paraoxonase-2 deficiency enhances pseudomonas aeruginosa quorum sensing in murine tracheal epithelia
TRANSCRIPT

Paraoxonase-2 Deficiency Enhances Pseudomonas aeruginosa Quorum Sensing in Murine Tracheal Epithelia
David A. Stoltz1*, Egon A. Ozer1*
, Carey J. Ng2,3, Janet M. Yu2, Srinivasa T. Reddy2,3, Aldons J. Lusis2, Noam Bourquard3, Matthew R. Parsek4, Joseph Zabner1#, Diana M. Shih2
* These authors contributed equally to this work.
1Department of Internal Medicine, Roy J. and Lucille A. Carver College of Medicine,
University of Iowa, Iowa City, IA 52242, USA2Department of Medicine, David Geffen School of Medicine at UCLA, Los Angeles, CA
90095, USA3Department of Molecular and Medical Pharmacology, David Geffen School of Medicine
at UCLA, Los Angeles, CA 90095, USA4Department of Microbiology, University of Washington, Seattle, 98195, USA
# To whom correspondence should be addressed at:
Department of Internal MedicineRoy J. and Lucille A. Carver College of MedicineUniversity of Iowa 440 EMRBIowa City, IA 52242, USATel: 319-335-7608Fax: 319-335-7623E-mail: [email protected]
Running Title:Paraoxonase and Murine Airway Epithelia
Page 1 of 43Articles in PresS. Am J Physiol Lung Cell Mol Physiol (November 22, 2006). doi:10.1152/ajplung.00370.2006
Copyright © 2006 by the American Physiological Society.

1
ABSTRACT
Pseudomonas aeruginosa is an important cause of nosocomial infections and is
frequently present in the airways of cystic fibrosis patients. Quorum sensing mediates P.
aeruginosa’s virulence and biofilm formation through density-dependent interbacterial
signaling with autoinducers. N-3-oxododecanoyl homoserine lactone (3OC12-HSL) is
the major autoinducer in P. aeruginosa. We have previously shown that human airway
epithelia and paraoxonases (PONs) degrade 3OC12-HSL. This study investigated the
role of PON1, PON2, and PON3 in airway epithelial cell inactivation of 3OC12-HSL.
All three PONs were present in murine tracheal epithelial cells, with PON2 and PON3
expressed at the highest levels. Lysates of tracheal epithelial cells from PON2 knockout
mice, but not PON1 or PON3 knockout mice, had impaired 3OC12-HSL inactivation
compared to wild type mice. In contrast, PON1, PON2, or PON3 targeted deletions did
not affect 3OC12-HSL degradation by intact epithelia. Overexpression of PON2
enhanced 3OC12-HSL degradation by human airway epithelial cell lysates, but not by
intact epithelia. Finally, using a quorum-sensing reporter strain of P. aeruginosa, we
found that quorum sensing was enhanced in PON2 deficient airway epithelia. In
summary, these results show that loss of PON2 impairs 3OC12-HSL degradation by
airway epithelial cells and suggests that diffusion of 3OC12-HSL into the airway cells
can be the rate-limiting step for degradation of the molecule.
Keywords:
N-3-oxododecanoyl homoserine lactone, 3OC12-HSL, cystic fibrosis, innate immunity,
paraoxonase
Page 2 of 43

2
INTRODUCTION
The lung is continuously exposed to a variety of infectious pathogens and foreign
substances. Airway epithelial cells are uniquely designed for monitoring and clearing
invading microbes through a number of mechanisms including: mucociliary clearance,
proinflammatory cytokine and chemokine production, and release of antimicrobial
peptides such as defensins, lactoferrin, and lysozyme (2). In the absence of impaired
innate immune function or underlying comorbid medical conditions, these mechanisms
are generally effective at preventing the development of pulmonary infections.
An uncommon, but serious, cause of respiratory tract infections is Pseudomonas
aeruginosa, a Gram-negative bacterial pathogen that is ubiquitously present in the
environment. Under suitable conditions and in a susceptible host this microorganism is a
frequent cause of acute and chronic infections. P. aeruginosa is a common cause of
hospital-acquired infections including cystitis, burn wound infections, and ventilator
associated pneumonia. In addition, cystic fibrosis (CF) patients are frequently colonized
with P. aeruginosa and pulmonary decline in CF is directly correlated to infection (22,
27). P. aeruginosa’s ability to produce chronic infections is, in part, explained by its
capacity to produce and exist in biofilms. Biofilms are communities of bacteria that are
characterized by a structured architecture consisting of bacteria contained within an
extracellular matrix. This surrounding matrix is composed of both bacterial and host
components. Biofilms typically display a marked resistance to antibiotic killing and
therefore infections associated with biofilm-forming bacteria are difficult to eradicate
(20).
Page 3 of 43

3
Many bacteria, including P. aeruginosa, utilize a cell-to-cell communication
system termed quorum sensing to regulate biofilm formation, under certain conditions,
and virulence factor production (12). Quorum sensing is dependent upon environmental
concentrations of small quorum-sensing signaling molecules produced by bacteria and
therefore bacterial cell density. Two well-studied quorum-sensing systems in P.
aeruginosa, las and rhl, utilize molecules that are acylhomoserine lactones (AHLs). LasI
is responsible for the production of N-3-oxododecanoyl homoserine lactone (3OC12-
HSL) and LasR is the transcription factor that responds to 3OC12-HSL. RhlI is an
enzyme that synthesizes N-butanoyl homoserine lactone (C4-HSL) and RhlR is the
transcription factor that responds to C4-HSL. Increasing concentrations of 3OC12-HSL
lead to production of C4-HSL. These two systems constitute a regulatory cascade, with
the las system controlling expression of the rhl system. Importantly, as many as 6% of
the approximately 6,000 genes expressed by P. aeruginosa are influenced by the las or
rhl quorum-sensing systems (10, 28, 33).
Paraoxonases (PONs) are enzymes originally described based upon their ability to
hydrolyze organophosphates. Since the initial descriptions of PON’s enzymatic activity,
three members of the PON family have been described: PON1, PON2, and PON3 (6).
The three genes encoding the enzymes are located next to each other on the long arm of
human chromosome 7. Human PON1 and PON3 are primarily produced in the liver and
found circulating in serum bound to HDL particles. The majority of research related to
PONs has been directed at the proposed protective role of PON1 in cardiovascular
Page 4 of 43

4
disease. PON2 is more ubiquitously expressed and is found in many tissues including
placenta, brain, lung, and kidney (19). More recent studies have suggested that the native
substrates for PONs are lactones (14). We have shown that human airway epithelial cells
can inactivate AHLs, including 3OC12-HSL (4). This ability to interfere with quorum
sensing may represent an additional function of airway epithelial cells in host defense. In
support of this concept, PON1 is known to inhibit biofilm formation by P. aeruginosa
(21). Our hypothesis is that PON activity in airway epithelial cells is important in
quorum-sensing modulation and functions as part of the host defense response to P.
aeruginosa. Here we describe the functional consequences of targeted deletion of PON1,
PON2, and PON3 on quorum-sensing regulation by murine airway epithelial cells.
Page 5 of 43

5
MATERIALS AND METHODS
Primary airway epithelial cell culture model.
Primary human airway epithelial cells were isolated from trachea and bronchi of
human donor lungs. The isolated cells were seeded onto collagen coated, semi-
permeable membranes (0.6 cm2 Millicell-HA; Millipore, Bedford, MA) and grown at the
air-liquid interface as previously described (13, 40). All experiments were performed on
well-differentiated human airway epithelia (HAE) (greater than 14 days after seeding of
cells). Samples were collected with approval of the University of Iowa Institutional
Review Board.
Murine tracheal epithelial cells were isolated and grown based upon the method
previously described by You et al. (38). Mice tracheas were isolated following
euthanasia. Tracheas were cut length-wise and immersed in dissociation media (15 mg
Pronase/10 ml Ham’s F-12 media) at 4°C. Twenty-four hours later, epithelial cell clumps
were harvested and resuspended in media containing a 50:50 ratio of DMEM:Ham’s F-12
supplemented with 4mM glutamine, 10 µg/ml insulin, 5 µg/ml transferrin, 62 µg/ml
cholera toxin, 5 ng/ml epidermal growth factor, 0.03 mg/ml bovine pituitary extract, 10-8
M retinoic acid, 5% fetal bovine serum, and 20 U/µg/ml Pen-Strep. Harvested cells were
pre-incubated in a culture dish at 37°C in 5% CO2 for 2-3 hours to remove fibroblasts.
The epithelial cells were then seeded onto rat tail collagen pre-coated transwell
membranes (0.33 cm2 insert membrane growth area, Co-Star; Corning, NY). Four days
post-seeding, an apical air interface was attempted daily until a confluent culture epithelia
Page 6 of 43

6
was formed. Once formed, the polarized culture epithelia was maintained in media
containing 50:50 DMEM:Ham’s F-12 supplemented with 4 mM glutamine, 10-8 M
retinoic acid, 2% V/V NuSerum, and 20 U/µg/ml Pen-Strep. Culture epithelia were
viable for 3-4 months.
Microscopic analysis of cultured epithelium.
For light microscopy, epithelial cell cultures were embedded in paraffin,
sectioned, and stained with hematoxylin and eosin. Samples were prepared for
Transmission Electron Microscopy imaging using routine methods. Briefly, the intact
filters were fixed in 2.5% gluteraldehyde in 0.1M cacodylate buffer pH 7.4, post fixed in
1% osmium tetroxide, en bloc stained in 2.5% uranyl acetate, and dehydrated in a graded
series of ethanol. After infiltration with Eponate 12 (Ted Pella, Inc., Redding, CA) the
samples were cured, sectioned at 70 nm and poststained with uranyl acetate and lead
citrate. Sections on formvar coated and uncoated copper grids were imaged in a JEOL
1230 TEM (Peabody, MA) using a Gatan 2k x 2k UltraScan 1000 camera (Pleasanton,
CA.).
3OC12-HSL degradation by intact epithelium and lysate preparations.
For intact epithelium experiments, the apical and basolateral surfaces of cultured
airway epithelium were washed twice with PBS containing calcium and magnesium (PBS
+/+, pH 7.1). 3OC12-HSL in acidified ethyl acetate (EtAc) was dried under a nitrogen
gas stream and then dissolved in PBS +/+ to achieve a final concentration of 10 µM
3OC12-HSL. Dissolved 3OC12-HSL was added to the apical (150 µl) and basolateral
Page 7 of 43

7
(250 µl) surface of human airway epithelia. For murine tracheal epithelial cultures, 75 µl
and 250 µl were added to the apical and basolateral surfaces, respectively. Treated
epithelia were incubated at 37°C in 5% CO2 and 6 µl samples of the apical media were
collected at 0, 15, 30, and 60 minutes for determination of remaining 3OC12-HSL using
a bioassay. These samples were added to 100 µl of EtAc and stored in airtight glass vials
at -20°C.
For epithelial cell lysate experiments, epithelia were first washed with cold PBS
+/+ and then 50 µl of lysis buffer (50 mM Tris·HCl, pH 6.9/150 mM NaCl/10 µM
leupeptin/10 µM aprotinin/1 µM pepstatin A/1 mM phenylmethylsulfonyl fluoride/0.1
mg/ml benzamidine ) was added to the apical surface. After 20 min of rocking in lysis
buffer at 4°C, the cells were scraped free from the membrane with a pipette tip and lysed
by sonication (10 pulses) (Branson Sonifier 250, Danbury, CT). The cellular debris was
cleared from the lysate by centrifugation (4500 g x 30 sec, 4°C). Relative protein
concentrations were determined by a Bio-Rad protein assay (Bio-Rad Laboratories,
Hercules, CA). Lysate preparations (10%) were diluted in PBS and incubated in the
presence of 10 µM 3OC12-HSL (in PBS +/+) at 37°C. At various timepoints, 6 µl
aliquots were collected and stored as described above.
3OC12-HSL and 3OC12-HSL quantitative assay.
All AHLs were obtained from Sigma-Aldrich Chemicals (St. Louis, MO), except
for 3OC12-HSL, which was obtained from Vertex Pharmaceuticals (Coralville, IA).
Page 8 of 43

8
3OC12-HSL was measured in a quantitative bioassay as previously described using
Escherichia coli MG4 (pKDT17) (23).
CHO and epithelial cell culture gene transfer experiments.
Chinese hamster ovary (CHO) cells were cultured as monolayers in 24-well
plates. Recombinant adenovirus serotype 5 (Ad5) expressing hPON1, hPON2, hPON3,
or GFP were produced by the University of Iowa Vector and Gene Targeting core (34).
CHO cells were infected with recombinant Ad5 by CaPi transfection (8) and allowed to
express for 48 hours. Synthetic AHLs were reconstituted in PBS +/+ to 20 µM and
incubated for one hour on transfected CHO cells. 3OC12-HSL lactone ring hydrolysis
was determined as described below.
Primary cultures of human airway epithelia were infected basolaterally with 100
MOI (multiplicity of infection) of Ad5 expressing hPON2 or GFP. The Millicells were
returned to fresh media and incubated at 37°C for 48 hours and used for intact epithelia or
lysate experiments as previously described above.
Western blot analysis of CHO cells.
CHO cell lysates were made by incubation in lysis buffer (50 mM Tris·HCl, pH
7.5, 138 mM NaCl, 5 mM EDTA, 1 mM EGTA, 1 mM NaF, phenylmethylsulfonyl
fluoride, leupeptin, pepstatin, and aprotinin) with 1% Triton X-100 for 20 min at 4°C on a
rocker. Cells were scraped from the tissue culture plates and transferred to a microfuge
tube. Sonication was performed for 10 sec and cell debris was cleared by centrifugation
Page 9 of 43

9
at 4,500 g for 30 sec, 4°C. The lysates were then combined with loading buffer and
separated by SDS-PAGE. Protein was then transferred to polyvinylidene difluoride
membranes (PVDF) (Immobilon-P; Millipore, Billerica, MA). PVDF membranes were
blocked overnight at 4° C in 5% bovine serum albumin in PBS. Membranes were then
incubated with the appropriate primary antibody for 2 h at room temperature and then
washed 3 times in 1X TTBS (137 mM NaCl, 2.7 mM KCl, 2.5 mM Tris, 0.05% Tween
20). Primary antibodies were: PON1 – monoclonal, anti-human PON1 at 1:1000
(AbCAM, Cambridge, MA), PON2 – rabbit, polyclonal anti-human PON2 (Orbigen, San
Diego, CA), and PON3 – rabbit, polyclonal anti-human PON3 (generated by Affinity
BioReagents, Golden, CO). Secondary antibodies were conjugated to horseradish
peroxidase (Amersham Biosciences, Buckingshire, England) and used at 1:20,000 for 1 h.
Following 3 washes with 1X TTBS, immunoreactive bands were detected with
SuperSignal solution (Pierce, Rockford, IL) and exposed to film.
Determination of unhydrolyzed 3OC12-HSL by HPLC.
3OC12-HSL was extracted from the culture media samples in acidified ethyl
acetate, dried under sterile nitrogen gas, and concentrated 10-fold in methanol.
Reconstituted samples were loaded onto a C18 HPLC column (Supelco, Bellefonte, PA)
and eluted in a 60%-90% methanol:water gradient. Eluent from the HPLC column was
ionized using positive electrospray and directed into a quadrupole ion trap mass
spectrometer (ThermoFinnigan, San Jose, CA) for analysis of lactone ring hydrolysis.
Page 10 of 43

10
P. aeruginosa quorum-sensing reporter experiments.
Overnight cultures of a reporter strain of P. aeruginosa expressing lacZ under
control of the qsc102 3OC12-HSL-responsive promoter (PAO1-qsc102-lacZ) were
subcultured for 2 hrs at 37°C (100 µl in 5 ml LB containing gentamycin 100 µg/ml). The
bacteria were washed twice with cold PBS +/+ and, following centrifugation, the
bacterial pellet was resuspended in 0.5 ml PBS +/+. A 2 µl suspension of the bacteria
(approximately 105 organisms) was applied to the apical surface of the epithelium and
cultures were incubated at 37°C in 5% CO2. At various timepoints, the extracellular
bacteria were collected from the apical surface by rinsing with 100 µl of PBS +/+.
Quantitative bacterial counts were performed with standard plating methods and β-
galactosidase expression was determined with Galacto-Light Plus (Applied Biosystems,
Bedford, MA).
RT-PCR of murine tracheal epithelia.
PON1, PON2, and PON3 mRNA levels in murine tracheal epithelial cells were
determined as previously described (21).
Microarray analysis of HAE cultures.
Total RNA was extracted from HAE cultures from eight separate donors using
TRIzol reagent (Invitrogen, Carlsbad, CA) according to manufacturer’s
recommendations. Following a 30 minute treatment with 1U/10µg RNA with DNase I
(Invitrogen), an additional RNA cleanup step was performed using the Qiagen RNeasy
Page 11 of 43

11
total RNA isolation kit (Chatsworth, CA). The RNA was processed for expression array
studies according to manufacturer’s recommended protocols and analyzed using the
Affymetrix human U133A GeneChip (Santa Clara, CA) (26, 39).
PON knockout animal generation.
PON1 and PON2 knockout animals were generated as previously described (18,
29). Generation of the PON3 knockout mouse will be described elsewhere (J. M. Yu, A.
J. Lusis, and D. M. Shih, unpublished data). After germ line transmission, backcrossing
was performed with C57BL/6J for 10, 6 and 4 generations for PON1, PON2, and PON3
knockout mice, respectively. Control mice were wild type littermates, for their respective
experimental group. All experiments were reviewed and approved by the Animal Care
and Use Committees at UCLA and the University of Iowa.
Statistical Analysis.
Statistical analysis was performed using the Student’s t test. Statistical
significance was set at p < 0.05.
Page 12 of 43

12
RESULTS
Airway epithelial cells express PON1, PON2, and PON3 mRNA.
We have previously shown that human airway epithelial cells are capable of
degrading 3OC12-HSL (4) and that PONs are also able to inactivate 3OC12-HSL (21).
Since a common site of P. aeruginosa infection is the lung, and PONs are capable of
inactivating AHLs, we wanted to define the role of PON1, PON2, and PON3 in the
airway epithelia response to quorum-sensing signals.
To investigate the expression profile of PON1, PON2, and PON3 in murine
airway epithelium, we first cultured murine tracheal epithelial cells at the air-liquid
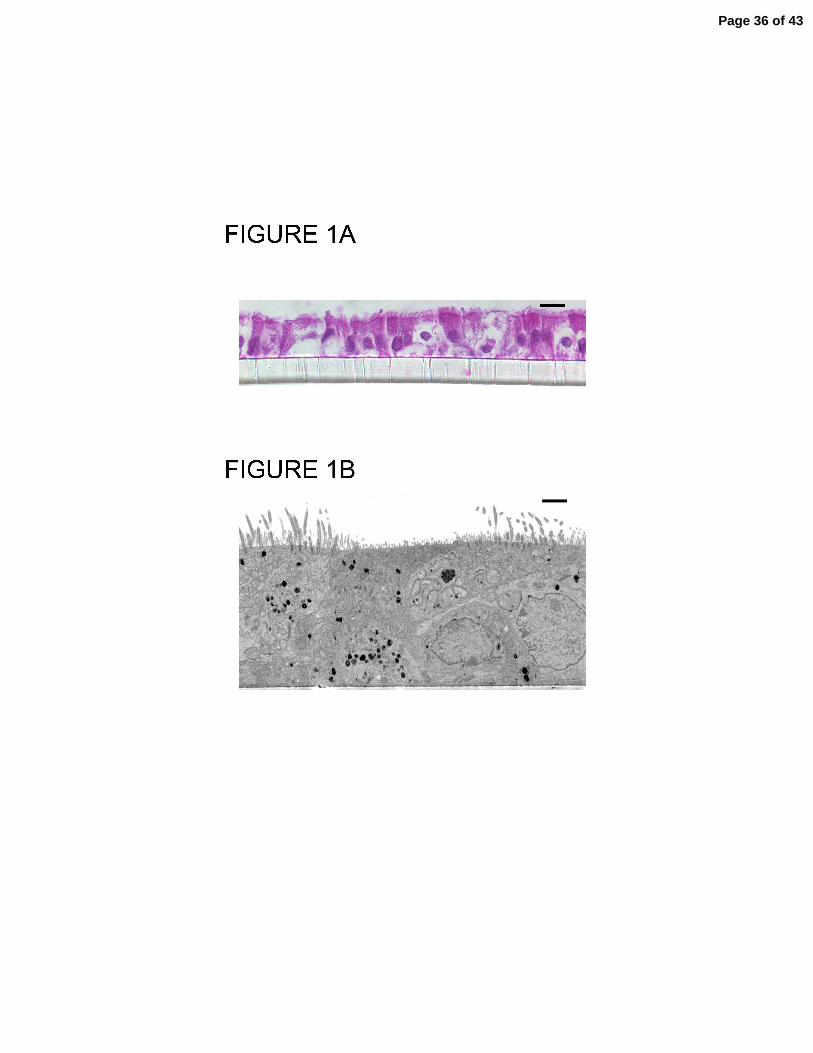
interface. Figures 1A and 1B show hematoxylin and eosin as well as TEM images of the
polarized epithelium grown at the air-liquid interface used for these studies. The cells
become well differentiated and form a pseudostratified epithelia with ciliated cells
present and an intact air-liquid interface. Transcripts for all three PONs were present.
However, PON2 and PON3 were expressed at the highest level (Figure 2A). This is in
contrast to human airway epithelia, that also express PON1, PON2, and PON3, but have
much greater expression of PON2 mRNA compared to PON1 or PON3 (Figure 2B).
PON2 preferentially degrades 3OC12-HSL.
PON is a promiscuous enzyme with multiple enzymatic activities including
paraoxonase, aryl esterase, and lactonase activity (6). Recent studies have suggested that
lactone molecules are likely to be PONs’ natural substrates (7, 14, 21). Since all three
members of the PON family are expressed in airway epithelia, we first examined whether
Page 13 of 43

13
substrate specificity was different among the three PONs. CHO cells were transduced
with adenoviruses encoding either PON1, PON2, or PON3 (Figure 3). Forty-eight hours
later, the cells were exposed to AHLs of varying side chain lengths and substitution
status. The active or closed-ring AHL forms in the cell media were determined by HPLC
following a 60 minute incubation. Figure 3 shows that all three PONs have similar
patterns of inactivating AHLs, with PON2 having the greatest lactonase activity
specifically for 3OC12-HSL. Prior studies have shown that PON2 has much greater
lactonase activity for AHLs, compared to PON1 or 3 (7, 21). PON2 specificity for
3OC12-HSL supports the hypothesis that this molecule may represent a natural substrate
for PON2. We also found that only AHLs with acyl side chains 8 carbons in length or
longer were susceptible to degradation by the PONs and the extent of lactonolysis
following incubation with the PON-expressing cells was proportional to the side-chain
length. Side-chain substitution also contributed to the amount of degradation in that
3OC12-HSL was more extensively degraded than C12-HSL, indicating that
hydroxylation of the third carbon of the acyl side chain augments PON’s lactonase
activity against the AHLs.
Murine tracheal epithelial cell lysates from PON2 knockout mice have impaired
3OC12-HSL inactivating ability.
We next asked whether loss of individual PONs would impair the ability of
murine tracheal epithelial cell lysates to inactivate 3OC12-HSL. PON1, PON2, and
PON3 knockout mice were generated by targeted deletion. Tracheas were harvested
Page 14 of 43

14
from knockout mice, and their corresponding wild type littermates, and then cultures of
murine tracheal epithelial cells were established.
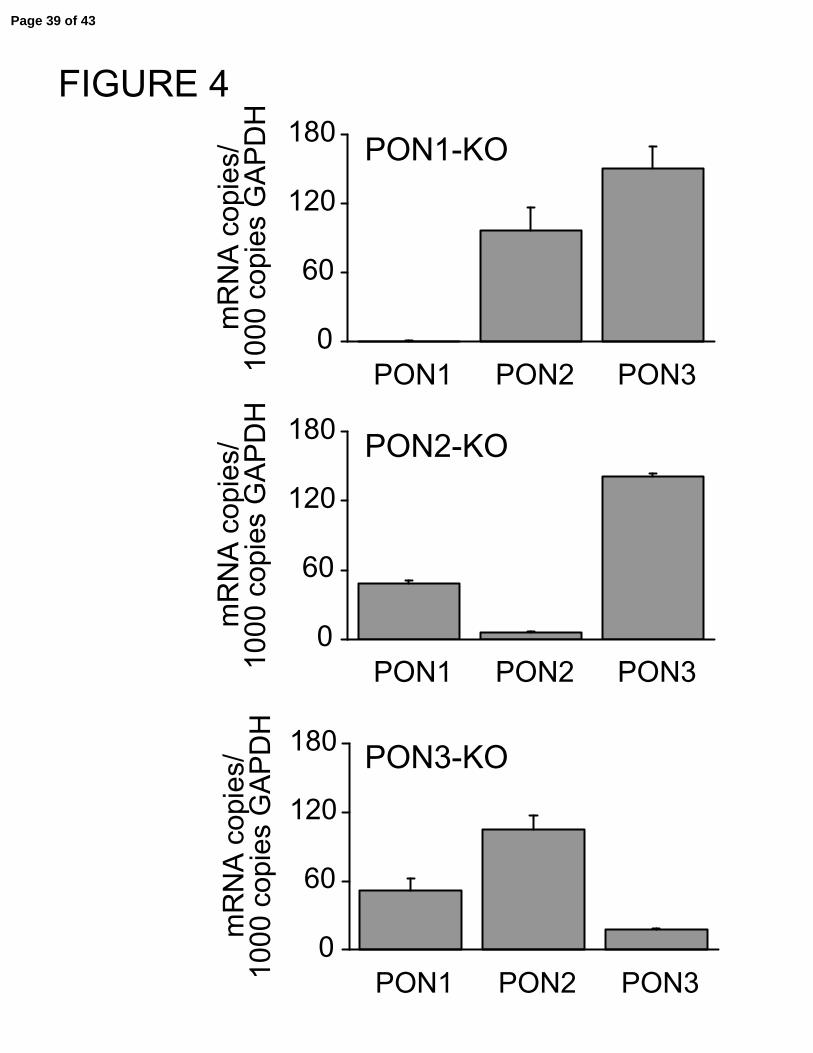
Figure 4 shows that PON1, PON2, and PON3 knockout murine tracheal epithelia
had little to no transcripts present for their respectively deleted PONs, compared to their
corresponding wild type epithelia (Figure 2A).
To investigate the ability of murine tracheal epithelial cells, individually disrupted
for each PON, to degrade 3OC12-HSL, we harvested cell lysates and exposed them to
3OC12-HSL. Wild type murine tracheal epithelial cell lysates degraded 3OC12-HSL
over time with less than 25% of active 3OC12-HSL remaining after a 60 minute
incubation. PON1 knockout and PON3 knockout tracheal epithelial cell lysates had
preserved 3OC12-HSL degrading capacity compared to littermate controls. However,
PON2 knockout epithelial cell lysates had significantly impaired ability to degrade
3OC12-HSL, with more than 75% of the unhydrolyzed 3OC12-HSL present at the end of
a 60 minute incubation (Figure 5). In order to determine if this defect could be reversed,
we added lysates from CHO cells transduced with AdPON2 to tracheal epithelial cell
lysates from PON2 knockout mice. 3OC12-HSL inactivation was significantly improved
with only 39.6 ± 1.7 % of unhydrolyzed 3OC12-HSL remaining after a 60 minute
incubation, compared to PON2 knockout lysate alone (79.1 ± 5.1%). These data show
that loss of PON1 or PON3 activity in murine tracheal epithelial cell lysates does not
affect AHL degradation, but the absence of PON2 impairs this activity.
Page 15 of 43

15
Intact murine tracheal epithelial cell cultures have preserved capacity to degrade
3OC12-HSL despite loss of PON1, PON2, or PON3.
In order to more closely simulate in vivo conditions, 3OC12-HSL hydrolysis by
an intact epithelia was determined. We hypothesized that intact epithelia from PON2
knockout mice would display impaired 3OC12-HSL degradation based upon our results
from the lysate experiments.
We first tested whether airway surface liquid (ASL) contained any AHL
degrading activity. Following collection of ASL from 12 transwells (in a total volume of
100 µl), we incubated the collected fluid with 3OC12-HSL. There was no degradation by
the ASL (data not shown), suggesting that PONs are either not secreted or not active in
the apical compartment. This is consistent with lack of lactonase activity of human and
mouse bronchoalveolar lavage fluid (4).
We then measured the hydrolysis of 10 µM 3OC12-HSL, following addition to
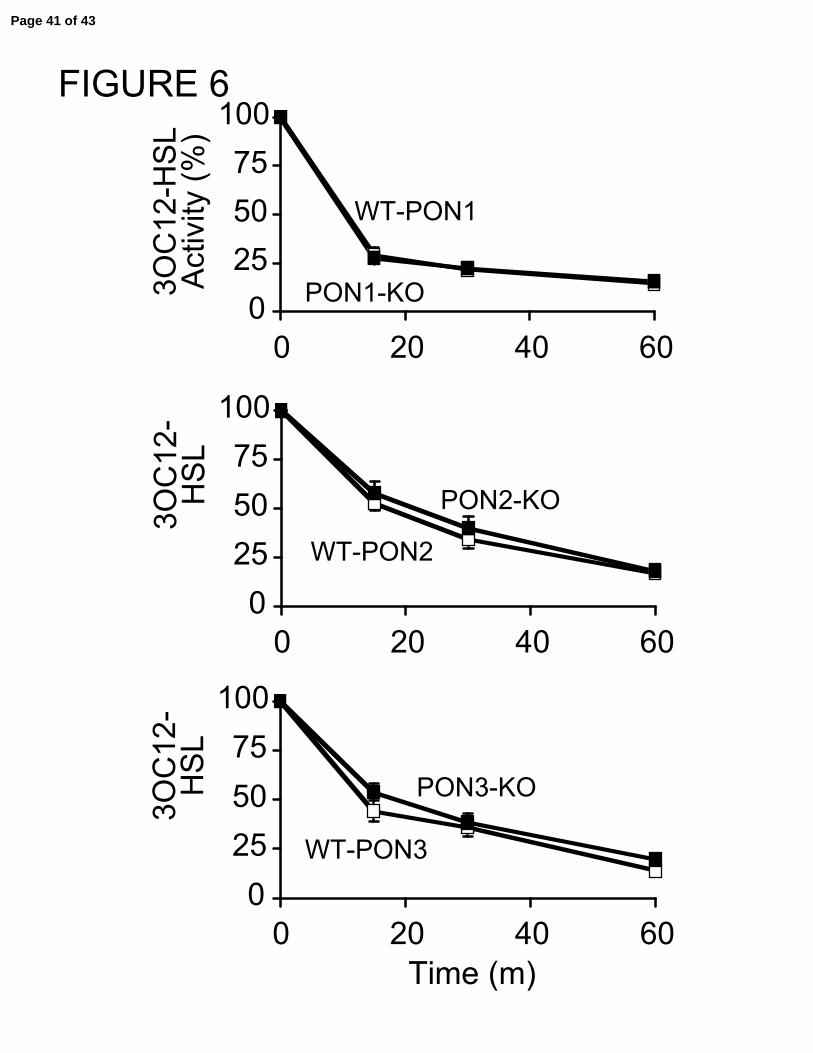
the apical and basolateral compartments of the murine epithelial transwells. Figure 6
shows that intact epithelia from both PON1 and PON3 deficient mice had preserved
capacity to degrade 3OC12-HSL. In contrast to our results from the PON2 knockout
lysate data, intact epithelia from PON2 knockout mice also degraded 3OC12-HSL at a
similar rate as control epithelia. This finding was surprising and raises several possible
explanations. First, differing cellular localizations of mouse PON1, PON2, and PON3
may account for the observed differences. Second, the lysate experimental conditions
may alter PON activity based upon yet undetermined factors including local calcium
Page 16 of 43

16
concentration, redox states, or endogenous inhibitors. Finally, diffusion of 3OC12-HSL
across the cell membrane may be the rate-limiting step to degradation at the
concentration tested.
Overexpression of PON2 in human airway epithelial cell cultures enhances 3OC12-
HSL degradation only in lysate preparations.
In order to test if diffusion of 3OC12-HSL across the cell membrane was the rate-
limiting step, we transduced HAEs with AdGFP or AdPON2. We predicted that if
diffusion across the membrane were rate limiting, increasing PON2 levels would result in
no change in 3OC12-HSL degradation, by intact epithelia, at the concentration tested.
Moreover, increasing concentrations of 3OC12-HSL should result in enhanced
degradation in the presence of higher PON2 concentrations. Alternatively, higher
3OC12-HSL concentrations in the setting of impaired lactonase activity (i.e, airway
epithelia from PON2 deficient mice) would lead to decreased 3OC12-HSL degradation.
We compared the rates of 3OC12-HSL degradation by hPON2 transduced epithelia in
lysate and intact epithelia preparations. Figure 7 shows that PON2 transduction
significantly enhanced inactivation of 3OC12-HSL in lysates. However, overexpression
of PON2 did not influence the rate of 3OC12-HSL degradation by intact epithelium.
These data suggest that at 10 µM 3OC12-HSL, diffusion across the cell membrane is
rate-limiting for inactivation of 3OC12-HSL.
Intact tracheal epithelial cultures from PON2 knockout mice show enhanced quorum-
sensing reporting by P. aeruginosa.
Page 17 of 43

17
We next wanted to test the hypothesis that increased concentrations of 3OC12-
HSL in the setting of diminished PON activity would result in impaired degradation.
However, concentrations of 3OC12-HSL greater than 10-20 µM are difficult to obtain
under experimental conditions due to the molecule’s limited solubility in aqueous
solutions. Therefore, to expose respiratory epithelium to greater 3OC12-HSL
concentrations and more closely mimic in vivo conditions, we developed a P. aeruginosa
β-galactosidase (β-gal) quorum-sensing reporter assay. We first developed and tested
this assay using serum from wild type and PON1 knockout mice. We used PON1
knockout sera because PON1 is the predominant PON present in sera. Figure 8A shows
that over a 6 hour period, β-gal expression increased in serum from both wild type and
PON1 knockout mice. However, PON1 knockout mouse serum had significantly higher
β-gal expression, compared to control serum. This shows that in the absence of PON1,
P. aeruginosa has enhanced quorum sensing as measured by increased β-gal expression.
We next tested this quorum-sensing assay in intact respiratory epithelium. A 2 µl
suspension of 105 cfu of PAO1-qsc-102-lacZ was applied to the apical surface of murine
tracheal epithelial cell cultures. Following varying lengths of incubation, the apical fluid
was collected for β-gal measurements (quorum-sensing activity) and bacterial
quantification. We hypothesized that PON2 knockout epithelia would show increased β-
gal levels compared to controls. At 4 and 6 hours following addition of the bacteria, β-
gal levels were increased with levels approximately doubled at the 6 hour timepoint,
compared to control epithelia (Figure 8B). Interestingly, the bacterial counts were similar
between the groups demonstrating that the observed differences in β-gal expression were
Page 18 of 43

18
caused by changes in quorum sensing and not due to changes in bacterial replication.
Page 19 of 43

19
DISCUSSION
The main results of this study show that murine tracheal epithelial cells are
capable of degrading the P. aeruginosa quorum-sensing molecule, 3OC12-HSL. In
addition, these cells express all three PON enzymes, with PON2 and PON3 being
expressed at the highest levels. While targeted deletion of PON1 and PON3 had no effect
on the ability of these cells to degrade 3OC12-HSL, loss of PON2 significantly impaired
this ability in lysate preparations. Finally, the absence of PON2 enhanced P. aeruginosa
quorum sensing in intact epithelia cultures. This study is the first to demonstrate the
individual contributions of PON1, PON2, and PON3 in airway epithelial cell 3OC12-
HSL degradation.
We previously reported that human airway epithelial cells are capable of
inactivating quorum-sensing molecules through an unknown mechanism (4). We, and
others, have subsequently shown that PONs can hydrolyze the lactone ring of AHL
quorum-sensing molecules, including 3OC12-HSL (7, 21, 37). This raised the question
of whether or not PON functions as an inhibitor of quorum-sensing signaling and
therefore as a regulator of P. aeruginosa virulence and, potentially, biofilm formation. In
response to this question, we demonstrated that recombinant PON1 and wild type murine
serum reduced biofilm formation, while biomass was enhanced in the presence of serum
deficient in PON1 (from PON1 knockout mice). Paradoxically, PON1 knockout mice
were protected in a P. aeruginosa peritonitis/sepsis model (21).
Page 20 of 43

20
The current study advances our knowledge regarding PON’s possible role in
airway host defense. The respiratory epithelium plays a very important role in the host
response to inhaled toxins and microbes that escape the upper airway clearance
mechanisms. These cells are uniquely situated to prevent colonization, and ultimately
infection, of the distal airways and alveoli. Airway epithelial cells can secrete a number
of mediators that are important for mobilization of immune system cells, repair of injured
epithelium, and direct pathogen killing with antimicrobial peptides such as β-defensins,
lactoferrin, lysozyme, LL-37, and secretory leukocyte proteinase inhibitor (2). We
propose that PONs may function as a defense mechanism against bacterial pathogens that
utilize lactone molecules to regulate their virulence. PONs are conserved from the
nematode Caenorhabditis elegans to mammals, but the endogenous substrate(s) is
unknown. Recently, we and others reported findings suggesting that the natural
substrates for PONs are in fact lactone molecules (7, 14, 21). Consistent with this
hypothesis is that while PON1, PON2, and PON3 all exhibit lactonase activity, there is
greater individual variability for other substrates depending upon the PON enzyme
examined (7, 32). Interestingly, in the human body, naturally occurring lactones are
limited and mainly consist of metabolites of fatty acid oxidation. Therefore, the question
remains whether exogenous lactones, possibly the AHL quorum-sensing molecules of P.
aeruginosa, represent a natural substrate for PON.
We showed in this study that murine tracheal epithelial lysates from PON2
deficient mice had impaired 3OC12-HSL degrading activity, compared to PON1 or
PON3 knockout cells. However, intact epithelium showed no differences. Several
Page 21 of 43

21
possible mechanisms may account for this discrepancy. First, although all three mouse
PONs are membrane bound (15), they may exhibit different subcellular localization
patterns. PON1 is bound to the extracellular membrane and transferred to HDL particles
(5). In HeLa cells, PON2 localizes to cytosolic and cell membrane fractions, but is
absent in the supernatant fraction (19). Human PON3 is also found on HDL, however, its
subcellular localization is not well understood (25). A global survey study of organ and
organelle protein expression in mouse provides evidence that mouse PON1, PON2, and
PON3 are all membrane bound without further information on subcellular localization
(website: http://tap.med.utoronto.ca/~mts/) (15). The method of lysate preparation (i.e.,
clearing centrifugation step) may affect which PONs are present in the reaction mixture
and how they individually contribute to 3OC12-HSL degradation. Secondly, the
chemical milieu that the PONs are exposed to may be different in the lysate versus intact
epithelium preparations. A number of conditions have been shown to affect PON activity
including: redox state, calcium concentration, lipid associations, and pH (1, 11, 17, 32).
Finally, diffusion of 3OC12-HSL across the cell membrane may be the rate determining
step for 3OC12-HSL inactivation. Findings from our intact epithelium studies, in which
individual deletion of PON1, PON2, or PON3 did not affect 3OC12-HSL inactivation
support the latter hypothesis. This suggests that diffusion of the molecule across the cell
membrane limits the rate of quorum-sensing molecule inactivation by PON. Also
supporting this hypothesis are the results of the experiments in which we over-expressed
PON2 in human airway epithelial cells. In the PON2 deficiency model, the remaining
two PONs, despite having relatively less lactonase activity compared to PON2, are still
able to adequately degrade 3OC12-HSL in the intact epithelia studies. The importance of
Page 22 of 43

22
PONs in the regulation of quorum-sensing control will ultimately require a murine model
with all three PONs deleted.
One advantage of this study is that with targeted deletion of the individual PONs,
the specific contributions of PON1, PON2, and PON3 towards airway epithelial cell
degradation of 3OC12-HSL could be examined. However, the true role of PON in
airway host defense and regulation of quorum sensing remains to be determined. PON2
may not be required, but redundant, with PON1 and PON3 each contributing to
lactonolysis of 3OC12-HSL. Alternatively, PON2 may be required but only for
intracellular signaling events or against microbes that become intracellularly located (9).
Finally, PON2 may be required, but only when extracellular concentrations of 3OC12-
HSL are markedly increased. AHL molecules are thought to be freely diffusible across
cell membranes, although to varying degrees based upon the carbon side chain length.
Very few studies have actually examined transport and kinetics of 3OC12-HSL
movement across cell membranes. Using a chimeric sensor protein based on LasR,
3OC12-HSL was found to enter COS-1 cells and activate these reporter constructs (35).
A study by Pearson et al. (24) reported that while C4-HSL reached steady state cell
concentrations in cell suspensions of P. aeruginosa by 30 seconds, 3OC12-HSL required
approximately 5 minutes to reach steady state levels. While these experiments were
conducted using bacterial cells, it does suggest that 3OC12-HSL is delayed in crossing
cell membranes.
Page 23 of 43

23
Airway epithelial cell paraoxonases may not only function as a component of the
innate immune system directly acting at the bacterial level, but these enzymes also may
be important for additional control of the inflammatory response by airway epithelial
cells responding to these signals. Quorum-sensing molecules have been shown to affect
many cells, including fibroblasts and epithelial cells. 3OC12-HSL induces an array of
inflammatory molecules including: prostaglandin E2, cyclooxygenase-2, interleukin-6,
and tumor necrosis factor-alpha (30, 31). Recently, Kravchenko et al. reported that
3OC12-HSL treatment of epithelial cells and macrophages leads to phosphorylation of
p38 and eukaryotic translation initiation factor 2α (eIF2a) (16), suggesting a possible
pathway for 3OC12-HSL’s effects on the epithelial cell’s inflammatory response. PΟΝ
may regulate these responses through 3OC12-HSL inactivation and inhibit uncontrolled
signaling of the inflammatory cascade.
If PON is a naturally occurring inhibitor of quorum sensing, then modulation of
its activity or overexpression may represent a potential therapeutic target. P. aeruginosa
infection of the lung is common and occurs in the acute setting, such as in nosocomial
pneumonia and burn wound infections, as well as in chronic infections such as in cystic
fibrosis patients. These infections are oftentimes difficult to treat and numerous
therapeutic options, besides antibiotics, have been proposed. Quorum-sensing inhibitors
have recently been investigated as a potential therapeutic agents with promising results.
Experimental data regarding PON’s role in the response to an infection is very limited.
Treatment of mice with a synthetic analogue of furanone, a naturally occurring quorum-
sensing inhibitor, improved bacterial clearance and mortality in a murine model of P.
Page 24 of 43

24
aeruginosa pulmonary infection (36). Similarly, mice treated with garlic extract, another
recently described quorum-sensing inhibitor, enhanced bacterial clearance in a
pneumonia model (3)
In summary, we have shown that murine tracheal epithelial cells express PON1,
PON2, and PON3. These cells can degrade 3OC12-HSL, but in the absence of PON2
this activity is impaired. Since PON2 inactivates 3OC12-HSL to the greatest extent and
is the most abundant PON in human airway epithelia, it is likely the primary enzyme
responsible for quorum-sensing molecule inactivation. The PONs may play a key role in
host defense to pathogens such as P. aeruginosa, but additional studies will be needed to
further address this question.
Page 25 of 43

25
ACKNOWLEDGEMENTS
We thank Lisa Jorgensen, Phil Karp, Thomas Moninger, Thomas Recker, Geri Traver,
Peter Taft, Xu-Ping Wang, Yu-Rong Xia, and Yi-Shou Shi for excellent assistance and
Michael Welsh, M.D. and Pete Greenberg, Ph.D. for insightful discussions. We
appreciate the support of the Central Microscopy Research Facility, the Gene Transfer
Vector Core, and the In Vitro Cell Models Core. These studies were supported in part by
NHLBI and NIH grants (RO1 HL71776 to STR, PO1 HL30568 to AJL and DMS, and
HL61234-08 to JZ).
Page 26 of 43

26
REFERENCES
1. Aviram M, Rosenblat M, Billecke S, Erogul J, Sorenson R, Bisgaier CL, Newton RS, and La Du B. Human serum paraoxonase (PON 1) is inactivated by oxidized low density lipoprotein and preserved by antioxidants. Free Radic Biol Med 26: 892-904, 1999.
2. Bals R and Hiemstra PS. Innate immunity in the lung: how epithelial cells fight against respiratory pathogens. Eur Respir J 23: 327-333, 2004.
3. Bjarnsholt T, Jensen PO, Rasmussen TB, Christophersen L, Calum H, Hentzer M, Hougen HP, Rygaard J, Moser C, Eberl L, Hoiby N, and Givskov M. Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology 151: 3873-3880, 2005.
4. Chun CK, Ozer EA, Welsh MJ, Zabner J, and Greenberg EP. Inactivation of a Pseudomonas aeruginosa quorum-sensing signal by human airway epithelia. Proc Natl Acad Sci U S A 101: 3587-3590, 2004.
5. Deakin S, Leviev I, Gomaraschi M, Calabresi L, Franceschini G, and James RW. Enzymatically active paraoxonase-1 is located at the external membrane of producing cells and released by a high affinity, saturable, desorption mechanism. J Biol Chem 277: 4301-4308, 2002.
6. Draganov DI and La Du BN. Pharmacogenetics of paraoxonase: a brief review. Naunyn Schmiedebergs Arch Pharmacol 369: 78-88, 2004.
7. Draganov DI, Teiber JF, Speelman A, Osawa Y, Sunahara R, and La Du BN.Human paraoxonases (PON1, PON2, and PON3) are lactonases with overlapping and distinct substrate specificities. J Lipid Res 46: 1239-1247, 2005.
8. Fasbender A, Lee JH, Walters RW, Moninger TO, Zabner J, and Welsh MJ.Incorporation of adenovirus in calcium phosphate precipitates enhances gene transfer to airway epithelia in vitro and in vivo. J Clin Invest 102: 184-193, 1998.
9. Garcia-Medina R, Dunne WM, Singh PK, and Brody SL. Pseudomonas aeruginosa acquires biofilm-like properties within airway epithelial cells. Infect Immun 73: 8298-8305, 2005.
10. Hentzer M, Wu H, Andersen JB, Riedel K, Rasmussen TB, Bagge N, Kumar N, Schembri MA, Song Z, Kristoffersen P, Manefield M, Costerton JW, Molin S, Eberl L, Steinberg P, Kjelleberg S, Hoiby N, and Givskov M. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. Embo J 22: 3803-3815, 2003.
Page 27 of 43

27
11. James RW and Deakin SP. The importance of high-density lipoproteins for paraoxonase-1 secretion, stability, and activity. Free Radic Biol Med 37: 1986-1994, 2004.
12. Juhas M, Eberl L, and Tummler B. Quorum sensing: the power of cooperation in the world of Pseudomonas. Environ Microbiol 7: 459-471, 2005.
13. Karp PH, Moninger T, Weber SP, Nesselhauf TS, Launspach J, Zabner J, and Welsh MJ. An in vitro model of differentiated human airway epithelia: methods and evaluation of primary cultures. In: Epithelial Cell Culture Protocols, edited by Wise C. Totowa, NJ: Humana Press, Inc., 2002, p. 115-137.
14. Khersonsky O and Tawfik DS. Structure-reactivity studies of serum paraoxonase PON1 suggest that its native activity is lactonase. Biochemistry 44: 6371-6382, 2005.
15. Kislinger T, Cox B, Kannan A, Chung C, Hu P, Ignatchenko A, Scott MS, Gramolini AO, Morris Q, Hallett MT, Rossant J, Hughes TR, Frey B, and Emili A. Global survey of organ and organelle protein expression in mouse: combined proteomic and transcriptomic profiling. Cell 125: 173-186, 2006.
16. Kravchenko VV, Kaufmann GF, Mathison JC, Scott DA, Katz AZ, Wood MR, Brogan AB, Lehmann M, Mee JM, Iwata K, Pan Q, Fearns C, Knaus UG, Meijler MM, Janda KD, and Ulevitch RJ. N-(3-OXO-ACYL) homoserine lactones signal cell activation through a mechanism distinct from the canonical pathogen-associated molecular pattern recognition receptor pathways. J Biol Chem281: 28822-28830, 2006.
17. Kuo CL and La Du BN. Calcium binding by human and rabbit serum paraoxonases. Structural stability and enzymatic activity. Drug Metab Dispos 26: 653-660, 1998.
18. Ng CJ, Bourquard N, Grijalva V, Hama S, Shih DM, Navab M, Fogelman AM, Lusis AJ, Young S, and Reddy ST. Paraoxonase-2 deficiency aggravates atherosclerosis in mice despite lower apolipoprotein-B containing lipoproteins -anti-atherogenic role for paraoxonase-2. J Biol Chem 281: 29491-29500, 2006.
19. Ng CJ, Wadleigh DJ, Gangopadhyay A, Hama S, Grijalva VR, Navab M, Fogelman AM, and Reddy ST. Paraoxonase-2 is a ubiquitously expressed protein with antioxidant properties and is capable of preventing cell-mediated oxidative modification of low density lipoprotein. J Biol Chem 276: 44444-44449, 2001.
Page 28 of 43

28
20. Nickel JC, Ruseska I, Wright JB, and Costerton JW. Tobramycin resistance of Pseudomonas aeruginosa cells growing as a biofilm on urinary catheter material. Antimicrob Agents Chemother 27: 619-624, 1985.
21. Ozer EA, Pezzulo A, Shih DM, Chun C, Furlong C, Lusis AJ, Greenberg EP, and Zabner J. Human and murine paraoxonase 1 are host modulators of Pseudomonas aeruginosa quorum-sensing. FEMS Microbiol Lett 253: 29-37, 2005.
22. Parad RB, Gerard CJ, Zurakowski D, Nichols DP, and Pier GB. Pulmonary outcome in cystic fibrosis is influenced primarily by mucoid Pseudomonas aeruginosa infection and immune status and only modestly by genotype. Infect Immun 67: 4744-4750, 1999.
23. Pearson JP, Gray KM, Passador L, Tucker KD, Eberhard A, Iglewski BH, and Greenberg EP. Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc Natl Acad Sci U S A 91: 197-201, 1994.
24. Pearson JP, Van Delden C, and Iglewski BH. Active efflux and diffusion are involved in transport of Pseudomonas aeruginosa cell-to-cell signals. J Bacteriol181: 1203-1210, 1999.
25. Reddy ST, Wadleigh DJ, Grijalva V, Ng C, Hama S, Gangopadhyay A, Shih DM, Lusis AJ, Navab M, and Fogelman AM. Human paraoxonase-3 is an HDL-associated enzyme with biological activity similar to paraoxonase-1 protein but is not regulated by oxidized lipids. Arterioscler Thromb Vasc Biol 21: 542-547, 2001.
26. Reese J, Das SK, Paria BC, Lim H, Song H, Matsumoto H, Knudtson KL, DuBois RN, and Dey SK. Global gene expression analysis to identify molecular markers of uterine receptivity and embryo implantation. J Biol Chem 276: 44137-44145, 2001.
27. Schaedel C, de Monestrol I, Hjelte L, Johannesson M, Kornfalt R, Lindblad A, Stradvik B, Wahlgren L, and Holmberg L. Predictors of deterioration of lung function in cystic fibrosis. Pediatr Pulmonol 33: 483-491, 2002.
28. Schuster M, Lostroh CP, Ogi T, and Greenberg EP. Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome analysis. J Bacteriol 185: 2066-2079, 2003.
29. Shih DM, Gu L, Xia YR, Navab M, Li WF, Hama S, Castellani LW, Furlong CE, Costa LG, Fogelman AM, and Lusis AJ. Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 394: 284-287, 1998.
Page 29 of 43

29
30. Smith RS, Harris SG, Phipps R, and Iglewski B. The Pseudomonas aeruginosa quorum-sensing molecule N-(3-oxododecanoyl)homoserine lactone contributes tovirulence and induces inflammation in vivo. J Bacteriol 184: 1132-1139, 2002.
31. Smith RS, Kelly R, Iglewski BH, and Phipps RP. The Pseudomonas autoinducer N-(3-oxododecanoyl) homoserine lactone induces cyclooxygenase-2 and prostaglandin E2 production in human lung fibroblasts: implications for inflammation. J Immunol 169: 2636-2642, 2002.
32. Teiber JF, Draganov DI, and La Du BN. Lactonase and lactonizing activities of human serum paraoxonase (PON1) and rabbit serum PON3. Biochem Pharmacol66: 887-896, 2003.
33. Wagner VE, Bushnell D, Passador L, Brooks AI, and Iglewski BH. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J Bacteriol 185: 2080-2095, 2003.
34. Welsh MJ, Zabner J, Graham SM, Smith AE, Moscicki R, and Wadsworth SC. Adenovirus-mediated gene transfer for cystic fibrosis: Part A. Safety of dose and repeat administration in the nasal epithelium. Part B. Clinical efficacy in the maxillary sinus. Hum Gene Ther 6: 205-218, 1995.
35. Williams SC, Patterson EK, Carty NL, Griswold JA, Hamood AN, and Rumbaugh KP. Pseudomonas aeruginosa autoinducer enters and functions in mammalian cells. J Bacteriol 186: 2281-2287, 2004.
36. Wu H, Song Z, Hentzer M, Andersen JB, Molin S, Givskov M, and Hoiby N.Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J Antimicrob Chemother 53: 1054-1061, 2004.
37. Yang F, Wang LH, Wang J, Dong YH, Hu JY, and Zhang LH. Quorum quenching enzyme activity is widely conserved in the sera of mammalian species. FEBS Lett 579: 3713-3717, 2005.
38. You Y, Richer EJ, Huang T, and Brody SL. Growth and differentiation of mouse tracheal epithelial cells: selection of a proliferative population. Am J Physiol Lung Cell Mol Physiol 283: L1315-1321, 2002.
39. Zabner J, Scheetz TE, Almabrazi HG, Casavant TL, Huang J, Keshavjee S, and McCray PB, Jr. CFTR DeltaF508 mutation has minimal effect on the gene expression profile of differentiated human airway epithelia. Am J Physiol Lung Cell Mol Physiol 289: L545-553, 2005.
Page 30 of 43

30
40. Zabner J, Wadsworth SC, Smith AE, and Welsh MJ. Adenovirus-mediated generation of cAMP-stimulated Cl- transport in cystic fibrosis airway epithelia in vitro: effect of promoter and administration method. Gene Ther 3: 458-465, 1996.
Page 31 of 43

31
FIGURE LEGENDS
FIGURE 1:
Imaging of murine tracheal epithelia. (A) Hematoxylin and eosin stained section of
well-differentiated murine tracheal epithelial cells on transwell membranes (0.33 cm2)
(bar = 10 µm); (B) Transmission electron microscopy of well-differentiated murine
tracheal epithelial cells on transwell membranes (0.33 cm2) (bar = 2 µm).
FIGURE 2:
Expression of PON1, 2, and 3 in murine tracheal and human airway epithelial cells.
RT-PCR using primers specific for murine PON 1, 2, and 3 was performed on tracheal
epithelial cell lysates. Microarray analysis, for PON1, 2, and 3 expression, was
performed on RNA samples extracted from human airway epithelia samples. (A) Murine
tracheal epithelia (B) human airway epithelia. Results are expressed as mRNA
copies/1000 copies GAPDH and signal intensity, respectively.
FIGURE 3:
Spectrum of inactivation of AHLs by PON1, PON2, and PON3. CHO cells were
transfected with an adenovirus expressing (A) hPON1, (B) hPON2, or (C) hPON3.
Unhydrolyzed AHLs were measured by HPLC following a 60 minute exposure. A
representative experiment is shown. Three independent experiments yielded qualitatively
similar results. Insets show Western blot analysis for PON1, PON2, or PON3 following
transduction of CHO cells.
Page 32 of 43

32
FIGURE 4:
Paraoxonase mRNA levels in murine tracheal epithelial cells following targeted
deletion of PON1, PON2, or PON3. Total RNA was isolated from murine tracheal
epithelial cells and subjected to RT-PCR using primers specific for murine paraoxonase
1, 2, and 3. Results are expressed as mRNA copies/1000 copies GAPDH.
FIGURE 5:
Impaired 3OC12-HSL degradation by murine tracheal epithelial cell lysates from
PON2 knockout mice. Following lysis of tracheal epithelial cell cultures from PON1,
PON2, and PON3 knockout mice, lysates (10% volume) were incubated with 10 µM
3OC12-HSL. Following a 15, 30, or 60 minute incubation, lysate samples were obtained
for measurement of inactivation of 3OC12-HSL. Data is expressed as the percentage of
active 3OC12-HSL remaining compared to initial levels. Data is mean ± SEM. n=2-3
experiments/group performed in quadruplicates. ∗ p < 0.05.
Page 33 of 43

33
FIGURE 6:
Preserved 3OC12-HSL degradation by intact murine tracheal epithelial cell cultures
from PON1, PON2, and PON3 knockout mice. Well-differentiated murine tracheal
epithelial cell cultures were incubated in the presence of 3OC12-HSL (final concentration
of 10 µM apically and basolaterally). Following a 15, 30, or 60 minute incubation,
samples of apical media were obtained for measurement of inactivation of 3OC12-HSL.
Data is expressed as the percentage of active 30C12-HSL remaining compared to initial
levels. Data is mean ± SEM. n=2-3 experiments/group performed in quadruplicates.
FIGURE 7:
Overexpression of PON2 in intact human airway epithelia cultures enhances
3OC12-HSL inactivation in lysate preparations, but not intact epithelia. Human
airway epithelia cultures were transfected with an adenovirus expressing PON2 or GFP.
48 hours later, 3OC12-HSL was added to lysate samples (2% volume) or intact epithelia.
30 minutes later 3OC12-HSL inactivation was measured. Data is expressed as the
percentage of active 3OC12-HSL remaining compared to initial levels. Data is mean ±
SEM. Experiments were performed in quadruplicates.
Page 34 of 43

34
FIGURE 8:
Loss of PON2 enhances P. aeruginosa quorum sensing. (A) PAO1-qsc-102-lacZ was
incubated in the presence of wild type or PON1 deficient serum. 6 hours later bacterial
counts and β-gal levels were measured. (B) a 2 µl sample of PAO1-qsc-102 lacZ (105
cfu) was applied to the apical surface of wild type or PON2 deficient murine tracheal
epithelia cultures. At 4 and 6 hours later bacterial counts and β-gal levels were
determined. Data is mean ± SEM. n=2-3 per group performed in triplicates. * p < 0.05.
Page 35 of 43
Page 36 of 43

FIGURE 2A
0
60
120
180
PON1 PON2 PON3
mR
NA
copi
es/
1000
copi
esG
AP
DH
010002000300040005000
PON1 PON2 PON3
Sig
nalI
nten
sity
FIGURE 2B
Page 37 of 43

0
50
100
150U
nhyd
roly
zed
AH
L(%
)
0
50
100
150
Unh
ydro
lyze
dA
HL
(%)
0
50
100
150
Unh
ydro
lyze
dA
HL
(%)
FIGURE 3
PON1
PON2
PON3
3OC12 C12 C10 C6C8 C4
3OC12 C12 C10 C6C8 C4
3OC12 C12 C10 C6C8 C4
AdPO
N1
AdG
FP
AdPO
N2
AdG
FP
AdPO
N3
AdG
FP
Page 38 of 43
0
60
120
180
PON1 PON2 PON3
mR
NA
copi
es/
1000
copi
esG
AP
DH
PON2KO
0
60
120
180
PON1 PON2 PON3
mR
NA
copi
es/
1000
copi
esG
AP
DH
PON3KO
FIGURE 4
0
60
120
180
PON1 PON2 PON3
mR
NA
copi
es/
1000
copi
esG
AP
DH
PON1KOPON1-KO
PON3-KO
PON2-KO
Page 39 of 43

0255075
100
0 20 40 60
3OC
12-H
SL
Act
ivity
(%)
0255075
100
0 20 40 60
3OC
12-H
SL
Act
ivity
(%)
FIGURE 5
WT-PON1
PON1-KO
WT-PON2
PON2-KO
WT-PON3
PON3-KO
* * *
0255075
100
0 20 40 60
3OC
12-
HS
L
Time (m)
Page 40 of 43
0255075
100
0 20 40 60
3OC
12-
HS
L
0255075
100
0 20 40 60
3OC
12-
HS
L
0255075
100
0 20 40 60
3OC
12-H
SL
Act
ivity
(%)
FIGURE 6
Time (m)
WT-PON1
PON1-KO
WT-PON2
PON2-KO
WT-PON3
PON3-KO
Page 41 of 43

0
20
40
60
80
100
3OC
12-H
SL
Act
ivity
(%) AdGFP
AdPON2
FIGURE 7
IntactEpithelium
Lysate
*
Page 42 of 43

0
4
8
12
16
0 2 4 6TIME (hrs)
0
1
2
3
4
0 2 4 6TIME (hrs)
FIG 8A
02468
log1
0C
FU/w
ell
PON1-KO
WT
PON2-KO
WT
FIG 8B
WT KO
SERUM
EPITHELIUM
Time (h)
Time (h)
*
*
Page 43 of 43