proteolytic systems of lactic acid bacteria
TRANSCRIPT

Appl Microbiol Biotechnol (2006) 71: 394–406DOI 10.1007/s00253-006-0427-1
MINI-REVIEW
Kirsi Savijoki . Hanne Ingmer . Pekka Varmanen
Proteolytic systems of lactic acid bacteria
Received: 30 November 2005 / Revised: 13 March 2006 / Accepted: 13 March 2006 / Published online: 21 April 2006# Springer-Verlag 2006
Abstract Lactic acid bacteria (LAB) have a very longhistory of use in the manufacturing processes of fermentedfoods and a great deal of effort was made to investigate andmanipulate the role of LAB in these processes. Today, thediverse group of LAB includes species that are amongthe best-studied microorganisms and proteolysis is oneof the particular physiological traits of LAB of whichdetailed knowledge was obtained. The proteolyticsystem involved in casein utilization provides cellswith essential amino acids during growth in milk and isalso of industrial importance due to its contribution tothe development of the organoleptic properties offermented milk products. For the most extensivelystudied LAB, Lactococcus lactis, a model for caseinproteolysis, transport, peptidolysis, and regulation there-of is now established. In addition to nutrient processing,cellular proteolysis plays a critical role in polypeptidequality control and in many regulatory circuits bykeeping basal levels of regulatory proteins low andremoving them when they are no longer needed. As partof the industrial processes, LAB are challenged byvarious stress conditions that are likely to affect meta-bolic activities, including proteolysis. While environ-mental stress responses of LAB have received increasinginterest in recent years, our current knowledge on stress-related proteolysis in LAB is almost exclusively based
on studies on L. lactis. This review provides the currentstatus in the research of proteolytic systems of LAB withindustrial relevance.
Introduction
Lactic acid bacteria (LAB), including members of the generaLactobacillus, Lactococcus, Leuconostoc, Pediococcus, andStreptococcus, are defined as gram-positive, nonsporulating,catalase-negative, and facultative anaerobic bacteria with afermentative metabolism (Axelsson 1998). LAB were foundto have applications in manufacturing various fermentedfoods, beverages, and feed products (Leroy and Devuyst2004). In addition, certain LAB strains, most notably thestrains from the genera Lactobacillus, are increasinglymarketed as health-promoting, i.e., probiotic bacteria(Saxelin et al. 2005), while certain Lactobacillus strainsare believed to produce bioactive health-beneficial peptidesfrom milk proteins (Meisel and Bockelman 1999; Korhonenand Pihlanto 2003). Another promising avenue for LAB is touse them as delivery vehicles for molecules with therapeuticvalue (Nouaille et al. 2003). Lactococcus lactis is the mostextensively studied LAB organism and the second moststudied gram-positive bacterium with respect to its genetics,physiology, and molecular biology.
Undoubtedly, the most important application of LAB istheir use as starter strains in the manufacture of variousfermented dairy products. In particular, Streptococcusthermophilus, L. lactis, Lactobacillus helveticus, andLactobacillus delbrueckii subsp. bulgaricus (hereafter L.bulgaricus) are widely used dairy starters and are of majoreconomic importance. In the milk fermentation processes,the proteolytic system of LAB plays the key role because itenables these bacteria to grow in milk, thereby ensuringsuccessful fermentation. LAB are fastidious microorgan-isms that require an exogenous source of amino acids orpeptides, which are provided by the proteolysis of casein,the most abundant protein in milk and the main source ofamino acids. In general, the exploitation of casein by LABis initiated by a cell-envelope proteinase (CEP) that
K. Savijoki . P. VarmanenDepartment of Basic Veterinary Sciences,Faculty of Veterinary Medicine,University of Helsinki,P.O. Box 66, Helsinki 00014, Finland
K. Savijoki (*)Institute of Biotechnology, University of Helsinki,P.O. Box 56, Helsinki 00014, Finlande-mail: [email protected].: +358-9-19159414
H. IngmerDepartment of Veterinary Pathobiology,The Royal Veterinary and Agricultural University,Stigbøjlen 4,Frederiksberg C 1870, Denmark

degrades the protein into oligopeptides that are subse-quently taken up by the cells via specific peptide transportsystems for further degradation into shorter peptides andamino acids by a concerted action of various intracellularpeptidases (Kunji et al. 1996; Christensen et al. 1999).While many LAB strains contain CEP, several of thesestrains, i.e., nonstarter LAB, do not, thus, they rely onstarter LAB for the production of peptides and amino acids.These pathways are also of industrial importance because inaddition to allowing growth, peptides, amino acids, andtheir derivatives are also known to contribute to theformation of texture and flavor of the fermented milkproducts. Therefore, a number of studies were undertakento unravel the pathways underlying this industriallyrelevant trait and excellent reviews have covered theseaspects of the most extensively utilized dairy starters (Kokand de Vos 1994; Poolman et al. 1995; Kunji et al. 1996;Mierau et al. 1997; Christensen et al. 1999; Doeven et al.2005). The proteolytic system of L. lactis was investigatedto the extent that a complete model for casein breakdown,transport, degradation of casein-derived peptides, andregulation thereof was established (Tynkkynen et al.1993; Hagting et al. 1994; Foucaud et al. 1995; Detmerset al. 1998; Guédon et al. 2001a,b; den Hengst et al. 2005a,b). In addition to this “casein exploitation machinery,”LAB, like any other cell, encode stress-inducible proteo-lytic enzymes that are active on casein in vitro, but are invivo known to be responsible for the processing ofabnormal proteins created by, e.g., heat shock (Frees andIngmer 1999).
To date, the published genome sequences of starter LABinclude L. lactis (Bolotin et al. 2001), S. thermophilus(Bolotin et al. 2004), and Lactobacillus sakei (Chaillou etal. 2005). Three other genome sequences were publishedfor probiotic LAB, Lactobacillus plantarum (Kleerebezemet al. 2003), Lactobacillus acidophilus (Altermann et al.2005), and Lactobacillus johnsonii (Pridmore et al. 2004).Furthermore, several other whole genome sequences ofLAB were recently completed and are expected to bepublished in the near future (Klaenhammer et al. 2005).Comparative genomics reveals some differences betweenthe proteolytic systems of LAB; differences that arethought to reflect the various environmental niches thesebacteria occupy (Boekhorst et al. 2004). For example, ofthe sequenced lactobacilli, only L. sakei and L. johnsoniiare equipped with CEPs. Compared to L. lactis, lactobacilliare largely deficient in amino acid biosynthetic capacity,which is compensated by an ability to encode a largenumber of peptidases, amino acid permeases, and multipleoligopeptide transport (Opp) systems (Klaenhammer et al.2005).
This review provides an overview of the componentscomprising the proteolytic system in LAB that areassociated with food fermentations, placing special em-phasis on components of the proteolytic system that weregenetically and biochemically characterized. Recent ad-vances in our understanding of the mechanisms employedby LAB to regulate proteolysis and the results of geneticengineering studies aiming to improve strain characteristics
in terms of proteolytic activity are also included in thisreview. High throughput “omics” technologies have alsoemerged in the field of dairy research and studies providingnovel information relevant to casein utilization arediscussed.
Proteinases
The first step in casein utilization by LAB is performed byCEPs. Five different types of these enzymes were clonedand characterized from LAB, including PrtP from L. lactisand Lactobacillus paracasei, PrtH from L. helveticus, PrtRfrom Lactobacillus rhamnosus, PrtS from S. thermophilus,and PrtB from L. bulgaricus (Kok et al. 1988; Holck andNaes 1992; Gilbert et al. 1996; Pederson et al. 1999; Siezen1999; Fernandez-Espla et al. 2000; Pastar et al. 2003). Inlactococci, prtP genes can either be plasmid- or genome-encoded, whereas CEPs characterized from lactobacilli aregenome-encoded. LAB typically possess only one CEP butthe presence of two CEPs was reported in strains of L.helveticus and L. bulgaricus (Stefanitsi et al. 1995;Pederson et al. 1999).
The CEPs of dairy LAB are typically synthesized aspreproproteins of approximately 2,000 residues and arecomposed of several distinct functional domains(Fernandez-Espla et al. 2000; Siezen 1999) (Fig. 1).From the N terminus the CEPs include: the prepro-domain (PP) corresponding to a signal sequence (∼40residues) required for secretion and a prosequence(∼150 residues) that is removed by autocatalyticprocessing; the catalytic serine protease domain (PR)(∼500 residues); an insert domain (I) (∼150 residues)that possibly modulates the substrate specificity ofCEPs; the A domain (∼400 residues) of unknownfunction; the B domain (∼500 residues) possiblyinvolved in stabilizing the CEP activity/specificity;the helix domain (H) (∼200 residues) involved inpositioning the A and B domains outside the bacterialcell; and a hydrophilic W domain (∼100 residues)functioning as a cell wall spacer. While the B domain isfound in most CEPs, it is not present in the PrtS of S.thermophilus. The W domain of PrtS contains thetypical amino acid composition of the cell wall domainof gram-positive bacteria (rich with Pro-Gly and Ser-Thr); however, it has no homologies with CEPs or withany other proteins from the database (Fernandez-Esplaet al. 2000). In contrast, the H domain is present only inPrtP (210 aa), PrtS (367 aa), and PrtH (72 aa). In PrtPand PrtS, the W domain is followed by a cell wallanchor domain (AN) carrying a sorting signal typicalfor many surface proteins of gram-positive bacteria(Navarre and Schneewind 1994). The L. helveticus andL. bulgaricus CEPs lack the AN domain and are insteadthought to bind the cell wall using the W domain itself(Pederson et al. 1999; Siezen 1999). In L. helveticus,PrtH is possibly attached to the cell wall via amechanism resembling that utilized by S-layer proteins(Pederson et al. 1999; Leenhouts et al. 1999). The PrtR
395

characterized from L. rhamnosus BGT10 is markedlydifferent from the CEPs of starter LAB (Pastar et al.2003). The B domain of PrtR is somewhat smaller anddifferent in that it lacks the helix and insert domains; itsW domain is more homologous to certain cell-surfaceantigens expressed by oral and vaginal streptococci andits AN domain contains an atypical sorting signal.
In L. lactis, the prtP gene is preceded by a divergentlytranscribed gene encoding a membrane-bound lipoprotein(PrtM) that was shown to be essential for autocatalyticmaturation of PrtP (Haandrikman et al. 1989, 1991). Thesame genetic organization for the two genes was demon-strated for L. paracasei (Holck and Naes 1992), whereas inL. helveticus, L. bulgaricus, or S. thermophilus, no prtMgene was identified in the regions flanking the genesencoding CEPs (Fernandez-Espla et al. 2000; Siezen1999). It was reported that the maturation process ofCEPs of L. bulgaricus does not require a PrtM-likechaperone (Gilbert et al. 1996; Germond et al. 2003).
CEPs have a strong preference for hydrophobic caseins,the most abundant proteins in milk (Swaisgood 1982).Caseins are divided into the αs1-, αs2-, β-, and κ-caseins;each contains a large number of proline residues that bothprevents the formation of α-helices and β-sheets andpromotes the formation of random coils. Together, thesesecondary structure characteristics lead to an unstructured,open molecule susceptible to action of CEPs (Fox 1989a).Lactococcus PrtPs are divided into PI- and PIII-typeenzymes, distinguished by their substrate specificity forαS1-, β-, and κ-caseins (Kunji et al. 1996). The PI-typeprimarily degrades β-casein that is cleaved into more than100 different oligopeptides ranging from 4 to 30 aminoacid residues (Juillard et al. 1995). κ-casein is cleaved to alesser extent by the PI-type enzyme, whereas the PIII-typeis able to cleave αS1-, β−, and κ-caseins equally well(Pritchard and Coolbear 1993). PrtPs are further classifiedinto seven groups (a, b, c, d, e, f, and g) by their specificitytoward the αS1-casein fragment from positions 1 to 23(f1–23) (Kunji et al. 1996). PrtP of L. lactiswas also shownto act on autolysin (Buist et al. 1998), which is required forcell separation and autolysis during the stationary phase ofgrowth. The rate and degree of autolysis was found todepend on the specificity, location, and the amount of the
PrtP. For Lactobacillus, CEPs PI-, PIII-, the intermediatePI/PIII-type, and some novel type substrate specificitieswere reported, whereas a CEP (PrtS) exhibiting theintermediate PI/PIII-type specificity was purified from S.thermophilus (Fernandez-Espla et al. 2000).
Peptide uptake systems
The second step in casein utilization includes transporta-tion of peptides generated by CEP into the cell by theaction of the Opp system; this was recently covered indepth in another review (Doeven et al. 2005) and istherefore only briefly discussed here. The Opp proteinsbelong to a superfamily of highly conserved ATP-bindingcassette transporters that mediate the uptake of casein-derived peptides (Higgins 1992). In L. lactis MG1363,genes encoding the oligopeptide-binding protein (OppA),two integral membranes (OppB and OppC) and thenucleotide-binding (OppD and OppF) proteins exist in anoperon (Tynkkynen et al. 1993). The Opp system of L.lactis transports peptides up to at least 18 residues and thenature of these peptides significantly affects the transportkinetics involved (Detmers et al. 1998; Juillard et al. 1998).Generally, the Opp systems of other LAB are not widelyinvestigated but it was reported that in S. thermophilus(Garault et al. 2002) and L. bulgaricus (Peltoniemi et al.2002), the composition of the Opp system is similar to thatdescribed for Lactococcus. It should be noted that S.thermophilus carries three paralogs of genes encodingOppA proteins, each of which is involved in oligopeptideinternalization (Garault et al. 2002).
Other peptide transporters identified in L. lactisMG1363 and IL1403 strains include a proton motiveforce (PMF)-driven dipeptide/tripeptide DtpT and an ATP-driven Dpp (previously referred to as DtpP) system(Hagting et al. 1994; Foucaud et al. 1995). Dpp is capableof transporting di-, tri-, and tetrapeptides containingrelatively hydrophobic branched-chain amino acids(BCAAs) and displays the highest affinity for tripeptides(Foucaud et al. 1995; Sanz et al. 2003), whereas DtpT has apreference for more hydrophilic and charged di- andtripeptides (Hagting et al. 1994). L. helveticus also
CW M C
PP PR
I
I
PP PR
A B H W
A B H W
L. lactisL. paracasei PrtP
L. helveticus PrtH
I
PP PR W A B L. bulgaricus PrtB
PP PR A H S. thermophilus PrtS
PP PR A B W L. rhamnosus PrtR
I
W
AN
AN
AN
Fig. 1 Schematic representationof CEPs of different LAB strains(model according to Siezen1999). CW Cell wall, M mem-brane, C cytoplasm, PP preprodomain, PR catalytic domain, Iinsert domain, A A domain, B Bdomain, H helix domain, W cellwall spacer domain, black dotsorting signal, and AN anchordomain
396
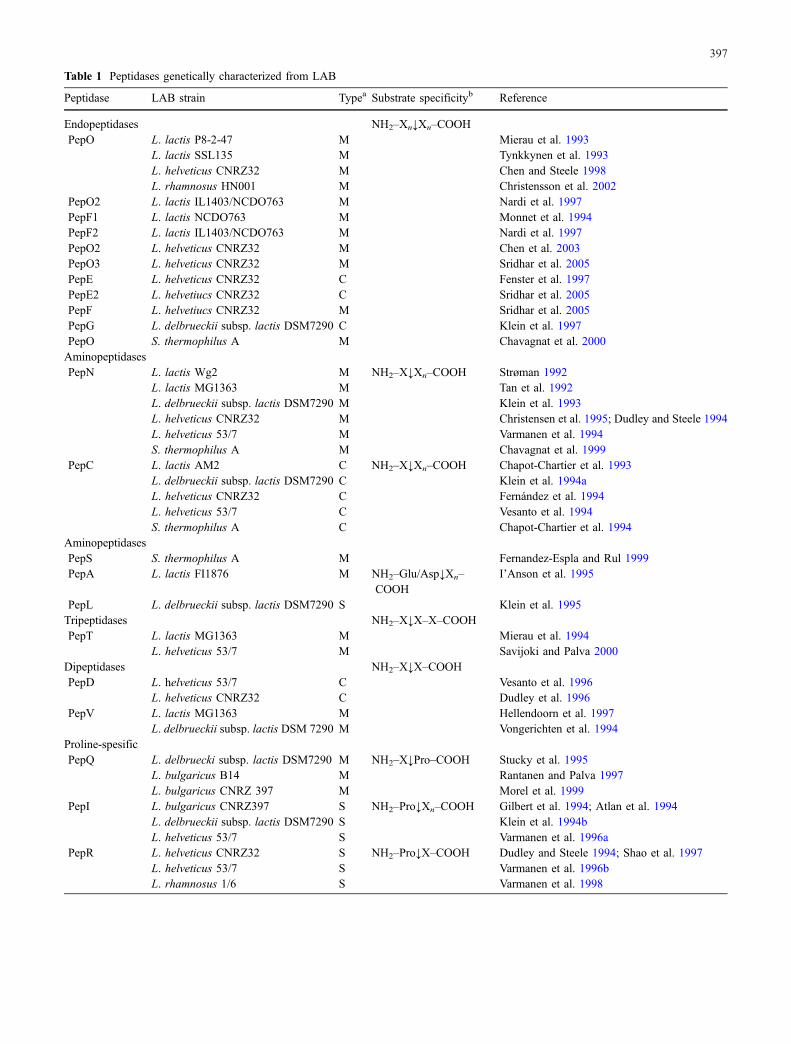
Table 1 Peptidases genetically characterized from LAB
Peptidase LAB strain Typea Substrate specificityb Reference
Endopeptidases NH2–Xn↓Xn–COOHPepO L. lactis P8-2-47 M Mierau et al. 1993
L. lactis SSL135 M Tynkkynen et al. 1993L. helveticus CNRZ32 M Chen and Steele 1998L. rhamnosus HN001 M Christensson et al. 2002
PepO2 L. lactis IL1403/NCDO763 M Nardi et al. 1997PepF1 L. lactis NCDO763 M Monnet et al. 1994PepF2 L. lactis IL1403/NCDO763 M Nardi et al. 1997PepO2 L. helveticus CNRZ32 M Chen et al. 2003PepO3 L. helveticus CNRZ32 M Sridhar et al. 2005PepE L. helveticus CNRZ32 C Fenster et al. 1997PepE2 L. helvetiucs CNRZ32 C Sridhar et al. 2005PepF L. helvetiucs CNRZ32 M Sridhar et al. 2005PepG L. delbrueckii subsp. lactis DSM7290 C Klein et al. 1997PepO S. thermophilus A M Chavagnat et al. 2000AminopeptidasesPepN L. lactis Wg2 M NH2–X↓Xn–COOH Strøman 1992
L. lactis MG1363 M Tan et al. 1992L. delbrueckii subsp. lactis DSM7290 M Klein et al. 1993L. helveticus CNRZ32 M Christensen et al. 1995; Dudley and Steele 1994L. helveticus 53/7 M Varmanen et al. 1994S. thermophilus A M Chavagnat et al. 1999
PepC L. lactis AM2 C NH2–X↓Xn–COOH Chapot-Chartier et al. 1993L. delbrueckii subsp. lactis DSM7290 C Klein et al. 1994aL. helveticus CNRZ32 C Fernández et al. 1994L. helveticus 53/7 C Vesanto et al. 1994S. thermophilus A C Chapot-Chartier et al. 1994
AminopeptidasesPepS S. thermophilus A M Fernandez-Espla and Rul 1999PepA L. lactis FI1876 M NH2–Glu/Asp↓Xn–
COOHI’Anson et al. 1995
PepL L. delbrueckii subsp. lactis DSM7290 S Klein et al. 1995Tripeptidases NH2–X↓X–X–COOHPepT L. lactis MG1363 M Mierau et al. 1994
L. helveticus 53/7 M Savijoki and Palva 2000Dipeptidases NH2–X↓X–COOHPepD L. helveticus 53/7 C Vesanto et al. 1996
L. helveticus CNRZ32 C Dudley et al. 1996PepV L. lactis MG1363 M Hellendoorn et al. 1997
L. delbrueckii subsp. lactis DSM 7290 M Vongerichten et al. 1994Proline-spesificPepQ L. delbruecki subsp. lactis DSM7290 M NH2–X↓Pro–COOH Stucky et al. 1995
L. bulgaricus B14 M Rantanen and Palva 1997L. bulgaricus CNRZ 397 M Morel et al. 1999
PepI L. bulgaricus CNRZ397 S NH2–Pro↓Xn–COOH Gilbert et al. 1994; Atlan et al. 1994L. delbrueckii subsp. lactis DSM7290 S Klein et al. 1994bL. helveticus 53/7 S Varmanen et al. 1996a
PepR L. helveticus CNRZ32 S NH2–Pro↓X–COOH Dudley and Steele 1994; Shao et al. 1997L. helveticus 53/7 S Varmanen et al. 1996bL. rhamnosus 1/6 S Varmanen et al. 1998
397

possesses a gene encoding a PMF-driven DtpT whosetransport specificity resembles its lactococcal counterpart(Nakajima et al. 1997).
Peptidases
A comprehensive review of the known specificities of LABpeptidases was published by Christensen et al. (1999). A
large number of these enzymes were purified andbiochemically characterized from S. thermophilus anddifferent Lactococcus and Lactobacillus strains; in mostcases, the corresponding gene was cloned and sequenced(Table 1). To date, no enzyme with carboxypeptidaseactivity was reported for any LAB.
After the casein-derived peptides are taken up by theLAB cells, they are degraded by a concerted action ofpeptidases with differing and partly overlapping specific-
PepN
PepCPepP
PepX
PepT
PepQ
PepR
PepI
PepDPepV
PepL
Free amino acids
Prt
M Prt
P
Op
p
Dp
p
Dtp
T
Ile
Leu
Val
prtP dtpT
A
B
Casein molecules Casein-derived peptides(4-18 amino acid residues) Peptides
(2-4 amino acid residues)
opp-pepO1 pepN/C/X/DA2prtM
CodYC
CWMC
codY
PepO PepF
Fig. 2 Simplified presentation of the function and regulation of theproteolytic system of lactococci in casein breakdown (data fromKunji et al. 1996; Christensen et al. 1999; Doeven et al. 2005). aPrtP Cell-envelope proteinase, Opp oligopeptide permease, DtpTthe ion-linked transporter for di- and tripeptides, and Dpp the ABCtransporter for peptides containing 2 to 9 amino acid residues.b Intracellural peptidases. PepO and PepF are endopeptidases,
PepN/PepC/PepP general aminopeptidases, PepX X-prolyl dipepti-dyl aminopeptidase, PepT tripeptidase, PepQ prolidase, PepRprolinase, PepI proline iminopeptidase, and PepD and PepVdipeptidases D and V. c The transcriptional repressor CodY sensesthe internal pool of branched-chain amino acids (isoleucin, leucine,and valine); using these residues as cofactors CodY represses theexpression of genes comprising the proteolytic system in L. lactis
Peptidase LAB strain Typea Substrate specificityb Reference
PepX L. lactis NCDO763 S NH2–X–Pro↓Xn–COOH Nardi et al. 1991L. delbrueckii DSM7290 S Meyer-Barton et al. 1993L. helveticus CNRZ32 S Yüksel and Steele 1996L. helveticus 53/7 S Vesanto et al. 1995L. rhamnosus 1/6 S Varmanen et al. 2000aS. thermophilus ACA-DC4 S Anastasiou et al. 2002
PepP L. lactis M NH2–X↓Pro–Xn–COOH Matos et al. 1998aCatalytic class of peptidase according to sequence analysis or biochemical characterizationbThe arrow indicates the cleavage siteM Metallopeptidase, C cysteine-peptidase, and S serine peptidase
Table 1 (continued)
398

ities (Kunji et al. 1996) (Fig. 2). The intracellularendopeptidases, general aminopeptidases (PepN andPepC), and the X-prolyl dipeptidyl aminopeptidase(PepX) are the first enzymes to act on oligopeptides.Several endopeptidases were characterized from LAB andassigned a variety of names (Table 1). All are metallo-peptidases with the exception of the L. helveticus PepE,which was shown to exhibit a thiol-dependent activity(Fenster et al. 1997). A common feature of endopeptidasesis their inability to hydrolyze intact casein but has theability to hydrolyze internal peptide bonds of casein-derived peptides. For example, the αs1-casein f1–23 and/orβ-casein f193–209 are the most preferred substrates for theendopeptidases of starter LAB origin. A unique cleavagespecificity on αs1-casein f1–23 and on postproline residuesof β-casein f203–209 was recently demonstrated for aPepO from a nonstarter strain, L. rhamnosus HN001 andfor PepO2 from the starter strain L. helveticus CNRZ32(Christensson et al. 2002; Chen et al. 2003). Besidescleaving oligopeptides from 7- to 17-residue-long, PepF isalso important for protein turnover under conditions ofnitrogen starvation in L. lactis (Monnet et al. 1994; Nardi etal. 1997).
Other peptidases capable of acting on oligopeptides arethe broad-specificity metallopeptidase PepN and cysteinepeptidase PepC proteins that were characterized fromdiverse LAB strains (Table 1). Collectively, these enzymescan remove the N-terminal amino acids from a peptide, thespecificity depending on the peptide length and the natureof the N-terminal amino acid residue (Kunji et al. 1996;Christensen et al. 1999). Di/tripeptides generated byendopeptidases, general aminopeptidases, and PepX arenext subjected to additional cleavage by the tripeptidase,PepT, and dipeptidases, PepV and PepD (Table 1). Theseenzymes prefer peptides containing hydrophobic aminoacids including leucine, methionine, phenylalanine, orglycine. An enzyme possessing specificity toward di/tripeptides with N-terminal leucine residues and dipeptidescontaining proline was biochemically characterized fromL. bulgaricus (Klein et al. 1995). Other peptidases withmore specific substrate specificities include: PepA, whichliberates N-terminal acidic residues from peptides that arethree- to nine-residue-long; PepP, which prefers tripeptidescarrying proline in the middle position; PepR and PepI,which act on dipeptides containing proline in thepenultimate position; PepQ, which cleaves dipeptidescarrying proline in the second position; and PepS, whichshows preference for peptides containing two to fiveresidues with Arg or aromatic amino acid residues in the N-terminal position (Kunji et al. 1996; Christensen et al.1999; Fernandez-Espla and Rul 1999) (Table 1).
Stress-related proteolytic systems of LAB
The cellular stress responses include the rapid and transientinduction of proteolytic activity to cope with theaccumulation of abnormally folded proteins caused by achange in the environmental conditions. During industrial
processes the starter bacteria are repeatedly exposed tostress conditions; in a recent report, several stress proteinsof starter LAB were identified in the ripening Emmentalcheese (Gagnaire et al. 2004). Knowing that several of thestress-inducible proteases are capable of degrading caseinin vitro (Katayama-Fujimura et al. 1987; Laskowska et al.1996), it is tempting to speculate that some of theseproteins could play roles in dairy fermentations at the stagewhere autolysis occurs. While Lon is the primary proteaseresponsible for the degradation of nonnative proteins inEscherichia coli (Gottesman 1996), homologues of Lonwere not identified in LAB. Thus, the stress-relatedproteases characterized to date include FtsH (Nilsson etal. 1994), HtrA (Smeds et al. 1998; Poquet et al. 2000), andthe Clp-protease (Frees and Ingmer 1999); it is interestingto note that all of these proteins are required for growth ofL. lactis under various stress conditions (Nilsson et al.1994; Frees and Ingmer 1999; Foucaud-Scheuneman andPoquet 2003). It is noteworthy that clpP of L. lactis, whichencodes the proteolytic subunit of Clp-protease, is centralin both the proteolysis of misfolded proteins (Frees andIngmer 1999) and in adjusting the level of key regulatoryproteins in the cell (Savijoki et al. 2003). Furthermore, asearch for mutations that suppressed the heat sensitivephenotype of a clpP mutant identified the regulatoryprotein, TrmA, whose inactivation stimulated the proteol-ysis of nonnative proteins in L. lactis (Frees et al. 2001).
Regulation of the proteolytic systems
LAB are likely to respond to changes in nitrogenavailability by regulating the activity of the proteolyticsystem to ensure proper nitrogen balance in the cell. It wassuggested that di/tripeptides with hydrophobic residues actas effector molecules in the transcriptional regulation of theOpp system and thereby impact the whole proteolyticsystem in L. lactis (Kunji et al. 1995, 1996; Detmers et al.1998). In a more recent study, the expression of sixtranscriptional units, including prtP, prtM, opp-pepO1,pepD, pepN, pepC, and pepX, were shown to be repressed 5to 150-fold upon the addition of casein hydrolysatecontaining 80% peptides and 20% amino acids to thegrowth medium and the expression was relieved only whencells encountered nitrogen-limiting conditions (Guédon etal. 2001a). Similarly, a proteomics approach revealed thatOpp, PepO1, PepN, PepC, PepF, and the putative substrate-binding protein (OptS) of a second oligopeptide ABCtransporter system are upregulated in L. lactis grown in amedium lacking free amino acids and peptides (Gitton et al.2005). It is now clear that the transcriptional regulatorCodY negatively regulates the expression of severalcomponents of the proteolytic system in lactis and thatthe strength of repression is modulated by the intracellularpool of BCAAs including isoleucine, leucine, and valine(Guedon et al. 2001b; Petranovic et al. 2004) (Fig. 2). Invitro assays indicate that CodY binds to the upstreamsequences preceding the opp-operon and that the BCAAsstimulate this binding (den Hengst et al. 2005a). The
399

conserved target sequence of CodY, the CodY-box(AATTTTCWGAAAATT), was recently discovered byden Hengst et al. (2005b) by combining a whole genomeexpression profiling and bioinformatic approaches withclassical DNA–protein interaction studies (den Hengst etal. 2005b). This study further demonstrated that CodYregulates its own synthesis and requires isoleucine, leucine,or valine to bind to the CodY box present at its promoter inL. lactis. In summary, in nitrogen-rich medium, theproteolytic system is repressed by CodYand the expressionis relieved when cells encounter limiting amounts ofBCAAs (Guedon et al. 2001b; den Hengst et al. 2005b)(Fig. 2).
The expression of pepF of L. lactis was suggested to beindependent of CodY (Gitton et al. 2005); it also appearsthat the regulation mechanisms of PepO1 and PepCexpression are only partly solved because the expressionof both of these enzymes is inducible by aeration andrespiratory permissive conditions also in the ΔcodYbackground (Vido et al. 2004). The carbon source in thegrowth medium was shown to affect the expression ofpepP in L. lactis (Guèdon et al. 2001a). This gene ispreceded by several potential catabolite-responsive ele-ment boxes, suggesting direct control by a CcpA-likeregulator. While the heat shock response was proposed tocontribute to the regulation of expression of somepeptidases in E. coli (Gottesman 1996), in L. lactis, heatstress had no effect on the expression of the peptidolyticsystem (Guèdon et al. 2001a). Of the stress-inducibleproteolytic enzymes, the expression of the proteolyticsubunit of Clp protease, clpP, and that of the regulatorysubunits clpC, clpE, and clpB are regulated by the stressresponse regulator CtsR (Varmanen et al. 2000b) that bindsto a directly repeated heptad located at upstream regions oftarget genes (Derre et al. 1999). Furthermore, inactivationof a gene named trmA partially complements a clpPmutation by stimulating Clp-independent degradation ofnonnative proteins, indicating that the gene product oftrmA is another negative regulator of proteolysis in L. lactis(Frees et al. 2001). Thus, at least three regulators, CodY,CtsR, and TrmA, are involved in regulating the proteolyticactivity in L. lactis.
The regulatory mechanisms of the proteolytic systems inlactobacilli are much less studied. The peptide concentra-tion in the growth medium was shown to control the PrtHand PrtR biosynthesis in L. helveticus and in L. rhamnosus,respectively (Hebert et al. 2000; Pastar et al. 2003). ThePrtH activity of L. helveticus CRL 1062 was shown to berepressed (11 to 32-fold) in a peptide-rich medium in whichdipeptide composed of leucine and proline was shown toplay an important role in regulation of PrtH activity (Hebertet al. 2000). The addition of peptides into the growthmedium also resulted in strong downregulation (17-fold) ofthe Opp and the DtpT expression in a sourdough starterLactobacillus sanfranciscensis DSM 20451, whereas theexpression of pepT was reduced to a lesser extent (two tofivefold) under the same conditions (Vermeulen et al.2005). Environmental factors are also known to affect theexpression of certain peptidase genes as was recently
demonstrated for a meat starter L. sakei where the pepRexpression was repressed during aerobic growth andinduced 20-fold upon anaerobic conditions (Champomier-Vergès et al. 2002). A recent systematic study on probioticL. acidophilus using expression profiling with wholegenome microarrays revealed that a two-component reg-ulatory system (2CRS) acts as a pleiotropic regulator incontrolling the expression of at least 80 genes, includingthose of the major components of the proteolytic system(Azcarate-Peril et al. 2005). However, the direct regulatoryrole of 2CRS was not apparent because no simple correla-tion of changes in gene expression level to proteolyticactivity or the loss thereof could be indicated (Azcarate-Perilet al. 2005). To date, only one regulatory protein directlycontrolling the expression of genes belonging to theproteolytic system was identified (Stucky et al. 1996;Schick et al. 1999). These in vitro studies have demonstratedthat PepR1 of L. delbrueckii subsp. lactis binds to thepromoter regions of the pepQ, pepX, and brnQ genes, whichencode a prolidase, an X-prolyl dipeptidyl aminopeptidase,and a BCAA transporter, respectively. More detailed studyon PepQ in L. bulgaricus has indicated that the pepR1 geneencodes a CcpA-like regulator that acts as a transcriptionalactivator of pepQ expression. The biosynthesis of PepR1 isautoregulated and the regulation depends on the glucoseconcentration in the culture medium (Morel et al. 1999,2001). The expression of PepQ was also shown to dependon the carbohydrate composition rather than peptideconcentration; this contrasts with the biosynthesis of othercomponents of the proteolytic system in L. bulgaricus(Morel et al. 1999).
Physiological role of proteolytic systems
In vitro assays were pivotal in resolving the role of theproteolytic system in amino acid liberation from casein andcasein-derived peptides. However, construction of variousmutant LAB strains lacking one or several genes hasenabled the study of the individual components of theproteolytic system and their coregulation under naturalconditions (Kunji et al. 1996, 1998; Christensen et al.1999; Varmanen et al. 2000a; Garault et al. 2002;Christensen and Steele 2003).
Mutant analyses were conducted on L. lactis, L.helveticus, L. rhamnosus, and S. thermophilus. The studieson L. lactis indicate that both the PrtP and Opp are crucialfor growth in a medium containing caseins as the solesource of amino acids; whereas the lack of the DtpT has noinfluence on the growth rate in milk (Kunji et al. 1996;Christensen et al. 1999). Also, in S. thermophilus, theinactivation of Opp results in a growth defect of thebacterium in milk (Garault et al. 2002). In conclusion, Oppis essential for nitrogen nutrition, whereas the di/tripeptidetransporters are not directly relevant to the utilization ofcasein-derived oligopeptides as no significant amounts offree amino acids, dipeptides or tripeptides, are formed byCEPs (Juillard et al. 1995). Instead, the di/tripeptidetransporters are suggested to play a role in the regulation
400

of the expression of genes involved in nitrogen metabolism(Marugg et al. 1995; Guedon et al. 2001a,b). NumerousLAB strains with mutations in single peptidase genes werealso examined for their ability to grow in milk (Kunji et al.1996; Christensen et al. 1999; Varmanen et al. 2000a).Although some minor changes in growth characteristicswere observed, it can be concluded that none of theindividual peptidases are essential for growth in milkbecause their activities can be compensated by the otherpeptidases (Christensen et al. 1999). In addition to singlemutants, multiple peptidase mutants were constructed inL. lactis (Mierau et al. 1996) and inL. helveticus (Christensenand Steele 2003). The L. lactis strain lacking pepX, pepO,pepT, pepC, and pepN grows equally well as the wild-typestrain in the complex medium (M17), suggesting that thesepeptidases have no housekeeping function, for instance, inprotein turnover (Mierau et al. 1996). However, the growthdeficiency of multiple mutant strains of L. lactis andL. helveticus in milk appears to be more severe whengreater number of peptidases are inactivated (Mierau et al.1996; Christensen and Steele 2003).
The availability of whole genome sequences is pavingthe way for global analyses of gene expression in LAB invivo. Recent proteomic approaches on LAB exploit theavailable sequence data for protein identification by massspectrometry (e.g., peptide mass mapping byMALDI-TOF,i.e., matrix-assisted laser desorption/ionization time-of-flight mass spectrometry). Recently, the protein expressionprofile of L. lactis growing in milk was published by Gittonet al. (2005). Using two-dimensional electrophoresis,Gitton et al. (2005) quantitatively detected about 900protein spots and identified more than 330 distinctproteins. While 12 of the identified proteins werepeptidases (PepA, PepC, PepDB, PepF, PepM, PepN,PepP, PepQ, PepO, PepO2, PepT, and PepV), only four ofthem (PepC, PepN, PepO, and PepF) were found to beupregulated in milk (Gitton et al. 2005). Of the genes inthe proteolytic machinery, the Opp system was found tobe strongly upregulated (Gitton et al. 2005). Gagnaire etal. (2004) utilized proteomic tools to characterize theEmmental cheese ripening process. This interestingapproach provided information on the peptidases re-leased into the cheese by the starter bacteria L. helveticusand S. thermophilus during ripening. Different peptidasesalso arose from L. helveticus (PepN, PepO, PepE, andPepQ) and S. thermophilus (PepN, PepX, and PepS).Remarkably, this work suggested the participation ofspecific peptidases of L. helveticus and S. thermophilusin the degradation of casein-derived peptides during theripening process of Emmental cheese (Gagnaire et al.2004).
Technological aspects of proteolysis
Proteolysis is considered one of the most importantbiochemical processes involved in manufacturing manyfermented dairy products (Fox 1989b). In cheese manu-facture, the proteolysis of casein is thought to play a pivotal
role because amino acids resulting from proteolysis are themajor precursors of specific flavor compounds, such asvarious alcohols, aldehydes, acids, esters, and sulfurcompounds (Smit et al. 2005). Bitterness, which resultsfrom the accumulation of hydrophobic peptides, i.e.,peptides rich with proline, is a serious quality concernfacing the manufacture of Gouda and Cheddar cheeses(Smukowski et al. 2003). The specificities of CEPs play anessential role in the production of bitter peptides (Visser etal. 1986; Rodriguez et al. 1996; Broadbent et al. 2002). Inaddition, the use of LAB strains deficient in peptidaseactivity have also indicated that peptidases, includingPepN, PepX, PepO2, and PepO3, are involved in thedegradation of bitter peptides; these peptidases thereforeimpact the development of the organoleptic quality of themilk product (Meyer and Spahni 1998, Chen and Steele1998; Chen et al. 2003; Christensen and Steele 2003;Sridhar et al. 2005). It was recently proposed that food-grade strains of L. lactis expressing L. helveticus CNRZ32PepO2 and PepO3, in combination with PepN, can be usedto reduce bitterness in cheese (Sridhar et al. 2005). Foreconomic reasons, several approaches were exploited toaccelerate the ripening process. These have includedmethods such as elevation of storage temperature, additionof proteinases, the use of bacteriophage-encoded lysin orlytic bacteriophages, and the addition of selected nonstarterLAB or lactobacilli adjuncts to cheese (Fox et al. 1996;Rodríguez et al. 1996; Morgan et al. 1997; Martínez-Cuesta et al. 1998; Meijer et al. 1998; Madkor et al. 2000).Enriching the L. lactis proteolytic potential by constructinga recombinant starter strain expressing peptidases derivedfrom L. helveticus or L. delbrueckii subsp. lactis under aconstitutive or inducible promoter can also be used toaccelerate cheese proteolysis and, hence, the ripeningprocess (Wegmann et al. 1999; Guldtfeldt et al. 2001;Luoma et al. 2001; Courtin et al. 2002; Henrich et al. 2002;Joutsjoki et al. 2002; Sridhar et al. 2005). In addition, thecheese trial accomplished by Guldtfeldt et al. (2001)demonstrated that overexpression of, e.g., pepN or pepC inL. lactis subsp. cremoris NM1 background, results in apositive effect on Cheddar cheese flavor and increases thelevel of specific free amino acids in cheese. However,while it can be concluded that balanced proteolysis isimportant for flavor formation and especially in preventionof bitterness in cheese, it is the conversion of the free aminoacids, rather than proteolysis/peptidolysis, that controls therate of flavor formation from proteins (Smit et al. 2005).Thus, engineering the proteolytic system alone is hardly thekey to accelerating flavor formation in cheese.
In addition to the proteolytic systems, the autolysis ofLAB starters is considered to be another important elementof cheese manufacture because this activity permits therelease of cytoplasmic peptidases into the curd, aprerequisite for flavor formation to proceed (Crow et al.1995). In this regard, a number of studies were sought tocontrol the rate and level of lysis of Lactococcus starterstrains; these include phage- and autolysin-based mecha-nisms and leaky lactococcal starter cultures overexpressingcertain L. lactis or L. helveticus peptidases (Buist et al.
401

1997; De Ruyter et al. 1997; Guldtfeldt et al. 2001; Tuler etal. 2002; Hickey et al. 2004). The preliminary resultssuggest the potential of such approaches to accelerate theripening process and to improve the sensory properties ofcheese.
Proteolytic activation of bioactive peptides
Besides the importance of the proteolytic/peptidolyticenzymes of LAB to organoleptic properties of the finalmilk product, certain LAB strains are known to contributeto the liberation of bioactive peptides that are thought topromote health beyond the basic nutrition (Pihlanto andKorhonen 2003; Meisel 2004). In this respect, the caseinmolecules of milk are of particular interest because they areknown to harbor bioactive peptides that are latent untilreleased by proteolysis. To date, LAB ascribed with suchactivity include strains like L. helveticus CP790, L.rhamnosusGG, L. bulgaricus SS1, and L. lactis subsp.cremoris FT4 (Gobbetti et al. 2002). Several reports haveindicated that bioactive peptides, besides existing innaturally ripened cheese and other fermented products,may also be produced in vivo after the intake of milkproteins (Meisel 2004). Manufacturing of such peptides onindustrial scale for use as dietary supplements andpharmaceutical preparations is currently receiving in-creased interest (Korhonen and Pihlanto 2003; Meisel2004).
Concluding remarks
The proteolytic system of the LAB model L. lactis wasstudied to the extent that a good understanding of the keycomponents involved in casein breakdown, uptake ofoligopeptides, and subsequent peptidolytic pathways ex-ists. While the corresponding research on lactobacillifocused mainly on L. helveticus and L. delbrueckii subsp. isalso relatively advanced, there are still many unansweredquestions, particularly regarding the regulation of proteo-lytic pathways. With the current knowledge and moleculartools, it has become possible to genetically engineer starterLAB to express desired proteolytic/peptidolytic activity.The increasing number of genome sequences available fordifferent LAB species will allow the integration offunctional genomic approaches, including transcriptomics,proteomics, and metabolomics, to define the molecularnature of metabolic networks within LAB and theregulatory mechanisms underlying the physiology traitsof these bacteria. This knowledge, aided by the develop-ment of novel food-grade genetic tools, will foster thedevelopment and screening of new LAB strains, whichperform better under different fermentation processes toprovide humans with healthy food with appealing textureand flavor.
References
Altermann E, Russell WM, Azcarate-Peril MA, Barrangou R, BuckBL, McAuliffe O, Souther N, Dobson A, Duong T, Callanan M,Lick S, Hamrick A, Cano R, Klaenhammer TR (2005)Complete genome sequence of the probiotic lactic acidbacterium Lactobacillus acidophilus NCFM. Proc Natl AcadSci USA 102:3906–3912
Anastasiou R, Papadelli M, Georgalaki MD, Kalantzopoulos G,Tsakalidou E (2002) Cloning and sequencing of the geneencoding X-prolyl-dipeptidyl aminopeptidase (PepX) fromStreptococcus thermophilus strain ACA-DC 4. J Appl Microbiol93:52–59
Atlan D, Gilbert C, Blanc B, Portalier R (1994) Cloning, sequencingand characterization of the pepIP gene encoding a prolineiminopeptidase from Lactobacillus delbrueckii subsp. bulgar-icus CNRZ 397. Microbiology 140:527–535
Axelsson L (1998) Lactic acid bacteria: classification and physiol-ogy. In: Salminen S, von Wright A (eds) Lactic acid bacteria.Microbiology and functional aspects. Marcel Dekker, NewYork, pp 1–72
Azcarate-Peril MA, McAuliffe O, Altermann E, Lick S, RussellWM, Klaenhammer TR (2005) Microarray analysis of a two-component regulatory system involved in acid resistance andproteolytic activity in Lactobacillus acidophilus. Appl EnvironMicrobiol 71:5794–5804
Boekhorst J, Siezen RJ, Zwahlen MC, Vilanova D, Pridmore RD,Mercenier A, KleerebezemM, de Vos WM, Brüssow H, DesiereF (2004) The complete genomes of Lactobacillusplantarum andLactobacillus johnsonii reveal extensive differences in chromo-some organization and gene content. Microbiology 150:3601–3611
Bolotin A, Wincker P, Mauger S, Jaillon O, Malarme K,Weissenbach J, Ehrlich SD, Soroki A (2001) The completegenome sequence of the lactic acid bacterium Lactococcuslactis ssp. lactis IL1403. Genome Res 11:731–753
Bolotin A, Quinquis B, Renault P, Sorokin A, Ehrlich SD,Kulakauskas S, Lapidus A, Goltsman E, Mazur M, PuschGD, Fonstein M, Overbeek R, Kyprides N, Purnelle B, ProzziD, Ngui K, Masuy D, Hancy F, Burteau S, Boutry M, DelcourJ, Goffeau A, Hols P (2004) Complete sequence andcomparative genome analysis of the dairy bacterium Strepto-coccus thermophilus. Nat Biotechnol 22:1554–1558
Broadbent JR, Barnes M, Brennand C, Strickland M, Houck K,Johnson ME, Steele JL (2002) Contribution of Lactococcuslactis cell envelope proteinase specificity to peptide accumula-tion and bitterness in reduced-fat cheddar cheese. Appl EnvironMicrobiol 68:1778–1785
Buist G, Karsens H, Nauta A, van Sinderen D, Venema G, Kok J(1997) Autolysis of Lactococcus lactis caused by inducedoverproduction of its major autolysin, AcmA. Appl EnvironMicrobiol 63:2722–2728
Buist G, Venema G, Kok J (1998) Autolysis of Lactococcus lactis isinfluenced by proteolysis. J Bacteriol 180:5947–5953
Chaillou S, Champomier-Vergès MC, Cornet M, Crutz-Le C, DudezAM, Martin V, Beaufils S, Darbon-Rongère E, Bossy R, LouxV, Zagorec M (2005) The complete genome sequence of themeat-borne lactic acid bacterium Lactobacillus sakei 23K. NatBiotechnol 23(12):1527–1533
Champomier-Vergès MC, Marceau A, Méra T, Zagorec M (2002)The pepR gene of Lactobacillus casei is positively regulated byanaerobisis at the transcriptional level. Appl Environ Microbiol68:3873–3877
Chapot-Chartier MP, Nardi M, Chopin MC, Chopin A, Gripon JC(1993) Cloning and sequencing of pepC, a cysteine aminopep-tidase gene from Lactococcus lactis subsp. cremoris AM2.Appl Environ Microbiol 59:330–333
Chapot-Chartier MP, Rul F, Nardi M, Gripon JC (1994) Genecloning and characterization of PepC, a cysteine aminopepti-dase from Streptococcus thermophilus, with sequence similarityto the eucaryotic bleomycin hydrolase. Eur J Biochem224:497–506
402

Chavagnat F, Casey MG, Meyer J (1999) Purification, characteriza-tion, gene cloning, sequencing, and overexpression of amino-peptidase N from Streptococcus thermophilus A. Appl EnvironMicrobiol 65:3001–3007
Chavagnat F, Meyer J, Casey MG (2000) Purification, character-isation, cloning and sequencing of the gene encodingoligopeptidase PepO from Streptococcus thermophilus A.FEMS Microbiol Lett 191:79–85
Chen YS, Steele JL (1998) Genetic characterization and physiolog-ical role of endopeptidase O from Lactobacillus helveticusCNRZ32. Appl Environ Microbiol 64:3411–3415
Chen YS, Christensen JE, Strickland M, Steele JL (2003) Identi-fication and characterization of Lactobacillus helveticus PepO2,an endopeptidase with post-proline specificity. Appl EnvironMicrobiol 69:1276–1282
Christensen JE, Steele JL (2003) Impaired growth rates in milk ofLactobacillus helveticus peptidase mutants can be overcome byuse of amino acid supplements. J Bacteriol 185:3297–3306
Christensen J, Lin D, Palva A, Steele J (1995) Sequence analysis,distribution and expression of an aminopeptidase gene fromLactobacillus helveticus CNRZ32. Gene 155:89–93
Christensen JE, Dudley EG, Pederson JA, Steele JL (1999)Peptidases and amino acid catabolism in lactic acid bacteria.Antonie Van Leeuwenhoek 76:217–246
Christensson C, Bratt H, Collins LJ, Coolbear T, Holland R,Lubbers MW, O’Toole PW, Reid JR (2002) Cloning andexpression of an oligopeptidase, PepO, with novel specificityfrom Lactobacillus rhamnosus HN001 (DR20). Appl EnvironMicrobiol 68:254–262
Courtin P, Nardi M, Wegmann U, Joutsjoki V, Ogier JC, Gripon JC,Palva A, Henrich B, Monnet V (2002) Accelerating cheeseproteolysis by enriching Lactococcus lactis proteolytic systemwith lactobacilli peptidases. Int Dairy J 12:447–454
Crow VL, Coolbear T, Gopal PK, Martley FG, McKay LL, Riepe H(1995) The role of autolysis of lactic acid bacteria in theripening of cheese. Int Dairy J 5:855–875
den Hengst CD, van Hijum SA, Geurts JM, Nauta A, Kok J,Kuipers OP (2005a) The lactococcus lactis CodY regulon:identification of a conserved cis-regulatory element. J BiolChem 280:34332–34342
den Hengst CD, Curley P, Larsen R, Buist G, Nauta A, van SinderenD, Kuipers OP, Kok J (2005b) Probing direct interactionsbetween CodY and the oppD promoter of Lactococcus lactis.J Bacteriol 187:512–521
Derre I, Rapoport G, Msadek T (1999) CtsR, a novel regulator ofstress and heat shock response, controls clp and molecularchaperone gene expression in gram-positive bacteria. MolMicrobiol 31:117–131
de Ruyter PG, Kuipers OP, Meijer WC, de Vos WM (1997) Food-grade controlled lysis of Lactococcus lactis for acceleratedcheese ripening. Nat Biotechnol 15:976–979
Detmers FJ, Kunji ER, Lanfermeijer FC, Poolman B, Konings WN(1998) Kinetics and specificity of peptide uptake by theoligopeptide transport system of Lactococcus lactis. Biochemistry37:16671–16679
Doeven MK, Kok J, Poolman B (2005) Specificity and selectivitydeterminants of peptide transport in Lactococcus lactis andother microorganisms. Mol Microbiol 57:640–649
Dudley EG, Steele JL (1994) Nucleotide sequence and distributionof the pepPN gene from Lactobacillus helveticus CNRZ32.FEMS Microbiol Lett 119:41–46
Dudley EG, Husgen AC, He W, Steele JL (1996) Sequencing,distribution, and inactivation of the dipeptidase A gene(pepDA) from Lactobacillus helveticus CNRZ32. J Bacteriol178:701–704
Fenster KM, Parkin KL, Steele JL (1997) Characterization of athiol-dependent endopeptidase from Lactobacillus helveticusCNRZ32. J Bacteriol 179:2529–2533
Fernández L, Bhowmik T, Steele J (1994) Characterization of theLactobacillus helveticus CNRZ32 pepC gene. Appl EnvironMicrobiol 60:333–336
Fernandez-Espla MD, Rul F (1999) A new member of theaminopeptidase T family of thermophilic bacteria. Eur JBiochem 263:502–510
Fernandez-Espla MD, Garault P, Monnet V, Rul F (2000) Strepto-coccus thermophilus cell wall-anchored proteinase: release,purification, and biochemical and genetic characterization.Appl Environ Microbiol 66:4772–4778
Foucaud C, Kunji ER, Hagting A, Richard J, Konings WN,Desmazeaud M, Poolman B (1995) Specificity of peptidetransport systems in Lactococcus lactis: evidence for athird system which transports hydrophobic di- and tripep-tides. J Bacteriol 177:4652–4657
Foucaud-Scheunemann C, Poquet I (2003) HtrA is a key factor inthe response to specific stress conditions in Lactococcus lactis.FEMS Microbiol Lett 224:53–59
Fox PF (1989a) The milk protein system. In: Fox P (ed)Developments in dairy chemistry, vol. 4. Elsevier AppliedScience, London, pp 1–53
Fox PF (1989b) Proteolysis during cheese manufacture andripening. J Dairy Sci 72:1379–1400
Fox PF, Wallace JM, Morgan S, Lynch CM, Niland EJ, Tobin J(1996) Acceleration of cheese ripening. Antonie VanLeeuwenhoek 70:271–297
Frees D, Ingmer H (1999) ClpP participates in the degradation ofmisfolded protein in Lactococcus lactis. Mol Microbiol31:79–87
Frees D, Varmanen P, Ingmer H (2001) Inactivation of a gene that ishighly conserved in gram-positive bacteria stimulates degrada-tion of non-native proteins and concomitantly increases stresstolerance in Lactococcus lactis. Mol Microbiol 41:93–103
Gagnaire V, Piot M, Camier B, Vissers JP, Jan G, Leonil J (2004)Survey of bacterial proteins released in cheese: a proteomicapproach. Int J Food Microbiol 94:185–201
Garault P, Le Bars D, Besset C, Monnet V (2002) Threeoligopeptide-binding proteins are involved in the oligopep-tide transport of Streptococcus thermophilus. J Biol Chem277:32–39
Germond JE, Delley M, Gilbert C, Atlan D (2003) Determination ofthe domain of the Lactobacillus delbrueckii subsp. bulgaricuscell surface proteinase PrtB involved in attachment to the cellwall after heterologous expression of the prtB gene inLactococcus lactis. Appl Environ Microbiol 69:3377–3384
Gilbert C, Atlan D, Blanc B, Portalier R (1994) Proline iminopepti-dase from Lactobacillus delbrueckii subsp. bulgaricus CNRZ397: purification and characterization. Microbiology 140:537–542
Gilbert C, Atlan D, Blanc B, Portalier R, Germond GJ, Lapierre L,Mollet B (1996) A new cell surface proteinase: sequencing andanalysis of the prtB gene from Lactobacillus debrueckii subsp.bulgaricus. J Bacteriol 178:3059–3065
Gitton C, Meyrand M, Wang J, Caron C, Trubuil A, Guillot A,Mistou MY (2005) Proteomic signature of Lactococcus lactisNCDO763 cultivated in milk. Appl Environ Microbiol71:7152–7163
Gobbetti M, Stepaniak L, De Angelis M, Corsetti A, Di Cagno R(2002) Latent bioactive peptides in milk proteins: proteolyticactivation and significance in dairy processing. CRC Crit RevFood Sci Nutr 42:223–239
Gottesman S (1996) Proteases and their targets in Escherichia coli.Annu Rev Genet 30:465–506
Guédon E, Renault P, Ehrlich D, Delorme C (2001a) Transcriptionalpattern of genes coding for the proteolytic system ofLactococcus lactis and evidence for coordinated regulation ofkey enzymes by peptide supply. J Bacteriol 183:3614–3622
Guédon E, Serror P, Ehrlich SD, Renault P, Delorme C (2001b)Pleiotropic transcriptional repressor CodY senses the intracel-lular pool of branched-chain amino acids in Lactococcus lactis.Mol Microbiol 40:1227–1239
Guldtfeldt LU, Sørensen KI, Strøman P, Behrndt H, Williams D,Johansen E (2001) Effect of starter cultures with a geneticallymodified peptidolytic or lytic system on cheddar cheeseripening. Int Dairy J 11:373–382
403

Haandrikman A, Kok J, Laan H, Soemitro S, Ledeboer A,Konings W, Venema G (1989) Identification of a generequired for maturation of an extracellular lactococcal serineproteinase. J Bacteriol 171:2789–2794
Haandrikman A, Kok J, Venema G (1991) Lactococcal proteinasematuration protein PrtM is a lipoprotein. J Bacteriol173:4517–4525
Hagting A, Kunji E, Leenhouts K, Poolman B, Konings W (1994)The di- and tripeptide transport protein of Lactococcus lactis. Anew type of bacterial peptide transporter. J Biol Chem269:11391–11399
Hebert EM, Raya RR, De Giori GS (2000) Nutritional requirementsand nitrogen-dependent regulation of proteinase activity ofLactobacillus helveticus CRL 1062. Appl Environ Microbiol66:5316–5321
Hellendoorn MA, Franke-Fayard BM, Mierau I, Venema G, Kok J(1997) Cloning and analysis of the pepV dipeptidase gene ofLactococcus lactis MG1363. J Bacteriol 179:3410–3415
Henrich B, Klein JR, Weber B, Delorme C, Renault P, Wegmann U(2002) Food-grade delivery system for controlled gene expres-sion. Appl Environ Microbiol 68:5429–5436
Hickey RM, Ross RP, Hill C (2004) Controlled autolysis and enzymerelease in a recombinant Lactococcal strain expressing themetalloendopeptidase enterolysin A. Appl Environ Microbiol70:1744–1748
Higgins CF (1992) ABC transporters: from microorganisms to man.Annu Rev Cell Biol 67–113
Holck A, Naes H (1992) Cloning, sequencing and expression of thegene encoding the cell-envelope-associated proteinase fromLactobacillus paracasei subsp. paracasei NCDO 151. J GenMicrobiol 138:1353–1364
I’Anson K, Movahedi S, Griffin H, Gasson M, Mulholland F (1995)A non-essential glutamyl aminopeptidase is required foroptimal growth of Lactococcus lactis MG1363 in milk.Microbiology 141:2873–2881
Joutsjoki V, Luoma S, Tamminen M, Kilpi M, Johansen E, Palva A(2002) Recombinant Lactococcus starters as a potential sourceof additional peptidolytic activity in cheese ripening. J ApplMicrobiol 92:1159–1166
Juillard V, Laan H, Kunji E, Jeronimus-Stratingh CM, Bruins A,Konings W (1995) The extracellular PI-type proteinase ofLactococcus lactis hydrolyzes β-casein into more than onehundred different oligopeptides. J Bacteriol 177:3472–3478
Juillard V, Guillot A, Le Bars D, Gripon JC (1998) Specificity ofmilk peptide utilization by Lactococcus lactis. Appl EnvironMicrobiol 64:1230–1236
Katayama-Fujimura Y, Gottesman S, Maurizi MR (1987) Amultiple-component, ATP-dependent protease from Escher-ichia coli. J Biol Chem 262:4477–4485
Klaenhammer TR, Barrangou R, Buck BL, Azcarate-Peril MA,Altermann E (2005) Genomic features of lactic acid bacteriaeffecting bioprocessing and health. FEMS Microbiol Rev29:393–409
Kleerebezem M, Boekhorst J, van Kranenburg R, Molenaar D,Kuipers OP, Leer R, Tarchini R, Peters SA, Sandbrink HM,Fiers MW, Stiekema W, Lankhorst RM, Bron PA, Hoffer SM,Groot MN, Kerkhoven R, de Vries M, Ursing B, de Vos WM,Siezen RJ (2003) Complete genome sequence of Lactobacillusplantarum WCFS1. Proc Natl Acad Sci U S A 100:1990–1995
Klein J, Klein U, Schad M, Plapp R (1993) Cloning, DNA sequenceand partial characterization of pepN, a lysyl aminopeptidasefrom Lactobacillus delbrueckii ssp. lactis DSM 7290. Eur JBiochem 217:105–114
Klein J, Henrich B, Plapp R (1994a) Cloning and nucleotide sequenceanalysis of the Lactobacillus delbrueckii ssp. lactis DSM7290cysteine aminopeptidase gene pepC. FEMS Microbiol Lett124:291–300
Klein J, Schmidt U, Plapp R (1994b) Cloning, heterologousexpression, and sequencing of a novel proline iminopeptidasegene, pepI, from Lactobacillus delbrueckii subsp. lactisDSM7290. Microbiology 140:1133–1139
Klein J, Dick A, Schick J, Matern H, Henrich B, Plapp R (1995)Molecular cloning and DNA sequence analysis of pepL, aleucyl aminopeptidase gene from Lactobacillus delbrueckiisubsp. lactis DSM7290. Eur J Biochem 228:570–578
Klein JR, Schick J, Henrich B, Plapp R (1997) Lactobacillusdelbrueckii subsp. lactis DSM7290 pepG gene encodes a novelcysteine aminopeptidase. Microbiology 143:527–537
Kok J, de Vos WM (1994) The proteolytic system of lactic acidbacteria. In: Gasson M, De Vos W (eds) Genetics andbiotechnology of lactic acid bacteria. Blackie Academic &Professional, Glasgow, pp 169–210
Kok J, Leenhouts KJ, Haandrikman AJ, Ledeboer AM, Venema G(1988) Nucleotide sequence of the cell wall proteinase gene ofStreptococcus cremoris Wg2. Appl Environ Microbiol54:231–238
Korhonen H, Pihlanto A (2003) Food-derived bioactive peptides—opportunities for designing future foods. Curr Pharm Des9:1297–1308
Kunji E, Hagting A, De Vries C, Juillard V, Haandrikman A,Poolman B, Konings WN (1995) Transport of β-casein-derivedpeptides by the oligopeptide transport system is a crucial step inthe proteolytic pathway of Lactococcus lactis. J Biol Chem270:1569–1574
Kunji ERS, Mierau I, Hagting A, Poolman B, Konings WN (1996)The proteolytic systems of lactic acid bacteria. Antonie VanLeeuwenhoek 70:187–221
Kunji ER, Fang G, Jeronimus-Stratingh CM, Bruins AP, Poolman B,Konings WN (1998) Reconstruction of the proteolytic pathwayfor use of beta-casein by Lactococcus lactis. Mol Microbiol27:1107–1118
Laskowska E, Kuczynska-Wisnik D, Skorko-Glonek J, Taylor A(1996) Degradation by proteases Lon, Clp and HtrA, ofEscherichia coli proteins aggregated in vivo by heat shock;HtrA protease action in vivo and in vitro. Mol Microbiol22:555–571
Leenhouts K, Buist G, Kok J (1999) Anchoring of proteins to lacticacid bacteria. Antonie Van Leeuwenhoek 76:367–376
Leroy F, Devuyst L (2004) Lactic acid bacteria as functional startercultures for the food fermentation industry. Trends Food SciTechnol 15:67–78
Luoma S, Peltoniemi K, Joutsjoki V, Rantanen T, Tamminen M,Heikkinen I, Palva A (2001) Expression of six peptidases fromLactobacillus helveticus in Lactococcus lactis. Appl EnvironMicrobiol 67:1232–1238
Madkor SA, Tong PS, El Soda M (2000) Ripening of cheddarcheese with added attenuated adjunct cultures of lactobacilli.J Dairy Sci 83:1684–1691
Martínez-Cuesta MC, Fernández de Palencia P, Requena T, Peláez C(1998) Enhancement of proteolysis by a Lactococcus lactisbacteriocin producer in a cheese model system. J Agric FoodChem 46:3863–3887
Marugg JD, Meijer W, van Kranenburg R, Laverman P, BruinenbergPG, de Vos WM (1995) Medium-dependent regulation ofproteinase gene expression in Lactococcus lactis: control oftranscription initiation by specific dipeptides. J Bacteriol177:2982–2989
Matos J, Nardi M, Kumura H, Monnet V (1998) Geneticcharacterization of pepP, which encodes an aminopeptidase Pwhose deficiency does not affect Lactococcus lactis growth inmilk, unlike deficiency of the X-prolyl dipeptidyl aminopep-tidase. Appl Environ Microbiol 64:4591–4595
Meijer WC, Dobbelaar C, Hugenholtz J (1998) Thermoinduciblelysis in Lactococcus lactis subsp. cremoris SK110: implicationsfor cheese ripening. Int Dairy J 8:275–280
Meisel H (2004) Multifunctional peptides encrypted in milkproteins. Biofactors 21:55–61
Meisel H, Bockelman W (1999) Bioactive peptides encrypted inmilk proteins: proteolytic activation and thropho-functionalproperties. Antonie Van Leeuwenhoek 76:207–215
404

Meyer J, Spahni A (1998) Influence of X-prolyl-dipeptidylamino-peptidase of Lactobacillus delbrueckii subsp. lactis on prote-olysis and taste of Swiss Gruyére cheese. Milchwissenschaft53:449–453
Meyer-Barton EC, Klein JR, Imam M, Plapp R (1993) Cloning andsequence analysis of the X-propyl-dipeptidyl-aminopeptidasegene (pepX) from Lactobacillus delbrueckii spp. lactisDSM7290. Appl Microbiol Biotechnol 40:82–89
Mierau I, Tan P, Haandrikman A, Mayo B, Kok J, Konings W,Venema G (1993) Cloning and sequencing of the gene for alactococcal endopeptidase, an enzyme with sequence similarityto mammalian enkephalinase. J Bacteriol 175:2087–2096
Mierau I, Haandrikman A, Velterop O, Tan P, Leenhouts K, Kok J,Venema G (1994) Tripeptidase gene (pepT) of Lactococcuslactis: molecular cloning and nucleotide sequencing of pepTand construction of a chromosomal deletion mutant. J Bacteriol176:2854–2861
Mierau I, Kunji ERS, Leenhouts KJ, Hellendoorn MA, HaandrikmanAJ, Poolman B, Konings WN (1996) Multiple-peptidase mutantsof Lactococcus lactis are severely impaired in their ability togrow in milk. J Bacteriol 178:2794–2803
Mierau I, Kunji ER, Venema G, Kok J (1997) Casein and peptidedegradation in lactic acid bacteria. Biotechnol Genet Eng Rev14:279–301
Monnet V, Nardi M, Chopin A, Chopin MC, Gripon JC (1994)Biochemical and genetic characterization of PepF, anoligopeptidase from Lactococcus lactis. J Biol Chem269:32070–32076
Morel F, Frot-Coutaz J, Aubel D, Portalier R, Atlan D (1999)Characterization of a prolidase from Lactobacillus delbrueckiisubsp. bulgaricus CNRZ 397 with an unusual regulation ofbiosynthesis. Microbiology 145:437–446
Morel F, Lamarque M, Bissardon I, Atlan D, Galinier A (2001)Autoregulation of the biosynthesis of the CcpA-like protein,PepR1, in Lactobacillus delbrueckii subsp. bulgaricus. J MolMicrobiol Biotechnol 3:63–66
Morgan S, Ross RP, Hill C (1997) Increasing starter cell lysis incheddar cheese using a bacteriocin-producing adjunct. J DairySci 80:1–10
Nakajima H, Hagting A, Kunji ER, Poolman B, Konings WN(1997) Cloning and functional expression in Escherichiacoli of the gene encoding the di- and tripeptide transportprotein of Lactobacillus helveticus. Appl Environ Microbiol63:2213–2217
Nardi M, Chopin MC, Chopin A, Cals M, Gripon J (1991) Cloningand DNA sequence analysis of an X-prolyl dipeptidyl amino-peptidase gene from Lactococcus lactis subsp. lactis NCDO763. Appl Environ Microbiol 57:45–50
Nardi M, Renault P, Monnet F (1997) Duplication of the pepF geneand shuffling of DNA fragments on the lactose plasmid ofLactococcus lactis. J Bacteriol 179:4164–4171
Navarre WW, Schneewind O (1994) Proteolytic cleavage and cellwall anchoring at the LPXTG motif of surface proteins in gram-positive bacteria. Mol Microbiol 14:115–121
Nilsson D, Lauridsen AA, Tomoyasu T, Ogura T (1994) A Lactococcuslactis gene encodes a membrane protein with putative ATPaseactivity that is homologous to the essential Escherichia coli ftsHgene product. Microbiology 140:2601–2610
Nouaille S, Ribeiro LA, Miyoshi A, Pontes D, Le Loir Y, OliveiraSC, Langella P, Azevedo V (2003) Heterologous proteinproduction and delivery systems for Lactococcus lactis. GenetMol Res 2:102–111
Pastar I, Tonic I, Golic N, Kojic M, van Kranenburg R, KleerebezemM, Topisirovic L, Jovanovic G (2003) Identification andgenetic characterization of a novel proteinase, PrtR, from thehuman isolate Lactobacillus rhamnosus BGT10. Appl EnvironMicrobiol 69:5802–5811
Pederson JA, Mileski GJ, Weimer BC, Steele JL (1999) Geneticcharacterization of a cell envelope-associated proteinase fromLactobacillus helveticus CNRZ32. J Bacteriol 181:4592–4597
Peltoniemi K, Vesanto E, Palva A (2002) Genetic characteriza-tion of an oligopeptide transport system from Lactobacillusdelbrueckii subsp. bulgaricus. Arch Microbiol 177:457–467
Petranovic D, Guedon E, Sperandio B, Delorme C, Ehrlich D,Renault P (2004) Intracellular effectors regulating the activityof the Lactococcus lactis CodY pleiotropic transcriptionregulator. Mol Microbiol 53:613–621
Pihlanto A, Korhonen H (2003) Bioactive peptides and proteins.Adv Food Nutr Res 47:175–276
Poolman B, Kunji E, Hagting A, Juillard V, Konings W (1995) Theproteolytic pathway of Lactococcus lactis. J Appl Bacteriol79:65–75
Poquet I, Saint V, Seznec E, Simoes N, Bolotin A, Gruss A (2000)HtrA is the unique surface housekeeping protease in Lacto-coccus lactis and is required for natural protein processing. MolMicrobiol 2000 35:1042–1051
Pridmore RD, Berger B, Desiere F, Vilanova D, Barretto C, PittetAC, Zwahlen MC, Rouvet M, Altermann E, Barrangou R,Mollet B, Mercenier A, Klaenhammer T, Arigoni F, Schell MA(2004) The genome sequence of the probiotic intestinalbacterium Lactobacillus johnsonii NCC 533. Proc Natl AcadSci USA 101:2512–2517
Pritchard GG, Coolbear T (1993) The physiology and biochemistryof the proteolytic system in lactic acid bacteria. FEMSMicrobiol Rev 12:179–206
Rantanen T, Palva A (1997) Lactobacilli carry cryptic genesencoding peptidase-related proteins: characterization of aprolidase gene (pepQ) and a related cryptic gene (orfZ) fromLactobacillus delbrueckii subsp. bulgaricus. Microbiology143:3899–3905
Rodríguez J, Requena T, Goudédranche H, Maubois JL, Juárez M(1996) Accelerated ripening of reduced fat semi-hard cheesefrom a mixture of cow’s, goat’s and ewe’s ultrafiltrated milk byusing a Lac-Prt-strain of lactococci. Lait 76:513–522
Sanz Y, Toldrá F, Renault P, Poolman B (2003) Specificity of thesecond binding protein of the peptide ABC-transporter (Dpp) ofLactococcus lactis IL1403. FEMS Microbiol Lett 227:33–38
Savijoki K, Palva A (2000) Purification and molecular characteriza-tion of a tripeptidase (PepT) from Lactobacillus helveticus.Appl Environ Microbiol 66:794–800
Savijoki K, Ingmer H, Frees D, Vogensen FK, Palva A, Varmanen P(2003) Heat and DNA damage induction of the LexA-likeregulator HdiR from Lactococcus lactis is mediated by RecAand ClpP. Mol Microbiol 50:609–621
Saxelin M, Tynkkynen S, Mattila-Sandholm T, de Vos W (2005)Probiotic and other functional microbes: from markets tomechanisms. Curr Opin Biotechnol 16:204–211
Schick J, Weber B., Klein JR, Henrich B (1999) PepR1, a CcpA-liketranscription regulator of Lactobacillus delbrueckii susbp.lactis. Microbiology 145:3147–3154
Shao W, Yuksel GU, Dudley EG, Parkin KL, Steele JL (1997)Biochemical and molecular characterization of PepR, a dipep-tidase, from Lactobacillus helveticus CNRZ32. Appl EnvironMicrobiol 63:3438–3443
Siezen RJ (1999) Multi-domain, cell-envelope proteinases of lacticacid bacteria. Antonie Van Leeuwenhoek 76:139–155
Smeds A, Varmanen P, Palva A (1998) Molecular characterization ofa stress-inducible gene from Lactobacillus helveticus. J Bacteriol180:6148–6153
Smit G, Smit BA, Engels WJM (2005) Flavour formation by lacticacid bacteria and biochemical flavour profiling of cheeseproducts. FEMS Microbiol Rev 29:591–610
Smukowski M, Wendorff WL, Ping Y, Rao RD (2003) Impact ofcheese defects on U.S. graded cheeses. J Dairy Sci 86:364
Sridhar VR, Hughes JE, Welker DL, Broadbent JR, Steele JL (2005)Identification of endopeptidase genes from the genomicsequence of Lactobacillus helveticus CNRZ32 and the role ofthese genes in hydrolysis of model bitter peptides. ApplEnviron Microbiol 71:3025–3032
Stefanitsi D, Sakellaris G, Garel JR (1995) The presence of twoproteinases associated with the cell wall of Lactobacillusbulgaricus. FEMS Microbiol Lett 128:53–58
405

Strøman P (1992) Sequence of a gene (lap) encoding a 95.3-kDaaminopeptidase from Lactococcus lactis ssp. cremoris Wg2.Gene 113:107–112
Stucky K, Klein J, Schüller A, Matern H, Henrich B, Plapp R (1995)Cloning and DNA sequence analysis of pepQ, a prolidase genefrom Lactobacillus delbrueckii subsp. lactis DSM7290 andpartial characterization of its product. Mol Gen Genet 247:494–500
Stucky K, Schick J, Klein JR, Henrich B, Plapp R (1996)Characterization of pepR1, a gene coding for a potentialtranscriptional regulator of Lactobacillus delbrueckii subsp.lactis DSM7290. FEMS Microbiol Lett 136:63–69
Swaisgood H (1982) Chemistry of milk protein. In: Fox P (ed)Developments in dairy chemistry, vol. 1. Applied Science,London, pp 1–59
Tan P, van Alen-Boerrigter I, Poolman B, Siezen R, de Vos W,Konings W (1992) Characterization of the LactococcuslactispepN encoding an aminopeptidase homologous to mam-malian aminopeptidase N. FEBS Lett 306:9–16
Tuler TR, Callanan MJ, Klaenhammer TR (2002) Overexpression ofpeptidases in Lactococcus and evaluation of their release fromleaky cells. J Dairy Sci 85:2438–2450
Tynkkynen S, Buist G, Kunji E, Kok J, Poolman B, Venema G,Haandrikman A (1993) Genetic and biochemical characteriza-tion of the oligopeptide transport system of Lactococcus lactis.J Bacteriol 175:7523–7532
Varmanen P, Vesanto E, Steele J, Palva A (1994) Characterizationand expression of the pepN gene encoding a general amino-peptidase from Lactobacillus helveticus. FEMS Microbiol Lett124:315–320
Varmanen P, Rantanen T, Palva A (1996a) An operon fromLactobacillus helveticus composed of a proline iminopepti-dase gene (pepI) and two genes coding for putative membersof the ABC transporter family of proteins. Microbiology142:3459–3468
Varmanen P, Steele J, Palva A (1996b) Characterization of aprolinase gene and its product and an adjacent ABC transportergene from Lactobacillus helveticus. Microbiology 142:809–816
Varmanen P, Rantanen T, Palva A, Tynkkynen S (1998) Cloning andcharacterization of a prolinase gene (pepR) from Lactobacillusrhamnosus. Appl Environ Microbiol 64:1831–1836
Varmanen P, Savijoki K, Åvall S, Palva A, Tynkkynen S (2000a) X-prolyl dipeptidyl aminopeptidase gene (pepX) is part of theglnRA operon in Lactobacillus rhamnosus. J Bacteriol182:146–154
Varmanen P, Ingmer H, Vogensen FK (2000b) ctsR of Lactococcuslactis encodes a negative regulator of clp gene expression.Microbiology 146:1447–1455
Vermeulen N, Pavlovic M, Ehrmann MA, Gänzle MG, Vogel RF(2005) Functional characterization of the proteolytic system ofLactobacillus sanfranciscensis DSM 20451 during growth insourdough. Appl Environ Microbiol 71:6260–6266
Vesanto E, Varmanen P, Steele J, Palva A (1994) Characterizationand expression of the Lactobacillus helveticuspepC geneencoding a general aminopeptidase. Eur J Biochem224:991–997
Vesanto E, Savijoki K, Rantanen T, Steele J, Palva A (1995) An X-prolyl dipeptidyl aminopeptidase (pepX) gene from Lactoba-cillus helveticus. Microbiology 141:3067–3075
Vesanto E, Peltoniemi K, Purtsi T, Steele J, Palva A (1996)Molecular characterization, over-expression and purification ofa novel dipeptidase from Lactobacillus helveticus. ApplMicrobiol Biotechnol 45:638–645
Vido K, Le Bars D, Mistou MY, Anglade P, Gruss A, Gaudu P(2004) Proteome analyses of heme-dependent respirationin Lactococcus lactis: involvement of the proteolyticsystem. J Bacteriol 186:1648–1657
Visser S, Exterkate F, Slangen C, de Veer GJCM (1986)Comparative study of action of cell wall proteinases fromvarious strains of Streptococcus cremoris on bovine αs1-, β-,and κ-casein. Appl Microbiol Biotechnol 29:61–66
Vongerichten KF, Klein JR, Matern H, Plapp R (1994) Cloning andnucleotide sequence analysis of pepV, a carnosinase gene fromLactobacillus delbrueckii subsp. lactis DSM7290, and partialcharacterization of the enzyme. Microbiology 140:2591–2600
Wegmann U, Klein R, Drumm I, Kuipers OP, Henrich B (1999)Introduction of peptidase genes from Lactobacillus delbrueckiisubsp. lactis into Lactococcus lactis and controlled expression.Appl Environ Microbiol 65:4729–4733
Yüksel GU, Steele JL (1996) DNA sequence analysis, expression,distribution, and physiological role of the Xaa-prolyl dipeptidylaminopeptidase gene from Lactobacillus helveticus CNRZ32.Appl Microbiol Biotechnol 44:766–773
406