replication of a chimeric origin containing elements from epstein–barr virus ori p and bovine...
TRANSCRIPT

Virus Research 75 (2001) 1–11
Replication of a chimeric origin containing elements fromEpstein–Barr virus ori P and bovine papillomavirus
minimal origin
Saul Kivimae, Aire Allikas, Reet Kurg, Mart Ustav *Department of Microbiology and Virology, Institute of Molecular and Cell Biology, Tartu Uni6ersity and Estonian Biocentre,
51010 Tartu, Estonia
Received 14 July 2000; accepted 01 November 2000
Abstract
The bovine papillomavirus E2 protein is a multifunctional protein that activates viral transcription, co-operates ininitiation of viral DNA replication, and is required for long-term episomal maintenance of viral genomes. TheEBNA1 protein of Epstein–Barr virus is required for synthesis and maintenance of Epstein–Barr virus genomes.Both viral proteins act through direct interactions with their respective DNA sequences in their origins of replication.The chimeric protein E2:EBNA1, which consists of an transactivation domain of E2 and DNA binding domain ofEBNA1 supported the replication of the chimeric origin that contained EBNA1 binding sites in place of the E2binding sites principally as full-length E2 did in the case of papillomavirus minimal origin. This indicates that thechimeric protein E2:EBNA1 is competent to assemble a replication complex similar to the E2 protein. These dataconfirm the earlier observations that the only part of E2 specifically required for its activity in replication is theN-terminal activation domain and the function of the DNA binding domain of E2 in the initiation of replication isto tether the transactivation domain of E2 to the origin of replication. © 2001 Elsevier Science B.V. All rightsreserved.
Keywords: DNA replication; E2 protein; Papillomavirus; EBNA1 protein; Epstein–Barr virus
www.elsevier.com/locate/virusres
1. Introduction
The papillomaviruses are small DNA virusesthat cause benign tumors in their hosts. Followingthe initial entry of the viral genome into the cell
nucleus, the papillomavirus DNA is amplified andviral copy number is increased up to 1000 perhaploid cell genome (Howley, 1996). At the next,maintenance, stage the viral DNA replicates insynchrony with the cellular DNA. It has beenshown that during both the amplificational andmaintenance stages of the viral life cycle, the E1and E2 proteins are the only viral factors requiredfor initiation of viral DNA replication andmaintenance of the genome in the proliferating
* Corresponding author. Tel.: +372-7-375047; fax: +372-7-420286.
E-mail address: [email protected] (M. Ustav).
0168-1702/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.
PII: S 0168 -1702 (01 )00218 -0

S. Ki6imae et al. / Virus Research 75 (2001) 1–112
cells (Ustav and Stenlund, 1991; Yang et al., 1991;Piirsoo et al., 1996). The papillomavirus E1protein is the origin recognition factor and pos-sesses ATP-dependent DNA helicase functionduring the elongation of the replication fork (Seoet al., 1993b; Yang et al., 1993). The viral E2protein is the master regulator of the viral geneexpression and genome replication. First, thisprotein is the origin recognition factor, whichserves as a specificity factor enhancing binding ofthe viral helicase E1 to the origin of replication(Yang et al., 1991; Seo et al., 1993a; Ustav et al.,1993; Sedman and Stenlund, 1995; Bonne-Andreaet al., 1997). Second, E2 may alleviate nu-cleosome-mediated repression of DNA replicationby binding to the specific binding sites near thereplication origin (Li and Botchan, 1994) andcould interact with cellular replication proteinssuch as replication protein A (RPA) (Li andBotchan, 1993), therefore facilitating the assemblyof the initiation complex.
The BPV-1 E2 protein is composed of twofunctional domains: an aminoterminal transcrip-tion and replication activation domain and a car-boxy-terminal DNA binding–dimerizationdomain that binds as a dimer to the ACCN6GGTrecognition sequence(Androphy et al., 1987; Giriand Yaniv, 1988; McBride et al., 1988, 1989).These domains of E2 are separated by a hingeregion of approximately 100 residues, whichserves as an unstructured linker between the twofunctional domains. The carboxy-terminal 85amino acids (aa) of BPV-1 E2 are sufficient forsequence-specific DNA binding and dimerization.A high-resolution crystal structure for this regionin the complex with DNA revealed that the DNA-binding domain is folded into a dimeric b-barrelwith flanking recognition helixes positioned in themajor groove of the DNA binding site (Hegde etal., 1992). However, recent data demonstrate theinvolvement of the flanking region (aa 310–325)in DNA–binding function and dimerization, too(Pepinsky et al., 1997; Veeraraghavan et al.,1998). Analysis of mutations which delete variousregions of the E2 protein demonstrate that, inmost cases, an intact transactivation domain andintact DNA binding domain are required forDNA replication (for review see Ustav and Ustav,1998).
The replication mechanism of the Epstein–Barrvirus (EBV) differs substantially from the papillo-maviruses, because EBV latent origin replicatesonly once per cell cycle and is not amplified afterentry into the cell nucleus (Yates and Guan,1991). Synthesis and maintenance of EBV DNArequires a viral cis-acting sequence, oriP, and asingle viral protein, the Epstein–Barr nuclearantigen 1 (EBNA1) (Yates et al., 1985). EBNA1 isa nuclear DNA binding protein, which does notpossess enzymatic activities that could contributedirectly to synthesis of oriP DNA (Middleton andSugden, 1992). There are 24 binding sites forEBNA-1 within oriP (Rawlins et al., 1985). Thesesites are organized into two clusters, referred to asthe family of repeats (FR) and the dyad symmetryelement (DS) (Reisman et al., 1985). The crystalstructures of E2 and EBNA1 DNA-binding do-mains have been found to be almost identical,though having no sequence similarity (Grossmanand Laimins, 1996). In addition to activities men-tioned above both these viral proteins mediateviral episome anchorage on the cellular chromo-somes during mitosis, thus ensuring proper trans-mission to daughter cells (Lehman and Botchan,1998; Skiadopoulos and McBride, 1998; Ilves etal., 1999; Marechal et al., 1999).
In the present work we made a fusion proteinbetween the transactivation domain of the BPV-1E2 protein and DNA-binding-dimerization do-main of EBNA 1 of Epstein–Barr virus. Thischimeric protein supported the replication of achimeric origin containing elements from Ep-stein–Barr virus ori P and BPV-1 minimal originsimilar to the E2 protein supporting the replica-tion of papillomavirus minimal origin. This workconfirmed that the major function of the DNAbinding domain of the E2 protein in the initiationof replication is to tether the transactivation do-main of E2 to the origin of replication.
2. Materials and methods
2.1. Plasmids
Bovine papillomavirus type 1 (BPV-1) E1 ex-pression vector pCGEag, E2 expression vector

S. Ki6imae et al. / Virus Research 75 (2001) 1–11 3
pCGE2 and minimal replication origin plasmidpUCAlu have been described previously (Ustavand Stenlund, 1991).
Chimeric protein E2:EBNA1 was expressedfrom CMV promoter in pUCE2:EBNA1 vectorand contained aa 1–248 from E2 protein and aa450–641 from EBNA1 protein. For constructingthe expression vector of E2:EBNA1 XmaI–XmaIfragment (6560–8289 bp) from the plasmidpEBO-LPP (Margolskee et al., 1988) (kind gift ofDr R. Margolskee) was inserted into the XmaIsite of pUC18. EcoRI–SmaI fragment frompCGE2 was cloned into previously constructedvector, which was cleaved with EcoRI andEcl136II. Plasmid pUCE2:EBNA1+NLS wasgenerated using previously constructed vectorpCGE2+NLS. XbaI–Acc65I fragment frompCGE2+NLS containing aa 1–283 from E2 withinserted NLS (Allikas et al., 2001) was recloned inthe pUCE2:EBNA1 vector. Chimeric origin plas-mids pUCMsp15DS and pUCMsp15FR wereconstructed using vectors pUCMsp15 (Ustav etal., 1993) and previously constructed pUCOriP,where oriP containing BamHI–AccI fragmentfrom pEBO-LPP was cloned into pUC18 betweenHincII and BamHI sites. EcoRI–EcoRV frag-ment from pUCOriP was cloned into pUCMsp15plasmid between EcoRI and SmaI sites resultingin a pUCMsp15DS. Whole oriP containingEcoRI–HindIII fragment was also cloned intopUCMsp15 plasmid between EcoRI and SmaIsites resulting in pUCMsp15OriP, from whichpUCMsp15FR was achieved by deleting theEcoRI and EcoRV fragment. Plasmids pCGE1KE439, pCGE1 DA497 and pCGE1 HL507 ex-pressing BPV E1 ATPase mutants are kind gift ofDr M. Botchan and have been described earlier(MacPherson et al., 1994).
2.2. Cells and transfections
The cell lines CHO and C127 were maintainedin Ham’s F12 medium supplemented with 10%fetal bovine serum. Electroporation experimentswere carried out as described earlier, using anInvitrogen ElectroPorator at capacitance setting960 mF. Voltage settings were 230 V for CHOcells and 220 V for C127 cells. Transient replica-
tion assays were performed as described previ-ously (Ustav and Stenlund, 1991).
2.3. Western blot analysis
The expression of chimeric proteins was verifiedby Western blot analysis of CHO cells transfectedwith 1 mg of expression plasmid. Forty eighthours after transfection cells were lysed in 200 mlof Laemmli buffer, separated by sodium dodecylsulfate–10% polyacrylamide gel electrophoresis(SDS–PAGE). After transfer, monoclonal anti-E2 antibodies 3F12 and 1E2 and goat anti-mouseantibody conjugated with horseradish peroxidaseas a secondary antibody were used for detectionof proteins. The signal was detected by use of anECL detection kit (Amersham) according to themanufacturer’s recommendations.
2.4. Immunofluoresence analysis
C127 cells transfected with expression vectorsof E2 protein or chimeric proteins were fixed at−20°C in acetone/methanol (1:1). Primary poly-clonal E2 specific antibodies were diluted in 1%BSA in PBS and incubated at room temperaturefor 1 h. After washing three times with PBS coverslips with cells were incubated with goat anti-rab-bit antibodies conjugated with FITC (LabAs,Tartu, Estonia). Cells were visualized and pho-tographed using Olympus Vanox microscope.
3. Results
3.1. The E2:EBNA1 chimeric protein supports thereplication of chimeric origin
DNA replication of bovine papillomavirus re-quires the viral E1 and E2 proteins (Ustav andStenlund, 1991). These proteins bind coopera-tively to the viral origin of replication, whichcontains binding sites for both proteins, formingan E1–E2-ori complex. The function of E2 inreplication is to serve as a specificity factor en-hancing binding of the viral helicase E1 to theorigin of replication (Yang et al., 1991; Seo et al.,1993a; Ustav et al., 1993; Sedman and Stenlund,
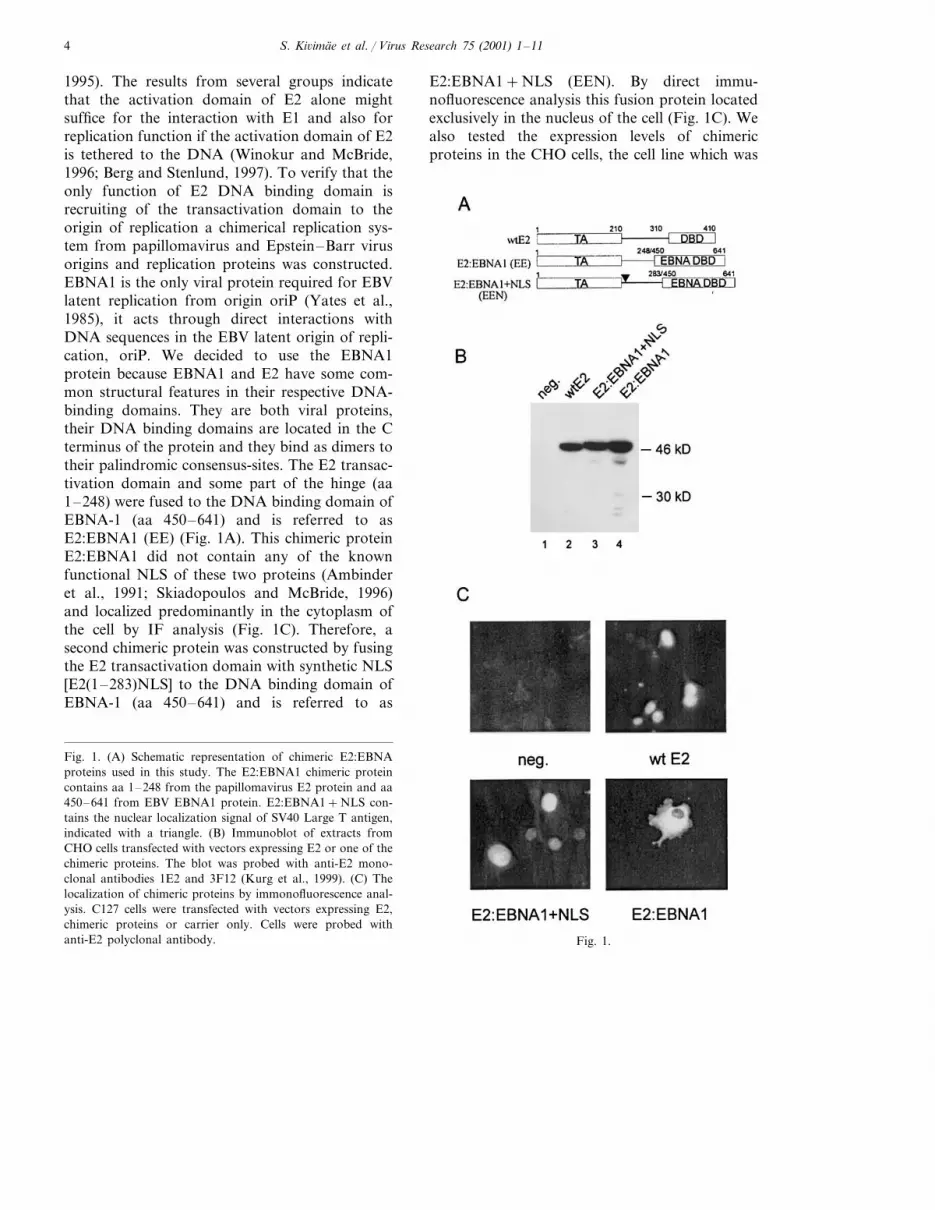
S. Ki6imae et al. / Virus Research 75 (2001) 1–114
1995). The results from several groups indicatethat the activation domain of E2 alone mightsuffice for the interaction with E1 and also forreplication function if the activation domain of E2is tethered to the DNA (Winokur and McBride,1996; Berg and Stenlund, 1997). To verify that theonly function of E2 DNA binding domain isrecruiting of the transactivation domain to theorigin of replication a chimerical replication sys-tem from papillomavirus and Epstein–Barr virusorigins and replication proteins was constructed.EBNA1 is the only viral protein required for EBVlatent replication from origin oriP (Yates et al.,1985), it acts through direct interactions withDNA sequences in the EBV latent origin of repli-cation, oriP. We decided to use the EBNA1protein because EBNA1 and E2 have some com-mon structural features in their respective DNA-binding domains. They are both viral proteins,their DNA binding domains are located in the Cterminus of the protein and they bind as dimers totheir palindromic consensus-sites. The E2 transac-tivation domain and some part of the hinge (aa1–248) were fused to the DNA binding domain ofEBNA-1 (aa 450–641) and is referred to asE2:EBNA1 (EE) (Fig. 1A). This chimeric proteinE2:EBNA1 did not contain any of the knownfunctional NLS of these two proteins (Ambinderet al., 1991; Skiadopoulos and McBride, 1996)and localized predominantly in the cytoplasm ofthe cell by IF analysis (Fig. 1C). Therefore, asecond chimeric protein was constructed by fusingthe E2 transactivation domain with synthetic NLS[E2(1–283)NLS] to the DNA binding domain ofEBNA-1 (aa 450–641) and is referred to as
E2:EBNA1+NLS (EEN). By direct immu-nofluorescence analysis this fusion protein locatedexclusively in the nucleus of the cell (Fig. 1C). Wealso tested the expression levels of chimericproteins in the CHO cells, the cell line which was
Fig. 1.
Fig. 1. (A) Schematic representation of chimeric E2:EBNAproteins used in this study. The E2:EBNA1 chimeric proteincontains aa 1–248 from the papillomavirus E2 protein and aa450–641 from EBV EBNA1 protein. E2:EBNA1+NLS con-tains the nuclear localization signal of SV40 Large T antigen,indicated with a triangle. (B) Immunoblot of extracts fromCHO cells transfected with vectors expressing E2 or one of thechimeric proteins. The blot was probed with anti-E2 mono-clonal antibodies 1E2 and 3F12 (Kurg et al., 1999). (C) Thelocalization of chimeric proteins by immonofluorescence anal-ysis. C127 cells were transfected with vectors expressing E2,chimeric proteins or carrier only. Cells were probed withanti-E2 polyclonal antibody.

S. Ki6imae et al. / Virus Research 75 (2001) 1–11 5
later used for assaying the replication supportingactivity of these proteins. As shown in Fig. 1B,chimeric proteins E2:EBNA1 and E2:EBNA1+NLS were expressed at similar levels comparedwith the wild-type E2 protein in CHO cells. Wealso constructed the chimeric Ori from E1 bindingsite and A/T rich region of papillomavirus originand EBNA1 binding sites of Epstein–Barr virusoriP (Fig. 2A). Dyad Symmetry element (DS)contains four EBNA-1 binding sites and family ofrepeats (FR) consists of 20 EBNA-1 binding sites(Rawlins et al., 1985; Reisman et al., 1985). So,two different chimeric origins were generated,where E2 binding sites in the minimal origin ofBPV-1 were replaced with DS element or FRelement, resulting in plasmids pUCMsp15DS andpUCMsp15FR, respectively (Fig. 2A). As shown
in Fig. 2B, the two fusion proteins, E2:EBNA1and E2:EBNA1+NLS were both capable of sup-porting replication of the origins containing theEBNA-1 binding sites (Fig. 2B, lanes 5, 6, 9 and10) but not of an origin containing E2 bindingsites (Fig. 2B, lanes 3 and 4). The replication ofchimeric origins was not supported by wild typeprotein (Fig. 2B, lane 7). These results demon-strate once more that tethering of the transactiva-tion domain of E2 to DNA is required for thereplicational activity and that this anchoring is themajor function of the DNA binding domain ofthe E2 protein in the initiation of the replication.Unexpectedly, we did not detect any differencebetween the activity of the E2:EBNA1 andE2:EBNA1+NLS proteins (compare lanes 5 and9 with lanes 6 and 10 in Fig. 2B). We suggest thatthe amount of E2 protein required for initiationof replication is very low and that there is stillenough E2:EBNA1 protein in the cell nucleus inspite of the data from indirect immunofluoresenceanalysis which shows mostly cytoplasmic localiza-tion. Most likely E2:EBNA1 is capable of bindingto the mitotic chromosomes when the nuclearmembrane is broken and could be used later,during the S-phase, for initiation of replication.
A transient replication assay with different con-centrations of expression vectors for E2:EBNA1and E2 was carried out to determine whether theactivation pattern of these two proteins is similar.These studies showed that similar concentrationsof E2:EBNA1 and E2 proteins were neededfor the maximum level of replication ofpUCMsp15DS or pUCAlu plasmids at constantE1 concentrations (data not shown). However,the replication signal with E2 and pUCAlu wasalways stronger than in the case of E2:EBNA1and chimeric origin. In both cases, higher-than-optimal protein concentrations resulted in a re-duction in replication, most likely caused by theself-squelching activity of E2, which is mediatedby the transactivation domain (Abroi et al., 1996;Ferguson and Botchan, 1996).
3.2. Replication modes of pUCAlu and chimericorigin pUCMsp15DS are similar
The two viruses — papillomavirus and Ep-
Fig. 2. (A) Schematic representation of chimeric origins usedin this study. (B) The ability of chimeric proteins to supportthe replication of BPV1 minimal origin and chimeric origins inthe transient replication assay. In this assay, 100 ng of pU-CAlu, pUCMsp15DS or pUCMsp15FR were co-transfectedwith 500 ng of pCGEag and 200 ng of pCGE2 or 30 ng ofpUCE2:EBNA1 (or pUCE2:EBNA1+NLS), respectively,into CHO cells. Low-molecular-weight DNA was extracted at48 and 72 h posttransfection, digested with HindIII and DpnI,and analyzed by Southern blotting using an ori probe.

S. Ki6imae et al. / Virus Research 75 (2001) 1–116
stein–Barr virus — have different modes of repli-cation. One difference between the replication oforiP and the replication of papillomavirus originis that oriP containing plasmids initiate replica-tion only once per cell cycle (Yates and Guan,1991). Papillomaviruses initiate replication manytimes per cell cycle (Gilbert and Cohen, 1987;Piirsoo et al., 1996) and as a result, a rapidamplification of their DNA follows the initialentry to the cell nucleus. The molecular mecha-nisms that regulate the mode of replication ofthese two viruses are not known. First we tried todetermine the number of replication events of thepUCMsp15DS origin in a single cell cycle. CHOcells were cotransfected with BPV origin plasmidpUCAlu or the fusion origin plasmidpUCMsp15DS and with E1 and E2 or E1 andE2:EBNA expression plasmids, respectively.Episomal DNA was collected 13, 18, 25 and 39 hafter transfection. The doubling time of CHOcells is approximately 16–18 h, so the first timepoint was harvested before the cells could gothrough one cell cycle. DNA was digested withHindIII which linearizes the pUCAlu and MspDSorigin plasmids and with DpnI (Fig. 3A, lanes1–8) or MboI (recognizes the same DNA se-
Fig. 3.
Fig. 3. (A) Chimeric origin pUCMsp15DS replicates manytimes in one cell cycle. In this assay, 100 ng of pUCAlu, 200ng of pCGE2 and 500 ng of pCGEag were transfected forBPV1 minimal origin pUCAlu replication, and 200 ng ofpUCMsp15DS, 30 ng of pUCE2:EBNA1 and 500 ng ofpCGEag were transfected for chimeric origin pUCMsp15DSreplication. Low-molecular-weight DNA was extracted at 13,18, 25 and 39 h posttransfection, digested with HindIII andDpnI, or with HindIII and MboI, and analyzed by Southernblotting using an ori probe. DNA larger in size than thereplicon marker is derived from MboI resistant HindIIIcleaved co-transfected protein expression plasmids. The arrowindicates replicated MboI fragments. (B) Replication ofchimeric origin pUCMsp15DS is dependent on E1 helicaseactivity. CHO cells were electroporated with 200 ng of reporterplasmid pUCMsp15DS, 30 ng of pUCE2:EBNA1 and 500 ngof pCGEag, expressing wild-type (Wt) or mutated E1 proteins.Cells were harvested 48 or 72 h (Ú) after electroporation.Episomal DNA was digested with DpnI and linearizing en-zyme HindIII and analyzed by Southern blotting. (C) Over-ex-pression of p53 supresses pUCMspDS replication. Southernblot analysis of transient replication of the pUCMspDS originin the presence of p53 and its N-terminal deletion mutants.Cells were harvested 48, 72 or 96 h (Ú) after electroporation.
quence as DpnI but cleaves only nonmethylatedsites) (lanes 9–16). Under our experiment condi-tions DpnI efficiently cleaves fully methylatedDNA, as well as hemimethylated DNA. There-

S. Ki6imae et al. / Virus Research 75 (2001) 1–11 7
fore, the appearance of the respective full sizeband on the Southern blot shows DNA which isfully demethylated i.e. has replicated at leasttwice. Thirteen hours after transfection the repli-cation signal of pUCAlu was readily detected(Fig. 3A, lane 1) consistent with the multiplereplications, per cell cycle, of BPV1 (Gilbert andCohen, 1987). Thirteen and 18 h after transfectionDpnI cleavage resistant pUCMsp15DS plasmidwas also detectable, but, at a very low level (Fig.3A, lanes 5 and 6). This indicates that two ormore rounds of replication have occurred. Thesedata allow us to conclude that the pUCMspDSorigin plasmid has a capacity to initiate replica-tion more than once during one cell cycle in orderto produce a population of molecules resistant toDpnI cleavage. Our data show that the DNAbinding domain of EBNA1 in the chimericprotein does not change the mode of replicationof this origin. Once per cell cycle replicationwould have required two cell cycles or approxi-mately 32–36 h after transfection to give rise toDpnI resistant DNA under these digestion condi-tions. These data allow us to conclude that thechimeric origin pUCMspDS has the capacity toreplicate more than once per cell cycle.
3.3. Replication of chimeric origin pUCMspDS isdependent on E1 helicase acti6ity
The other difference between papillomavirusand EBV is the usage of the helicase activity inreplication. Papillomaviruses encode their own he-licase, which is carried by the E1 polypeptide,while some cellular helicase is probably function-ing in the EBV oriP. To examine whether thepUCMsp15DS origin requires the viral helicaseactivity, the replication assay was performed usingmutant E1 proteins, which are defective in ATPhydrolysis but able to bind DNA (MacPherson etal., 1994). The replication of plasmid containingchimeric origin pUCMsp15DS was not detectedin the absence of E1 (Fig. 3B, lane 2) or in thepresence of E1 ATPase mutants (KE439, DA497,HL507) and E2:EBNA1 (Fig. 3B, lanes 4–6). Thepresence of wild type E1 restored pUCMsp15DSreplication (Fig. 3B, lane 3), showing the absolute
requirement for E1 as a helicase in pUCMsp15DSreplication.
3.4. O6er-expression of p53 supressespUCMspDS replication
Recently it has been shown that overexpressionof the tumor supressor protein p53 was able toblock papillomavirus DNA amplificational repli-cation while not interfering with Epstein–Barrvirus oriP once-per-cell cycle replication (Lepik etal., 1998). The mechanism of this inhibition isunknown. To find out whether the expression ofp53 has an effect on chimeric originpUCMsp15DS replication the transient replica-tion assay was carried out. As shown in Fig. 3Cthe replication of pUCMsp15DS is suppressed bywild type 53 as well as by mutant p53 proteinsthat lack the N-terminal activation domain (mu-tant DN39) or the C-terminal regulatory domain(mutant DC362) or both (mutant DN39DC362),but not suppressed by mutant p53, which is defi-cient in DNA binding (mutant Trp248). The sameresults were obtained for the pUCAlu origin by us(data not shown) and are in agreement with theresults of Lepik et al. These data indicate oncemore that the mode of replication of originpUCMsp15DS is more similar to papillomavirusminimal origin pUCAlu than to ori P, which onlyreplicates once per cell cycle.
In conclusion, our results with the E2:EBNA1fusion protein and chimeric origins demonstratedthat E2:EBNA1 fusion protein supported replica-tion of the origin pUCMsp15DS principally asdid full-length E2 in the case of origin pUCAlu.This indicates that E2:EBNA1, which consists ofan transactivation domain of E2 and DNA bind-ing domain of EBNA1, is competent to assemblea replication complex in a manner similar to theE2 protein.
4. Discussion
Our studies with chimeric origin and chimericprotein showed that the E2:EBNA1 fusionprotein supported replication of the originpUCMsp15DS, as did full-length E2 in the case of

S. Ki6imae et al. / Virus Research 75 (2001) 1–118
origin pUCAlu. This indicates that E2:EBNA1,which consists of an transactivation domain of E2and DNA binding domain of EBNA1, is com-petent to assemble a replication initiation similarto the E2 protein. The pUCMsp15DS ori replica-tion was found to depend on the viral E1 initiatorprotein and the chimeric activator E2:EBNA1,similar to the way in which pUCAlu requires E1and E2 in its replication. Titration of activatorproteins gave a similar pattern of activation andsimilar quantities of proteins were required formaximal activation of replication. However, thereplication signal with E2 and pUCAlu was al-ways stronger than in the case of E2:EBNA1 andpUCMsp15DS. There can be several reasons forthe lower replication level of the pUCMsp15DSorigin. EBNA1 is known to interact with its bind-ing sites with high stability (Milman and Hwang,1987) and occupy oriP throughout the cell cycle(Hsieh et al., 1993). Such stable interaction of theactivator with its binding sites might inhibit theformation of the pre-initiation complex on thepapillomavirus origin. The role of E2 in replica-tion is only to enhance binding of the viral heli-case E1 to the origin of replication, in thereplication pre-initiation complex the E2 proteinis displaced by additional E1 molecules. The re-sulting E1–ori complex is able to unwind DNAfrom the ori site and therefore is essential forinitiation of DNA replication (Sanders and Sten-lund, 1998; Fouts et al., 1999). Second, the originconfiguration might also influence the level of thereplication signal. In the chimeric originpUCMsp15DS, the distance between the DS andthe E1 binding site is 35bp plus the AT-richregion, so it is an origin with distal binding siteand differs from the BPV ori Alu fragment (aproximal E2 binding site). Stenlund and co-work-ers have shown that cooperative binding of the E1and E2 proteins to the BPV1 origin of DNAreplication consists of two components, in which(i) the DNA binding domains of the two proteinsinteract with each other and (ii) the E2 transacti-vation domain interacts with the helicase domainof E1 (Chen and Stenlund, 2000). In the case ofthe chimeric origin, the first component of coop-erative binding is eliminated, which can explainthe weaker replication signal.
The chimeric origin pUCMsp15DS replicatedmore than once per cell cycle, which means thatthe mode of replication has not changed by re-placing E2BS in the papillomavirus ori with DSor FR elements of oriP. This result was expected,as oriP in cis has been shown to not imposecellular regulation on runaway DNA synthesisfrom an SV40 ori present on the same plasmid(Heinzel et al., 1988). Our results are also consis-tent with the recent work by Aiyar and Sugden(1998), who showed that fusions between EBNA1of EBV and the large T-antigen of SV-40 replicatetheir cognate origins, but a fusion protein whichcontains intact large T-antigen and only the DNAbinding domain of EBNA1, replicates the SV-40ori in a manner indistinguishable from large T-antigen. So, using oriP in cis and DNA bindingdomain of EBNA1 in trans we do not change themode of replication of papillomavirus.
The results from this study show that the onlypart of E2 essential for initiation of replication isthe N-terminal activation domain, while otherdomains can be replaced. Although theE2:EBNA1 fusion proteins contained some partof the hinge region, our recent data show that noparticular amino acid sequence of the hinge re-gion is required for the ability of E2 to supportreplication (Allikas et al., 2001). The mutationaland structural analysis have shown that the acti-vation domain of E2 is highly integrated, wheredeterminants interacting with transcriptional andreplicational machinery are at least partially sepa-rated (Abroi et al., 1996; Breiding et al., 1996;Brokaw et al., 1996; Ferguson and Botchan, 1996;Kurg et al., 1999; Sakai et al., 1996). This domaininteracts with the E1 protein (Benson and How-ley, 1995; Yasugi et al., 1997) and probably withmany different cellular factors, which are used inthe process of initiation of replication or activa-tion of transcription. The interaction between thetransactivation domain of E2 and viral helicaseE1 is specific as the chimeric protein VP16-E2,which contains the activation domain from theherpes simplex virus protein VP16 fused to theDNA binding domain of E2, fails to supportreplication of papillomavirus origin (Ustav et al.,1991). However, the transactivation domain ofBPV1 E2 can be replaced with the activation

S. Ki6imae et al. / Virus Research 75 (2001) 1–11 9
domain of HPV-11 E2 as they both interact withBPV1 E1 protein and are able to support replica-tion at comparable levels (Berg and Stenlund,1997; Chiang et al., 1992). The function of theC-terminal DNA-binding–dimerization domainin replication is to transport and tether the activa-tion domain to the origin of replication. TheDNA binding domain of E2 is not specificallyrequired for replication of papillomaviruses as wecan replace this with the heterologous DNA bind-ing domain, if the respective DNA binding sitesare present within the origin. Our data showconvincingly that the EBNA1 DNA binding–dimerization domain efficiently replaced the E2DNA binding–dimerization domain. Ananalogous result was obtained recently by Bergand Stenlund (1997) using the E2–GCN4 fusionprotein and artificial origin with the distal bindingsite for GCN4. However, in the case of BPV1 wildtype ori, where the E1 and E2 binding sites areadjacent to each other, the DNA-binding–dimer-ization domain of BPV1 E2 is also required forcomplex formation with BPV1 E1 and for replica-tion (Berg and Stenlund, 1997; Chen and Sten-lund, 2000). In our study, the DNA bindingdomain of E2 was replaced by the DNA bindingdomain of EBNA1, so the interaction between theDNA binding domains of E1 and E2 was elimi-nated, in this case only the activation domain ofE2 participated in the formation of the replicationinitiation complex on the origin of replication.Therefore, the only part of E2 specifically re-quired for its activity in replication is the N-termi-nal activation domain and the main function ofthe DNA binding domain of E2 in the initiationof replication is to tether the transactivation do-main of E2 to the origin of replication.
Acknowledgements
We thank R. Margolskee for providing pEBO-LPP and Bill Sugden for providing EBV oriPplasmid; also Anne Kalling for excellent technicalassistance. This study was supported by grants4475 and 4476 from the Estonian Science Foun-dation, grant INTL55000339 from the HowardHughes Medical Institute, CIPA-CT94-015 and
CT96-0918 from the European Union and theTumor Biology grant from the CitrinaFoundation.
References
Abroi, A., Kurg, R., Ustav, M., 1996. Transcriptional andreplicational activation functions in the bovine papillo-mavirus type 1 E2 protein are encoded by different struc-tural determinants. J. Virol. 70, 6169–6179.
Aiyar, A., Sugden, B., 1998. Fusions between Epstein–Barrviral nuclear antigen-1 of Epstein–Barr virus and the largeT-antigen of simian virus 40 replicate their cognate origins.J. Biol. Chem. 273, 33073–33081.
Allikas A., O8 rd D., Kurg R., Kivi S., Ustav M., 2001. Roles ofthe hinge region and the DNA binding domain of thebovine papillomavirus type 1 E2 protein in initiation ofDNA replication. VirusResearch. Submitted.
Ambinder, R., Mullen, M., Chang, Y., Hayward, G., Hay-ward, S., 1991. Functional domains of Epstein–Barr virusnuclear antigen EBNA-1. J. Virol. 65, 1466–1478.
Androphy, E., Lowy, D., Schiller, J., 1987. Bovine papillo-mavirus E2 trans-activating gene product binds to specificsites in papillomavirus DNA. Nature 325, 70–73.
Benson, J., Howley, P., 1995. Amino-terminal domains of thebovine papillomavirus type 1 E1 and E2 proteins partici-pate in complex formation. J. Virol. 69, 4364–4372.
Berg, M., Stenlund, A., 1997. Functional interactions betweenpapillomavirus E1 and E2 proteins. J. Virol. 71, 3853–3863.
Bonne-Andrea, C., Tillier, F., McShan, G., Wilson, V., Cler-tant, P., 1997. Bovine papillomavirus type 1 DNA replica-tion: the transcriptional activator E2 acts in vitro as aspecificity factor. J. Virol. 71, 6805–6815.
Breiding, D., Grossel, M., Androphy, E., 1996. Genetic analy-sis of the bovine papillomavirus E2 transcriptional activa-tion domain. Virology 221, 34–43.
Brokaw, J., Blanco, M., McBride, A., 1996. Amino acidscritical for the functions of the bovine papillomavirus type1 E2 transactivator. J. Virol. 70, 23–29.
Chen, G., Stenlund, A., 2000. Two patches of amino acids onthe E2 DNA binding domain define the surface for interac-tion with E1, J. Virol. 74, 1506–1512.
Chiang, C., Ustav, M., Stenlund, A., Ho, T., Broker, T.,Chow, L., 1992. Viral E1 and E2 proteins support replica-tion of homologous and heterologous papillomaviralorigins. Proc. Natl. Acad. Sci. USA. 89, 5799–5803.
Ferguson, M., Botchan, M., 1996. Genetic analysis of theactivation domain of bovine papillomavirus protein E2: itsrole in transcription and replication. J. Virol. 70, 4193–4199.
Fouts, E.T., Yu, X., Egelman, E.H., Botchan, M.R., 1999.Biochemical and electron microscopic image analysis of thehexameric E1 helicase. J. Biol. Chem. 274, 4447–4458.

S. Ki6imae et al. / Virus Research 75 (2001) 1–1110
Gilbert, D., Cohen, S., 1987. Bovine papilloma virus plasmidsreplicate randomly in mouse fibroblasts throughout Sphase of the cell cycle. Cell 50, 59–68.
Giri, I., Yaniv, M., 1988. Structural and mutational analysis ofE2 trans-activating proteins of papillomaviruses revealsthree distinct functional domains. EMBO. J. 7, 2823–2829.
Grossman, S., Laimins, L., 1996. EBNA1 and E2: a newparadigm for origin-binding proteins? Trends. Microbiol.4, 87–89.
Hegde, R., Grossman, S., Laimins, L., Sigler, P., 1992. Crystalstructure at 1.7 A, of the bovine papillomavirus-1 E2DNA-binding domain bound to its DNA target. Nature359, 505–512.
Heinzel, S.S., Krysan, P.J., Calos, M.P., DuBridge, R.B.,1988. Use of simian virus 40 replication to amplify Ep-stein–Barr virus shuttle vectors in human cells. J. Virol.62, 3738–3746.
Howley, P.M., 1996. Papillomavirinae: the viruses and theirreplication. In: Fields, B.C., Knipe, D.M., Howley, P.M.(Eds.), Virology, 2nd edn. Lippincott-Raven Publishers,Philadelphia, pp. 2045–2076.
Hsieh, D., Camiolo, S., Yates, J., 1993. Constitutive bindingof EBNA1 protein to the Epstein–Barr virus replicationorigin, oriP, with distortion of DNA structure duringlatent infection. EMBO. J. 12, 4933–4944.
Ilves, I., Kivi, S., Ustav, M., 1999. Long-term episomalmaintenance of bovine papillomavirus type 1 plasmids isdetermined by attachment to host chromosomes, which Ismediated by the viral E2 protein and its binding sites. J.Virol. 73, 4404–4412.
Kurg, R., Parik, J., Juronen, E., Sedman, T., Abroi, A., Liiv,I., Langel, U., Ustav, M., 1999. Effect of bovine papillo-mavirus E2 protein-specific monoclonal antibodies on pa-pillomavirus DNA replication. J. Virol. 73, 4670–4677.
Lehman, C.W., Botchan, M.R., 1998. Segregation of viralplasmids depends on tethering to chromosomes and isregulated by phosphorylation. Proc. Natl. Acad. Sci. USA95, 4338–4343.
Lepik, D., Ilves, I., Kristjuhan, A., Maimets, T., Ustav, M.,1998. p53 protein is a suppressor of papillomavirus DNAamplificational replication. J. Virol. 72, 6822–6831.
Li, R., Botchan, M., 1993. The acidic transcriptional activa-tion domains of VP16 and p53 bind the cellular replicationprotein A and stimulate in vitro BPV-1 DNA replication.Cell 73, 1207–1221.
Li, R., Botchan, M., 1994. Acidic transcription factors allevi-ate nucleosome-mediated repression of DNA replication ofbovine papillomavirus type 1. Proc. Natl. Acad. Sci. USA91, 7051–7055.
MacPherson, P., Thorner, L., Parker, L., Botchan, M., 1994.The bovine papilloma virus E1 protein has ATPase activityessential to viral DNA replication and efficient transforma-tion in cells. Virology 204, 403–408.
Marechal, V., Dehee, A., Chikhi-Brachet, R., Piolot, T.,Coppey-Moisan, M., Nicolas, J.-C., 1999. MappingEBNA-1 domains involved in binding to metaphase chro-mosomes, J. Virol. 73, 4385–4392.
Margolskee, R.F., Kavathas, P., Berg, P., 1988. Epstein–Barrvirus shuttle vector for stable episomal replication ofcDNA expression libraries in human cells, Mol. Cell. Biol.8, 2837–2847.
McBride, A., Byrne, J., Howley, P., 1989. E2 polypeptidesencoded by bovine papillomavirus type 1 form dimersthrough the common carboxyl-terminal domain: transacti-vation is mediated by the conserved amino-terminal do-main. Proc. Natl. Acad. Sci. USA 86, 510–514.
McBride, A., Schlegel, R., Howley, P., 1988. The carboxy-ter-minal domain shared by the bovine papillomavirus E2transactivator and repressor proteins contains a specificDNA binding activity. EMBO. J. 7, 533–539.
Middleton, T., Sugden, B., 1992. EBNA1 can link the en-hancer element to the initiator element of the Epstein–Barrvirus plasmid origin of DNA replication. J. Virol. 66,489–495.
Milman, G., Hwang, E.S., 1987. Epstein–Barr virus nuclearantigen forms a complex that binds with high concentra-tion dependence to a single DNA-binding site. J. Virol. 61,465–471.
Pepinsky, R., Prakash, S., Corina, K., Grossel, M., Barsoum,J., Androphy, E., 1997. Sequences flanking the core DNA-binding domain of bovine papillomavirus type 1 E2 con-tribute to DNA-binding function. J. Virol. 71, 828–831.
Piirsoo, M., Ustav, E., Mandel, T., Stenlund, A., Ustav, M.,1996. Cis and trans requirements for stable episomalmaintenance of the BPV-1 replicator. EMBO. J. 15, 1–11.
Rawlins, D.R., Milman, G., Hayward, S.D., Hayward, G.S.,1985. Sequence-specific DNA binding of the Epstein–Barrvirus nuclear antigen (EBNA-1) to clustered sites in theplasmid maintenance region. Cell 42, 859–868.
Reisman, D., Yates, J., Sugden, B., 1985. A putative origin ofreplication of plasmids derived from Epstein–Barr virus iscomposed of two cis-acting components. Mol. Cell. Biol. 5(8), 1822–1832.
Sakai, H., Yasugi, T., Benson, J., Dowhanick, J., Howley, P.,1996. Targeted mutagenesis of the human papillomavirustype 16 E2 transactivation domain reveals separable tran-scriptional activation and DNA replication functions. J.Virol. 70, 1602–1611.
Sanders, C.M., Stenlund, A., 1998. Recruitment and loadingof the E1 initiator protein: an ATP-dependent processcatalysed by a transcription factor. Embo. J. 17 (23),7044–7055.
Sedman, J., Stenlund, A., 1995. Co-operative interaction be-tween the initiator E1 and the transcriptional activator E2is required for replicator specific DNA replication ofbovine papillomavirus in vivo and in vitro. EMBO. J. 14,6218–6228.
Seo, Y., Muller, F., Lusky, M., Gibbs, E., Kim, H., Phillips,B., Hurwitz, J., 1993a. Bovine papilloma virus (BPV)-en-coded E2 protein enhances binding of E1 protein to theBPV replication origin. Proc. Natl. Acad. Sci. USA 90,2865–2869.
Seo, Y., Muller, F., Lusky, M., Hurwitz, J., 1993b. Bovinepapilloma virus (BPV)-encoded E1 protein contains multi-

S. Ki6imae et al. / Virus Research 75 (2001) 1–11 11
ple activities required for BPV DNA replication. Proc.Natl. Acad. Sci. USA 90, 702–706.
Skiadopoulos, M., McBride, A., 1996. The bovine papillo-mavirus type 1 E2 transactivator and repressor proteinsuse different nuclear localization signals. J. Virol. 70,1117–1124.
Skiadopoulos, M., McBride, A., 1998. Bovine papillomavirustype 1 genomes and the E2 transactivator protein areclosely associated with mitotic chromatin. J. Virol. 72,2079–2088.
Ustav, M., Ustav, E., Szymanski, P., Stenlund, A., 1991.Identification of the origin of replication of bovine papillo-mavirus and characterization of the viral origin recognitionfactor E1. EMBO. J. 10, 4321–4329.
Ustav, E., Ustav, M., 1998. E2 protein as the master regulatorof extrachromosomal replication of the papillomaviruses.Papillomavirus Report 9, 145–152.
Ustav, E., Ustav, M., Szymanski, P., Stenlund, A., 1993. Thebovine papillomavirus origin of replication requires a bind-ing site for the E2 transcriptional activator. Proc. Natl.Acad. Sci. USA 90, 898–902.
Ustav, M., Stenlund, A., 1991. Transient replication of BPV-1requires two viral polypeptides encoded by the E1 and E2open reading frames. EMBO. J. 10, 449–457.
Veeraraghavan, S., Mello, C.C., Lee, K.M., Androphy, E.J.,Baleja, J.D., 1998. 1H, 15N, and 13C NMR resonanceassignments for the DNA-binding domain of the BPV-1 E2protein. J. Biomol. NMR 11, 457–458.
Winokur, P., McBride, A., 1996. The transactivation andDNA binding domains of the BPV-1 E2 protein havedifferent roles in cooperative origin binding with the E1protein. Virology 221, 44–53.
Yang, L., Li, R., Mohr, I., Clark, R., Botchan, M., 1991.Activation of BPV-1 replication in vitro by the transcrip-tion factor E2. Nature 353, 628–632.
Yang, L., Mohr, I., Fouts, E., Lim, D., Nohaile, M., Botchan,M., 1993. The E1 protein of bovine papilloma virus 1 is anATP-dependent DNA helicase. Proc. Natl. Acad. Sci. USA90, 5086–5090.
Yasugi, T., Benson, J., Sakai, H., Vidal, M., Howley, P., 1997.Mapping and characterization of the interaction domainsof human papillomavirus type 16 E1 and E2 proteins. J.Virol. 71, 891–899.
Yates, J., Guan, N., 1991. Epstein–Barr virus-derived plas-mids replicate only once per cell cycle and are not am-plified after entry into cells. J. Virol. 65, 483–488.
Yates, J.L., Warren, N., Sugden, B., 1985. Stable replicationof plasmids derived from Epstein–Barr virus in variousmammalian cells. Nature 313, 812–815.
.