photosynthetic hydrogen production
TRANSCRIPT

R
P
SSMa
b
c
d
e
f
a
ARRAA
KCHPNH
C
T
1d
Journal of Photochemistry and Photobiology C: Photochemistry Reviews 11 (2010) 101–113
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology C:Photochemistry Reviews
journa l homepage: www.e lsev ier .com/ locate / jphotochemrev
eview
hotosynthetic hydrogen production
uleyman I. Allakhverdieva,b,∗, Velmurugan Thavasi c, Vladimir D. Kreslavskib,ergey K. Zharmukhamedovb, Vyacheslav V. Klimovb, Seeram Ramakrishnac, Dmitry A. Losa,amoru Mimurod, Hiroshi Nishiharae, Robert Carpentier f
Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya Street 35, Moscow 127276, RussiaInstitute of Basic Biological Problems, Russian Academy of Sciences, Pushchino, Moscow Region 142290, RussiaNanoscience and Nanotechnology Initiative, National University of Singapore, 2, Engineering Drive 3, Singapore 117576, SingaporeGraduate School of Human and Environmental Studies, Kyoto University, Kyoto 606-8501, JapanDepartment of Chemistry, School of Science, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, JapanGroupe de recherche en biologie végétale, Université du Québec à Trois-Rivières, CP 500, Trois-Rivières, Québec G9A 5H7, Canada
r t i c l e i n f o
rticle history:eceived 25 May 2010eceived in revised form 23 July 2010ccepted 24 July 2010
a b s t r a c t
Among various technologies for hydrogen production, the use of oxygenic natural photosynthesis hasa great potential as can use clean and cheap sources—water and solar energy. In oxygenic photosyn-thetic microorganisms electrons and protons produced from water and redirected by the photosyntheticelectron-transport chain via ferredoxin to the hydrogen-producing enzymes hydrogenase or nitrogenase.
vailable online 10 August 2010
eywords:lean energyydrogen photoproduction
By these enzymes, e− and H+ recombine and form molecular hydrogen. Hydrogenase activity can be veryhigh but is extremely sensitive to the photosynthetically evolved O2 that leads to reduced and unstable H2
production. However, presently, several approaches are developed to improve the energetic efficiencyto generate H2. This review examines the main available pathways to improve the photosynthetic H2
hotosynthetic organismsitrogenaseydrogenase
production.© 2010 Elsevier B.V. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1022. Oxygenic organisms (cyanobacteria and microalgae) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 104
2.1. The use of photosynthesis for H2 production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1042.2. Enzymes for biohydrogen production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1042.3. Structures of hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1052.4. Pathways for H2 photoproduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1062.5. Separation of O2 and H2 production. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1072.6. Pathways of increasing light to H2 photon conversion efficiency . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 108
2.6.1. Reduced antenna size and increased PQ pool . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1082.6.2. Immobilization of microbial cultures. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1082.6.3. The use of mimics of water oxidation systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1082.6.4. Enhanced resistance to environmental stress conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1082.6.5. Genetic engineering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1092.6.6. Role of photosystems in H2 photoproduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110
3. Anoxygenic organisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Abbreviations: Chl, chlorophyll; Fd, ferredoxin; PQ, plastoquinone; PS, photosystem;∗ Corresponding author: Institute of Basic Biological Problems, Russian Academy of Scieel.: +7 4967731837; fax: +7 4967330532.
E-mail addresses: [email protected], [email protected] (S.I. A
389-5567/$20.00 © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.jphotochemrev.2010.07.002
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111
RC, reaction center.nces, ul. Institutskaya 2, Pushchino, Moscow Region 142290, Russian.
llakhverdiev).

1 Photo
natase
02 S.I. Allakhverdiev et al. / Journal of Photochemistry and
Suleyman I. Allakhverdiev is head of the Laboratoryof “Controlled Photobiosynthesis” at the K.A. TimiryazevInstitute of Plant Physiology, Russian Academy of Sciences(RAS), Moscow, and Chief Research Scientist (highest/topposition) at the Institute of Basic Biological Problems,RAS, Pushchino, Moscow Region, Russia. He obtained hisDr.Sci. degree in photobiochemistry and plant physiologyfrom K.A. Timiryazev Institute of Plant Physiology (2002,Moscow), and Ph.D. in physics and mathematics, fromInstitute of Biophysics (1984, Pushchino). He graduatedfrom Azerbaijan State University, Department of Physicswith B.S./M.S. in physics (Baku). He has been guest-editorand is a member of the Editorial Board of several inter-
ational journals. He also acts as a referee for major international journals. Heuthored above 300 papers. He has been cited ca. 3500. He organized several interna-ional conferences on photosynthesis. His research interests include the structurend function of photosystem II, water oxidizing complex, artificial photosynthe-is, hydrogen photoproduction, catalytic conversion of solar energy, plant undernvironmental stresses, photoreceptor signaling.
Dr Velmurugan Thavasi is a research fellow at the NUSNanoscience and Nanotechnology Initiative. He obtainedan M.Eng. in chemical engineering and Ph.D. in chemistryfrom National University of Singapore. His research inter-ests include materials synthesis and interface engineeringfor energy and electronic devices.
Vladimir D. Kreslavski is a senior researcher. He is thehead of group in Laboratory of Ecology and Physiologyof phototrophic organisms at Institute of Basic Biolog-ical Problems, RAS, Pushchino, Moscow Region, Russia.The field of interests: photoreceptor signaling, molecu-lar mechanisms of plant stress resistance and acclimationof photosynthetic apparatus as well as the pathways ofphotosynthetic improvement.
Sergey K. Zharmukhamedov is a senior researcher atthe Institute of Basic Biological Problems, RAS, Pushchino,Russia. He defended Ph.D. thesis in biology on the sub-ject “Revealing and investigation of the mechanism ofaction of new chemical inhibitors of electron transfer inphotosystem II of plants” (1993, Pushchino). He gradu-ated from Kazakhstan State University, Department ofBiochemistry and Biophysics (1983). The field of interests:biophysics and biochemistry of photosynthesis (mecha-nism of inhibitory action of herbicides and stress factors).
Dr Seeram Ramakrishna is a professor of MechanicalEngineering and Vice-President (Research Strategy) at theNational University of Singapore. He is an acknowledgedglobal leader on nanofibers. He authored more than 330journal papers, five books, and 25 book chapters. He nur-tured ten technology entrepreneurs. He is among theworld’s top fifty materials scientists according to the ISIEssential Science Indicators. He is an elected Fellow ofRoyal Academy of Engineering, UK. He is also a fellow ofall major disciplenary professional societies in Singapore,UK and USA.
Dmitry A. Los is a professor at the K.A. Timiryazev Insti-tute of Plant Physiology, Moscow, Russia. He obtainedhis Dr.Sci. degree in plant physiology and biochemistryin 1997 (Moscow). His research interests are in the areaof stress sensors and transduction pathways in photosyn-thetic cells.
biology C: Photochemistry Reviews 11 (2010) 101–113
Mamoru Mimuro is a professor of biology in the Gradu-ate School of Human and Environmental Studies, KyotoUniversity. He received a Ph.D. degree in botany fromthe University of Tokyo in 1978. He graduated the Uni-versity of Tokyo, Botany Department. He has been anEditorial Board member of photosynthesis research, andalso served as a guest-editor of several journals. His inter-est is an oxygen-evolving reaction and also photosystemII reactions. In 1992, he started a satellite meeting on“Pigment system and energy transfer” on the occasion ofPhotosynthesis Congress held in Nagoya.
Hiroshi Nishihara is a professor of Department of Chem-istry, School of Science at The University of Tokyo. Hereceived his B. Sc. degree in 1977 and D. Sc. in 1982from The University of Tokyo. He was appointed researchassociate of Department of Chemistry, Faculty of Scienceand Technology at Keio University in 1982, and promotedassociate professor in 1992. Since 1996, he has been a pro-fessor at The University of Tokyo. He also worked as avisiting research associate at The University of North Car-olina at Chapel Hill (1987–1989), and as a researcher ofPRESTO, JRDC (1992–1996). His research has been focusedon creation of new electro- and photo-functional materi-als comprising both transition metals and pi-conjugated
chains, invention of unidirectional electron transfer systems utilizing molecularlayer interfaces, and bio-conjugated photo-electronic devices.
Robert Carpentier is professor at Université du Québecà Trois- Rivières, Québec, Canada. He obtained his Ph.D.in biochemistry from Laval University (1983, Québec).He is editor of Journal of Photochemistry and PhotobiologyB: Biology, associate editor of the journal PhotosynthesisResearch and was the chair of the XIIIth Interna-tional Congress on Photosynthesis (Montréal, 2004). Hisresearch interests concern the influence of environmentalstresses on electron transport pathways in photosystemsI and II and energy dissipation in photosynthesis.
1. Introduction
Solar energy stored by photosynthesis over billions of yearsconstitutes the main source of energy available on Earth. It cantherefore be argued that one of the most important events in theEarth’s history is the evolution of photosynthetic organisms capa-ble of water oxidation. In these organisms, photosystem II (PS II)uses sunlight to split water, an energetically unfavorable reaction,where electrons and protons are extracted from water, and oxygenis released as a by-product. In turn, accumulation of the oxygenevolved resulted in an aerobic atmosphere. Formation of an ozonelayer allowed organisms to move from the ocean to the land.
Thanks to photosynthesis, captured solar energy is accumulatedin the form of coal, oil and gas. These fuels have been intensivelyused and are becoming limited. Out of the global energy consump-tion in 2005, 86% was obtained from fossil fuels (oil, 37%; coal, 23%;natural gas, 26%) [http://www.eia.doe.gov/oiaf/ieo/index.html].Moreover, global energy consumption will increase from thecurrent level of 12.8 TW to 27 TW by 2050, which will leadto a global warming of the Earth [1]. During the last 50years, the concentration of atmospheric CO2 increased by18% [http://www.esrl.noaa.gov/gmd/ccgg/trends/] and the surfacetemperature of the Earth increased by 0.64 ◦C [2]. It is still debat-able how much the activity of human industry is responsible forthis global change of climate because there are so many variables.Nevertheless, there is one thing for which human activity is defi-
nitely responsible for: the invention of many ways to liberate CO2,and hardly anyway to assimilate it. This is problematic because weare distorting the balance of the carbon cycle by consumption ofour inherited carbon resource, without much hope for its renewal.Therefore, renewable and clean energy sources must be developed
S.I. Allakhverdiev et al. / Journal of Photochemistry and Photobiology C: Photochemistry Reviews 11 (2010) 101–113 103
Fig. 1. Schematic view of light-powered H2 production during oxygenic photosynthesis and subsequent formation of carbohydrates in microalgae. The photosyntheticprocesses are driven by the light energy captured by the light-harvesting complexes of photosystem I (PS I) and photosystem II (PS II). Electrons derived from H2O by wateroxidation at the oxygen-evolving complex (OEC) of PS II passed along the photosynthetic electron-transport chain via plastoquinone (PQ), the cytochrome b6/f complex (Cytb6/f), plastocyanin (PC), photosystem I (PS I), and ferredoxin (Fd). Then, ferredoxin–NADP+ oxidoreductase transfers the electrons to NADP+ with final production of NADPH.H+ are released into the thylakoid lumen by the OEC and the PQ/PQH2 cycle and used to produce ATP via the ATP synthase. The ATP and NADPH generated during the primaryphotosynthetic processes are consumed for CO2 fixation in the Calvin–Benson cycle, which produces sugars and ultimately starch. Under anaerobic conditions, hydrogenasecan accept electrons from reduced Fd molecules and use them to reduce protons to molecular hydrogen (H2). Anaerobic conditions allow to use starch as a source of H+ ande Thylac
A
icwedf
btaaitbh
fDcari[ettot
gecnt
− for H2 production (via NADPH, PQ, cytb6/f, PC and PS I) using the hydrogenase.olor and proton with brown color.
dapted from Refs. [5,10,11,15,16].
n the near future. In this regard, photosynthesis provides a suc-essful example of how solar energy can be converted into fuelhen electrons are extracted from water by using light as the only
nergy input. It would be wise to look into photosynthesis in furtheretail because the photosynthetic processes contain many cluesrom which we could learn.
Biological production of hydrogen using photosynthesis mayecome a valuable alternative to chemical and electrochemicalechnologies. Firstly, sun energy and water are cheap and renew-ble energy sources. Secondly, burning H2 is clean, emitting waters its end product, and a renewable process [3,4]. Photosynthesiss at the basis of all biological solar-driven methods of H2 produc-ion. Therefore, the development of approaches examining the linketween photosynthetic efficiency, photosynthetic products, andydrogen production is promising.
The fixation of CO2 by photosynthetic organisms and its trans-ormation into carbohydrates like starch include two phases.uring the light phase, light-harvesting pigments, (bacterio)-hlorophylls and carotenoids, as well as other pigments, act asntenna to collect solar energy. The absorbed light energy theneaches photochemical reaction centers (RC), where it is convertednto chemical energy by a series of electron transfer steps (Fig. 1)5–11]. Under normal physiological conditions of the dark phase,lectrons, protons, and ATP generated during the light-driven elec-ron flow are consumed in the Calvin–Benson cycle that fixes CO2o carbohydrate, and the appropriate (perhaps slight excess) ratiof ATP to NADPH is maintained by cyclic electron flow around pho-osystem I (PS I) [12–14].
Photosynthesis can be either oxygenic (O2 producing) or anoxy-
enic [5–11,15,16]. Oxygenic organisms use solar energy to extractlectrons and protons from water mainly for the CO2 assimilationycle, and to produce oxygen (Fig. 1). Anoxygenic organisms doot possess the necessary oxidizing potential to oxidize H2O andherefore take electrons from electron donor substrates like H2Skoid membrane is denoted with green color. Electron transfer is shown with blue
or organic acids. Oxygenic photosynthesis takes place in higherplants, algae and cyanobacteria, whereas anoxygenic photosynthe-sis occurs in organisms such as green sulfur and purple non-sulfurbacteria. Some of the anoxygenic organisms are able to generatehydrogen quite efficiently. However, as they cannot get electronsfrom water, their use is not commercially viable for the photopro-duction of hydrogen on a large scale. This review thus focuses onoxygenic organisms such as algae and cyanobacteria that are ableto split water and evolve H2.
All oxygenic phototrophs extract electrons and protons fromwater and use them to reduce plastoquinone and NADP+ asenergy sources for the metabolism. However, oxygenic pho-totrophs including cyanobacteria and microalgae can transientlyproduce H2 under anaerobic conditions via proton reductioncatalyzed by hydrogenase (or nitrogenase) in competition withother intracellular processes. In this case the electrons and pro-tons, ultimately produced by water oxidation, are transducted viaferredoxin/NADPH to hydrogenase. Thus, the photosyntheticallyreduced ferredoxin (or NADPH) can serve as the physiological elec-tron donor to hydrogenase and links hydrogenase (nitrogenase) tothe electron-transport chain [4,10,11,15].
Although the conversion efficiency under ideal conditions (lowirradiation, red light) could be as high as 27% (21% for photosyn-thetic active radiation (PAR) [9–11,17]), there is a crucial problemassociated with the use of microorganisms for hydrogen productionowing to the simultaneous production of oxygen and hydrogen bythe two photosystems. The evolved O2 inhibits hydrogenase activ-ity and suppresses its synthesis. This leads to unstable and reducedphotoproduction of H2. Melis et al. [18] proposed a solution to
this problem using sulfur deficiency and photoheterotrophic con-ditions. Such conditions lead to anaerobiosis, which in turn inducesthe synthesis of hydrogenase. In this way, the microalgae Chlamy-domonas reinhardtii were able to assume prolonged H2 productioneven though the efficiency is limited by the strongly reduced activ-
1 Photo
idfatw
2
2
otpit(
ioo[fwctcc
atfttNvaoi
FiQc
04 S.I. Allakhverdiev et al. / Journal of Photochemistry and
ty of PS II [19]. The progress based on microalgal research hasemonstrated the possibility to use photoautotrophic conditionsor effective H2 bioproduction [10,11,19] and improve its efficiencynd durability [10,11,20–25]. The role of molecular hydrogen inheir metabolism and the pathways of H2 evolution and oxidationere considered earlier by Kondratieva and Gogotov [26].
. Oxygenic organisms (cyanobacteria and microalgae)
.1. The use of photosynthesis for H2 production
The light reactions of photosynthesis involve a sequential chainf events that includes light absorption, water splitting, electronransport, reduction of NADP+, and creation of a transmembraneroton gradient for ATP synthesis. Several main complexes are
nvolved in the process of oxygenic photosynthesis: PS II, includinghe water oxidation complex, PS I, and the cytochrome b6/f complexFig. 2).
The PS II RC contains several redox components (see Fig. 2),ncluding a special form of Chl a, P680 (the primary electron donorf PS II), pheophytin (Pheo) a as well as the primary and sec-ndary plastoquinone electron acceptors (QA and QB, respectively)6,8–11,17,27]. The electron from excited P680 in PS II RC is trans-erred through a number of carriers to the cytochrome complex,hich consists of several subunits including cytochrome f and
ytochrome b6. Electrons derived from water are transferred tohe oxidized P680. Finally, a series of redox reactions within theytochrome complex ultimately transfer the electrons to a plasto-yanin (PC) that acts as a shuttle to PS I.
In PS I, electrons are transferred from PS I RC to ferredoxin (Fd),protein that carries electrons to other reaction pathways outside
he thylakoid. The reaction center replaces the electrons trans-erred to Fd by accepting electrons from the PC located betweenhe cytochrome complex and PS I. Normally, Fd shuttle’s electronso the enzyme ferredoxin–NADP+–oxidoreductase that reduces
ADP+ to NADPH, an important source of reduction needed to con-ert CO2 to carbohydrates in the Calvin–Benson cycle. As electronsre transported through the electron-transport chain, protons (H+)utside the thylakoid are carried to the inner thylakoid space form-ng a proton gradient across the thylakoid membrane (Fig. 2).ig. 2. Schematic representation of the thylakoid membrane complexes of PS II, PS I andn Fig. 1 with their polypeptide composition. cyt b559, cytochrome b559; YZ and YD, tyro
B, primary and secondary quinone acceptors; cyt f, cytochrome f; b6, cytochrome b6; Fhlorophyll a; A1, phylloquinone; FX, FA and FB, FeS centres; Fd, ferredoxin; FNR, ferredox
biology C: Photochemistry Reviews 11 (2010) 101–113
Under anaerobic conditions, hydrogenase (nitrogenase) canaccept electrons from reduced Fd molecules and use them to reduceprotons to molecular hydrogen. Thus, under anaerobic conditions,the photosynthetic reducing power can be partitioned between atleast two pathways: CO2 reduction and H2 production. CO2 reduc-tion requires ATP, whereas H2 production by hydrogenase, that isthe desired pathway for renewable energy production, does not useATP. Under normal conditions, the competition for electron donorsfavors the CO2 fixation pathway. However, in the absence or limi-tation of CO2, the favorable pathway is the H2 production, which isdown-regulated due to the non-dissipation of the �pH caused bythe lack of ATP utilization.
There are two major research challenges related to the con-version of protons and electrons by light energy into H2: (i) thelevel of light intensities that can be efficiently utilized to drivephotosynthesis should be optimized for microorganisms; (ii) forhydrogenases, there are kinetic limitations on electron transportto the enzyme under H2-producing conditions. Nitrogenase usesATP during the production of H2, therefore, the efficiency of thenitrogenase system is lower than that of hydrogenase.
2.2. Enzymes for biohydrogen production
In contrast to terrestrial plants, many microalgae and cyanobac-teria are able to produce hydrogen, converting the protons releasedduring photosynthesis into H2 [10,11,17,22,28,29]. One of the mostwiderspread enzymes to be found in many of the microalgae andcyanobacteria is hydrogenase [10,11,21,22,30]. Two major classesof hydrogenases [NiFe]-hydrogenases and [FeFe]-hydrogenases arevery effective; their reaction rates usually lie in the range of103–104 turnovers per second [30]. The metal-clusters of hydroge-hases are unique in having CO and CN ligands sensitive to O2 and CO.[NiFe]-hydrogenases and [FeFe]-hydrogenases can be inactivatedby these agents and, especially in the latter case, the inactivationby O2 is irreversible [10,11,21].
Among microalgae, many unicellular green algae have the high-est rate of H2 production. Green algal hydrogenases that belongto the class of [FeFe]-hydrogenases are involved in much higherspecific activities than cyanobacterial [NiFe]-hydrogenases [31].Whereas [NiFe]-hydrogenases tend to be involved in H2 consump-
cytochrome b6/f responsible for electron transfer from H2O to NADP+ as describedsines Z and D; P680, primary electron donor of PS II; Pheo, pheophytin a; QA andeS, FeS Rieske centre; PC, plastocyanin; P700, primary electron donor of PS I; AO,in–NADP+–oxidoreductase.

S.I. Allakhverdiev et al. / Journal of Photochemistry and Photo
Fig. 3. Hydrogen metabolism by nitrogenase and hydrogenase. Nitrogenase com-prises 3 subunits, 2�-subunits (light gray ovals), 2�-subunits (dark gray oval) and 4dinitrogenase reductase subunits (light gray balls). Nitrogenase reduces N2 to NH3
and reduces electrons to H2, consuming ATP. The uptake hydrogenase comprisestwo subunits, HupL (the larger light gray oval) and HupS (the smaller dark grayoy
A
t[gAnremtafflrovttp
rtdw
icacgccfnstaerstghttplm
to an Fe(CN)2CO moiety by two conserved cysteine residues; (ii)
val), which regenerate electrons from H2. The actual structure of HupSL has notet been determined.
dapted from Refs. [34,35].
ion, [FeFe]-hydrogenases are usually involved in H2 production32]. Along with [NiFe]-hydrogenases, cyanobacteria also use nitro-enases that produce H2 contaminent with N2 fixation [16,17].
nitrogenase enzyme consists of two parts: one is dinitroge-ase (MoFe protein, encoded by the genes nifD and nifK, � and �espectively) and the other is dinitrogenase reductase (Fe protein,ncoded by nifH). Dinitrogenase is a �2�2 heterotetramer, havingolecular weight of about 220–240 kDa respectively, breaks apart
he atoms of nitrogen. Dinitrogenase reductase is a homodimer ofbout 60–70 kDa and plays the specific role of mediating the trans-er of electrons from the external electron donor (a ferredoxin or aavodoxin) to the dinitrogenase [29]. Several forms of cyanobacte-ial nitrogenases are known to date, with molybdenum, vanadiumr iron at the active site [29,33]. These enzymes do not have aery rapid catalytic cycle, with typical turnover numbers of lesshan 10 s−1 and need additional ATP consumption for H2 produc-ion that lowers the efficiency of the nitrogenase systems for H2hotoproduction [4].
The [FeFe]-hydrogenase enzyme is known to catalyze theeduction of protons into hydrogen according to the equa-ion: 2H+ + 2Fd− → H2 + 2Fd. Ferredoxin, being the natural electrononor, transports the electrons to the algal [FeFe]-hydrogenase asell as to nitrogenase (Fig. 1).
Hydrogenases are oxygen-labile enzymes, which make themncompatible with the oxygen-evolving photosynthesis present inyanobacteria; hence hydrogenase activity is restricted to cells withnoxic conditions. Based on current knowledge, various strains ofyanobacteria may have no hydrogenase, only an uptake hydro-enase, only a reversible hydrogenase, or both hydrogenases. Allyanobacterial hydrogenases are NiFe enzymes [29,34,35]. Manyyanobacteria that fix nitrogen have an uptake hydrogenase, whoseunction is to recover the electrons lost to hydrogen production byitrogenase (Fig. 3) [34]. The uptake hydrogenase comprises twoubunits encoded by hupS and hupL. The large subunit containshe four cysteines of the active site, two of which bridge the Fend Ni atoms. The small subunit has three FeS clusters that passlectrons from the active center to the electron acceptor in theespiratory electron-transport chain, thus producing ATP and con-uming oxygen, both beneficial for nitrogen fixation. In bacteriahat use hydrogen as an energy source, the small subunit of hydro-enase donates electrons to another protein encoded by hupC oroxZ that anchors the hydrogenase in the membrane and transfershe electrons to a respiratory electron-transport chain. In cyanobac-
eria there are no close homologs of hupC; thus, the anchoringrotein is unknown. The uptake hydrogenase in cyanobacteria isocalized on the cytoplasmic side of the cytoplasmic or thylakoidembrane [35]. The biological role of bidirectional or reversible
biology C: Photochemistry Reviews 11 (2010) 101–113 105
hydrogenase, is poorly understood and thought to control ion lev-els in the organism. Reversible hydrogenase is associated with thecytoplasmic membrane and likely functions as an electron acceptorfrom both NADH and H2 [36].
H2 production is often limited mainly because of the extremesensitive nature of hydrogenases to oxygen. A reduced O2 sensitiv-ity may be obtained by genetic modification of the hydrogenases.For genetic modification of the hydrogenases it is important tounderstand, why the hydrogenases are O2 sensitive [37]. The activesite of [FeFe]-hydrogenases, a complex structure known as the“H-cluster”, consists of a binuclear Fe center (2FeH) linked to a[4Fe4S] cluster [38,39]. The mechanism of oxygen inactivation ofthe [FeFe]-hydrogenase CrHydA1 from C. reinhardtii has been inves-tigated. X-ray absorption spectroscopy shows that reaction withoxygen results in destruction of the [4Fe4S] domain of the activesite H-cluster while leaving the di-iron domain (2FeH) essentiallyintact.
Schematic structures of the active site of [NiFe]- and [FeFe]-hydrogenases [32,40–43] are presented now days. Many ofthe bacterial [FeFe]-hydrogenases [32] have been characterizedbiochemically, and their genes have also been cloned and character-ized at the molecular level. In eukaryotes, the genes are located inthe nucleus, whereas the enzyme is localized to organelles (chloro-plast or hydrogenosome) of endosymbiotic origin. As it shownin many green algae, the enzyme is located in the chloroplaststroma and is linked via ferredoxin to the photosynthetic electron-transport chain [31,44].
2.3. Structures of hydrogenases
Hydrogenases are found in many organisms and catalyzereversible conversion between H2 and H+. In the active centerof a hydrogenase enzyme, there are two metal ions (FeFe orNiFe) that cooperate during the catalytic cycle for production(and consumption) of H2. [FeFe]-hydrogenases are small, monoor dimeric enzymes of 45–65 kDa. The active site cofactor is aunique [FeS] compound commonly referred to as “H-cluster” (seeabove). In vivo, [FeFe]-hydrogenases are usually found in hydro-gen evolution. Enzyme activity is easily inactivated by oxygenand CO although the characteristics of inactivation differ in thereduction (evolution) and oxidation (uptake) directions. Whileprokaryotic hydrogenases are usually part of the fermentativemetabolism, [FeFe]-hydrogenases in algae receive reducing equiv-alents at the end of the photosynthetic electron transfer chain viaferredoxin. Therefore, chlorophytatype [FeFe]-hydrogenases havebeen termed “photosynthetic hydrogenases” (see Refs. [44–48]).
Recently, much research progress was made on the structureand function of the hydrogenases by X-ray analysis, spectroscopictechniques, theoretical methods, and model studies (for review seeRefs. [10,11]). [FeFe]-hydrogenases from organisms like Clostrid-ium pasteurianum and acetobutylicum, Desulfovibrio desulfuricans,Megasphaera elsdenii, C. reinhardtii, S. obliquus, Chlamydomonasmoewusii, Chlorococcum submarinum and Chlorella fusca, have beendescribed in detail in Refs. [44–47]. The schematic view of the activecenters of two types of hydrogenases is shown in Fig. 4.
The hydrogenases catalyze the reversible oxidation of molecularhydrogen. Sequence comparisons and biochemical analyses haveclearly shown that [Fe]-hydrogenases and Ni-containing hydro-genases are not related. Three forms of hydrogenases are knownwhich differ in their active site structure (Fig. 5): (i) in the [NiFe]-hydrogenases, the active site contains a Ni ion that is bridged
the active site of the [FeFe]-hydrogenases consists of a 2Fe sub-cluster in which two Fe(CO)(CN) moieties are bridged by a COligand and a dithiol (S–X–S) moeity of unknown composition. The2Fe subcluster is tethered to the protein by a thiolate from con-
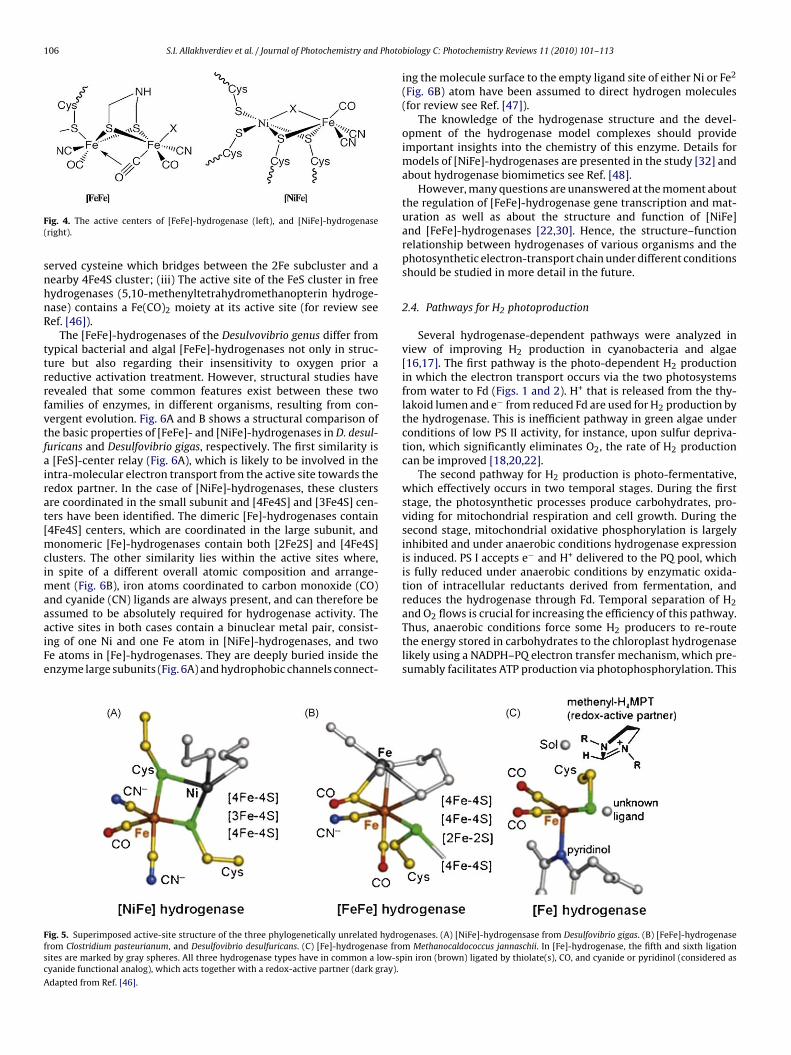
106 S.I. Allakhverdiev et al. / Journal of Photochemistry and Photo
F(
snhnR
ttrrfvtfairat[mcimaaaiFe
Ffsc
A
ig. 4. The active centers of [FeFe]-hydrogenase (left), and [NiFe]-hydrogenaseright).
erved cysteine which bridges between the 2Fe subcluster and aearby 4Fe4S cluster; (iii) The active site of the FeS cluster in freeydrogenases (5,10-methenyltetrahydromethanopterin hydroge-ase) contains a Fe(CO)2 moiety at its active site (for review seeef. [46]).
The [FeFe]-hydrogenases of the Desulvovibrio genus differ fromypical bacterial and algal [FeFe]-hydrogenases not only in struc-ure but also regarding their insensitivity to oxygen prior aeductive activation treatment. However, structural studies haveevealed that some common features exist between these twoamilies of enzymes, in different organisms, resulting from con-ergent evolution. Fig. 6A and B shows a structural comparison ofhe basic properties of [FeFe]- and [NiFe]-hydrogenases in D. desul-uricans and Desulfovibrio gigas, respectively. The first similarity is[FeS]-center relay (Fig. 6A), which is likely to be involved in the
ntra-molecular electron transport from the active site towards theedox partner. In the case of [NiFe]-hydrogenases, these clustersre coordinated in the small subunit and [4Fe4S] and [3Fe4S] cen-ers have been identified. The dimeric [Fe]-hydrogenases contain4Fe4S] centers, which are coordinated in the large subunit, and
onomeric [Fe]-hydrogenases contain both [2Fe2S] and [4Fe4S]lusters. The other similarity lies within the active sites where,n spite of a different overall atomic composition and arrange-
ent (Fig. 6B), iron atoms coordinated to carbon monoxide (CO)nd cyanide (CN) ligands are always present, and can therefore be
ssumed to be absolutely required for hydrogenase activity. Thective sites in both cases contain a binuclear metal pair, consist-ng of one Ni and one Fe atom in [NiFe]-hydrogenases, and twoe atoms in [Fe]-hydrogenases. They are deeply buried inside thenzyme large subunits (Fig. 6A) and hydrophobic channels connect-ig. 5. Superimposed active-site structure of the three phylogenetically unrelated hydrorom Clostridium pasteurianum, and Desulfovibrio desulfuricans. (C) [Fe]-hydrogenase froites are marked by gray spheres. All three hydrogenase types have in common a low-spyanide functional analog), which acts together with a redox-active partner (dark gray).
dapted from Ref. [46].
biology C: Photochemistry Reviews 11 (2010) 101–113
ing the molecule surface to the empty ligand site of either Ni or Fe2
(Fig. 6B) atom have been assumed to direct hydrogen molecules(for review see Ref. [47]).
The knowledge of the hydrogenase structure and the devel-opment of the hydrogenase model complexes should provideimportant insights into the chemistry of this enzyme. Details formodels of [NiFe]-hydrogenases are presented in the study [32] andabout hydrogenase biomimetics see Ref. [48].
However, many questions are unanswered at the moment aboutthe regulation of [FeFe]-hydrogenase gene transcription and mat-uration as well as about the structure and function of [NiFe]and [FeFe]-hydrogenases [22,30]. Hence, the structure–functionrelationship between hydrogenases of various organisms and thephotosynthetic electron-transport chain under different conditionsshould be studied in more detail in the future.
2.4. Pathways for H2 photoproduction
Several hydrogenase-dependent pathways were analyzed inview of improving H2 production in cyanobacteria and algae[16,17]. The first pathway is the photo-dependent H2 productionin which the electron transport occurs via the two photosystemsfrom water to Fd (Figs. 1 and 2). H+ that is released from the thy-lakoid lumen and e− from reduced Fd are used for H2 production bythe hydrogenase. This is inefficient pathway in green algae underconditions of low PS II activity, for instance, upon sulfur depriva-tion, which significantly eliminates O2, the rate of H2 productioncan be improved [18,20,22].
The second pathway for H2 production is photo-fermentative,which effectively occurs in two temporal stages. During the firststage, the photosynthetic processes produce carbohydrates, pro-viding for mitochondrial respiration and cell growth. During thesecond stage, mitochondrial oxidative phosphorylation is largelyinhibited and under anaerobic conditions hydrogenase expressionis induced. PS I accepts e− and H+ delivered to the PQ pool, whichis fully reduced under anaerobic conditions by enzymatic oxida-tion of intracellular reductants derived from fermentation, andreduces the hydrogenase through Fd. Temporal separation of H2
and O2 flows is crucial for increasing the efficiency of this pathway.Thus, anaerobic conditions force some H2 producers to re-routethe energy stored in carbohydrates to the chloroplast hydrogenaselikely using a NADPH–PQ electron transfer mechanism, which pre-sumably facilitates ATP production via photophosphorylation. Thisgenases. (A) [NiFe]-hydrogensase from Desulfovibrio gigas. (B) [FeFe]-hydrogenasem Methanocaldococcus jannaschii. In [Fe]-hydrogenase, the fifth and sixth ligationin iron (brown) ligated by thiolate(s), CO, and cyanide or pyridinol (considered as

S.I. Allakhverdiev et al. / Journal of Photochemistry and Photobiology C: Photochemistry Reviews 11 (2010) 101–113 107
Fig. 6. (A) Structures of (1) Desulfovibrio desulfuricans [FeFe]-hydrogenase and (2) Desulfovibrio gigas [NiFe]-hydrogenase. The small subunits are in green and the largesubunits in orange. Carbon atoms are black, iron atoms are gray, sulfur atoms are yellow, the nickel atom is light blue, the nitrogen atoms are dark blue and oxygen atomsare red. (B) Structure of the (1) [FeFe] and (2) [NiFe]-hydrogenase active centers. The color code is the same as in (A). In the case of the [FeFe] center, each proximal (Fe1) anddistal (Fe2) iron atom is coordinated by one cyanide (CN) molecule, one carbon monoxide (CO) molecule and two bridging thiolates from the 1,3-propanedithiol molecule.The proximal iron atom possesses one additional bridging cysteine thiolate ligand, which is also involved in the coordination of the proximal iron sulfur center. In the caseo eine ta , the t
A
td
(aric
2
Ii[tHneIdni
Cooot[
f the [NiFe] center, the nickel atom is coordinated by two simple coordinating cysts an oxygen in the enzyme oxidized state. Two CN molecules and one CO molecule
dapted from Ref. [47].
wo stages pathway seems to be the most effective for H2 biopro-uction [17].
The third pathway uses the nitrogenase of cyanobacteriasee Refs. [6,10,11,15–17,20–25,44]). Here, electrons and protonsre delivered from photosynthesis. However, this pathway thatequires the largest numbers of photons results in lower efficiencyn comparison with other pathways and hence, makes it economi-ally impractical.
.5. Separation of O2 and H2 production
The majority of hydrogenases are very sensitive to O2 [4,22].t is an important issue, and the pathways for suppressing O2 andmproving H2 production yield were discussed in several reviews16,17,20,49]. Due to the fact that hydrogenase is hypersensitiveo oxygen and is located in the chloroplast where PS II releases O2,2 production rate is usually low. There are several natural mecha-isms that could be used for lowering the concentration of O2: thenhancement of respiration, photochemical reduction of O2 by PS, and reversible inactivation of O2-evolution in PS II [17]. Modularevices were also constructed, which consist only of componentsecessary for photo-induced hydrogen production instead of whole
ntact cells [50].It is known that the lack of sulfur from the growth medium of
. reinhardtii causes a specific but reversible decline in the rate
f oxygenic photosynthesis; however, it does not affect the ratef mitochondrial respiration [18,51]. This led to an inactivationf PS II so that cellular respiration draws to anaerobiosis, induc-ion of hydrogenase, and sustained hydrogen evolution in the light18,22,24,52]. Thus, sulfur deprivation is used as a physiologicalhiolates, two bridging cysteine thiolates and one bridging atom that was identifiedwo bridging cysteine thiolates and the bridging atom coordinate the iron atom.
switch, to switch over aerobic photosynthetic growth mode toan anaerobic H2-producing mode. Under sulfur-deprivation con-ditions the rate of turnover of D1, one of the main protein subunitof the PS II RC, decreases due to the lack of availability of sulfury-lated amino acid residues [34]. However, it is now clear that PS II isnot completely inactivated under sulfur deprivation, and the elec-trons reducing protons at the hydrogenase are still mainly comingfrom water, with concomitant oxygen evolution [4].
One of the most effective pathways for the generation of H2 isindirect biophotolysis that intends to eliminate the oxygen sensi-tivity of the hydrogenases by separating the hydrogen-producingreactions from the oxygen-evolving ones [30]. This is usually imag-ined as a system that grows algae in inexpensive open ponds,where they evolve oxygen and store carbohydrates. The organ-isms are then harvested and placed in an anaerobic reactorwhere they inactivate PS II and induce hydrogenase. Illumina-tion oxidizes the stored carbohydrates and lipids, and produceshydrogen, either directly or after an anaerobic dark fermentation[53]. Cyanobacteria developed several pathways to protect thenitrogenase from inactivation by oxygen: (a) heterocyst-containingcyanobacteria physically separate oxygen evolution from nitro-genase activity by segregating oxygenic photosynthetic activityin vegetative cells and nitrogenase activity in heterocystis withreduced oxygen-permeability [54], and (b) non-heterocystouscyanobacteria separate oxygen evolution from nitrogenase activ-
ity by performing these functions during light and dark periodsrespectively [55].Green algae such as C. reinhardtii synthesize an [FeFe]-hydrogenase that is highly active in hydrogen evolution. However,the extreme sensitivity of [FeFe]-hydrogenases to oxygen presents

1 Photo
aamCstttuoinedtctt(a
2e
2
tmr[gAsys
atodecfl
2
atotlslwiuofiotbwwbt
08 S.I. Allakhverdiev et al. / Journal of Photochemistry and
major challenge for exploiting these organisms to achieve sustain-ble photosynthetic hydrogen production [39]. In this study, theechanism of inactivation of [FeFe]-hydrogenase CrHydA1 from
. reinhardtii by oxygen has been investigated. X-ray absorptionpectroscopy shows that reaction with oxygen results in destruc-ion of the [4Fe4S] domain of the active site H-cluster while leavinghe di-iron domain (2FeH) essentially intact. By protein film elec-rochemistry the authors determined the order of events leadingp to this destruction. Carbon monoxide, a competitive inhibitorf CrHydA1 which binds to an Fe atom of the 2FeH domain ands otherwise not known to attack FeS clusters in proteins, reactsearly two orders of magnitude faster than oxygen and protects thenzyme against oxygen damage. These results therefore show thatestruction of the [4Fe4S] cluster is initiated by binding and reduc-ion of oxygen at the di-iron domain—a key step that is blocked byarbon monoxide. The relatively slow attack by oxygen comparedo carbon monoxide suggests that a very high level of discrimina-ion can be achieved by subtle factors such as electronic effectsspecific orbital overlap requirements) and steric constraints at thective site [39].
.6. Pathways of increasing light to H2 photon conversionfficiency
.6.1. Reduced antenna size and increased PQ poolThe efficiency of light utilization is one of the important factors
hat determine the H2 production yield. Enhanced H2 productionay be achieved by engineering the antenna size to suppress fluo-
escence and heat dissipation that causes a reduction in efficiency17,20,56]. Genes that regulate the Chl antenna size in the modelreen alga C. reinhardtii were identified and characterized [20].nalysis of the tla1 and tlaX mutants with decreased Chl antennaize in comparison with the wild type (WT) demonstrated higherields of photosynthesis in microalgae with a truncated Chl antennaize.
An increased PQ pool capacity and strong proton buffer capacitylso can improve light utilization since it is able to accelerate elec-ron transport to PS I, slow down the back reactions in PS II, andxidize reducing equivalents stored during CO2 fixation. Besides,own regulation of competing pathways can redirect the fluxes oflectrons via PS I and ferredoxin into hydrogenase (Fig. 1). Suchompetitive pathways, for example, can be blocking cyclic electronow from PS I into PQ pool [57].
.6.2. Immobilization of microbial culturesThe reported rates of H2 production by sulfur-deprived cultures
re still far below the maximum potential rate of H2 photoproduc-ion for an algal system [58] mainly due to the partial inactivationf photosynthetic water oxidation [23]. However, it was shownhat sulfur-deprived cultures of C. reinhardtii cells can be immobi-ized by inexpensive matrices and sulfur-deprivation stress can beuccessfully applied to immobilized algal cells. The improved andonger-term H2 photoproduction by immobilized green alga cells
as demonstrated in several studies [22–25]. On the other hand,mmobilized cyanobacteria can produce H2 at much higher vol-metric rates than suspension cultures [59]. Cyanobacterial cellsf Synechocystis sp. PCC 6803 were also encapsulated success-ully in silica sol–gel using glycerol and polyethylene glycol toncrease gel porosity in order to reduce the immobilization andsmotic stress [60]. These preparations also presented H2 produc-ion levels comparable or above the free cells. Hydrogen evolution
y Rhodobacter sphaeroides cells immobilized in calcium alginateas enhanced by 2–3-fold compare to the free cells using wasteater as energy source [61]. Several techniques and matrices cane used to immobilize microalgal species that greatly improveheir stability. The same techniques applied for the preparation of
biology C: Photochemistry Reviews 11 (2010) 101–113
biosensor devices may be applied for H2 evolution [62,63]. Goodexamples of immobilization techniques are the co-reticulation inan albumin–glutaraldehyde crosslinked matrix [64] or the phys-ical entrapment in photo-crosslinkable polyvinylalcohol polymer[62,64]. Rouillon et al. [65] showed that the immobilization ofthe mixture of chloroplasts and photosynthetic membranes in thePVA-SbQ increased considerably the storage stability. After 90-day storage at +4 ◦C in the dark and in a dry state, the polymerswith 1700 and 3500 (betaine form) as degree of polymerisa-tion retained 20% of their initial activity and the polymers 2300and 3500, 10% and 12%, respectively, whereas the membranesentrapped in polymers 600 and 1200 were not active. The stabil-ity of the thylakoids entrapped in PVA-SbQ 2300 after 24 h dryingat +4 ◦C and under storage at −18 ◦C was checked. After 427-day storage, the membranes still retained 20% activity [65]. Theprocedure for such immobilization is very simple and is outlinein Fig. 7.
2.6.3. The use of mimics of water oxidation systemsAnother approach to overcome the partial inactivation of the
photosynthetic water oxidation system leading to low efficiencyand instability of H2 photoproduction is the use of mimics of thenatural Mn-cluster. It is well known that the water oxidation com-plex is composed of a special tetra manganese cluster forming aMn4Ca complex (Fig. 8) which is very important in photosyntheticoxygen evolution. However, the oxygen-evolving center of PS II isnot suitable for engineering an application such as H2 photopro-duction due to its limited stability. It is believed that performinga directed molecular design and broad synthesis of different arti-ficial metal–organic complexes with different ligand spheres andmatrices that mimic the natural Mn-cluster of PS II might avoidthe problems associated with low H2 photoproduction rates andscale-up of bioreactors [30,66–76]. Such systems would have moreversatility and might split water with sunlight and produce hydro-gen and oxygen, with a high efficiency and long-term stability.Many synthetic Mn complexes with different ligands have beensynthesized and examined with various degrees of restoration ofthe original function of PS II, including oxygen evolution in Mn-depleted PS II complexes [66–76]. Some of them even producedhydrogen peroxide [73]. If protons from a water oxidation com-plex can be captured and reduced to H2 in the above-reconstructedphotosynthetic systems, it could provide an interesting approachfor future developments. The following step could be thecombination of an artificial Mn-containing water oxidation com-plex with a hydrogenase system stabilized by inexpensivematrices.
2.6.4. Enhanced resistance to environmental stress conditionsTo increase productivity, algal cells must be maintained in a
healthy, active state during H2 production for a longer period oftime. The tolerance of cell cultures to environmental stresses, firstof all, to photoinhibition, salt stress and high temperatures is nec-essary for sustainable photosynthesis and, hence, H2 production[10,11,16,17]. The efficiency of the recovery of PS II, especially theoxygen-evolving complex (OEC), from damage induced by highlight or environmental stress is one of the key factors in photo-synthetic resistance [77–84]. Other factors might be engeneeringcells with higher antioxidant activity, by antenna adjasment, etc.
Besides optimal light conditions and sufficient ATP, other impor-tant factors are the synthesis of different low-molecular weight
protective compounds such as glycinebetaine, bioflavonoids,carotenoids, stress proteins, PS II stabilizing factors (extrinsic pro-teins of PS II, the availability of bicarbonate and intracellular Mn2+),as well as improvement of thylakoid membrane stress resistanceand CO2 consumption conditions [82–93].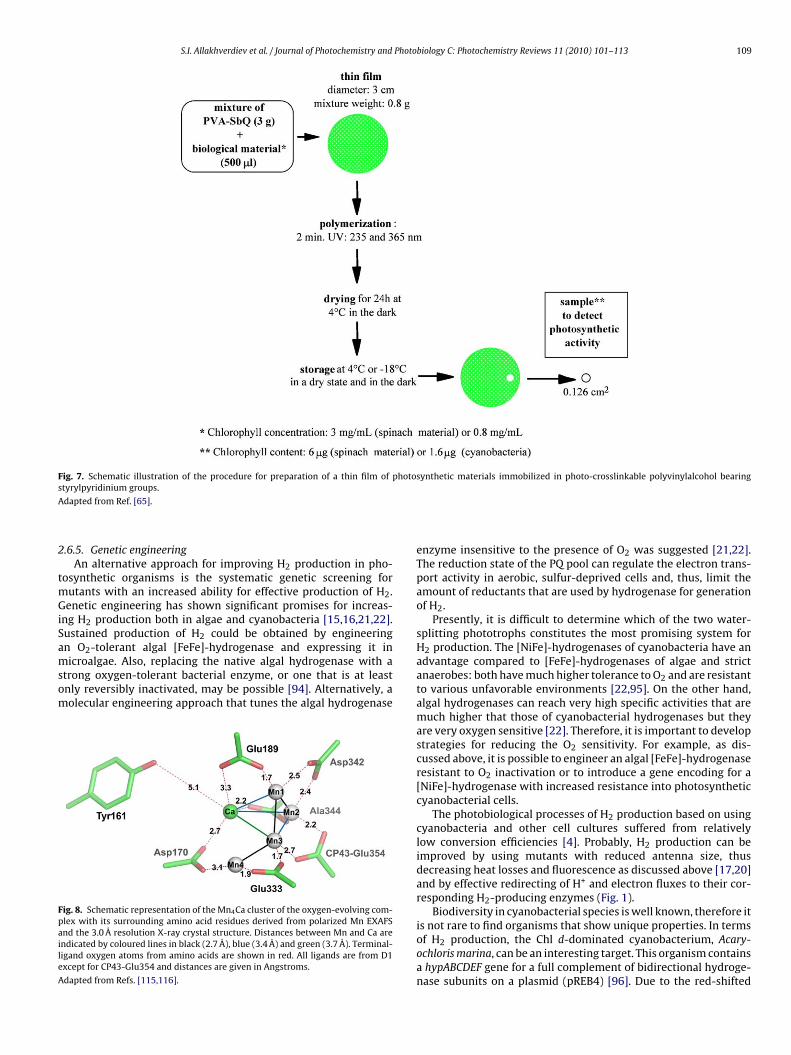
S.I. Allakhverdiev et al. / Journal of Photochemistry and Photobiology C: Photochemistry Reviews 11 (2010) 101–113 109
Fig. 7. Schematic illustration of the procedure for preparation of a thin film of photosynthetic materials immobilized in photo-crosslinkable polyvinylalcohol bearings
A
2
tmGiSamsom
Fpaile
A
tyrylpyridinium groups.
dapted from Ref. [65].
.6.5. Genetic engineeringAn alternative approach for improving H2 production in pho-
osynthetic organisms is the systematic genetic screening forutants with an increased ability for effective production of H2.enetic engineering has shown significant promises for increas-
ng H2 production both in algae and cyanobacteria [15,16,21,22].ustained production of H2 could be obtained by engineering
n O2-tolerant algal [FeFe]-hydrogenase and expressing it inicroalgae. Also, replacing the native algal hydrogenase with atrong oxygen-tolerant bacterial enzyme, or one that is at leastnly reversibly inactivated, may be possible [94]. Alternatively, aolecular engineering approach that tunes the algal hydrogenase
ig. 8. Schematic representation of the Mn4Ca cluster of the oxygen-evolving com-lex with its surrounding amino acid residues derived from polarized Mn EXAFSnd the 3.0 A resolution X-ray crystal structure. Distances between Mn and Ca arendicated by coloured lines in black (2.7 A), blue (3.4 A) and green (3.7 A). Terminal-igand oxygen atoms from amino acids are shown in red. All ligands are from D1xcept for CP43-Glu354 and distances are given in Angstroms.
dapted from Refs. [115,116].
enzyme insensitive to the presence of O2 was suggested [21,22].The reduction state of the PQ pool can regulate the electron trans-port activity in aerobic, sulfur-deprived cells and, thus, limit theamount of reductants that are used by hydrogenase for generationof H2.
Presently, it is difficult to determine which of the two water-splitting phototrophs constitutes the most promising system forH2 production. The [NiFe]-hydrogenases of cyanobacteria have anadvantage compared to [FeFe]-hydrogenases of algae and strictanaerobes: both have much higher tolerance to O2 and are resistantto various unfavorable environments [22,95]. On the other hand,algal hydrogenases can reach very high specific activities that aremuch higher that those of cyanobacterial hydrogenases but theyare very oxygen sensitive [22]. Therefore, it is important to developstrategies for reducing the O2 sensitivity. For example, as dis-cussed above, it is possible to engineer an algal [FeFe]-hydrogenaseresistant to O2 inactivation or to introduce a gene encoding for a[NiFe]-hydrogenase with increased resistance into photosyntheticcyanobacterial cells.
The photobiological processes of H2 production based on usingcyanobacteria and other cell cultures suffered from relativelylow conversion efficiencies [4]. Probably, H2 production can beimproved by using mutants with reduced antenna size, thusdecreasing heat losses and fluorescence as discussed above [17,20]and by effective redirecting of H+ and electron fluxes to their cor-responding H2-producing enzymes (Fig. 1).
Biodiversity in cyanobacterial species is well known, therefore it
is not rare to find organisms that show unique properties. In termsof H2 production, the Chl d-dominated cyanobacterium, Acary-ochloris marina, can be an interesting target. This organism containsa hypABCDEF gene for a full complement of bidirectional hydroge-nase subunits on a plasmid (pREB4) [96]. Due to the red-shifted
1 Photo
auos
shtqrtiDimiItatampeP
2
rtmwtcTt
IdPadece3ttot(oIsppp(iroo
pcs
10 S.I. Allakhverdiev et al. / Journal of Photochemistry and
bsorption maximum of Chl d, this organism can utilize the lightp to a near infra-red region. Due to the unique redox-potentialsf its electron transfer components in photosynthesis [97,98] thispecies might be referred to as a natural mutant in this aspect.
In the study of Bernát et al. [99] three Synechocystis PCC 6803trains with different levels of phycobilisome antenna-deficiencyave been investigated for their impact on photosynthetic electronransport and response to environmental factors (i.e. light-quality, -uantity and composition of growth media). Oxygen yield and P700eduction kinetic measurements showed enhanced linear electronransport rates, especially under photoautotrophic conditions, withmpaired antenna size, starting from wild type (full antenna) overapcE- (phycobilisomes functionally dissociated) and Olive (lack-
ng phycocyanin) up to the PAL (phenylalanine ammonium liase)utant (lacking the whole phycobilisome). Antenna-deficiency
nvolves higher linear electron transport rates by tuning the PS II/PSratio from 1:5 in wild type up to 1:1 in the PAL mutant. While stateransitions were observed only in the wild type and Olive mutant,further 30% increase in the PS II/PS I ratio was achieved in all
he strains by long-term adaptation to far red light (720 nm).Theuthors conclude that the highest rates can be expected ifutants deficient in phycobilisome antennas are grown under
hotoautotrophic conditions in combination with uncoupling oflectron transport and an illumination which excites preferablyS I.
.6.6. Role of photosystems in H2 photoproductionBesides the activity of PS I, at least some activity of PS II is
equired to sustain H2 photoproduction. Indeed, recent observa-ions using electron transfer inhibitors indicated that the vast
ajority of the electrons driving H2 production originate fromater oxidation [100]. The progressive impairment of PS II pho-
ochemical activity in sulfur-deprived C. reinhardtii D1-R323oincided with the progressive decrease in O2-evolution [25].hese mutants exhibited lower H2 yield compared to the wildype.
Another interesting aspect is the direct evolution of H2 by PSI. Early studies have indeed demonstrated that H2 can be pro-uced from PS II under certain conditions, both in mutants lackingS I [101] and in preparations of PS II particles [102]. Wild typend mutants of the green alga C. reinhardtii lacking PS II pro-uced H2 with high efficiency. However, a mutant lacking PSIxhibited low efficiency in H2 production [101]. Conversely, sub-hloroplast preparations enriched in PSII in the presence of thelectron donor TMPD exhibited higher H2 evolution rates (up to0 nmol mg Chl−1 h−1) than preparations enriched in PS I underhe same conditions [102]. It is interesting that H2 photoproduc-ion in PS II preparations was stimulated by 10-fold after removalf manganese (by tris-treatment) from the OEC and this reac-ion was suppressed by DCMU (5 �M), dinoseb (10 �M), atrazine10 �M), o-phenanthroline (10 �M) or CO (0.4%) [102]. The datan the suppression of H2 evolution by well known inhibitors of PSI (DCMU, dinoseb, atrazine) proved that H2 photoproduction wasensitized by the reaction center of PS II. Becides, Mn2+ stimulatedhotoreduction of NADP+ was demonstrated in Mn-depleted PSIIreparations [103]. It has been shown that the mid-point redox-otential of the intermediate electron acceptor of PS II, pheophytinPheo) a, is less than −0.6 V [104,105]. Theoretically, this potentials sufficient to allow PS II to photoreduce electron acceptors withedox-potential of ca. −0.4 V (Fd, NADP+, methylviologen, benzylvi-logen, NO2
−, NO3−, SO4
2−, etc.) typical for PS I, and photoreduction
f H2 (−0.42 V) [103].These results demonstrate that theoretically, isolated PS II canroduce H2 under sunlight. The detailed characterization and appli-ation of this unique approach of H2 photoproduction by PS IIhould be the subject of further research in near future.
biology C: Photochemistry Reviews 11 (2010) 101–113
3. Anoxygenic organisms
Anoxygenic photosynthetic bacteria can use the sun energy togenerate ATP, substantial amounts of which are needed to drivethe enzyme nitrogenase that catalyzes the reaction 2H+ + 2e− → H2.The nitrogenase-encoding gene responsible for H2 production inpurple non-sulfur and green sulfur photosynthetic bacteria isexpressed in the light under anaerobic conditions and in theabsence of ammonium.
Electrons for the parallel reduction of N2 and H+ bythe nitrogenase are provided upon the catabolism of smallorganic acids or carbohydrates, according to the reaction:N2 + 8e− + 8H + + 16ATP → 2NH3 + H2 + 16ADP + 16Pi. Both NH3and H2 are products of the nitrogenase enzyme. Under conditionsof N2 limitation, the H2/NH3 product ratio increases. Approachesusing molecular genetics allowed improving the H2 productionefficiency in purple non-sulfur and green sulfur bacteria. Suchapproaches include, especially, optimization of culture parametersand suppression of electron sink pathways alternative to H2production [106].
At present, the most profitable approach from both economicaland ecological point of view in using carbohydrate-rich wastewaterobtained as a result of food processing for anoxygenic photosyn-thetic H2 production [106,107]. Pathways of H2 production andtheir relationship with photosynthesis in anoxygenic organismshave been reviewed in the literature [4,10,11,16,26,31]. Thus, pho-tosynthetic hydrogen production could play a leading role in directsolar conversion, but research is needed to improve its efficiency,capacity, and reliability.
4. Conclusion
Photosynthetic microorganisms using sunlight as the princi-pal driving force are capable of effective hydrogen production[4,10,11,16,30]. The actual photoproduction of hydrogen will haveto be carried out in a sealed photobioreactor and also requirescareful reactor designs for the substantial improvement of hydro-gen production rates and yields [10,11,16,108–111]. A prerequisitechallenge is to improve current systems at the biochemical levelso that they can clearly generate hydrogen at a rate and efficiency,which surpasses the 11% energy efficiency indicated in photoelec-trical systems [10,11,112–115].
The optimization of photosynthetic organisms for effectiveH2 production by genetic modification is one of the promisingpathways. Further research will focus on increasing the O2-tolerance and turnover frequency of [FeFe]-hydrogenases, theuse of immobilized microbial cultures as well as engineeringmore active nitrogenase. Improved light access, nutrient medium,enhanced resistance to environmental stress conditions and costof bioreactors should also be considered for increased hydrogenphotoproduction efficiency.
Acknowledgements
This work was supported, in part, by grants from theRussian Foundation for Basic Research (Nos. 08-04-00241, 09-04-01074, 09-04-91219-CT), from the Molecular and Cell BiologyProgram of the Russian Academy of Sciences; by Japanes Soci-ety for the Promotion of Science (JSPS) Invitation Fellowshipfor Research in Japan, to SIA; by a Grant-in-Aid for Cre-
ative Scientific Research (No. 17GS0314) from the JSPS and byScientific Research on Priority Areas “Comparative Genomics”(Nos: 17018022 and 18017016) from the Ministry of Educa-tion, Sports, Culture, Science, and Technology, Japan, to MM;VT and SR acknowledge the NUSNNI, Singapore. RC was sup-
Photo
pC
R
S.I. Allakhverdiev et al. / Journal of Photochemistry and
orted by Natural Sciences and Engineering Research Council ofanada.
eferences
[1] N.S. Lewis, D.G. Nocera, Powering the planet: chemical challengesin solar energy utilization, Proc. Natl. Acad. Sci. U.S.A. 103 (2006)15729–15735.
[2] S. Solomon, D. Qin, M. Manning, Climate change, in: S. Solomon, D. Qin, M.Manning (Eds.), The Physical Science Basis. Contribution of Working Group Ito the Fourth Assessment Report of the Intergovernmental Panel on ClimateChange, IPCC, 2007, pp. 64–109.
[3] P. Hoffmann, Tomorrow’s Energy. Hydrogen, Fuel Cells and Prospects forCleaner Planet, The MIT Press, Cambridge, UK, 2002, p. 224.
[4] R.C. Prince, H.S. Kheshgi, The photobiological production of hydrogen: poten-tial efficiency and effectiveness as a renewable fuel, Crit. Rev. Microbiol. 31(2005) 19–31.
[5] R.E. Blankenship, Molecular Mechanisms of Photosynthesis, Blackwell Sci-ence, Oxford, UK, 2002, p. 321.
[6] W.S. Chow, Photosynthesis: from natural towards artificial, J. Biol. Phys. 29(2003) 447–459.
[7] D.A. LaVan, J.N. Cha, Approaches for biological and biomimetic energy con-version, Proc. Natl. Acad. Sci. U.S.A. 103 (2006) 5251–5255.
[8] S.I. Allakhverdiev, Recent perspectives of photosystem II: structure, functionand dynamics, Photosynth. Res. 98 (2008) 1–5.
[9] S.I. Allakhverdiev, J.J. Casal, T. Nagata, Photosynthesis from molecular per-spectives: towards future energy production, Photochem. Photobiol. Sci. 8(2009) 137–138.
[10] S.I. Allakhverdiev, V.D. Kreslavski, V. Thavasi, S.K. Zharmukhamedov, V.V.Klimov, T. Nagata, H. Nishihara, S. Ramakrishna, Hydrogen photoproductionby use of photosynthetic organisms and biomimetic systems, Photochem.Photobiol. Sci. 8 (2009) 148–156.
[11] S.I. Allakhverdiev, V.D. Kreslavski, V. Thavasi, S.K. Zharmukhamedov, V.V.Klimov, S. Ramakrishna, H. Nishihara, M. Mimuro, R. Carpentier, T. Nagata,Photosynthetic energy conversion: hydrogen photoproduction by naturaland biomimetic systems, in: A. Mukherjee (Ed.), Biomimetics, Learning fromNature, In-Tech (intechweb.org), Vukovar, Croatia, 2010, pp. 49–76.
[12] P. Joliot, A. Joliot, Cyclic electron transfer in plant leaf, Proc. Natl. Acad. Sci.U.S.A. 99 (2002) 10209–10214.
[13] N.G. Bukhov, R. Carpentier, Alternative PSI-driven electron transport routes:mechanisms and functions, Photosynth. Res. 82 (2004) 17–33.
[14] S. Govindachary, C. Bigras, J. Harnois, D. Joly, R. Carpentier, Changes in themode of electron flow to photosystem I following chilling-induced photoin-hibition in a C3 plant, Cucumis sativus L., Photosynth. Res. 94 (2007) 333–345.
[15] O. Kruse, J. Rupprecht, K.P. Bader, S. Thomas-Hall, P.M. Schenk, G. Finazzi,B. Hankamer, Improved photobiological H2 production in engineered greenalgal cells, J. Biol. Chem. 280 (2005) 34170–34177.
[16] J. Rupprecht, B. Hankamer, J.H. Mussgnug, G. Ananyev, G.C. Dismukes, O.Kruse, Perspectives and advances of biological H2 production in microorgan-isms, Appl. Microbiol. Biotechnol. 72 (2006) 442–449.
[17] O. Kruse, J. Rupprecht, J.H. Mussgnug, G.C. Dismukes, B. Hankamer, Pho-tosynthesis: a blue print for energy capture and conversion technologies,Photochem. Photobiol. Sci. 4 (2005) 957–970.
[18] A. Melis, L. Zhang, M. Forestier, M.L. Ghirardi, M. Seibert, Sustained photo-biological hydrogen gas production upon reversible inactivation of oxygenevolution in the green alga Chlamydomonas reinhardtii, Plant Physiol. 122(2000) 127–133.
[19] A.A. Tzygankov, S.N. Kosourov, I.V. Tolstygina, M.L. Ghirardi, M. Seibert,Hydrogen production by sulfur-limited Chlamydomonas reinhardtii underphotoautotrophic conditions, Int. J. Hydrogen Energy 31 (2006) 1574–1584.
[20] A. Melis, Bioengineering of green algae to enhance photosynthesis and hydro-gen production, in: A.F. Colling, C. Critchley (Eds.), Artificial Photosynthesis:From Basic Biology to Industrial Application, Wiley-Verlag and Co. KGaA,Weinheim, 2005, pp. 229–240.
[21] M.L. Ghirardi, Development of algal systems for hydrogen photoproduc-tion: addressing the hydrogenise oxygen sensitivity problem, in: A.F.Colling, C. Critchley (Eds.), Artificial Photosynthesis: From Basic Biology toIndustrial Application, Wiley-Verlag and Co. KGaA, Weinheim, 2005, pp.213–227.
[22] M.L. Ghirardi, M.C. Posewitz, P.-C. Maness, A. Dubini, J. Yu, M. Seibert,Hydrogenase and hydrogen photoproduction in oxygenic photosyntheticorganisms, Annu. Rev. Plant Biol. 58 (2007) 71–91.
[23] T.V. Laurinavichene, A.S. Fedorov, M.L. Ghirardi, M. Seibert, A.A. Tsygankov,Demonstration of sustained hydrogen photoproduction by immobilized,sulfur-deprived Chlamydomonas reinhardtii cells, Int. J. Hydrogen Energy 31(2006) 659–667.
[24] T.V. Laurinavichene, S.N. Kosourov, M.L. Ghirardi, M. Seibert, A.A. Tsygankov,Prolongation of H2 photoproduction by immunobilized, sulfur-limitedChlamydomonas reinhardtii cultures, J. Biotechnol. 134 (2008) 275–277.
[25] V.V. Makarova, S. Kosourov, T.E. Krendeleva, B.K. Semin, G.P. Kukarskikh, A.B.Rubin, R.T. Sayre, M.L. Ghirardi, M. Seibert, Photoproduction of hydrogen bysulfur-deprived C. reinhardtii mutants with impaired photosystem II photo-chemical activity, Photosynth. Res. 94 (2007) 79–89.
[26] E.N. Kondratieva, I.N. Gogotov, Molecular Hydrogen in Microbial Metabolism,Nauka, Moscow, 1981, p. 344.
biology C: Photochemistry Reviews 11 (2010) 101–113 111
[27] J.P. McEvoy, G.W. Brudvig, Water-splitting chemistry of photosystem II, Chem.Rev. 106 (2006) 4455–4483.
[28] V.A. Boichenko, E. Greenbaum, M. Seibert, Hydrogen production by photo-synthetic microorganisms, in: M.D. Archer, J. Barber (Eds.), Photoconversionof Solar Energy, Molecular to Global Photosynthesis, vol. 2, Imperial CollegePress, London, UK, 2004, pp. 397–452.
[29] D. Dutta, D. De, S. Chaudhuri, S.K. Bhattacharya, Hydrogen production bycyanobacteria, Microb. Cell. Fact. 4 (2005) 36–106.
[30] W. Lubitz, E.J. Reijerse, J. Messinger, Solar water-splitting into H2 and O2
design principles of photosystem II and dehydrogenases, Energy Environ. Sci.1 (2008) 15–31.
[31] L. Florin, A. Tsokoglou, T. Happe, A novel type of iron hydrogenase in the greenalga Scenedesmus obliquus is linked to the photosynthetic electron transportchain, J. Biol. Chem. 276 (2001) 6125–6132.
[32] P.M. Vignais, B. Billoud, Occurrence, classification, and biological function ofhydrogenases: an overview, Chem. Rev. 107 (2007) 4206–4272.
[33] J.P. Zehr, B.D. Jenkins, S.M. Short, G.F. Steward, Nitrogenase gene diversityand microbial community structure: a cross-system comparison, Environ.Microbiol. 5 (2003) 539–554.
[34] P. Tamagnini, J.L. Costa, L. Almeida, M.J. Oliveira, R. Salema, P. Lindblad, Diver-sity of cyanobacterial hydrogenases, a molecular approach, Curr. Microbiol.40 (2000) 356–361.
[35] P. Tamagnini, R. Axelsson, P. Lindberg, F. Oxelfelt, R. Wunschiers, P. Lindblad,Hydrogenases and hydrogen metabolism of cyanobacteria, Microbiol. Mol.Biol. Rev. 66 (2002) 1–20.
[36] G. Boison, H. Bothe, A. Hansel, P. Lindblad, Evidence against a common useof the diaphorase subunits by the bidirectional hydrogenase and by therespiratory complex I in cyanobacteria, FEMS Microbiol. Lett. 174 (1999)159–165.
[37] S.T. Stripp, G. Goldet, C. Brandmayr, O. Sanganas, K.A. Vincent, M. Hau-mann, F.A. Armstrong, T. Happe, How oxygen attacks [FeFe] hydrogenasesfrom photosynthetic organisms, Proc. Natl. Acad. Sci. U.S.A. 106 (2009)17331–17336.
[38] Y. Nicolet, C. Piras, P. Legrand, C. Hatchikian, J.C. Fontecilla-Camps, Desulfovib-rio desulfuricans iron hydrogenase: the structure shows unusual coordinationto an active site Fe binuclear center, Structure 7 (1999) 13–23.
[39] S. Stripp, O. Sanganas, T. Happe, M. Haumann, The structure of the active siteH-cluster of [FeFe] hydrogenase from the green alga Chlamydomonas rein-hardtii studied by X-ray absorption spectroscopy, Biochemistry 48 (2009)5042–5049.
[40] J.C. Fontecilla-Camps, A. Volbeda, C. Cavazza, Y. Nicolet, Structure and func-tion relationships of [NiFe]- and [FeFe]-hydrogenases, Chem. Rev. 107 (2007)4273–4303.
[41] P.M. Vignais, Hydrogenases and H+-reduction in primary energy conserva-tion, Results Probl. Cell Differ. 45 (2008) 223–252.
[42] F.A. Armstrong, N.A. Belsey, J.A. Cracknell, G. Goldet, A. Parkin, E. Reisner,K.A. Vincent, A.F. Wait, Dynamic electrochemical investigations of hydrogenoxidation and production by enzymes and implications for future technology,Chem. Soc. Rev. 38 (2009) 36–51.
[43] B.E. Barton, T.B. Rauchfuss, Terminal hydride in [FeFe]-hydrogenase modelhas lower potential for H2 production than the isomeric bridging hydride,Inorg. Chem. 47 (2008) 2261–2263.
[44] A. Melis, T. Happe, Hydrogen production. Green algae as a source of energy,Plant Physiol. 127 (2001) 740–748.
[45] S.T. Stripp, T. Happe, How algae produce hydrogen—news from the photosyn-thetic hydrogenase, Dalton Trans. 45 (2009) 9960–9969.
[46] S. Shima, O. Pilak, S. Vogt, M. Schick, M.S. Stagni, W. Meyer-Klaucke, E.Warkentin, R.K. Thauer, U. Ermler, The crystal structure of [Fe]-hydrogenasereveals the geometry of the active site, Science 321 (2008) 572–575.
[47] L. Casalot, M. Rousset, Maturation of the [NiFe] hydrogenases, Trends Micro-biol. 9 (2001) 228–237.
[48] A.M. Kluwer, R. Kapre, F. Hartl, M. Lutz, A.L. Spek, A.M. Brouwer, P.W.N.M.van Leeuwen, J.N.H. Reek, Self-assembled biomimetic [2Fe2S]-hydrogenase-based photocatalyst for molecular hydrogen evolution, Proc. Natl. Acad. Sci.U.S.A. 106 (2009) 10460–10465.
[49] L. Zhang, P. Happe, A. Melis, Biochemical and morphological characterizationof sulfur-deprived and H2-producing Chlamydomonas reinhardtii (green alga),Planta 214 (2002) 552–561.
[50] S.-O. Wenk, D.-J. Quan, T. Wakayama, C. Nakamura, N. Zorin, M. Rögner, J.Miyake, Biomolecular device for photoinduced hydrogen production, Int. J.Hydrogen Energy 27 (2002) 1489–1493.
[51] D.D. Wykoff, J.P. Davies, A. Melis, A.R. Grossman, The regulation of photo-synthetic electron transport during nutrient deprivation in Chlamydomonasreinhardtii, Plant Physiol. 117 (1998) 129–139.
[52] T.V. Laurinavichene, I.V. Tolstygina, R.R. Galiulina, M.L. Ghirardi, M. Seibert,A.A. Tsygankov, Different methods to deprive Chlamydomonas reinhardtii cul-tures of sulfur for subsequent hydrogen photoproduction, Int. J. HydrogenEnergy 27 (2002) 1245–1249.
[53] P. Hallenbeck, J.R. Benemann, Biological hydrogen production: fundamentalsand limiting processes, Int. J. Hydrogen Energy 27 (2002) 1185–1193.
[54] P. Fay, Oxygen relations of nitrogen fixation in cyanobacteria, Microbiol. Rev.56 (1992) 340–373.
[55] B. Bergman, J.R. Gallon, A.N. Rai, L.J. Stal, N2 fixation by non-heterocystouscyanobacteria, FEMS Microbiol. Rev. 19 (1997) 139–185.
[56] A. Melis, J. Neidhardt, J.R. Benemann, Dunaliella salina (Chlorophyta) withsmall chlorophyll antenna sizes exhibit higher photosynthetic productivities

1 Photo
12 S.I. Allakhverdiev et al. / Journal of Photochemistry andand photon use efficiencies than normally pigmented cells, J. Appl. Phycol. 10(1999) 515–525.
[57] L. Cournac, G. Guedeney, G. Peltier, P.M. Vignais, Sustained photoevolutionof molecular hydrogen in a mutant of Synechocystis sp. strain PCC 6803 defi-cient in the type I NADPHdehydrogenase complex, J. Bacteriol. 186 (2004)1737–1746.
[58] M.L. Ghirardi, W. Amos, Renewable hydrogen from green algae, Biocycle 45(2004) 59–62.
[59] K.K. Rao, D.O. Hall, Hydrogen production by cyanobacteria: potential, prob-lems and prospects, J. Mar. Biotechnol. 4 (1996) 10–15.
[60] D.J. Dickson, C.J. Page, R.L. Ely, Photobiological hydrogen production fromSynechocystis sp. PCC 6803 encapsulated in silica sol–gel, Int. J. HydrogenEnergy 34 (2009) 204–215.
[61] H. Zhu, T. Suzuki, A.A. Tsygankov, Y. Asada, J. Miyake, Hydrogen productionfrom tofu wastewater by Rhodobacter sphaeroides immobilized in agar gels,Int. J. Hydrogen Energy 24 (1999) 305–310.
[62] R. Rouillon, M. Toscabens, R. Carpentier, A photoelectrochemical cell fordetecting pollutant-induced effects on the activity of immobilized cyanobac-terium Synechococcus sp. PCC 7942, Enzyme Microb. Technol. 25 (1999)230–235.
[63] R. Carpentier, Immobilization of photosynthetic membranes in analbumin–glutaraldehyde crosslinked matrix, in: G.F. Bickerstaff (Ed.),Methods in Biotechnology, Immobilization of Enzymes and Cells, vol. 1,Humana Press Inc., Totowa, NJ, USA, 1997, pp. 101–108.
[64] R. Rouillon, P. Euzet, R. Carpentier, Stabilization of photosynthetic materi-als, in: R. Carpentier (Ed.), Methods in Molecular Biology, PhotosynthesisResearch Protocols, vol. 274, Humana Press Inc., Totowa, NJ, USA, 2004, pp.261–269.
[65] R. Rouillon, J.J. Mestres, J.L. Marty, Entrapment of chloroplasts and thylakoidsin polyvinylalcohol-SbQ. Optimization of membrane preparation and storageconditions, Anal. Chim. Acta 311 (1995) 437–442.
[66] V.V. Klimov, S.I. Allakhverdiev, V.A. Shuvalov, A.A. Krasnovsky, Effect ofextraction and readdition of manganese on light reactions of photosystemII preparations, FEBS Lett. 148 (1982) 307–312.
[67] V.V. Klimov, G.M. Ananyev, S.I. Allakhverdiev, S.K. Zharmukhamedov, M.Mulay, U. Hedge, S. Padhye, Photoreaction and photoinactivation of photo-system II after a complete removal ofmanganese from pea subchloroplastparticles, in: M. Baltscheffsky (Ed.), Current Research in Photosynthesis,Kluwer Academic Publishers, Dordrecht, 1990, pp. 247–254.
[68] S.I. Allakhverdiev, M.S. Karacan, G. Somer, N. Karacan, E.M. Khan, S.Y. Rane, S.Padhye, V.V. Klimov, G. Renger, Reconstitution of the water-oxidizing com-plex in manganese depleted photosystem II complexes by using syntheticbinuclearmanganese complexes, Biochemistry 33 (1994) 12210–12214.
[69] S.I. Allakhverdiev, M.S. Karacan, G. Somer, N. Karacan, E.M. Khan, S.Y. Rane,S. Padhye, V.V. Klimov, G. Renger, Binuclear manganese (III) complexesas electron donors in D1/D2/cytochrome b559 preparations isolated fromspinach photosystem II membrane fragments, Z. Naturforsch. 49c (1994)587–592.
[70] S. Hotchandani, U. Ozdemir, C. Nasr, S.I. Allakhverdiev, N. Karacan, V.V.Klimov, P.V. Kamat, R. Carpentier, Redox characterization of schiff basemanganese and cobalt complexes related to water-oxidizing complex of pho-tosynthesis, Bioelectrochem. Bioenerg. 48 (1999) 53–59.
[71] S. Hotchandani, U. Ozdemir, S.I. Allakhverdiev, N. Karacan, V.V. Klimov,P.V. Kamat, R. Carpentier, Redox characteristics of manganese and cobaltcomplexes obtained from pyridine N-oxide, Bioelectrochemistry 51 (2000)175–180.
[72] R. Lomoth, A. Magnuson, M. Sjödin, P. Huang, S. Styring, L. Hammarström,Mimicking the electron donor side of photosystem II in artificial photosyn-thesis, Photosynth. Res. 87 (2006) 25–40.
[73] T. Nagata, T. Nagasawa, S.K. Zharmukhamedov, V.V. Klimov, S.I. Allakhverdiev,Reconstitution of the water-oxidizing complex in manganese-depletedphotosystem II preparations using synthetic binuclear Mn(II) and Mn(IV)complexes: production of hydrogen peroxide, Photosynth. Res. 93 (2007)133–138.
[74] T. Nagata, S.K. Zharmukhamedov, A.A. Khorobrykh, V.V. Klimov, S.I.Allakhverdiev, Reconstitution of the water-oxidizing complex in manganese-depleted.photosystem II preparations using synthetic Mn complexes: afluorine-19NMRstudy of the reconstitution process, Photosynth. Res. 98(2008) 277–284.
[75] V.N. Kurashov, S.I. Allakhverdiev, S.K. Zharmukhamedov, T. Nagata, V.V.Klimov, A.Y. Semenov, M.D. Mamedov, Electrogenic reactions on the donorside of Mn-depleted photosystem II core particles in the presence of MnCl2and synthetic trinuclear Mn-complexes, Photochem. Photobiol. Sci. 8 (2009)162–166.
[76] T. Nagasawa, S.I. Allakhverdiev, T. Nagata Kimura, Photooxidation of alcoholsby a porphyrin/quinone/TEMPO system, Photochem. Photobiol. Sci. 8 (2009)174–180.
[77] S.I. Allakhverdiev, N. Murata, Environmental stress inhibits the synthesis denovo of proteins involved in the photodamage–repair cycle of Photosystem IIin Synechocystis sp. PCC 6803, Biochim. Biophys. Acta 1657 (2004) 23–32.
[78] Y. Nishiyama, S.I. Allakhverdiev, N. Murata, Inhibition of the repair of pho-tosystem II by oxidative stress in cyanobacteria, Photosynth. Res. 84 (2005)1–7.
[79] Y. Nishiyama, S.I. Allakhverdiev, N. Murata, A new paradigm for the actionof reactive oxygen species in the photoinhibition of photosystem II, Biochim.Biophys. Acta 1757 (2006) 742–749.
biology C: Photochemistry Reviews 11 (2010) 101–113
[80] N. Murata, S. Takahashi, Y. Nishiyama, S.I. Allakhverdiev, Photoinhibitionof photosystem II under environmental stress, Biochim. Biophys. Acta 1767(2007) 414–421.
[81] P. Mohanty, S.I. Allakhverdiev, N. Murata, Application of low tempera-tures during photoinhibition allows characterization of individual steps inphotodamage and the repair of photosystem II, Photosyn. Res. 94 (2007)217–224.
[82] V.D. Kreslavski, R. Carpentier, V.V. Klimov, N. Murata, S.I. Allakhverdiev,Molecular mechanisms of stress resistance of the photosynthetic apparatus,Membr. Cell Biol. 1 (2007) 185–205.
[83] S.I. Allakhverdiev, V.D. Kreslavski, V.V. Klimov, D.A. Los, R. Carpentier, P.Mohanty, Heat stress: an overview of molecular responses in photosynthesis,Photosynth. Res. 98 (2008) 541–550.
[84] V.D. Kreslavski, R. Carpentier, V.V. Klimov, S.I. Allakhverdiev, Transductionmechanisms of photoreceptor signals in plant cells, J. Photochem. Photobiol.C: Photochem. Rev. 10 (2009) 63–80.
[85] V.V. Klimov, S.I. Allakhverdiev, Y.M. Feyziev, S.V. Baranov, Bicarbonaterequirement for the donor side of photosystem II, FEBS Lett. 363 (1995)251–255.
[86] S.I. Allakhverdiev, Y.M. Feyziev, A. Ahmed, H. Hayashi, J.A. Aliev, V.V. Klimov,N. Murata, R. Carpentier, Stabilization of oxygen evolution and primaryelectron transport reactions in photosystem II against heat stress with glycin-betaine and sucrose, J. Photochem. Photobiol. B 34 (1996) 149–157.
[87] H. Wincencjusz, S.I. Allakhverdiev, V.V. Klimov, H.J. van Gorkom, Bicarbonate-reversible formate inhibition at the donor side of photosystem II, Biochim.Biophys. Acta 1273 (1996) 1–3.
[88] S.I. Allakhverdiev, Y. Yruela, R. Picorel, V.V. Klimov, Bicarbonate is an essen-tial constituent of the water-oxidizing complex of photosystem II, Proc. Natl.Acad. Sci. U.S.A. 94 (1997) 5050–5054.
[89] V.V. Klimov, S.I. Allakhverdiev, B. Hulsebosch, H. Wincencjusz, H.J. vanGorkom, A. Hoff, Bicarbonate may be required for ligation of manganesein the oxygen-evolving complex of photosystem II, Biochemistry 36 (1997)16277–16281.
[90] V.V. Klimov, S.V. Baranov, S.I. Allakhverdiev, Bicarbonate protects the donorside of photosystem II against photoinhibition and thermoinactivation, FEBSLett. 418 (1997) 243–246.
[91] S.I. Allakhverdiev, H. Hayashi, Y. Nishiyama, A.G. Ivanov, J.A. Aliev, V.V. Klimov,N. Murata, R. Carpentier, Glycinebetaine protects the D1/D2/Cytb559 complexof photosystem II against photo-induced and heat-induced inactivation, J.Plant Physiol. 160 (2003) 41–49.
[92] J.L. Roose, K.M. Wegener, H.B. Pakrasi, The extrinsic proteins of photosystemII, Photosynth. Res. 92 (2006) 369–387.
[93] T. Shutova, H. Kenneweg, J. Buchta, J. Nikitina, V. Terentyev, S. Chernyshov,B. Andersson, S.I. Allakhverdiev, V.V. Klimov, H. Dau, W. Junge, G. Samuels-son, The photosystem II-associated Cah3 in Chlamydomonas enhances the O2
evolution rate by proton removal, EMBO J. 27 (2008) 782–791.[94] A.K. Jones, S.E. Lamle, H.R. Pershad, K.A. Vincent, S.P. Albracht, F.A. Armstrong,
Enzyme electrokinetics: electrochemical studies of the anaerobic intercon-versions between active and inactive states of Allochromatium vinosum[NiFe]-hydrogenase, J. Am. Chem. Soc. 125 (2003) 8505–8514.
[95] M.L. Ghirardi, R.K. Togasaki, M. Seibert, Oxygen sensitivity of algal H2-production, Appl. Biochem. Biotechnol. 63–65 (1997) 141–151.
[96] W.D. Swingley, M. Chen, P.C. Cheung, A.L. Conrad, L.C. Dejesa, J. Hao, B.M.Honchak, L.E. Karbach, A. Kurdoglu, S. Lahiri, S.D. Mastrian, H. Miyashita, L.Page, P. Ramakrishna, S. Satoh, W.M. Sattley, Y. Shimada, H.L. Taylor, T. Tomo,T. Tsuchiya, Z. Wang, J. Raymond, M. Mimuro, R.E. Blankenship, J.W. Touch-man, Niche adaptation and genome expansion in the chlorophyll d-producingcyanobacterium Acaryochloris marina, Proc. Natl. Acad. Sci. U.S.A. 105 (2008)2005–2010.
[97] T. Tomo, Y. Kato, T. Suzuki, S. Akimoto, T. Okubo, K. Hasegawa, T. Noguchi, T.Tsuchiya, K. Tanaka, M. Fukuya, N. Dohmae, T. Watanabe, M. Mimuro, Charac-terization of highly purified PS I complexes from the chlorophyll d dominatedcyanobacterium Acaryochloris marina MBIC 11017, J. Biol. Chem. 283 (2008)18198–18209.
[98] T. Tomo, T. Okubo, S. Akimoto, M. Yokono, H. Miyashita, T. Tsuchiya, T.Noguchi, M. Mimuro, Identification of the special pair of photosystem II inthe chlorophyll d-dominated cyanobacterium, Proc. Natl. Acad. Sci. U.S.A. 104(2007) 7283–7288.
[99] G. Bernát, N. Waschewski, M. Rögner, Towards efficient hydrogen pro-duction: the impact of antenna size and external factors on electrontransport dynamics in Synechocystis PCC 6803, Photosynth. Res. 99 (2009)205–216.
[100] S. Kosourov, M. Seibert, M.L. Ghirardi, Effects of extracellular pH on themetabolic pathways in sulfur-deprived, H2-producing cultures, Plant CellPhysiol. 44 (2003) 146–155.
[101] V.A. Boichenko, S.I. Allakhverdiev, V.G. Ladygin, V.V. Klimov, Functional con-junction of hydrogenase with photosystem II in whole cells of Chlamydomonasreinhardtii mutants, Dokl. Acad. Nauk. SSSR 290 (1986) 995–998.
[102] S.V. Mal’tsev, S.I. Allakhverdiev, V.V. Klimov, A.A. Krasnovsky, Hydrogen evo-lution by subchloroplast preparations of photosystem II from pea and spinach,
FEBS Lett. 240 (1988) 1–5.[103] S.I. Allakhverdiev, V.V. Klimov, Photoreduction of NADP+ in photosystem IIof higher plants: requirement for manganese, Z. Naturforsch. 47c (1992)57–62.
[104] V.V. Klimov, S.I. Allakhverdiev, S. Demeter, A.A. Krasnovsky, Photoreduc-tion of pheophytin in the photosystem 2 of chloroplasts depending on the

Photo
[
[
[
[
[
[
[
[117] http://www.eia.doe.gov/oiaf/ieo/index.html: International Energy Outlook2008, Report # DOE/EIA-0484 (2008), Energy Information Administration,U.S. Department of Energy, Washington [accessed 07.09].
[118] http://www.esrl.noaa.gov/gmd/ccgg/trends/: Pieter Tans, Trends in Atmo-
S.I. Allakhverdiev et al. / Journal of Photochemistry and
oxidation–reduction potential of the medium, Dokl. Acad. Nauk. SSSR 249(1979) 227–230.
105] S.I. Allakhverdiev, T. Tomo, Y. Shimada, H. Kindo, R. Nagao, V.V. Klimov, M.Mimuro, Redox potential of pheophytin a in photosystem II of two cyanobac-teria having the different special pair chlorophylls, Proc. Natl. Acad. Sci. U.S.A.107 (2010) 3924–3929.
106] E. Franchi, C. Tosi, G. Scolla, G. Della Penna, F. Rodriguez, P.M. Pedroni,Metabolically engineered Rhodobacter sphaeroides RV strains for improvedbiohydrogen photoproduction combined with disposal of food wastes, Mar.Biotechnol. 6 (2004) 552–565.
107] C. Lee, P. Chen, C. Wang, Y. Tung, Photohydrogen production using purple non-sulfur bacteria with hydrogen fermentation reactor effluent, Int. J. HydrogenEnergy 27 (2002) 1309–1313.
108] H. Koku, I. Eroglu, U. Gunduz, M. Yucel, L. Turker, Aspects of the metabolismof hydrogen production by Rhodobacter sphaeroides, Int. J. Hydrogen Energy27 (2002) 1315–1329.
109] A.A. Tsygankov, A.S. Fedorov, S.N. Kosourov, K.K. Rao, Hydrogen productionby cyanobacteria in an automated outdoor photobioreactor under aerobicconditions, Biotechnol. Bioeng. 80 (2002) 777–783.
110] S.N. Kosourov, A.A. Tsygankov, M. Seibert, M.L. Ghirardi, Sustained hydrogenphotoproduction by Chlamydomonas reinhardtii: effects of culture parame-ters, Biotechnol. Bioeng. 78 (2002) 731–740.
111] J. Moreno, M.A. Vargas, H. Rodriguez, J. Rivas, M.G. Guerrero, Outdoor cultiva-tion of a nitrogen-fixing marine cyanobacterium, Anabaena sp. ATCC 33047,Biomol. Eng. 20 (2003) 191–197.
biology C: Photochemistry Reviews 11 (2010) 101–113 113
[112] A. Shah, P. Torres, R. Tscharner, N. Wyrsch, H. Keppner, Photovoltaic technol-ogy: the case for thin-film solar cells, Science 285 (1999) 692–698.
[113] M. Grätzel, Photoelectrochemical cells, Nature 414 (2001) 338–344.[114] H. Berberoglu, J. Jay, L. Pilon, Effect of nutrient media on photobiological
hydrogen production by Anabaena variabilis ATCC 29413, Int. J. HydrogenEnergy 33 (2008) 1172–1184.
[115] A. Gauthier, R. Carpentier, Disorganization of the Mn4Ca complex of photo-system II by ruthenium red: a thermoluminescence study, Luminescence 24(2009) 108–114.
[116] M. Campàs, R. Carpentier, R. Rouillon, Plant tissue and photosynthesis-basedbiosensors, Biotechnol. Adv. 26 (2008) 370–378.
Web references
spheric Carbon Dioxide—Mauna Loa, NOAA/ESRL [accessed 07.09].