mechanistic study of microbial control of hydrogen sulfide production in oil reservoirs
TRANSCRIPT

Mechanistic Study of Microbial Controlof Hydrogen Sulfide Production inOil Reservoirs
M. Nemati,1 G. E. Jenneman,2 G. Voordouw1
1Department of Biological Sciences, University of Calgary, Calgary,Alberta, Canada, T2N 1N4; telephone: 1 (403) 2203138;fax: 1 (403) 2899311; e-mail: [email protected] Petroleum Company, 224 GB, Bartlesville, Oklahoma 74004, USA
Received 26 October 2000; accepted 2 March 2001
Abstract: Microbial control of biogenic production of hy-drogen sulfide in oil fields was studied in a model systemconsisting of pure cultures of the nitrate-reducing, sul-fide-oxidizing bacterium (NR-SOB) Thiomicrospira sp.strain CVO and the sulfate-reducing bacterium (SRB)Desulfovibrio sp. strain Lac6, as well as in microbial cul-tures enriched from produced water of a Canadian oilreservoir. The presence of nitrate at concentrations up to20 mM had little effect on the rate of sulfate reduction bya pure culture of Lac6. Addition of CVO imposed a stronginhibition effect on production of sulfide. In the absenceof added nitrate SRB we were able to overcome this ef-fect after an extended lag phase. Simultaneous additionof CVO and nitrate stopped the production of H2S imme-diately. The concentration of sulfide decreased to a neg-ligible level due to nitrate-dependent sulfide oxidationactivity of CVO. This was not prevented by raising theconcentration of Na-lactate, the electron donor for sul-fate reduction. Similar results were obtained with enrich-ment cultures. Enrichments of produced water with sul-fide and nitrate were dominated by CVO, whereas enrich-ments with sulfate and Na-lactate were dominated bySRB. Addition of an NR-SOB enrichment to an SRB en-richment inhibited the production of sulfide. Subsequentaddition of sufficient nitrate caused the sulfide concen-tration to drop to zero. A similar response was seen inthe presence of nitrate alone, although after a pro-nounced lag time, it was needed for emergence of a siz-able CVO population. The results of the present studyshow that two mechanisms are involved in microbialcontrol of biogenic sulfide production. First, addition ofNR-SOB imposes an inhibition effect, possibly by in-creasing the environmental redox potential to levelswhich are inhibitory for SRB. Second, in the presence ofsufficient nitrate, NR-SOB oxidize sulfide, leading to itscomplete removal from the environment. Successful mi-crobial control of H2S in an oil reservoir is crucially de-pendent on the simultaneous presence of NR-SOB (ei-ther indigenous population or injected) and nitrate in theenvironment. © 2001 John Wiley & Sons, Inc. Biotechnol Bio-eng 74: 424–434, 2001.Keywords: oil reservoir; hydrogen sulfide; souring; ni-trate; Thiomicrospira sp. strain CVO; reverse sample ge-nome probing
INTRODUCTION
Souring, the formation of H2S, often results when an oilreservoir is subjected to water flooding for secondary oilrecovery. Souring increases the cost of oil production and isbelieved to be mediated by the activity of sulfate-reducingbacteria (SRB) in the reservoir. SRB are strict anaerobes,which use hydrogen or simple organic acids or alcohols aselectron donors for sulfate reduction. These are formed asend-products of fermentative metabolism or through chemi-cal transformation of buried organic matter at high tempera-ture and pressure (Shock, 1988). Recent studies have shownthat hydrocarbons in petroleum may also serve as electrondonors in bioreduction of sulfate. Toxicity of H2S, acceler-ated corrosion rates, and decreased efficiency of secondaryoil recovery due to plugging of the reservoir by SRB bio-mass and precipitated metal sulfides are some of the prob-lems associated with souring. This dictates control of H2Sproduction, of its release into the environment, and of itscontact with iron and steel parts in tanks, pipelines, valves,and pumps. Different approaches can be used to controlsulfide contamination. Sulfide production can be reduced byinhibiting SRB activity by treatment of injection water withbiocides such as glutaraldehyde, cocodiamines, or molyb-dates. The use of biocide is most successful in controllingSRB in surface facilities but is of limited effectiveness inthe reservoir, because chemical components in the reservoirmay scavenge biocide through reaction (Reinsel et al.,1996). Elimination of sulfate from the injection water isanother option to control SRB activity (Bakke et al., 1992).It is also possible to remove H2S from sour water by treat-ment with physicochemical methods (Montgomery et al.,1990). Finally, sulfide production can be controlled micro-bially. For this option it is important to note that oil fields atmoderate subsurface depth and a moderate resident tem-perature harbor a complex microbial community. Stimula-tion and control of the metabolism of this community canprevent or revert souring (Telang et al., 1997). Addition ofnitrate and/or nitrite and introduction of nitrate-reducing,sulfide-oxidizing bacteria (NR-SOB) such asThiobacillusdenitrificansin injection water have been shown to controlsulfide accumulation (Jenneman et al., 1986, 1997; McIn-
Correspondence to:M. NematiContract grant sponsors: the Natural Sciences and Engineering Research
Council of Canada and the Phillips Petroleum Company.
© 2001 John Wiley & Sons, Inc.

erney et al., 1992, 1996). Earlier workers have proposedvarious explanations for the decrease in biogenic sulfideproduction in oil reservoirs due to nitrate addition. How-ever, the exact mechanism underlying this phenomenon isnot fully understood and by no means settled.
In the work presented here two model systems consistingof 1) an SRB and an NR-SOB, and 2) microbial culturesenriched from oil field produced water are used to elucidatethe mechanism involved in the nitrate control of biogenicH2S production. Surprisingly, the data indicate that NR-SOB not only controls the level of H2S through sulfideoxidation but also imposes a strong and immediate inhibi-tion effect on production of sulfide by SRB.
MATERIALS AND METHODS
Microorganisms and Media
Pure cultures of the SRBDesulfovibriosp. strain Lac6 andthe NR-SOBThiomicrospirasp. strain CVO, both isolatedfrom western Canadian oil fields (Voordouw et al., 1996;Gevertz et al., 2000), were used. The stock culture of Lac6was maintained on saline Postgate’s medium C (Postgate,1984). Coleville synthetic brine (CSB) developed by Ge-vertz et al. (2000) was used for the growth and maintenanceof CVO. The composition of the media is given in Table I.Trace element mixture contained (per L): 2 g nitrilotriaceticacid, 0.0058 g FeCl3, 1.2 g CaSO4 ? 2H2O, 2.0 gMgSO4 ? 7H2O, 0.16 g NaCl, 1.4 g Na2HPO4, 0.72 gKH2PO4, and 10 mL of micronutrient solution. Micronutri-ent solution contained (per L): 0.5 mL H2SO4, 2.28 gMnSO4 ? H2O, 0.5 g ZnSO4 ? 7H2O, 0.5 g H3BO3, 0.025 gCuSO4 ? 5H2O, 0.025 g Na2MoO4 ? 2H2O, and 0.045 gCoCl2 ? 6H2O. All medium components, except the sodium
sulfide in CSB medium, were combined and the pH wasadjusted to 7.0 using either 2M HCl or 2M NaOH. Mediawere dispensed in 100 mL aliquots in 120 mL serum bottles.The bottles were sealed, flushed with mixed gas (85% N2,5% H2, 10% CO2 by volume), and autoclaved. Sulfide wasadded aseptically to sterilized CSB medium from a sterilestock solution of 1 M Na2S. Stock cultures of Lac6 andCVO were used as inoculum (3% v/v). The cultures weremaintained at 30°C and were subcultured on a weekly basis.
To simulate the microbial communities which could pre-vail in an oil reservoir under different conditions, namelypopulations dominated by SRB and NR-SOB, two differentmedia were used. CSB medium was used to enrich a culturedominated by NR-SOB and a modified CSB medium con-taining 14 mM of sulfate, 30 mM of Na-lactate, no nitrate,and no sulfide was employed to achieve an SRB-dominatedculture. Coleville produced water was used as an inoculum(5% v/v) in both cases. The preparation of media, mainte-nance of the cultures, and all other conditions were similarto those described for pure cultures.
Field Sample
A produced water sample (4 L) was obtained from the Cole-ville oil field, located near Kindersley, Saskatchewan,Canada. In this field oil is produced by water flooding.Upon arrival the sample was stored in a Coy anaerobicchamber equilibrated with mixed gas.
Effects of Nitrate Concentration andCVO Injection
Modified CSB media containing 14 mM sulfate, 10 mMlactate, no sulfide, and nitrate concentrations in the range0–20 mM were used to investigate the effect of nitrate onthe activity of Lac6. Serum bottles containing 100 mL ofmodified CSB medium were inoculated with 3 mL of aLac6 culture grown in sPGC medium. The cultures wereincubated at 30°C. To assess the effect of CVO on H2Sproduction, to each exponentially growing culture (100 mL;2 × 108 cells/mL) 3 mL of a CVO culture (108 cells/mL)was added. The concentrations of sulfide, nitrite, and sulfatewere monitored during the course of the experiment. Thefractions of CVO and Lac6 in co-cultures were determinedby reverse sample genome probing (RSGP) at the end of theexperiment.
Effect of Na-Lactate Concentration
Different concentrations of Na-lactate ranging from 10–50mM were added to a modified CSB medium containing 14mM sulfate and 10 mM nitrate. The medium bottles werethen inoculated with 3 mL of a Lac6 culture (109 cells/mL)and incubated at 30°C. During exponential growth of Lac6,3 mL of a CVO culture (108 cells/mL) was added to each ofa set of Lac6 cultures. The concentrations of sulfide andsulfate were monitored as a function of time. The fractions
Table I. Composition of the media used for growth ofDesulfovibriostrain Lac6 andThiomicrospirastrain CVO.
Component1 sPGC2 CSB2
NaCl 7.0 7.0MgCl2 ? 6H2O 1.2 0KH2PO4 0.5 0.027NH4Cl 1.0 0.02Na2SO4 4.5 0CaCl2 ? 2H2O 0.042 0.24MgSO4 ? 7H2O 0.03 0.68FeSO4 ? 7H2O 0.004 0(NH4)2SO4 0 0.13KNO3 0 1.0sodium citrate 0.28 0sodium lactate (70% w/v) 8.6 0sodium acetate 0 0.681 M Na2S ? 9H2O 0 2.5 mLresazurin (0.1% w/v) 1 mL 1 mLNaHCO3 0 1.90trace elements 0 50 mLyeast extract 1.0 0
1All amounts are in g/L except where stated otherwise.2sPGC, saline Postgate’s medium C; CSB, Coleville synthetic brine.
NEMATI ET AL.: MICROBIAL CONTROL OF SOURING IN OIL RESERVOIRS 425

of CVO and Lac6 in the co-cultures were determined byRSGP.
Studies with CVO Washed Cells and Nitrate
The effect of addition of washed CVO cells, or of additionof both washed CVO cells and nitrate (final concentrations0.2–10 mM) on the activity of Lac6 were studied.Desulfo-vibrio sp. strain Lac6 was grown in modified CSB mediumcontaining 14 mM sulfate, 30 mM lactate, no sulfide, and nonitrate. During the exponential growth phase, 0.25 ml ofCVO cell suspension (1010 cells/mL) or both 0.25 ml ofCVO cell suspension and different amounts of nitrate wereadded to a set of Lac6 cultures. To verify whether metabo-lites formed by CVO have any effect on biogenic produc-tion of H2S, 5 mL of a CVO culture filtrate was added to agrowing culture of Lac6. The culture filtrate was preparedby passing the CVO culture through a 0.2-mm membranefilter (Millipore, Bedford, MA, USA). The concentrationsof sulfide and sulfate were monitored during the experi-ment. A suspension of CVO cells in distilled water wasprepared by centrifugation of 200 mL of CVO culture at17,000 rpm for 1 h. The resulting pellets were washed twice,resuspended in 2 mL of distilled water, and used in differentexperiments.
Studies with Produced Water Enrichments
The efficiency of the microbial control of H2S production ina microbial consortium similar to that in the oil reservoirwas studied using a microbial culture enriched from pro-duced water. To simulate the microbial communities whichcould prevail in an oil reservoir under different conditions,namely, populations dominated by SRB and NR-SOB, twodifferent media were used. CSB medium was used to enricha culture dominated by NR-SOB and a modified CSB me-dium containing 14 mM of sulfate, 30 mM of Na-lactate, nonitrate, and no sulfide was employed to achieve an SRBdominated culture. Coleville produced water was used as aninoculum (5% v/v) in both cases.
The effects of addition of: 1) nitrate at a concentration of10 mM (in the culture); 2) NR-SOB enrichment (5% v/v); 3)combination of nitrate (10 mM) and NR-SOB enrichment(5% v/v), on the activity of a growing population of SRB(modified CSB inoculated with produced water) were in-vestigated. The additions were carried out during the expo-nential phase of SRB growth. The concentrations of sulfideand sulfate were monitored during the course of the experi-ments. The composition of the microbial community wasdetermined for each enrichment after complete utilization ofeither sulfide or sulfate, while for co-cultures this was car-ried out at various stages of the experiment.
Analysis of Microbial Community Composition
Composition of the microbial community was analyzed byRSGP. DNA was isolated from the mixed cultures accord-
ing to a procedure described previously (Telang et al.,1994). The concentration of purified DNA was determinedfluorimetrically (Voordouw et al., 1993). Purified DNA(100 ng) was mixed with 2.5 ng of bacteriophagel DNAand labeled with the random hexamer procedure using [a32-P] dCTP (Voordouw et al., 1992). The denatured, labeledprobe was hybridized with the oil field master filter for 16h under stringent conditions at 68°C (Voordouw et al.,1989). Following washing and drying, the filters were ex-posed to BAS-III type Image Plates which were scannedwith a BAS 1000 Bioimage Analyzer. The measured hy-bridization intensities were used to calculate the fraction ofeach microorganism in the microbial community. The mas-ter filter contained the denatured chromosomal DNA of oilfield bacteria standards, including 30 SRB, two sulfide-oxidizing bacteria, and 16 heterotrophs (Table II). Standardsare defined as bacteria with genomes that have no or limitedcross hybridization (Voordouw, 1998). A batch of 200 mas-ter filters had been prepared in previous work (Telang et al.,1997) by spotting denatured chromosomal DNA of stan-dards in a known amount (usually 100 ng) at known loca-tions on 8 × 7 cmHybond-N grid. Denatured bacteriophagel DNA (5, 10, 20, 30, 50, 100, 200, and 400 ng) was alsoincluded on the filters. Following covalent linkage of theDNAs on the filter, the filters were stored at −20°C. Thesewere then used for analysis of microbial communities.
Chemical Analysis
The concentration of sulfide was determined using a color-imetric method (Cord-Ruwisch, 1985). To 0.1 mL of thesample, 0.9 mL of an acidic copper sulfate solution wasadded. The absorbance of the resulting mixture was mea-sured at 480 nm. A calibration curve, prepared using stan-dard solutions of sulfide, was used to calculate the concen-tration of sulfide in the sample. A turbidimetric method wasused to measure the concentration of sulfate (APHA, 1992).To 0.1 mL of a centrifuged sample, 0.9 mL of a condition-ing agent (0.85 mL glycerol, 0.5 mL concentrated HCl, 1.3g Na Cl, 1.7 mL ethanol and 1L distilled water) and anexcess amount of finely ground BaCl2 was added. Thesewere then mixed and the absorbance of the mixture wasmeasured at 420 nm. Using a calibration curve, the concen-tration of sulfate was calculated. Nitrite was measured by acolorimetric method (APHA, 1992). To 25 mL of a dilutedsample, 0.5 mL of 0.1% sulfanilamide solution was added.The reagent reacted with sample for 5 min and then 0.5 mLof NED-dihydrochloride was added. The absorbance of theresulting solution was measured at 543 nm after 10 min. Acalibration curve was used to determine the concentration ofnitrite in the sample.
RESULTS
Effects of Nitrate Concentration andCVO Injection
Nitrate at concentrations up to 7.5 mM did not have anyeffect on the production of sulfide by Lac6 (Fig. 1A). With
426 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 74, NO. 5, SEPTEMBER 5, 2001
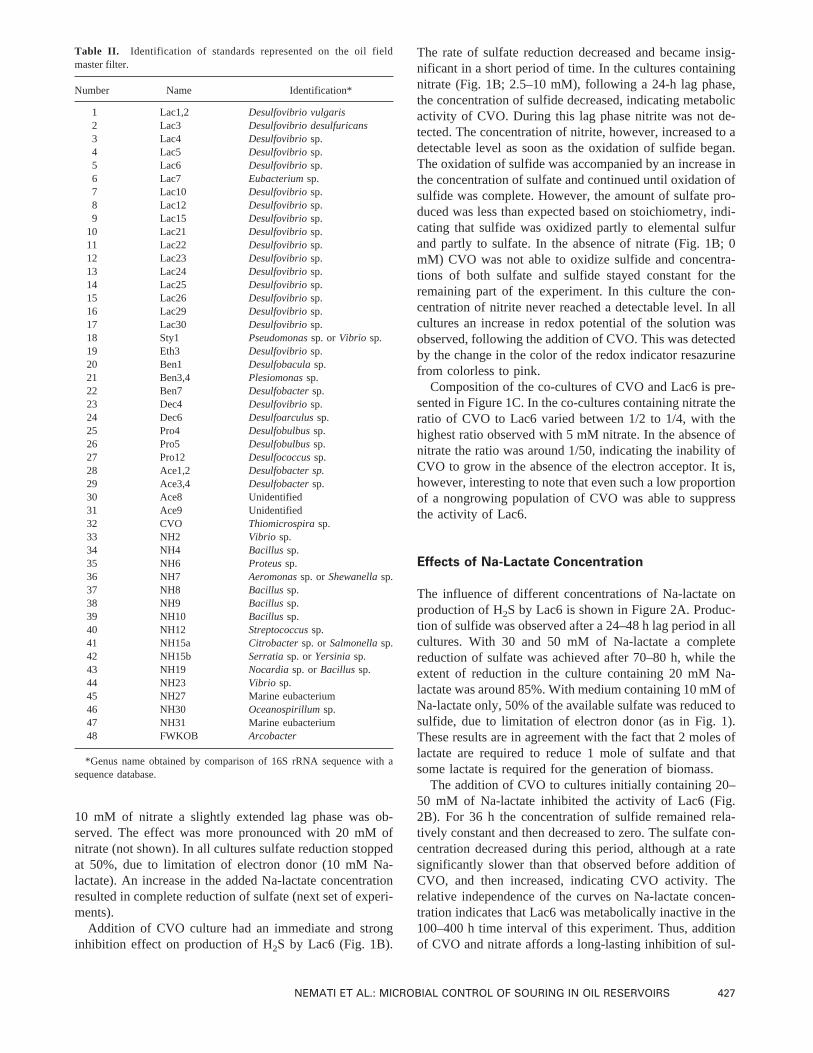
10 mM of nitrate a slightly extended lag phase was ob-served. The effect was more pronounced with 20 mM ofnitrate (not shown). In all cultures sulfate reduction stoppedat 50%, due to limitation of electron donor (10 mM Na-lactate). An increase in the added Na-lactate concentrationresulted in complete reduction of sulfate (next set of experi-ments).
Addition of CVO culture had an immediate and stronginhibition effect on production of H2S by Lac6 (Fig. 1B).
The rate of sulfate reduction decreased and became insig-nificant in a short period of time. In the cultures containingnitrate (Fig. 1B; 2.5–10 mM), following a 24-h lag phase,the concentration of sulfide decreased, indicating metabolicactivity of CVO. During this lag phase nitrite was not de-tected. The concentration of nitrite, however, increased to adetectable level as soon as the oxidation of sulfide began.The oxidation of sulfide was accompanied by an increase inthe concentration of sulfate and continued until oxidation ofsulfide was complete. However, the amount of sulfate pro-duced was less than expected based on stoichiometry, indi-cating that sulfide was oxidized partly to elemental sulfurand partly to sulfate. In the absence of nitrate (Fig. 1B; 0mM) CVO was not able to oxidize sulfide and concentra-tions of both sulfate and sulfide stayed constant for theremaining part of the experiment. In this culture the con-centration of nitrite never reached a detectable level. In allcultures an increase in redox potential of the solution wasobserved, following the addition of CVO. This was detectedby the change in the color of the redox indicator resazurinefrom colorless to pink.
Composition of the co-cultures of CVO and Lac6 is pre-sented in Figure 1C. In the co-cultures containing nitrate theratio of CVO to Lac6 varied between 1/2 to 1/4, with thehighest ratio observed with 5 mM nitrate. In the absence ofnitrate the ratio was around 1/50, indicating the inability ofCVO to grow in the absence of the electron acceptor. It is,however, interesting to note that even such a low proportionof a nongrowing population of CVO was able to suppressthe activity of Lac6.
Effects of Na-Lactate Concentration
The influence of different concentrations of Na-lactate onproduction of H2S by Lac6 is shown in Figure 2A. Produc-tion of sulfide was observed after a 24–48 h lag period in allcultures. With 30 and 50 mM of Na-lactate a completereduction of sulfate was achieved after 70–80 h, while theextent of reduction in the culture containing 20 mM Na-lactate was around 85%. With medium containing 10 mM ofNa-lactate only, 50% of the available sulfate was reduced tosulfide, due to limitation of electron donor (as in Fig. 1).These results are in agreement with the fact that 2 moles oflactate are required to reduce 1 mole of sulfate and thatsome lactate is required for the generation of biomass.
The addition of CVO to cultures initially containing 20–50 mM of Na-lactate inhibited the activity of Lac6 (Fig.2B). For 36 h the concentration of sulfide remained rela-tively constant and then decreased to zero. The sulfate con-centration decreased during this period, although at a ratesignificantly slower than that observed before addition ofCVO, and then increased, indicating CVO activity. Therelative independence of the curves on Na-lactate concen-tration indicates that Lac6 was metabolically inactive in the100–400 h time interval of this experiment. Thus, additionof CVO and nitrate affords a long-lasting inhibition of sul-
Table II. Identification of standards represented on the oil fieldmaster filter.
Number Name Identification*
1 Lac1,2 Desulfovibrio vulgaris2 Lac3 Desulfovibrio desulfuricans3 Lac4 Desulfovibriosp.4 Lac5 Desulfovibriosp.5 Lac6 Desulfovibriosp.6 Lac7 Eubacteriumsp.7 Lac10 Desulfovibriosp.8 Lac12 Desulfovibriosp.9 Lac15 Desulfovibriosp.
10 Lac21 Desulfovibriosp.11 Lac22 Desulfovibriosp.12 Lac23 Desulfovibriosp.13 Lac24 Desulfovibriosp.14 Lac25 Desulfovibriosp.15 Lac26 Desulfovibriosp.16 Lac29 Desulfovibriosp.17 Lac30 Desulfovibriosp.18 Sty1 Pseudomonassp. orVibrio sp.19 Eth3 Desulfovibriosp.20 Ben1 Desulfobaculasp.21 Ben3,4 Plesiomonassp.22 Ben7 Desulfobactersp.23 Dec4 Desulfovibriosp.24 Dec6 Desulfoarculussp.25 Pro4 Desulfobulbussp.26 Pro5 Desulfobulbussp.27 Pro12 Desulfococcussp.28 Ace1,2 Desulfobacter sp.29 Ace3,4 Desulfobactersp.30 Ace8 Unidentified31 Ace9 Unidentified32 CVO Thiomicrospirasp.33 NH2 Vibrio sp.34 NH4 Bacillus sp.35 NH6 Proteussp.36 NH7 Aeromonassp. orShewanellasp.37 NH8 Bacillus sp.38 NH9 Bacillus sp.39 NH10 Bacillus sp.40 NH12 Streptococcussp.41 NH15a Citrobactersp. orSalmonellasp.42 NH15b Serratiasp. orYersiniasp.43 NH19 Nocardiasp. orBacillus sp.44 NH23 Vibrio sp.45 NH27 Marine eubacterium46 NH30 Oceanospirillumsp.47 NH31 Marine eubacterium48 FWKOB Arcobacter
*Genus name obtained by comparison of 16S rRNA sequence with asequence database.
NEMATI ET AL.: MICROBIAL CONTROL OF SOURING IN OIL RESERVOIRS 427
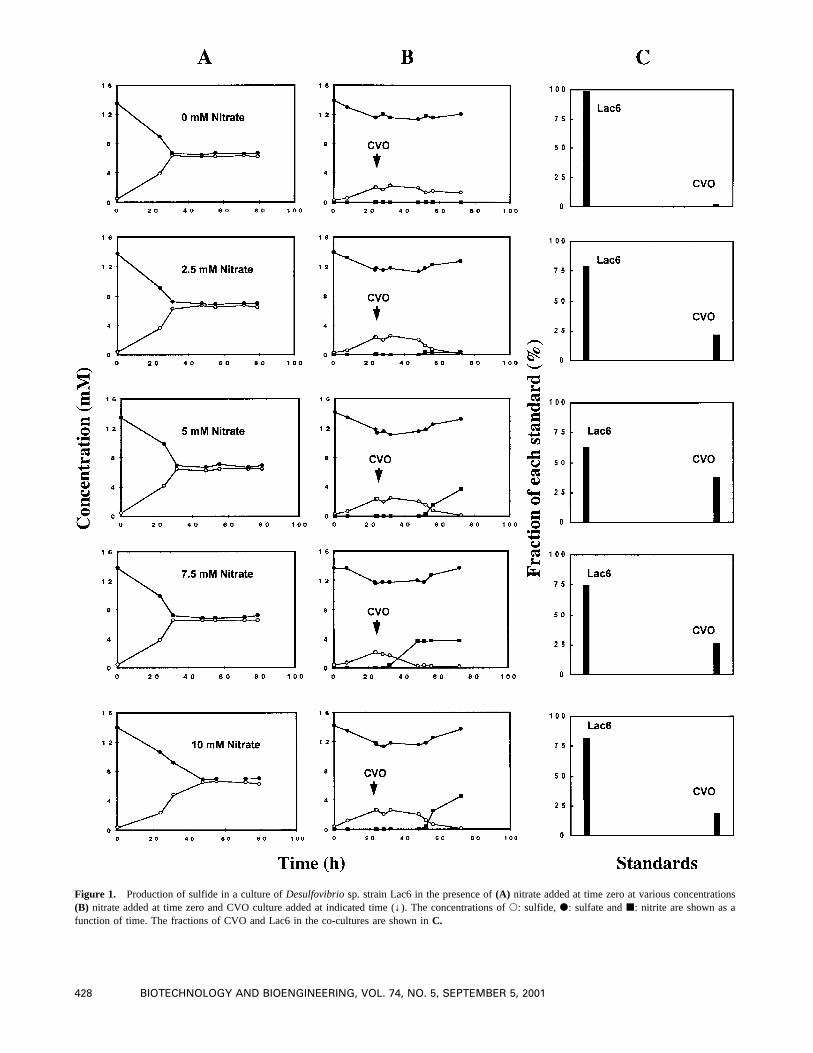
Figure 1. Production of sulfide in a culture ofDesulfovibriosp. strain Lac6 in the presence of(A) nitrate added at time zero at various concentrations(B) nitrate added at time zero and CVO culture added at indicated time (↓). The concentrations ofs: sulfide, d: sulfate andj: nitrite are shown as afunction of time. The fractions of CVO and Lac6 in the co-cultures are shown inC.
428 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 74, NO. 5, SEPTEMBER 5, 2001
fide production by Lac6 and an increase in concentration ofelectron donor did not revert this effect.
Composition of the co-cultures containing different con-centrations of Na-lactate at 140 h following complete oxi-dation of sulfide is presented in Figure 2C. The ratio ofadded CVO to Lac6 at 50 h was around 1/70. Following 90h of growth this ratio reached values between 1/10 to 1/2.5.
Studies with CVO Washed Cells and Nitrate
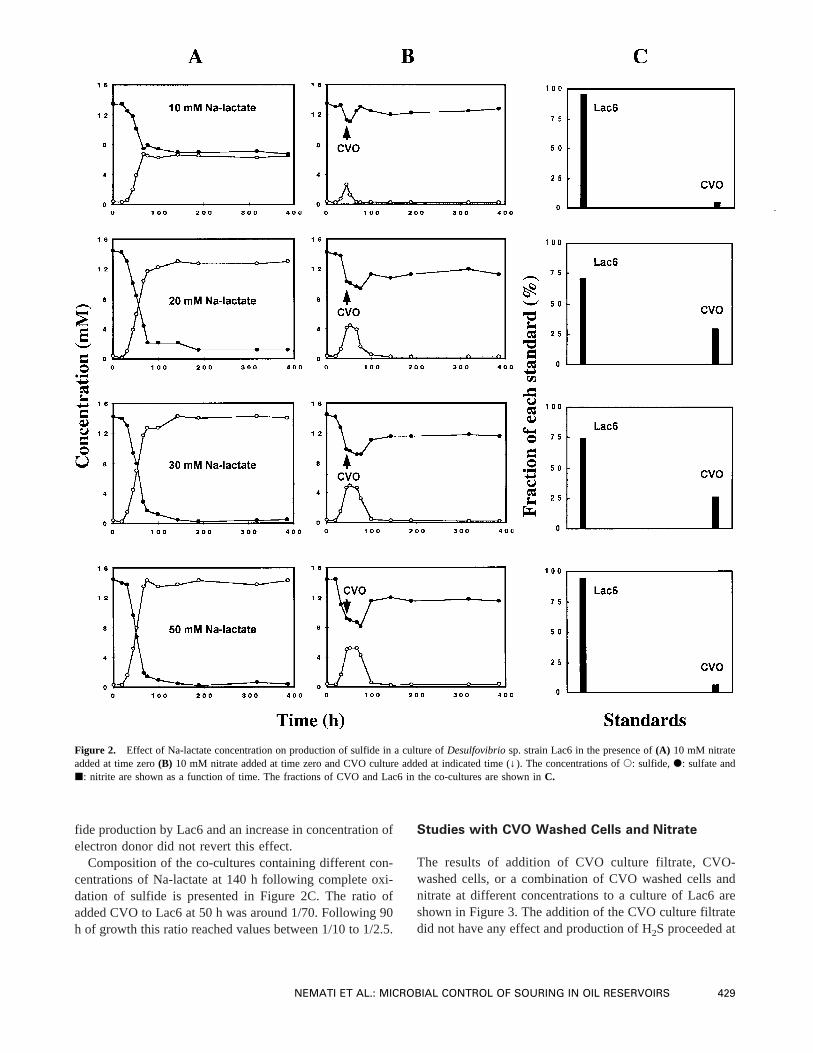
The results of addition of CVO culture filtrate, CVO-washed cells, or a combination of CVO washed cells andnitrate at different concentrations to a culture of Lac6 areshown in Figure 3. The addition of the CVO culture filtratedid not have any effect and production of H2S proceeded at
Figure 2. Effect of Na-lactate concentration on production of sulfide in a culture ofDesulfovibriosp. strain Lac6 in the presence of(A) 10 mM nitrateadded at time zero(B) 10 mM nitrate added at time zero and CVO culture added at indicated time (↓). The concentrations ofs: sulfide,d: sulfate andj: nitrite are shown as a function of time. The fractions of CVO and Lac6 in the co-cultures are shown inC.
NEMATI ET AL.: MICROBIAL CONTROL OF SOURING IN OIL RESERVOIRS 429
a rate similar to that achieved in the control culture (notshown). Addition of CVO-washed cells inhibited the activ-ity of Lac6 and a decrease in the rate of sulfate reductionwas seen. SRB, however, was able to overcome this inhi-bition effect and after 100 h a marked increase in the re-duction rate was detected. This reduction rate was slightlyslower than that observed in the absence of CVO cells.Complete reduction of sulfate was obtained after 180 h.Simultaneous addition of CVO-washed cells and nitrate,even at concentrations as low as 0.1 mM, imposed a strongand immediate inhibition effect on reduction of sulfate.With 0.1 and 0.2 mM nitrate the production of sulfidestopped and the concentration of sulfide remained constantfor the remaining part of the experiment. In the culturecontaining 0.5 mM of nitrate a slight decrease in the sulfideconcentration was measured. With higher concentrations ofnitrate the activity of CVO led to complete removal of sul-fide from the system. In the cultures containing 1 and 2 mMnitrate, despite the complete removal of sulfide, the concen-tration of sulfate remained unchanged, whereas in the pres-ence of 5 and 10 mM nitrate an increase in concentration of
sulfate was observed. This indicates that in the absence ofsufficient nitrate elemental sulfur is the only end-product ofsulfide oxidation (Eq. 1). Complete oxidation of sulfide tosulfate is only achieved when excess electron acceptor isavailable (Eq. 2):
HS− + 0.4 NO3 + 1.4 H+ → S° + 0.2 N2 + 1.2 H2O (1)
HS− + 1.6 NO3 + 0.6 H+ → SO421 + 0.8 N2 + 0.8 H2O
(2)
Studies with Produced Water Enrichments
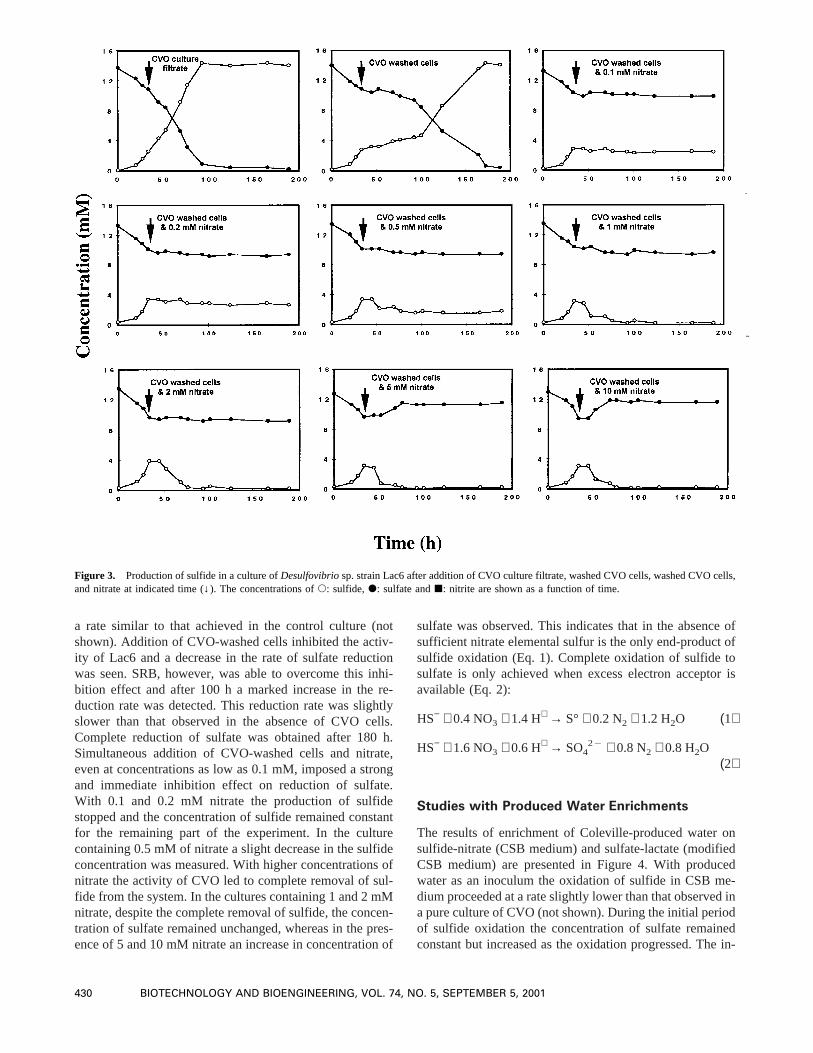
The results of enrichment of Coleville-produced water onsulfide-nitrate (CSB medium) and sulfate-lactate (modifiedCSB medium) are presented in Figure 4. With producedwater as an inoculum the oxidation of sulfide in CSB me-dium proceeded at a rate slightly lower than that observed ina pure culture of CVO (not shown). During the initial periodof sulfide oxidation the concentration of sulfate remainedconstant but increased as the oxidation progressed. The in-
Figure 3. Production of sulfide in a culture ofDesulfovibriosp. strain Lac6 after addition of CVO culture filtrate, washed CVO cells, washed CVO cells,and nitrate at indicated time (↓). The concentrations ofs: sulfide, d: sulfate andj: nitrite are shown as a function of time.
430 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 74, NO. 5, SEPTEMBER 5, 2001
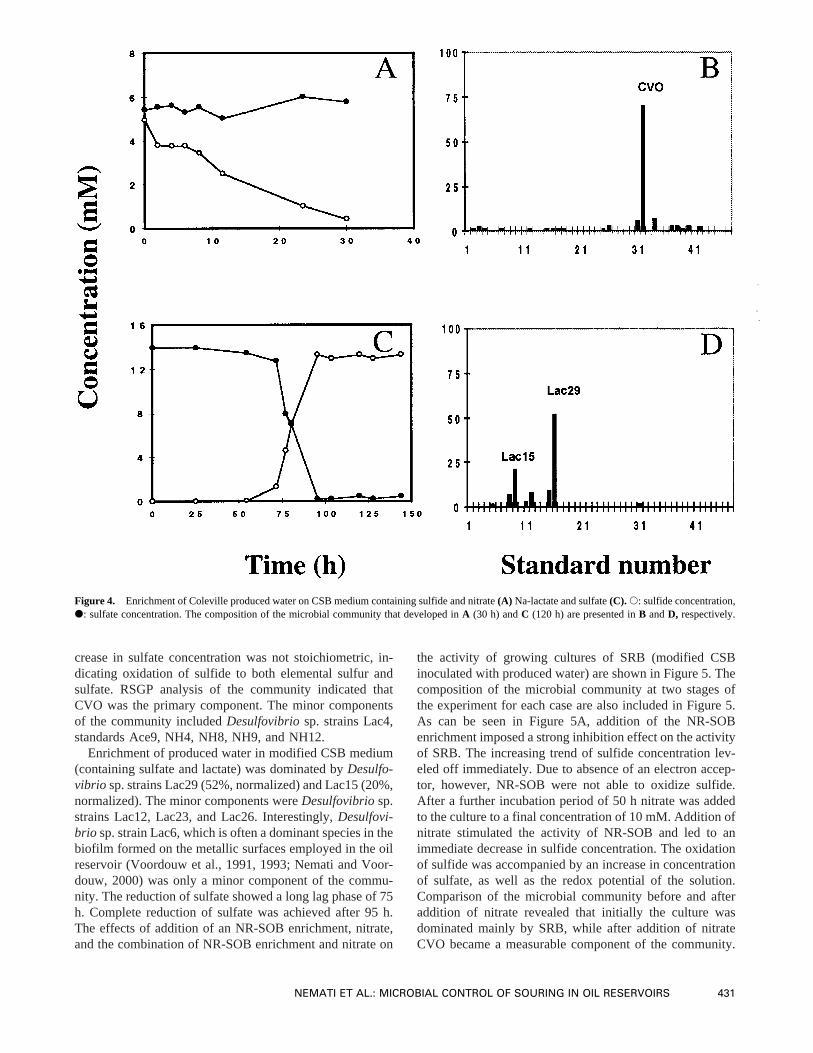
crease in sulfate concentration was not stoichiometric, in-dicating oxidation of sulfide to both elemental sulfur andsulfate. RSGP analysis of the community indicated thatCVO was the primary component. The minor componentsof the community includedDesulfovibriosp. strains Lac4,standards Ace9, NH4, NH8, NH9, and NH12.
Enrichment of produced water in modified CSB medium(containing sulfate and lactate) was dominated byDesulfo-vibrio sp. strains Lac29 (52%, normalized) and Lac15 (20%,normalized). The minor components wereDesulfovibriosp.strains Lac12, Lac23, and Lac26. Interestingly,Desulfovi-brio sp. strain Lac6, which is often a dominant species in thebiofilm formed on the metallic surfaces employed in the oilreservoir (Voordouw et al., 1991, 1993; Nemati and Voor-douw, 2000) was only a minor component of the commu-nity. The reduction of sulfate showed a long lag phase of 75h. Complete reduction of sulfate was achieved after 95 h.The effects of addition of an NR-SOB enrichment, nitrate,and the combination of NR-SOB enrichment and nitrate on
the activity of growing cultures of SRB (modified CSBinoculated with produced water) are shown in Figure 5. Thecomposition of the microbial community at two stages ofthe experiment for each case are also included in Figure 5.As can be seen in Figure 5A, addition of the NR-SOBenrichment imposed a strong inhibition effect on the activityof SRB. The increasing trend of sulfide concentration lev-eled off immediately. Due to absence of an electron accep-tor, however, NR-SOB were not able to oxidize sulfide.After a further incubation period of 50 h nitrate was addedto the culture to a final concentration of 10 mM. Addition ofnitrate stimulated the activity of NR-SOB and led to animmediate decrease in sulfide concentration. The oxidationof sulfide was accompanied by an increase in concentrationof sulfate, as well as the redox potential of the solution.Comparison of the microbial community before and afteraddition of nitrate revealed that initially the culture wasdominated mainly by SRB, while after addition of nitrateCVO became a measurable component of the community.
Figure 4. Enrichment of Coleville produced water on CSB medium containing sulfide and nitrate(A) Na-lactate and sulfate(C). s: sulfide concentration,d: sulfate concentration. The composition of the microbial community that developed inA (30 h) andC (120 h) are presented inB andD, respectively.
NEMATI ET AL.: MICROBIAL CONTROL OF SOURING IN OIL RESERVOIRS 431
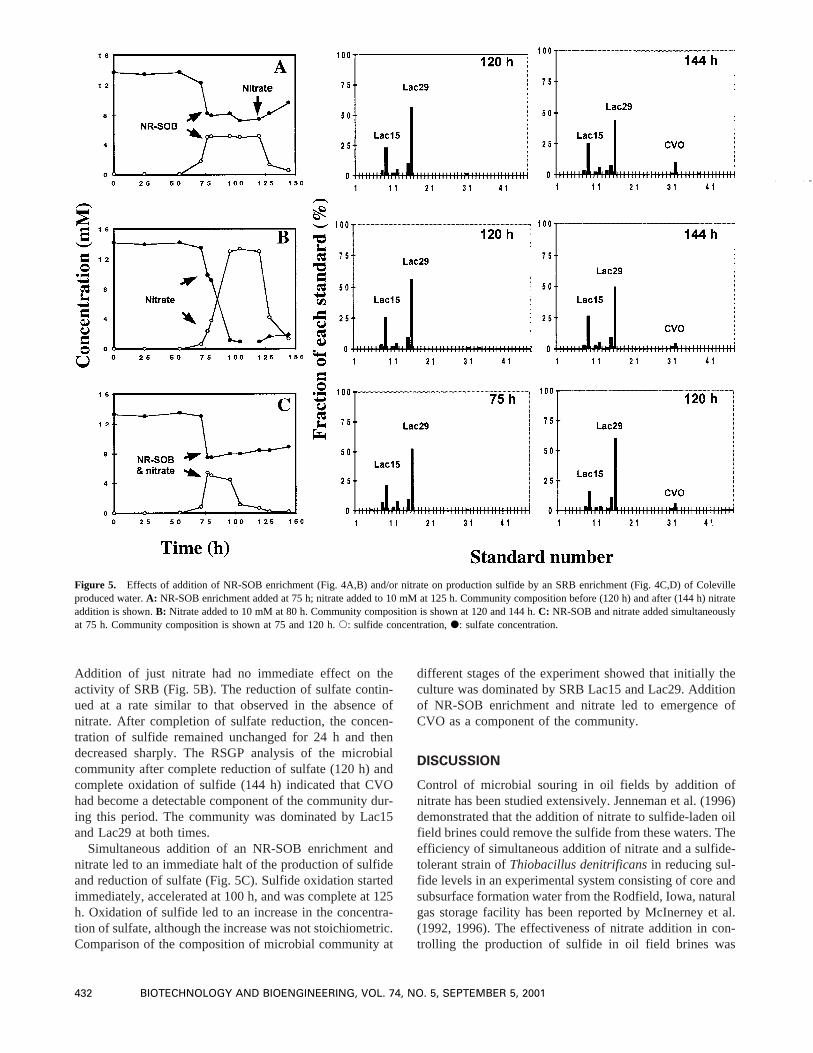
Addition of just nitrate had no immediate effect on theactivity of SRB (Fig. 5B). The reduction of sulfate contin-ued at a rate similar to that observed in the absence ofnitrate. After completion of sulfate reduction, the concen-tration of sulfide remained unchanged for 24 h and thendecreased sharply. The RSGP analysis of the microbialcommunity after complete reduction of sulfate (120 h) andcomplete oxidation of sulfide (144 h) indicated that CVOhad become a detectable component of the community dur-ing this period. The community was dominated by Lac15and Lac29 at both times.
Simultaneous addition of an NR-SOB enrichment andnitrate led to an immediate halt of the production of sulfideand reduction of sulfate (Fig. 5C). Sulfide oxidation startedimmediately, accelerated at 100 h, and was complete at 125h. Oxidation of sulfide led to an increase in the concentra-tion of sulfate, although the increase was not stoichiometric.Comparison of the composition of microbial community at
different stages of the experiment showed that initially theculture was dominated by SRB Lac15 and Lac29. Additionof NR-SOB enrichment and nitrate led to emergence ofCVO as a component of the community.
DISCUSSION
Control of microbial souring in oil fields by addition ofnitrate has been studied extensively. Jenneman et al. (1996)demonstrated that the addition of nitrate to sulfide-laden oilfield brines could remove the sulfide from these waters. Theefficiency of simultaneous addition of nitrate and a sulfide-tolerant strain ofThiobacillus denitrificansin reducing sul-fide levels in an experimental system consisting of core andsubsurface formation water from the Rodfield, Iowa, naturalgas storage facility has been reported by McInerney et al.(1992, 1996). The effectiveness of nitrate addition in con-trolling the production of sulfide in oil field brines was
Figure 5. Effects of addition of NR-SOB enrichment (Fig. 4A,B) and/or nitrate on production sulfide by an SRB enrichment (Fig. 4C,D) of Colevilleproduced water.A: NR-SOB enrichment added at 75 h; nitrate added to 10 mM at 125 h. Community composition before (120 h) and after (144 h) nitrateaddition is shown.B: Nitrate added to 10 mM at 80 h. Community composition is shown at 120 and 144 h.C: NR-SOB and nitrate added simultaneouslyat 75 h. Community composition is shown at 75 and 120 h.s: sulfide concentration,d: sulfate concentration.
432 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 74, NO. 5, SEPTEMBER 5, 2001

demonstrated by a field test conducted at Coleville in Sas-katchewan, Canada (Jenneman et al., 1997). Continuous ad-dition of 500 ppm of ammonium nitrate to injected brineover a period of 50 days resulted in complete removal ofsulfide at one of the two injectors employed, as well as50–60% reduction of sulfide levels in the co-produced brineat two adjacent producing wells. Using RSGP, it was shownthat the decrease in sulfide level was accompanied by asignificant increase in population of CVO, previously iso-lated from Coleville brine (Telang et al., 1997).
Several mechanisms have been proposed previously toexplain the decrease in biogenic production of sulfide in oilfields after addition of nitrate. These include the ability ofresident nitrate-reducing bacteria to outcompete SRB forcommon electron donors (Sandbeck and Hitzman, 1995),the preferential use of nitrate as an electron acceptor bySRB (Dalsgaard and Bak, 1994), the oxidation of sulfide bychemolithotrophic bacteria such asThiobacillusspecies orThiomicrospirasp. strain CVO, which could use nitrate aselectron acceptor (McInerney et al., 1996; Telang et al.,1997), and the inhibition of SRB activity by intermediateproducts of nitrate reduction such as nitrite or nitrous oxide(Reinsel et al., 1996). The results of the present study showthat two mechanisms are involved in microbial control ofbiogenic production of sulfide. First, addition of CVO and asmall amount of nitrate (0.1 mM in a batch system) imposesa strong and immediate inhibition effect on the activity ofSRB and prevents the production of H2S by these bacteria.Second, the metabolic activity of NR-SOB in the presenceof sufficient nitrate removes the H2S already present in thesystem. In this work the inhibition effect of CVO on theactivity of SRB was observed in the cultures initially con-taining a high level of Na-lactate in which the system wasnot electron-donor limited. In addition, RSGP analysis ofmicrobial community in the co-cultures revealed that evenafter completion of sulfide oxidation, SRB were still thedominant community component (NR-SOB/SRB: 1/2.5–1/15). Moreover, CVO is an autotroph that uses only CO2 asthe carbon source (Gevertz et al., 2000). Thus, the cessationof sulfate reduction is not due to competition of CVO andSRB for Na-lactate as a common electron donor. Owing tothe fact that the production of H2S by Lac6 in the presenceand absence of nitrate was similar (Fig. 1A), the possibilityof preferential use of nitrate by SRB can also be excluded.We have also shown that the addition of a CVO culturefiltrate did not have a significant effect on biogenic produc-tion of H2S by Lac6 (Fig. 3). AlthoughDesulfovibriosp.Strain Lac6 is unaffected by nitrate, its activity is inhibitedby very low concentration of nitrite (Nemati et al., 2001).However, monitoring the nitrite concentration revealed thatthe concentration of nitrite did not increase to a measureablelevel after addition of CVO to a Lac6 culture and during thelag phase in activity of CVO, while a strong inhibition effecton reduction of sulfate was observed (Fig. 1B). Nitrite wasonly formed as a result of CVO metabolism when excessnitrate (5 mM or more) was added. Hence, metabolites ofCVO, for example, intermediate products of nitrate reduc-
tion (especially nitrite), were not responsible for the imme-diate inhibition of H2S production by Lac6.
The sensitivity of SRB to environmental redox potentialis well documented. In culturing the SRB it is a prerequisitethat the redox potential of the environment starts at around−100 mV. In fact, the presence of a redox-poising agentsuch as H2S is a necessity for successful cultivation of SRB(Postgate, 1984). Activity of SRB and further production ofH2S by these bacteria could reduce the redox potential tovalues as low as −400 mV (Reinsel et al., 1996). On thecontrary, the activity of NR-SOB and consequent removalof H2S from the environment is accompanied by an increaseof the redox potential. This phenomenon has been reportedby previous researchers (Mueller, 1994; Reinsel et al., 1996)and was noticed in the present work. Therefore, the sup-pression of biogenic production of H2S could be attributedto the fact that after addition of CVO and nitrate, due tometabolic activity of CVO, the environmental redox poten-tial is increased to levels which are inhibitory for SRB. Thisincrease in environmental redox potential can be achieved ata very low ratio of CVO to Lac6 cells (estimated at 1:70) ata very low concentration of nitrate (0.1 mM).
The results of our study with cultures enriched from pro-duced water confirmed that the microbial control of bio-genic production of H2S in a consortium of bacteria is gov-erned by mechanisms similar to those observed in pure cul-tures. In other words, addition of an NR-SOB enrichmentimposes an immediate inhibition effect on the activity ofSRB which led to a decrease or cessation of sulfide produc-tion (Fig. 5A). Addition of nitrate alone did not inhibit SRBactivity (Fig. 5B) because an inhibitory number of CVOcells was not yet present. In the presence of an electronacceptor such as nitrate, the NR-SOB population is able tooxidize sulfide already present in the system and as a resulta significant decrease in the level of H2S is observed (Fig.5A,B,C).
In conclusion, the successful microbial control of H2Slevel in an oil reservoir is crucially dependent on the simul-taneous presence of NR-SOB (either an indigenous or aninjected population) and nitrate in the environment. Addi-tion of nitrate in the absence of NR-SOB has no effect onthe activity of SRB and biogenic production of sulfide.Equally, the activity of NR-SOB with respect to oxidationof sulfide depends significantly on the availability of anelectron acceptor such as nitrate. In the absence of an elec-tron acceptor NR-SOB are not able to derive energy fromoxidation of sulfide which leads to their exclusion from theenvironment. Addition of nitrate to oil field production wa-ter may lead to induction of a sufficient amount of NR-SOB.If in a system induction of NR-SOB cannot be achieved bynitrate injection, then a pregrown NR-SOB population mustbe added to achieve immediate inhibition of sulfide produc-tion by SRB and a rapid removal of sulfide by microbialnitrate-dependent sulfide oxidation.
The authors thank Pat McCarron and Andrew Richardson fromPetrovera Resources for providing production water samples.
NEMATI ET AL.: MICROBIAL CONTROL OF SOURING IN OIL RESERVOIRS 433

The technical assistance of A. J. Telang (Department of Biologi-cal Sciences, University of Calgary) is gratefully acknowledged.
References
APHA (American Public Health Association). 1992. Standard methods forthe examination of water and waste water, 18 ed. Washington, DC.
Bakke RB, Rivedal B, Mehan S. 1992. Oil reservoir biofouling control.Biofouling 6:53–60.
Cord-Ruwisch R. 1985. A quick method for determination of dissolved andprecipitated sulfides in cultures of sulfate-reducing bacteria. J Micro-biol Meth 4:33–36.
Dalsgaard T, Bak F. 1994. Nitrate reduction in a sulfide-reducing bacte-rium, Desulfovibrio desulfuricans,isolated from rice paddy soil, sul-fide inhibition, kinetics and regulation. Appl Environ Microbiol 60:291–297.
Gevertz D, Telang AJ, Voordouw G, Jenneman GE. 2000. Isolation andcharacterization of strains CVO and FWKO B: two novel nitrate-reducing, sulfide oxidizing bacteria isolated from oil field brine. ApplEnviron Microbiol 66:2491–2501.
Jenneman GE, McInerney MJ, Knapp RM. 1986. Effect of nitrate onbiogenic sulfide production. Appl Environ Microbiol 51:1205–1211.
Jenneman GE, Wright M, Gevertz D. 1996. Sulfide bioscavenging of sourproduced water by natural microbial population. Proceeding of 3rd
International Petroleum Environmental Conference. Albuquerque,NM.
Jenneman GE, Moffitt PD, Bala GA, Webb RH. 1997. Field demonstrationof sulfide removal in reservoir brine by bacteria indigenous to a Ca-nadian reservoir. Proceeding of the SPE Annual Technical Conferenceand Exhibition. San Antonio, TX.
McInerney MJ, Bhupathiraju VK, Sublette KL. 1992. Evaluation of amicrobial method to reduce hydrogen sulfide levels in a porous rockbiofilm. J Indust Microbiol 11:53–58.
McInerney MJ, Wofford NQ, Sublette KL. 1996. Microbial control ofhydrogen sulfide production in a porous medium. Appl Biochem Bio-technol 57/58:933–944.
Montgomery AD, McInerney MJ, Sublette KL. 1990. Microbial control ofthe production of hydrogen sulfide by sulfate reducing bacteria. Bio-technol Bioeng 35:533–539.
Mueller RF. 1994. Microbial dynamics in souring oil reservoirs. PhD dis-sertation, Montana State University, Bozeman, MT.
Nemati M, Voordouw G. 2000. Identification and characterization of sul-fate reducing bacteria involved in microbially influenced corrosion inoil fields. 55th Annual Corrosion Conference. Orlando, FL. Paper 126.
Nemati M, Mazutinec T, Jenneman GE, Voordouw G. 2001. Control ofbiogenic H2S production by nitrite and molybdate. Ind Microbiol Bio-technol (in press).
Postgate JR. 1984. The sulfate-reducing bacteria, 2nd ed. Cambridge:Cambridge University Press.
Reinsel MA, Sears JT, Stewart PS, McInerney MJ. 1996. Control of mi-crobial souring by nitrate, nitrite or glutaraldehyde injection in a sandstone column. J Ind Microbiol 17:128–136.
Sandbeck KA, Hitzman DO. 1995. Biocompetitive exclusion technology.A field system to control reservoir souring and increase production. In:Byrant R, Sublette K, editors. Proceeding 5th international conferenceon microbial enhanced oil recovery and related biotechnology for solv-ing environmental problems. Richardson, TX. p 311–320.
Shock EL. 1988. Organic acid metastability in sedimentary basins. Geol-ogy 16:886–890.
Telang AJ, Voordouw G, Ebert S, Sifeldeen N, Foght JM, Fedorak PM,Westlake DWS. 1994. Characterization of the diversity of sulfate-reducing bacteria in soil and mining waste water environments bynucleic acid hybridization techniques. Can J Microbiol 40:955–964.
Telang AJ, Ebert S, Foght JM, Westlake DWS, Jenneman GE, Gevertz D,Voordouw G. 1997. Effect of nitrate injection on the microbial com-munity in an oil field as monitored by reverse sample genome probing.Appl Environ Microbiol 63:1785–1793.
Telang AJ, Jenneman GE, Voordouw G. 1999. Sulfur cycling in mixedcultures of sulfide-oxidizing and sulfate- or sulfur-reducing oil fieldbacteria. Can J Microbiol 45:905–913.
Voordouw G. 1998. Reverse sample genome probing of microbial com-munity dynamics. ASM News 64:627–633.
Voordouw G, Strang JD, Wilson FR. 1989. Organization of genes encod-ing [Fe] hydrogenase inDesulfovibrio vulgarissubsp.oxamicusMon-ticello. J Bacteriol 171:3881–3889.
Voordouw G, Voordouw JK, Karkhoff-Schweizer RR, Fedorok PM, West-lake DWS. 1991. Reverse sample genome probing, a new techniquefor identification of bacterial in environmental samples by DNA hy-bridization, and its application to the identification of sulfate reducingbacteria in oil field samples. Appl Environ Microbiol 57:3070–3078.
Voordouw G, Voordouw JK, Jack TR, Foght JM, Fedorak PM, WestlakeDWS. 1992. Identification of distinct communities of sulfate-reducingbacteria in oil fields by reverse sample genome probing. Appl EnvironMicrobiol 58:3542–3552.
Voordouw G, Shen Y, Harrington CS, Telang AJ, Jack TR, WestlakeDWS. 1993. Quantitative reverse sample genome probing of microbialcommunities and its application to oil field production waters. ApplEnviron Microbiol 59:4101–4114.
Voordouw G, Armstrong SM, Reimer MF, Fouts B, Telang AJ, Shen Y,Gevertz D. 1996. Characterization of 16S rRNA genes from oil fieldmicrobial communities indicates the presence of a variety of sulfate-reducing, fermentative, and sulfide-oxidizing bacteria. Appl EnvironMicrobiol 62:1623–1629.
Wright M, Jenneman GE, Gevertz D. 1997. Effect of nitrate on sulfidebio-scavenging by indigenous bacteria in produced brines from WestTexas oil fields. In: Proceedings of the 4th International PetroleumEnvironmental Conference. San Antonio, TX.
434 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 74, NO. 5, SEPTEMBER 5, 2001