rugose corals from the upper viséan (carboniferous) of the jerada massif (ne morocco): taxonomy,...
TRANSCRIPT

RESEARCH PAPER
Rugose corals from the upper Visean (Carboniferous)of the Jerada Massif (NE Morocco): taxonomy, biostratigraphy,facies and palaeobiogeography
Markus Aretz
Received: 14 September 2009 / Accepted: 24 October 2009 / Published online: 25 November 2009
� Springer-Verlag 2009
Abstract The Visean succession of the Jerada Massif
contains a relatively diverse rugose coral fauna, which
comprises 12 genera. Only two of these (Siphonodendron
and Lithostrotion) are reported to include more than one
species. Most taxa show distinctive facies dependencies.
Small solitary corals are found in shaly environments of the
Oued Es-Sassi Formation. On the northern flank a coral
patch reef formed on an oolitic shoal in the Koudiat Es-
Senn Formation. Its core mainly consists of Lithostrotion
vorticale colonies, whereas in its surroundings literally
hundreds of specimens of Siphonophyllia samsonensis
occur. Associated with these dominant taxa occur colonial
and further large solitary taxa (Palaeosmilia, Dibunophyl-
lum). Tizraia and Pareynia are restricted to microbial-
dominated buildup facies. Their presence might be strongly
controlled by the development of this buildup type, because
further occurrences in Algeria, Morocco, and Belgium are
all in the same facies. The coral fauna at Jerada is a typical
Late Visean assemblage for the Western European Prov-
ince. The Eastern Moroccan Meseta may be an important
pathway within the province for the connection between
the Central Saharan basins and NW Europe. The bio-
stratigraphic ages of the coral fauna partly contradict ages
based on carbonate microfossils; the coral ages are slightly
older and typical Brigantian coral taxa are absent.
Keywords Visean � Rugose corals � Jerada Massif �Eastern Moroccan Meseta � Palaeobiogeography
Kurzfassung Die viseische Abfolge des Jerada-Massivs
enthalt eine relativ diverse Fauna. Diese Fauna umfasst 12
Gattungen, aber mehrere Arten sind nur in zwei dieser
Gattungen (Siphonodendron und Lithostrotion) vertreten.
Die meisten Taxa zeigen eine deutliche Fazies-
Abhangigkeit. Kleine solitare Formen sind nur aus schiefr-
igen Gesteinen der Oued Es-Sassi-Formation bekannt. Auf
dem Nordflugel des Jerada-Synklinoriums bildete sich in
der Koudiat Es-Senn-Formation an einer oolitischen Barre
ein Korallen-Patch-Riff.Sein Kern besteht im Wesentlichen
aus Lithostrotion vorticale-Kolonien. Das Riff wird von zah-
lreichen (einigen hundert) Exemplaren von Siphonophyllia
samsonensis umgeben. Assoziiert mit diesen beiden domi-
nierenden Taxa sind weitere großwuchsige solitare (Pala-
eosmilia, Dibunophyllum) und koloniale Taxa. Tizraia und
Pareynia sind nur in mikrobiell-dominierter Buildup-Fazies
zu finden. Ihr Vorkommen scheint generell auf diesen
Builduptyp beschrankt, wie weitere Vorkommen in gleicher
Fazies in Algerien, Marokko und Belgien zeigen. Die
Korallenfauna von Jerada ist typisch fur Assoziationen des
spaten Viseums innerhalb der westeuropaischen Faunen-
provinz. Die Ostliche Marokkanische Meseta stellt einen
moglichen Verbindungsweg innerhalb dieser Provinz
zwischen den Becken der zentralen Sahara und Westeuropa
dar. Die biostratigraphischen Alter der Korallen stehen in
einem gewissen Widerspruch zu den auf kalkigen Mikro-
fossilien beruhenden Altern. Die Korallen-Alter sind etwas
alter, typische Vertreter des Brigantiums fehlen.
Schlusselworter Viseum � Rugose Korallen �Jerada Massiv � Ostliche Marokkanische Meseta �Palaobiogeographie
M. Aretz (&)
Universite de Toulouse (UPS), LMTG (OMP),
14 Avenue Edouard Belin, 31400 Toulouse, France
e-mail: [email protected]
M. Aretz
CNRS, LMTG, 31400 Toulouse, France
M. Aretz
IRD, LMTG, 31400 Toulouse, France
123
Palaontol Z (2010) 84:323–344
DOI 10.1007/s12542-009-0046-0

Introduction
Rugose corals of Carboniferous age are relatively poorly
studied in NW Africa. Diverse shallow water faunas have
been described from the Bechar Basin (Semenoff-Tian-
Chanksy 1974, 1985) and the Eastern Central Moroccan
Meseta (Said et al. 2007; Said and Rodrıguez 2007, 2008,
Aretz and Herbig 2009). However, limited numbers of
specimens from various regions and facies found during
reconnaissance work and mapping (e.g. Termier and
Termier 1950; Conrad 1984) clearly indicate the likelihood
of comparable abundances and diversities in NW Africa
(Fig. 1) as in contemporaneous tropical-shelf systems (e.g.
western USA: Sando 1980, Belgium: Poty 1981). The only
briefly described fauna of the Anti-Atlas (Wendt et al.
2001) shows a possible mixture of taxa of deeper and
shallow water environments. The unpublished material
of the Bechar Basin and further Algerian Basins (coll.
Semenoff-Tian-Chansky, Museum Nationale d’Histoire
Naturelle, Paris) contains numerous small undissepimented
solitary rugose corals, which probably represent deeper
water environments and environments similar to the
Anti-Atlas.
The record of Carboniferous corals in the Eastern
Moroccan Meseta is very poor. Owodenko (1946, 1976)
mentioned Caninia juddi Nicholson (? = Pseudazphren-
toides juddi), Dibunophyllum sp., Zaphrentis sp. and
Favosites parasitica d’Orbigny (? = Sutherlandia) from
the Visean succession of the Jerada Massif. However, the
depository for this material is unclear, and a revision of
those specimens is inevitable.
It is the purpose of this paper:
1. to describe newly collected rugose corals from the
upper Visean of the Jerada Massif;
2. to place the specimens in a facies context; and
3. to briefly discuss stratigraphic and palaeobiogeograph-
ic implications.
Geological setting
The Jerada Massif is part of the Palaeozoic basement of the
Eastern Moroccan Meseta deformed during the Variscan
Orogeny and today widely covered by Mesozoic–Caino-
zoic strata. It belongs to the inner zones within the tectonic
framework of the northwest African Variscan domain
(Pique et al. 1993).
The simple-structured WSW–ENE synclinorium is sit-
uated about 50 km south of the town of Oujda near the
Moroccan–Algerian border (Fig. 2). It is about 30 km in
strike and about 15 km normal to strike. The dip of the
northern flank is gentle (15–30�S), whereas the southern
flank dips steeply (80�N), and may locally be overturned.
The synclinorium contains an exclusively Carboniferous
succession (Owodenko 1946, 1976). A Visean basin fill
sequence (Herbig et al. 2006) is unconformably overlain by
a Namurian to Westphalian paralic sequence (Desteucq
et al. 1988). It ends with coal-bearing strata of Westphalian
B–C age below the Variscan unconformity.
Herbig et al. (2006) established a lithostratigraphic
subdivision of the Visean succession (Fig. 3), which
substituted the formations of Berkhli et al. (1999) defined
in repeated succession of tectonic slices. The Cafcaf For-
mation mainly consists of bedded cherts. The succeeding
Oued El Koriche Formation is lithological variegated,
which results from the deposition of debris flows, turbidites
Fig. 1 Geological map of NW Africa (after Pique 2001) with
location of the main coral occurrence. See text for main references for
coral data
324 M. Aretz
123

(siliciclastic and carbonate dominated ones), and olistoliths
in mud- to silt-dominated lower and middle slope facies.
The Oued Es-Sassi Formation comprises mainly silty
shales. The general lithology and thickness of all forma-
tions varies between northern and southern flank, but also
on the individual flanks. Probably the most striking dif-
ference between the two flanks is the composition of the
Koudiat Es-Senn Formation, which tops the Oued Es-Sassi
Formation. On the southern flank it contains a single sed-
imentary cycle, which includes microbial-sponge buildups
and small microbial-metazoan buildups (Aretz and Herbig
2008); whereas on the northern flank the formation is twice
as thick, consists of two sedimentary cycles (Berkhli et al.
1999), and only contains a single rugose coral patch reef in
the uppermost beds (Aretz et al. 2006). All sedimentary
cycles in the Koudiat Es-Senn Formation consist of a lower
siliciclastic-dominated part and a carbonate-dominated
upper part. The formation is locally topped by an ignim-
brite (Berkhli et al. 2000) indicating subaerial exposure of
the region around the Visean/Namurian boundary. Sub-
sequent Namurian strata are dominated by fine-grained
siliciclastics facies (‘‘Namurian-Westphalian shales’’).
Aretz and Herbig (2007) showed that generally more
distal facies occurs in the NE of the synclinorium, whereas
the most proximal facies is reported from the SW (Oued
Agaia area), thus indicating a NE-wards tilted basin. Hence
local and regional palaeogeography markedly affected the
sedimentation patterns in the Jerada Massif.
Materials and methods
Most corals are from limestones of the Oued El Koriche
and Koudiat Es-Senn formations, but a few records are
from the Cafcaf and Oued Es-Sassi formations (Fig. 3).
They have been collected during two field campaigns in
2004 and 2006, which focussed on mapping, buildup, and
reef facies and Mississippian basin development. Rare
single section of tabulate corals (michelinids, auloporids)
and heterocorals (Hexaphyllia), not described herein, were
found in petrographic thin sections.
A more focussed collection on corals may result in a
more diversified fauna, and would enable, in many cases,
more precise determination of intra-specific variability.
The quality of the material and thus the availability of thin
sectioned rugose corals is limited, because not all of the
material studied has been collected for this purpose.
However, at least one vertical section is always present. In
a few cases well preserved specimens could be found, but
the majority is either enclosed in limestone or outer parts
have been eroded, as typical for many African specimens
(Semenoff-Tian-Chansky 1974). The terminology of mor-
phological characters follows Hill (1981) and Poty (1981).
The attribution of genera to the higher taxonomic levels of
Hill (1981) is largely followed in this work. All material
is stored in the Institut fur Geologie und Mineralogie,
Universitat zu Koln under the numbers GIK 2156-2240.
Institutional abbreviations: BM, British Museum (Natural
History) London; DPM, Departamento de Paleontologıa,
Universidad Complutense de Madrid; HM Hunterian Museum
Glasgow; MNHNP, Museum National d’Histoire Naturelle,
Paris; NMD, National Museum of Irland, Dublin; RSM, Royal
Scottish Museum, Edinburgh; SM, Sedgwick Museum,
University of Cambridge; UMO, University Museum Oxford.
Systematic palaeontology
Systematic descriptions are presented for all taxa. Some
descriptions of well-known species are limited to key
Fig. 2 Geological map of the
Jerada synclinorium and
location of the sample localities
(P1-P10), with inset showing
the Palaeozoic massifs of the
Eastern Moroccan Meseta. Inset
after Berkhli et al. (1993)
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 325
123

points and less common described characters. To reduce
the lengths of the synonymy lists, the only references added
are those not included in recent synonymy list which the
author agrees with (cum. syn.).
Class Anthozoa Ehrenberg, 1834
Subclass Rugosa Milne-Edwards and Haime, 1850
Order Stauriida Verrill, 1865
Suborder Caniniina Wang, 1950
Family Cyathopsidae Dybowski, 1873
Genus Siphonophyllia Scouler in McCoy, 1844
Type species: Siphonophyllia cylindrica Scouler in
McCoy, 1844.
Siphonophyllia samsonensis (Salee, 1913) (Fig. 4a–f)
*1913 Caninia samsonensis Salee: 48, pl. D fig. 1.
1975 Siphonophyllia benburbensis (Lewis, 1927).—
Hoffmann et al.: pl. 4 fig. 3.
1981 Siphonophyllia samsonensis (Salee, 1913).—Poty:
52, fig. 49 pl. 25, fig. 4, pl. 26, fig. 3, 4. (cum. syn.)
?1992 Siphonophyllia cf. samsonensis (Salee, 1913).—
Rodrıguez and Falces: 190, pl. 16 fig. 1, 2.
2004 Siphonophyllia samsonensis (Salee, 1913).—Rod-
rıguez et al.: 74, fig. 2.5, figs. 3.1-2, 3.4.
2005 Siphonophyllia samsonensis (Salee, 1913).—Cozar
et al.: fig. 12.12.
2006 Siphonophyllia samsonensis (Salee, 1913).—Galla-
gher et al.: fig. 13.4.
Holotype: The specimen illustrated by Salee could not
be located by Poty (1981) in the collections at Louvain-la-
Neuve, thus it might be considered lost.
Locus typicus and stratum typicum: Nameche, Bel-
gium; lower Upper Visean.
Diagnosis: See Poty (1981)
Remark: S. benburbensis (Lewis 1927) is a synonym of
S. samsonensis (Salee 1913), but still widely used in the
literature.
Material: Four fragments from P1 (GIK 2172-2175), 16
fragments from P4; thin sectioned specimens (2156-2163),
uncut specimens (GIK 2164-2171).
Description: Literally hundreds of fragments from this
large straight cylindrical solitary coral can be found in the
two studied localities. However, younger stages of the
species are absent, as in many other outcrops (Semenoff-
Tian-Chansky 1974).
The state of preservation is very different. The better
preserved specimens show 2–3 growth rings per cm, and
well differentiated furrows and ridges (10–15 per cm).
Rejuvenesence occurs occasionally. The thin undulating
wall of most corallites is eroded, and this may also
include large parts of the dissepimentarium. Corallite
diameters, partly reconstructed, vary between 4.0 and
6.3 cm. There are 55–64 septa in two series. The major
septa are of variable length, but always leave a large
open axial space. They are straight and thickened in the
tabularium. Steroplasma may completely fill the space
between major septa. Within the dissepimentarium the
major septa become sinuous. Minor septa are mostly
restricted to septal spines within the dissepimenta-
rium, and only occur very rarely in the tabularium. A
cardinal fossula is always present. The counter septum
may be:
Fig. 3 Lithostratigraphy and simplified schematic lithological logs of
the Visean succession on the two flanks of the Jerada synclinorium
(not to scale)
326 M. Aretz
123

1. slightly longer, as the other major septa;
2. inconspicuous; or
3. in a counter fossula (very rare).
The dissepimentarium is variable in thickness. It con-
sists of an inner more regular part and an outer dissepi-
mentarium with transeptal dissepiments of both orders. In
the longitudinal section there are 4–9 rows of elongated
and variably inclined dissepiments (3–8 per cm). Maxi-
mum elongation is 1 cm. Tabulae are complete (9–12 per
cm). In the axial part they are flat and continue into a
peripheral depression. Within this depression stereoplas-
ma may be observed. Occasionally axial tabellae are
present.
Discussion: The Jerada specimens fit into the variability
described for this species, and almost perfectly match the
drawings of Dixon (1970) for specimens from Streedagh
Point (Ireland). A number of species of Siphonophyllia
are known from the uppermost Famennian to the early
Visean (e.g. S. garwoodi Ramsbottom and Mitchell 1980,
S. gigantea (Michelin 1843), whereas only a few species
are reported from younger strata. Vaughan (1915) has
already made the case that S. samsonensis is the end
member of one phylogentic linage within the genus char-
acterised by the arrangement of the dissepiments. S. siblyi
(Semenoff-Tian-Chansky 1974) differs from S. samsonensis
by its thicker major septa, the extreme reduction of the
minor septa, and a higher diameter/septa ratio. Dixon
(1970) showed the differences between S. samsonensis and
S. cylindrica Scouler in McCoy 1844, but, because of the
characteristic high morphological variability, single speci-
mens of S. samsonensis may show a habit of the second
species.
Distribution: Well known species in the middle and late
Visean of Europe and North Africa.
Family Hapsiphyllidae Grabau, 1928
Genus Zaphrentites Hudson, 1941
Type species: Zaphrentis curvulena Thomson, 1881
?Zaphrentites sp. (Fig. 4g)
Material: Three fragments from P4 (GIK 2177-2179),
one fragment from P8 (GIK 2176).
Description: Small solitary, curvated, trochoid coral.
The preservation is variable, calices and mature stages are
absent, walls are locally eroded. The maximum diameter is
15 mm. There are 34–37 major septa, which are thick and
confluent at the axial ends to close the marked cardinal
fossula above the centre. Septa in the cardinal half of the
corallite are less thick than those in the counter half. Ste-
roplasma is found between individual septa along the wall
preferentially in the counter half. Although the longitudinal
section is dominated by the thickened septa, few tabulae
rising towards the centre are seen. Dissepiments and minor
septa seem to be absent.
Discussion: The specimens belong to the group of
zaphrentoid corals. Studies of British coral workers (Vaughan
1906; Carruthers 1910; Hudson 1942) clearly showed the
need for detailed phylogenitic studies of the single coral and
of populations. This is not possible with the Jerada material
because of the small number of specimens and their bad
preservation. However, the transverse sections of these
specimens are similar to section in medial parts of Am-
plexizaphrentis enniskilleni (Milne-Edwards and Haime
1851) specimens (Lewis 1930; Hill 1940). However, the
characteristic variability of the fossula of that species can-
not be observed in the Jerada specimen, because the mature
stages are absent. The specimens are, in some respects,
provisionally assigned to the genus Zaphrentites. Further
specimens may help to elucidate their taxonomic position.
Suborder Aulophyllina Hill, 1981
Family Palaeosmiliidae Hill, 1940
Genus Palaeosmilia Milne-Edwards and Haime, 1848
Type species: Palaeosmilia murchisoni Milne-Edwards
and Haime, 1848
Palaeosmilia murchisoni Milne-Edwards and Haime,
1848 (Fig. 4h)
*1848 Palaeosmilia murchisoni Milne-Edwards and
Haime: 261.
?1999 Palaeosmilia murchisoni Milne-Edwards and
Haime.—Liao and Rodrıguez: 551, figs. 5.4–5.5.
2005 Palaeosmilia murchisoni Milne-Edwards and
Haime.—Aretz and Nudds: 172, pl. 1 fig. 5. (cum. syn.)
Holotype: specimen BM 48398, Bowerbank Collection.
Locus typicus and stratum typicum: Frome, Somerset,
UK; Visean. Doubts about the age and locality are sum-
marized in Hill (1940).
Diagnosis: See Semenoff-Tian-Chansky (1974)
Material: Three specimens from P1 (GIK 2183), P4
(GIK 2181) and P6 (GIK 2182).
Description: All fragments fit well into the variability
of this species (Semenoff-Tian-Chansky 1974). The best
preserved specimen is cylindrical and the calice is well
visible, but lacks most of the apical parts and the wall. The
calicular platform (max. 2.3 cm wide) surrounds a deep
U-shaped calicular pit (3.0 cm in width), which is sur-
mounted by a gentle mounted calicular boss (\0.5 cm
high, 1.5 cm wide). The 0.5 cm deep cardinal fossula is
well visible. Maximum diameter is up to 7.0 cm. In the
mature stages there are two rows of 82–94 septa. Many
septa show a marked thickening in the dissepimentarium
when arrangements of the dissepiments become more
regular, probably corresponding to the inner boundary of
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 327
123

328 M. Aretz
123

the calicular platform. Minor septa are 1/2 to 2/3 as long as
the major septa. Septa are straight to slightly sinuous in the
dissepimentarium, whereas in the tabularium the major
septa become more sinuous and may touch each other near
the axis. In the transverse section the dissepimentarium is
three-folded, an outer zone of lonsdaleoide and sometimes
naotic dissepiments, is followed by a middle zone of
more irregular dissepiments, and finally an inner zone of
numerous rows of often elongated, regular dissepiments.
Discussion: Semenoff-Tian-Chansky (1974) showed the
large variability of the species and skipped the formerly
defined sub-species (Hill 1938–1941). The Jerada speci-
mens differ by a lower diameter/septa ratio from P. mul-
tispetata Semenoff-Tian-Chansky 1974, which might be a
more compact variant of P. murchisoni. P. ressoti Menc-
hikoff and Hsu 1935 differs by its smaller size, a more open
dissepimentarium, and a large open axial space.
Distribution: This species is well known from the lower
Visean to Serpukhovian in Europe and North Africa
(Semenoff-Tian-Chansky 1974; Mitchell 1989; Aretz 2002)
Family Aulophyllidae, Dybowski, 1873
Genus Axoclisia Semenoff-Tian-Chansky, 1974
Type species: Axoclisa cuspiforma, Semenoff-Tian-
Chansky, 1974
?Axoclisa sp. (Fig. 4i, k)
Material: Large fragment of a solitary coral from P6
(GIK 2184).
Description: The solitary coral is enclosed in a limestone
block. Its maximum diameter is 2.7 cm. There are two series
of 62 septa. Major septa are long and thickened at the base of
the tabularium. Only the cardinal septum (?) is attached to
the axial structure. The minor septa are very short and often
persist discontinuously in the outer dissepimentarium (septal
spines). The wall is thin. The axial structure (clisophyllid)
consists of a thickened axial plate, which is surrounded by
numerous lamellae and tabellae. Its maximum length is
1.0 cm. The dissepimentarium is up to 5 mm thick and very
irregular. Herringbone dissepiments are observed in the
outer rows, but also occasionally transeptal dissepiments
develop next to the wall. A longitudinal section, oblique to
the axial structure, shows numerous rows of globose to
elongated dissepiments. Tabulae are incomplete.
Discussion: The attribution of this single specimen is
not free from doubts. The axial structure is clearly cliso-
phyllid, but transeptal dissepiments are uncommon in
Clisiophyllum Dana, 1846. Semenoff-Tian-Chansky (1974)
established the genus Axoclisia for those specimens
between the two genera Clisiophyllum and Axophyllum,
thus having a clisophyllid axial structure and transeptal
dissepiments. The genus is so far monospecific and only
three specimens are known; the holotype from the Bechar
Basin (Algeria), and two specimens from SW Spain
(Rodrıguez et al. 2001b). The Jerada specimen differs from
these specimens described as Axoclisia cuspiforma Seme-
noff-Tian-Chansky 1974 in much shorter minor septa, a
higher number of septa-to-diameter ratio, less numerous
developed transeptal dissepiments, a considerably more
complicated dissepimentarium, and a larger axial structure.
A definitive attribution to the genus and/or the known
species requires more material.
Genus Dibunophyllum Thomson and Nicholson, 1876
Type species: Clisiophyllum bipartitum McCoy 1849
Dibunophyllum bipartitum (McCoy 1849) (Fig. 4l)
*1849 Clisiophyllum bipartitum McCoy: 2.
1992 Dibunophyllum bipartitum (McCoy).—Rodrıguez
and Falces: 193, pl. 17 fig. 1.
?1999 Dibunophyllum bipartitum (McCoy).—Liao and
Rodrıguez: 546, figs. 6.1–6.2.
2005 Dibunophyllum bipartitum (McCoy).—Aretz and
Nudds: 172, pl. 2 figs. 1–6. (cum. syn.)
2005 Dibunophyllum bipartitum (McCoy).—Cozar and
Somerville: figs. 12, 13.
2007 Dibunophyllum bipartitum (McCoy).—Rodrıguez
and Somerville: pl. 1 fig. 1.
2007 Dibunophyllum bipartitum (McCoy).—Aretz and
Nudds: pl. 1 fig. 2.
2008 Dibunophyllum bipartitum (McCoy).—Said and
Rodrıguez: 31, figs. 6g–i, 8.
Holotype: specimen SM A 1971, W. Hopkins Collec-
tion (chosen by Hill 1938).
Locus typicus and stratum typicum: Derbyshire, UK;
Lower Carboniferous.
Diagnosis: See Hill (1938) and Poty (1981)
Material: Single specimen from P4 (GIK 2185).
Description: The calice of this *6 cm long partly
eroded solitary coral is not preserved. Corallum curved in
early stages, upper part almost cylindrical. The maximum
diameter is 2.5 cm. A single transverse section is from the
mature stage of the corallite.
There are two series of 58 septa. The major septa are
long and reach towards the axial structure. They are
thickened at their base (forming parts of the wall?). An
open cardinal fossula is developed. Minor septa are very
Fig. 4 a–f Siphonophyllia samsonensis (Salee, 1913); a–e transverse
sections, f longitudinal section, all specimens 91, P4; a GIK 2163,
b GIK 2160, c GIK 2157, d GIK 2159, e GIK 2162, f GIK 2161.
g ?Zaphrentites sp.; transverse section, 93, GIK 2201. h Palaeosmiliamurchisoni Milne-Edwards and Haime, 1848; transverse section, 91,
P4, GIK 2181. i–k ?Axoclisia sp.; transverse sections, 92, P7, GIK
2184. l Dibunophyllum bipartitum (McCoy 1849); transverse section,
92, P4, GIK 2185. m–o Rylstonia sp.; m, n transverse sections, olongitudinal section, all sections 92, P8, GIK 2186
b
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 329
123

short, and persist only into the outer rows of dissepimen-
tarium. There are 6–8 rows of mainly irregular dissepi-
ments. The oval axial structure comprises about one-third
of the corallite diameter. It consists of a long medial plate,
five septal lamellae on each side, and numerous axial
tabellae.
Discussion: The specimen fits well into the character-
istic variability of this well-known species. Federowski
(1971) described a series of species from the same genus,
but they may either fall into the variability of D. biparti-
tum, as currently defined, or show marked differences to
the Jerada specimen in the diameter/septa ration, the
complexity of the axial structure, and the arrangement of
the dissepimentarium.
Distribution: The species is abundant in many late
Visean shallow water carbonate platforms of Europe.
Records for NW Africa are from the Bechar Basin and
Adarouch area (Semenoff-Tian-Chansky 1974; Said and
Rodrıguez 2008).
?Genus Rylstonia Hudson and Platt, 1927
Type species: Rylstonia benecompacta Hudson and
Platt, 1927
Rylstonia sp. (Fig. 4m–o)
Material: One specimen from P8 (GIK 2186).
Description: The ceratoid solitary coral is 43 mm high
and up to 19 mm in diameter. Calice, apex, and wall are
absent. There are up to 33 thick septa in two series. The
major septa are strongly thickened, especially in the car-
dinal half of the corallite. Minor septa are short. A marked
cardinal fossula is developed. The centre of the corallite is
composed of a prominent, compact axial structure to
which most major septa are fused. The axial structure is
5 mm long and 2–4 mm wide, and consists of a complex
amalgamation of an axial plate and axial lamellae. In
transverse section the dissepimentarium is 2.0–4.0 mm
thick and consists of 3–7 rows of steeply declined elon-
gated dissepiments. The tabulae are incomplete and axially
domed.
Discussion: This specimen shows similarities with the
specimens of the Bechar Basin attributed to this genus by
Semenoff-Tian-Chansky (1974). However, the character-
istic variability of the axial structure, which specimens
from the UK show, has not been observed in the Jerada
specimen.
Family Lithostrotionidae d’Orbigny, 1852
Subfamily Lithostrotioninae d’Orbigny, 1852
Genus: Lithostrotion Fleming, 1828
Type species: Lithostrotion striatum Fleming, 1828
(=Madrepora vorticale Parkinson, 1808).
Lithostrotion maccoyanum Milne-Edwards and Haime,
1851 (Fig. 5a)
*1851 Lithostrotion M’Coyanum Milne-Edwards and
Haime: 444.
2005 Lithostrotion maccoyanum Milne-Edwards and
Haime.—Aretz and Nudds: 174. (cum. syn.)
2005 Lithostrotion maccoyanum Milne-Edwards and
Haime.—Cozar and Somerville: fig. 12, 9.
Lectotype: specimen Z 87 illustrated by Milne-Edwards
and Haime (1852: pl. 17 fig. 2a), MNHNP; (chosen by
Semenoff-Tian-Chansky and Nudds 1979).
Locus typicus and stratum typicum: Oswestry, UK;
lower Carboniferous (probably Brigantian).
Diagnosis: See Semenoff-Tian-Chansky and Nudds
(1979)
Material: One colony and a fragment from P4 (GIK
2187, 2188).
Description: The maximal diameter of the colony is
25 cm and it is 10 cm in height. The shape of the colony is
two-folded into a bulbous lower half and a tabular upper
half. The epitheca is usually absent, but on the bottom of
the colony concentric growth indicates the growth centre.
The calicular pit is, on average, 1–2 mm deep, the calicular
boss consists of a single elongated axial plate. Attached
septa may rise towards the columella
Single corallites are of polygonal shape, with prefer-
ences of pentagonal to hexagonal corallites. There are 9–
12, average 10, septa in two series. The major and minor
septa are long. Major septa are often attached to the axial
plate, thus forming a spider-web structure. There are 3–7
rows of flat-lying regular to elongated relatively small
dissepiments. The diameter of the tabularium is less than
2 mm. Tabulae are incomplete and rise towards the axis.
Discussion: L. maccoyanum is the smallest cerioid
species of the genus in the West European faunal province,
and thus its recognition is relatively easy. The Jerada
specimens represent a morphotype with relatively small
corallite diameters within the variability of this species
described by Semenoff-Tian-Chansky and Nudds (1979),
Poty (1981) and Aretz (2002).
Distribution: The species is known from the late As-
bian-early Brigantian interval in NW Europe (Aretz and
Nudds 2005). In Morocco Said et al. (2007) listed it for the
Asbian of the Adarouch area.
Lithostrotion vorticale (Parkinson, 1808) (Fig. 5b)
*1808 Madrepora vorticalis Parkinson: 45, pl. 5,
figs. 3, 6.
2005 Lithostrotion vorticale (Parkinson).—Aretz and
Nudds: 175, pl. 2 fig. 8. (cum. syn.)
330 M. Aretz
123

2005 Lithostrotion vorticale (Parkinson).—Cozar et al.:
fig. 12/3
Types: The Parkinson specimens are regarded as lost. A
lectotype has not yet been chosen.
Diagnosis: See Poty (1981).
Material: One colony fragment from P1 (GIK 2189),
one colony fragment from P3 (GIK 2190), 18 colony
fragments (GIK 2191-2207) from P4, one from P5 (GIK
2208) and one from P7 (GIK 2209).
Description: A well preserved tabular colony enables
description of several outer characters. The corallum is
15 cm in diameter and 7 cm high. The growth is mainly
peripheral. The bottom surface shows a growth centre of
seven corallites. Usually the single corallites are of polyg-
onal shape, with preference of hexagonal outlines. Where
corallites are not in direct contact with each other corallite
walls become curved. This is not only seen on the bottom
surface of the corallum, but also in areas within the colony
where corallites were partly killed by sediment supply.
Especially in those latter areas corallites look like a Siphono-
dendron corallite. Fragments of further colonies show a
relatively deep calice surmounted by a thin axial structure.
There are two series of 18–21 major septa in mature
stages. Cardinal and counter septum are differentiated by
the orientation of the styliform columella, and are usually
attached to it. Further major septa are only occasionally
attached. Minor septa persist shortly into the tabularium.
Major and minor septa are more sinuous in the dissepi-
mentarium than in the tabularium. The tabularium is 3.0–
4.4 mm wide and consists of incomplete tabulae, which
increase towards the axis. The dissepimentarium consists
of 2–7 rows of globose to elongated dissepiments, which
are slightly declined. Transeptal dissepiments may occur in
the peripheral parts of corallites, especially in corners and
in offset structures.
Discussion: Based on morphometric criteria (e.g. width
of tabularium, number of septa) the Jerada specimens can
be easily placed in this species; L. maccoyanum and
L. decipiens (McCoy 1849) are smaller, L. araneum larger.
Distribution: In Europe the species extends from the
Holkerian/Livian to the Late Brigantian (Mitchell 1989,
Rodrıguez et al. 2002). In Morocco Said et al. (2007)
reported it from the Brigantian of the Adarouch area and
Semenoff-Tian-Chansky (1985) from the Visean of the
Algerian Bechar Basin.
Lithostrotion araneum (McCoy 1844) (Fig. 5c)
*1844 Astrea aranea McCoy: 187, pl. 27 fig. 6.
1981 Lithostrotion araneum (McCoy).—Poty: 20,
Figs. 10, 15, 16, pl. 4 figs. 1–4. (cum. syn.)
1992 Lithostrotion araneum (McCoy).—Rodrıguez and
Falces: 198.
2002 Lithostrotion araneum (McCoy).—Rodrıguez
et al.: 22, figs. 5, 8.
2005 Lithostrotion araneum (McCoy).—Cozar et al.:
fig. 12/4.
2005 Lithostrotion araneum (McCoy).—Cozar and
Somerville: fig. 12, 8.
2006 Lithostrotion araneum (McCoy).—Gallagher et al.:
fig. 13, 1.
Syntypes: specimen F7467/1-3, NMD. In Mitchell
(1989) specimen 50-1926 is called holotype.
Locus typicus and stratum typicum: Ireland; Lower
Carboniferous.
Diagnosis: See Poty (1981)
Material: Single specimen from P9 (GIK 2210).
Description: A fragment of a colony, which is 10 cm
wide and 2.5 cm high. Corallites are polygonal (up to
15 mm wide). There are, on average, 24 septa in two series.
The major septa are long, slightly to highly sinuous. The
may or may not attain the axial structure. In some corallites
major septa are fused next to the axial structure. Minor
septa persist a short distance into the tabularium. The axial
structure is a lenticular plate of variable thickness. The
orientation of the axial plate defines cardinal and counter
septum, which are otherwise not recognizable. The tabu-
larium is larger than 4.2 mm, and consists of incomplete
tabulae. Numerous rows of dissepiments are observed.
Discussion: This fragment has by far the largest values
for corallite diameter, number of septa, and tabularium
width of all recovered Lithostrotion specimens from
Jerada. It well fits into the morphological variability of
L. araneum described from Europe (Poty 1981, Rodrıguez
et al. 2002).
Distribution: In Europe L. araneum appears in the
middle Visean (Corphalie Member of the Lives Formation;
Livian) and continues into the Asbian. Said et al. (2007)
listed it for the upper limestone horizon of the Tizra
Formation.
Genus Siphonodendron McCoy, 1849
Type species: Siphonodendron pauciradiale (McCoy,
1844)
Siphonodendron junceum (Fleming, 1828) (Fig. 5d)
*1828 Caryophyllia juncea Fleming: 508.
2005 Siphonodendron junceum (Fleming).—Aretz and
Nudds: 176, pl. 2 figs. 9–10. (cum. syn.)
2007 Siphonodendron junceum (Fleming).—Rodrıguez
and Somerville: pl. 1 fig. 2.
Lectotype: specimen C4203, Ure Collection, HM.
Locus typicus and stratum typicum: Rutherglen,
Lanarkshire, UK; Lower Carboniferous.
Diagnosis: See Kato (1971) and Poty (1981).
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 331
123
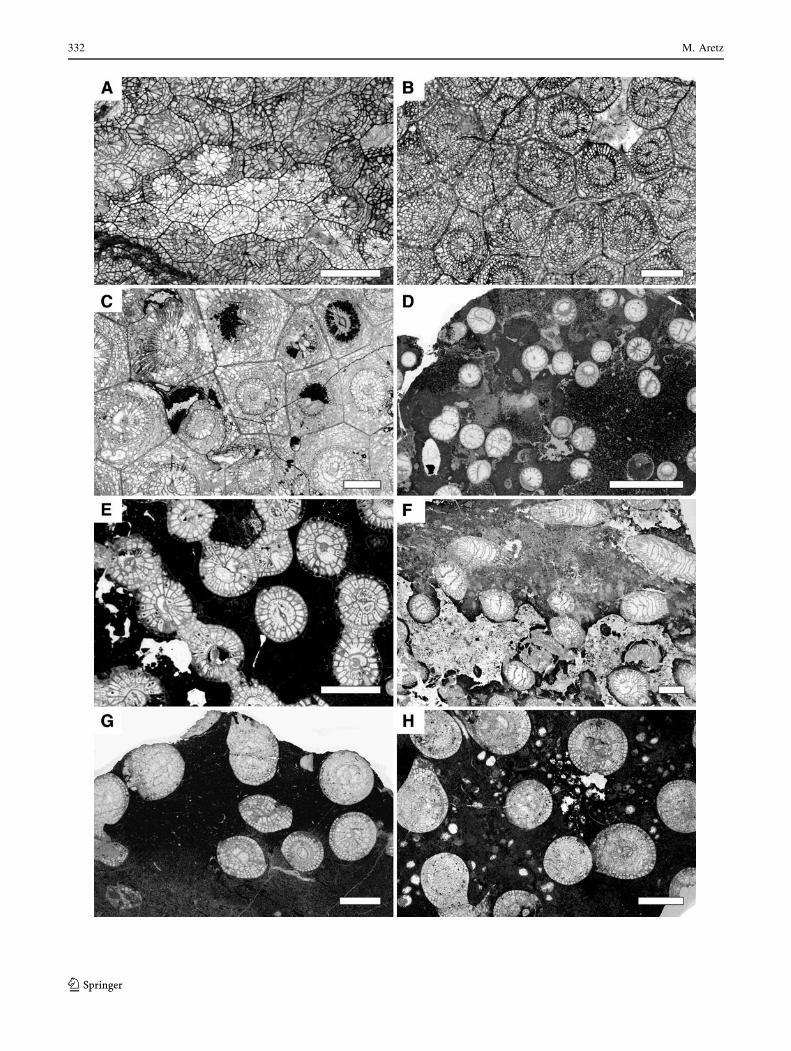
332 M. Aretz
123

Material: Nine colony fragments from P4 (GIK 2211-
2219).
Description: Fragments of phaceloid colonies up to
15 cm in length and 10 cm in height. The largest fragment
shows the existence of a spreading centre in the middle of
the corallum, thus indicating medial growth and a bulbous
colony shape. The distances between corallites are, in
general, up to few millimetres. The length of individual
corallites can attain several centimetres. The increase is
lateral.
Mature corallites are 2.0–3.2 mm in diameter, and there
are, on average, 13–14 major septa; the maximum value is
16. Minor septa, if developed, are very short spines. The
major septa are variable in length and thickness. However,
in many cases they leave an almost open space in the
central part of the corallite, and only cardinal and counter
septum may attach to the columella. The columella is a
single axial plate, but not present in all corallites. These
diphymorphic corallites are randomly distributed. A dis-
sepimentarium is not observed. The presence of a colu-
mella defines the morphology of the tabularium.
Incomplete tabulae generally rise convex towards the
columella. A depression of the tabulae is often developed
near the wall. In diphymorphic corallites the tabulae are
complete and arranged horizontally in the corallite centre.
Discussion: The Jerada specimens fit well into the
recorded variability of that taxon, which is the most
unusual Siphonodendron species, because of the lack of
dissepiments. As typical for many occurrences the intra-
specific variability in a single outcrop is very limited.
Compared with specimens from Royseux, Belgium, and
Little Asby Scar, Britain (Aretz 2002, Aretz and Nudds
2005) the corallite sizes at Jerada are slightly smaller, and
thus the average number of septa shifts to lower values.
Distribution: This species is characteristic of the late
Asbian and Brigantian of NW Europe and SW Spain (Poty
1981, Mitchell 1989, Rodrıguez and Somerville 2007). In
Morocco Said et al. (2007) listed it for the Adarouch area.
Siphonodendron pauciradiale (McCoy, 1844) (Fig. 5e)
*1844 Lithodendron pauciradialis McCoy: 189, pl. 27
fig. 7.
2002 Siphonodendron pauciradiale (McCoy).—Poty: pl.
1 fig. 9.
2005 Siphonodendron pauciradiale (McCoy).—Aretz
and Nudds: 176. (cum. syn.)
2005 Siphonodendron pauciradiale (McCoy).—Cozar
and Somerville: fig. 12, 10.
2007 Siphonodendron pauciradiale (McCoy).—Rodrı-
guez and Somerville: pl. 1 fig. 8.
Lectotype: specimen 82-1925, Griffith Collection,
NMD.
Locus typicus and stratum typicum: Magheramore,
Tobercurry, Co. Sligo, Ireland; Glencar Limestone For-
mation, Asbian.
Diagnosis: See Poty (1981).
Material: Single fragment from P3 (GIK 2220).
Description: The fragment of a phaceloid colony is
strongly weathered. The upper surface of the sample shows
well-separated cylindrical corallites, whereas the transverse
thin section shows many offsets and connections between
individual corallites. Corallites have an average diameter
of 4.0 mm (3.7–4.5 mm). There are two series of 20 major
septa (rarely 19–21). Major septa are variable in length.
Cardinal and counter septum may be attached to columella.
The minor septa are short in the tabularium. The columella
is an axial plate which may be thickened. There is one row
of globose dissepiments. The incomplete tabulae increase
towards the axis.
Discussion: The Jerada specimen is a rather large rep-
resentative within the reported species variability. The
single row of dissepiments is a sufficient character to dif-
ferentiate the Jerada specimens from small specimens of S.
irregulare and S. intermedium, which may have similar
number of septa and diameters, but have two rows of
dissepiments.
A badly preserved fragment from P4 (GIK 2221) shares
many characteristics of S. pauciradiale, but has inferior
corallite diameters and, often, a number of septa around the
lower limit for this species. However, because at least one
row of dissepiments is present and the largest and most
complete corallites are clearly within the normal size for the
species, the specimen is determinate as S. cf. pauciradiale.
Distribution: S. pauciradiale appears in Europe in the
early Asbian and persists into the Brigantian. Semenoff-
Tian-Chansky (1985) mentioned it from the late Visean of
the Bechar Basin, and Said et al. (2007) reported it from the
Brigantian of the Adarouch area.
Siphonodendron irregulare (Phillips, 1836) (Fig. 5f, g)
*1836 Lithostrotion irregulare Phillips: 202, pl. 2,
figs. 14, 15.
1852 Lithostrotion irregulare Phillips.—Milne-Edwards
and Haime: 198, pl. 11 fig. 1a–e.
Fig. 5 a Lithostrotion maccoyanum Milne-Edwards and Haime,
1851; transverse section, P4, GIK 2187. b Lithostrotion vorticale(Parkinson, 1808); transverse section, P3, GIK 2190. c Lithostrotionaraneum (McCoy 1844); transverse section, P9, GIK 2210. dSiphonodendron junceum (Fleming, 1828); transverse section, P4,
GIK 2212. e Siphonodendron pauciradiale (McCoy 1844); transverse
section, P3, GIK 2220. f Siphonodendron irregulare (Phillips, 1836);
transverse section, P4, GIK 2222. g Siphonodendron irregulare(Phillips 1836); transverse section, P2, GIK 2227. h Siphonodendronmartini (Milne-Edwards and Haime, 1851); transverse section, P4,
GIK 2229. All scale bars = 5 mm
b
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 333
123

e.p. 1940 Lithostrotion pauciradiale McCoy.—Hill: 169.
?1962 Lithostrotion cf. pauciradiale (McCoy).—Cald-
well and Charlesworth: 377, pl. 14 fig. 4.
1981 Siphonodendron irregulare (Phillips).—Poty: 30,
pl. 12 figs. 1–4.
1986 Siphonodendron irregulare (Phillips).—Herbig:
202, Abb. 4, Abb. 11 figs. 1, 5.
1992 Siphonodendron irregulare (Phillips).—Rodrıguez
and Falces: 203, pl. 19, figs. 1–3.
1997 Siphonodendron martini ecotype 2; Nudds and
Day: 96, fig. 1.
2002 Siphonodendron irregulare (Phillips).—Aretz:
112, pl. 1 fig. 8, pl. 5 fig. 4, pl. 10 fig. 4.
2002 Siphonodendron irregulare (Phillips).—Rodrıguez
et al.: 26, fig. 13a.
2002 Siphonodendron irregulare (Phillips).—Poty: pl. 1
fig. 8.
2005 Siphonodendron sp.—Aretz and Nudds: 178, pl. 3
fig. 1.
2007 Siphonodendron sp.—Aretz and Nudds: pl. 1
fig. 4.
Type material: The specimens of Phillips are regarded
as lost. Poty (1993) described topotypic material from the
locality Ashfell, Cumbria.
Locus typicus and stratum typicum: There is some
confusion on this issue, because Phillips did not indicate a
single locality, and those named contain different Si-
phonodendron species (Hill 1940). If the definition of Poty
(1981, 1993) is accepted, the Middle Visean exposed in the
road cut next to Ashfell, Cumbria, is the type area.
Diagnosis: See Poty and Hannay (1994).
Material: Single colony fragment from P2 (GIK 2222),
and six fragments from P4 (GIK 2223-2228).
Description: Phaceloid corallum with long cylindrical
corallites. Two types of corallite sizes can be differentiated.
The smaller type has corallite diameters of 4.6–6.2 mm and
there are two series of 23–24 septa. The larger type com-
prises specimens with corallite diameters 5.8–6.5 mm and
septal numbers of 21–23.
Both types have a thin wall. The major septa are thin and
straight or sinuous. Some attain a simple axial plate. Its
orientation distinguishes the cardinal and counter septum.
The minor septa are short and persist into the tabularium.
The dissepimentarium consists of one or two rows of
regular dissepiments. In the specimen from P2 diphymor-
phic stages are observed.
Discussion: Aretz and Nudds (2005) discussed the main
problems of the correct specific affiliation of Siphonoden-
dron specimens between the two well defined species
S. pauciradiale (McCoy 1844) and S. martini (Milne-Edwards
and Haime 1851). In this study S. irregulare (Phillips, 1836) is
used as defined by Poty and Hannay (1994).
In the case of the small type it differs from S. martini by
its smaller size, from S. pauciradiale by its larger size and
the second rows of dissepiments, and from S. intermedium
Poty, 1981 by the long major septa attaining the axial plate.
The larger type has the corallite diameter of typical S.
martini, but the number of septa is much lower. The Jerada
specimens do not show an isolated axial plate as commonly
observed in S. intermedium.
Distribution: Because of the confusion on the correct
specific affiliations, the precise occurrence of these species
is uncertain. In Europe and North Africa the name has been
used for specimens of middle and late Visean age.
Siphonodendron martini (Milne-Edwards and Haime
1851) (Fig. 5h)
*1851 Lithostrotion martini Milne-Edwards and Haime:
436.
2005 Siphonodendron martini (Milne-Edwards and
Haime).—Aretz and Nudds: 178, pl. 3 fig. 2. (cum. syn.)
Lectotype: specimen E 1446, Phillips Collection, UMO.
Locus typicus and stratum typicum: Yorkshire, UK,
possibly Teesdale or Ribblesdale; Lower Carboniferous.
Diagnosis: See Semenoff-Tian-Chansky and Nudds
(1979).
Material: One colony fragment from P4 (GIK 2229).
Description: Phaceloid corallum with long cylindrical
corallites. The corallite diameter is 6.0–7.3 mm. There are
23–25 septa in two series. Major septa are long, cardinal
and counter septum attain the styliform columella. The
latter is a single axial plate thickened in its central part. The
minor septa are 1/5 to 1/2 as long as the major septa, often
only entering the tabularium. The septa are straight to
slightly sinuous. There are 2–3 rows of regular globose to
elongated dissepiments. Tabulae are tent-shaped, incom-
plete and in two series.
Discussion: The Jerada specimen is a relatively small
representative within the variability of this species (Se-
menoff-Tian-Chansky and Nudds 1979, Poty 1981).
However, it also shares many similarities with specimens
of S. irregulare from the same locality, except its corallite
size and size-related characters.
Distribution: S. martini is perhaps the most common
species of Siphonodendron in the middle and late Visean of
Europe as Aretz and Nudds (2005) stated. Records in NW
Africa are so far from the late Visean (Semenoff-Tian-
Chansky 1985, Said et al. 2007).
Subfamily Diphyphyllinae Dybowski, 1873
Genus Tizraia Said and Rodrıguez, 2007
Type species: Tizraia berkhlii Said and Rodrıguez,
2007
334 M. Aretz
123

Tizraia berkhlii Said and Rodrıguez, 2007 (Fig. 6a–d)
?2005 Lublinophyllum sp.—Cozar and Somerville:
figs. 12, 6–7.
*2007 Tizraia berkhlii.—Said and Rodrıguez: 27,
figs. 3–8.
Holotype: specimen DPM-TIZ2/5-6.
Locus typicus and stratum typicum:, Adarouch area,
Morocco; upper beds of the Tizra Formation, upper Visean
(Brigantian).
Diagnosis: See Said and Rodrıguez (2007).
Material: Two fragments, one from P7 (GIK 2230), one
from P10 (GIK 2231).
Description: The fragments are from phaceloid colonies.
The increase is parricidal and may occur axial and lateral.
Mature corallites are 7–12 mm in diameter (mean 8.8 mm),
and there are 30–32 major septa; maximum value is 33.
The variably thick corallite wall is generally undulose, but
may also be smooth. The lengths and thickness of the
septa are variable, but they always leave an open space in
the axis of the corallite. Sinuosity of septa is higher in the
dissepimentarium with no difference between major and
minor septa. In general there is a tendency of thickened
septa in the dissepimentarium and/or at the limit of the
dissepimentarium/tabularium. However, in single corallites
there is no thickening at all or it occurs in the tabularium.
Lengths of major septa are 1/3 to 3/4 of the corallite diam-
eter. Minor septa are comparable, variable, and, in general,
1/3 to 1/2 of the major septa long. The dissepimentarium is
variable. It mainly consists of regular dissepiments, but
transeptal dissepiments of both orders regularly occur.
These transeptal dissepiments rarely form a complete circle
in the outer tabularium. There are mature corallites without
any trace of transeptal dissepiments and others with only one
or two transeptal dissepiments of 2nd order.
In the longitudinal section there are 3–4 rows of small
globose to elongated dissepiments forming a relatively thin
dissepimentarium. The tabularium mainly consists of
dome-shaped incomplete tabellae and outer inclined
smaller peripheral tabellae.
Discussion: The Jerada specimens slightly enlarge the
species variability described by Said and Rodrıguez (2007).
Especially the inconsistency of the transeptal dissepiments
and the variability of the septal shapes are noteworthy. This
clearly points to the need for detailed comparison of the
variabilities in Tizraia and Lublinophyllum Khoa, 1977,
because at least some characters used to differentiated
these two genera (Said and Rodrıguez 2007) may fall into
the intra-specific variability. Kubassophyllum Dobrolyub-
ova in Dobrolyubova et al. (1966) should also be included,
because it has many similarities with the genera under
question.
Distribution: Only the type material from the Brig-
antian of the Adarouch area and the two Jerada specimens
of this study can so far be surely assigned to Tizraia
berkhlii. Said and Rodrıguez (2007) mentioned the exis-
tence of specimens in the Jerada Basin in the Djerada
Formation (Eastern Morocco). The term Djerada Forma-
tion might be deduced from older work concerning the
Jerada Coal Basin (Owodenko 1946, 1976), but conse-
quently it must be used for the productive coal seams of
the Westphalian age, where no carbonates are recorded.
Carbonates and also corals are restricted to the interval
Oued El-Koriche Formation to Koudiat Es-Senn Forma-
tion (= upper Visean). Hence, it can be concluded that
Said and Rodrıguez (2007) intended to indicate this
stratigraphic interval.
Subfamily Aulininae Hill, 1981
Genus Aulokoninckophyllum Sando, 1976
Type species: Campophyllum carinatum Carruthers,
1909.
Aulokoninckophyllum carinatum (Carruthers, 1909)
(Fig. 6e, f)
*1909 Campophyllum carinatum.—Carruthers: 150, pl.
1 figs. 3–6.
1998 Aulokoninckophyllum carinatum (Carruthers,
1909).—Niko and Yamagiwa: 139, fig. 7.
2001 Aulokoninckophyllum carinatum (Carruthers,
1909).—Rodrıguez et al.: 90, figs: 3, 4. (cum. syn.)
Holotype: syntypes 1954.6.8-12, RSM (according to
Semenoff-Tian-Chansky 1974).
Locus typicus and stratum typicum: Cap Chernyi,
Nowaja Semlja, Russia; upper Visean.
Diagnosis: see Rodrıguez et al. (2001a)
Material: Four specimens from P4 (GIK 2232-2235),
one specimen from P8 (GIK 2236).
Description: Fragments trochoid solitary corals, which
lack apex, calice, and a large part of the wall. Maximum
height is 3.0 cm, maximum diameter is 18 mm. Areas of
intact wall show the presence of 1 mm thick horizontal
bands possibly growth bands. There are 36–47 septa of two
series. Major septa are up to 7 mm long, and thus leave an
open central space. The minor septa are 3–5 mm long.
They persist some mm or as short septal spines into the
tabularium. All septa are sinuous, carinate (zigzag), and
thickened in the dissepimentarium. A cardinal fossula is
developed; the cardinal septum is about 3/4 as long as the
other major septa. The outer part of the dissepimentarium
is irregular and rarely contains transeptal dissepiments. In
the longitudinal section there are numerous rows of glo-
bose to slightly elongated dissepiments. The tabulae are
incomplete. An aulos is discontinuously developed. In the
axial area/aulos they are almost horizontal, and axial
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 335
123
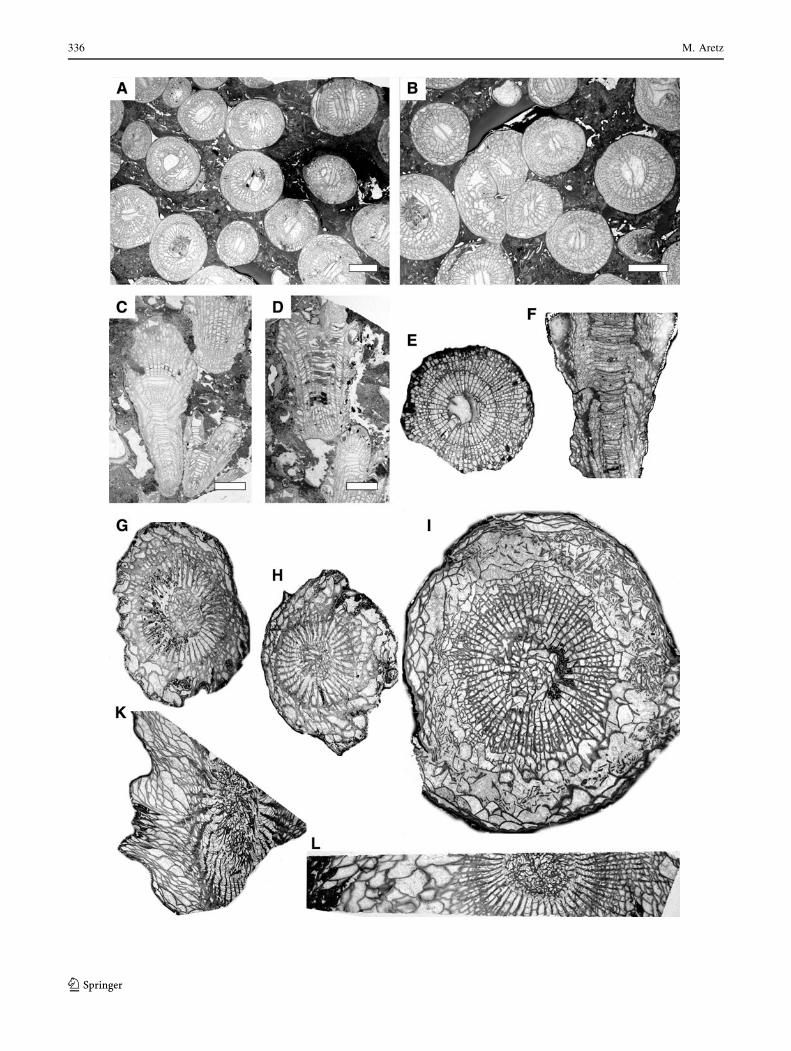
336 M. Aretz
123

tabellae occasionally occur; in the peripheral zone they are
replaced by declined or curved tabellae.
Discussion: The Jerada specimens fit well into the
variability described by Rodrıguez et al. (2001a). The
largest specimen has a diameter/septa ratio comparable
with that of the monotypic A. amarensis (Semenoff-Tian-
Chansky 1974), but the latter differs by fewer axial tabellae
and a narrower dissepimentarium.
Distribution. The species is known from the late Vis-
ean—Serpukhovian of Europe, Asia, and North Africa,
possibly also Queensland (Hill 1981).
Suborder Lonsdaleiina Spasskiy, 1974
Family Axophyllidae Milne-Edwards and Haime, 1851
Genus Axophyllum Milne-Edwards and Haime, 1850
Type species: Axophyllum expansum Milne-Edwards
and Haime, 1850
Axophyllum aff. pseudokirsopianum Semenoff-Tian-
Chansky, 1974 (Fig. 6g, h)
Material: One specimens from P6, GIK 2237.
Description: A fragment of a solitary corals, the rela-
tively thick wall is only partly preserved. The diameter is
about 2.5 cm. There are two series of 42 septa. The major
septa are long and slightly sinuous. The minor septa are not
more than half as long as the major septa, and only persist
few mm into the tabularium. The axial structure comprises
about one-third of the corallite. It consists of a slightly
sinuous partly thickened axial plate, several thickened,
curved axial lamellae, and somewhat thinner but also
curved axial tabellae. The dissepimentarium comprises
large transeptal dissepiments of two orders, which are
elongated and declined in the longitudinal section. Tabulae
are incomplete.
Discussion: The Jerada specimen is a typical larger
sized Axophyllum. It differs from A. pseudokirsopianum
Semenoff-Tian-Chansky, 1974 by its shorter minor septa,
and a slightly larger dissepimentarium. A large dissepi-
mentarium is characteristic for A. parkinsoni (Ryder,
1930), but that of the Jerada specimen is less thick. A
similar length of minor septa is known from A. kirsopia-
num (Thomson, 1880), but naotic septa have not been
observed as in the latter.
Genus Pareynia Semenoff-Tian-Chansky, 1974
Type species: Pareynia splendens Semenoff-Tian-
Chansky, 1974
Pareynia splendens Semenoff-Tian-Chansky, 1974
(Fig. 6i, k, l)
*1974 Pareynia splendens splendens Semenoff-Tian-
Chansky: 241, Figs. 93, 94, 96, pl. 62 fig. 7, pl. 63 figs. 1,
2, pl. 64 figs. 1–4, pl. 65 figs. 2, 3, pl. 73 figs. 1, 2.
?1974 Pareynia gangamophyllides Semenoff-Tian-
Chansky: 248, Fig. 95, 96, pl. 63 fig. 3, pl. 65 fig. 4,
pl. 66 fig. 1-6, pl. 70 fig. 1, 2, pl. 74 fig. 1-4.
cf. 1978 Pareynia splendens Semenoff-Tian-Chansky.—
Poty in Kimpe et al.: pl. 6 fig. 6.
1981 Pareynia splendens Semenoff-Tian-Chansky.—
Poty: 64, pl. 31 figs. 1, 2.
?1998 Pareynia? sp. indet.—Niko and Yamagiwa: 141,
fig. 8.
Holotype: specimen PAR 382/93. Semenoff-Tian-
Chansky collection, MNHNP.
Locus typicus and stratum typicum: Piton a l’Est du
Meharez el Kebir, Bechar Basin, Algeria; Ioucha 21
Member, upper Visean.
Diagnosis: (Semenoff-Tian-Chansky 1974 and Poty
1981). Pareynia with large dissepimentarium, which may
comprise up to the half of the corallite; short, but well
developed minor septa; variable axial structure, especially
the thickness of the axial plate; thin to medium-thick wall
present; variable corallite diameter throughout growth
(common rejuvenesence?); lateral increase possible.
Material: One specimens from P6 (GIK 2238), two
from P7 (GIK 2239-2240).
Description: The large solitary corals are completely
surrounded by calcareous matrix, thus the external char-
acters of the specimens are not observed. The maximum
diameter is 5.6 cm, however during growth it strongly
varies between zone of extension and contraction, resulting
in a very irregular corallite shape. A thin to medium-thick
undulating wall is present, but may be less visible in the
areas where the growth style changes; additionally these
areas are preferentially eroded.
There are two series of up to 55 septa. The major septa
are long and slightly sinuous. In the outer dissepimentari-
um they are interrupted by transeptal dissepiments of 1st
order. The maximum length of minor septa is half that of
the major septa. The axial structure is about 1 cm in
diameter. There are numerous tabellae and strongly curved
axial lamellae. The thickness of the central axial plate is
variable, thus resulting in some sections in a gangamo-
phyllid appearance of the axial structure. The dissepi-
mentarium can be separated in a vast outer zone of
Fig. 6 a–d Tizraia berkhlii Said and Rodrıguez, 2007; a, b transverse
sections, GIK 2230, c, d longitudinal sections, all scale bars = 5 mm,
P7, GIK 2231. e, f Aulokoninckophyllum carinatum (Carruthers,
1909); transverse sections, 92, P8, GIK 2236. g, h Axophyllum aff.
pseudokirsopianum Semenoff-Tian-Chansky, 1974; transverse sec-
tions, 92, P6, GIK 2237. i, k, l Pareynia splendens Semenoff-Tian-
Chansky, 1974; i, l transverse sections, 92, I: GIK 2239, l GIK 2240,
k longitudinal section, 91, P7, GIK 2240
b
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 337
123

transeptal dissepiments of both orders, and an inner dis-
sepimentarium of 3–5 rows of regular dissepiments.
Tabulae are incomplete, clinotabulae are developed. Ele-
ments of the axial structure are much thicker than dissep-
iments and tabulae. Tabellae rise towards the axis.
Discussion: The genus Pareynia is rarely reported and
thus the morphological variabilities are poorly known. The
irregular growth observed in the Jerada specimens combined
with partly eroded areas of the corallites may explain some of
the difficulties with interpretation of this genus. However, the
Jerada specimens significantly increase the likelihood that
the axial structure is more variable, especially the thickness
and shape of the axial plate, as currently stated for the genus.
Hence P. gangamophylloides may fall in the variability of
P. splendens. For the same reason, use of subspecies defined
by Semenoff-Tian-Chansky (1974) is questionable.
Occurrence: Upper Visean of Algeria, Belgium and
probably Japan.
Discussion
Biostratigraphic position
Owodenko (1946, 1976) dated the studied interval as late
Visean based on brachiopods and ammonoids. Vachard and
Berkhli (1992) used carbonate microfossils and concluded
an almost exclusive Brigantian age with a possible Asbian
age for the base of the succession, and a Serpukhovian age
for a 10 m thick limestone package in the Oued Agaia area
(SW end of synclinorium). The corals collected in this
study are used to discuss these ages and to indicate pos-
sibilities for refinements (Fig. 7).
The basal volcanic rocks cannot be dated by biostrati-
graphic tools; the primitive carbonate microfossils
(Pachysphaerina and Diplosphaerina) found in thin pack-
ages of intercalated calcareous shales (Vachard and Berkhli
1992, Berkhli et al. 1999) do not have a biostratigraphic
value. The Siphonodendron irregulare colony recovered
from the lower third of the Cafcaf Formation on the
northern flank indicates a maximum age of middle Visean
(Livian/Holkerian) and a minimum age of late Visean.
Most corals recorded from the succeeding formations have
limited biostratigraphic potential. They are all known from
late Visean coral faunas, but first appearance dates (FADs)
are often uncertain. It seems to be reasonable to assume at
least a middle Visean age for the Cafcaf Formation,
especially because the accumulation of bedded cherts in
that thickness (up to 40 m), requires a substantial amount
of time for deposition. The middle/late Visean boundary is
hard to trace and might still be placed in the bedded chert
facies.
The age of the fauna of the Oued El-Koriche Formation
is not unequivocal. The biostratigraphic range of Litho-
strotion vorticale is rather large (Holkerian–Brigantian).
Lithostrotion araneum is known in the Holkerian–Asbian,
but not from the Brigantian. Tizraia is so far only known
from the Brigantian (Said and Rodrıguez 2007). Thus, in
any case, the biostratigraphic range of Lithostrotion ara-
neum or Tizraia has to be extended.
The fauna of the Oued Es-Sassi Formation contains
Aulokoninckophyllum carinatum, which is late Asbian/
Brigantian in age. The composition of the coral faunas in
the entire Koudiat Es-Senn Formation indicates at least an
Asbian age. A clear late Asbian FAD has Dibunophyllum
bipartitum found in the upper Koudiat Es-Senn Formation
Fig. 7 Distribution of rugose
corals in the Visean succession
and resulting stratigraphic
divisions, with comparison with
ages based on carbonate
microfossils (Vachard and
Berkhli 1992; Berkhli et al.
1999). HOL, Holkerian; LAS,
lower Asbian; UAS, upper
Asbian; AS, Asbian; LBR, lower
Brigantian; UBR, upper
Brigantian; ARN, Arnsbergian
(E2)
338 M. Aretz
123

(Aretz and Nudds 2005). Pareynia is a taxon, which is
generally thought to appear ‘‘high’’ in the late Visean
(Brigantian) (Poty 1981), but its FAD is very uncertain
because of its rarity. Two species recovered from the
youngest levels in the Koudiat Es-Senn Formation,
Siphonodendron junceum and Lithostrotion maccoyanum,
have a late Asbian FAD and both persist into the Brigan-
tian. L. maccoyanum is unknown from the upper Brigantian
in NW Europe (Aretz and Nudds 2005). Note that Rodrıguez
and Somerville (2007) do not differentiate the Brigantian in
their distribution pattern. Thus if the temporal distribution of
these two species in NW Africa is the same as in NW Europe,
the top of the Koudiat Es-Senn Formation is not older than
late early Brigantian.
The interpretation of these coral data is critical, because
diagnostic taxa are rare (Fig. 7). However, it is obvious that
taxa characteristic for many Brigantian faunas (e.g. colo-
nial axophyllids, Palastrea, Corwenia; Mitchell 1989,
Rodrıguez and Somerville 2007) do not occur in the faunas
collected in the Jerada Basin. Their absence might be best
explained by three possibilities:
1. so far these taxa have been overlooked;
2. the succession is mainly older than the Brigantian; or
3. an impoverished fauna has been recovered, which
lacks the typical marker.
None of these three arguments can definitely be ruled
out, but it must be stressed that those taxa indicating a
Brigantian age, for example Tizraia and Pareynia, are both
only reported from few localities. Additionally their
stratigraphic range is strongly questionable, because their
occurrence might be strongly facies-controlled (see below).
Hence a conservative age model based on the corals
(Fig. 7) indicates as possible age for the Cafcaf Formation
the Middle Visean and possibly basal late Visean, early
and/or, late Asbian for the Oued El-Koriche and Oued Es-
Sassi formations, and late Asbian and/or early Brigantian
age for the Koudiat-Es-Senn Formation. The hiatus around
the Visean/Serpukhovian boundary would at least comprise
the upper Brigantian and the basal Serpukhovian.
This interpretation contradicts the data of Vachard and
Berkhli (1992) and Berkhli et al. (1999), and thus either the
postulated ranges for corals or carbonate microfossils have
to be reconsidered (Fig. 7). This is of special importance
because Jerada is the key region for the Brigantian (Cfm 7)
in the Moroccan biozonation of carbonate microfossils
(Vachard in Berkhli et al. 1993).
New ammonoid data of Korn and Ebbighausen (2008)
from pelites in the Chebket el Hamra area, in the NE of the
Jerada synclinorium, partly help to elucidate this problem.
These data are from a somewhat deeper and almost car-
bonate-free part of the basin. A basal late Asbian assem-
blage is followed by four assemblages of early and late
Brigantian age, thus showing that at least in deeper parts of
the basin sedimentation continued into the later Brigantian.
The stratigraphic correlations into the shallow parts of the
basin are so far not well constrained, because of tectonic
complication and the partial absence of good lithostrati-
graphic and biostratigraphic marker horizons. However, the
succession studied by Korn and Ebbighausen (2008) is
lithologically similar to the interval Cafcaf to Oued
Es-Sassi formations, the absence of limestone beds makes
the Koudiat Es-Senn Formation unrecognizable.
The coral fauna recorded from the Adarouch area (Said
et al. 2007, Said and Rodrıguez 2007, 2008) is more
diverse than those at Jerada, but especially the assemblages
from the lower part of that succession (Tizra Formation)
show a comparable composition. Said et al. (2007) indi-
cated an Asbian age for most of the Tizra Formation (the
uppermost beds are Brigantian in age), and thus these
results possibly strengthen the coral-based interpretation at
Jerada. It is important to mention that L. maccoyanum is
restricted to the Asbian (Said et al. 2007), and that the late
Visean-Serpukhovian succession of the Adarouch area
contains similar depositional environments as those of the
Jerada Massif.
However, all these considerations have to take into
account that the Visean succession of the Jerada Massif
comprises very different depositional environments. Facies
dependencies of the studied organisms are inevitable, and
may largely control the biostratigraphic interpretation.
The composition of the coral faunas recovered from the
individual outcrops clearly shows these facies dependen-
cies (Table 1), and these must also be suspected for groups
such as carbonate microfossils and ammonoids.
Facies
The smallest coral individuals, Aulokoninckophyllum,
Rylstonia and ?Zaphrentitis, have been found in shales of
the Oued Es-Sassi Formation (P8). A specialised fauna,
including Pareynia, Tizraia, Palaeosmilia and ?Axoclisia,
is known from the microbial buildup environments on the
southern flank of the synclinorium (P6, 7, 10). So far all
known occurrences of Tizraia are restricted to this partic-
ular facies. The same can be suspected for Pareynia. In its
type area, Bechar Basin, it was described (Semenoff-Tian-
Chansky 1974) from a region dominated by large sponge-
fenestellid bryozoan buildups (Bourque et al. 1995) with
abundant microbial boundstones, and also the Belgian
specimens (Poty 1981) are very likely to be from the same
kind of environment (Aretz and Chevalier 2007).
A marked differentiation in the composition of a coral
fauna is known from the uppermost Koudiat Es-Senn
Formation (P4) on the northern flank. Here, mass occur-
rences of Siphonophyllia samsonensis are only found in the
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 339
123

vicinity of a coral patch reef which is mainly formed by
Lithostrotion vorticale. Associated is a diverse coral fauna,
which comprises several species of Siphonodendron and
Lithostrotion, and larger solitary corals as Palaeosmilia
and Dibunophyllum. The small solitary corals (Au-
lokoninckophyllum and ?Zaphrentites) recovered from P4
are most probably associated with calcareous shales locally
intercalated and also present some meters below the patch
reef, hence confirming the observations from the Oued Es-
Sassi Formation. The patch reef formed next to an oolitic
shoal which contains Siphonodendron pauciradiale and
Lithostrotion decipiens (P3). The composition of the fauna
in P1 has some resemblance to that of P4, especially the
dominance of S. samsonensis and L. decipiens, but fewer
specimens were collected, and (thus) the fauna is less
diverse. However, the presence of a further coral patch reef
of the type found at P4 may be envisaged for P1.
In P2, P5, and P8 all corals are reworked and represent
more shallow facies than the depositional environment they
are embedded in. Their distribution pattern is random and
not controlled by any ecological preferences.
Overall, the compositions of assemblages, in particular
facies, show that large solitary corals and most colonial
taxa have a preference for shallow and turbulent water
whereas smaller corals represent unfavourable conditions
(siliciclastic input, deeper water?).
Palaeobiogeography
The Jerada fauna comprises taxa similar to the coral faunas
described from the late Visean of the Eastern Central
Meseta, which display comparable depositional environ-
ments (Said et al. 2007; Aretz and Herbig 2009). Differ-
ences mainly result from the total number of samples for
individual localities and slightly different ages. Although
the fauna of the Bechar Basin (Semenoff-Tian-Chansky
1974, 1985) is only partly well studied, the available data
indicate a substantial overlap with the Jerada fauna. The
later might be characterised as an impoverished Bechar
fauna. A new aspect for Morocco is the presence of
Pareynia and ?Axoclisa in the Jerada fauna, which have
been described in NW Africa so far only from the Bechar
Basin (Semenoff-Tian-Chansky 1974). Thus, in the late
Visean the Jerada fauna can be interpreted as a link
between the coral faunas of the Central Sahara basins and
the region north of the Central Atlas Fault in Morocco.
Table 1 Overview of the
distribution of rugose corals in
the different late Visean
environments of the succession
in the Jerada Massif
Environment Coral Taxa Formation
Coral patch-reef (core) Lithostrotion vorticale possibly
further lithostrotionids
Koudiat Es-Senn
Formation (P 4)
Coral patch-reef (surroundings and off-reef
facies)
Siphonophyllia samsonensis
Palaeosmilia murchisoni
Dibunophyllum bipartitum
Lithostrotion maccoyanum
Lithostrotion vorticale
Siphonodendron junceum
Siphonodendron irregulare
Siphonodendron martini
Koudiat Es-Senn
Formation (P1, 4)
Oolitic shoal Siphonodendron pauciradiale
Lithostrotion vorticale
Koudiat Es-Senn
Formation (P3)
Silty shale Aulokoninckophyllum carinatum
Rylstonia sp.
Zaphrentites sp.
Oued Es-Sassi
Formation (P8)
Microbial-sponge buildup (core and capping
beds) and microbial-metazoan buildup
Palaeosmilia murchisoni
?Axoclisia sp.
Lithostrotion vorticale
Tizraia berkhlii
Axophyllum aff.
pseudokirsopianum
Pareynia splendens
Koudiat Es-Senn
Formation (P6, 7)
Oued El-Koriche
Formation (P10)
Calciturbidite Lithostrotion vorticale
Lithostrotion araneum
Oued El-Koriche
Formation (P5, 9)
Bedded chert (turbiditic layer?) Siphonodendron irregulare Cafcaf Formation
(P2)
340 M. Aretz
123

Comparison with regions such as Anti-Atlas, western
Central Meseta, and Jebilet are so far impossible because
of the absence of solid data from these regions.
The Jerada fauna comprises many taxa, which are (well-)
known in NW Europe north of the Variscan Orogen. Its
composition clearly indicates its belonging to the West
European faunal province which includes North Africa
(Sando 1990). Comparison of the faunas from NW Europe,
SW Spain, and NW Africa shows that the Eastern Moroccan
Meseta links NW Europe with the central Saharan basins.
Hence the appearance of ‘‘exotic’’ taxa in NW Europe, for
example Pareynia in the Vise area (Poty 1981) or Axoclisia
in SW Spain, might be explainable by migration along a
pathway through the Eastern Moroccan Meseta.
The Jerada material cannot help elucidation of the
relationships of the West-European province to palaeoge-
ographic units further east (circum-Mediterranean, Minor
Asia), because contemporaneous faunas of these regions
are unknown or too poorly studied. However, contempo-
raneous European faunas from the southern side of the
Variscan Orogen (Herbig 1986; Aretz 2002) have a dif-
ferent composition, and contain taxa, for example Kizilia,
which are literally absent from NW Africa. Thus, in the late
Visean an important palaeobiogeographic barrier hampered
faunal exchange between NW Africa and regions south of
the Variscan Orogen, today neighboured as the Betic
Cordillera. The present geographic position may be partly a
result of Late Variscan tectonic activity along important
shear zones (Vai 1991).
Conclusions
1. The lithologically variegated succession of the Late
Visean of the Jerada Massif contains a diversified
rugose coral fauna. It comprises well-known and
geographically widely distributed taxa, for example
Palaeosmilia murchisoni or Siphondendron martini,
but also rare taxa, for example Pareynia splendens and
Tizraia berkhlii.
2. The fauna of the Adarouch area is similar to those of
the herein studied fauna from the Jerada Basin with
regard to composition and age. However, the Jerada
fauna is less diversified, which may be partly the result
of significantly fewer collected specimens.
3. The biostratigraphic ages of the Visean succession
(Cafcaf–Koudiat Es-Senn formations) are difficult to
establish because of missing or poor fauna and facies
changes and dependencies. Carbonate microfossils and
possibly also ammonoids indicate a Brigantian age at
least for parts of the succession. However, the coral-
based biostratigraphic interpretation shows some
differences from these ages. Typical Brigantian taxa,
for example colonial axophyllids and Palastrea, are
absent, and, consequently, coral ages are slightly older.
4. Overall an improved and more detailed biostrati-
graphic framework is needed for NW Africa to
precisely date the first appearance dates of coral taxa.
The facies dependencies described in this study clearly
demonstrate possible complications, but also may help
to explain differences between specific regions in NW
Africa.
5. The appearance of most taxa can be correlated with
distinctive facies. Small solitary corals occur in silty
shales, locally calcareous, whereas larger solitary
corals are found in limestones surrounding a coral
patch reef and on top of large microbial-sponge
buildups. The massive colonial coral Lithostrotion
vorticale is the main framework builder of that small
coral patch reef, which is a relatively unique feature in
Visean reefs. Overall, colonial corals occur in very
different environments. Their higher abundance in
buildup facies at Jerada is linked with the predomi-
nance of this facies as the main carbonate facies in
many parts of the Jerada Basin.
6. Palaeobiogeographically, the fauna recovered at Jerada
is part of the Western European coral province (Sando
1990), which comprises NW Africa. The presence of
taxa, for example Pareynia, Tizraia, and ?Axoclisia,
which are commonly not found in NW Europe,
indicate the position and importance of the Eastern
Moroccan Meseta as a pathway between the Central
Saharan basins and NW Europe. However, more coral
data from NW Africa are needed to reveal the precise
timing and directions of migration patterns.
Acknowledgments This study benefited from the help of H.-G.
Herbig (Koln), D. Korn (Berlin), V. Ebbighausen (Odenthal), and
E. Poty (Liege). Special thanks to Sebastian, Stefan, Arne, and Florian
(all Koln) for the corals they collected during their mapping projects.
A. Mihli and J. El Mokhtari (Direction Regional du Ministere de
l’Energie et des Mines a Oujda) are thanked for permission to work in
the Jerada Basin in the autumns of 2004 and 2006. All officials from
the ‘‘Eau et Foret’’ authority in Jerada are greatly acknowledged for
their hospitality and help during our field campaigns. R. Baumler
(Koln) prepared the thin sections for this study.
Appendix
Location of sample localities based on the topographic
maps (1/50,000) of the region (Jerada, Wad Al Himar) (see
Fig. 2 for approximate positions).
A: Jerada Synclinorium, northern flank:
P1. Eastern slope of Jbel Bou Keltoum, off-patch reef
facies in the upper part of Koudiat Es-Senn Formation;
34�21042N; 2�03023W.
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 341
123

P2. Valley flank of the creek east of abandoned barite
mine, lower third of Cafcaf Formation; 34�23015N;
2�00027W.
P3. Eastern side of the valley east of Koudiat Es-Senn;
upper Koudiat Es-Senn Formation; 34�22014N;
2�00014W.
P4. Coral patch reef on the southern slope of the hill east
of Koudiat Es-Senn; upper Koudiat Es-Senn; 34�22015N;
2�00009W.
P5. SW of Small hill (1022 m); Oued El Koriche
Formation; 34�23025N; 1�56023W.
B: Jerada Synclinorium, southern flank:
P6. Top and capping beds of the most western microbial-
sponge buildup and microbial-metazoan buildups in the
Koudiat Es-Senn Formation, west of P7; 34�17058N;
2�03021W.
P7. Large microbial-sponge buildup in the Koudiat
Es-Senn Formation west of the road Oujda-Ain Beni
Mathar and Oued Defla; 34�17059N; 2�03009W.
P8. Silty shales of the Oued Es-Sassi Formation south of
P7; west of the road Oujda-Ain Beni Mathar and Oued
Defla; 34�17055N; 2�03009W.
P9. Calciturbidites within the Oued El Koriche Forma-
tion east Oued Defla, west of P 10; 34�17058N;
2�03001W.
P10. Large olistolite mainly consisting of microbial-
sponge buildup facies east of the road Oujda-Ain Beni
Mathar and Oued Defla; 34�17056N; 2�02057W.
References
Aretz, M. 2002. Habitatanalyse und Riffbildungspotential kolonialer
rugoser Korallen aus dem Unterkarbon (Mississippium) von
Westeuropa. Kolner Forum fur Geologie und Palaontologie 10:
1–155.
Aretz, M., and E. Chevalier. 2007. After the collapse of stromatoporid
sponge—coral reefs—The Famennian and Dinantian reefs of
Belgium: Much more than Waulsortian Mounds. In PalaeozoicReefs and Bioaccumulations: Climatic and Evolutionary Con-trols, eds. Alvaro, J.-J., M. Aretz, F. Boulvain, A., Munnecke, D.
Vachard and E. Vennin, 275: 163–188. Geological Society
London, Special Publications.
Aretz, M., and H.-G. Herbig. 2007. The Visean of the Jerada
synclinorium (NE Morocco)—succession, basin architecture and
palaeogeographic implications. Journal of Stratigraphy 31(Sup-
pl. 1): 109.
Aretz, M., and H.-G. Herbig. 2008. Microbial-sponge and microbial-
metazoan buildups in the Late Visean basin-fill sequence of the
Jerada Massif (Carboniferous, NE Morocco). In Proceedings ofthe Carboniferous conference cologne 2006. From Platform toBasin. eds. Aretz, M., H.-G. Herbig, and I. Somerville.
Geological Journal 43: 307–336.
Aretz, M., and H.-G. Herbig. 2009. Corals from the upper Visean of
the southern Azrou-Khenifra Basin (Carboniferous, Central
Moroccan Meseta). Coral Research Bulletin (in press).
Aretz, M., and J. Nudds. 2005. The coral fauna of the Holkerian/
Asbian boundary stratotype section (Carboniferous) at Little
Asby Scar (Cumbria, England) and implications for the bound-
ary. Stratigraphy 2: 167–190.
Aretz, M., and J. Nudds. 2007. Palaeoecology of the Late Visean
(Dinantian) coral-chaetetid biostrome at Little Asby Scar
(Cumbria, Great Britain). In Fossil corals and sponges.Proceedings of the 9th international symposium on FossilCnidaria and Porifera, Graz 2003. eds. Hubmann, B., and W.
Piller, Schriftenreihe der Erdwissenschaftlichen Kommission 17:
365–381.
Aretz, M., H.-G. Herbig, and S. Huck. 2006. Les bioconstructions du
Viseen superieur (Carbonifere) du Bassin de Jerada (Maroc
oriental). Resumees Reunion Sciences de la Terre Dijon: S-16.
Berkhli, M., J.-C. Paicheler, and D. Vachard. 1993. Donnees
nouvelles sur la stratigraphie des terrains carboniferes de la
Meseta orientale marocaine (boutonnieres de Debdou, Mekam et
Jerada). Geologische Rundschau 82: 84–100.
Berkhli, M., D. Vachard, A. Tahiri, and J.-C. Paicheler. 1999.
Stratigraphie sequentielle du Viseen superieur du bassin de Jerada
(Maroc Oriental). Eclogae geologicae Helvetiae 92: 285–294.
Berkhli, M., D. Vachard, A. Tahiri, and J.-C. Paicheler. 2000.
Caracteres sedimentologiques et structuraux des depots volcan-
oclastiques du Viseen superieur–Serpoukhovien de la Meseta
orientale marocaine. Bulletin de l’Institut Scientifique Rabat 22:
1–8.
Bourque, P.-A., A. Madi, and B.L. Mamet. 1995. Waulsortian-type
bioherm development and response to sea-level fluctuations:
Upper Visean of Bechar basin, western Algeria. Journal ofSedimentary Research B65: 80–95.
Caldwell, W.G.E., and H.A.K. Charlesworth. 1962. Visean coral reefs
in the Bricklieve Mountains of Ireland. Proceedings of theGeological Association 73: 359–381.
Carruthers, R.G. 1909. Notes on the corals. In A Carboniferous faunafrom Novaja Semlja collected by Dr. W.S. Bruce. ed. Lee, G.W.
Transactions of the Royal Society of Edinburgh 47: 148–156.
Carruthers, R.G. 1910. On the evolution of Zaphrentis delanouei in
Lower Carboniferous Times. Quarterly Journal of the Geolog-ical Society London 66: 523–538.
Conrad, J. 1984. Les series carboniferes du Sahara central algerien,
324 p. These d’Etat, Universite d’Aix-Marseille.
Cozar, P., and I.D. Somerville. 2005. Stratigraphy of upper Visean
rocks in the Carlow area, southeast Ireland. Geological Journal40: 35–64.
Cozar, P., I.D. Somerville, M. Aretz, and H.-G. Herbig. 2005.
Biostratigraphic dating of the Bricklieve Limestone Formation
(NW-Ireland) using foraminifers, calcareous algae and rugose
corals. Irish Journal of Earth Sciences 23: 1–23.
d’Orbigny, A. 1852. Cours elementaires de paleontologie et degeologie stratigraphique. Vol. 2, 1, 349 p. Paris: Victor Masson.
Dana, J.D. 1846. Structure and classification of zoophytes. U.S.Exploring Expedition during the years 1838-1842 under thecommand of Charles Wilkes, U.S.N., Vol. 7, 740 p. Philadelphia:
Lea and Blanchard.
Desteucq, C., A. Izart, and P. Potherat. 1988. Etude sedimentologique
du Carbonifere du bassin de Jerada (Maroc oriental). Annales dela Societe geologique du Nord 107: 203–210.
Dixon, O.A. 1970. The variation in the Visean coral Caniniabenburbensis from north-west Ireland. Palaeontology 13: 52–63.
Dobrolyubova, T.A., N.V. Kabakovich, and T.A. Sayutina. 1966.
Korally nizhnego karbona Kuznetskoj kotloviny. Trudy Paleon-tological Institut Akademii Nauk SSSR 111: 1–276.
Dybowski, W.N. 1873. Monographie der Zoantharia Sclerodermata
Rugosa aus der Silurformation Estlands, Nordlivlands und der
Insel Gotland. Archiv fur Naturkunde Liv-, Ehst- und Kurlands 5:
257–414.
342 M. Aretz
123

Ehrenberg, C.G. 1834. Beitrage zur physiologischen Kenntniss der
Corallenthiere im allgemeinen, und besonders des rothen
Meeres, nebst einem Versuche zur physiologischen Systematik
derselben. Abhandlungen der Koniglichen Akademie der Wis-senschaften zu Berlin 1832: 225–380.
Federowski, J. 1971. Aulophyllidae (Tetracoralla) from the upper
Visean of Sudetes and Holy Cross Mountains. PaleontologicaPolonica 24: 1–137.
Fleming, J. 1828. A history of British animals, XXIII: 565 p.
Edinburgh and London: Bell and Bradfute.
Gallagher, S.J., C.V. MacDermot, I.D. Somerville, M. Pracht, and
A.G. Sleeman. 2006. Biostratigraphy, microfacies and deposi-
tional environments of upper Visean limestones from the Burren
region, County Clare, Ireland. Geological Journal 41: 61–91.
Grabau, A.W. 1928. Palaeozoic corals of China. Part. I, Tetraseptata,
II, Second contribution to our knowledge of the streptelasmoid
corals of China and adjacent territories. Palaeontologica Sinica(serie B) 2: 1–175.
Herbig, H.-G. 1986. Rugosa und Heterocorallia aus Obervise-
Gerollen der Marbella-Formation (Betische Kordilliere, Sudspa-
nien). Palaontologische Zeitschrift 60: 189–225.
Herbig, H.-G., M. Aretz, A. Aurag, F. Leis, S. Rautenberg, A.
Schiefer, and A. Milhi. 2006. The Visean of the Jerada Massif
(NE Morocco): lithostratigraphy, facies, and depositional setting.
In Carboniferous conference Cologne. from platform to basin,September 4–10, 2006. Program and Abstracts, eds. Aretz, M.,
and H.-G. Herbig, Kolner Forum fur Geologie und Palaontol-ogie 15: 36–38.
Hill, D. 1938, 1939, 1940, 1941. A monograph on the Carboniferous
rugose corals of Scotland. Palaeontographical Society of Lon-don, 1938: 1–78, 1939: 79–114, 1940: 115–204, 1941: 205–215.
Hill, D. 1981. Coelenterata: Anthozoa. Subclasses Rugosa, Tabulata.
In Treatise on invertebrate paleontology, part F, Coelenterata,Suppl. 1, Rugosa and Tabulata. 762 p. ed. Teichert, C., Boulder/
Lawrence: Geological Society of America/University of Kansas
Press.
Hoffmann, N., W. Lindert, D. Weyer, and K.-H. Illers. 1975. Zum
Unterkarbon-Vorkommen auf den Inseln Rugen und Hiddensee.
Zeitschrift fur Geologische Wissenschaften 3: 851–873.
Hudson, R.G.S. 1941. On the Carboniferous corals, Zaphrentiscarruthersi sp. nov., from the Mirk Fell beds and its relation to
the Z. delanouei species group. Proceedings of the YorkshireGeological Society 24: 290–311.
Hudson, R.G.S. 1942. Fasciculophyllum Thomson and other genera
of the ‘‘Zaphrentis’’ omaliusi group of Carboniferous corals.
Geological Magazine 79: 257–263.
Hudson, R.G.S., and M.I. Platt. 1927. On the Lower Carboniferous
corals: the development of Rylstonia benecompacta, gen. and sp.
nov. Annals and Magazine of Natural History (series 9) 19: 39–48.
Kato, M. 1971. J. Fleming’s species of the British Lower Carbon-
iferous. Transactions and Proceedings of the PaleontologicalSociety of Japan (new series) 81: 1–10.
Khoa, N.D. 1977. Carboniferous Rugosa and Heterocorallia form
boreholes in the Lublin region (Poland). Acta PalaeontologicaPolonica 22: 301–404.
Kimpe, W.F.M., M.J. Bless, J. Bouckaert, R. Conil, E. Groessens,
J.P.M.T. Meesen, E. Poty, M. Streel, J. Thorez, and M.
Vanguestaine. 1978. Paleozoic deposits east of the Brabant
Massif in Belgium and the Netherlands. Mededelingen RijksGeologische Dienst (Nieuwe Serie) 30(2): 37–103.
Korn, D., and V. Ebbighausen. 2008. The Early Carboniferous
(Mississippian) ammonoids from the Chebket el Hamra (Jerada
Basin, Morocco). Fossil Record 11: 83–156.
Lewis, H.P. 1927. Caninia cylindrica Scouler and other large
Caninias from the Carboniferous limestone of Ireland. ScientificProceedings of the Royal Dublin Society 18: 373–382.
Lewis, H.P. 1930. The Avonian succession in the south of the Isle of
Man. Quarterly Journal of the Geological Society London 86:
234–290.
Liao, W.H., and S. Rodrıguez. 1999. Lower Carboniferous corals
from the southwestern margin of the Tarim Basin, NW China.
Geobios 32(4): 539–559.
McCoy, F. 1844. A synopsis of the characters of the Carboniferouslimestone fossils of Ireland, 207 p. Dublin: Dublin University
Press.
McCoy, F. 1849. On some new genera and species of Paleozoic corals
and foraminifera. Annals and Magazine of Natural History(series 2) 3: 1–20, 119–136.
McCoy, F. 1851–1855. In A synopsis of the classification of theBritish Paleozoic rocks with a systematic description of theBritish Palaeozoic fossils in the geological museum of theUniversity of Cambridge, ed. Sedgwick, A., 1851: 1–184, 185–
406, 1855: I–XCVIII, 407–661. London: W. Parker and Son and
Cambridge University Press.
Menchikoff, N., and T.Y. Hsu. 1935. Les polypiers carboniferes du
Sahara occidental. Bulletin de la Societe geologique de France(serie 5) 5: 229–261.
Michelin, H. 1840–1847. Iconographie Zoophytologique. Descriptionpar localites et terrains des polypiers fossiles de France et paysenvironnants, 348 p. Paris: E. Bertrand.
Milne-Edwards, H., and J. Haime. 1848. Recherches sur les polypiers.
Quatrieme memoire. Monographie des Astreides. Annales desSciences Naturelles Paris (serie 3) 10: 209–320.
Milne-Edwards, H., and J. Haime. 1850–1855. A monograph of the
British fossil corals. Palaeontographical Society Monographs1850: i–lxxxv, 1–71, 1851: 73–145, 1852: 147–210, 1853: 211–
244, 1855: 245–299.
Mitchell, M. 1989. Biostratigraphy of Visean (Dinantian) rugose coral
faunas of Britain. Proceedings of the Yorkshire GeologicalSociety 47: 233–247.
Niko, S., and N. Yamagiwa. 1998. Early Carboniferous corals from
the ‘‘Omi non-calcareous group’’, Niigata Prefecture. Bulletin ofthe National Science Museum Tokyo (C) 24(3,4): 129–150.
Nudds, J., and A. Day. 1997. The effects of clastic sedimentation on a
fasciculate rugose coral from the Lower Carboniferous of
Northern England. Boletın de la Real Sociedad Espanola deHistoria Natural (Seccion Geologica) 91: 93–97.
Owodenko, B. 1946. Memoire explicatif de la carte geologique du
bassin houiller de Djerada et de la region au sud d’Oujda. SocieteGeologique de Belgique, Memoire 70: 1–168.
Owodenko, B. 1976. Le bassin houiller de Jerada (Maroc oriental)—
Essai de synthese et de paleogeographie. Service geologique duMaroc, Notes et Memoire 207: 1–133.
Parkinson, J. 1808. Organic remains of a former world. Vol. 2Zoophytes, 286 p. London: Sherwood, Nelly & Jones.
Phillips, J. 1836. Illustrations of the geology of Yorkshire, Part 2: TheMountain Limestone, 253. London: John Murray.
Pique, A. 2001. Geology of Northwest Africa, 310 p. Stuttgart:
Gebruder Borntraeger (Beitrage zur regionalen Geologie der
Erde 29).
Pique, A., G. Bossiere, J.-P. Bouillin, A. Chalouan, and C. Hoepffner.
1993. The southern margin of the Variscan belt. The northwest-
ern Gondwana mobile zone (Eastern Morocco and Northern
Algeria). Geologische Rundschau 82: 432–439.
Poty, E. 1981. Recherches sur les Tetracoralliaires et les Heteroco-
ralliaires du Viseen de la Belgique. Mededelingen RijksGeologische Dienst 36: 1–161.
Poty, E. 1993. Heterochronic processes in some lower Carboniferous
rugose corals. Courier Forschungsinstitut Senckenberg 164:
141–152.
Poty, E. 2002. Stratigraphy and paleobiogeography of Carboniferous
Rugose Corals of Nova Scotia. In Carboniferous and Permian of
Rugose corals from the upper Visean (Carboniferous) of the Jerada Massif 343
123

the world. eds. Hills, L.V., C.M. Henderson, and E.W. Bamber,
Canadian Society of Petroleum Geologists, Memoir 19: 580–
587.
Poty, E., and D. Hannay. 1994. Stratigraphy of rugose corals in the
Dinantian of the Boulonnais (France). Memoires de l’InstitutGeologique de l‘Universite de Louvain 35: 51–82.
Ramsbottom, W.H.C., and M. Mitchell. 1980. The recognition and
division of the Tournaisian Series in Britain. Journal of theGeological Society London 137: 61–63.
Rodrıguez, S., and I. Somerville. 2007. Comparisons of rugose corals
from the upper Visean of SW Spain and Ireland: implications for
improved resolution in Late Mississippian coral biostratigraphy.
In Fossil corals and sponges. Proceedings of the 9th interna-tional symposium on Fossil Cnidaria and Porifera, Graz 2003.Eds. Hubmann, B., and W. Piller, 17: 275–305. Schriftenreiheder Erdwissenschaftlichen Kommission.
Rodrıguez, S., and S. Falces. 1992. Corales rugosos. Coloquios dePaleontologia 44: 159–218.
Rodrıguez, S., J.M. Hernando, and I. Said. 2001a. Estudio de los
corales con aulos del Viseense (Carbonıfero) de la Unidad de la
Sierra del Castillo (Area del Guadiato, SO de Espana). Coloquiosde Paleontologıa 52: 85–94.
Rodrıguez, S., L. Rodrıguez-Curt, and J.M. Hernando. 2001b. Estudio
de los Aulophyllidae (Rugosa) from the Sierra del Castillo Unit
(Cordoba, Spain). Coloquios de Paleontologıa 52: 47–78.
Rodrıguez, S., J.M. Hernando, and L. Rodrıguez-Curt. 2002. Estudio
de los coralos lithostrotionidos del Viseense (Misisipense) de la
Unidad de la Sierra del Castillo (Cordoba, Espana). RevistaEspanola de Paleontologıa 17: 13–36.
Rodrıguez, S., J.M. Hernando, and L. Rodrıguez-Curt. 2004. Estudio
de los corales cyathopsidos (Rugosa) del Viseense superior de la
Unidad de la Sierra del Castillo (Cordoba, Espana). Coloquios dePaleontologıa 54: 69–82.
Ryder, T.A. 1930. Notes on ‘‘Carcinophyllum’’, Nich. and Thom.,
with descriptions of two new species. Annals and Magazine ofNatural History London (series 10) 5: 337–351.
Said, I., and S. Rodrıguez. 2007. Description of Tizraia berkhlii gen.
et sp. nov. (Rugosa) from Adarouch Area (Brigantian, NE
Central Morocco). Coloquios de Paleontologıa 57: 23–35.
Said, I., and S. Rodrıguez. 2008. Descripcion de los corales
aulophyllidos del Viseense y Serpujoviense del area de Ada-
rouch (Marruecos). Coloquios de Paleontologıa 58: 13–40.
Said, I., S. Rodrıguez, and M. Berkhli. 2007. Preliminary data on the
coral distribution in the upper Visean (Mississippian) succession
from Adarouch area (NE Central Morocco). In Fossil Corals andSponges. Proceedings of the 9th international symposium onFossil Cnidaria and Porifera, Graz 2003. Eds. Hubmann, B.,
and W. Piller, Schriftenreihe der Erdwissenschaftlichen Kom-mission 17: 353–364.
Salee, A. 1913. Contribution a l’etude des polypiers du calcaire
carbonifere de la Belgique. Memoire Institut Geologique del’Universite de Louvain 1: 177–293.
Sando, W.J. 1976. Revision of the Carboniferous genus Aulina Smith
(Coelenterata, Anthozoa). Journal of Research, US GeologicalSurvey 4: 421–435.
Sando, W.J. 1980. The paleoecology of Mississippian corals in the
western conterminous United States. Acta PalaeontologicaPolonica 25: 619–631.
Sando, W.J. 1990. Global Mississippian coral zonation. CourierForschungsinstitut Senckenberg 130: 173–187.
Scouler, J. 1844. In A synopsis of the characters of the Carboniferouslimestone Fossils of Ireland, ed. McCoy, F., 207. Dublin: Dublin
University Press.
Semenoff-Tian-Chansky, P. 1974. Recherches sur les Tetracoralli-
aires du Carbonifere du Sahara occidental. Centre de Recherchesdes Zones arides (serie geologie) 21: 1–316.
Semenoff-Tian-Chansky, P. 1985. Corals. In The Carboniferous ofthe world, II, Australia, Indian subcontinent, South Africa, SouthAmerica and North Africa. eds. Wagner, R.H., C.F. Winkler-
Prins, and L.F. Granados, 20: 374–381. IUGS Publication.
Semenoff-Tian-Chansky, P., and J. Nudds. 1979. Revision de
quelques especes de Lithostrotion des ıles Britanniques decrites
par Milne-Edwards et Haime (Tetracorallaires carboniferes).
Bulletin du Museum Nationale d0Histoire naturelle (serie 4) 1:
245–283.
Spasskiy, N.Y. 1974. Dialekticheskoe edinstvo prostranstvennorvre-
mennykh zakonomernostey evolyutsii (na primere chet-
yrekhluchevykh korallov). Leningrad Gornyi Institut, Zapiski67: 127–135.
Termier, H., and G. Termier. 1950. Paleontologie Marocaine. II.
Invertebres de l’ere primaire. Fasc. I. Foraminiferes, spongiaires
et coelenteres. Notes et Memoires du Service de la cartegeologique du Maroc 73: 1–219.
Thomson, J. 1880. Contributions to our knowledge of the rugose
corals from the Carboniferous Limestone of Scotland. Proceed-ings of the Royal Philosophical Society Glasgow 12: 225–261.
Thomson, J. 1881. On the genus Alveolites, Amplexus and Zaphren-
tis, from the Carboniferous system of Scotland. Proceedings ofthe Royal Philosophical Society Glasgow 13: 194–237.
Thomson, J., and H.A. Nicholson. 1876. Contribution to the study of
the chief generic types of the Paleozoic corals. Annals andMagazine of Natural History (series 3) 17: 60–70, 123–138,
290–305, 451–461.
Vachard, D., and M. Berkhli. 1992. Importance des coupes du bassin
de Jerada (Maroc) pour la connaissance du Viseen terminal.
Revue de Micropaleontologie 35: 307–328.
Vai, G.B. 1991. Palaeozoic strike-slip rift pulses and palaeogeography
in the circum-Mediterranean Tethyan realm. Palaeogeography,Palaeoclimatology. Palaeoecology 87(1–4): 223–252.
Vaughan, A. 1906. The Carboniferous limestone series (Avonian) of
the Avon Gorge. Proceedings of the Bristol Natural Society 1:
73–166.
Vaughan, A. 1915. Correlation of Dinantian and Avonian. QuarterlyJournal of the Geological Society London 71: 1–52.
Verrill, A.E. 1865. Classification of polyps (Extract condensed from a
synopsis of the polypi of the North Pacific exploring Expedition,
under captains Ringgold and Rodgers). Essex Institution Pro-ceedings 4: 145–149.
Wang, H.C. 1950. A revision of the zonantharie rugose in the light of
their minute skeletal structures. Philosophical TransactionsRoyal Society London (series B no. 611) 234: 175–246.
Wendt, J., B. Kaufmann, and Z. Belka. 2001. An exhumed Palaeozoic
underwater scenery: the Visean mud mounds of the eastern Anti-
Atlas (Morocco). Sedimentary Geology 145: 215–233.
344 M. Aretz
123