peptidomic analysis of human acquired enamel pellicle
TRANSCRIPT

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1107ORIGINAL RESEARCHORIGINAL RESEARCH
Copyright © 2007 John Wiley & Sons, Ltd.
BIOMEDICAL CHROMATOGRAPHY
Biomed. Chromatogr. 21: 1107–1117 (2007)
Published online 21 May 2007 in Wiley InterScience
(www.interscience.wiley.com) DOI: 10.1002/bmc.830
Peptidomic analysis of human acquired enamel pellicle
Rui Vitorino,1,2 Maria João Calheiros-Lobo,3 Jason Williams,4 António J. Ferrer-Correia,1 Kenneth B. Tomer,4José A. Duarte,2 Pedro M. Domingues1 and Francisco M. L. Amado1*1Department of Chemistry, University of Aveiro, Aveiro, Portugal2CIAFEL, Faculty of Sport, University of Porto, Porto, Portugal3High-School of Health-North, Medical School of Dentistry, Portugal4Laboratory of Structural Biology, National Institute of Environmental Health Sciences, National Institutes of Health, DHHS, Research Triangle Park,
NC 27709, USA
Received 12 December 2006; revised 6 February 2007; accepted 7 February 2007
ABSTRACT: Human acquired enamel pellicle is the result of a selective interaction of salivary proteins and peptides with the
tooth surface. In the present work, the characterization of the peptides as well as the type of interactions established with the
enamel surface was performed. Peptides from in vivo bovine enamel implants in the human oral cavity were sequentially
extracted using guanidine and trifluoroacetic acid solutions and the fractions obtained were analysed by LC-MS and LC-MS/MS.
Based on the LC-MS data, six phosphorylated peptides were identified in an intact form, strongly adsorbed to the enamel surface.
Data from the LC-MS/MS analyses allowed us to identified 30 fragment peptides non-covalently bonded to enamel [basic proline-
rich proteins, histatins (1 and 3) and acidic proline-rich protein classes]. The tandem mass spectrometry experiments showed the
existence of a pattern of amide bond cleavage for the different identified peptide classes suggesting a selective proteolytic activity.
For histatins, a predominance of cleavage at Arg, Lys and His residues was observed, while for basic proline-rich proteins,
cleavage at Arg and Pro residues prevailed. In the case of acidic proline-rich proteins, a clearly predominance of cleavage of the
Gln–Gly amide bond was evident. Copyright © 2007 John Wiley & Sons, Ltd.
KEYWORDS: acquired enamel pellicle; whole saliva; LC-MS/MS; salivary peptides; proteolytic activity
*Correspondence to: F. M. L. Amado, Department of Chemistry,
University of Aveiro, 3810-193, Aveiro, Portugal.
E-mail: [email protected]
Abbreviations used: PRP, proline-rich protein; TFA, trifluoroacetic
acid.
Contract/grant sponsor: Intramural Research Program of the NIH,
National Institute of Environmental Health Sciences.
Contract/grant sponsor: Fundação para a Ciência e Tecnologia; Con-
tract/grant number: SFRH/BPD/14968/2004 and POCTI/QUI/5890/
2004.
INTRODUCTION
The acquired enamel pellicle is a protein film originat-
ing in the continuous exposure of enamel to whole
saliva. The adsorption of proteins and peptides and
subsequent pellicle formation is considered a dynamic
process influenced by several factors inherent to the
individual, such as the circadian cycle, oral microflora,
proteolytic activity and the tooth’s physical and chemi-
cal properties, as well as the location of teeth in the
mouth (Lendenmann et al., 2000; Carlen et al., 1998;
Vacca Smith and Bowen, 2000; Rykke and Sonju, 1991;
Li et al., 2003). As the enamel pellicle constitutes an
interface between teeth and the oral environment, it is
expected that its origin and composition should be of
major importance for the function it plays within the
oral cavity. The acquired enamel pellicle has been pro-
posed to be involved in several oral functions, such as
lubrication to prevent abrasive damage, acting as a selec-
tive permeable barrier that regulates the mineralization/
demineralization processes, and control of the dental
plaque microbial flora composition (Lendenmann et al.,
2000; Carlen et al., 1998; Vacca Smith and Bowen,
2000; Rykke and Sonju, 1991; Li et al., 2003). Using dif-
ferent methodologies, several authors have studied the
acquired enamel pellicle composition, which allowed
identification of mucins MG1 and MG2, amylase, albu-
min, IgA, S-IgA, proline-rich proteins (PRPs), cystatins,
lysozyme, carbonic anhydrase, lactoferrin and histatins
as major pellicle components (Carlen et al., 1998; Vacca
Smith and Bowen, 2000; Rykke and Sonju, 1991; Li
et al., 2003, 2004; Schupbach et al., 2001; Yao et al.,
2001, 2003; Vitorino et al., 2004a; Leinonen et al., 1999).
It is assumed that the formation of the acquired
enamel pellicle takes place in two stages, the first one
being characterized by an almost instantaneous adsorp-
tion of proteins to the enamel surface (Carlen et al.,
1998; Jensen et al., 1991; Lamkin et al., 1996). In this
phase, the initial adsorption of salivary components
occurs by electrostatic attraction forces developed

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
1108 R. Vitorino et al.ORIGINAL RESEARCH
between the charged groups of the macromolecules
and the phosphate and calcium ions of enamel. Indeed,
calcium ions present in the crystal construction of
enamel showed a stronger tendency to migrate to saliva
than did phosphate ions, contributing to the creation of
a double-layer: negative charges of phosphate ions on
the tooth surface and the opposite charged calcium ions
at the boarder phase (Yin et al., 2003; Gorbunoff and
Timasheff, 1984; Moreno et al., 1984). Therefore, basic
proteins are bound primarily by electrostatic interac-
tions between the amino groups and the phosphate ions
of the hydroxyapatite, whereas the acidic proteins are
bound specifically by complexation of their carboxyl
and/or phosphate groups to calcium sites. The predomi-
nant groups of salivary proteins which show selective
adsorption to enamel include small components such as
acidic PRPs, statherin and histatins, usually thought of
as pellicle precursors (Jensen et al., 1991; Moreno et al.,
1984; Bennick et al., 1983). A strong adsorption is
exhibited by the phosphorylated forms of those small
proteins. It has been described that these components
bind immediately when in contact with the enamel sur-
face and do not increase thereafter, while molecules
such as histatins bind slowly and progressively over
time (Vacca Smith and Bowen, 2000). Those peptides
are capable of phosphate or carboxyl-terminal anion
exchange reactions with the enamel surface as described
for statherin (Hay and Moreno, 1979; Schlesinger and
Hay, 1977; Stayton et al., 2003). In association with the
process of adsorption to hydroxyapatite, soluble pellicle
precursor proteins are transformed into a less soluble
or even insoluble protein film. This insolubility may
also arise from post-secretory processing of pellicle pro-
teins, which includes cross-linking and proteolysis (Yao
et al., 1999, 2000; Lamkin et al., 2001). In fact, in vitro
studies showed the existence of a proteolytic pattern
for adsorbed salivary proteins (Lamkin et al., 2001) and
that proteolysis can occur after protein adsorption to
hydroxyapatite, as shown by Bennick et al. (1983) and
Jensen et al. (1991). To date, several authors have stud-
ied salivary peptides in order to elucidate possible
cleavage pathways and post-translational modifications
that occur after its secretion from salivary glands. From
these studies, Castagnola and coworkers described a
possible cascade of fragmentation on histatins which
follows a trypsin-like pathway, and characterized a
complex of several isoforms from basic PRPs and acidic
PRPs (Castagnola et al., 2004; Messana et al., 2004;
Inzitari et al., 2005). In another study, Hardt et al.
(2005) characterized the peptidome of parotid saliva
by ultrafiltration, finding mainly peptides belonging to
acidic PRPs and histatin 1 classes.
Because of the importance of salivary peptides on
pellicle formation, the main goal of the present study is
the in vivo elucidation of peptide pellicle composition
and how salivary peptides interact with the enamel
surface. To achieve this, a combination of LC-MS and
LC-MS/MS was used for the separation and charac-
terization of human acquired enamel pellicle peptides
using a sequential extraction with guanidine (6 M) and
trifluoroacetic acid (TFA, 2%) from enamel implants.
MATERIAL AND METHODS
Chemicals. HPLC-grade acetonitrile (Riedel, Seelze, Germany),
TFA (Fluka, Buchs, Switzerland), guanidine (Sigma-Aldrich,
Madrid, Spain) and Milli-Q grade water were used.
In situ pellicle formation. For in situ pellicle formation,
two healthy, non-medicated male subjects (aged 24 and 28)
were selected according to visual oral examination performed
by an experienced dentist. The subjects showed no presence
of caries or signs of other oral pathologies. The subjects
refrained from eating and drinking for 3 h before and during
pellicle formation. Small rectangular pieces of enamel (about
10 mm long and 4 mm high) were prepared from labial
surfaces of bovine incisors (Hannig et al., 2005). The enamel
pieces were bonded on the labial surfaces of the upper and
lower first molars according to the accepted adhesion pro-
tocols for tooth restoration, as close as possible to the
gingival. The enamel implants were then thoroughly cleaned
with fluoride-free paste with a synthetic brush and copious
rinsed with an air–water spray to eliminate any possible con-
tamination with saliva previous to pellicle formation. After
2 h, the enamel pieces were removed from the mouth with
proper bracket debonding forceps used in orthodontics.
Peptide extraction. Sequential extractions were performed
as described previously for in vitro experimental conditions
(Vitorino et al., 2005), using solutions of guanidine (6 M)
and TFA (2%). Briefly, after enamel implant wash with
water, 300 µL of guanidine (6 M) were added to the enamel
pieces. Pellicle peptide extraction was performed by vigorous
vortexing for 120 s. Then the extract was centrifuged at
6000 rpm, 4°C, for 15 min, the resultant supernatant was
removed and the enamel pieces were further washed with
water under vigorous vortexing followed by a spin at
6000 rpm. After this, 300 µL of 2% TFA were added to
enamel pieces and after 2 h the suspension was centrifuged
and the supernatant recovered for further analysis. Adsorbed
salivary peptides were isolated by passing the supernatant
extracts through a 30 kDa cut-off membrane (Centricom 30,
Millipore, USA). All fractions were subsequently analysed by
LC-MS. Blanks were obtained with bovine enamel pieces
treated with the same sequential extraction procedure without
performing the incubation step.
Liquid chromatography and mass spectrometry analysis.LC-MS was performed using a Waters 2690 (Alliance, Waters
Corporation, Saint-Quentin, France) HPLC system at a flow
rate of 0.30 mL/min connected to a splitter (Acurate, LC
Packings, Amsterdam, Netherlands) to obtain a flow rate of
15 µL/min, compatible with the column used and the mass
spectrometer (QTOF2, Micromass, UK) electrospray source
operated in the positive mode at 3.0 kV. The HPLC column

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1109ORIGINAL RESEARCH
was a Discovery C18
15 ! 0.32 mm, 5 µm particle size
(Supelco, Sigma-Aldrich, Madrid, Spain). The eluents were:
(A) water–0.05% TFA and (B) acetonitrile–0.05% TFA).
Peptides were separated using the following gradient pro-
gram: 0–5 min, 5% B; 5–60 min, linear increase to 44% B;
60–70 min, linear increase to 70% B. For each run 5 µg of
peptide extract from each subject were loaded onto the system.
LC-MS/MS analyses were performed using an Agilent
Nanoflow Proteomics Solution (Agilent Technologies, Cali-
fornia, USA). Twenty microlitres of each sample (5 µg of
peptide extract) were injected onto a C18
trapping column
(Zorbax 300SB-C18
, 5 µm particle size, 5 ! 0.3 mm, Agilent
Technologies) using an autosampler. The sample was washed
over the trapping column for 10 min with 95% buffer A
(water–0.1% formic acid), 5% buffer B (acetonitrile–0.1%
formic acid) at a flow rate of 10 µL/min. The flow was then
reversed over the trapping column, and the sample was eluted
onto a 150 ! 75 µm Zorbax 300SB capillary analytical C18
column with 3.5 µm particle size (Agilent Technologies) at a
flow rate of 0.3 µL/min. The analytical column was directly
plumbed into a nanoelectrospray ionization source (operated
at 1.9 kV), which was sampled into the quadrupole ion trap
mass spectrometer. A linear gradient of 5% buffer B to 65%
buffer B was run over a period of 65 min. The column was
then washed with a 10 min gradient from 65 to 95% buffer B,
followed by an 10 min hold at 95% buffer B. The column was
then re-equilibrated in 5% buffer B prior to future analyses.
Peptides were analysed by data-dependent acquisition of full-
scan mass spectra and MS/MS scans for the most abundant
ions. Spectra were collected in Standard-Enhanced MS mode,
with automated switching for MS/MS acquisition in Ultrascan
mode with automated selection of precursor ions.
Peptide identification
The data obtained from the LC/MS analyses were processed
using Masslynx software (Micromass, Manchester, UK). Spec-
tra corresponding to the detected peaks were deconvoluted
using MaxEnt 1 and 3 programs to yield monoisotopic masses.
Experimental masses were further compared with available
known theoretical peptide masses in the Swiss-Prot (http://
www.expasy.ch) and/or NCBI (http://www4.ncbi.nlm.nih.gov/
entrez/query.fcgi) databases. Identifications were considered
positive when differences between theoretical and experimen-
tal mass values were less than to 100 ppm.
The LC-MS/MS obtained data were processed by search-
ing the MS/MS spectra against the NCBI non-redundant
database (NCBInr, 3 January 2005) in Homo sapiens using
SpectrumMill software (Agilent Technologies, California,
USA) without enzyme specified or modifications. Mass toler-
ance was set to 0.15 Da for MS and MS/MS. The algorithm
used to create peak lists was Data Extractor, which is from
Agilent. The settings for this extractor are: all scans were
processed; scans were merged if they were within ±1.5 m/z
and ±240 s. MH+ values were allowed from 300 to 6000 and
sequence tag lengths from MS/MS were required to be >2.
The precursor charge assignment was set to ‘find’ with a
maximum charge of 6, a minimum signal/noise of 10 and
the ‘find 12C’ function enabled. Positive identifications
where accepted when the confidence level was greater than
95%.
RESULTS
LC-MS has been used for the analysis of salivary
peptides (Hardt et al., 2005; Vitorino et al., 2004b;
Messana et al., 2003; Castagnola et al., 2003). This
approach has allowed the identification of peptides
belonging to the basic PRPs, acidic PRPs, histatins and
statherins. As previously reported for in vivo and
in vitro experimental conditions (Vitorino et al., 2005),
we performed the analysis of the guanidine- and
TFA-extracted fractions of the acquired enamel pellicle
peptides (accounting for ca. 38.5 ± 3% of the total
adsorbed protein). In general, LC-MS experiments
show mainly peptides belonging to two classes: histatins
and proline-rich proteins. A large number of unidenti-
fied low-molecular-weight components were detected
on the guanidine extracts: It was also possible to detect,
although in trace amounts, the presence of the follow-
ing intact peptides: II-2, histatin 1, PRP3 and statherin
(Table 1). Some unidentified low-molecular-weight
components were also detected in the TFA extract, but
the same intact peptides found in the guanidine extract
(Fig. 1) predominate in this fraction. IB1 and PRP1
were also found with low relative abundance in this
fraction.
Table 1. Identified salivary peptides on acquired enamel pellicle by LC-MS on guanidine and TFA extracts. (a) Guanidine extract
and (b) TFA extract
Salivary Experimental Theoretical Acession Sequence
peptides MW MW pI number fraction
Histatin 1 4926.17 4926.23 9.1 123,134 a/b
Histatin 5 3035.53 3035.554 10.55 123,143 a/b
IB-1 9548.46 9588.51 10.1 131,006 b
II-2 7604.4 9.81 a/b
PRP1 15,514.2 15,514.1 4.63 131,008 b
PRP3 11,161.15 11,161 4.11 131,008 a/b
Statherin 5348.69 5378.45 6.64 134,947 b
a refers to guanidine extraction.
b refers to TFA extraction.

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
1110 R. Vitorino et al.ORIGINAL RESEARCH
peptide fragments (18 out of 30) were found only in the
guanidine extract, 12 were found in both fractions and
two were observed only in the TFA extract. Generally,
the low-molecular-weight peptides found in the TFA
extracts were of low relative abundances, as shown for
the peptide of m/z 1819.0 (6-fold increase on guanidine
extracts compared with TFA extracts, obtained from
the ratio of the ion currents for the ion of m/z 1819.0 in
both fractions) or for the peptide of m/z 1565.5 (25-fold
increase on guanidine extracts when compared with
TFA extracts).
Several peptides were identified in the MS/MS
experiments as fragments of basic PRPs, with diverse
points of cleavage along the main chain (Fig. 4). The
observed peptide fragments arise via fragmentations of
the C-terminal and of the middle region of the primary
sequence in the case of IB8a and IB9 and of the
middle region of the N-terminus of IB1. Owing to the
sequence similarity among different b-PRPs, some of
the fragments detected can derive also from other
b-PRPs. Similarly to basic PRPs, the fragmentation of
Figure 2. Total ion chromatogram of the small peptides on guanidine and TFA extract obtained by LC-MS/MS. A similar
chromatogram profile is observed for both extracts.
Figure 1. Total ion chromatograms of acquired enamel pellicle peptides on guanidine and TFA
extract obtained by 1D-LC-MS.
The bulk of the unknown low-molecular-weight com-
ponents (<2200 Da), which were observed as doubly
and triply charged ions, were detected at a retention
time between 30 and 35 min in the LC-MS experi-
ments. To achieve a better separation and consequently
an increase the number of possible identified peptides,
we used a longer, linear gradient program in the LC-
MS/MS experiments. The LC-MS/MS chromatograms
for both the guanidine and TFA extracts are similar
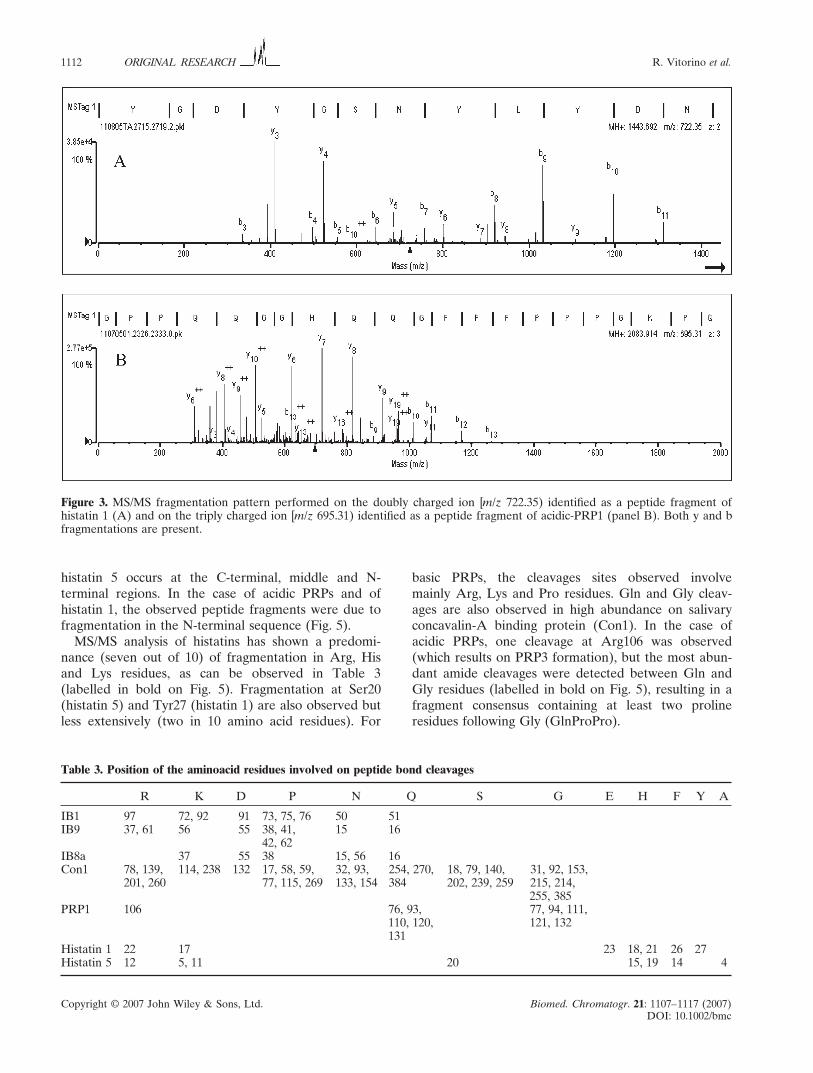
(Fig. 2). All the peptides detected were sequenced by
MS/MS and identified by automated interpretation of
the peptide fragmentation pattern. The CID experi-
ments performed on doubly charged ions led to the
observation of peaks corresponding to y and b fragment
ions while the experiments performed on peptides with
more than 2 charges led to the detection mainly of y
fragments (Fig. 3). As a result, 30 peptides were identi-
fied and assigned with high confidence as fragments
belonging to basic PRPs, acidic PRPs and histatins
(Table 2). Comparing the guanidine and TFA extracts,
it is possible to observe that most of the identified
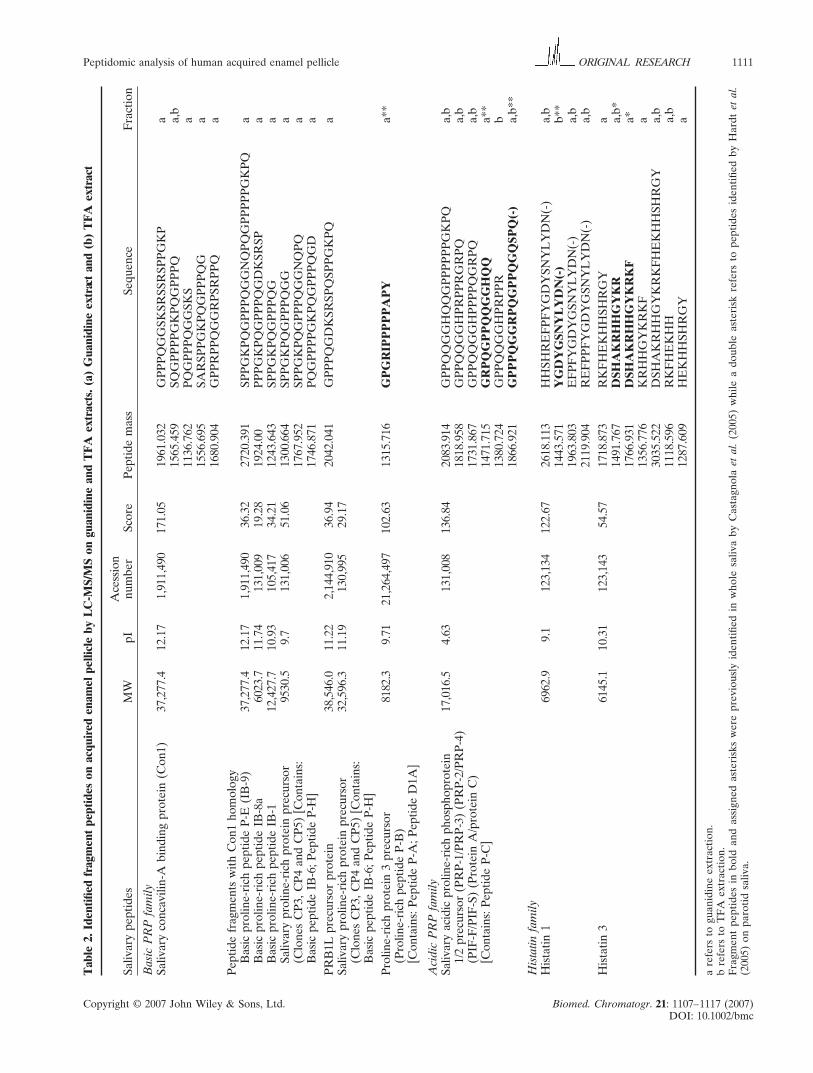
Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1111ORIGINAL RESEARCH
Tab
le 2.
Id
en
tifi
ed
fragm
en
t p
ep
tid
es o
n acq
uir
ed
en
am
el
pell
icle
b
y L
C-M
S/M
S o
n gu
an
idin
e an
d T
FA
extracts.
(a) G
uan
idin
e extract an
d (b
) T
FA
extract
Acessio
n
Sali
vary p
ep
tid
es
MW
pI
nu
mb
er
Sco
re
Pep
tid
e m
ass
Seq
uen
ce
Fractio
n
Basic
P
RP
fam
ily
Sali
vary co
ncavil
in-A
b
ind
ing p
ro
tein
(C
on
1)
37,2
77.4
12.1
71,9
11,4
90
171.0
51961.0
32
GP
PP
QG
GS
KS
RS
SR
SP
PG
KP
a
1565.4
59
SQ
GP
PP
PG
KP
QG
PP
PQ
a,b
1136.7
62
PQ
GP
PP
QG
GS
KS
a
1556.6
95
SA
RS
PP
GK
PQ
GP
PP
QG
a
1680.9
04
GP
PR
PP
QG
GR
PS
RP
PQ
a
Pep
tid
e fragm
en
ts w
ith
C
on
1 h
om
olo
gy
Basic
p
ro
lin
e-ric
h p
ep
tid
e P
-E
(IB
-9)
37,2
77.4
12.1
71,9
11,4
90
36.3
22720.3
91
SP
PG
KP
QG
PP
PQ
GG
NQ
PQ
GP
PP
PP
GK
PQ
a
Basic
p
ro
lin
e-ric
h p
ep
tid
e IB
-8a
6023.7
11.7
4131,0
09
19.2
81924.0
0P
PP
GK
PQ
GP
PP
QG
DK
SR
SP
a
Basic
p
ro
lin
e-ric
h p
ep
tid
e IB
-1
12,4
27.7
10.9
3105,4
17
34.2
11243.6
43
SP
PG
KP
QG
PP
PQ
Ga
Sali
vary p
ro
lin
e-ric
h p
ro
tein
p
recu
rso
r9530.5
9.7
131,0
06
51.0
61300. 6
64
SP
PG
KP
QG
PP
PQ
GG
a
(C
lon
es C
P3,
CP
4 an
d C
P5) [C
on
tain
s:
1767.9
52
SP
PG
KP
QG
PP
PQ
GG
NQ
PQ
a
Basic
p
ep
tid
e IB
-6;
Pep
tid
e P
-H
]1746.8
71
PQ
GP
PP
PG
KP
QG
PP
PQ
GD
a
PR
B1L
p
recu
rso
r p
ro
tein
38,5
46.0
11.2
22,1
44,9
10
36.9
42042.0
41
GP
PP
QG
DK
SR
SP
QS
PP
GK
PQ
a
Sali
vary p
ro
lin
e-ric
h p
ro
tein
p
recu
rso
r32,5
96.3
11.1
9130,9
95
29.1
7
(C
lon
es C
P3,
CP
4 an
d C
P5) [C
on
tain
s:
Basic
p
ep
tid
e IB
-6;
Pep
tid
e P
-H
]
Pro
lin
e-ric
h p
ro
tein
3 p
recu
rso
r8182.3
9.7
121,2
64,4
97
102.6
31315.7
16
GP
GR
IP
PP
PP
AP
Ya**
(P
ro
lin
e-ric
h p
ep
tid
e P
-B
)
[C
on
tain
s:
Pep
tid
e P
-A
; P
ep
tid
e D
1A
]
Acid
ic P
RP
fam
ily
Sali
vary acid
ic p
ro
lin
e-ric
h p
ho
sp
ho
pro
tein
17,0
16.5
4.6
3131,0
08
136.8
42083.9
14
GP
PQ
QG
GH
GP
PP
PP
PG
KP
Qa,b
1/2
p
recu
rso
r (P
RP
-1/P
RP
-3) (P
RP
-2/P
RP
-4)
1818.9
58
GP
PQ
QG
GH
PR
PP
RG
RP
Qa,b
(P
IF
-F
/PIF
-S
) (P
ro
tein
A
/pro
tein
C
)1731.8
67
GP
PQ
QG
GH
PP
PP
QG
RP
Qa,b
[C
on
tain
s:
Pep
tid
e P
-C
]1471.7
15
GR
PQ
GP
PQ
QG
GH
a**
1380.7
24
GP
PQ
QG
GH
PR
PP
Rb
1866.9
21
GP
PP
QG
GR
PQ
GP
PQ
GQ
SP
Q(-)
a,b
**
His
tati
n fam
ily
His
tatin
1
6962.9
9.1
123,1
34
122.6
72618.1
13
HH
SH
RE
FP
FY
GD
YS
NY
LY
DN
(-)
a,b
1443.5
71
YG
DY
GS
NY
LY
DN
(-)
b**
1963.8
03
EF
PF
YG
DY
GS
NY
LY
DN
(-)
a,b
2119.9
04
RE
FP
PF
YG
DY
GS
NY
LY
DN
(-)
a,b
His
tatin
3
6145.1
10.3
1123,1
43
54.5
71718.8
73
RK
FH
EK
HH
SH
RG
Ya
1491.7
67
DS
HA
KR
HH
GY
KR
a,b
*
1766.9
31
DS
HA
KR
HH
GY
KR
KF
a*
1356.7
76
KR
HH
GY
KR
KF
a
3035.5
22
DS
HA
KR
HH
GY
KR
KF
HE
KH
HS
HR
GY
a,b
1118.5
96
RK
FH
EK
HH
a,b
1287.6
09
HE
KH
HS
HR
GY
a
a refers to
gu
an
idin
e extractio
n.
b refers to
T
FA
extractio
n.
Fragm
en
t p
ep
tid
es in
b
old
an
d assig
ned
asteris
ks w
ere p
revio
usly
id
en
tifi
ed
in
w
ho
le sali
va b
y C
astagn
ola
et
al.
(2005) w
hil
e a d
ou
ble
asteris
k refers to
p
ep
tid
es id
en
tifi
ed
b
y H
ard
t et
al.
(2005) o
n p
aro
tid
sali
va.
Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
1112 R. Vitorino et al.ORIGINAL RESEARCH
Figure 3. MS/MS fragmentation pattern performed on the doubly charged ion [m/z 722.35) identified as a peptide fragment of
histatin 1 (A) and on the triply charged ion [m/z 695.31) identified as a peptide fragment of acidic-PRP1 (panel B). Both y and b
fragmentations are present.
Table 3. Position of the aminoacid residues involved on peptide bond cleavages
R K D P N Q S G E H F Y A
IB1 97 72, 92 91 73, 75, 76 50 51
IB9 37, 61 56 55 38, 41, 15 16
42, 62
IB8a 37 55 38 15, 56 16
Con1 78, 139, 114, 238 132 17, 58, 59, 32, 93, 254, 270, 18, 79, 140, 31, 92, 153,
201, 260 77, 115, 269 133, 154 384 202, 239, 259 215, 214,
255, 385
PRP1 106 76, 93, 77, 94, 111,
110, 120, 121, 132
131
Histatin 1 22 17 23 18, 21 26 27
Histatin 5 12 5, 11 20 15, 19 14 4
histatin 5 occurs at the C-terminal, middle and N-
terminal regions. In the case of acidic PRPs and of
histatin 1, the observed peptide fragments were due to
fragmentation in the N-terminal sequence (Fig. 5).
MS/MS analysis of histatins has shown a predomi-
nance (seven out of 10) of fragmentation in Arg, His
and Lys residues, as can be observed in Table 3
(labelled in bold on Fig. 5). Fragmentation at Ser20
(histatin 5) and Tyr27 (histatin 1) are also observed but
less extensively (two in 10 amino acid residues). For
basic PRPs, the cleavages sites observed involve
mainly Arg, Lys and Pro residues. Gln and Gly cleav-
ages are also observed in high abundance on salivary
concavalin-A binding protein (Con1). In the case of
acidic PRPs, one cleavage at Arg106 was observed
(which results on PRP3 formation), but the most abun-
dant amide cleavages were detected between Gln and
Gly residues (labelled in bold on Fig. 5), resulting in a
fragment consensus containing at least two proline
residues following Gly (GlnProPro).

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1113ORIGINAL RESEARCH
Figure 4. Amino acid sequences of basic salivary proline-rich peptides IB1, IB8a and
IB9 (top panel) and histatin 3 (bottom panel). The peptide fragments observed in the
LC-MS/MS experiment are aligned below the protein sequence. All peptide bond cleav-
age sites are indicated in bold and underlined. Phosphorylated sites are indicated in
italic bold and underlined. < represents a pyroglutamic residue. Diverse points of cleav-
age along the main chain of the peptides were observed for basic-PRP and Con1.
Figure 5. Amino acid sequences of salivary proline-rich phosphoprotein 1/2 (top panel),
histatin 1 and histatin 5 (bottom panel). The peptide fragments observed in the LC-MS/
MS experiment are aligned below the protein sequence. All peptide bond cleavage sites
are indicated in bold and underlined. Phosphorylated sites are indicated in italic bold
and underlined. < represents a pyroglutamic residue. Predominant cleavages near the C-
terminal from the primary sequence of acidic-PRP and histatin 1 were observed.

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
1114 R. Vitorino et al.ORIGINAL RESEARCH
DISCUSSION
In the present study, an experimental approach based
on the recovery of bovine teeth enamel implants after
2 h in the oral cavity of two human subjects was used
for in vivo pellicle collection. Bovine enamel presents
a high structural compatibility with human dental
hard tissues (Hannig et al., 2004) and was used recently
by Hannig et al. (2004, 2005), who have quantified
lyzosyme and amylase on in vivo enamel pellicle on
similar experimental conditions to those described in
this work. Moreover, to test possible contributions from
bovine enamel implants, blank LC-MS experiments
were performed using the same experimental proce-
dure, which showed the absence of contaminating
peptides (data not shown), confirming this strategy as a
valuable tool for in vivo experiments.
In order to dissociate pellicle protein and simultane-
ously characterize the bonding nature established between
salivary peptides and enamel surface, a sequential ex-
traction was performed based on guanidine cahotropic
properties (Del Vecchio et al., 2002) and on hydr-
oxyapatite solubilization by TFA, to remove the
remaining adsorbed proteins (Vitorino et al., 2005).
In most of the early studies, both in vivo and in vitro,
the characterization of acquired enamel pellicle was
performed based on bulk extractions procedures. As
shown by Moreno et al. (1984) and Gorbunoff and
Timasheff (1984), the nature of the interactions of
saliva components with enamel is crucial to the under-
standing of the mechanisms of its formation and its
function in tooth protection and bacterial adhesion.
The results found in this in vivo experiment clearly
show different compositions between the TFA and the
guanidine extracts and lead to the hypothesis that there
is a simple structural explanation for the different
strengths of interaction corresponding to the substitu-
tion of negative enamel ions and its complexation with
positive charges integrating the solid surface structure
(Vitorino et al., 2005).
In recent studies of whole saliva, Inzirati et al. (2005)
proteolysed with trypsin the isolated salivary peptides
obtained by HPLC pool collection. Several peptides
from basic and acidic PRP classes were then identified
by MS and MS/MS. This procedure allowed identifica-
tion of 18 different acidic PRPs isoforms (including
phosphorylated forms) and 15 different basic PRP
isoforms. All these peptides have molecular weights
over 3500 Da. Using HPLC separation and MS/MS,
Castagnola et al. (2004) analysed the low molecular
weight peptides (<3500 Da) whole saliva, and Hardt
et al. (2005) analysed parotid saliva samples. A cascade
of fragments resulting mainly from histatin 3, some
fragments resulting from PRPs (Hardt et al., 2005) and
the intact form of histatin 1 were observed (Csatagnola
et al., 2004). Only a few reports concerning pellicle
peptide analysis have been described. Lamkin et al.
(2001) reported the sequencing of several peptide
fragments in in vitro experiments belonging mainly
to histatins, statherin and cystatins classes. Yao et al.
(2001) reported the analysis of pellicle peptides by LC-
MS, finding components with a molecular weight in
the 1500–5400 Da range, attributed to statherin variant
V1 and of cystatin fragments. Following this, and taking
into consideration the importance of peptides as pellicle
precursors, we adopted a strategy based on the applica-
tion of LC-MS and LC-MS/MS for the analysis of pel-
licle extracts. This strategy allowed the separation and
identification of small peptides (by MS/MS pattern
search) as well as large peptides (through the com-
parison of the theoretical molecular weight with the
experimentally observed molecular weight). Our ex-
periments showed that guanidine extracts present
higher levels of fragmented peptides, which most
likely interact non-covalently with the enamel surface,
while mainly ‘intact’ peptides strongly interacting with
enamel surface are detected in TFA extracts.
With the exception of histatin 5, all the peptides
found in the TFA extract correspond to the phos-
phorylated species. These phosphorylated peptides,
statherin acidic PRPs, histatin 1, IB1 and II-2 have
been extensively studied with respect to their ability to
adsorb to enamel surface participating on its pellicle
formation (Lendenmann et al., 2000; Carlen et al., 1998;
Vacca Smith and Bowen, 2000; Jensen et al., 1991; Yin
et al., 2006; Al-Hashimi and Levine, 1989). The high
affinity of these species for enamel is most probably
due to the negative N-terminal. For example, in the se-
quence of acidic PRPs, the first 11 N-terminal residues
of the acidic PRPs contain eight negative charges
including a phosphorylated serine residue at position 8
and in the case of statherin the first five N-terminal
residues are three negative acidic amino acid residues
and two phosphorylated serines, Ser2 and Ser3. A simi-
lar trend with negative charges and phosphorylated
serines is observable for the other species. The negative
N-terminal is extremely important for interactions with
hydroxyapatite, as reported by Boackle et al. (1999),
who found that, after cleavage of the N-terminal of the
acidic PRPs by human leukocyte elastase, the affinity
decreased by more than 70%. Studies investigating the
conformation of statherin on the hydroxyapatite surface
showed a helical conformation after its adsorption to
the surface of hydroxyapatite. The statherin N-terminal
pentapetide recognizes the hydroxyapatite surface and
establishes a strong interaction between the surface and
the phosphoserines and carboxylate side-chains groups.
Dynamics studies suggest a weak interaction for the
remainder of the helix with hydroxyapatite (Schlesinger
and Hay, 1977; Stayton et al., 2003). Despite the
absence of reports concerning the other phosphorylated
peptides, a similar mechanism could be suggested for

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1115ORIGINAL RESEARCH
hydroxyapatite interaction by carboxylate groups and
phosphoserines with hydroxyapatite (Yin et al., 2006).
The histatin 5 sequence shows an aspartic residue on
the N-terminus (Asp1) (Fig. 5). The charge on the Asp
side chain is similar to that of phosphorylated residues
present on the N-terminal region and supports a similar
mechanism for the strong interaction of histatins with
the hydroxyapatite surface.
In this in vivo study we have found several fragments
arising from fragmentation of basic PRPs, acidic PRPs
and histatins non-covalently bound to the enamel sur-
face. In their study of histatin 3 fragmentation in saliva,
Castagnola et al. (2004) reported a cascade of frag-
ments from this peptide. In our work several histatin
fragments were observed; however, these belonged
only to histatin 5. Taking into account that the precur-
sor of histatin 5 is histatin 3, the absence of pellicle
fragments not shared between histatin 3 and 5 suggests
that the small fragments arise only from histatin 5 frag-
mentation. Underlining this suggestion, the observed
fragments cover the entire sequence of histatin 5. In the
case of basic PRPs, peptide fragments identified by
LC-MS/MS could be formed through fragmentation of
several protein precursors (at least four possibilities can
be assigned; Fig. 4) because this is a highly homologous
family (Table 2). These peptides, as observed for
histatin 5, arise from diverse cleavage points along their
primary sequence. For histatin 1, only C-terminal
fragmentations of the primary sequence were detected.
In this study, when compared with PRP3 relative
abundances, low relative abundances of PRP1 were
found strongly adsorbed (LC-MS of TFA extracts;
Fig. 1). This is in agreement with the work of Bennick
et al. (1983), who observed by crossed immuno-
electrophoresis the degradation of PRP1 to PRP3 and
smaller peptides after adsorption to enamel.
Comparing the identified peptide fragments with
those early observed on whole (Castagnola et al., 2004;
Messana et al., 2004; Inzitari et al., 2005) and parotid
saliva (Hardt et al., 2005), only six of the 30 fragments
identified have been previously observed. Our results
indicate that only a few classes of proteins and amino
acid residues are involved in proteolysis. Based on the
fragments identified, specific cleavage sites could be
assigned for the different classes of peptides. For
example, 80% (eight cleavage sites of the total of 10
observed for histatins) of the amide bond cleavages in
histatins occur at the Arg, His and Lys residues, while
for basic PRPs, the predominant cleavage sites occur
at the Lys, and Pro residues (four cleavage sites of
the total of six observed for this class). Some 85% of
the cleavage sites of acidic PRPs are located between
Gln and Gly residues. Altogether, this illustrates
the specificity of the proteolytic enzymes which pro-
mote the degradation of peptides on enamel pellicle.
Castagnola et al. (2004), in their characterization of
peptides in whole saliva, also observed a dominant
cleavage at Arg and Lys in histatin 3 and suggested that
a trypsin-like digestion pathway generated the histatin
3 cascade fragmentation. Proteolytic activity has been
reported in whole saliva in the last few years in associa-
tion with oral pathologies such as Sjögren’s syndrome,
and periodontitis (Dickinson, 2002; Pozo et al., 2005).
Among the enzymes implicated are metalloproteins
(MMP1 and 9; Chaussain-Miller et al., 2006), cathepsins
(B, H and L; Dickinson, 2002) and kallikrein. LC-MS/
MS of trypsin digests of whole saliva showed the pres-
ence of proteolytic enzymes including kallikrein (1 and
11), cathepsins (D and L) and carboxipeptidase H
(Wilmarth et al., 2004; Hu et al., 2005). The fragments
observed in our work could be attributed not only to
one enzyme, but to a combination of different enzymes.
The high content on lysine and arginine residues on
histatins makes these proteins extremely susceptible to
tryptic-like digestion. Most of the resulting fragments
found from the cleavage at the Arg and Lys residues
could be attributed to a trypsin-like pathway. Cleavage
of the Lys-His bond at Lys-17 of histatin 1 promotes
the release of the C-terminal peptide with a molecular
weight of 2618.11. After this, the cleavage at Arg–Xaa
between residues 22 and 23 of the primary sequence of
histatin 1 releases the peptide with a molecular weight
of 1963.8, or the cleavage between the residues Arg37
and Pro38 of the primary sequence of IB9 results in the
peptide with a molecular weight of 1746.87. Minor
cleavages at the Ser and Phe residues in the primary
sequence of histatins can also occur. Wong et al. (1983)
have reported that kallikreins are the responsible
enzymes associated with the generation of acidic PRP
C (now known as PRP3) before and after its secretion
from glandular tissues by a cleavage at the Arg106 resi-
due of acidic PRP A (now known as PRP1). Enzymes
which are able to cleave amide bond at His residues
(the other predominant cleavage on histatins) are also
required. It was previously reported by Xu et al. (1993)
that there existed a chymotryptic pathway which ad-
dressed the peptide-bond cleavage at the His residues.
Enzymes such as kallikreins or cathepsin members,
which work similarly to pepsin A via the chymotrypitic
pathway, may be involved in cleavages of histatin 3
leading to the observation of peptides with molecular
weights 1287.61, 1356.78 and 1766.93. The predominant
cleavages of acidic PRPs detected in LC-MS/MS ex-
periments occur between the Gln–Gly residues, which
is in agreement with the previous report by Jonsson
et al. (2001). This suggests that the presence of a cyclic
structure of the Pro residue which follows Gly could be
important for the stabilization of the reactive conforma-
tion. Furthermore, it should be emphasized that there
are no known enzymes able to cleave acidic PRPs at
Gln–Gly bonds, indicating the presence/activities of an
as yet unidentified enzyme.

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
1116 R. Vitorino et al.ORIGINAL RESEARCH
In conclusion, we have used a strategy based on
LC-MS/MS to characterize the low-molecular-weight
peptides of in vivo enamel pellicle. Based on these
data, a pattern of proteolytic pathways for salivary
peptides on enamel pellicle was observed with pre-
dominant cleavages at Arg, Lys, Ser and Phe residues
for the majority of the peptides (except acidic PRPs,
where the predominant cleavages are at the Gln–Gly
bond). These data suggest the involvement of a com-
bination of proteolytic enzymes which selectively
degrades the salivary proteins and contributes to modu-
lation of the acquired enamel pellicle.
Comparing all detected peptides in both extracts
(guanidine and TFA) with the peptides identified in
whole saliva described in previous works (some with
similar experimental conditions: Castagnola et al., 2004;
Messana et al., 2004; Vitorino et al., 2004a, 2004b;
Hardt et al., 2005), it is possible to observe a small
number of species adsorbed to the enamel. This sug-
gests the existence of a selective adsorption process
and the predominance of the adsorption of intact
forms. The presence of carboxyl groups or phosphate
groups in some adsorbed peptides should contribute to
a strong interaction with the enamel surface. This may
also be a protection factor against proteolytic activity
involving the formation of covalent/non-covalent com-
plexes with other proteins. Additionally, this study
shows that the bulk of peptide fragments were derived
from proteins other than the most abundant known
salivary components also present on enamel pellicle
(albumin, amylase, mucins) (Carlen et al., 1998;
Li et al., 2003, 2004; Hannig et al., 2005). These results
are in agreement with Hardt et al. (2005) for the
low-molecular-weight species isolated from parotid
saliva by passage through a 10 kDa membrane fol-
lowed by their characterization using tandem mass
spectrometry.
Although based on only two subjects, this study con-
tributes to a better understanding of the composition of
acquired enamel pellicle because previous studies
(Rykke and Sonju, 1991; Li et al., 2003; Yao et al., 2001;
Al-Hashimi and Levine, 1989) have shown that in vivo
pellicle composition exhibits considerable inter-subject
consistency, and that the overall protein composition
of the pellicles displayed characteristics typical of the
saliva prevailing in the area of the mouth where the
pellicles were formed (Vacca Smith and Bowen, 2000).
Based on the results reported here, we hypothesize that
the general mechanism for pellicle formation involves
a strong interaction of peptides containing carboxyl
and/or phosphate groups and enamel surface and elec-
trostatic interactions of other protein species forming
several layers, as suggested already by other authors
(Gorbunoff and Timasheff, 1984; Moreno et al., 1984;
Hay and Moreno, 1979). Further research concerning
inter-subject variability and the importance of tooth
location for pellicle composition could be the key to the
understanding of diseases involving tooth destruction.
Acknowledgements
This research was supported in part by the Intramural
Research Program of the NIH, National Institute of
Environmental Health Sciences. The authors would like
to express their appreciation for the financial support
provided by the Fundação para a Ciência e Tecnologia
(FCT, grant nos SFRH/BPD/14968/2004 and POCTI/
QUI/5890/2004).
REFERENCES
Al-Hashimi I and Levine MJ. Characterization of in vivo salivary-
derived enamel pellicle. Archives of Oral Biology 1989; 34: 289–
295.
Bennick A, Chau G, Goodlin R, Abrams S, Tustian D and
Madapallimattam G. The role of human salivary acidic proline-rich
proteins in the formation of acquired dental pellicle in vivo and
their fate after adsorption to the human enamel surface. Archives
of Oral Biology 1983; 28: 19–27.
Boackle RJ, Dutton SL, Robinson WL, Vesely J, Lever JK, Su HR
and Chang NS. Effects of removing the negatively charged N-
terminal region of the salivary acidic proline-rich proteins by
human leucocyte elastase. Archives of Oral Biology 1999; 44: 575–
585.
Carlen A, Borjesson AC, Nikdel K and Olsson J, Composition of
pellicles formed in vivo on tooth surfaces in different parts of the
dentition, and in vitro on hydroxyapatite. Caries Research 1998; 32:
447–455.
Castagnola M, Cabras T, Inzitari R, Zuppi C, Rossetti DV,
Petruzzelli R, Vitali A, Loy F, Conti G and Fadda MB. Determina-
tion of the post-translational modifications of salivary acidic proline-
rich proteins. European Journal Morphology 2003; 41: 93–98.
Castagnola M, Inzitari R, Rossetti DV, Olmi C, Cabras T, Piras V,
Nicolussi P, Sanna MT, Pellegrini M, Giardina B and Messana I.
A cascade of 24 histatins (histatin 3 fragments) in human saliva.
Suggestions for a pre-secretory sequential cleavage pathway. Jour-
nal of Biological Chemistry 2004; 279: 41436–41443.
Chaussain-Miller C, Fioretti F, Goldberg M and Menashi S. The role
of matrix metalloproteinases (MMPs) in human caries. Journal of
Dental Research 2006; 85: 22–32.
Del Vecchio P, Graziano G, Granata V, Barone G, Mandrich L,
Rossi M and Manco G. Denaturing action of urea and guanidine
hydrochloride towards two thermophilic esterases. Biochemistry
Journal 2002; 367: 857–863.
Dickinson DP. Cysteine peptidases of mammals: their biological roles
and potential effects in the oral cavity and other tissues in health
and disease. Critical Reviews in Oral Biology and Medicine 2002;
13: 238–275.
Gorbunoff MJ and Timasheff SN. The interaction of proteins with
hydroxyapatite. III. Mechanism. Analytical Biochemistry 1984; 136:
440–5.
Hannig C, Attin T, Hannig M, Henze E, Brinkmann K and Zech R.
Immobilisation and activity of human alpha-amylase in the
acquired enamel pellicle. Archives of Oral Biology 2004; 49: 469–
475.
Hannig C, Hoch J, Becker K, Hannig M and Attin T. Lysozyme
activity in the initially formed in situ pellicle. Archives of Oral
Biology 2005; 50: 821–828.
Hardt M, Thomas LR, Dixon SE, Newport G, Agabian N,
Prakobphol A, Hall SC, Witkowska HE and Fisher SJ. Toward
defining the human parotid gland salivary proteome and
peptidome: identification and characterization using 2D SDS-
PAGE, ultrafiltration, HPLC, and mass spectrometry. Biochemistry
2005; 44: 2885–2899.

Copyright © 2007 John Wiley & Sons, Ltd. Biomed. Chromatogr. 21: 1107–1117 (2007)
DOI: 10.1002/bmc
Peptidomic analysis of human acquired enamel pellicle 1117ORIGINAL RESEARCH
Hay DI and Moreno EC. Differential adsorption and chemical affini-
ties of proteins for apatitic surfaces. Journal of Dental Research
1979; 58: 930–942.
Hu S, Xie Y, Ramachandran P, Ogorzalek Loo RR, Li Y, Loo JA
and Wong DT. Large-scale identification of proteins in human
salivary proteome by liquid chromatography/mass spectrometry
and two-dimensional gel electrophoresis–mass spectrometry.
Proteomics 2005; 5: 1714–1728.
Inzitari R, Cabras T, Onnis G, Olmi C, Mastinu A, Sanna MT,
Pellegrini MG, Castagnola M and Messana I. Different isoforms
and post-translational modifications of human salivary acidic
proline-rich proteins. Proteomic. 2005; 5: 805–815.
Jensen JL, Lamkin MS, Troxler RF and Oppenheim FG. Multiple
forms of statherin in human salivary secretions. Archives of Oral
Biology 1991; 36: 529–534.
Jonsson AP, Bergman T, Jornvall H and Griffiths WJ. Gln-Gly
cleavage: a dominant dissociation site in the fragmentation of
protonated peptides. Journal of American Society of Mass
Spectrometry 2001; 12: 337–342.
Lamkin MS, Arancillo AA and Oppenheim FG. Temporal and
compositional characteristics of salivary protein adsorption to
hydroxyapatite. Journal of Dental Research 1996; 75: 803–808.
Lamkin MS, Migliari D, Yao Y, Troxler RF and Oppenheim FG.
New in vitro model for the acquired enamel pellicle: pellicles
formed from whole saliva show inter-subject consistency in protein
composition and proteolytic fragmentation patterns. Journal of
Dental Research 2001; 80: 385–388.
Leinonen J, Kivela J, Parkkila S, Parkkila AK and Rajaniemi H.
Salivary carbonic anhydrase isoenzyme VI is located in the human
enamel pellicle. Caries Research 1999; 33: 185–190.
Lendenmann U, Grogan J and Oppenheim FG. Saliva and dental
pellicle. Advances in Dental Research 2000; 14: 22–28.
Li J, Helmerhorst EJ, Corley RB, Luus LE, Troxler RF and
Oppenheim FG. Characterization of the immunologic responses to
human in vivo acquired enamel pellicle as a novel means to investig-
ate its composition. Oral Microbiology and Immunology 2003; 18:
183–191.
Li J, Helmerhorst EJ, Troxler RF and Oppenheim FG. Identification
of in vivo pellicle constituents by analysis of serum immune
responses. Journal of Dental Research 2004; 83: 60–64.
Messana I, Loffredo F, Inzitari R, Cabras T, Giardina B, Onnis G,
Piludu M and Castagnola M. The coupling of RP-HPLC and ESI-
MS in the study of small peptides and proteins secreted in vitro by
human salivary glands that are soluble in acidic solution. European
Journal of Morphology 2003; 41: 103–106.
Messana I, Cabras T, Inzitari R, Alessandro L, Zuppi C, Olmi C,
Fadda B, Cordaro M, Giardina B and Castagnola M. Characteriza-
tion of the human salivary basic proline-rich protein complex by a
proteomic approach. Journal of Proteome Research 2004; 3: 792–
800.
Moreno EC, Kresak M and Hay DI. Adsorption of molecules of bio-
logical interest onto hydroxyapatite. Calcified Tissue International
1984; 36: 48–59.
Pozo P, Valenzuela MA, Melej C, Zaldivar M, Puente J, Martinez B
and Gamonal J. Longitudinal analysis of metalloproteinases, tissue
inhibitors of metalloproteinases and clinical parameters in gingival
crevicular fluid from periodontitis-affected patients. Journal of
Periodontal Research 2005; 40: 199–207.
Rykke M and Sonju T. Amino acid composition of acquired enamel
pellicle collected in vivo after 2 hours and after 24 hours.
Scandinavian Jounal of Dental Research 1991; 99: 463–469.
Schlesinger DH and Hay DI. Complete covalent structure of
statherin, a tyrosine-rich acidic peptide which inhibits calcium phos-
phate precipitation from human parotid saliva. Journal of Biologi-
cal Chemistry 1977; 252: 1689–1695.
Schupbach P, Oppenheim FG, Lendenmann U, Lamkin MS,
Yao Y and Guggenheim B. Electron-microscopic demonstration of
proline-rich proteins, statherin, and histatins in acquired enamel
pellicles in vitro. European Journal of Oral Sciences 2001; 109: 60–
68.
Stayton PS, Drobny P, Shaw WJ, Long JR and Gilbert M. Molecular
recognition at the protein-hydroxyapatite interface. Critical Reviews
in Oral Biology and Medicine 2003; 14: 370–376.
Vacca Smith AM and Bowen WH. In situ studies of pellicle forma-
tion on hydroxyapatite discs. Archives of Oral Biology 2000; 45:
277–291.
Vitorino R, Lobo MJ, Duarte J, Ferrer-Correia AJ, Tomer KB,
Dubin JR, Domingues PM and Amado FM. In vitro hydroxya-
patite adsorbed salivary proteins. Biochemistry Biophysics and
Research Communications 2004a; 320:! 342–346.
Vitorino R, Lobo MJ, Duarte JA, Ferrer-Correia AJ, Domingues PM
and Amado FM. Analysis of salivary peptides using HPLC–
electrospray mass spectrometry. Biomedical Chromatography
2004b; 18: 570–575.
Vitorino R, Lobo MJ, Duarte J, Ferrer-Correia AJ, Tomer KB,
Dubin JR, Domingues PM and Amado FM. The role of salivary
peptides in dental caries. Biomedical Chromatography 2005; 19:
214–222.
Wilmarth PA, Riviere MA, Rustvold DL, Lauten JD, Madden TE
and David LL. Two-dimensional liquid chromatography study of
the human whole saliva proteome. Journal of Proteome Research
2004; 3: 1017–1023.
Wong RS, Madapallimattam G and Bennick A. The role of glandular
kallikrein in the formation of a salivary proline-rich protein A by
cleavage of a single bond in salivary protein C. Biochemistry Jour-
nal 1983; 211: 35–44.
Xu L, Lal K, Santarpia RP III and Pollock JJ. Salivary proteolysis of
histidine-rich polypeptides and the antifungal activity of peptide
degradation products. Archives of Oral Biology 1993; 38: 277–283.
Yao Y, Lamkin MS and Oppenheim FG. Pellicle precursor proteins:
acidic proline-rich proteins, statherin, and histatins, and their
crosslinking reaction by oral transglutaminase. Journal of Dental
Research 1999; 78: 1696–1703.
Yao Y, Lamkin MS and Oppenheim FG. Pellicle precursor protein
crosslinking characterization of an adduct between acidic proline-
rich protein (PRP-1) and statherin generated by transglutaminase.
Journal of Dental Research 2000; 79: 930–938.
Yao Y, Grogan J, Zehnder M, Lendenmann U, Nam B, Wu Z,
Costello CE and Oppenheim FG. Compositional analysis of human
acquired enamel pellicle by mass spectrometry. Archives of Oral
Biology 2001; 46: 293–303.
Yao Y, Berg EA, Costello CE, Troxler RF and Oppenheim FG.
Identification of protein components in human acquired enamel
pellicle and whole saliva using novel proteomics approaches. Jour-
nal of Biological Chemistry 2003; 278: 5300–5308.
Yin A, Margolis HC, Grogan J, Yao Y, Troxler RF and Oppenheim
FG. Physical parameters of hydroxyapatite adsorption and effect
on candidacidal activity of histatins. Archives of Oral Biology 2003;
48: 361–368.
Yin A, Margolis HC, Yao Y, Grogan J and Oppenheim FG.
Multi-component adsorption model for pellicle formation: the
influence of salivary proteins and non-salivary phospho proteins on
the binding of histatin 5 onto hydroxyapatite. Archives of Oral
Biology 2006; 51: 102–110.