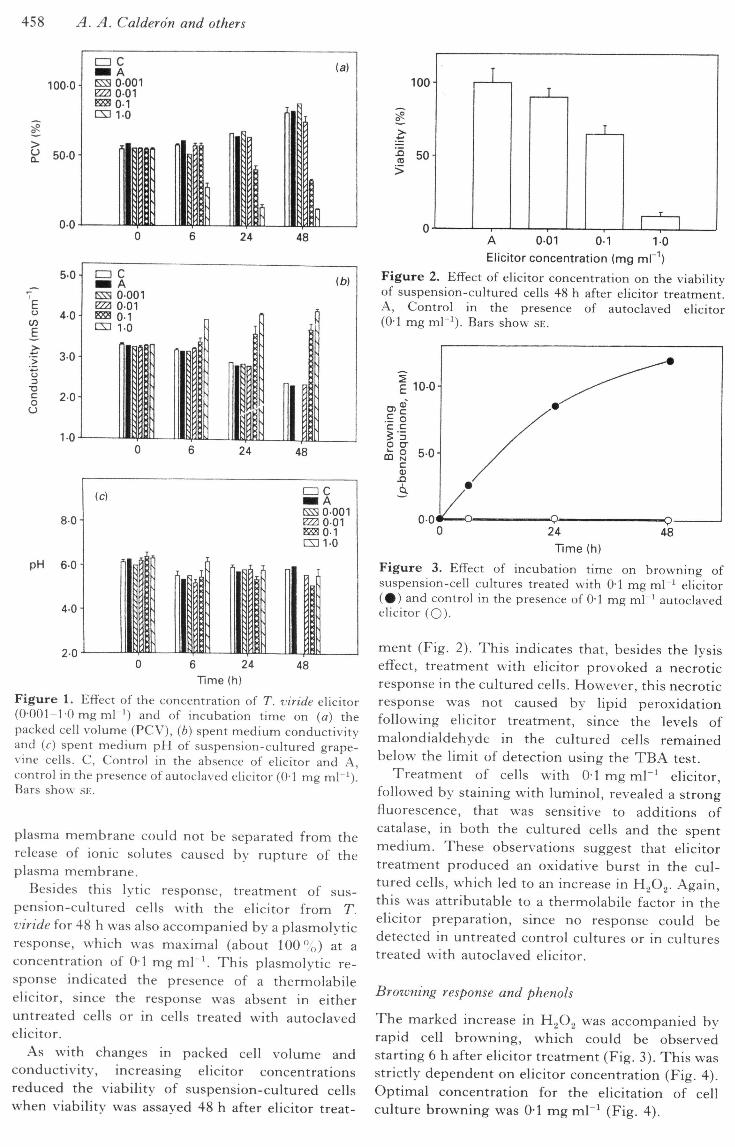
458 A. A. C alder on and others
6 24Time (h)
Figure 1. Effect of the concentration of T. viride elicitor(0-001-1-0 mg ml ') and of incubation time on {a) thepacked cell volume (PCV), (b) spent medium conductivityand (f) spent medium pH of suspension-cultured grape-vine cells. C, Control in the absence of elicitor and A,control in the presence of autoclaved elicitor (0-1 mg ml ').Bars show SE.
plasma membrane could not be separated from therelease of ionic solutes caused by rupture of theplasma membrane.
Besides this lytic response, treatment of sus-pension-cultured cells with the elicitor from T.inride for 48 h was also accompanied by a plasmolyticresponse, which was maximal (about 100%) at aconcentration of 0-1 mg ml ^ This plasmolytic re-sponse indicated the presence of a thermolabileelicitor, since the response was absent in eitheruntreated cells or in cells treated with autoclavedelicitor.
As with changes in packed cell volume andconductivity, increasing elicitor concentrationsreduced the viability of suspension-cultured cellswhen viability was assayed 48 h after elicitor treat-
100
50-j-
IT
T
A 0-01 0-1 1-0
Elicitor concentration (mg mf^)
Figure 2. Effect of elicitor concentration on the viabilityof suspension-cultured cells 48 h after elicitor treatment.A, Control in the presence of autoclaved elicitor(0-1 mg mr^). Bars show SE.
Figure 3. Effect of incubation time on browning ofsuspension-cell cultures treated with 0-1 mg ml ' elicitor(#) and control in the presence of 0-1 mg ml"^ autoclavedelicitor (O)-
ment (Fig. 2). This indicates that, besides the lysiseffect, treatment with elicitor provoked a necroticresponse in the cultured cells. However, this necroticresponse was not caused by lipid peroxidationfollowing elicitor treatment, since the levels ofmalondialdehyde in the cultured cells remainedbelow the limit of detection using the TBA test.
Treatment of cells with 01 mg ml"' elicitor,followed by staining with luminol, revealed a strongfluorescence, that was sensitive to additions ofcatalase, in both the cultured cells and the spentmedium. These observations suggest that elicitortreatment produced an oxidative burst in the cul-tured cells, which led to an increase in H^O. . Again,this was attributable to a thermolabile factor in theelicitor preparation, since no response could bedetected in untreated control cultures or in culturestreated with autoclaved elicitor.
Browning response and phenols
The marked increase in HgOg was accompanied byrapid cell browning, which could be observedstarting 6 h after elicitor treatment (Fig. 3). This wasstrictly dependent on elicitor concentration (Fig. 4).Optimal concentration for the elicitation of cellculture browning was 0-1 mg mV^ (Fig. 4).
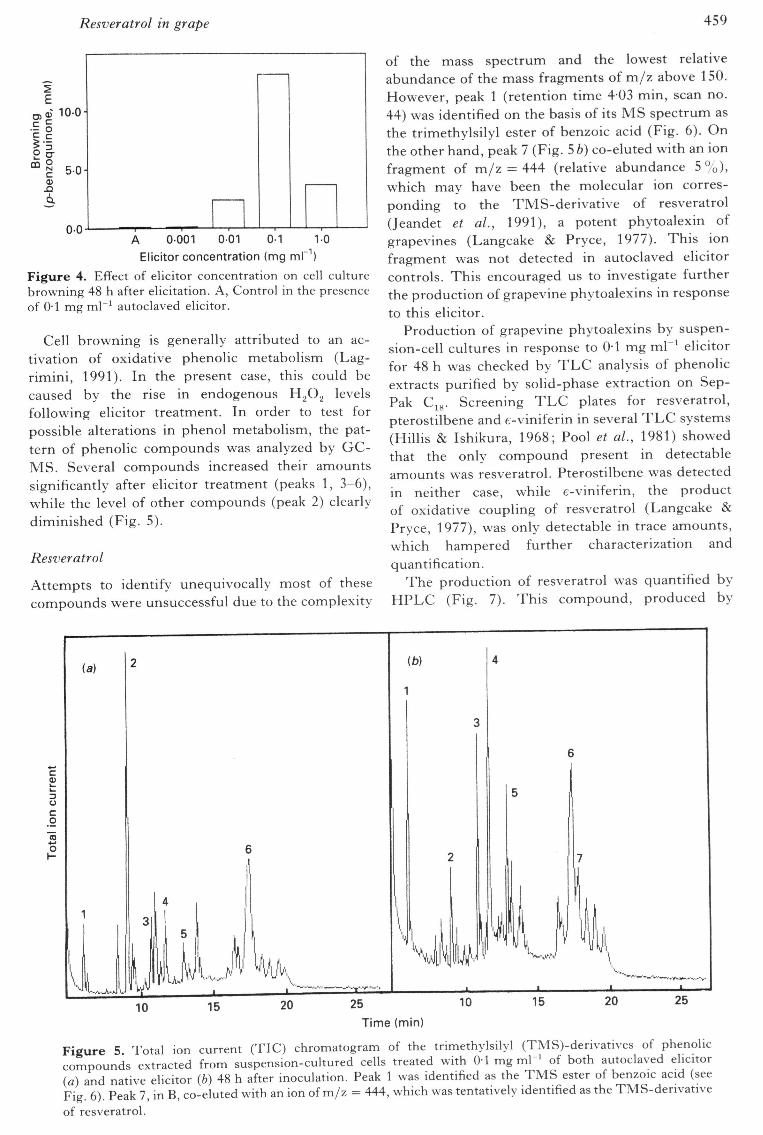
Resveratrol in grape 459
Elicitor concentration (mg m\Figure 4. Effect of elicitor concentration on cell culturebrowning 48 h after elicitation. A, Control in the presenceof 0-1 mg mr^ autoclaved elicitor.
Cell browning is generally attributed to an ac-tivation of oxidative phenolic metabolism (Lag-rimini, 1991). In the present case, this could becaused by the rise in endogenous H.2O2 levelsfollowing elicitor treatment. In order to test forpossible alterations in phenol metabolism, the pat-tern of phenolic compounds was analyzed by GC-MS. Several compounds increased their amountssignificantly after elicitor treatment (peaks 1, 3-6),while the level of other compounds (peak 2) clearlydiminished (Fig. 5).
Resveratrol
Attempts to identify unequivocally most of thesecompounds were unsuccessful due to the complexity
of the mass spectrum and the lowest relativeabundance of the mass fragments of m/z above 150.However, peak 1 (retention time 4-03 min, scan no.44) was identified on the basis of its MS spectrum asthe trimethylsilyl ester of benzoic acid (Fig. 6). Onthe other hand, peak 7 (Fig. 5 b) co-eluted with an ionfragment of m/z = 444 (relative abundance 5%),which may have been the molecular ion corres-ponding to the TMS-derivative of resveratrol(Jeandet et al, 1991), a potent phytoalexin ofgrapevines (Langcake & Pryce, 1977). This ionfragment was not detected in autoclaved elicitorcontrols. This encouraged us to investigate furtherthe production of grapevine phytoalexins in responseto this elicitor.
Production of grapevine phytoalexins by suspen-sion-cell cultures in response to 01 mg ml"^ elicitorfor 48 h was checked by T L C analysis of phenolicextracts purified by solid-phase extraction on Sep-Pak Cig. Screening T L C plates for resveratrol,pterostilbene and e-viniferin in several T L C systems(Hillis & Ishikura, 1968; Pool et al,, 1981) showedthat the only compound present in detectableamounts was resveratrol. Pterostilbene was detectedin neither case, while e-viniferin, the productof oxidative coupling of resveratrol (Langcake &Pryce, 1977), was only detectable in trace amounts,which hampered further characterization andquantification.
The production of resveratrol was quantified byHPLC (Fig. 7). This compound, produced by
Time (min)
Figure 5 Total ion current (TIC) chromatogram of the trimethylsilyl (TMS)-derivatives of phenoliccompounds extracted from suspension-cultured cells treated with 0-1 mg ml ' of both autoclaved elicitor{a) and native elicitor {b) 48 h after inoculation. Peak 1 was identified as the TMS ester of benzoic acid (seeFig. 6). Peak 7, in B, co-eluted with an ion of m/z = 444, which was tentatively identified as the TMS-derivativeof resveratrol.
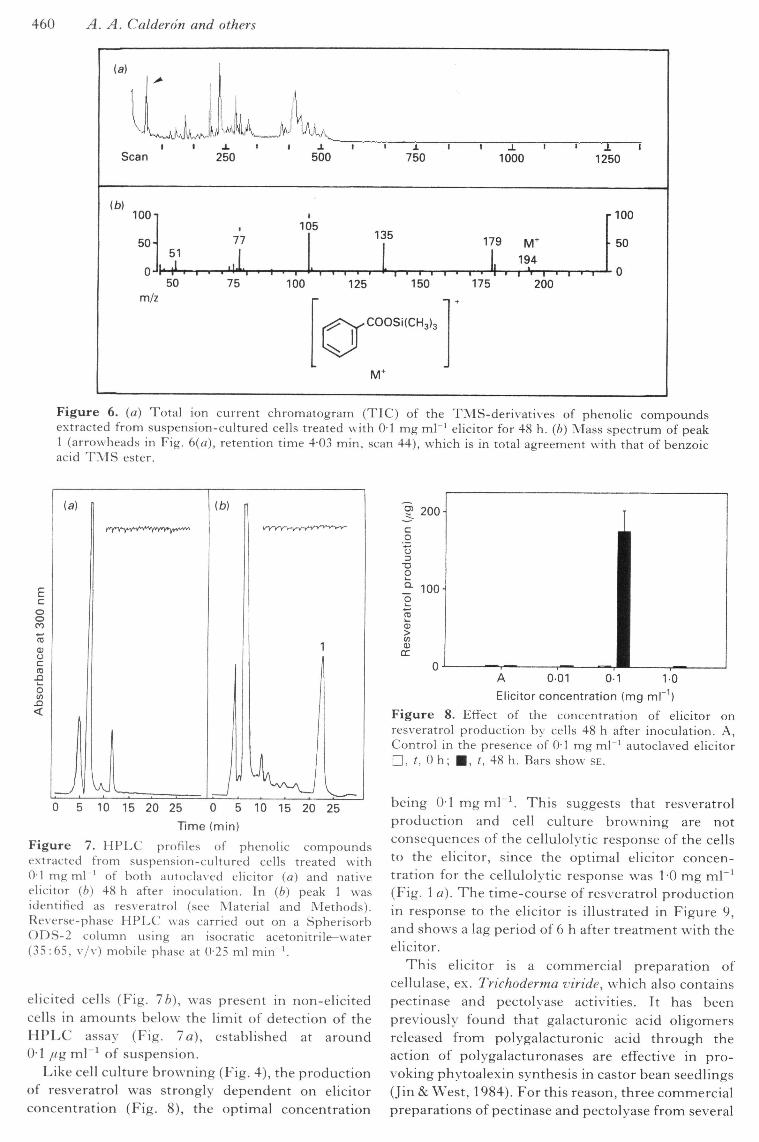
460 A. A. Calder on and others
Figure 6. (a) Total ion current chromatogram (TIC) of the TMS-derivatives of phenolic compoundse.xtracted from suspension-cultured cells treated with 01 mg ml'^ elicitor for 48 h. {b) Mass spectrum of peak1 (arrowheads in Fig. 6(a), retention time 4-03 min, scan 44), which is in total agreement with that of benzoicacid T M S ester.
EcooCO
uc.ao
(a) ? 200-
0 5 10 15 20 25 0 5 10 15 20 25Time (min)
Figure 7. HPLC profiles of phenolic compoundsextracted from suspension-cultured cells treated with0-1 mg ml ' of both autoclaved elicitor (a) and nativeelicitor (b) 48 h after inoculation. In (b) peak 1 wasidentified as resveratrol (see Material and Methods).Reverse-phase HPLC was carried out on a SpherisorbODS-2 column using an isocratic acetonitrile-water(35:65, v/v) mobile phase at 025 ml min '.
elicited cells (Fig. lb), was present in non-elicitedcells in amounts below the limit of detection of theHPLC assay (Fig. la), established at around01 fig m\'^ of suspension.
Like cell culture browning (Fig. 4), the productionof resveratrol was strongly dependent on elicitorconcentration (Fig. 8), the optimal concentration
co
f- 100-oCD
>d)
DC0
001
Elicitor concentration (mg mFigure 8. Effect of the concentration of elicitor onresveratrol production by cells 48 h after inoculation. A,Control in the presence of 0-1 mg ml™' autoclaved elicitorn, ^ 0 h; • , /, 48 h. Bars show SE.
being 0-1 mg mT^ This suggests that resveratrolproduction and cell culture browning are notconsequences of the cellulolytic response of the cellsto the elicitor, since the optimal elicitor concen-tration for the cellulolytic response was 1-0 mg mL^(Fig. 1 a). The time-course of resveratrol productionin response to the elicitor is illustrated in Figure 9,and show s a lag period of 6 h after treatment with theelicitor.
This elicitor is a commercial preparation ofcellulase, ex. Trichoderma viride, which also containspectinase and pectolyase activities. It has beenpreviously found that galacturonic acid oligomersreleased from polygalacturonic acid through theaction of polygalacturonases are effective in pro-voking phytoalexin synthesis in castor bean seedlings(Jin & West, 1984). For this reason, three commercialpreparations of pectinase and pectolyase from several
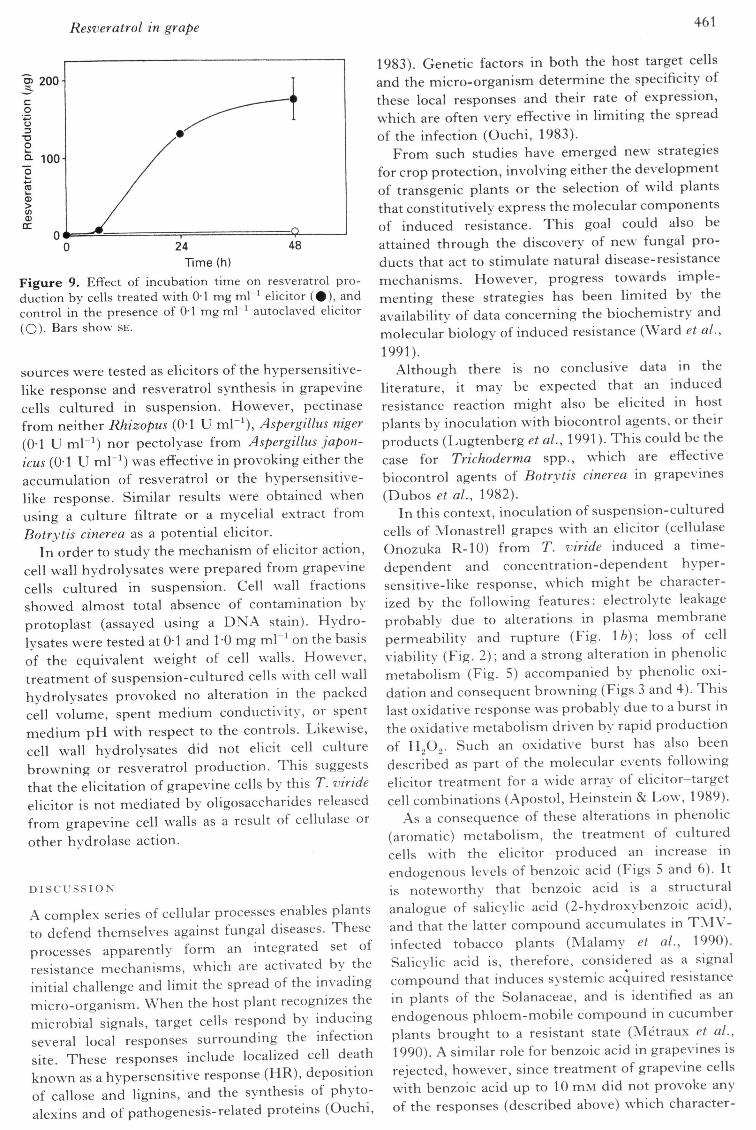
Resveratrol in grape 461
200-
couD
T3O
100-
CE0<
0 24Time (h)
Figure 9. Effect of incubation time on resveratrol pro-duction by cells treated with 0-1 mg ml" elicitor ( # ) , andcontrol in the presence of O'l mg(O)- Bars show SE.
autoclaved elicitor
sources were tested as elicitors of the hypersensitive-like response and resveratrol synthesis in grapevinecells cultured in suspension. However, pectinasefrom neither Rhizopus (0-1 U ml"^), Aspergillus niger(0-1 U mr^) nor pectolyase from Aspergillus japon-icus (0-1 U mr^) was effective in provoking either theaccumulation of resveratrol or the hypersensitive-like response. Similar results were obtained whenusing a culture filtrate or a mycelial extract fromBotrytis cinerea as a potential elicitor.
In order to study the mechanism of elicitor action,cell wall hydrolysates were prepared from grapevinecells cultured in suspension. Cell wall fractionsshowed almost total absence of contamination byprotoplast (assayed using a DNA stain). Hydro-lysates were tested at 0-1 and 10 mg ml"^ on the basisof the equivalent weight of cell walls. However,treatment of suspension-cultured cells with cell wallhydrolysates provoked no alteration in the packedcell volume, spent medium conductivity, or spentmedium pH with respect to the controls. Likewise,cell wall hydrolysates did not elicit cell culturebrowning or resveratrol production. This suggeststhat the elicitation of grapevine cells by this T. virideelicitor is not mediated by oligosaccharides releasedfrom grapevine cell walls as a result of cellulase orother hvdrolase action.
DISCUSSION
A complex series of cellular processes enables plantsto defend themselves against fungal diseases. Theseprocesses apparently form an integrated set ofresistance mechanisms, which are activated by theinitial challenge and limit the spread of the invadingmicro-organism. When the host plant recognizes themicrobial signals, target cells respond by inducingseveral local responses surrounding the infectionsite. These responses include localized cell deathknown as a hypersensitive response (HR), depositionof callose and lignins, and the synthesis of phyto-alexins and of pathogenesis-related proteins (Ouchi,
1983). Genetic factors in both the host target cellsand the micro-organism determine the specificity ofthese local responses and their rate of expression,which are often very effective in limiting the spreadof the infection (Ouchi, 1983).
From such studies have emerged new strategiesfor crop protection, involving either the developmentof transgenic plants or the selection of wild plantsthat constitutively express the molecular componentsof induced resistance. This goal could also beattained through the discovery of new fungal pro-ducts that act to stimulate natural disease-resistancemechanisms. However, progress towards imple-menting these strategies has been limited by theavailability of data concerning the biochemistry andmolecular biology of induced resistance (Ward et al.,1991).
Although there is no conclusive data in theliterature, it may be expected that an inducedresistance reaction might also be elicited in hostplants by inoculation with biocontrol agents, or theirproducts (Lugtenberg et al., 1991). This could be thecase for Trichoderma spp., which are effectivebiocontrol agents of Botrytis cinerea in grapevines(Dubos et al., 1982).
In this context, inoculation of suspension-culturedcells of Monastrell grapes with an elicitor (cellulaseOnozuka R-10) from T. viride induced a time-dependent and concentration-dependent hyper-sensitive-like response, which might be character-ized by the following features: electrolyte leakageprobably due to alterations in plasma membranepermeability and rupture (Fig. Ib); loss of cellviability (Fig. 2); and a strong alteration in phenolicmetabolism (Fig. 5) accompanied by phenolic oxi-dation and consequent browning (Figs 3 and 4). Thislast oxidative response was probably due to a burst inthe oxidative metabolism driven by rapid productionof H,O.j. Such an oxidative burst has also beendescribed as part of the molecular events followingelicitor treatment for a wide array of elicitor-targetcell combinations (Apostol, Heinstein & Low, 1989).
As a consequence of these alterations in phenolic(aromatic) metabolism, the treatment of culturedcells with the elicitor produced an increase inendogenous levels of benzoic acid (Figs 5 and 6). Itis noteworthy that benzoic acid is a structuralanalogue of salicylic acid (2-hydroxybenzoic acid),and that the latter compound accumulates in TMV-infected tobacco plants (Malamy et al., 1990).Salicylic acid is, therefore, considered as a signalcompound that induces systemic acquired resistancein plants of the Solanaceae, and is identified as anendogenous phloem-mobile compound in cucumberplants brought to a resistant state (Metraux et al.,1990). A similar role for benzoic acid in grapevines isrejected, however, since treatment of grapevine cellswith benzoic acid up to 10 mM did not provoke anyof the responses (described above) which character-

462 A. A. Calderdn and others
ize the hypersensitive response. Nevertheless, ben-zoic acid, alone or in conjunction with ascorbic acidand/or citric acid, has been described as an effectivecompound in the control of Botrytis cinerea invarious crops (Elad, 1992). This may be due to itscapacity to inhibit the polygalacturonase and poly-galacturonic acid lyase activities of several fungi (seeElad 1992) and, thus, to inhibit fungal penetration.
Grapevine cells also respond to the T. virideelicitor through a plasmolytic response. The litera-ture on this plasmolytic response of target cells tofungal elicitors is scant (Kamoen, 1984). However, ithas been shown that such a response may bemediated by the synthesis of osmotin, a proteindirectly related to osmotic regulation and adaptationto low water potentials, and which may also form apart of signal transduction during plant defenceresponses (Linthorst, 1991). To date, the significanceof this plasmolytic response in plant defence isunclear (Linthorst, 1991 ; Ye et al., 1992).
Together with these alterations in cell physiologyand metabolism, elicitation induced the synthesisand accumulation of resveratrol (Figs 7-9), a potentphytoalexin of grapevines (Langcake & Pryce, 1977).Resveratrol is an inhibitor of both the germination ofconidia and mycelial growth of B. cinerea (Hoos &Blaich, 1990). Elicitation reached a plateau 24-48 hafter inoculation (Fig. 9). Elicitation of stilbenesynthase, the key enzyme in resveratrol synthesis(Schroder & Schroder, 1990), in suspension-cul-tured cells of Vitis vinifera, reaches maximal values10-20 h after elicitation by fungal elicitors (Lis-widowati et al., 1991), which is in accordance withthe time-course of the appearance of resveratrol inthe cell culture media (Fig. 9).
Barber & Ride (1988) observed that the elicitoractivity of cellulase from T. viride in the inducedlignification of wounded wheat leaves was heat-stable. However, observations on the activity of thiselicitor in grapevine suggest that it is thermolabile,since it was destroyed by autoclaving (Figs 7-9).Similar results were found by Brooks, Watson &Freer (1986) and Threlfall & Whitehead (1988) forthe induction of capsidiol accumulation in Capsicumannum and Nicotiana tabacum in response to cellulaseex. T. viride.
The responses described above were specific, sincenone was obtained with pectolytic enzymes fromseveral sources, or with the inocula of either mycelialextracts or culture filtrates of Botrytis cinerea.Elicitation of both the HR response and phytoalexinsynthesis in plant cells in response to fungalhydrolytic enzymes is generally believed to bemediated by oligosaccharide signals produced by thepartial and controlled degradation of plant cell walls(especially from the pectin fraction) brought aboutby enzymes secreted by the microbial agent (West,1981; Darvill & Albersheim, 1984). However, noelicitation of the HR or of resveratrol synthesis in
suspension-cultured grapevine cells was observed inresponse to inoculation with hydrolysates of grape-vine cell walls treated with the cellulase from T.viride. This poses a question as regards the nature ofthe elicitor which deserves further attention. It maybe relevant that toxic glycoproteins with elicitor-likeactivity have been reported in the culture filtrates ofRhynchosporium secalis (Mazars et al., 1990) and incell walls of Puccinia graminis (Kogel et al., 1988).
ACKNOWLEDGEMENTS
This work was partly supported by a grant from theCICYT Spain (project 296/89). A. A. Calderon holds afellowship (BPP) from the MEC Spain.
REFERENCES
Adams PB. 1990. The potential of mycoparasites for biologicalcontrol of plant diseases. Annual Review of Phytopathology 28-59-72.
Apostol I, Heinstein PF, Low PS. 1989. Rapid stimulation of anoxidative burst during elicitation of cultured plant cells. PlantPhysiology 9Q: 109-116.
Barber MS, Ride JP. 1988. A quantitative assay for inducedlignification in wounded wheat leaves and its use to surveypotential elicitors of the response. Physiological and MolecularPlant Pathology Z2: 185-197.
Brooks CJW, Watson DG, Freer IM. 1986. Elicitation ofcapsidiol accumulation in suspended callus cultures of Capsicumannuum. Phytochemistry 25: 1089-1092.
Darvill AG, Albersheim P. 1984. Phytoalexins and theirelicitors. A defense against microbial infection in plants. AnnualReview of Plant Physiology 35: 243-275.
Dubos B, Jailloux F, Bulit J. 1982. Protection du vignoble contrela pourriture grise: les proprietes antagonistes du Trichodermaa r e g a r d d u Botrvtis cinerea. Les Colloques de I'INRA 1 1 -205-219.
Elad Y. 1992. The use of antioxidants (free radical scavengers) tocontrol grey mould {Botrytis cinerea) and white mould (Sclero-tina sclerotiorum) in various crops. Plant Pathology 41 •417-426.
Greenaway W, English S, May J, Whatley FR. 1991. Analysisof phenolics of bud exudate of Populus sieboldii by GC-MS.Phytochemistry 30: 3005-3008.
Hillis WE, Ishikura N. 1968. The chromatographic and spectra!properties of stilbene derivatives. Journal of Chromatography32: 323-336.
Hoos G, Blaich R. 1990. Influence of resveratrol on germinationof conidia and mycelial growth oi Botrytis cinerea and Phomopsisviticola. Journal of Phytopathology 129: 102-110.
Ingham JL. 1976. 3,5,4'-Trihydroxystilbene as a phytoalexinfrom groundnuts {Arachis hypogaea). Phytochemistry 15: 1791-1793.
Jeandet P, Bessis R, Gautheron B. 1991. The production ofresveratrol (3,5,4'-trihydroxystilbene) by grape berries indifferent developmental stages. American Journal of Enologyand Viticulture 42: 41-46.
Jin DF, West CA. 1984. Characteristics of galacturonic acidoligomers as elicitors of casbene synthetase activity in castorbean seedlings. Plant Physiology 74: 989-992.
Kamoen O. 1984. Secretions from Botrytis cinerea as elicitors ofnecro.sis and defence. Rezme de Cytologie et Biologie vegetale -Botanique 7: 241-248.
Kogel G, Beissmann B, Reisener HJ, Kogel KH. 1988. Asingle glycoprotein from Puccinia graminis f. sp. tritici cell wallselicits the hypersensitive lignification response in wheat.Physiological and Molecular Plant Pathology 33: 173-185.
Lagrimini LM. 1991. Wound-induced deposition of polyphenolsin transgenic plants overexpressing peroxidase. Plant Physiology96: 577-583.
Langcake P, Cornford CA, Pryce RJ. 1979. Identification ofpterostilbene as a phytoalexin from Vitis vinifera leaves.Phytochemistry 18: 1025-1027.

Resveratrol in grape 463
Langcake P, Pryce RJ. 1976. The production of resveratrol byVitis vinifera and other members of the Vitaceae as a responseto infection or injury. Physiological Plant Pathology 9: 77—86.
Langcake P, Pryce RJ. 1977. A new class of phytoalexins fromgrapevines. Experientia 33: 151-152.
Linthorst HJM. 1991. Pathogenesis-related proteins of plants.Critical Reviews in Plant Sciences 10: 123-150.
Liswidowati, Hohmann F, Schwer B, Kindl H. 1991.Induction of stilbene synthase by Botrytis cinerea in culturedgrapevine cells. Planta 183: 307-314.
Lugtenberg BJJ, de Weger LA, Bennett JW. 1991. Microbialstimulation of plant growth and protection from disease.Current Opinion in Biotechnology 2: 457-464.
Malamy J, Carr JP, Klessig DF, Raskin I. 1990. Salicylic acid:a likely endogenous signal in the resistance response of tobaccoto viral infection. Science 250: 1002-1004.
Mazars, C, LafHte C, Marquet PY, Rossignol M, Auriol P.1990. Elicitor-like activity of the toxic glycoprotein isolatedfrom Rhynchosporium secalis (Oud.) Davis culture filtrates.Plant Science 69: 11-17.
Metraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M,Gaudin J, Raschdorf K, Schmid E, Blum W, Inverardi B.1990. Increase in salicylic acid at the onset of the systemicacquired resistance in cucumber. Science 250: 1004-1006.
Ouchi S. 1983. Induction of resistance or susceptibility. AnnualReview of Phytopathology 21: 289-315.
Papavizas GC. 1985. Trichoderma and Gliocladium: biology,ecology, and potential for biocontrol. Annual Review ofPhytopathology 23: 23-54.
Placer ZA, Cushman LL, Johnson BC. 1966. Estimation ofproduct of lipid peroxidation (malonyl dialdehyde) in bio-chemical systems. Analytical Biochemistry 16: 359-364.
Pool RM, Creasy LL. Frackelton AS. 1981. Resveratrol and the
viniferins, their application to screening for disease resistance ingrape breeding programs. Vitis 20: 136-145.
Schroder J, Schroder G. 1990. Stilbene and chalcone synthases:related enzymes with key functions in plant-specific pathways.Zeitschrift fiir Naturforschung 45: 1-8.
Shimada M, Akagi N, Nakai Y, Goto H, Watanabe M,Watanabe H, Nakanishi M, Yoshimatsu S, Ono C. 1991.Eree radical production by the red tide alga, Chattonellaantiqua. Histochemical Journal 23: 361-365.
Threlfall DR, Whitehead IM. 1988. Co-ordinated inhibition ofsqualene synthase and induction of enzymes of sesquiterpenoidphytoalexin biosynthesis in cultures of Nicotiana tahacum.Phytochemistry 27: 2567-2580.
Ward ER, Uknes SJ. Williams SC, Dincher SS, WiederholdDL, Alexander DC, Ahl-Goy P, Metraux JP, Ryals JA.1991. Coordinate gene activity in response to agents that inducesystemic acquired resistance. The Plant Cell 3: 1085-1094.
West CA. 1981. Eungal elicitors of the phytoalexin response inhigher plants. Naturwissenschaften 68: 447-457.
Ye XS, Jarlfors U, Tuzun S, Pan SQ, Kuc J. 1992. Biochemicalchanges in cell walls and cellular responses to tobacco leavesrelated to systemic resistance to blue mold {Peronosporatabacina) induced by tobacco mosaic virus. Canadian Journal ofBotany 70: 49-57.
Yoden K, Iio T. 1989. Determination of thiobarbituric acid-reactive substances in oxidized lipids by high-performanceliquid chromatography with a postcolumn reaction system.Analytical Biochemistry 182: 116-120.
Zapata JM, Salinas C, Calderon AA, Munoz R, Ros BarceloA. 1991. Reduction of 2,3,5-triphenyltetrazolium chloride bythe KCN-insensitive, salicylhydroxamic acid-sensitive alterna-tive respiratory pathway of mitochondria from cultured grape-vine cells. Plant Cell Reports 10: 579-582.
