pentadinium alabamensis: a new, unusual dinoflagellate from the early oligocene of the gulf coast,...
TRANSCRIPT

Review of Palaeobotany and Palynology 175 (2012) 47–54
Contents lists available at SciVerse ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r .com/ locate / revpa lbo
Research paper
Pentadinium alabamensis: A new, unusual dinoflagellate from the early Oligocene ofthe Gulf Coast, Alabama, USA
Willemijn Quaijtaal a,b,⁎, Henk Brinkhuis a
a Marine Palynology, Laboratory of Palaeobotany and Palynology, Department of Earth Sciences, Faculty Geosciences, Utrecht University, Budapestlaan 4, 3584 CD Utrecht, The Netherlandsb Research Unit Palaeontology, Department Geology and Soil Science, Ghent University, Krijgslaan 281 S8/WE13, B-9000 Ghent, Belgium
⁎ Corresponding author at: Research Unit PalaeontologScience, Ghent University, Krijgslaan 281 S8/WE13, B-90264 46 10.
E-mail address: [email protected] (W. Qu
0034-6667/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.revpalbo.2012.03.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 August 2011Received in revised form 9 March 2012Accepted 12 March 2012Available online 20 March 2012
Keywords:organic walled dinoflagellate cystsEocene/Oligocene transitionSt. Stephens QuarryAlabamaUSA
The Eocene–Oligocene Transition (EOT, ~34 Ma)marks the onset of major Antarctic ice sheets. The environmentalconsequences of the transition included major changes in e.g., sea level, temperature, and ocean circulation, com-plicating biostratigraphic correlations in this interval. Organic walled dinoflagellate cysts (dinocysts) however doshow potential for EOT biostratigraphy, especially for ancient shallow marine settings.At St. Stephens Quarry, Alabama, USA, we found a new, extremely suturocavate dinocyst, Pentadinium alabamensissp. nov., described herein. The range of the new species spans the critical EOT magnetosubchron C13n, makingthis taxon a useful biostratigraphic marker for this interval in the Gulf Coast region. The species appears to be as-sociated with shallow marine, euryhaline conditions.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The Eocene–Oligocene transition (EOT, ~34 Ma) reflects the tran-sition from the early Paleogene Greenhouse into the Icehouse world,marking the onset of major Antarctic glaciation (e.g., Zachos et al.,1996, 2001; Coxall et al., 2005; Zachos et al., 2008). The EOT paleocli-matic and correlated paleoceanographic changes often complicatestraightforward biostratigraphic interpretations in this interval.Steepened latitudinal temperature gradients, surface water reorgani-zations, corrosive deep ocean currents and sea level changes causeddiachronous range tops and first appearances, besides issues withpreservation and reworking (see Coxall and Pearson, 2007 for anoverview). Biostratigraphy based on the organic walled remains ofdinoflagellates (dinocysts) has shown potential for the EOT intervalin e.g., the Mediterranean, North Atlantic, and Tasman Sea regions(e.g., Brinkhuis and Biffi, 1993; Brinkhuis, 1994; Bujak and Mudge,1994; Sluijs et al., 2003; Eldrett et al., 2004).
Because many organic-cyst-forming dinoflagellates are ecologicallyadapted to relatively marginal, shallow marine settings, this group isparticularly useful when correlating ancient in- to offshore settings(see e.g., Pross and Brinkhuis, 2005). While analyzing the relativelyshallowmarine deposits of the Gulf Coast region at St. Stephens Quarry(SSQ), Alabama, USA (see Fig. 1; see alsoWade et al., 2012)we recordedan unusual dinocyst within lower Oligocene sediments calibrated
y, Department Geology and Soil00 Ghent, Belgium. Tel.: +32 9
aijtaal).
rights reserved.
againstmagnetosubchron C13n (Wade et al., 2012). It appearsmorpho-logically related to representatives of the genus Pentadinium but differsby displaying extraordinarilywide separation of the outerwall from theinner wall. This taxon was reported earlier from multiple localitieswithin the lower Oligocene of the Gulf Coast region (Fig. 1) byJaramillo and Oboh-Ikuenobe (1999) as ‘Pentadinium sp. A’. Our studynow confirms its consistent presence in the lower Oligocene in thisregion and the morphological stability of the species. The latter aspectwarrants a separate taxonomic position. Here, we thus describe anddocument this biostratigraphically useful, new species, and proposeplacing it within the genus Pentadinium.
2. Material and methods
2.1. Material
The St. Stephens Quarry borehole (SSQ, St. Stephens, WashingtonCounty, Alabama, USA; 31°33′ N lat., 88°02′ W long., see Fig. 1) wascontinuously cored by ARCO Oil and Gas Company in 1987 (Milleret al., 1993; Wade et al., 2012). The cored interval mainly consists ofsilts, clays and sands with varying carbonate content. This successionis interrupted by thin siliciclastic and glauconitic beds (Miller et al.,2008). Details of the lithology can be found in Miller et al. (2008).The studied interval of the SSQ bore hole contains the followingtypical Gulf Coast lithostratigraphic units: the Jackson Group andthe Vicksburg Group. These units can be subdivided into severalformations and their respective members (Fig. 2). The JacksonGroup is composed of the Moodys Branch Formation and theYazoo Clay. The Yazoo Clay can furthermore be subdivided in the

0 20 km
#1 Wayne
#1 Young
St. Stephens Quarry
Washington County
Wayne County
89°W
89°W90°W
90°W
31°N 31°N
35°N 35°N
0 50 km
ALABAMA
MISSISSIPPI
tt
Atlantic Ocean
Gulf of Mexico
Fig. 1. Locations of the localities where Pentadinium alabamensis has been recorded. t0:paleoshoreline during accumulation of Shubuta Clay and equivalents, t1: paleoshorelineduring accumulation of Red Bluff Clay and equivalents (adapted from Jaramillo andOboh-Ikuenobe, 1999, paleoshorelines after Tew and Mancini, 1995).
48 W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
following members: 1) the North Twistwood Creek Member, 2) theCocoa Sand Member, 3) the Pachuta Marl Member and 4) the Shu-buta Member. The Vicksburg Group at SSQ contains the BumpnoseLimestone, the Red Bluff Clay, the Marianna Limestone and theByram Formation. The Glendon Limestone Member is the lowestmember of the Byram Formation.
Magnetic inclination
-90 0 90
15
20
25
30
35
40
45
50
55
60
Dep
th (
m)
HO Pseudoh
HO R.
HO H
Base C12n
31 32
Mag
neto
subc
hron
s
C12r
C13n
C13r
C15r
C16r
C12n
Sam
ples
NT
CP
MC
SM
MS
MG
L
Jack
son
Gro
upV
icks
burg
Gro
up
Yazo
o C
lay
Mar
iann
a Li
mes
tone
BLS
RB
BF
MLi
thos
trat
ogra
phic
units Oligocene
Fig. 2. Magnetic inclination, magnetostratigraphy, lithostratigraphic units, samples analyzesamples with erratic paleomagnetic behavior. Age model is based onmagnetostratigraphy (blanannoplankton, stars: planktonic foraminifera) (Miller et al., 2008). Tie points used for the agelines, the black dashed line in Yazoo Clay indicates a parasequence boundary. Highest and lowOi-1 event and corresponding hiatus are located at the base of C13n. HO: highest occurrence, LOClay, BFM: Byram Formation, NTC: North Twistwood Creek Member, C: Cocoa Sand MembeGL: Glendon Limestone Member.
A total of 59 samples have generally been taken every ~1.5 m,and ~0.3–0.6 m for the EOT interval (see Fig. 2).
For the age model we follow Wade et al. (2012), which providesan update from the earlier data presented by e.g., Miller et al.(2008) (see Fig. 2). This update mainly regards the identification ofa ~200 kyr hiatus – associated with the Oligocene Isotope-1 Event(Oi-1) – near the base of magnetosubchron C13n, whereas Milleret al. (2008) thought SSQ to be complete at this point. The agemodel is based on magnetostratigraphy, δ18O correlations andbiostratigraphy (see Fig. 2).
2.2. Methods
Standard palynological techniques have been used to process thesamples. Briefly: samples were cleaned and crushed before oven dry-ing at 60 °C. Dried samples were then weighed. Material was firstrehydrated with the wetting agent Agepon® (1:200). Then, to re-move carbonates, hydrochloric acid (HCl, 10%) was added. Next, todissolve silicates, 38% hydrofluoric acid (HF) has been used, followedby shaking at ~250 rpm for 2 h and addition of a surplus of 30% HCl toremove fluoride gels. Samples were washed twice by decanting aftera 24 h settling and filling up with water after each acid step. Sampleswere first sieved with a 250-μm nylon mesh sieve; the filtrate wasagain sieved with a 15-μm nylon mesh sieve. The sample was shortlyplaced in an ultrasonic bath to break up clumped residue. The sievedresidue was transferred into a glass test tube. Test tubes were centri-fuged at 2000 rpm for 5 min without brake. Water surplus was re-moved and the residue was transferred into a vial with addition ofglycerin water. After homogenization, one drop of the residue wasmounted on a microscopic slide together with some glycerin jellyand stirred. Slides were covered with a cover slip and sealed withnail polish. Per sample two slides have been prepared. A minimumnumber of 200 dinocysts was counted; afterwards the uncounted
Depth (m
)
60
15
20
25
30
35
40
45
50
55
astigerina spp.
Base C16n.2nTop C16n.1n
HO E. formosa
umbilicus
HO R. reticulata
FAD I. recurvusHO D. saipanensisantkeninidae
HO T. cerroazulensis group
HO P. micra
Base C13n
Top C13n
33 34 35 36 37
Age (Ma, Berggren et al., 1995)
LO P. alabamensis
HO P. alabamensis
Eocene
d for palynology and age-depth plot for the St. Stephens Quarry core. Plusses indicateck squares), δ13C correlations (gray circles) and biostratigraphy (white circles: calcareousmodel are connected by a solid black line. Hiatuses are indicated by horizontal red dashedest occurrences of Pentadinium alabamensis have been indicated by black dotted lines. The: lowest occurrence, FAD:first appearance datum, BLS: Bumpnose Limestone, RB: Red Bluffr, PM: Pachuta Marl Member, SM: Shubuta Member, MSM: Mint Spring Marl Member,
49W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54

50 W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
part of the slide was scanned for important biostratigraphic markers.Taxonomy, unless indicated otherwise, follows that cited in Fensomeet al. (2008).
Light microscopic images have been made with a Leica DFC320camera on a Leica DM LB2 microscope using Adobe Photoshop, addi-tional scanning electron microscope (SEM) photography has beenperformed as well. For this purpose, splits of the palynological resi-dues were sieved with warm water over a 15 μm nylon mesh sieveto remove glycerin and then transferred to a tray table. Each traytable was coated with 12 nm platinum using a sputter coater (Cres-sington 208 h). Samples have been examined using a Philips XL30SFEG device, located at Utrecht University. Material is stored in the col-lection of the Laboratory of Palaeobotany and Palynology, UtrechtUniversity, The Netherlands.
The terminology for describing fossil dinocysts we use follows Evittet al. (1977) and Evitt (1985). However, we refrain from using “para-”as a prefix of morphological features referring to the cyst (followingFensome et al., 1993), to avoid confusion with terminology solelyused for exceptional situations, e.g. “parasutural ridges”. Furthermore,we do not use the terms “autophragm” and “ectophragm” since in Pen-tadinium there is no true, single layered autophragm, but an endo-phragm and a periphragm that are appressed in the central plate areaand separated at the plate boundaries.
3. Systematic paleontology
Division DINOFLAGELLATA (Bütschli, 1885) Fensome et al., 1993Subdivision DINOKARYOTA Fensome et al., 1993Class DINOPHYCEAE Pascher, 1914Subclass PERIDINIPHYCIDAE Fensome et al., 1993Order GONYAULACALES Taylor, 1980Suborder GONYAULACINAE AutonymFamily GONYAULACACEAE Lindemann, 1928Subfamily GONYAULACOIDEAE AutonymGenus Pentadinium Gerlach, 1961, emend. Benedek et al., 1982
Pentadinium alabamensis, sp. nov., Plate 1, A–D1999 Pentadinium sp. A, Jaramillo and Oboh-Ikuenobe, plate 2, fig. 9
Etymology: Named after the state of Alabama (USA), home of thetype species.Holotype: Sample SSQ 17 (42.56 m core depth), slide 1, EnglandFinder (EF) coordinates N33/2, Plate 1, A–DParatype 1: Sample SSQ 17(42.56 m core depth), slide 2, EF G31/1,Plate 1, E–GParatype 2: Sample SSQ 17(42.56 m core depth), slide 2, EF H30/4,Plate 1, H–JType locality: St. Stephens Quarry, Alabama, USAType stratum: Red Bluff Clay
Repository: Laboratory of Palaeobotany and Palynology, UtrechtUniversity, Budapestlaan 4, 3584CD Utrecht, The Netherlands.
Diagnosis: A species of Pentadinium displaying a gonyaulacoidtabulation (3–4′, 6″, 6c, 6‴, 1p, 1⁗; Evitt, 1985) with a 1P (3″) arche-opyle, operculum free. The species shows several characteristics typ-ical for Pentadinium: two walls that appear as a single layer at the
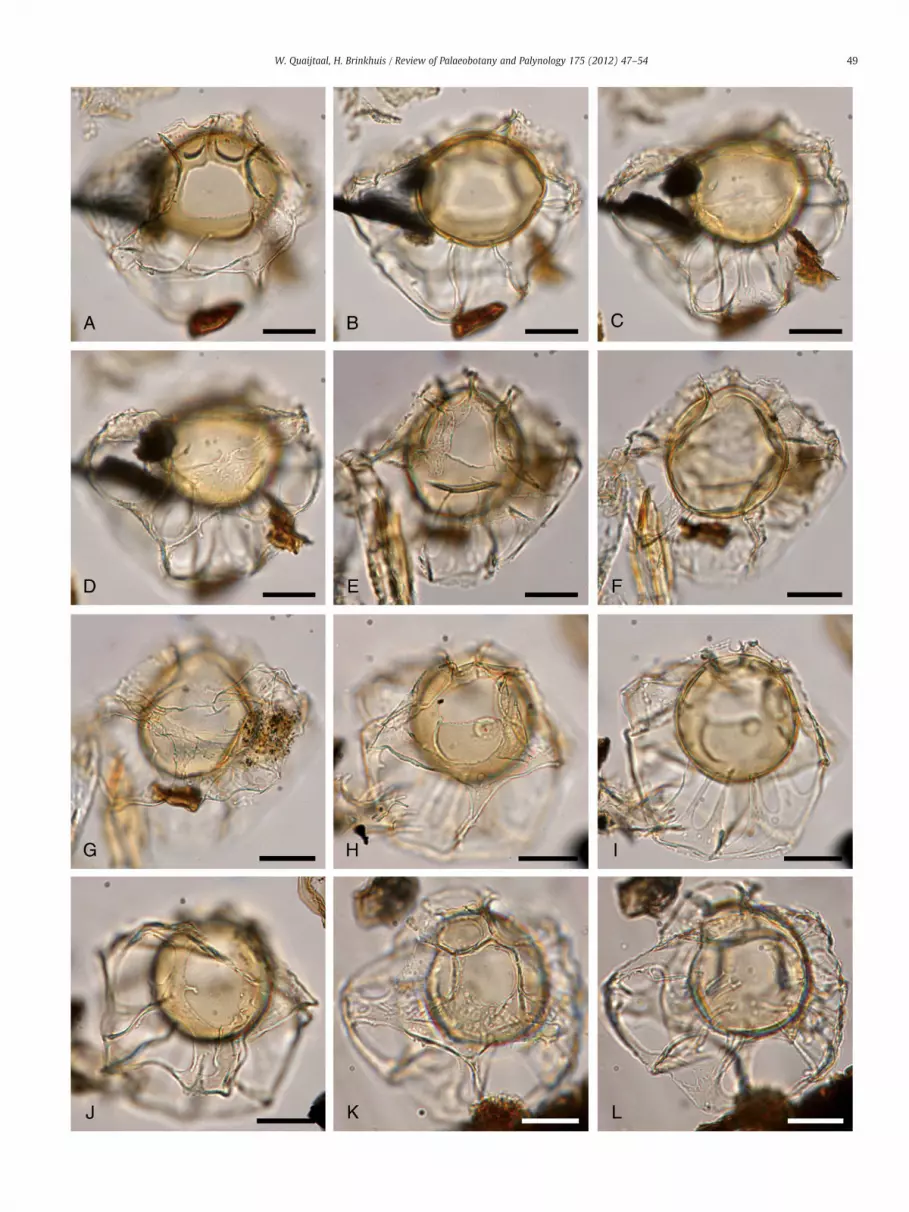
Plate 1. Light microscopy pictures of Pentadinium alabamensis sp. nov. The scale bar is 20 μ
A–D: Holotype (sample “SSQ 17, slide 1”, EF Reference: N33/2). A: dorsal view, hiD: same specimen with lowest focus on the ventral side.
E–G: Paratype 1 (sample “SSQ 17, slide 2”, EF Reference: G31/1). E: dorsal view,
H–J: Paratype 2 (sample “SSQ 17, slide 2”, EF Reference: H30/4). H: dorsal view,
K, L: “sample SSQ 17, slide 1”, EF Reference: K36/3, K: lowest focus on right la
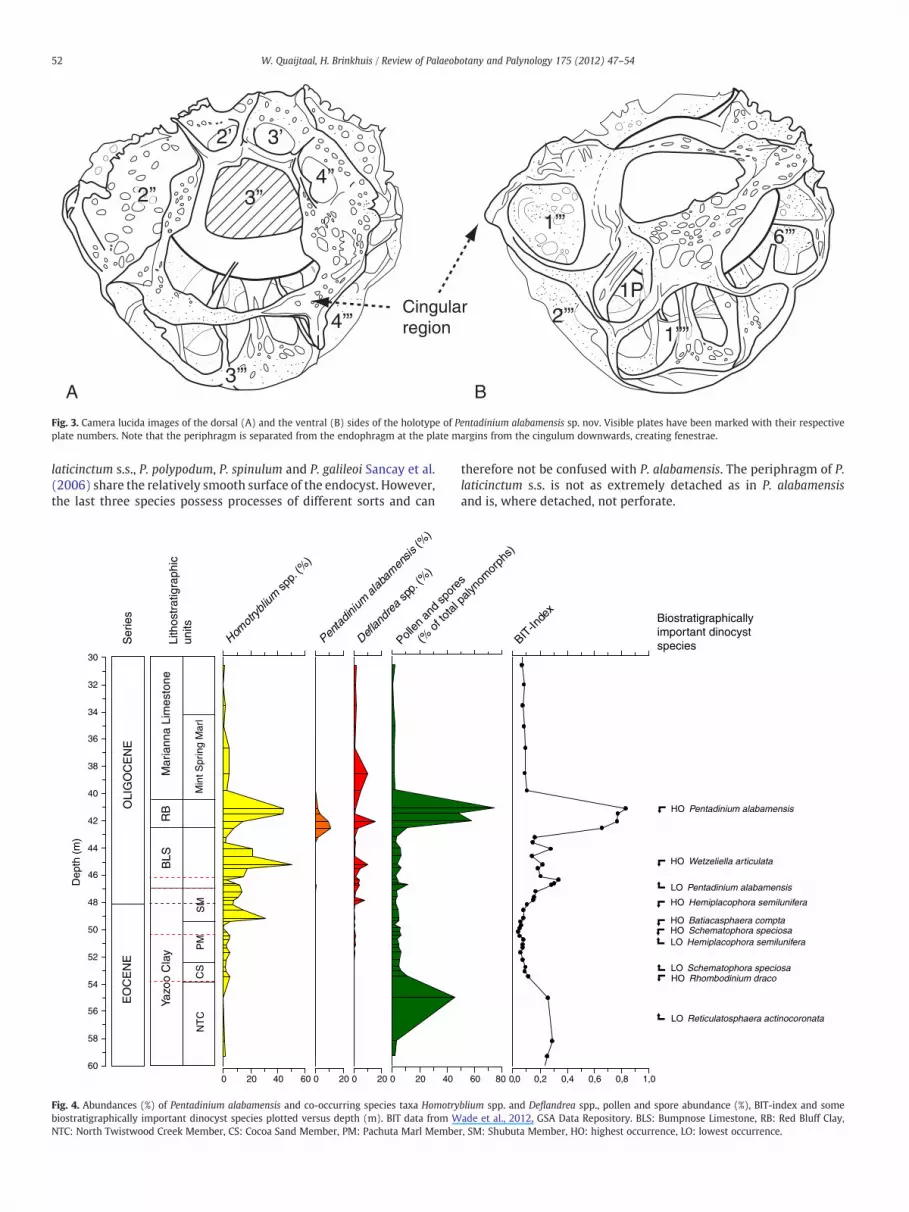
plate interior and show separation at the plate margins. The endo-phragm is thicker than the periphragm. Furthermore, it shows cava-tion around the cingulum. Plates 1′ and 4′ are more or less fusedand plate 6″ is poorly delimited (see Plate 2A,E).
The endophragm is subsphaerical, scabrate or microgranulate. Theperiphragm is thin, perforate and widely extends outwards. The tab-ulation is expressed by these extreme outgrowths of the periphragm,considered to principally reflect extreme suturocavation. Outgrowthsform septa at the apical plates and a fenestrate membrane on the restof the cyst. The sulcal plates are not reflected.
Description: A medium sized (60–70 μm), subsphaerical to ellipti-cal species of Pentadinium, showing extreme suturocavation. The cystpossesses two wall layers. The endophragm is thick and scabrate tomicrogranulate (see Plate 2D). It does not show any tabulation,except for the precingular, 1P (3″) archeopyle, operculum free. Theperiphragm is thin, perforate and closely appressed to the endo-phragm at the central areas of the plates. At these appressed areasthe periphragm is not perforate. At the plate boundaries the peri-phragm is detached and extends outwards exceptionally (suturoca-vation). This aspect in Pentadinium alabamensis sp. nov. is soextreme that it appears to bear large distal crest-like structures, afenestrate periphragm supported by slender processes (see Fig. 3;Plate 2B, F). We consider these to be exceptional outfolds of thesuturocavate nature of the periphragm. Pentadinium alabamensis sp.nov. displays a gonyaulacoid tabulation: 3–4′, 6″, 6c, 6‴, 1p, 1⁗with a 1P(3″) archeopyle, operculum free (see Fig. 3; Plate 2A, E).The sulcal region is not reflected by the periphragm (see Fig. 3B).The crest-like structures on the cyst are mostly found to representthe apical plates series (plates 1–4′, see Plate 2A), while from theprecingular plates downward the periphragm is almost completelydetached, reflecting the precingular, cingular, postcingular andantapical plates. The cingular plates may be difficult to see underthe light microscope (see Fig. 3).
Dimensions: Average diameter of the endocyst was 55 μm, averagediameter of the pericyst 91 μm (n=15). Holotype: diameter ofendocyst 53 μm, diameter of pericyst 95 μm; paratype 1: endocyst54 μm, pericyst 95 μm; paratype 2: endocyst 50 μm, pericyst87 μm.
Comparison: Pentadinium alabamensis sp. nov. can be distinguishedfrom all species of Pentadinium in showing extreme separation ofthe perforate periphragm from the endophragm. Although Pentadi-nium favatum also might show extreme separation of these layers,this is not a consistent feature as in P. alabamensis. More importantly,P. favatum has its characteristic honeycomb texture. Pentadinium?circumsutum, P. corium Schiøler (2005), P. goniferum, P. granulatum(Gocht, 1969) Fensome et al. (2009), P. imaginatum, P. membranaceum,P. sabulum Fensome et al. (2009) and P. taenagrium can all be distin-guished from the new species P. alabamensis in having a granular to ver-miculate endocyst surface structure, whereas P. lophophorum can beseparated based on its typical surface ornamentation. P. alabamensiscan be distinguished from P. netangei based on its size and having a per-forate rather than a microgranulate periphragm. The camocavate P.omasum has, unlike P. alabamensis, a pseudopunctate endocyst struc-ture and cavation is restricted to the dorsal part of the cyst. P.
m.
gh focus. B: dorsal view, middle focus. C: same specimen with lower focus on ventral side.
high focus. F: dorsal view, lower focus. G: same specimen, lowest focus on ventral side.
high focus. I: dorsal view, lower focus. J: same specimen, lowest focus on ventral view.
teral side. L: low focus on left lateral side.

Plate 2. Scanning electron microscope (SEM) pictures of Pentadinium alabamensis sp. nov.
A: Apical view (sample “SSQ 17”). Note the reduction of apical plates 1′ and 4′.
B: Right lateral view (sample “SSQ 17”). This sample shows the restriction of the septa-like outfolds of the periphragm to the apical plates.
C: Oblique antapical view (sample “SSQ 17”).
D: Detail (sample “SSQ 17”) of the endophragm structure. Note the splitting of the base of the ‘process’, showing the suturocavate origin.
E: Apical view (sample “SSQ 17”). Note the merged apical plates 1′ and 4′.
F: Detail of Plate 2E (sample “SSQ 17”) showing the perforate, suturocavate nature of the periphragm.
51W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
3’’4’’
3’2’
Cingularregion
3’’’
4’’’
2’’1’’’
6’’’
1P2’’’
1’’’’
A B
Fig. 3. Camera lucida images of the dorsal (A) and the ventral (B) sides of the holotype of Pentadinium alabamensis sp. nov. Visible plates have been marked with their respectiveplate numbers. Note that the periphragm is separated from the endophragm at the plate margins from the cingulum downwards, creating fenestrae.
52 W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
laticinctum s.s., P. polypodum, P. spinulum and P. galileoi Sancay et al.(2006) share the relatively smooth surface of the endocyst. However,the last three species possess processes of different sorts and can
30
32
34
36
38
40
42
44
46
48
50
52
54
56
58
60
Dep
th (
m)
0 20 40 60
Homotr
ybliu
msp
p. (%
)
0 20
Penta
dinium
alaba
men
sis (%
)
0 20
Deflan
drea
spp.
(%)
0 20 40
Pollen
and sp
or
(
% o
f tot
a
Mar
iann
a Li
mes
tone
Min
t Spr
ing
Mar
l
Yazo
o C
lay
NT
CC
SP
MS
M
BLS
RB
cihpargitartsohtiLstinu
EO
CE
NE
OLI
GO
CE
NE
seireS
Fig. 4. Abundances (%) of Pentadinium alabamensis and co-occurring species taxa Homotrybiostratigraphically important dinocyst species plotted versus depth (m). BIT data from WNTC: North Twistwood Creek Member, CS: Cocoa Sand Member, PM: Pachuta Marl Membe
therefore not be confused with P. alabamensis. The periphragm of P.laticinctum s.s. is not as extremely detached as in P. alabamensisand is, where detached, not perforate.
60 80
es
l paly
nom
orph
s)
0,0 0,2 0,4 0,6 0,8 1,0
BIT-In
dex
HO Wetzeliella articulata
HO Hemiplacophora semilunifera
HO Batiacasphaera comptaHO Schematophora speciosaLO Hemiplacophora semilunifera
LO Schematophora speciosaHO Rhombodinium draco
LO Reticulatosphaera actinocoronata
HO Pentadinium alabamensis
LO Pentadinium alabamensis
Biostratigraphically important dinocyst species
blium spp. and Deflandrea spp., pollen and spore abundance (%), BIT-index and someade et al., 2012, GSA Data Repository. BLS: Bumpnose Limestone, RB: Red Bluff Clay,r, SM: Shubuta Member, HO: highest occurrence, LO: lowest occurrence.

P. fa
vatu
m
P. g
onife
rum
P. p
olyp
odum
P. m
embr
anac
eum
P. la
ticin
ctum
P. g
ranu
latu
m
TALLAHATTA
LISBON
GOSPORT&
EQUIVALENTS
MOODYSBRANCH
YAZOO
MID
DLE
EO
CE
NE
UP
PE
RE
OC
EN
E
P. a
laba
men
sis
LOW
ER
OLI
GO
CE
NE
YAZOO
BUMPNOSE
RED BLUFF
MINT SPRING
Fig. 5. Ranges of Pentadinium species in western and central Alabama, including the newspecies Pentadinium alabamensis. Dashed lines on P. laticinctum laticinctum and P. laticinc-tum granulatum indicate the occurrences in the Piney Point Formation (middle Eocene) inVirginia (adapted from Edwards, 1982).
53W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
Geographic and stratigraphic distribution: Pentadinium alabamensissp. nov. has so far been reported from four localities; Jaramillo andOboh-Ikuenobe (1999) recorded it in two cores in Mississippi State(Mobil Exploration and Production Services #1 Young and #1Wayne cores), as well as from the SSQ outcrop (see Fig. 1). Wefound P. alabamensis sp. nov. in the SSQ core, taken ca 1.6 kmfrom the SSQ outcrop analyzed by Jaramillo and Oboh-Ikuenobe(1999). The species has never been mentioned or shown in otherregions.
Quantitatively, Pentadinium alabamensis sp. nov. may contributesubstantially to the dinocyst assemblages (up to 10% in the SSQcore). Furthermore, P. alabamensis sp. nov. co-occurs with taxa suchas Homotryblium spp. and Deflandrea spp. at all localities (see Fig. 4and fig. 9 of Jaramillo and Oboh-Ikuenobe, 1999). These taxa are typ-ically found in relatively restricted, shallow marine environments,characterized by a relatively large range in salinity (e.g., Brinkhuis,1994; Pross and Schmiedl, 2002; Röhl et al., 2004; Pross andBrinkhuis, 2005). At the SSQ core, samples in which P. alabamensissp. nov. occurs are characterized by high relative abundances ofterrestrial palynomorphs (i.e., pollen and spores) and a highBranched and Isoprenoid Tetraether (BIT)-index as well (Wade etal., 2012, GSA Data repository; see Fig. 4). The BIT-index is a tracerfor terrestrially derived soil bacterial lipids and a high BIT maytherefore point toward a high influx of terrestrial organic matterin sediments (e.g., Hopmans et al., 2004). In addition, Miller et al.(2008) characterized the Red Bluff Clay, where P. alabamensis hasits highest occurrence (see Fig. 4), as a shelf environment with adepth of 75±15 m, based on the benthic foraminiferal Hanzawaiabiofacies (Miller et al., 2008, and references therein). Altogether,this suggests that P. alabamensis sp. nov. is associated with proxi-mal environments.
As can be seen in Fig. 4 and implied from table 8 and fig. 17 ofJaramillo and Oboh-Ikuenobe (1999) Pentadinium alabamensis sp.nov. typically has its highest relative abundance in the OligoceneRed Bluff Clay and upper Forest Hill Sand (not present at SSQ)(see Fig. 4). The lowest occurrence of P. alabamensis sp. nov. in allfour locations may differ slightly (Fig. 4; Jaramillo and Oboh-Ikuenobe, 1999), but it always occurs above the Eocene–Oligoceneboundary (not be confused with the younger Oi-1 isotope event).The oldest first occurrence out of all 4 locations is an occurrenceof a single specimen within the SSQ core at 46.74 m, at a horizoncalibrated to be against the very top of, but still within, magneto-chron C13r. At all locations, P. alabamensis sp. nov. has its highestoccurrence at the Red Bluff Clay/Forest Hill Sand–Mint Spring Marlcontact, except for the location of the #1 Wayne core. However, inthat location it concerns only a single specimen, occurring 0.9 mabove the Forest Hill Sand–Mint Spring Marl contact; it may repre-sent reworking. The boundary of the Red Bluff Clay–Mint SpringMarl contact at SSQ is calibrated to be against the subchronC13n–C12r boundary. Accordingly, we place the last occurrenceof P. alabamensis sp. nov. at this boundary, dated at ~33.058 Ma inBerggren et al. (1995). This implies that the range of P. alabamensissp. nov. essentially straddles magnetosubchron C13n (see Fig. 2).In Fig. 4 the lowest and highest occurrences (LO; HO) of P. alaba-mensis can be compared to other biostratigraphically importantspecies. The LO of P. alabamensis is ca. 1.3 m above the HO ofHemiplacophora semilunifera, a species that can be associatedwith the Eocene–Oligocene boundary (Brinkhuis and Biffi, 1993).This further confirms the early Oligocene age of the range ofP. alabamensis.
In 1982, Edwards already stated that Pentadinium speciesoccur widely throughout the Eocene and the Oligocene and thatthey might provide good biostratigraphic markers. She introduceda scheme of ranges of middle and upper Eocene Pentadiniumspecies. We can now expand this scheme up into the Oligocene(see Fig. 5).
4. Concluding remarks
The distinctive new dinocyst species Pentadinium alabamensis sp.nov. appears to be a useful regional biostratigraphic marker speciesfor magnetosubchron C13n in the Gulf Coast region. In addition,we surmise that the taxon reflects shallow marine, euryhaline set-tings. Future analysis at other locations in and outside the Gulf ofMexico will have to further confirm the age ranges given, as wellas further elucidate the environmental preferences of P. alabamensissp. nov.
Acknowledgments
N.L.D.Welters, L.P.M. Bik and J. J. van Tongeren are thanked for tech-nical support. Prof. Ken Miller and co-workers (Rutgers University, NJ,USA) have kindly provided the samples. They, as well as dr. BridgetWade (University of Leeds, UK), are kindly thanked for cooperation.Dr. C. Jaramillo (Smithsonian) is thanked for providing additional infor-mation on Pentadinium sp. A. The two reviewers, dr. Lucy E. Edwardsand dr. Karin Zonneveld, as well as Sander Houben MSc are thankedfor carefully reading the manuscript and suggesting improvements.
References
Benedek, P.N.v., Gocht, H., Sarjeant, W.A.S., 1982. The dinoflagellate cyst genus Pentadi-nium Gerlach: a re-examination. Neues Jahrbuch für Geologie und Paläontologie,Abhandlungen 162, 265–295.
Berggren, W.A., Kent, D.V., Swisher, C.C., Aubry, M.-P., 1995. A revised Cenozoic geochro-nology and chronostratigraphy. In: Berggren, W.A., Kent, D.V., Aubry, M.-P.,Hardenbol, J. (Eds.), Geochronology, time scales and stratigraphic correlation: Societyof Economic Paleontologists and Mineralogists Special Publication, 54, pp. 129–212.
Brinkhuis, H., 1994. Late to Early Oligocene dinoflagellate cysts from the Priabionantype-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation.Palaeogeography, Palaeoclimatology, Palaeoecology 107, 121–163.
Brinkhuis, H., Biffi, U., 1993. Dinoflagellate cyst stratigraphy of the Eocene/Oligocenetransition in Central Italy. Marine Micropaleontology 22, 131–183.

54 W. Quaijtaal, H. Brinkhuis / Review of Palaeobotany and Palynology 175 (2012) 47–54
Bujak, J.P., Mudge, D., 1994. A high-resolution North Sea dinocyst zonation. Journal ofthe Geological Society of London 151, 449–462.
Bütschli, O., 1885. Erster Band. Protozoa. Dr. H.G. Bronn's Klassen und Ordnungen desThier-Reichs, wissenschaftlich dargestellt in Wort und Bild. C.F. Winter'scheVerlagsbuchhandlung, Leipzig and Heidelberg, pp. 865–1088.
Coxall, H.K., Pearson, P.N., 2007. The Eocene–Oligocene transition. In: Williams, M.,Haywood, A.M., Gregory, F.J., Schmidt, D.N. (Eds.), Deep-time perspectives on climatechange: marrying the signal from computer models and biological proxies: TheMicropalaeontologcal Society, Special Publications, London, pp. 351–387.
Coxall, H.K., Wilson, P.A., Pälike, H., Lear, C.H., Backman, J., 2005. Rapid stepwise onset ofAntarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature433, 53–57.
Edwards, L.E., 1982. Biostratigraphically important species of Pentadinium Gerlach1961 and a likely ancestor, Hafniasphaera goodmanii n. sp., from the Eocene ofthe Atlantic and Gulf Coastal Plains. Palynology 6, 105–117.
Eldrett, J.S., Harding, I.C., Firth, J.V., Roberts, A.P., 2004. Magnetostratigraphic calibration ofEocene–Oligocene dinoflagellate cyst biostratigraphy from the Norwegian–GreenlandSea. Marine Geology 204, 91–127.
Evitt, W.R., 1985. Sporopollenin Dinoflagellate Cysts: Their Morphology and Interpreta-tion. American Association of Stratigraphic Palynologists Foundation, Dallas. 300 pp.
Evitt, W.R., Lentin, J.K., Millioud, M.E., Stover, L.E., Williams, G.L., 1977. Dinoflagellatecyst terminology. Geological Survey of Canada Paper 76 (24), 1–11.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I., Williams, G.L.,1993. A classification of fossil and living dinoflagellates. Micropaleontology PressSpecial Paper, vol. 7. 351 pp.
Fensome, R.A., MacRae, R.A., Williams, G.L., 2008. DINOFLAJ2, Version 1. American Asso-ciation of Stratigraphic Palynologists, Data Series no. 1.
Fensome, R.A., Williams, G.L., MacRae, R.A., 2009. Late Cretaceous and Cenozoic fossildinoflagellates and other palynomorphs from the Scotian Margin, offshore easternCanada. Journal of Systematic Palaeontology 7 (1), 1–79.
Gerlach, E., 1961. Mikrofossilien aus dem Oligozän und Miozän Nordwestdeutschlands,under besonderer Berücksichtigung der Hystrichosphaeren und Dinoflagellaten.Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 112, 143–228.
Gocht, H., 1969. Formengemeinschaften alttertiären Mikroplanktons aus Bohrprobendes Erdölfeldes Meckelfeld bei Hamburg. Palaeontographica, Abteilung B 126,1–100.
Hopmans, E.C., Weijers, J.W.H., Schefuß, E., Herfort, L., Sinninghe Damsté, J.S., Schouten, S.,2004. A novel proxy for terrestrial organic matter in sediments based on branchedand isoprenoid tetraether lipids. Earth and Planetary Science Letters 224, 107–116.
Jaramillo, C.A., Oboh-Ikuenobe, F.E., 1999. Sequence stratigraphic interpretations frompalynofacies, dinocyst and lithological data of Upper Eocene–Lower Oligocenestrata in southern Mississippi and Alabama, U.S., Gulf Coast. Palaeogeography,Palaeoclimatology, Palaeoecology 145, 259–302.
Lindemann, E., 1928. Abteilung Peridineae (dinoflagellatae). In: Engler, A., Prantl, K.(Eds.), Die Natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigerenArten insbesondere den Nutzpflanzen. Zweite stark vermehrte und verbesserte
Auflage herausgegeben von A. Engler. 2 Band. Wilhelm Engelmann, Leipzig,pp. 3–104.
Miller, K.G., Thompson, P.R., Kent, D.V., 1993. Integrated late Eocene–Oligocene stratigra-phy of the Alabama coastal plain: correlation of hiatuses and strata surfaces toglacioeustatic lowerings. Paleoceanography 8 (2), 313–331.
Miller, K.G., Browning, J.V., Aubry, M.-P., Wade, B.S., Katz, M.E., Kulpecz, A.A., Wright, J.D.,2008. Eocene–Oligocene global climate and sea-level changes: St. Stephens Quarry,Alabama. Geological Society of America Bulletin January/February 120 (1–2), 34–53.
Pascher, A., 1914. Über Flagellaten und Algen. Deutsche Botanische Gesellachaft, Berichte32, 136–160.
Pross, J., Brinkhuis, H., 2005. Organic-walled dinoflagellate cysts as paleoenvironmentalindicators in the Paleogene; a synopsis of concepts. Paläntologische Zeitschrift 79(1), 53–59.
Pross, J., Schmiedl, G., 2002. Early Oligocene dinoflagellate cysts from the Upper RhineGraben (SW Germany): paleoenvironmental and paleoclimatic implications. MarineMicropaleontology 45, 1–24.
Röhl, U., Brinkhuis, H., Stickley, C.E., Fuller, M., Schellenberg, S.A., Wefer, G., Williams, G.L.,2004. Sea level and astronomically induced environmental changes in middle andlate sediments from the East Tasman Plateau. In: Exon, N.F., Kennett, J.P., Malone,M.J. (Eds.), The Cenozoic Southern Ocean: tectonics, sedimentation and climatechange between Australia and Antarctica: American Geophysical Union, GeophysicalMonograph Series, 151, pp. 127–151.
Sancay, R.H., Bati, Z., Edwards, L.E., Ertug, K.I., 2006. A new species of Pentadinium fromEastern Anatolia, Turkey, Pentadinium galileoi. Micropaleontology 52 (6), 537–543.
Schiøler, P., 2005. Dinoflagellate cysts and acritarchs from the Oligocene-Lower Mio-cene interval of the Alma-1X well, Danish North Sea. Journal of Micropalaeontol-ogy 24, 1–37.
Sluijs, A., Brinkhuis, H., Stickley, C.E., Warnaar, J., Williams, G.L., Fuller, M., 2003. Dinofla-gellate cysts from the Eocene–Oligocene transition in the Southern Ocean: resultsfrom ODP Leg 189. In: Exon, N.F., Kennett, J.P., Malone, M.J. (Eds.), Proceedings ofthe Ocean Drilling Program: Scientific Results, 189, pp. 1–42.
Taylor, F.J.R., 1980. On dinoflagellate evolution. Biosystems 13, 65–108.Tew, B.H., Mancini, E.A., 1995. An integrated stratigraphic model for paleogeographic
reconstruction: examples from the Jackson and Vicksburg Groups of the easternGulf Coastal Plain. Palaios 10, 133–153.
Wade, B.S., Houben, A.J.P., Quaijtaal, W., Schouten, S., Rosenthal, Y., Katz, M.E., Miller, K.G.,Wright, J.D., Brinkhuis, H., 2012. Multiproxy record of abrupt sea-surface coolingacross the Eocene–Oligocene transition in the Gulf of Mexico. Geology 40 (2),159–162.
Zachos, J.C., Quinn, T.M., Salamy, K.A., 1996. High-resolution (104 years) deep-sea forami-niferal stable isotope records of the Eocene–Oligocene climate transition. Paleocea-nography 11 (3), 251–266.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001. Trends, rhythms, and aber-rations in global climate 65 Ma to present. Science 292, 686–693.
Zachos, J.C., Dickens, G.R., Zeebe, R.E., 2008. An early Cenozoic perspective on greenhousewarming and carbon-cycle dynamics. Nature 451, 279–283.