paterson, j.r., 2004. palaeobiogeography of the ordovician trilobite prosopiscus, with a new species...
TRANSCRIPT

Palaeobiogeography of the Ordovic ian trilobite Prosopiscus, with a new species from western New South Wales
JOHN R. PATERSON
PATERSON, J. R., 30.4.2004. Palaeobiogeography of the Ordovician trilobite Prosopiscus, with a new species from western New South Wales. Alcheringa 28, 65-76. ISSN 0311- 5518.
Prosopiscus is particularly important in Ordovician palaeobiogeography because of its wide geographic distribution in Gondwana and peri-Gondwanan regions. It appears to have been confined to low palaeolatitudes, representing a characteristic member of the warm water eastern Gondwanan shelf faunas. Trends in the distribution of the Ordovician genus can be observed due to its long stratigraphic range. Prosopiscus was restricted to, and may have originated in, Australia during the late Early Ordovician (Bendigonian- Chewtonian). By the Middle Ordovician (Darriwilian), Prosopiscus had dispersed to other parts of Gondwana and peri-Gondwana, including the North and South China blocks, Tarim, central Himalayas, and the Argentine Precordillera (South America). Possible explanations for the distribution of Prosopiscus are that: (1) there were no oceanic barriers preventing dispersal of trilobites between different regions of Gondwana, thus permitting uninhibited migration over vast distances; (2) Prosopiscus was not restricted to a specific biofacies; (3) a major eustatic transgression during the early Darriwilian may have facilitated the dispersal of Prosopiscus in allowing further devel- opment and expansion of marine environments; and (4) a prolonged planktonic larval stage may have permitted wide dispersal. Prosopiscus lauriei sp. nov. is described from the late Early Ordovician (Bendigonian- Chewtonian) Tabita Formation at Mount Arrowsmith, northwestern New South Wales, Australia. The new species is closely related to P praecox, from the Nora Formation, Georgina Basin, central Australia, and to P magicus from northwest China.
John R. Paterson [[email protected]], Centre for Ecostratigraphy & Palaeobiology, Department of Earth & Planetary Sciences, Macquarie University, North Ryde, NSW 2109, Australia; received 1.8.2002, revised 11.11.2002.
Key words: Prosopiscus, Trilobita, Ordovieian, palaeobiogeography, palaeogeography, New South Wales, Australia, Gondwana.
THE IMPORTANCE of Prosopiscus as a potentially useful taxon in Ordovician palaeobiogeography has been discussed by several authors (Webby et al. 2000, Zhou & Dean 1989, Cocks & Fortey 1990). The first real attempt to develop a biogeographic model for Prosopiscus was that of Webby et al. (2000), with minor treatment of the genus in previous studies by Zhou & Dean (1989), Fortey & Owens (1990), Edgecombe et al. (1999), Cocks & Fortey (1990) and Burrett & Stait (1987). In an attempt 0311/5518/2004/01065-12 $3.00 ©AAP
to establish a workable, biogeographic-ally distinct, faunal nomenclature for Australasian and neighbouring regions, Webby et al. (2000, p. 95) proposed the 'Prosopiscus fauna' - the name-bearer chosen to 'reflect only the temporal range of the specific time interval of the proposed fauna [in this case, Darriwilian], but in a few cases these key provincial elements have more extended stratigraphic ranges (e.g. Hysterolenus, Prosopiscus, Pliomerina)' . In relation to provincial nomenclature, Webby (1971) had proposed the 'Pliomerina fauna' based on the
Downloaded By: [Macquarie University] At: 02:07 6 December 2008
66 JOHN PATERSON ALCHERINGA
L..
CG
0 co
0 20 40 km I I I I I
Mt Poole A
Mt Browne •
Mt Shannon
"Mt Arrowsmith"
q f
q.
, . - . . ,
Tibooburra
l 30 ° s
Packsaddle
N
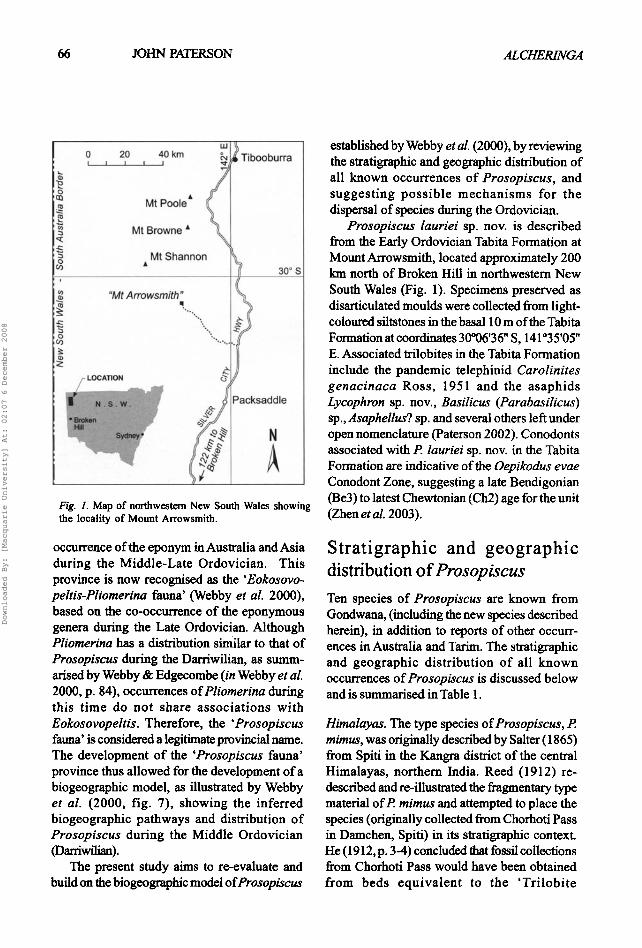
A Fig. 1. Map of northwestern New South Wales showing the locality of Mount Arrowsrnith.
occurrence of the eponym in Australia and Asia during the Middle-Late Ordovician. This province is now recognised as the 'Eokosovo- peltis-Pliomerina fauna' (Webby et al. 2000), based on the co-occurrence of the eponymous genera during the Late Ordovician. Although Pliomerina has a distribution similar to that of Prosopiscus during the Darriwilian, as summ- arised by Webby & Edgecombe (in Webby et al. 2000, p. 84), occurrences of Pliomerina during this time do not share associations with Eokosovopeltis. Therefore, the "Prosopiscus fauna' is considered a legitimate provincial name. The development of the 'Prosopiscus fauna' province thus allowed for the development of a biogeographic model, as illustrated by Webby et al. (2000, fig. 7), showing the inferred biogeographic pathways and distribution of Prosopiscus during the Middle Ordovician ( D a r r i ~ ) .
The present study aims to re-evaluate and build on the biogeographic model of Prasopiscus
established by Webby et aL (2000), by reviewing the stratigraphic and geographic distribution of all known occurrences of Prosopiscus, and suggesting possible mechanisms for the dispersal of species during the Ordovician.
Prasopiscus lauriei sp. nov. is described from the Early Ordovician Tabita Formation at Mount An'owsmith, located approximately 200 km north of Broken Hill in northwestern New South Wales (Fig. 1). Specimens preserved as disarticulated moulds were collected from light- coloured siltstones in the basal 10 m of the Tabita Formation at coordinates 30006'36 " S, 141°35'05 " E. Associated trilobites in the Tabita Formation include the pandemic telephinid Carolinites genacinaca Ross, 1951 and the asaphids Lycophron sp. nov., Basilicus (Parabasilicus) sp., Asaphellus? sp. and several others left under open nomenclature (Paterson 2002). Conodonts associated with P lauriei sp. nov. in the Tabita Formation are indicative of the Oepikodus evae Conodont Zone, suggesting a late Bendigonian (Be3) to latest Chewtonian (Ch2) age for the unit (Zhen etaL 2003).
Stratigraphic and geographic distribution o f Prosopiscus
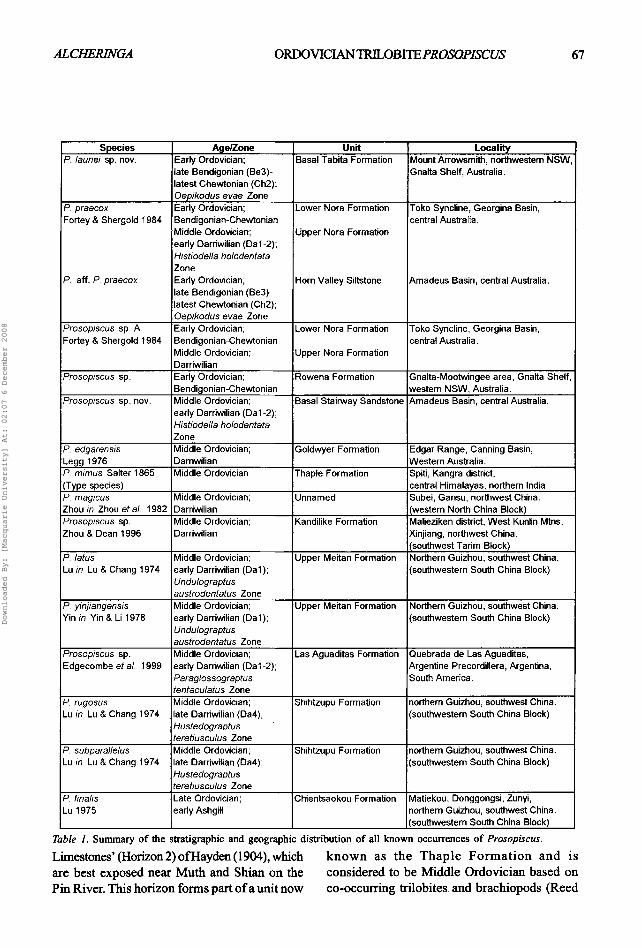
Ten species of Prosopiscus are known from Gondwana, (including the new species described herein), in addition to reports of other occurr- ences in Australia and Tarim. The stratigraphic and geographic distribution of all known occurrences of Prosopiscus is discussed below and is summarised in Table 1.
Himalayas. The type species of Prosopiscus, P.. mimus, was originally described by Salter (1865) from Spiti in the Kangra district of the central Himalayas, northern India. Reed (1912) re- described and re-illustrated the fragmentary type material ofP mimus and attempted to place the species (originally collected from Chorhoti Pass in Damchen, Spiti) in its stratigraphic context. He (1912, p. 3-4) concluded that fossil collections from Chorhoti Pass would have been obtained from beds equivalent to the 'Trilobite
Downloaded By: [Macquarie University] At: 02:07 6 December 2008
AL CHERINGA ORDO VICIAN TRILOBITE PR OSOPISCUS 6 7
Species P. ~unei sp. nov.
P. praecox Fortey & Shergold 1984
P. aft. P. praecox
Age/Zone Early Ordovician; late Bendigonian (Be3) latest Chewtonian (Ch2); Oepikodus evae Zone Early Ordovician; Bendigonian-Chewtonian
Unit Basal Tabita Formation
Lower Nora Formation
Prosopiscus sp. A Fortey & Shergold 1984
iProsopiscus sp.
Prosopiscus sp. nov.
!P. edgarensis Legg 1976 P. mimus Salter 1865 (Type species) P. magicus Zhou in Zhou etal. 1982 Prosopiscus sp. Zhou & Dean 1996
P latus Lu in Lu & Chang 1974
P. yinjiangensis Yin in Yin & Li 1978
Prosopiscus sp. Edgecombe et al. 1999
P. rugosus Lu in Lu & Chang 1974
P. subparallelus Lu in Lu & Chang 1974
P. finalis Lu 1975
Middle Ordovician; early Dardwilian (Dal-2); Histiodella holodentata Zone Early Ordovician; late Bendigonian (Be3)- latest Chewtonian (Ch2); Oepikodus evae Zone Early Ordovician; Bendigonian-Chewtonian Middle Ordovician; Darriwilian Early Ordovician; Bendigonian-Chewtonian Middle Ordovician; early Darriwilian (Dal-2); Histiodella holodentata Zone Middle Ordovician; Darriwilian Middle Ordovician
Middle Ordovician; Darriwilian Middle Ordovician; Darriwilian
Middle Ordovician; early Darriwilian (Dal); Undulograptus austrodentatus Zone Middle Ordovician; early Darriwilian (Dal); Undulograptus austrodentatus Zone Middle Ordovician; eady Darriwilian (Dal-2); Paraglossograptus tentaculatus Zone Middle Ordovician; late Darriwilian (Da4); Hustedograptus teretiusculus Zone Middle Ordovician; late Darriwilian (Da4); Hustedograptus teretiusculus Zone Late Ordovician; early Ashgill
Upper Nora Formation
Horn Valley Siltstone
Lower Nora Formation
Upper Nora Formation
Rowena Formation
Basal Stairway Sandstone
Goldwyer Formation
Thaple Formation
Unnamed
Kandilike Formation
Upper Meitan Formation
Upper Meitan Formation
Las Aguaditas Formation
Shihtzupu Formation
Shihtzupu Formation
Chientsaokou Formation
Locality Mount Arrowsmith, northwestern NSW !Gnalta Shelf, Australia.
Toko Syncline, Georgina Basin, central Australia.
Amadeus Basin, central Australia.
Toko Syncline, Georgina Basin, central Australia.
Gnalta-Mootwingee area, Gnalta Shelf western NSW, Australia. Amadeus Basin, central Australia.
Edgar Range, Canning Basin, Western Australia. Spiti, Kangra district, central Himalayas, northern India. Subei, Gansu, northwest China. (western North China Block) Malieziken district, West Kunlin Mtns, Xinjiang, northwest China. (southwest Tarim Block) Northern Guizhou, southwest China. (southwestern South China Block)
Northern Guizhou, southwest China. (southwestern South China Block)
Quebrada de Las Aguaditas, Argentine Precordillera, Argentina, South America.
northern Guizhou, southwest China. (southwestern South China Block)
northern Guizhou, southwest China. (southwestern South China Block)
Matiekou, Donggongsi, Zunyi, iorthern Guizhou, southwest China. (southwestern South China Block)
Table 1. Summary of the stratigraphic and geographic distlibution of all known occurrences of Prosopiscus.
Limestones' (Horizon 2) ofHayd~ (1904), which known as the Thaple Formation and is are best exposed near Muth and Shian on the considered to be Middle Ordovician based on Pin River. This horizon forms part of a unit now co-occurring trilobites and brachiopods (Reed
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

68 JOHN PATERSON ALCHERINGA
1912, Baud et aL 1984, Gaetani et aL 1986). A reconstruction of the cephalon of the type species was illustrated by Fortey & Shergold (1984, p. 359) based on fragments in the type collection housed in the Natural History Museum, London. Syntypes of P mimus were subsequently re- illustrated by Morris & Fortey (1985, pl. 4, figs 2-4).
Australia. Prosopiscus is known from several Early and Middle Ordovician cratonic successions across Australia. Fortey & Shergold (1984) described P praecox and Prosopiscus sp. A from the Nora Formation, Georgina Basin, central Australia. Both species range from the lower to upper Nora Formation, which is considered to span the Early to Middle Ordovician. Conodonts reported from the lower Nora Formation by Stait & Druce (1993) suggest a late Early Ordovician (Bendigonian- Chewtonian) age; this is further supported by the restricted occurrence of Carolinites genacinaca in the lower Nora Formation (Fortey & Shergold 1984). Age constraints on the upper Nora Formation are lacking; however Nicoll (in Shergold 1991) reported Histiodella holodentata from the unit, implying a Middle Ordovician (Darriwilian) age.
Documentation of Ordovician trilobite faunas from the Amadeus Basin of central Australia remains incomplete, with the most recent report being that by Shergold & Laurie (in Shergold 1991). Two species of Prosopiscus are known from the Amadeus Basin: P. aft. P praecox from the Horn Valley Siltstone, and a new species (similar to P mimus) from the overlying basal Stairway Sandstone (Dr John Laurie, pets. comm., 2002). Conodont studies by Cooper ( 1981 ), NicoU (in Shergold 1991), and Zhen et al. (2001) suggest a late Early Ordovician (Bendigonian- Chewtonian) age for the Horn Valley Siltstone, whereas the Stairway Sandstone was considered by Shergold (1991) and Young & Laurie (1996) to be Middle Ordovieian (early Darriwilian).
Early Ordovician trilobite faunas from the Gnalta Shelf of western New South Wales are also poorly documented. Occurrences of Prosopiscus include P. lauriei sp. nov. (described herein) from the basal Tabita Formation at Mount
Arrowsmith, and an undescribed species from the middle Rowena Formation in the Gnalta- Mootwingee area (Shergold 1971, Webby et al. 1981, Shergold et al. 1982, Webby 1983). Prosopiscus lauriei sp. nov. is considered to be late Early Ordovician (Bendigonian-Chewtonian) in age based on co-occurring conodont assemblages (Zhen et al. 2003). The occurrence of Carolinites genacinaca in the Rowena Formation is indicative of a late Early Ordovician (Bendigonian-Chewtonian) age for the unit.
Legg (1976) described P edgarensis from the Middle Ordovician Goldwyer Formation in the Canning Basin, Western Australia. Legg (1976, 1978) recorded specimens from the upper part of the unit, associated with the graptolites Didymograptus artus, D. stabilis, Undulo- graptus situlus, Archiclimacograptus confertus and Hallograptus hirtus, which imply an early- mid Darriwilian age (VandenBerg & Cooper 1992, Chen et al. 2001 a). The Goldwyer Formation is considered to be Darriwilian, based on conodont and chitinozoan studies (Watson 1988, Nicoll et al. 1993, Winchester-Seeto et al. 2000). Watson (1988) inferred a Darriwilian (Da3) age for conodonts described from the upper Goldwyer and Nita formations. Subsequently, Nicoll et al. ( 1993 ) considered the lower Goldwyer Formation to fall within the Histiodella holodentata Conodont Zone and the upper Goldwyer and Nita formations within the Phragmodus plectodina Conodont Zone. Winchester-Seeto et al. (2000, p. 280) concluded that 'the chitinozoan assemblage from the upper Goldwyer and Nita formations can be correlated with the presently "undefined zone" immediately above the Cyathochitina jenkinsi Zone in the Laurentian biozonation', suggesting a latest Darriwilian (Da4) age.
North China. The only occurrence of Prosopiscus in North China is that ofP magicus, described by Zhou (in Zhou et al. 1982) from a single cranidium. It occurs in an unnamed rock unit of littoral to shallow-water elastics in Subei, Gansu, northwest China, associated with asaphids and the brachiopod Xizangostrophia (Dr Zhou Zhiyi, pers. comm., 2002). Zhou et al.
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

ALCHERINGA ORDOVICIAN TR/LOBITE PROSOPISCUS 69
(1982) and Zhou & Dean (1989) considered the species to be ofLlanvirn (=Darriwilian) age.
South China. Five species of Prosopiscus have been described from northern Guizhou, southwest China. Prosopiscus rugosus Lu (in Lu & Chang 1974) andP subparallelus Lu (in Lu & Chang 1974) are based on cranidia from the Middle Ordovician Shihtzupu Formation, both species occurring at the type locality in Feilaishi, north of Zunyi City. However, P subparallelus is also known from Hsinglungchang, Suiyang district (Lu & Chang 1974, Lu 1975, Yin & Li 1978, Zhou et al. 1984). Zhou et al. (1984) noted that sparse graptolites occurring throughout the Shihtzupu Formation type locality at Feilaishi are probably attributable to the teretiusculus Zone, hence suggesting a late Darriwilian (Da4) age for the unit.
Two species of Prosopiscus have been described from the Meitan Formation of northern Guizhou: P. latus Lu (in Lu & Chang 1974) andP yingjiangensis Yin (in Yin & Li 1978). Although the species are probably synonymous, they are treated as separate species here. Both occur in the upper part of the formation (Dr Zhou Zhiyi, pers. comm., 2002), within the Undulograptus austrodentatus graptolite Zone, indicating a basal Darriwilian (Dal) age (Chen et al. 1995).
Lu (1975) described P. finalis from the Chientsaokou (=Jiantsaokou) Formation at Matiekou, Donggongsi, north of Zunyi City. This species is associated with the graptolites Dicellograptus cf. D. johnstrupi and Recto- graptuspauperatus, implying a Late Ordovician (earlyAshgill) age (Chen et al. 1995, 2000, 2001b).
Tarim. Zhou & Dean (1996, p. 169) reported an undescribed species of Prosopiscus from the lower part of the Kandilike Formation in the Malieziken district of the West Kunlin Mountains, Xinjiang. The associated cephalo-pods Wutinoceras and Adamsoceras from the lower part of the unit suggest a probable Middle Ordovician age.
Argentina. Edgecombe et al. (1999) documented a species of Prosopiscus, (left under open nomenclature due to a paucity of holaspid
material), from the Argentine Precordillera (South America), representing the first record of protaspid larvae and a hypostome for the genus. These silicified specimens were recovered from the Las Aguaditas Formation, Quebrada de Las Aguaditas, Argentina, and are considered to be early-mid Darriwilian (Paraglossograptus tentaculatus graptolite Zone).
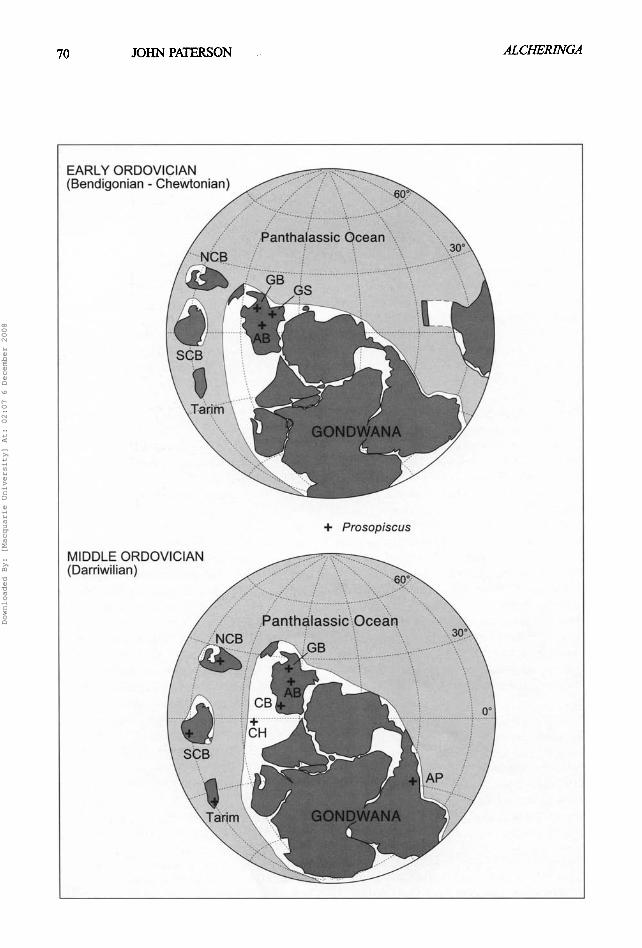
Palaeobiogeographic implications In their synthesis of the palaeobiogeography of Australasian Ordovician faunas, Webby et al. (2000) presented a detailed review of Ordovician palaeogeography, particularly for Gondwana and the surrounding crustal blocks, such as North and South China and Tarim. The global maps used by Webby et al. (2000) for the Early and Middle Ordovician are employed here (Fig. 2) to illustrate the distribution of Prosopiscus during the Ordovician.
Based on stratigraphic data (Table 1), it appears that Prosopiscus was restricted to, and may have originated in Australia during the late Early Ordovician (Bendigonian-Chewtonian). Although the phylogeny of Prosopiscus has only been briefly discussed by Fortey & Shergold (1984, p. 357-358) and Edgecombe et al. (1999, p. 1149-1153), the biogeographic hypothesis presented herein assumes that phylogenetic patterns within the genus mirror the temporal distribution of species. Australian Early Ordovician shallow marine successions containing Prosopiscus were deposited during a transgressive episode (the Oepikodus evae transgression: Nielsen 1992, Barnes et al. 1995), resulting in the development of a trans-cratonic (epicontinental) seaway called the Larapintine Sea (Webby 1978, Nicoll et al. 1988). The development of this seaway and its continuity into the Middle Ordovician allowed for a major expansion of shallow marine depositional conditions, connecting cratonic basins such as the Amadeus and Georgina basins with shelf margins (Gnalta Shelf) and the Panthalassic Ocean to the east, and the Canning Basin to the west (Fig. 3). The Larapintine Sea provided the
Downloaded By: [Macquarie University] At: 02:07 6 December 2008
70 JOHN PATERSON ALCI~RINGA
E A R L Y O R D O V I . . . . . ( B e n d i g o n i a n - C
M I D D L E ORDO~ . . . . . . . (Dar r iw i l ian)
+ Prosopiscus
Downloaded By: [Macquarie University] At: 02:07 6 December 2008
A L CH ERING A ORDOVICIAN TRILOBITE PROSOPISCUS 71
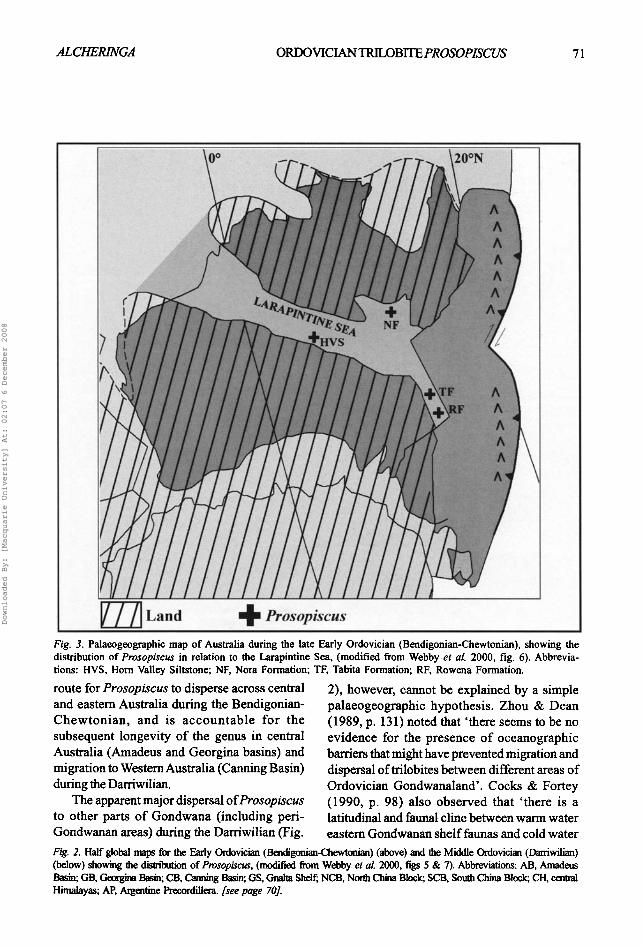
~--~ Land 4= Prosopiscus Fig. 3. Palaeogeographie map of Australia during the late Early Ordovieian (Bendigonian-Chewtonian), showing the distribution of Prosopiscus in relation to the Larapintine Sea, (modified from Webby et al. 2000, fig. 6). Abbrevia- tions: HVS, Horn Valley Siltstone; NF, Nora Formation; TF, Tabita Formation; RF, Rowena Formation.
route for Prosopiscus to disperse across central and eastern Australia during the Bendigonian- Chewtonian , and is accoun tab le for the subsequent longevity of the genus in central Australia (Amadeus and Georgina basins) and migration to Western Australia (Canning Basin) during the Darriwilian.
The apparent major dispersal o f Prosopiscus to other parts o f Gondwana (including peri- Gondwanan areas) during the Darriwilian (Fig.
2), however, cannot be explained by a simple palaeogeographic hypothesis. Zhou & Dean (1989, p. 131) noted that 'there seems to be no evidence for the presence o f oceanographic barriers that might have prevented migration and dispersal of trilobites between different areas of Ordovician Gondwanaland'. Cocks & Fortey (1990, p. 98) also observed that ' there is a latitudinal and faunal cline between warm water eastern Gondwanan shelf faunas and cold water
Fig. 2. Half global maps for the Early Ordovieian (Bendigonia~-Chewtonian) (above) and the Middle Ordoviei~a (Darriwilian) (below) showing the distribution of Prosopiscus, (mo~ed from Webby et aL 2000, figs 5 & 7). Abbreviations: AB, Amadens Basin; GB, G ~ Basin; CB, C a ~ Basin; GS, Gnalla Shelt~ NCB, Norlh China Block; SCB, Soulh China Block; CH, central Himalayas; AP, Argentine Prceordillera. [see page 70].
Downloaded By: [Macquarie University] At: 02:07 6 December 2008
72 JOHN PATERSON ALCHERINGA
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

AL CHERINGA ORDO VI CIAN TRILOBITE PR OSOPISCUS 73
western Gondwanan she l f faunas, but no evidence of the disjunct distribution which might indicate a vanished oceanic barrier'. More recently, Zhou et al. (1998a, b) suggested that the peri-Gondwanan North and South China blocks and the Tarim block were situated close to each other at low palaeolatitudes, were not separated by significant oceanic barriers, and may have formed part of Gondwana.
Prosopiscus seems to have inhabited a variety of marine settings, from shallow-water, inshore facies (e.g. P praecox from the Nora Formation) to deeper-water platform or slope facies (e.g. P edgarensis from the Goldwyer Formation). Therefore Prosopiscus was probably not restricted to a particular biofacies like the majority of benthic trilobites in platform settings, thus allowing migration over vast distances and various facies belts. This hypothesis corres- ponds to one of the informal biogeographic groups of Ordovician trilobites (Group B) established by Zhou & Dean (1989). Prosopiscus differs from other members assigned to this group in seemingly being confined to low palaeolatitudes (Fig. 2).
As mentioned above, the Larapintine Sea persisted well into the Middle Ordovician. Following a short-lived regressive episode, a major eustatic marine transgression during the early Darriwilian (Nicoll et al. 1988, Nielsen 1992, Barnes et aL 1995) may have induced the dispersal of Prosopiscus to Western Australia, the Himalayas, Tarim, and the North and South China blocks, as illustrated by Webby et al. (2000, fig. 7). Whether the occurrence in the Argentine Precordillera (South America) was the result of a Chinese or Australian influence is uncertain. This sudden dispersal of Prosopiscus species during the Daniwilian ~ to correspond to lhe Basal Llanvim Bio-Event (13 'Ira) ofBames eta/. (1995), charactefis~ by the basal Llanvim (or Darriwilian) regressive- transgressive couplet, and marks a period of rapid
diversification across most faunal groups. Fortey & Shergold (1984) have suggested
that widely dis t r ibuted genera (such as Annamitella and Presbynileus) may be the result of efficient larval dispersal. Studies by Chatterton & Speyer (1989, 1990, 1997) and Speyer & Chatterton (1989, 1990) on the larval ecology of t r i lobi tes have shown that species with 'p lanktonic , nonadul t - l ike la rvae ' had a widespread palaeogeographic distribution. Although the protaspides o f Prosopiscus illustrated in Edgecombe et al. (1999, figs 5.1- 5.4) probably represent an initial benthic stage (P2), they bear close similarity to the equivalent stage in the pterygometopid Calyptaulax (as discussed in Edgecombe et aL 1999), of which the smallest protaspid instar represents a nonadult-like form (Chatterton & Speyer 1997, fig. 146.4). If Prosopiscus did indeed have a nonadult-like larval stage, the genus could be placed within 'Life-history strategy II' o f Chatterton & Speyer (1997). In this strategy the first calcified protaspid (or larval) stage is p lanktonic and later stages are benthic. Chatterton & Speyer (1989) also observed that the proportion of taxa with planktonic larvae increased towards the equator. This trend also supports the idea o f Prosopiscus having planktonic larvae, since all known occurrences of Prosopiscus are confined to low palaeo- latitudes (Fig. 2). Therefore the wide distribution of Prosopiscus is due, in part, to the likelihood of the genus having a prolonged planktonic larval stage.
Systematic palaeontology Specimens are catalogued in the palaeontological collections of the Australian Museum, Sydney (pretixAMF).
Class TRILOBITAWalch, 1771
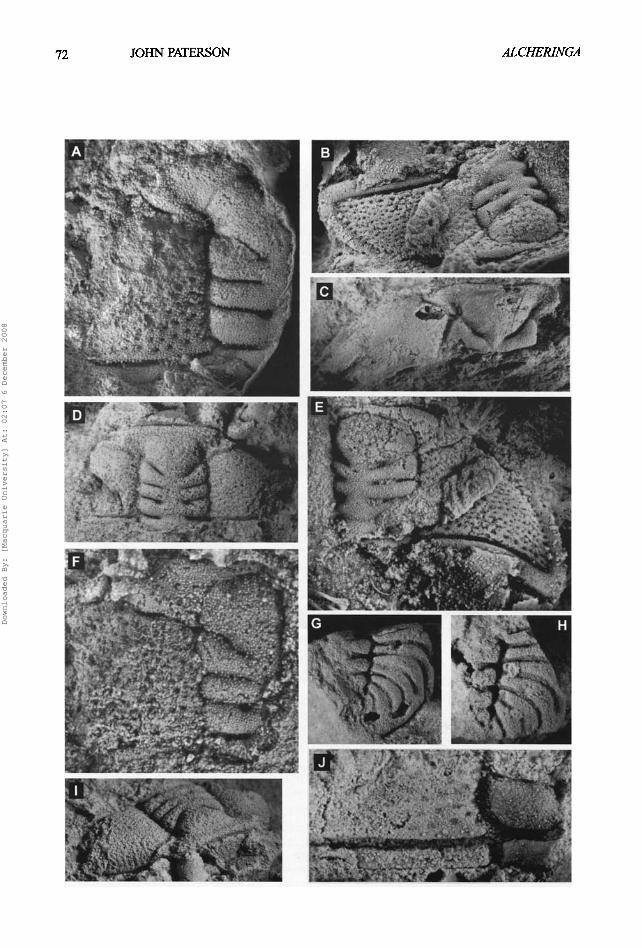
Fig. 4A-J. Prosopiscus lauriei sp. nov. A-B, E-F are latex casts of external moulds and C-D, G-J are internal moulds. A, dorsal view of paratyp¢ partial cranidium, AM F120750, x7; B, E, anterolateml and dorsal views of holotype cranidium, AM F120751, x8; C, dorsal view of paratyp¢ partial cranidium, AM F120752, x4; D, l, dorsal and anterolateral views of paratype cranidium, AM F120753, x7, F, dorsal view of paratype partial cranidium, AM F120754, x12.5; G, dorsal view of paratypo partial pygidium, AM F120755, x5.5; H, dorsal view of paratype partial pygidium, AM F120756, x5.5; J, dorsal view of posterior portion of paratyp¢ cranidium, AM F120757, x9. [see page 72, opposite].
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

74 JOHN PATERSON ALCHER1NGA
Order PHACOPIDA Salter, 1864 Suborder PHACOPINA Struve, 1959 Family PROSOPISCIDAE Fortey & Shergold, 1984
Discussion. The higher-level classification of the Prosopiscidae has been discussed in detail by Edgecombe et al. (1999, p. 1149).
Prosopiscus Salter, 1865
Type species. Prosopiscus mimus Salter, 1865, by original designation.
Prosopiscus lauriei sp. nov. (Figs 4A-J)
Etymology. For Dr John R. Laurie (Geoscience Australia, Canberra), for his work on Early Ordovician trilobites of Australia.
Diagnosis. Prosopiscus with a parallel-sided glabella, subpentagonal frontal glabellar lobe, clearly defined eye ridges, and three pygidial axial rings. Genal pitting coarse and evenly spaced.
Type material. Holotype AM F 120751 (cranidium; Figs 411, E). Paratypes are: cranidia- AM F 120750, 120752-120754, 120757 (Figs 4A, C-D, F, I-J); and pygidia - AM F 120755-120756 (Figs 4G-H). All specimens are disarticulated moulds collected from light-coloured siltstones from the basal 10 m of the Tabita Formation, Mount Arrowsmith, at coordinates 30006'36 '' S, 141035'05 '' E, approximately 200 km north of Broken Hill in northwestern New South Wales (Fig. 1).
Description. Cranidium approximately semi- circular in outline, convex, 40% as long (sag.) as wide, with maximum width along posterior border. Glabella elongate, parallel-sided, about 70% as wide as long (sag.), with glabella length approximately 90% cranidial length (sag.); fronted lobe wide with subpentagonal outline. Axial furrow well developed laterally, weak around anterior of glabella; preglabellar furrow well developed. Occipital furrow well developed laterally, shallow over mid-part of axis. S 1 and $2 straight (inner end of S 1 deepened into an apodeme), parallel, directed slightly posteriorly from axial furrow; L2 slightly shorter (exsag.) than
L1.S3 gently sinuous, strongly directed posteriorly from axial furrow, diverging anteriorly from each other at about 100 °, intersecting the axial furrow at the same point as the eye ridge; L3 expanding abaxially. Eye ridges extend anterolaterally from axial furrow, widening abaxiaUy, defined by shallow anterior furrow and deep, sigmoidal posterior furrow. Area in front of eye ridge (including preglabellar field) continuous with rounded cranidial anterior border. Lateral border narrow, separated from genal field by a shallow border furrow; posterior border wider than lateral border, defined by a deep, wide, posterior border furrow; genal angle rounded. Surface of cranidium densely covered in coarse granules; pitting on genal field is coarse and evenly spaced.
Pygidium about three-quarters as long as wide, convex (tr.) with steeply downsloping lateral margins. Axis relatively short, with three (possibly four) rings. Pleural field contains three segments, each of which are separated by a long, wide interpleural furrow. Long, very wide pleural furrows divide each segment into anterior and posterior bands, both of which merge with the convex border. Border widens posteriorly.
Discussion. This species is similar to P praecox and Prosopiscus sp. A from the Nora Formation (Georgina Basin) in centralAus~alia (Fortey & Shergold 1984), and to P magicusfrom northwest China (Zhou et al. 1982). Prosopiscus praecox differs from P lauriei in having a semicircular frontal lobe on the glabella, narrow posterior border furrow, dense genal pitting, and seven axial rings on the pygidium. Prosopiscus sp. A differs in having a much broader lateral cephalic border, narrow posterior border furrow, and four pygidial axial rings. Prosopiscus magicus, known from a single cranidium, is nearly identical to P. lauriei, however, B mag/cus has an occipital furrow which is deep across the entire width of the glabeUa.
Other known species of Prosopiscus which possess a parallel-sided glabella include the Chinese species P. subparallelus Lu (in Lu & Chang 1974), P. latus Lu (in Lu & Chang 1974), and P. yinjiangensis Yin (in Yin & Li 1978), in addition to P. edgarensis Legg (1976) from the Middle Ordovician Goldwyer Formation
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

AL CHERINGA ORDOVICIAN TRILOBITEPROSOPISCUS 75
(Canning Basin) in Western Australia. Due to the lack of representative specimens and poor preservation of these species, only general comparisons can be made. Prosopiscus subparallelus is known from four cranidia (Lu & Chang 1974, Lu 1975, Zhou etal. 1984) and differs from P. lauriei in having a rounded frontal glabellar lobe, a wider lateral cephalic border, and smaller L3. Prosopiscus latus is known from a single complete specimen (Lu & Chang 1974), and differs in having a rounded frontal glabellar lobe and a uniformly deep occipital furrow. Prosopiscus yinjiangensis was described from a single, very poorly preserved cranidium (Yin & Li 1978), which make comparisons difficult. Prosopiscus edgarensis is known from two small and flattened cranidia (Legg 1976); the major differences (not caused by poor preservation) are that P. edgarensis has a rounded frontal glabellar lobe and dense genal pitting.
Acknowledgments I am indebted to John Laurie for invaluable discussions and advice during various stages of this research. Barry Webby kindly donated specimens ofP. lauriei for study. Yongyi Zhen provided information on conodonts from the Tabita Formation, and made available a preprint on the Early Ordovician conodonts from western New South Wales. I thank: Zhou Zhiyi for providing additional information on occurrences of Prosopiscus from China; Glenn Brock for reviewing an early draft of the manuscript and providing constructive comments; Dean Oliver who drafted Figs 1 and 2; and Richard Fortey and Greg Edgecombe who provided helpful reviews. Funding for this research was provided by the Betty Mayne Scientific Research Fund of the Linnean Society of N.S.W. This paper is a contribution to IGCP410: 'The Great Ordovician Biodiversification Event'.
Refe r ences
BARNES, C.R., FORTEY, R.A. & WmLL~MS, S.H., 1995. The pattern of global biG-events during the Ordovieian Period. In Global events and event stratigraphy in the Phanerozoic, O.H. WALUSER, ed., Springer, Berlin, 139-172.
BAUD, A., GAETANI, M., GARZANTI, E., FOIS, E., NICORA, A. & Tl~rroRI, A., 1984. Geological observations in south- eastern Zanskar and adjacent Lahul area (northwest- ern Himalaya). Eclogae Geologicae Helvetiae 77, 171-197.
BURRErr, C.F. & STArt, B., 1987. China and southeast Asia as part of the Tethyan margin of Cambro-Ordovician Gondwanaland. In Shallow Tethys 2, K.G McKENzm, ed., Balkema, Rotterdam, 65-77.
CHAtteRTON, B.D.E. & Sem¢~, S.E., 1989. Larval ecology, life history strategies, and patterns of extinction and survivorship among Ordovician trilobites. Paleobiology 15, 118-132.
CHAa'rERrON, B.D.E. & SPEVER, S.E., 1990. Applications of the study of trilobite ontogeny. In Short Courses in Paleontology, No. 3: Arthropod Paleobiology, Paleontological Society, Knoxville, 116-136.
CHATTERTON, B.D.E. & SPEYER, S,E., 1997. Ontogeny. In Treatise on Invertebrate Paleontology, Part O, Trilobita, Revised, R.L. KAESLER, ed., Geological Society of America and University of Kansas Press, Lawrence, 173-247.
CHEN, X., RO~G, J.-Y., MITCHELL, C.E., HARPER, D.A.T., FAN, J.-X., ZHAH, R.-B., ZHANG, Y.-D., LI, R.-Y. & WANG, Y., 2000. Late Ordovician to earliest Silurian graptolite and brachiopod biozonation from the Yangtze region, South China, with a global correlation. Geological Magazine 137, 623-650.
CHEN, X., RONG, J.-Y., WANG, X.-F., WANG, Z.-H., ZHANG, Y.-D. & ZHAN, R.-B., 1995. Correlation of the Ordovieian Rocks of China. International Union of Geological Sciences, Publication 31, 104 p.
CHEN, X., ZHANG, Y.-D. & MITCHELL, C.E., 2001a. Early Darriwilian graptolites from central and western China. Alcheringa 25, 191-210.
Ca~q, X., Zaou, Z.-Z., R~t;, J.-Y. & Ll, J., 200lb. Ordovician series and stages in Chinese stratigraphy: steps toward a global usage. Alcheringa 25, 131-141.
Coczs, L.R.M. & FORTEV, R.A., 1990. Biogeography of Ordovieian and Silurian faunas. In Palaeozoic Palaeogeography and Biogeography, W.S. McKEImow & C.R. SCOTESE, eds, Geological Society of London Memoir 12, 97-104.
COOPER, B.J., 1981. Early Ordovician conodonts from the Horn Valley Siltstone, central Australia. Palaeontology 24, 147-183.
EOGECOMBE, GD., CHAttERTON, B.D.E., WAISFr.LD, B.G. & VACCARI, N.E., 1999. Ordovician pliomerid and prosopiscid trilobites from Argentina. Journal of Paleontology 73, 1144-1154.
FORTEY, R.A. & OWENS, R.M., 1990. Evolutionary radiations in the Trilobita. In Major Evolutionary Radiations, P.D. TAYLOR & G,P. LARWOOD, eds, Systematics Association Special Volume 42, Clarendon Press, Oxford, 139-164.
FORTEY, R.A. & SHERGOLD, J.H., 1984. Early Ordovician trilobites, Nora Formation, central Australia. P alaeontology 27, 315-366.
GAETANI, M., CASNEDI, R., Fols, E., Gm, ZAN'n, E., JAOOUL, F., NICORA, A: & Trs-rova, A., 1986. Stratigraphy of the Tethys Himalaya in Zanskar, Ladakh. Rivista Italiana di Paleontologia e Stratigrafia 91, 443-478.
HAVOEN, H.H., 1904. The geology of Spiti with parts of Bashahr and Rupshu. Memoirs of the Geological Survey of India 36, 129 p.
I2GG, D.P., 1976. Ordovician trilobites and graptolites
Downloaded By: [Macquarie University] At: 02:07 6 December 2008

76 J O H N PATERSON ALCHERINGA
from the Canning Basin, Western Australia. Geologica et Palaeontologica 10, 58 p.
l_~c~, D.P., 1978. Ordovieian bioslratigraphy of the Canning Basin, Western Australia. Alcheringa 2, 321-334.
Lu, Y.-H., 1975. Ordovician trilobite faunas of central and southwestern China. Palaeontologica Sinica 152, 463 p.
Lu, Y.-H. & CHANG, W.-T., 1974. Ordovieian trilobites. In A handbook of stratigraphy and palaeontology in southwest China, Science Press, Beijing, 124-136.
Mortals, S.E & Fomzv, R.A., 1985. Catalogue of the type and figured specimens of Trilobita in the British Museum (Natural History). British Museum (Natural History), London, 183 p.
NICOLL, R.S., L A t ~ , J.R. & Rocrm, M.T., 1993. Revised stratigraphy of the Ordovieian (Late Tremadoc- Arenig) Prices Creek Group and Devonian Poulton Formation, Lermard Shelf, Canning Basin, Western Australia. AGSO Journal of Australian Geology & Geophysics 14, 65-76.
NICOLL, R.S., OWEN, M., SHERC, OLD, J.H., LAU~E, J.R. & GORTER, J.D., 1988. Ordovician event stratigraphy and the development of a Larapintine seaway, central Australia. Bureau of Mineral Resources, Geology and Geophysics, Record 1988/42, 72-77.
NIELSEN, A.T., 1992. Intercontinental correlation of the Arenigian (Early Ordovieian) based on sequence and ecostratigraphy. In Global Perspectives on Ordovician Geology, B.D. WEaaY & J.R. LAt~IE, edS, Balkema, Rotterdam, 367-379.
PATERSOr~, J.R., 2002. Early Ordovieian trilobites from Mount Arrowsmith, northwestern N.S.W., Australia: Biostratigraphic and biogeographic implications. Geological Society of Australia, Abstracts 68, 128.
REED, ER.C., 1912. Ordovician and Silurian fossils from the central Himalayas. Memoirs of the Geological Survey of lndia, Palaeontologica Indica 7, 168 p.
SALTER, J.W., 1865. In J.W. SALTER 8~ H.G BLANDFORD, Palaeontology of Niti in the northern Himalaya. Military Orphan Press, Calcutta. ! 12 p.
SHERGOLD, J.H., 1971. Resum6 of data on the base of the Ordovician in northern and central Australia. M$moire de la Bureau de Recherches G$olo~que et Mineralogique 73, 391-402.
SHEROOLO, J.H., 1991. Late Protetozoie and Early Palaeozoic palaecntology and biostrafigraphy of the Amadem Bask Bureau of Mineral Resources, Geology & Geophysics (Australia), Bulletin 236, 97-1 ! 1.
Sn~OOLD, J.H., COOPER, R.A., DRUCE, E.C. & WEaaY, B.D., 1982. Synopsis of selected sections at the Cambrian- Ordovician boundary in Australia, New Zealand, and Antarctica. In The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations, M.G. BASSETT & W.T. DEAN, eds, National Museum of Wales, Geological Series No. 3, Cardiff, 211-227.
SPEVErt, S.E. & CrIATTEa~TON, B.D.E., 1989. Trilobite larvae and larval ecology. Historical Biology 3, 27-60.
SPErm, S.E. & CHArmRTON, B.D.E., 1990. Trilobite larvae, larval ecology and developmental paleobiology. In Short Courses in Paleontology, No. 3: Arthropod Paleobiology, Paleontological Society, Knoxville, 137-156.
STAIT, K. & DRUCE, E.C., 1993. Conodonts from the Coolibah Formation, Georgina Basin, central Australia. AGSO Journal of Australian Geology & Geophysics 13, 293-322.
VANDENBERG, A.H.M. & COOPER, R.A., 1992. The Ordovician graptolite sequence of Australasia.
Alcheringa 16, 33-85. WATSON, S.T., 1988. Ordovician conodonts from the
Canning Basin (W. Australia). Palaeontographica 203A, 91-147.
WEBBV, B.D., 1971. The trilobite Ptiomerina Chugaeva from the Ordovician of New South Wales. Palaeontology 14, 612-622.
WEBaY, B.D., 1978. History of the Ordovician continental platform shelf margin of Australia. Journal of the Geological Society of Australia 25, 41-63.
WEBBY, B.D., 1983. Lower Ordovician arthropod trace fossils from western New South Wales. Proceedings of the Linnean Society of New South Wales 107, 59-74.
WEaBV, B.D., PERCIVAL, I.G, EOGECOMBE, GD., COOPER, R.A., VANDENBERG, A.H.M., PICKETT, J.W., POJETA JR, J., PLAYFORD, GL, WINCHESTER-SEETO, T., YOUNG, G.C., ZHEN, Y.Y., NICOLL, R.S., Ross, J.R.E & SCHALLREUTER, R., 2000. Ordovician palaeobiogeography of Australasia. Memoir of the Association of Australasian Palaeontologists 23, 63-126.
WEaav, B.D., VANDENBERG, A.H.M., COOPER, R.A., BANKS, M.R., BURRETr, C.E, HENDERSON, R.A., CLARKSON, ED., HUGHES, C.P., LAURm, J.R., S rAIT, B.A., THOMSON, M.R.A. & WEBERS, G.F., 1981. The Ordovician System in Australia, New Zealand and Antarctica. International Union of Geological Sciences, Publication 6, 64 p.
WINCHESTER-SEETO, T., FOSTER, C.B. &: O'LEARY, T., 2000. Chitinozoans from the Middle Ordovician (Darriwilian) Goldwyer and Nita Formations, Canning Basin (Western Australia). Acta Palaeontologica Polonica 45, 271-300.
YIs, G.-Z. & LI, S.-J., 1978. Trilobita. In Handbook of Palaeontology of southwest China, Guizhou, Part 1, Geological Press, Beijing, 385-594.
YOUNG, G.C. &: LAURIE, J.R. (eds), 1996. An Australian Phanerozoic Timescale. Oxford University Press, Melbourne, 279 p.
tHEN, Y.-Y., NICOLL, R.S., PERCIVAL, I.G., HAMEDI, M.A. & STEWART, I., 2001. Ordovician rhipidognathid conodonts from Australia and Iran. Journal of Paleontology 75, 186-207.
ZHEN, Y.-Y., PERCIVAL, I.G. & WE[laY, B.D., 2003. Early Ordovician conodonts from far western New South Wales, Australia. Records of the Australian Museum 55, 169-220.
ZHOU, Z.-Q., LI, J.-S. & Qu, X.-G., 1982. Trilobita. In Paleontological Atlas of northwest China, Shaanxi- Gansu-Ningxia, Volume 1, Precambrian and Early Paleozoic, Geological Publishing House, Beijing, 215-294.
Znou, Z.-Y. & DEAN, W.T., 1989. Trilobite evidence for Go analand in cast Affaa during the Ordovici~a. Journal of Southeast Asian Earth Sciences 3, 131-140.
Znou, Z.-Y. & DEAN, W.T. (eds), 1996. Phanerozoic geology of Northwest China. Science Press, Beijing, 316 p.
Znou, Z.-Y., DEAN, W.T. & LUO, H.-L., 1998a. Early Ordovician trilobites from Dali, West Yunnan, China, and their palaeogeograpbical significance. Palaeontology 41, 429-460.
Z8oo, Z.-Y., DEAN, W.T., YUAN, W.-W. & ZHOU, T.-R., 1998b. Ordovician trilobites from the Dawangou Formation, Kalpin, Xinjiang, north-west China. Palaeontology 41, 693-735.
Znou, Z.-Y., YIN, G-Z. & TRIPP, R.P., 1984. Trilobites from the Ordovician Sbihtzupu Formation, Zunyi, Guizhou Province, China. Transactions of the Royal Society of Edinburgh: Earth Sciences 75, 13-36.
Downloaded By: [Macquarie University] At: 02:07 6 December 2008