morphologie et évolution ontogénétique de trimerocephalus lelievrei nov. sp., premier trilobite...
TRANSCRIPT

l MORPHOLOGIE ET I VOLUTION
ONTOGI NI TIQUE DE TRIMEROCEPHA LUS LELIEVREI NOV. SP.,
PREMIER TRILOBITE PHACOPID] AVEUGLE DE FAMENNIEN NORD-AFRICAIN
CATHERINE CRONIER & RAIMUND FEIST
CRONIER C. & FEIST R. 1997. Morphologie et ~volution ontog~ngtique de Trimerocephalus lelievrei nov. sp., pre- mier trilobite phacopidg aveugle du Famennien nord-africain. [Morphology and ontogeny of Trimerocephalus lelie- vrei nov. sp., first blind phacopid trilobite from the Famennian of North Africa]. GEOBIOS, M.S. n ° 20: 161-170.
Rt~SUM]~ - Les exceptionnelles richesse et conservation des exuvies silicifi~es de nombreux stades tardi-larvaires et juveniles, permettent d'filucider de fa~on trgs dgtaillge la morphologie et la morphog~ngse du genre Trimerocephalus, phacopidg aveugle caractgrisant le Famennien inf~rieur. Le matSriel appartient/~ la nouvelle esp~- ce lelievrei, premier repr~sentant du genre en Afrique. Ses relations avec les esp~ces proches de l'Europe sont ~ta- blies. Les changements ontoggngtiques rappellent ceux de Phacops confirmant ainsi une relation phyldg~n~tique ~troite entre ces deux genres.
MOTS-CLt~S: TRILOBITA, PHACOPIDAE, FAMENNIEN, AFRIQUE DU NORD, MORPHOLOGIE, ONTOGt~NIE.
ABSTRACT - Exceptionally rich and well preserved silicified exuvia comprising numerous late larval and early juvenile stages allow retracing detailed morphological and morphogenetical features of Trimerocephalus, a blind early Famennian phacopid. The materiel belongs to the new species lelievrei which is the first described represen- tative of the genus in Africa. Its affinities with related species from Europe are established. Ontogenetic changes are similar to those of Phacops emphasizing close phyletic relationships between both genera.
KEYWORDS: TRILOBITA, PHACOPIDAE, FAMENNIAN, NORTH AFRICA, MORPHOLOGY, ONTOGENY.
INTRODUCTION Le d~veloppement ontog~n~tique des phacopidSs est encore mal connu. Notamment , nous ne connaissons de la p~riode larvaire que quelques pisces isol~es du stade meraspis appar tenant au genre Phacops (Jahnke 1969; Chat ter ton 1971; Alberti 1972). La seule investigation sur l'~volu- tion tardi- larvaire et juvenile d'un Trimeroce- phalus McCoY, 1949 a ~tg entreprise par Osmdls- ka en 1963. Son matgriel trilobitique comprend de nombreux moules in te rnes et ex ternes com- presses de carapaces entigres, preserves dans des mudstones d~ealcifi~s. Les donn6es d'Osmdlska peuvent ~tre v~rifi~es et compl6t~es apr~s la rgcente dgcouverte d'un assemblage d'exuvies de diffgrentes tailles, exeep t ionne l lement bien conserv6es en 3-D par silicification et ne pr~sen- rant aucune d6formation. Ce mat6riel tr~s abon- dant appar t ient ~ une nouvelle esp~ce: Trime- rocephalus lelievrei nov. sp., le premier repr~sen- rant du genre en Afrique. I1 a 6t5 d~couvert par H. Leli~vre (Mus6um national d'Histoire Naturelle, Paris) lors de l 'a t taque acide d'un nodule marno-
calcaire pour en d6gager des restes osseux d'un grand arthrodire Dunkleosteus marsaisi LEHMAN, 1956 (d6termina t ion Leli~vre). Ce nodule arthrodires et trilobites a ~t6 pr~lev~ par Leli~vre sur la cr~te de Khorb-el-Atil au SE d'Erfoud (Tafilalet, SE marocain) ~ une v ingta ine de kilom~tres ~ I'ESE de Dar Kaoua. Les conodontes Palmatolepis marginifera trouv~s darts ce mSme nodule lui conf'erent un age Famennien inf~rieur ~lev~ (Zone inf6rieure ~ marginifera, to III).
ETUDE SYSTt MATIQUE T e r m i n o l o g i e - La terminologie morphologique des trilobites phacopid6s adoptSe est bas~e essen- t ie l lement sur les t r avaux d'Hup~ 1953; d'Harrington et al. in Treatise 1959; de Chlupfi5 1977 et de Clarkson 1979. Par ailleurs, quelques abr~viations et symboles ont ~t~ utilis~s: sag = sagittal, t r = transversal, L = longueur, 1 = largeur maximale, b = largeur minimale, C = c~phalon, G = glabelle, R = rachis, P = pygidium.

Ordre PHACOPIDA Hawle & Corda, 1847 Sous ordre PHACOPINA Struve, 1959
Super famille PHACOPACEA Hawle & Corda, 1847 Famil le PHACOPIDAE Hawle & Corda, 1847
Sous famil le PHACOPINAE Hawle & Corda, 1847
Genre Tr imeroeepha lus McCoY, 1849
Trimerocephalus lelievrei nov. sp. P1.1, fig. 1-5
Holotypus - C6phalon UM2-CCt26; P1. 1, fig. la, lb.
Locus typicus - Khorb-el-Atil au SE d'Erfoud, Tafilalet, Maroc.
Stratum typicum - Marno-calcaire ~ miches ~ arthrodires, Famennien to III, Zone inf6rieure A marginifera.
Derivatio nomin i s - D'apr6s Herv6 Leli6vre (Paris), sp6cia- Iiste de vert6br6s du D6vonien sup6rieur.
D i a g n o s e - Esp6ce du genre Trimerocephalus Mc CoY avec les t r a i t s carac t6r is t iques suivants: gla- belle et joues peu bomb6es. Glabelle surbaiss6e, subpen tagona le et large, ~ contour f ronta l large- m e n t parabol ique. Lobe f ronta l su rp lomban t pe ine le sillon frontal . $2 et $3 distincts. S u tu r e faciale en ta i l l an t la pen te du sillon ant6ro- la t6ral sans r e m o n t e r sur la joue. Marge pos t6r ieure du pyg id ium rect i l igne avec une 16g6re inf lexion m6diane. Pi6ce t e rmina l e du rachis surbaiss6e et d61imit6e du l imbe. T u b e r c u l a t i o n dense sur c6phalon, g ranu la t ion fine sur pygidium.
M a t 6 r i e l - Le mat6r ie l d '6tude d6calcifi6 ~ l 'acide formique (10%) a pe rmis l 'obtent ion de nom- b r e u s e s exuv ies sil icifi6es e t d6sa r t i cu l6es in tac tes soit: I p ro topygid ium merasp is M9, 6 pro-
162
topygidia merasp i s M10, 206 pygidia holaspis, 64 cranidia et c6phalons, 9 doublures , 12 hypos tomes et de n o m b reu x segments thoraciques . L 'ensemble du mat6r ie l est d6pos6 dans les collections du Labora to i re de Pal~ontologie de Montpe l l i e r (col- lect ion UM2-CC).
D e s c r i p t i o n - * C6phalon - Rappor t L/1 a u t o u r de 0,54, une m o y en n e p a rm i les g rands sp6cimens ( reconst i tu t ion Fig. 1). Vue dorsale - La glabelle est di lat6e en avant . Le r appor t L/1 est au tou r de 0,76. Le contour f ron ta l est l a rgemen t parabol ique. La glabelle es t d61i- mit6e pa r des sillons dorsaux larges et t r6s pro- fonds. Les sillons dorsaux, concaves 16g6rement vers l 'ext6rieur, d ive rgen t d 'envi ron 55 ° d a n s leur por t ion pos t6r ieure et de 80" dans leur por t ion ant6r ieure , Le lobe frontal , non s6par6 des lobes glabellaires L2 et L3, couvre le bord frontal ; il p r& sente darts sa pa r t i e an t6ro-cen t ra le u n tr~s 16ger b o m b e m e n t chez cer ta ins sp6cimens. Le sillon glabel la i re $3 est compos6: une b ranche ant6r ieu- re peu marqu6e, r e l a t i v e m e n t longue, subparal l6- le au sillon dorsal; une b ranche pos t6 r ieure tr6s courte, peu marqu6e, parall61e au sillon occipital; elle n ' a t t e in t pas le sillon dorsal . Le sillon glabel- laire $2 est tr6s court , peu marque , parall61e au sillon occipital, dispos6 ob l iquement vers l 'avant; il ne jo in t pas le sillon dorsal. La por t ion m6diane du sillon glabel laire S1 (sillon pr6occipital) est courb6e mod6r6men t en avant , peu marqu6e; tes port ions distales sont profondes, dispos6es 16g6re- m en t ob l iquement vers l 'avant . L ' anneau pr6occi- pi tal es t tr~s 16g~rement bomb6. Les lobes la t& r au x pr6occipi taux sont ~ pe ine percept ibles . Le sillon occipital est plus n e t t e m e n t diff6renci6,
PLANCHE 1
Fig. 1-16 - Trimerocephalus lelievrei nov. sp., Khorb-el-Atil, Tafilalet, Maroc, Famennien, D6vonien sup6rieur. Photographies au MEB d'exuvies silicifi6es. Les sp6cimens sont d6pos6s dans la collection UM2-CC du Laboratoire de Pal6ontologie de Montpel'lier (France). la, holotype, vue frontale du c6phalon: UM2-CCt26, x 7; lb, holotype, vue dorsale du c6phalon: UM2-CCt26, × 7; 2a, vue ventrale de c6phalon: UM2-CCt11, ×7; 2b, vue lat& rale de c6phalon: UM2-CCt11, x 9; 3, rue dorsale de doublure c6phalique: UM2-CCt2, x13; 4, rue ventrale de dou- blure c6phalique: UM2-CCt46, x 8; 5a, vue ventrale de cranidium: UM2-CCt19, x 8; 5b, vue frontale de cranidium: UM2-CCt19, x 10; 5c, vue lat6rale de cranidium: UM2-CCt19, x 10; 6, vue dorsale de cranidium: UM2-CCt48, x 16; 7, vue dorsale d'hypostome: UM2-CCt8, x 30; 8, vue lat6rale de pygidium: UM2-CCt51, x 20; 9, rue d'un seg- ment thoracique: UM2-CCt4, x 8; 10, vue dorsale de cranidium larvaire: UM2-CCt39, x 40; 11, vue ventrale de la bordure protopygidiale M10 et des deux tubercules: UM2-CCt47, x 120; 12, vue frontale de pygidium: UM2- CCt52, × 8; 13, vue dorsale de pygidium: UM2-CCt55, x 12; 14, vue ventrale de pygidium: UM2-CCtl, x 20; 15a, vue dorsale de protopygidium M9: UM2-CCt7, x 40; 15b, vue ventrale de protopygidium M9: UM2-CCt7, x 40; 16, rue ant6ro-lat6rale du c6phalon et de la suture faciale: UM2-CCt32, x 22. la, holotype, frontal view ofcepha- !on; lb, holotype, dorsal view of cephalon; 2a, ventral view of cephalon; 2b, lateral view of cephalon; 3, dorsal view of cephalic doublure; 4, ventral view of cephalic doublure; 5a, ventral view of cranidium; 5b, frontal view of crani- dium; 5c, lateral view of cranidium; 6, dorsal view of cranidium; 7, dorsal view of hypostome; 8, lateral view of pygidium; 9, view of one thoracic segment; 10, dorsal view of larval cranidium; 11, ventral view of protopygidial border M10 and its two tubercules; 12, frontal view of pygidium; 13, dorsal view of pygidium; 14, ventral view of pygidiam; 15a, dorsal view of protopygidium M9; 15b, ventral view of protopygidium M9; 16, antero-lateral view of cephalon and facial suture view.

G e o b i o s P1. 1 M.S . n ° 2(} C. C r S n i e r & R . F e i s t

164
fi' i l a ,o l c
.... ,. :~y:. ' ,~F. :~i& ~, / ~ ' , ~:,~ ~:~,~'~ ,~ ' ~ ~$:~ :b ,~ ,~ ' ~. '~t~!~ ' i~!~L~ :
2b ld
FIGURE 1 - Trimerocephalus lelievrei nov. sp. Khorb-el-Atil, Tafilalet, Maroc, Famennien, D6vonien sup6neur. Dessins "au points" d'exuvies silicifi6es, r6alis6s ~ la chambre claire. Reconstitution d'un c~phalon d'holaspis, UM2-CCt29: l a , r u e dorsale; l b , vue lat6rale; lc, vue ventrale; ld , r ue frontale. Reconstitution d'un pygidium d'ho]aspis, UM2-CCt54: 2a, vue dorsale; 2b, vue ventra- le. Echelle: t mm. Upper Devonian. Pictures of silicified moults. Reconstruction of a holaspid cephalon: la, dorsal view; lb, lateral view; lc, ventral view; ld, frontal view. Reconstruction of a holaspid pygidium: 2a, dorsal view; 2b, ventral view.
courb6 mod6r6ment en avant dans sa portion m6diane; les port ions distales 16g6rement obliques vers l 'avant sont tr6s profondes. L'anneau occipital est deux fois plus large (sag) que l 'anneau pr6occipital. La portion m6diane est fortement enfl6e. Uenflure diminue dans leE por- tions distales. Les lobes lat6raux occipitaux ne sont pas diff6renci6s. Les d6pressions musculaires sont tr6s profondes. Aucune protub6rance oculai- re ni champ visuel ne sont d6velopp6s. La suture faciale submarginale d6tectable ~ partir de la mi- longueur du bourrelet lat6ral, traverse le bourre- let lat6ral obliquement puis parcourt le sillon lat6ral. Elle remonte ~ l'int6rieur de ce sillon dans sa partie la plus ant6rieure, vers les joues mais sans les entailler. Les joues sont uniform6ment peu bomb6es. Le sillon frontal est bien marqu6 mais ne t tement moins profond que lee sillons la t6raux et post6rieurs, non in ter rompus au niveau de l'angle g6nal, seulement un peu r6duits en profondeur. Les bourrelets lat6raux et post6- rieurs, peu enfl6s, au niveau de leur jonction, s'6- largissent. Les angles g6nanx sont arrondis. Vue latgrale - Le contour frontal de la glabelle est mod6r6ment arqu6. I1 est suivi d'un contour dor- sal 16g~rement convexe. Le contour de l 'anneau pr6occipital est droit. Celui de l 'anneau occipital est for tement convexe. L'anneau occipital est aussi hau t que la glabelle ~ son maximum de convexit6. Le bord frontal est fortement inclin6 vers l'int6riem: La bande ant6rieure de la doublu- re c6phalique, longue, est inclin6e vers l'arri6re et l'int6rieur. La bande post6rieure, quant ~ elle, est courte, dispos6e en retrai t et inclin~e vers l'arri6- re et l'ext6rieur. Par cons6quent, le trac6 du sillon
de fermeture est large et profond. Vue frontale - La glabelle a un contour semi-circu- laire. Le sillon frontal pr6sente une courbure plus accentu6e que celle du sillon de fermeture danE sa portion m~diane. Le bord frontal pr6sente des por- tions distales l~g6rement enfl6es situ6es dane un plan horizontal. Progressivement, dans le plan vertical, ce bord, dans sa portion m6diane, devient plan od il atteint sa largeur (sag) maximale. Vue ventrale - Transversalement, le sillon de fer- meture est large, profond, constant, simple, d61i- mit6 par des crates aigu~s. I1 pr6sente une cour- bure vers l'arri6re, de par t et d 'autre de sa portion transversale, dont les angles ant6ro-lat6raux sont d'environ 130 °. Le sillon s'61argissant lat6rale- ment progressivement vers l'arri6re comporte des fossettes de fermeture contigu~s. Sa profondeur est diminu6e. La doublure est 6troite lat6rale- ment, s'61argit dans la portion m6diane: la bande ant6rieure est longue, rejoignant le milieu du bourrelet post6rieur (face vent ra l@ la bande post6rieure est plus courte, elle n 'at teint pas l'angle g6nal. Le rapport entre la largeur maxi- male de la bande post6rieure et la largeur maxi- male du sillon de fermeture est autour de 0,48.
* Hypostome - Triangulaire ~ ~troitement trap6- zoidal. Le corps central est 16g6rement enfl6. La marge ant6rieure est droite. Ant~rieurement, de chaque c6t6 du corps central existe une 16g~re d6pression. Post6rieurement, ce corps central est d61imit6 par un sillon post~rieur superficiel, qui s'61argit dans sa portion m6diane. LeE ailes ant6- rieures sont d6velopp6es, orient6es vers l'int6- rieur. Les aries post6rieures sont r6duites ~ de

165
petites pointes. Les maculae sont absentes. Le bourrelet post@ieur pr6sente une marge post~- rieure denticul6e (5 dents).
* Thorax - Repr6sent6 sous forme de segments thoraciques isol6s. Les sp@imens complets du genre Trimeroce- phalus connus mais absents dans notre mat@iel montrent que la morphologie des segments varie graduel lement le long de l'axe thoracique et qu'une diff@enciation existe entre les segments ant~rieurs et post@ieurs: sur l 'extr6mit6 distale de la doublure pleurale, existe un tubercule pro6- minent qui sur les segments ant@ieurs est 16ger et, sur les segments post@ieurs est distinct. Aucun des 61~ments consti tuant le thorax n'est fixe, il n'est donc pas possible d'identifier la posi- tion d'origine des segments d6sarticul6s. Vue dorsale - L'anneau thoracique est fortement enfi6 dans sa portion m6diane. Distalement, l'en- flure dispara~t. Les lobes lat6raux ne sont pas diff~renci6s. I ' anneau axial est s6par6 du demi- anneau articulaire par des sillons distaux diver- gents d'environ 45 ° . Les pl~vres thoraciques g~ni- cul6es darts leur portion m6diane sont inclin6es vers l'arri~re, formant un angle de 130 °. Les sillons pleuraux sont bien distincts, profonds, jusque dans la part ie m6diane o/~ ils disparais- sent. La bande ant@ieure est deux fois plus 6troi- te (exsag) que la bande post@ieure (exsag). Les pl~vres sont arrondies aux extr6mit6s distales. La facette articulaire large (exsag) se d6veloppe dbs la portion distale de la pl~vre. Vue ventrale - Les d6pressions musculaires sont profondes. Sur l'extr6mit~ distale de la doublure pleurale, localis~e dans la portion distale des pl~vres, existe un tubercule qui, selon les anneaux sensiblement de mgme taille, est plus ou moins distinct. PrOs de la marge ant@ieure de la dou- blure pleurale, existent des organes de Pander en forme de petites protub6rances allong6es situ6es ant@ieurement sur la doublure. La doublure est peine plus large (tr) que la pl~vre (exsag).
* Pygidium - Transverse et lenticulaire fi trap6zo~- dal. Rapport IN autour de 0,33, une moyenne parmi les grands sp6cimens (reconstitution Fig. 1). Vue dorsale - La marge ant@ieure est droite (tr). Les sillons dorsaux sont bien distincts: larges plus ou moins profonds. Le rachis en relief pr6sente une enflure diminuant progressivement de l'avant vers l'arri~re. Le rachis est court (sag) et large (tr). Son extr~mit6 post@ieure est arrondie. I1 poss~de 4 anneaux axiaux bien diff@enci6s, bien d61imit~s par des sillons rachidiens larges, surtout au niveau des sillons dorsaux, mod@6ment profonds. La piece terminale est ddimit6e par des sillons peu mar- qu6s. Les sillons pleuraux et interpleuraux nette- merit distincts fi ravant, deviennent un peu moins distincts darts la partie post@ieure du pygidium.
lls d61imitent des demi-c6tes distinctes, mod@6- ment enfl6es. Ces sillons n'atteignent pas la marge post@ieure, ils d61imitent un limbe s'61argissant lat~ralement. Le limbe est denx fois plus large (sag) que la piece terminale du rachis. Les pl~vres soud6es sont g6nicu16es, inclin6es vers le bas. La marge post@ieure enti~re, sans pointe caudale ni ~pine pleurale, pr6sente une 16g~re inflexion vers ravant dans sa portion m6diane. Les angles des bords lat@aux et post@ieurs sont autour de 80 °. Vue ventrale - La doublure pygidiale ~troite dans la portion m6diane s'~largit p@iph@iquement. Sur cette doublure, la bordure post@ieure forme un butoir continu. Ce butoir, au niveau des extr6- mit6s distales, se prolonge par un tubercule. Ces deux tubercules distaux, pr6sents sur la bordure pygidiale, correspondent aux deux premieres fos- settes de la doublure c~phalique, permet tan t la coaptation du trilobite lors de renroulement .
* Sculpture - Uensemble du c@halon pr6sente deux sortes de tubercules: des petits dispers6s parmi des larges coniques. Ils sont localis6s sur la glabelle, les joues et m~me sur les anneaux occipi- tal et pr6occipital (deux tubercu les larges coniques sur l 'anneau pr6occipital, un tubercule large conique au centre de l 'anneau occipital). Les angles des joues pr6sentent un tr~s ]6ger tubercu- le, r~miniscence d'une 6pine g6nale larvaire. La doublure est d6pourvue de tubercules. Uhyposto- me est lisse. Les plus petits tubercules coniques sont localis6s sur le pygidium.
* Variabilit~ - Des diff@ences individuel les concernent le contour ant@ieur de la glabelle: l 'arrondi diff'ere depuis des formes au contour lar- gement parabolique aux formes au contour un peu plus anguleux (le lobe frontal pr6sentant une protub@ance large et ]6g~re). La diff@ence des angles divergents des sillons dorsaux est environ de 5 °. Des diff@ences apparaissent 6galement au niveau de la profondeur de $2 et $3 qui est plus ou moins nette. La course, la distance entre les branches ant@ieure et post@ieure de $3 varient. Le trac6 du sillon de fermeture et celui du sillon frontal ne montrent pas de variabilit6. Ils parais- sent constituer ainsi des caract~res stables. I1 en est de m~me pour le trac6 de la suture faciale. Les pygidiums ne montrent pas de variabilit6 intra- sp6cifique. Au niveau de la sculpture, aucune variation intrasp@ifique n'est ob.serv6e.
C o m p a r a i s o n s - La nouvelle esp~ce se distingue de la plupart des autres Trimerocephalus par le trac~ de la suture faciale qui n'entaille pas les joues, trac6 qui est interm~diaire entre celui caract@istique du genre Trimerocephalus et celui du genre Dianops.
Cependant, cette nouvelle esp~ce pr6sente des similitudes avec Trimerocephalus dianopsoides

FIGURE 2 - Dimensions (en
166
mm) de quelques (a) era- (a) C LC 1G LG bG LC/1C LG/1G 1G/]C bG/1G nidia-c6phalons et (b) protopygidia-pygidia de UM2-CCt39 1,55 0,98 0,76 0,84 0,28 0,63 1,10 0,49 0,36 Trimerocephalus lelievrei UM2-CCt6 2,17 1,33 1,12 1,07 0,40 0,61 0,96 0,52 0,36 nov. sp. du Famennien UM2-CCt38 2,69 1,74 1,66 1,48 0,71 0,64 0,89 0,62 0,43 d'Afrique du Nor& (L = longueur, 1 = largeur UM2-CCt23 3,88 2,38 2,44 2,00 1,06 0,61 0,84 0,63 0,43 maximale, b = largeur UM2-CCt19 5,63 3,18 3,13 2,68 1,44 0,56 0,84 0,56 0,46 minimale, C = cranidium- UM2-CCtll 8,00 4,19 4,38 3,38 2,25 0,52 0,77 0,55 0,51 c6phalon, P = protopygi- UM2-CCt29 10,5 5,69 6,13 4,63 2,80 0,54 0,76 0,58 0,46 dium-pygidium, R = rachis). Measurements (in ram) of a few (a) cranidia- cephalons and (b) proto-
(b) 1P LP 1R LR LP/1P L R / 1 R LR/LP 1R/1P
pygidia-pygidia of Trime- UM2-CCt7 1,72 0,64 0,43 0,57 0,37 1,33 0,89 0,25 rocephalus lelievrei sp. UM2-CCt22 2,28 0,83 0,62 0,71 0,36 1,15 0,86 0,27 nov. from Famennian of North Africa. (L = length, UM2-CCt19 3,25 1,07 0,91 0,84 0,32 0,92 0,79 0,28 l = maximum width, b = UM2-CCtll 4,00 1,25 1,13 1,00 0,31 0,88 0,80 0,28 minimum width, C =cra- UM2-CCt29 5,88 2,00 1,69 1,63 0,34 0,96 0,82 0,29 nidium-cephalon, P = UM2-CCt52 6,75 2,31 2,13 1,94 0,34 0,91 0,84 0,32 protopygidium-pygidium, tt = rachis). UM2-CCt54 9,00 3,00 2,75 2,50 0,33 0,91 0,83 0,31
le
1¢1
Ib
l a
2e
G 2a
2b
2a
FIGURE 3 - Trimerocephalus lelievrei nov. sp. Fig. la-lg, s6rie ontog6n6tique croissante de pygidium, vue dorsa]e, la, UM2- CCt7; lb, UM2-CCt22; 1c, UM2-CCtl9; ld, UM2-CCtll; le, UM2-CCt29; lf, UM2-CCt52; lg, UM2-CCt54. Fig. 2a-2g, s6rie ontog6n6tique croissante de c6phalon, vue dorsale. 2a, UM2- CCt39; 2b, UM2-CCt6; 2c, UM2-CCt38; 2d, UM2-CCt23; 2e, UM2-CCt19; 2f, UM2-CCtll; 2g, UM2-CCt29. Figs. la-lg, ontogenetic series of pygidium, dorsal view. Figs. 2a-2g, ontoge- netie series of cephalon, dorsal view. Scale: 1 ram.
OSMOLSKA, 1963: (i) trac6 de la suture faciale, (ii) contour frontal de la glabelle parabolique, (iii) lobe frontal non pointu ant6rieurement. Mais dia- nopso ides poss6de des protub6rances oculaires (r6miniscences du lobe oculaire ancestral) au niveau de l'angle g6nal ant6rieur, un pygidium plus long (sag) indistinctement segment6. Par ailleurs, $2 et $3 sont absents.
L'esp6ce la plus proche de lel ievrei est caecus (G~)RIC~, 1896): (i) trac6 de ta suture faciale, (ii) absence de protub6rances oculaires, (iii) pr6sence de $2 et $3, (iv) m~me type de sculpture. Cependant, chez caecus le c6phalon est plus 6troit. Les joues fortement convexes sont tir6es vers l'arri6re. Le bourrelet lat6ral disparalt ainsi bien avant l'angle ant6rieur de la glabelle, en vue dorsale. La nouvelle esp6ce pr6sente des joues uniform6ment enfl6es et tr6s 6tal6es dans le plan horizontal. Notons 6galement la pr6sence chez caecus d'un $1 plus prononc6 dans sa partie m6diane, d'un sillon occipital moins anguleux au niveau de l'emplacement des lobes lat6raux. En vue frontale, la largeur (tr) des joues est nette- ment inf6rieure ~ la 1/2 largeur de la glabelle (tr) chez caecus. Chez la nouvelle esp6ce, la largeur (tr) des joues est au moins 6gale ~ la 1/2 largeur (tr) de la glabelle. Par ailleurs, les sillons dorsaux du pygidium sont plus superficiels chez caecus (Osm51ska 1958). La longueur maximale des pl6vres (tr) par rapport ~ la largeur (tr) du rachis est plus importante chez la nonvelle esp6ce. La largeur (tr) depuis l'angle ant6ro-tat6ral du pygi- dium au sillon dorsal par rapport ~ celle depuis l'angle post6ro-lat6ral du pygidium au sillon dor- sal est plus importante chez lelievrei. D'autre part, la bordure post6rieure semble plus parabo- lique chez caecus alors que, chez lelievrei, la marge post6rieure du pygidium pr6sente une
167
l~g~re inflexion vers l 'avant dans sa portion m~diane.
R ~ p a r t i t i o n - Toutes les esp~ces appartiennent au D~vonien terminal: Famennien to II-IV (acrn~ au Famennien to II) (Chlup~5 1977; Becker & Schreiber 1994). La nouvelle esp~ce est probable- ment un peu plus jeune (Famennien to III, Zone inf~rieure ~ marginifera) que dianopsoides (to II, to III?) et caecus (to II-III) dont l'~ge par rapport ~ la zonation des conodontes n'est toutefois pas ~tablie.
Trimerocephalus est repr~sent~ en Asie Mineure, au Kazakhstan, darts l 'Oural et largement en Europe: il est connu depuis longtemps en Angleterre, dans les Massifs rh~nan et thurin- gien, dans le Harz et en Montagne Sainte-Croix (Richter & Richter 1926). Plus r~cemment, il a ~t~ d~couvert dans le MassifArmoricain (Morzadec & Babin 1976) et la Cha~ne Cantabrique (Arbizu 1985). Jusqu'ici, sa presence a ~t~ mentionn~e, sans description ni figuration, darts le Tafilalet (Maroc) par Alberti en 1970. I1 pourrai t s'agir d 'une forme voisine de la nouvelle esp~ce. L'affinit~ entre les esp~ces proches: dianopsoides, present en Pologne, et caecus, present en Pologne, en Allemagne, en Oural et dans le Mass i f Armoricain, indiquerait des communications pos- sibles entre les marges de la Laurobaltica et le Nord-Gondwana.
Cet te diversit~ des phacopid~s adapt~s aux domaines de plates-formes externes coincide avec l 'expansion des facies p~lagiques au D~vonien sup~rieur (Chlup~5 1975).
D~veloppement ontog~n~tique - Le materiel comprenant diff~rents stades ontog~n~tiques,
partir des stades meraspis, a permis d'aborder par t ie l lement la morphog~n~se de lelievrei (tableau de mesures Fig. 2; reconsti tution Fig. 3). Selon les modes d'exuviation, les pi~ces c~pha- liques se pr~sentent sans doublure (stades meras- pis et juveniles pr~coces) ou avec doublures (stades juveniles tardifs et adultes). En effet, le genre Trimerocephalus pr~sente, au cours de son ontog~nie, deux types d'exuviation successifs . Tout d'abord, un mode d'exuviation o~ la suture faciale marginale, ligne de moindre r~sistance, est fonctionnelle comme chez la plupart des trilobites. Ce mode a ~t~ obser-v~ darts les jeunes stades ontog~n~tiques de Trimerocephalus (Maksimova 1955; OsmSlska 1963). Celui-ci est suivi d'un mode d'exuviation of~ les sutures c~phaliques sont ankylos~es, la fente d'exuviation se produisant au niveau de rart iculat ion c~phalo-thoracique. Ce mode caract~rise les stades holaspis (Richter 1937; Maksimova 1955; OsmSlska 1963).
STADES MERASPIS
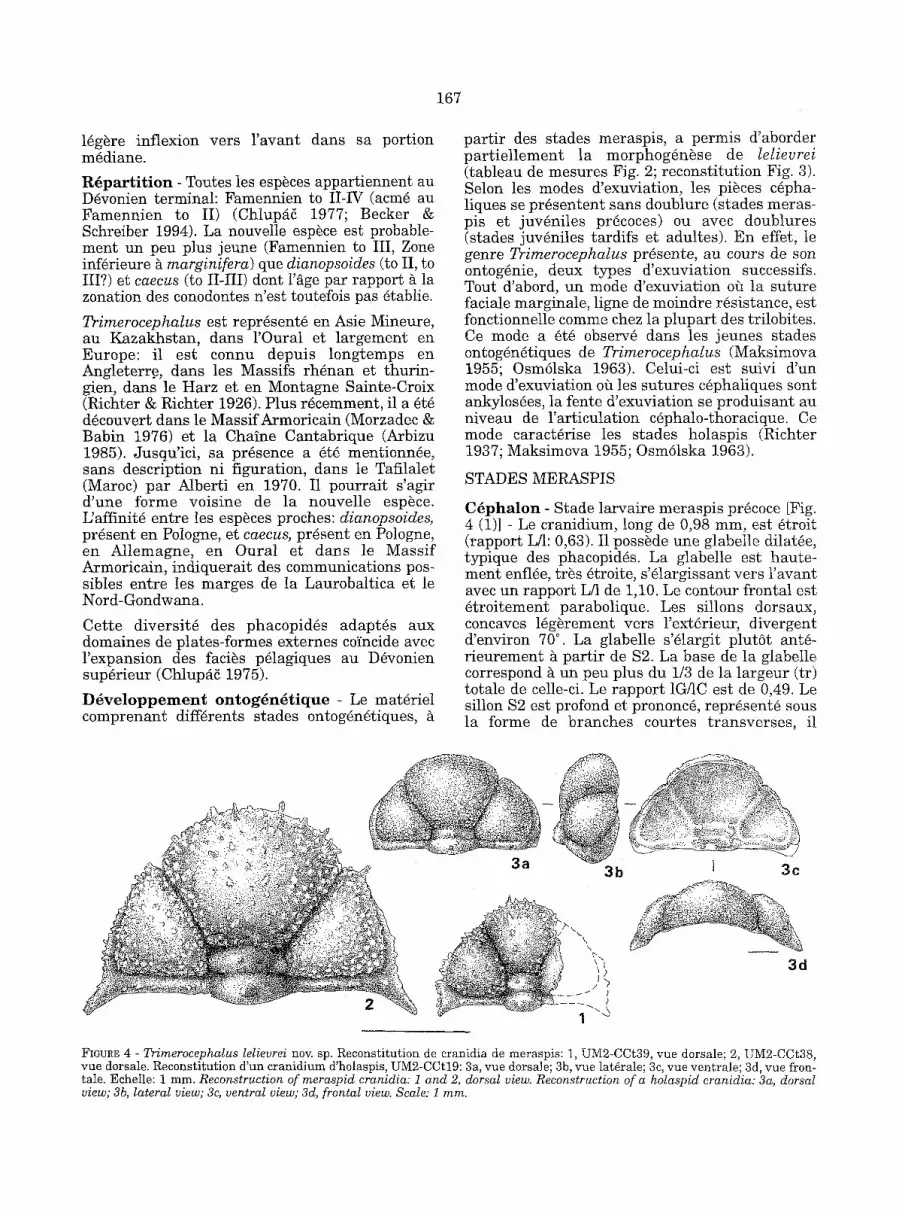
C ~ p h a l o n - Stade larvaire meraspis pr~coce [Fig. 4 (1)] - Le cranidium, long de 0,98 ram, est ~troit (rapport L/l: 0,63). I1 poss~de une glabelle dilat~e, typique des phacopid~s. La glabelle est haute- ment enfl~e, tr~s ~troite, s '~largissant vers l 'avant avec un rapport L/1 de 1,10. Le contour frontal est ~troitement parabolique. Les sillons dorsaux, concaves l~g~rement vers l'ext~rieur, divergent d'environ 70 ° . La glabelle s'~largit plutSt ant~- r ieurement h part ir de $2. La base de la glabelle correspond ~ un peu plus du 1/3 de la largeur (tr) totale de celle-ci. Le rapport 1G/1C est de 0,49. Le sillon $2 est profond et prononc~, repr~sent~ sous la forme de branches courtes t ransverses , il
1 I 3c
3d
FIGURE 4 - Trimerocephalus lelievrei nov. sp. Recons t i tu t ion de eranidia de meraspis : 1, UM2-CCt39, vue dorsale; 2, UM2-CCt38, r u e dorsale. Recons t i tu t ion d 'un c ran id ium d'holaspis, UM2-CCt19: 3a, vue dorsale; 3b, vue lat~rale; 3c, r u e vent ra le ; 3d, vue fron- tale. Echelle: 1 ram. Reconstruction of meraspid cranidia: 1 and 2, dorsal view. Reconstruction of a holaspid cranidia: 3a, dorsal view; 3b, lateral view; 3c, ventral view; 3d, frontal view. Scale: 1 mm.
168
atteint les sillons dorsaux. Ce dernier est parall~- le ~t un S1 continu, profond lat~ralement formant un apod~me. $3 semble ~tre absent. L'anneau pr~occipital est fortement enflS, peu large (tr), presqu'aussi long (sag) et enfl~ que l 'anneau occi- pital. L'anneau pr~occipital poss~de deux ~pines; quant ~ ranneau occipital, une importante ~pine m~diane s'y situe. Les lobes lat~raux pr~occipi- taux et occipitaux ne sont pas diff~renci~s. Les joues sont ~troites, fortement entices, plongeantes vers le bourrelet externe. Une l~g~re protuberan- ce existe au niveau de l'angle gSnal ant~rieur, marquee par des ~pines plus fortes que celles pr~- sentes sur le reste de la joue. La suture faciale propariale est fonctionnelle: ]es librig~nes sont d~tachSes. L'angle g~nal poss~de une pro~minente et longue ~pine g~nale se projetant vers l'arri~re. En vue lat~rale, le contour de l 'anneau pr~occipi- tal est convexe.
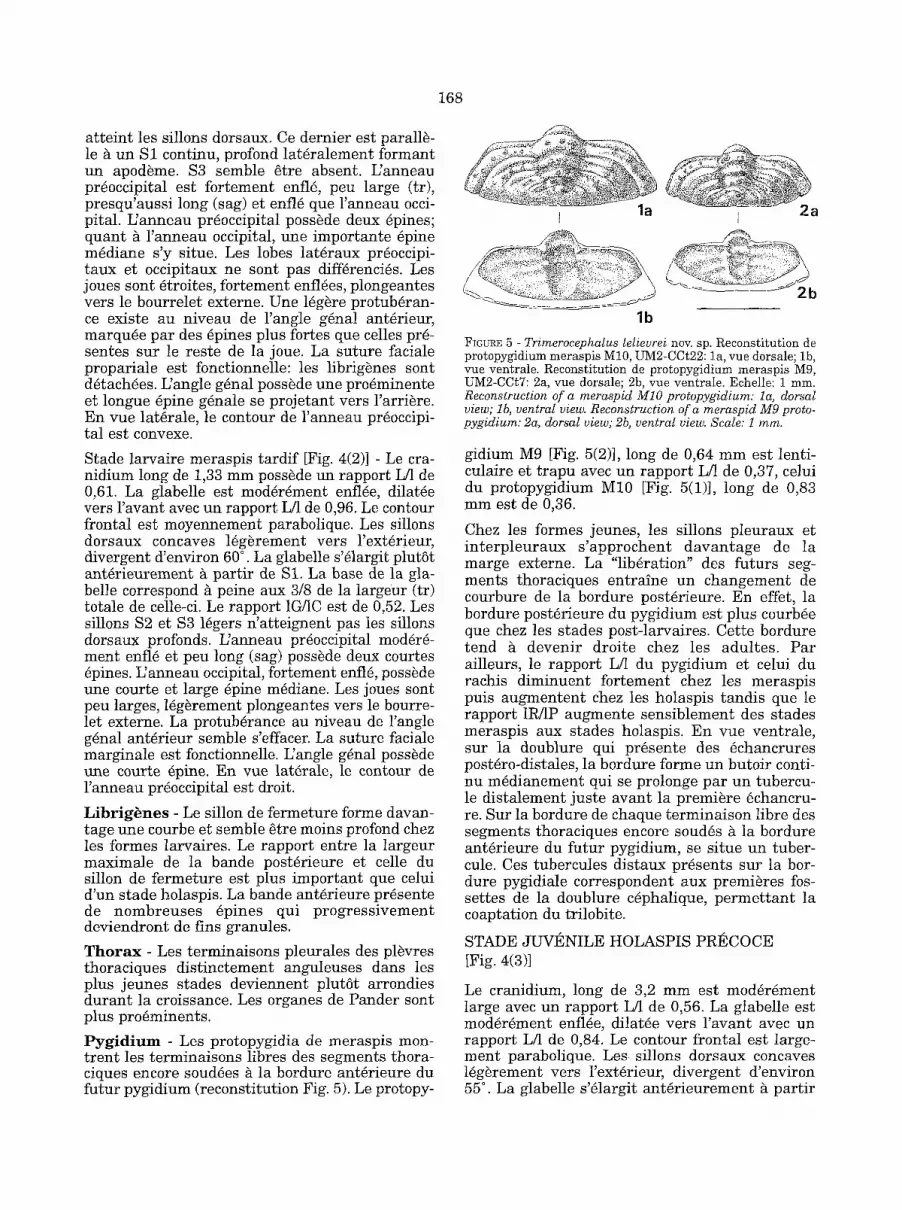
Stade larvaire meraspis tardif [Fig. 4(2)] - Le cra- nidium long de 1,33 mm poss~de un rapport L/1 de 0,61. La glabelle est mod~r~ment enfl~e, dilat~e vers l 'avant avec un rapport L/1 de 0,96. Le contour frontal est moyennement parabolique. Les sillons dorsaux concaves ]~g~rement vers l'ext~rieur, divergent d'environ 60 ° . La glabelle s'~largit plut~t ant~rieurement ~ partir de $1, La base de la gla- belle correspond ~ peine aux 3/8 de la largeur (tr) tota]e de celle-ci. Le rapport 1G/1C est de 0,52. Les sillons $2 et $3 l~gers n'atteignent pas les sfilons dorsaux profonds. L'anneau pr5occipital modSr~- ment enfl~ et pen long (sag) poss~de deux courtes 6pines. L'anneau occipital, fortement enflS, poss~de une courte et large ~pine m~diane. Les joues sont peu larges, l~g~rement plongeantes vers le bourre- let externe. La protuberance au niveau de l'angle g~nal ant~rieur semble s'effacer. La suture faciale marginale est fonctionnelle. L'angle g~nal poss~de une courte ~pine. En rue lat~rale, le contour de ranneau pr~occipital est droit.
L i b r i g ~ n e s - Le sillon de fermeture forme davan- tage une courbe et semble ~tre moins profond chez les formes larvaires. Le rapport entre la largeur maximale de la bande post~rieure et celle du sillon de fermeture est plus important que celui d'un stade holaspis. La bande ant6rieure pr~sente de nombreuses 6pines qui progress ivement deviendront de fins granules.
T h o r a x - Les terminaisons pleurales des pl~vres thoraciques dist inctement anguleuses dans les plus jeunes stades deviennent plutSt arrondies durant la croissance. Les organes de Pander sont plus pro6minents.
P y g i d i u m - Les protopygidia de meraspis mon- trent les terminaisons libres des segments thora- ciques encore soud~es ~ la bordure ant6rieure du futur pygidium (reconstitution Fig. 5). Le protopy-
t la l 2 a
l b
L ~
FIGURE 5 - Trimerocephalus lelievrei nov. sp. Reconstitution de protopygidium meraspis M10, UM2-CCt22: la, vue dorsale; lb, vue ventrale. Reconstitution de protopygidium meraspis M9, UM2-CCt7: 2a, vue dorsale; 2b, vue ventrale. Echelle: 1 ram. Reconstruction of a meraspid MIO protopygidium: la, dorsal view; lb, ventral view. Reconstruction of a meraspid M9 proto- pygidium: 2a, dorsal view; 2b, ventral view. Scale: 1 ram.
gidium M9 [Fig. 5(2)], long de 0,64 m m e s t lenti- culaire et t rapu avec un rapport L/1 de 0,37, celui du protopygidium M10 [Fig. 5(1)], long de 0,83 mm est de 0,36.
Chez les formes jeunes, les sillons pleuraux et in te rp leuraux s 'approchent davan tage de la marge externe. La "lib6ration" des futurs seg- ments thoraciques entraine un changement de courbure de la bordure post6rieure. En effet, la bordure post6rieure du pygidium est plus courb6e que chez les stades post-larvaires. Cette bordure tend /~ devenir droite chez les adultes. Par ailleurs, le rapport L/1 du pygidium et celui du rachis diminuent fortement chez les meraspis puis augmentent chez les holaspis tandis que le rapport 1R/1P augmente sensiblement des stades meraspis aux stades holaspis. En vue ventrale, sur la doublure qui pr6sente des ~chancrures post6ro-distales, la bordure forme un butoir conti- nu m~dianement qui se prolonge par un tubercu- le distalement juste avant la premiere ~chancru- re. Sur la bordure de chaque terminaison libre des segments thoraciques encore soud~s g la bordure ant6rieure du futur pygidium, se situe un tuber- cule. Ces tubercules distaux presents sur la bor- dure pygidiale correspondent aux premieres fos- settes de la doublure c~phalique, permet tant la coaptation du trilobite.
STADE JUVENILE HOLASPIS PRI~COCE [Fig. 4(3)]
Le cranidium, long de 3,2 mm est mod~r6ment large avec un rapport L/1 de 0,56. La glabelle est mod6r6ment enfl~e, dilat6e vers l 'avant avec un rapport Lfl de 0,84. Le contour frontal est large- ment parabolique. Les sillons dorsaux concaves l~g~rement vers l'ext~rieur, divergent d'environ 55 °. La glabelle s'61argit ant6rieurement fi partir

169
de $1. La base de la glabelle correspond au 3/7 de la largeur (tr) totale de celle-ci. Le rapport 1G/1C est de 0,56. Les sillons $2 et $3 16gers n'attei- gnent pas les sillons dorsaux profonds. L'anneau pr6occipital, peu enfl6 et peu long (sag), poss6de deux 16gers tubercules. L'anneau occipital, forte- ment enfl6 et deux lois plus long (sag), poss6de un gros tubercule m6dian. Lea joues sont larges, 6tal6es dans le plan horizontal, ne pr~sentant aucune protub6rance oculaire. La suture faciale marginale est fonctionnelle: les librig6nes li6es la doublure sont s6par6es. L'angle g6nal poss~de un tubercule assez pro6minent qui diminue pro- gressivement jusqu 'au stade adulte.
CONCLUSION ET DISCUSSION
A partir des donn6es du nouveau mat6riel, il appa- ra~t que: (i) les dimensions relativement plus larges du c6phalon, chez les formes plus ag6es, r6sultent d'un changement de convexit6 et de forme de la gla- belle qui s'61argit lat6ralement: le rapport IA du c6phalon diminue ainsi que celui de la glabelle, tan- dis qne le rapport b/1 de la glabelle augmente; (ii) la 16g~re protub6rance (t6moin d'une r6miniscence du lobe oculaire ancestral), soulign6e par des 6pines plus marqu6es au niveau de l'angle g6nal ant6rieur, disparait totalement dans un stade mature; (iii) dans les stades jeunes, la suture faciale est fonc- tionnelle, l 'ankylose n'apparait que dans un stade holaspis tardif; (iv) les nombreuses petites 6pines distribu6es sur l'ensemble du cranidium, les deux 6pines de l 'anneau pr6occipital, l'6pine m6diane de l 'anneau occipital se r6duisent progressivement des tubercules plus ou moins distincts; (v) la lon- gueur de l'6pine g6nale se r6duit 6galement ~ un tubercule qui disparait ~ son tour. La d6croissance de l'6pine g6nale semble ~tre de l'ordre allom6- trique. Ceci avait 6galement 6t6 observ6 dans le mat6riel trilobitique d'Osm61ska (1963) et chez les exemplaires peu nombreux, d6calcifi6s et d6form6s de Morzadec & Babin (1976). La pr6sence de cette 6pine g6nale dans les stades jeunes de divers genres atteste sa g6n6ralisation chez les phaco- pid6s (Osm61ska 1963; Jahnke, 1969; Chatterton 1971; Alberti 1972) et peut 6tre consid6r6e comme crit6re immature distinctif.
La similitude entre les stades pr6coces meraspis de Phacops spedeni CHATTERTON, 1971 et ceux de Trimerocephalus lelievrei sugg6re une relation phylog6n6tique 6troite entre les genres Phacops et Trimerocephalus. Ils soulignent des change- ments ontog6n6tiques similaires incluant: (i) l'6- largissement de la glabelle, (ii) la d6croissance de l'6pine g6nale en longueur, (iii) la d6croissance des 6pines recouvrant le cranidium. Cependant, alors que chez Fanc~tre Phacops spedeni, l'oeil se d6ve-
loppe et migre vers l'arri6re et vers la glabelle, chez le descendant Trimerocephalus lelievrei, le stade de l'oeil n'est jamais atteint, il se d6veloppe une protub6rance oculaire sans champ visuel qui s'efface au cours du d6veloppement ontog6n~- tique. Cet effet re tard consti tue un exemple d'h6t6rochronie du d6veloppement (Feist 1995).
D'autre part, les changements de forme interve- nant au d6but de l'ontog6nie entre les s tades lar- vaires pr6coces et tardifs seraient probablement dus ~ une modification du mode de vie (Speyer & Chatterton 1989). Les larves minuscules orn6es d'6pines devaient mener une vie planctonique. Ceci est probablement l'une des raisons de l'ab- sence des stades protaspis dans notre mat6riel constitu6 de formes benthiques.
Notons 6galement que l 'extraordinaire accumula- tion de ces exuvies silicifi6es imbriqu6es dans les os phosphat6es d'arthrodires contraste avec la faible densit6 des trilobites calcaires pr6sents lat6- ralement dans le m6me banc dans les endroits d6pourvus d'arthrodires. I1 s'agit probablement d'un site pr6f6rentiel afin de permettre ~ ces orga- nismes benthiques d'effectuer leur mue ~ proxi- mit6 d'une source de nourriture et dans un milieu extr~mement calme (s6diment marno-calcaire fin).
La reconstitution de l'ontog6nie tardi-larvaire et juv6nile de cette nouvelle esp6ce pourrait ~tre consid6r6e comme une r6f6rence parmi les phaco- pid6s du D6vonien sup6rieur.
R e m e r c i e m e n t s - Nous remercions tout particuli6rement H. Leli~vre qui a d6couvert le mat6riel utilis6 dans cette 6tude et d6termin6 l 'arthrodire associ6 aux trilobites, L. Meslin pour la r6alisation des dessins et son aide technique, M. Datas pour les photographies au MEB et J. Martin et M. Pons pour les tirages photographiques. Ceci est une contribution de I'UMR 5554, Montpellier (ISE 96/127).
R]~Ft~RENCES
ALBERTI H. 1970 - Neue Trilobiten-Faunen aus dem Ober-Devon Marokkos. GSttinger Arbeiten zur Geologie und Paliiontologie, 5: 15-29.
ALBERTI H. 1972 - Ontogenie des Trilobiten Phacops accipitrinus. Neues Jahrbuch fi~r Geologie und Pal(tontologie. Abhandlungen, 141: 1-36.
A~BIZU M. 1985 - Trilobites Phacopinae de la Formaci6n Vidrieros, en el ~rea de Gildar-Mont6 (Le6n, NO de Espafia) y su distribuci6n estratigr~fica. Trabajos de Geologia, Universidad de Oviedo, 15: 67-75.
BECKER R.T. & SCHREIBER G. 1 9 9 4 - Zur Trilobiten - Stratigraphie im Letmather Famennium (n6rdliches Rheinisches Schiefergebirge). Berliner geowissen schaftliche Abhandlungen (E), 13: 369-387.
CHATTERTON B.D.E. 1971 - Taxonomy and ontogeny of Siluro-Devonian trilobites from near Yass, New South Wales. Palaeontographica (A), 137: 1-108.

170
CHLUPA5 I. 1975 - The d is t r ibu t ion of phacopid t r i lobi tes in space and t ime. Fossils and Strata, 4: 399-408.
CHLUP~C I. 1977 - The phacopid t r i lobi tes of the S i lu r ian and Devonian of Czechoslovakia. Vydal []stredni ~tstav geologicky, 43: 1-164.
CLARKSON E.N.K. 1979 - Invertebrate palaeontology and evolution. 323 p., George Al len & Unwin Ldt, London.
FEIST R. 1995 - Effect of paedomorphosis in eye reduc- t ion on pa t t e rn s of evolution and ext inct ion in tri lo- bites. In McNAMARA K.J. (ed.): Evolutionary change and heterochrony. John Whi ley & Sons Ltd: 225-244.
GURICH G. 1896 - Das Palaeozoicum im Polnischen Mit telgebirge. Verhandlungen der Russisch-Kaiser- lichen Mineralogischen Gesellschaft, 32: 1-539.
HARRINGTON.J. 1959 - Genera l descr ipt ion of Trilobita. In MOORE R.C. (ed.): T r e a t i s e on I n v e r t e b r a t e Paleontology. P a r t O, A r t h r o p o d a 1. Geological Society of America & University of Kansas Press, Lawrence: O38-Ol17.
HuPI~ P. 1953 - Classif icat ion des tr i lobites. Annales de Paldontologie, 39: 1-110.
JAHNKE H. 1969 - Phacops zinkeni F. A. ROEMER 1843 - ein Beispiel ftir eine ontogenetische Entwicklung bei Phacopiden (Trilobitae, Unterdevon). Neues Jahrbuch fiir Geologie und Pali~tontologie. Abhandlungen, 133: 309-324.
LEHMAN J.P. 1956 - Les a r th rod i res du D6vonien sup6- r i eu r du Tafi la le t (sud marocain). Notes et Mdmoires du Service Gdologique de Rabat, 129:70 p.
MAKSIMOVA Z.A. 1955 - Tri lobi ty srednego i verchnego devona U r a l a i severnych Mugodzar. Trudy vseso-
jusn. nautschno-issled, geol. Inst. (VSEGEI), 3: 1-263, Moskva.
MORZADEC P. & BABIN C. 1976 - Pr6sence de Trimero- cephalus caecus (GORICH, 1896) (Trilobita, Phaco- pidae) dans le Famenn ien inf~rieur du Finist~re: in t6r~t s t r a t i g r a p h i q u e et pa l6og6ograph ique . Geobios, 9 (3): 367-371.
OSMOLSKA H. 1958 - F a m e n n i a n Phacopidae from Holy Cross Mts (Poland) .Acta Palaeontologica Polonica, 3: 119-148.
OSMOLSKA H. 1963 - On some F a m e n n i a n P h a c o p i n a e (T r i l ob i t a ) f rom the H o l y Cross M o u n t a i n s (Poland). Acta Palaeontologica Polonica, 8 (4): 495- 523.
RICHTER R. 1937 - Von Bau und Leben der Tri lobiten. 8. Die "Sal tersche Einbe t tung" als Folge und Kenn- zeichen des H~utungs-Vorgangs. Senckenbergiana, 19: 413-431.
RICHTER n. • RICHTER E. 1926 - Die Tr i lobi ten des Oberdevons. Beitr~ige zur K e n n t r i s devon i scher Tri lobi ten IV. Abhandlungen der preussischen geolo- gischen Landesanstalt, 99: 1-314.
SPEYER S.E. & CHATTERTON B.D.E. 1989 - Tri lobi te l a rvae and la rva l ecology. Historical Biology, 3: 27-60.
C. CR(}NIER & R. F E I S T Laboratoire de Pal6ontologie
Institut des Sciences de l'Evolution, UMR 5554 Universit6 Montpellier II C 64, Place E. Bataillon
F-34095 Montpellier cedex 05 E-Mail: [email protected], fr
rfeist@is em.univ-montp2, fr