constriction in tracheal smooth muscle
TRANSCRIPT

Semenov, et al. 1
BK Channel β1 Subunit Regulation of Calcium Handling and
Constriction in Tracheal Smooth Muscle
Iurii Semenov, Bin Wang, Jeremiah T. Herlihy, Robert Brenner*
Department of Physiology, University of Texas Health Science Center at San Antonio,
San Antonio, Texas 78229.
Running Title: BK Channel β1 and Airway
Correspondence to:
Robert Brenner, Ph.D.Department of PhysiologyUniversity of Texas Health Science Center San Antonio7703 Floyd Curl DriveSan Antonio, Texas 78229Phone: (210)-567-4343Fax: (210)-567-4410E-mail: [email protected],
Page 1 of 36Articles in PresS. Am J Physiol Lung Cell Mol Physiol (April 21, 2006). doi:10.1152/ajplung.00104.2006
Copyright © 2006 by the American Physiological Society.

Semenov, et al. 2
ABSTRACT
The large conductance, calcium-activated (BK-type) potassium channels are
regulators of voltage-dependent calcium entry in many cell types. The BK channel
accessory β1 subunit promotes channel activation in smooth muscle and is required for
proper tone in the vasculature and bladder. However, although BK channels have also
been implicated in airway smooth muscle function, their regulation by the β1 subunit had
not been investigated. We have investigated the role of the β1 subunit in tracheal smooth
muscle by utilizing the gene targeted mice for the β1 subunit gene. Mice containing the
β1 knockout allele had significantly reduced BK channel activity in excised tracheal
smooth muscle patches, and had reduced spontaneous BK currents in whole tracheal
smooth muscle cells. The consequence of the β1 knockout was an increase in resting
calcium levels, and an increase in the sustained component of calcium influx following
cholinergic signaling. Tracheal constriction studies demonstrate that the β1 knockout
have the relatively same level of constriction as compared to BK channel block with
paxilline, indicating that BK channels contribute little to airway relaxation in the absence
of the β1 subunit. Utilizing nifedipine, we found that the increase constriction caused by
the β1 knockout could be accounted for by an increased recruitment of L-type voltage-
dependent calcium channels. These results indicate that the β1 subunit is required in
airway smooth muscle for BK channels to control voltage-dependent calcium influx
during rest and following cholinergic signaling.
KEYWORDS: BK Channels, Airway, β1 Subunit
Page 2 of 36

Semenov, et al. 3
INTRODUCTION
In many smooth muscle cells, voltage-dependent calcium channels are the major
conduits for calcium mediating constriction (21). Potassium channels generally function
to oppose constriction by hyperpolarizing membranes, which deactivates voltage-
dependent calcium channels and reduces calcium influx. Among these, the large-
conductance (BK-type) calcium activated potassium channels are broadly expressed and
have been studied in many cell types. These channels are composed of a pore-forming
alpha subunit, and a tissue-specific beta subunit. Binding of intracellular calcium and
depolarizing voltage gate BK channels to open. When open, BK channels have a very
large outward potassium conductance (>200 pS) and are therefore very effective in
hyperpolarizing the membrane. As such, they are important regulators of membrane
voltage in a number of cell types (6, 15, 20).
BK channels bind calcium at low affinity (around 10 µM) (9, 28) and require
colocalization with a calcium source or the contribution of voltage to open the channel at
physiological calcium concentrations (8, 30, 31). In vascular smooth muscle, large
voltage changes do not occur, and BK channels have an enhanced apparent calcium
sensitivity that is conferred by association with the one member of the accessory BK
channel beta subunit family (β1−β4 subunits), the β1 subunit (23, 42). Previously, the
important role of the β1 subunit was demonstrated utilizing targeted gene knockout of the
β1 locus in mice. Knockout mice demonstrated BK channels with reduced opening in
vascular smooth muscle, and increased vascular tone and essential hypertension (4, 37).
Similarly, the β1 subunit has been shown to have an important role was in regulating
bladder and colon smooth muscle tone (16, 36).
Page 3 of 36

Semenov, et al. 4
In airway smooth muscle (ASM) constriction generally occurs following receptor-
coupled activation of IP3 receptors, which releases calcium from internal calcium stores.
This has been termed pharmacomechanical coupling (41), in contrast to the
electromechanical coupling that is mediated through voltage-dependent calcium
channels. The predominant role of pharmacomechanical coupling in ASM is strongly
supported by the fact that voltage-dependent calcium channel blockers such as nifedipine
only partially relax following cholinergic evoked constriction (14) and are poor bronchial
dilators themselves (1, 25). Yet, there is considerable evidence that the control of
membrane voltage by potassium channels is an important factor affecting the relative
constriction of ASM. For example, depolarization of ASM with potassium chloride
causes constriction (47) and hyperpolarization with potassium channel agonists can relax
ASM (33). In addition, the most common acute treatments for asthma, the β–adrenergic
agonists confer much of their effects through activation of large conductance calcium-
activated (BK-type) potassium channels (24). Iberiotoxin, a highly specific BK channel
blocker reduces the bronchial dilatory effect of β-adrenergic agonists (7). It has
frequently been hypothesized that BK channels may serve an important role in
controlling bronchial constriction, and are potential pharmaceutical targets for regulating
ASM constriction during asthma (17). Yet the role of potassium channels in general, and
BK channels specifically in calcium signaling in ASM is poorly understood. Here, we
have utilized the β1 knockout to evaluate the role of BK channel β1 subunit in tracheal
constriction smooth muscle contraction during cholinergic signaling. We show that
modulation by the β1 subunit does have effects on airway constriction, and these effects
are mediated by regulation of voltage-dependent calcium channels.
Page 4 of 36

Semenov, et al. 5
METHODS
Tracheal Constriction Studies
The BK channel β1 subunit knockout mice are congenic by seven generations of
inbreeding to the C57BL/6 line of Jackson Labs (strain C57BL/6J) and maintained as
homozygous lines. Control animals used in these studies also were the background
C57BL/6 mice strain from Jackson Labs. For tracheal constriction studies, animals were
deeply anesthetized with isoflurane and then sacrificed with cervical dislocation. Trachea
were quickly removed and then dissected clean of surrounding tissues in ice-cold normal
physiological saline solution (PSS) according to compositions given below. The tracheal
tube was cut below the pharynx and above the primary bronchus bifurcation. Two metal
wires (attached to a force transducer and micrometer) were threaded into the lumen of the
trachea and the trachea was placed into an oxygenated (95% oxygen, 5% CO2, pH =
7.35), at 37oC organ bath. Resting tension was continuously readjusted to 1 gram for one
hour and then challenged with high potassium PSS solution (KCl substituted for NaCl on
an equimolar basis) until a reproducible constriction response was obtained. Subsequent
experimental challenges with drugs were normalized to the constriction response in the
high potassium PSS solution. In high potassium PSS, the potassium reversal potential is
depolarized and therefore potassium currents are unlikely to play a role in controlling
membrane voltage and tone. This was consistent with our finding that there was no
significant difference in the potassium evoked constriction between wild type and
knockout mice (N= 14). Normal PSS used was (mM) 119 NaCl, 4.7 KCl, 2 CaCl2, 1.18
KH2PO4, 1.17 MgSO4*7H20, 18 NaHCO3, 0.026 EDTA, 11 glucose, and 12.5 sucrose.
Page 5 of 36

Semenov, et al. 6
The high potassium PSS substituted 56.7 mM NaCl with equimolar KCl, all other salts
were unchanged.
Tracheal Myocytes Isolation
Tracheas were isolated as described above. The dorsal muscle layer was cut away
from the hyaline cartiladge rings and minced into approximately 1 mm pieces in calcium-
free Hepes buffered Krebs/BSA solution (in mM: 140 NaCl, 4.7 KCl, 1.13 MgCl, 10
Hepes, 10 glucose, 1 mg/ml bovine serum albumin fraction V, pH 7.3). After 2.5
units/ml papain (MP Biomedicals) and 1 mg/ml dithiothreitol were added to cells, cells
were dissociated at 37oC on a rocking platform for 20 minutes. The tissue was washed
once with the calcium-free Krebs solution and digested with 12.5 units/ml of type VII
collagenase (Sigma Chemical) for 10 minutes at 37oC. The tissue was washed 3X in
calcium-free Krebs, BSA and gently triturated to disburse single tracheal myocytes.
Tracheal myocytes were stored on ice in calcium-free Krebs/BSA and used the same day.
Measurement of BK Channel Activity.
Isolated tracheal myocytes were attached to german glass coverslips
(Bioindustrial Products, San Antonio, Texas) for 30 minutes at room temperature. For
inside/out excised patches, isotonic recording solutions consisted of 140 mM K-
Methanesulfonate, 2 mM KCl and 10 mM Hepes (pH 7.2). Internal solutions (bath
solution, cytosolic side of membrane) included calcium buffered with 5 mM HEDTA to
give a final calcium concentrations of 7 µM as predicted by the Maxchelator programs
Page 6 of 36

Semenov, et al. 7
(32), and confirmed with a calcium sensitive electrode. To reduce contraction of muscle
during patch clamping the internal solution was supplemented with 20 micromolar
cytochalasin D. External (pipet, external side of membrane) solution was supplemented
with 2 mM MgCl2. Patch clamp pipets were pulled to resistance of 4 to 6 megaohm.
Single channels were recorded using a HEKA electronics EPC8 patch clamp amplifier
and Pulse software and filtered at 3.3 KHz. Single channel open probability and open
channel dwell times was determined using the TAC analysis program (Bruxton Corp).
For recording of transient outward currents (whole cell mode), tracheal smooth muscle
cells were plated in Krebs solution and voltage clamped with the amphoterecin B
perforated patch clamp technique for 60 seconds time steps at a given voltage (34).
Spontaneous outward currents were analyzed with a 10 pA threshold detection using the
Minianalysis Program. (Synaptosoft Software, Leonia, NJ).
Fura-2 measurement of calcium.
Cells were loaded with 1 µM Fura-2 AM ester and 40 micromolar pluronic acid in
Hepes buffered Krebs solution. The loading was at room temperature for 30 minutes in
the dark. We found that loading for longer incubation times or at 37oC allowed the Fura-2
to load into subcellular compartments. This was apparent as a larger baseline Fura-2 ratio
and smaller response to cholinergic stimulation. Cells were washed free of Fura-2 AM
and placed into perfusion chamber in a Nikon TE2000S (Nikon USA) microscope fitted
with an Ion Optix Bialkali phototube (IonOptix Corporation, Milton, MA) to assay
fluorescence intensity. Excitation wavelength was controlled with a Sutter Instruments
DG5 high speed filter changer (Sutter Instrument Company, Novato, CA) using 100 ms
Page 7 of 36

Semenov, et al. 8
windows for the 340 and 380 wavelength excitations, and monitoring the 510 nM
emission at a frequency of 1 Hz during the experiment. The Fura-2 ratio was calibrated to
calcium concentration using culture cells with β-escin to make the membrane permeable
to calcium, and EGTA buffered calcium calibration solutions purchased from Molecular
Probes (Invitrogen Corporation, Carlsbad, CA).
RESULTS
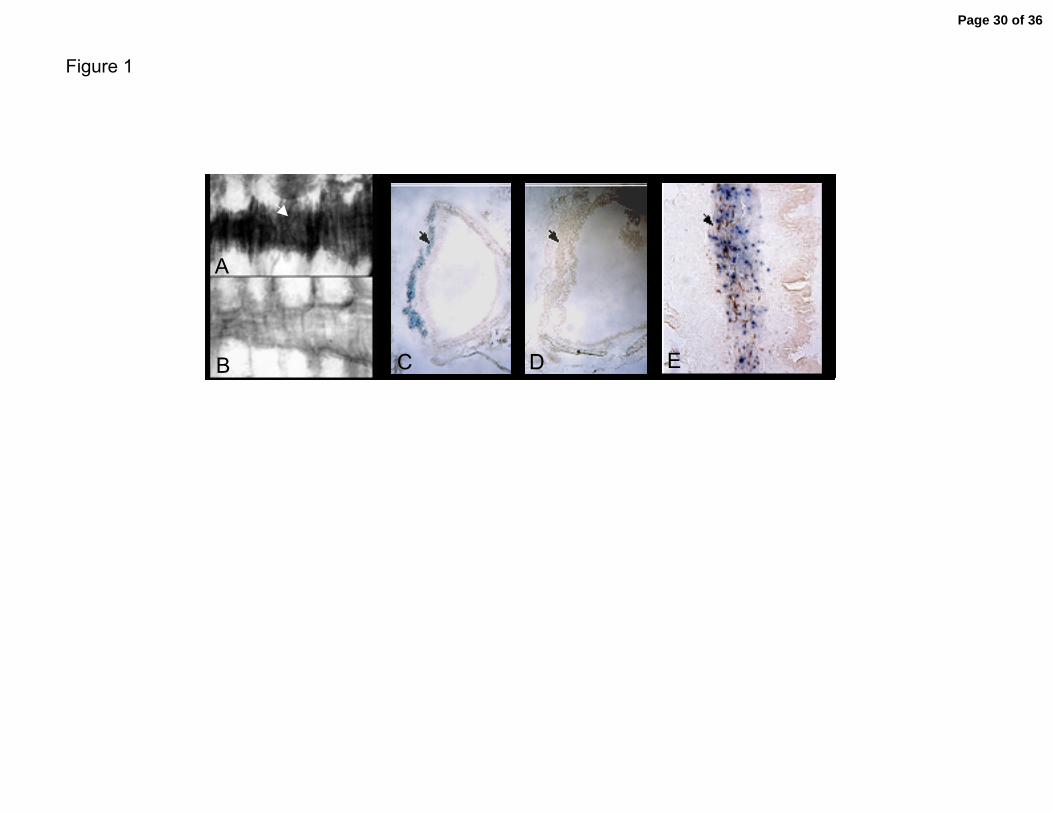
β1 Subunit Expression in Airway Smooth Muscle
BK channel activity is readily detected in both tracheal and bronchial smooth
muscle cells utilizing patch clamp techniques (40). Direct evidence for the accessory β1
subunit has been shown by biochemical copurification of the β1 subunit from bovine
tracheal smooth muscle in a 1:1 ratio with the pore-forming alpha subunit (22, 23). We
utilizing the β-galactosidase reporter that was gene targeted to the β1 subunit translation
initiation site in mice to directly determine the cell type that expresses the β1 gene in
airway tissues. Reporter activity is detected in trachea in the gene targeted animals
(Figure 1A, 1C, 1E), but not in control animals that lack the reporter (Figure 1B, 1D)
Figure 1A shows low magnification picture of β−galactosidase activity in the posterior
wall of the trachea (region of muscle between hyaline cartiledge rings). Consistent with
previous reports showing that β1 gene expression is smooth muscle-specific (4, 46),
expression in trachea is observed only in the smooth muscle layers (arrows in Figure 1C
and 1E).
BK Channels have a reduced opening in β1 knockout smooth muscle cells.
Page 8 of 36

Semenov, et al. 9
The β1 subunit confers an increased open probability and slow gating kinetics to
BK channels expressed in heterologous expression systems (11, 27, 28). The knockout of
the β1 subunit causes a dramatic reduction in BK channel openings and faster gating
kinetics of a number of smooth muscle tissues (4, 10, 36). However, BK channels alpha
subunits in different smooth muscle tissues undergo alternative splicing (26, 44, 45) and
the functional properties conferred by the β1 subunit can be dependent on the tissue-
specific splice product of the alpha subunit (12, 38). We investigated the functional
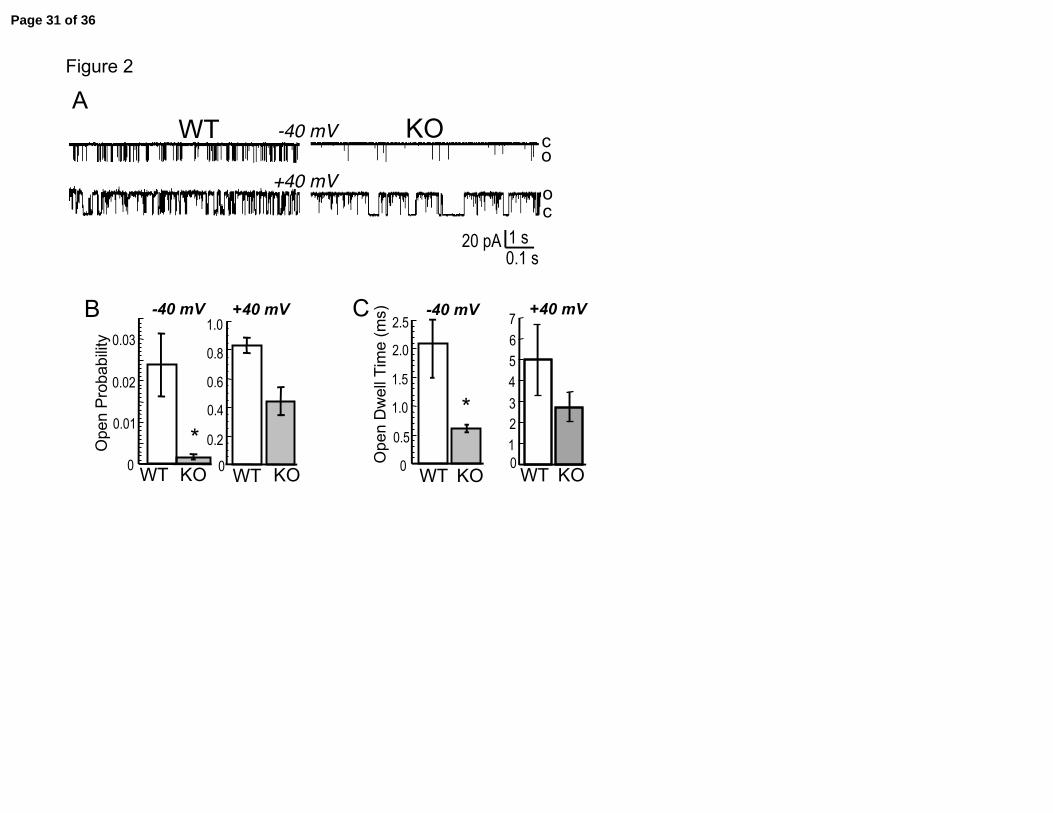
consequences of the lack of β1 subunit in tracheal smooth muscle. Figure 2A shows BK
single channel activity after excision in 7 micromolar calcium buffered internal solution.
At both –40 and +40 mV, wild type BK channels show increased activity over knockout
BK channels. As composite data in Figure 2B and Figure 2C show, wild type BK
channels at +40 mV have an approximately two fold increase in open probability and two
fold increase in open channel dwell time, respectively, as compared to BK channels
lacking β1 subunits (WT Po = 0.83 ± 0.06 vs. KO Po = 0.44 ± 0.1; WT open dwell time =
5 ± 1.7 ms vs. KO dwell time = 2.7 ± 0.7 ms). In tracheal smooth muscle, membrane
potentials generally do not overshoot 0 mV, and cholinergic signaling alters membrane
potentials within negative voltages ranges (19). At –40 mV we see a greater than 10 fold
reduction in open probability in the β1 knockout as compared to wild type cells (Figure
2B, WT = 2.4e-2 ± 0.76e-2 vs. KO = 0.17e-2 ± 0.07e-2, P < 0.05). These results indicate
that BK channel opening is severely perturbed in the β1 knockout at physiological
voltages. The mechanism by which BK channels increases open probability is by
increasing the open channel burst duration (30). As expected, channel burst durations in
Page 9 of 36

Semenov, et al. 10
the β1 knockout are reduced as compared to wild type BK channels (Figure 2C, -40 mV
WT = 2.1 ± 0.6 ms vs. KO = 0.54 ± 0.08 ms, P < 0.05).
Physiological changes in BK channel activity can be assessed by measuring
spontaneous transient outward currents (STOC) in non-dialyzed cells using perforated
patch clamp recording (34). Transient outward currents are mediated by BK currents
following calcium-induced calcium release events (so called “calcium sparks”), that
colocalize with, and activate, BK channels (18). Gene knockout of the pore-forming α
subunit has been shown to eliminate transient outward currents in bladder and vascular
smooth muscle (29, 39). Whereas in the knockout of the β1 subunit, spontaneous outward
currents are not absent but reduced in frequency in vascular smooth muscle (4, 37). In
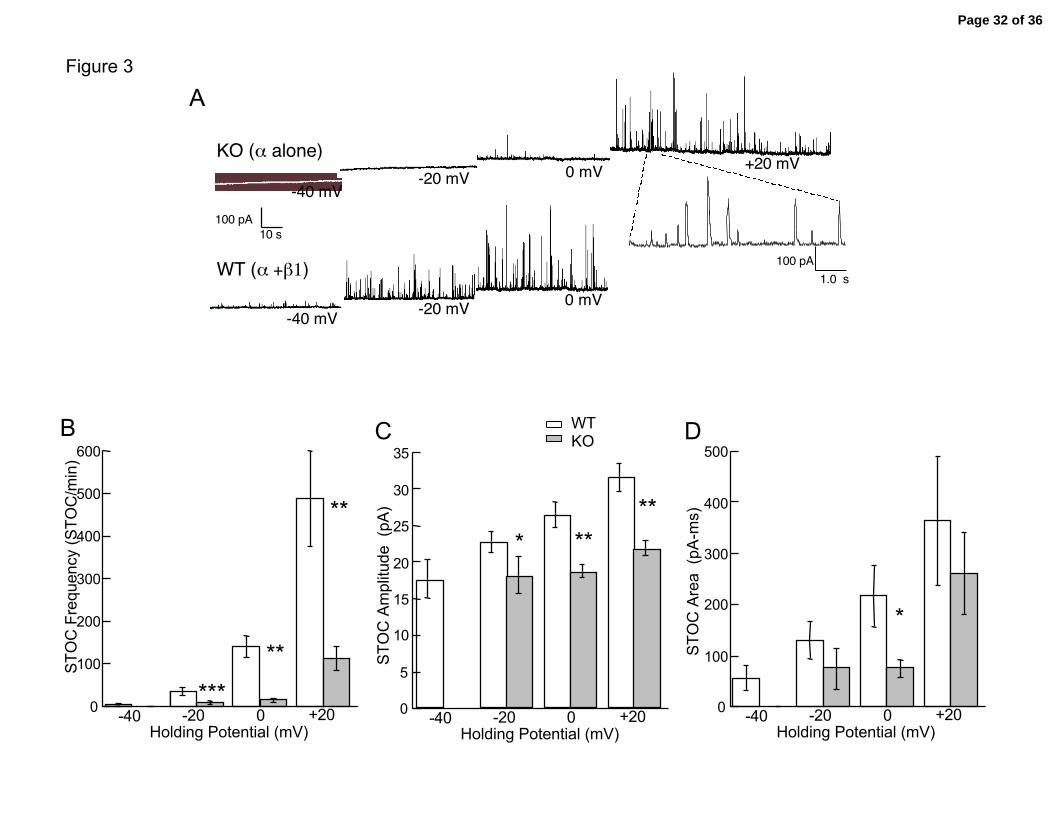
tracheal smooth muscle, we see a similar effect. Figure 3A shows that at -40 mV, wild
type cells (lowest trace) have significant BK current activity (seen in 6/6 cells at –40
mV), whereas β1 knockout smooth muscle cells (upper trace) show no (0/5 cells) or very
little activity (-20 mV, see Figure 3B). Nevertheless, as in the single channel
measurements (Figure 2B), given sufficient depolarization β1 knockout airway smooth
muscle demonstrate spontaneous transient BK currents (Figure 3A), albeit at a lower
frequency. Figure 3B depicts composite data for frequency; over the whole range of
holding potentials the knockout cells exhibited significantly lower transient BK currents
(Figure 3B, WT = 487 ± 112 STOC/min vs. KO = 113 ± 29 STOC/min at +20 mV, P <
0.01).
The effects of the β1 knockout on STOC amplitudes and areas (Figure 3C and
3D) were less dramatic than that seen with frequency (Figure 3B). Given that the STOC
amplitude should represents the number of channels opening following a spark event
Page 10 of 36

Semenov, et al. 11
(50), these data suggest that the β1 knockout causes a somewhat smaller recruitment of
BK channels during a calcium spark event (Figure 3C). In addition, wild type tracheal
BK channels appear to have a tendency towards more sustained opening as indicated by a
greater STOC area (Figure 3D). This is consistent with the longer open dwell times of
wild type BK channels (Figure 2C). In summary, these results indicate that BK channel
activity is significantly greater in wild type trachea relative to β1 knockout trachea, and
therefore would be expected to play a greater role at physiological voltages in controlling
calcium influx and constriction.
Knockout of the β1 subunit increases resting calcium and cholinergic evoked
calcium release.
In airway smooth muscle, calcium release via IP3 receptors initiates contraction.
Following calcium release, contraction is maintained by calcium influx channels (3).
Acutely isolated tracheal smooth muscle cells were loaded with Fura-2 to determine the
influence of the β1 subunit on calcium influx. We found that tracheal smooth muscle
have an approximately 50 nM increase in resting calcium levels in the β1 knockout (Inset
A in Figure 4A, 177 ± 11 nM in KO vs. 124 ± 0.8 nM in WT, P < 0.01). This indicates
that BK channels contribute to regulation of basal calcium.
During cholinergic evoked excitation, calcium increases from an early phase that
is mainly contributed by calcium release from the SR (calcium transient, B in Figure 4A)
and late sustained components that underlie sustained calcium influx (C, early sustained,
and D, late sustained, in Figure 4A). Because the β1 knockouts have an increased
Page 11 of 36

Semenov, et al. 12
baseline calcium, we quantified the relative calcium increases (subtracted from resting
calcium) at different phases of the calcium transient to determine the changes in evoked
calcium. Example traces where baseline calcium is normalized is shown in Figure 4A and
data is summarized in Figure 4B. In addition to the peak transient, we averaged the
sustained component between 15-17 seconds (component C) and 77-80 seconds
(component D) after carbachol addition (see methods for more detailed description) in
anticipation that the β1 knockout may have different effects of the components of the
calcium transients. The major effect of the β1 knockout is to increase the late sustained
component of the calcium influx (component D, KO = 131 ± 16 nM [Ca2+] vs. WT =
77.2 nM ± 12 nM [Ca2+], P < 0.005), with no significant effect on the calcium transient
and early sustained component (Figure 4A and 4C). In addition, although there was no
statistical difference between early sustained components (component C), we saw a
tendency for the β1 knockout to exhibit calcium oscillations much more frequently than
wild type cells (example in Figure 4B, Oscillations in KO = 12/26 cells vs. WT = 3/29
cells). Because very few wild type cells exhibited calcium oscillations, it would not be
meaningful to quantify calcium differences between knockout oscillating cells and wild
type cells. Therefore the oscillating cells were not included in further analysis.
Since a major function of potassium channels is to control membrane voltage, a
reasonable assumption is that increased calcium in the β1 knockouts arises from
depolarization and recruitment of voltage sensitive calcium channels. To these ends, we
utilized the L-type voltage-dependent calcium channel blocker, nifedipine (1 µM), to
observe the contribution of these channels to each component of cholinergic evoked
calcium transients. Indeed, nifedipine caused a substantial drop in all components of the
Page 12 of 36

Semenov, et al. 13
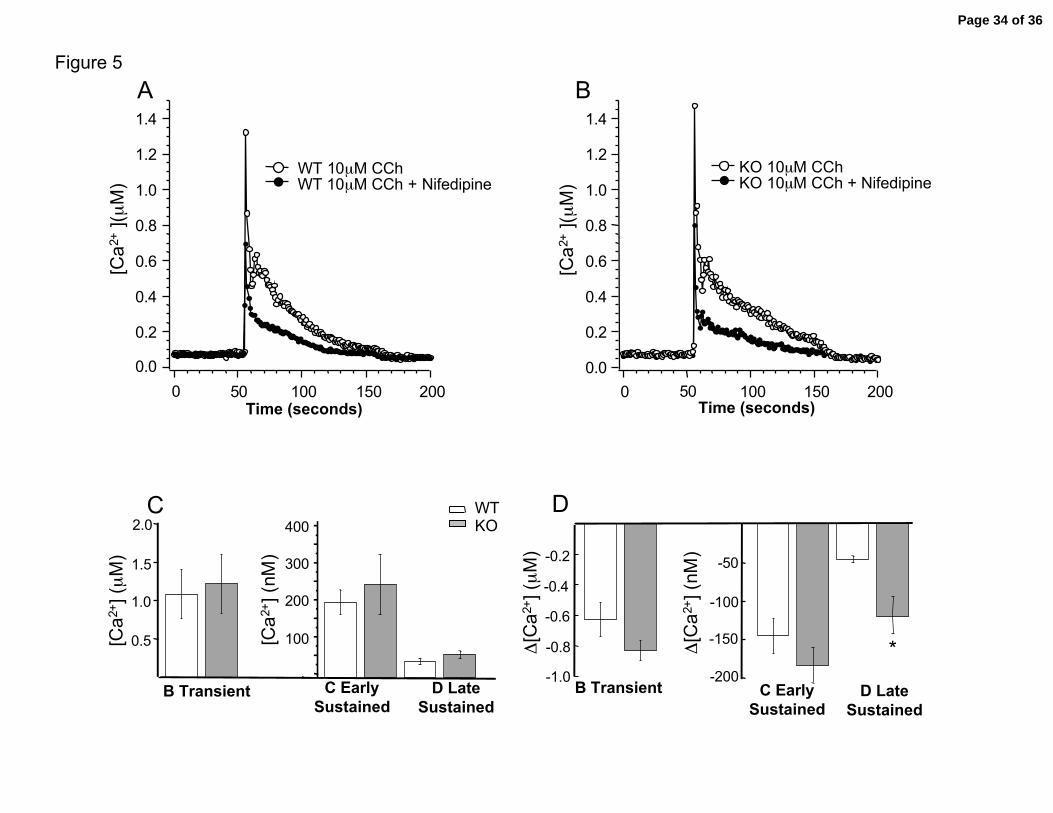
calcium transient (Figure 5A and 5B) in both wild type and knockout cells. Figure 5C
summary shows that the average response of all components are similar between wild
type and knockout muscle blocked with nifedipine. Particularly important was the
observation that nifedipine eliminated the difference in the late sustained calcium influx
(component D) observed between wild type and β1 knockout cells (summary in Figure
4B, nifedipine and Figure 5C, P = 0.22 WT vs. KO). This can also be seen when
measuring the relative difference in calcium evoked by carbachol before and after block
with nifedipine (Figure 5A and 5B, data are paired with the same tracheal cell). The
relative changes are summarized in panel 5D. Consistent with the above results,
knockouts show a larger nifedipine sensitive late sustained component than wild type
trachea cells (Figure 5D, WT = 48 nM ± 5 vs. KO = 120 nM ± 33, P < 0.02). A similar
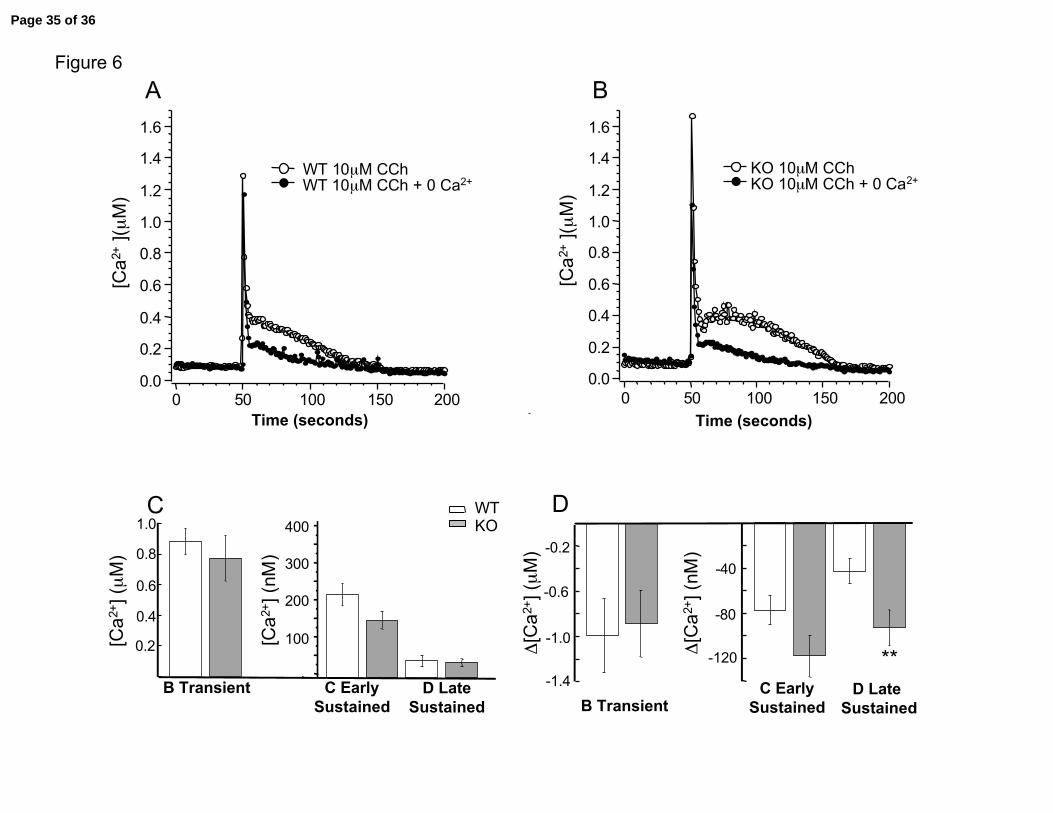
result was obtained with perfusion of zero extracellular calcium (with 1 mM EGTA
calcium chelator) (Figure 6A and 6B). As in nifedipine experiments, the significant
difference between wild type and knockout calcium transients during late sustained
component seen in 2 mM calcium solutions (Figure 4B) was eliminated with zero
calcium (Figure 6C, WT = 46 nM ± 13 vs. KO = 41 nM ± 10). As well, the largest
change in calcium mediated by the β1 subunit is seen during the late sustained
component (Figure 6D, WT = 43 nM ± 11 vs. KO = 93 nM ± 16 ). In summary, these
results indicate that increased calcium in the knockout trachea can be accounted for by
increased activation of voltage-dependent calcium channels. It was also observed that
although the β1 knockout showed an increased frequency of cells showing calcium
oscillations, block of voltage-dependent calcium channels or 0 calcium solutions did not
eliminate calcium oscillations from cells (data not shown).
Page 13 of 36

Semenov, et al. 14
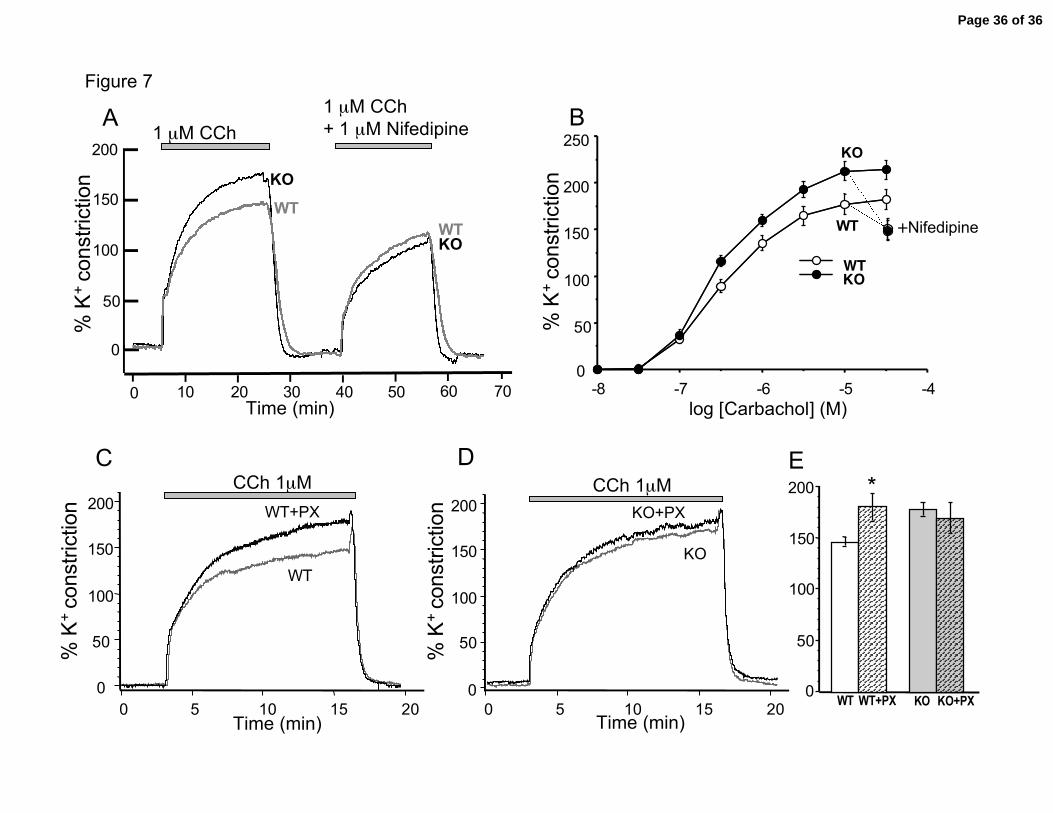
BK Channel β1 Knockout Causes Increased Constriction of Trachea
Figure 7A shows cholinergic evoked constrictions normalized to trachea
responses in high potassium. Consistent with the expectation that BK channels control
airway constriction, the data shows that the β1 knockouts have a larger carbachol-
induced constriction than the wild type mice. Summary dose-response curves (Figure 7B)
indicate that the main effect of the β1 knockout is an increase in the maximal response,
rather than a shift in the dose-response curve. Of note, although some knockouts showed
calcium oscillations, we did not see oscillation in carbachol-induced constrictions.
The increased constriction of the β1 knockouts is accounted for by increased
contribution of voltage-dependent calcium channels during sustained contractions.
The most likely role of BK potassium channels is to oppose membrane
depolarization during agonist induced constriction. A reasonable hypothesis is that
membrane depolarization recruits voltage-dependent calcium channels and increased
calcium influx and constriction. Consistent with this hypothesis, we found that the β1
knockouts relax to the same relative constriction as wild type mice (Figure 7A, right
panel, Figure 7B dose-response + nifedipine). This indicates that the increased
constriction of the knockouts is largely accounted for by increased calcium influx through
L-type voltage-dependent calcium channels.
It is fairly well established that two sources of calcium underlie tracheal
constriction; an early transient component that is dependent on agonist-induced calcium
release from ER calcium stores and a subsequent sustained component that is dependent
on calcium influx from plasma membrane channels. Consistent with the nifedipine
Page 14 of 36

Semenov, et al. 15
experiments above (Figure 5), the β1 knockouts show the greatest effect on constriction
during the sustained component where voltage-dependent calcium channels are expected
to play a role. This is shown in representative trace in Figure 7A. Nifedipine block of
voltage-dependent calcium channels brings constriction during the sustained component
to similar levels between the knockout and wild-type (Figure 7A and 7B).
In vascular smooth muscle, block of BK channels in wild type cells shows similar
levels of constriction as β1 knockouts (4). This indicates that the absence of the β1
subunit renders BK channels nonfunctional in controlling constriction. In tracheal smooth
muscle, we see a similar effect. Figure 7D and 7E shows that the KO trachea had no
significant difference in cholinergic evoked constriction in KO in the presence or absence
of paxilline block, indicating BK channel activity do not significantly contribute to
relaxation in the absence of the β1 subunit. In contrast, wild type trachea show a
significant difference with and without paxilline block (Figure 7C and 7E).
DISCUSSION
Airway smooth muscle presents a medically important model for studying the role
of ion channels during receptor-coupled activation of calcium transients. Unlike vascular
smooth muscle, where voltage-dependent calcium channel are a major calcium conduit
mediating contraction, the role of voltage-dependent calcium channels and other calcium
influx pathways in airway smooth muscle is poorly defined. As such, pharmaceuticals
targeted to ion channels in airway have not been utilized to treat asthma.
These data demonstrate that BK channels in β1 knockout airway smooth muscle
exhibit a large reduction in channel openings as indicated by single channel activity and
Page 15 of 36

Semenov, et al. 16
STOC frequency. As a consequence, BK channel contribution to relaxation is perturbed
to a sufficient extent that pharmacological block of BK channels shows that same relative
constriction as the β1 knockout trachea (Figure 7C-E). This is similar to vascular smooth
muscle wherein iberiotoxin block of wild type vessels induced constriction, but had no
further constrictive effect on β1 knockout vessels (4). This is in contrast to bladder
smooth muscle where the β1 knockout shows a partial effect on constriction as compared
to iberiotoxin block (36). A distinction between tonic smooth muscles such as vascular
and trachea muscle, as compared to bladder smooth muscle that is phasic in excitation, is
that large depolarizations occur in bladder that are likely to complement BK activation in
the β1 knockout. Indeed, we see much smaller differences in single channel and STOC
activity of WT and KO at depolarizing voltages.
Tao et al (43) demonstrated that cholinergic evoked constriction in bovine airway
is largely independent of voltage-activated calcium channels unless sarcoplasmic
reticulum calcium stores are depleted or BK potassium channels are blocked. In the latter
case, the mode of contraction coupling switches to an excitation, voltage-dependent
coupling. Our findings utilizing the β1 knockout in mouse trachea indicate that there is
more of a partial contribution, rather than a switch in coupling between pharmaco- and
excitation evoked contraction. In wild type mice, voltage-dependent calcium channels
appear to contribute approximately 17 % of the carbachol induced contraction at
saturating carbachol doses (Figure 7B). In the absence of the β1 subunit, BK channel
activity decreases and the contribution of voltage-dependent calcium channels increase to
approximately 30% of the constriction (Figure 7B). This is a smaller contribution than β1
subunits appear to have in aortic smooth muscle. In those studies, maximal contraction is
Page 16 of 36

Semenov, et al. 17
increased approximately 65% by the β1 knockout (37). Thus, BK potassium channels in
tracheal smooth muscle make a significant, but smaller contribution, to agonist induced
relaxation than in vascular muscle.
The calcium pathways downstream of voltage-dependent calcium channels that
are regulated by BK channels are likely to be complex. The consequence of the β1
knockout was to increase resting calcium levels and increases in the sustained component
of calcium influx during cholinergic evoked signaling. These were translated to increased
cholinergic evoked constriction. A conclusion that may be drawn is that the regulation of
membrane voltage by the BK channel is important, albeit less apparent, until these
channels are blocked or by deletion of the β1 subunit. The hyperpolarizing activity of BK
channels is likely to hold membrane voltage during cholinergic contractions below the
operational voltages where voltage-dependent calcium channels and excitation-
contraction coupling contribute to airway constriction (Bourreau 1993). This may
reconcile the fact that voltage-dependent calcium channels are regarded to provide a
minority of the calcium for contraction (Janssen 2002), yet the potassium channels would
have an important role such as is observed during β-adrenergic mediated relaxation.
An interesting finding was that the β1 knockout mice had a high frequency of
cells showing oscillatory calcium changes following cholinergic activation (Figure 4B).
However we did not see a similar effect on constriction, which were all tonic in response
to high potassium or cholinergic evoked constriction. Thus, it is as yet unclear how the
β1 knockout-induced calcium oscillations influence the muscle contractile properties in
the trachea. Other examples exist where perturbing BK channels promotes oscillations in
trachea. Oscillatory contractions have been described for guinea pig tracheal smooth
Page 17 of 36

Semenov, et al. 18
muscle cells treated with the BK channel blockers charybdotoxin and iberiotoxin (48,
49). However, these studies did not investigate calcium changes that may underlie the
oscillatory contractions. More recent studies in bronchioles have observed slow
oscillatory calcium transients in response to depolarization with potassium (35). This may
be considered analogous to BK channel block, or the β1 knockout since blocking of
potassium channels presumably depolarizes the cell. However, slow calcium oscillations
were correlated with twitching rather than constriction of airway (35).
The fact that the calcium oscillations are not affected by nifedipine or 0 external
calcium suggests that the β1 subunit has other effects in addition to their actions on
membrane voltage or on calcium influx through plasmallema channels. In this regard, the
increase in resting calcium concentration observed in the β1 knockout could play a role.
Increases in global calcium have been shown to promote SR calcium loading and thereby
sustain calcium release (5). As well, increases in cytosolic calcium concentrations and SR
calcium stores promote IP3 receptor activation and calcium oscillations (2, 13). Indeed,
the mechanism by which knockout of the β1 subunit causes calcium oscillations and
perhaps affects other currents requires further study.
In summary, these findings show that tracheal smooth muscle require the
accessory β1 subunit to promote normal BK channel activation. Despite the predominant
role of agonist induced signaling in trachea, BK channels have a significant role in
controling resting calcium and cholinergic evoked calcium influx contributed by voltage-
dependent calcium channels.
ACKNOWLEDGEMENTS
Page 18 of 36

Semenov, et al. 19
This work was supported by the Sandler Program for Asthma Research to R.B.
and J.T.H., and a National American Heart Association grant 0335007N to R.B. We
thank Dr. Qing H. Chen and David Petrik for critical reading of the manuscript.
REFERENCES
1. Barnes PJ. Clinical studies with calcium antagonists in asthma. British Journal of
Clinical Pharmacology 20 Suppl 2: 289S-298S, 1985.
2. Bezprozvanny I, Watras J, and Ehrlich BE. Bell-shaped calcium-response
curves of Ins(1,4,5)P3- and calcium-gated channels from endoplasmic reticulum of
cerebellum. Nature 351: 751-754, 1991.
3. Bourreau JP. Cross talk between plasma membrane and sarcoplasmic reticulum
in canine airway smooth muscle. Biol Signals 2: 272-283, 1993.
4. Brenner R, Perez GJ, Bonev AD, Eckman DM, Kosek JC, Wiler SW,
Patterson AJ, Nelson MT, and Aldrich RW. Vasoregulation by the beta1 subunit of the
calcium-activated potassium channel. Nature 407: 870-876., 2000.
5. Burdyga T and Wray S. Action potential refractory period in ureter smooth
muscle is set by Ca sparks and BK channels. Nature 436: 559-562, 2005.
6. Calderone V. Large-conductance, ca(2+)-activated k(+) channels: function,
pharmacology and drugs. Current Medicinal Chemistry 9: 1385-1395, 2002.
7. Corompt E, Bessard G, Lantuejoul S, Naline E, Advenier C, and Devillier P.
Inhibitory effects of large Ca2+-activated K+ channel blockers on beta-adrenergic- and
NO-donor-mediated relaxations of human and guinea-pig airway smooth muscles.
Naunyn-Schmiedebergs Archives of Pharmacology 357: 77-86, 1998.
Page 19 of 36

Semenov, et al. 20
8. Cox DH and Aldrich RW. Role of the beta1 subunit in large-conductance
Ca(2+)-activated K(+) channel gating energetics. Mechanisms of enhanced Ca(2+)
sensitivity. J Gen Physiol 116: 411-432., 2000.
9. Cox DH, Cui J, and Aldrich RW. Allosteric gating of a large conductance Ca-
activated K+ channel. J Gen Physiol 110: 257-281, 1997.
10. Dick GM and Sanders KM. (Xeno)estrogen sensitivity of smooth muscle BK
channels conferred by the regulatory beta1 subunit: a study of beta1 knockout mice. J
Biol Chem 276: 44835-44840, 2001.
11. Dworetzky SI, Boissard CG, Lum-Ragan JT, McKay MC, Post-Munson DJ,
Trojnacki JT, Chang CP, and Gribkoff VK. Phenotypic Alteration of a Human BK
(hSlo) Channel by hSlobeta Subunit Coexpression: Changes in Blocker Sensitivity,
Activation/Relaxation and Inactivation Kinetics, and Protein Kinase A Modulation. J
Neurosci 16: 4543-4550, 1996.
12. Erxleben C, Everhart AL, Romeo C, Florance H, Bauer MB, Alcorta DA,
Rossie S, Shipston MJ, and Armstrong DL. Interacting effects of N-terminal variation
and strex-exon splicing on slo potassium channel regulation by calcium, phosphorylation
and oxidation. J Biol Chem, 2002.
13. Falcke M, Li Y, Lechleiter JD, and Camacho P. Modeling the dependence of
the period of intracellular Ca2+ waves on SERCA expression. Biophys J 85: 1474-1481,
2003.
14. Farley JM and Miles PR. The sources of calcium for acetylcholine-induced
contractions of dog tracheal smooth muscle. Journal of Pharmacology & Experimental
Therapeutics 207: 340-346, 1978.
Page 20 of 36

Semenov, et al. 21
15. Gribkoff VK, Starrett JE, Jr., and Dworetzky SI. The pharmacology and
molecular biology of large-conductance calcium- activated (BK) potassium channels. Adv
Pharmacol 37: 319-348, 1997.
16. Hagen BM, Bayguinov O, and Sanders KM. Beta 1-subunits are required for
regulation of coupling between Ca2+ transients and Ca2+-activated K+ (BK) channels by
protein kinase C. Am J Physiol Cell Physiol 285: C1270-1280, 2003.
17. Jackson WF. Ion channels and vascular tone. Hypertension 35: 173-178., 2000.
18. Jaggar JH, Wellman GC, Heppner TJ, Porter VA, Perez GJ, Gollasch M,
Kleppisch T, Rubart M, Stevenson AS, Lederer WJ, Knot HJ, Bonev AD, and
Nelson MT. Ca2+ channels, ryanodine receptors and Ca(2+)-activated K+ channels: a
functional unit for regulating arterial tone. Acta Physiol Scand 164: 577-587, 1998.
19. Janssen LJ. Ionic mechanisms and Ca(2+) regulation in airway smooth muscle
contraction: do the data contradict dogma? American Journal of Physiology - Lung
Cellular & Molecular Physiology 282: L1358-1365, 2002.
20. Kaczorowski GJ, Knaus HG, Leonard RJ, McManus OB, and Garcia ML.
High-conductance calcium-activated potassium channels; structure, pharmacology, and
function. J Bioenerg Biomembr 28: 255-267, 1996.
21. Karaki H, Ozaki H, Hori M, Mitsui-Saito M, Amano K, Harada K,
Miyamoto S, Nakazawa H, Won KJ, and Sato K. Calcium movements, distribution,
and functions in smooth muscle. Pharmacological Reviews 49: 157-230, 1997.
22. Knaus HG, Folander K, Garcia-Calvo M, Garcia ML, Kaczorowski GJ,
Smith M, and Swanson R. Primary sequence and immunological characterization of
Page 21 of 36

Semenov, et al. 22
beta-subunit of high conductance Ca(2+)-activated K+ channel from smooth muscle. J
Biol Chem 269: 17274-17278, 1994.
23. Knaus HG, Garcia-Calvo M, Kaczorowski GJ, and Garcia ML. Subunit
composition of the high conductance calcium-activated potassium channel from smooth
muscle, a representative of the mSlo and slowpoke family of potassium channels. J Biol
Chem 269: 3921-3924, 1994.
24. Kotlikoff MI and Kamm KE. Molecular mechanisms of beta-adrenergic
relaxation of airway smooth muscle. Annual Review of Physiology 58: 115-141, 1996.
25. Lofdahl CG and Barnes PJ. Calcium channel blockade and asthma--the current
position. European Journal of Respiratory Diseases 67: 233-237, 1985.
26. McCobb DP, Fowler NL, Featherstone T, Lingle CJ, Saito M, Krause JE,
and Salkoff L. A human calcium-activated potassium channel gene expressed in vascular
smooth muscle. Am J Physiol 269: H767-777, 1995.
27. McManus OB, Helms LM, Pallanck L, Ganetzky B, Swanson R, and
Leonard RJ. Functional role of the beta subunit of high conductance calcium- activated
potassium channels. Neuron 14: 645-650, 1995.
28. Meera P, Wallner M, Jiang Z, and Toro L. A calcium switch for the functional
coupling between alpha (hslo) and beta subunits (KV,Ca beta) of maxi K channels. FEBS
Lett 382: 84-88, 1996.
29. Meredith AL, Thorneloe KS, Werner ME, Nelson MT, and Aldrich RW.
Overactive bladder and incontinence in the absence of the BK large conductance Ca2+-
activated K+ channel. J Biol Chem 279: 36746-36752, 2004.
Page 22 of 36

Semenov, et al. 23
30. Nimigean CM and Magleby KL. The beta subunit increases the Ca2+ sensitivity
of large conductance Ca2+-activated potassium channels by retaining the gating in the
bursting states. J Gen Physiol 113: 425-440, 1999.
31. Nimigean CM and Magleby KL. Functional coupling of the beta(1) subunit to
the large conductance Ca(2+)-activated K(+) channel in the absence of Ca(2+). Increased
Ca(2+) sensitivity from a Ca(2+)-independent mechanism. J Gen Physiol 115: 719-736.,
2000.
32. Patton C, Thompson S, and Epel D. Some precautions in using chelators to
buffer metals in biological solutions. Cell Calcium 35: 427-431, 2004.
33. Pelaia G, Gallelli L, Vatrella A, Grembiale RD, Maselli R, De Sarro GB, and
Marsico SA. Potential role of potassium channel openers in the treatment of asthma and
chronic obstructive pulmonary disease. Life Sciences 70: 977-990, 2002.
34. Perez GJ, Bonev AD, Patlak JB, and Nelson MT. Functional coupling of
ryanodine receptors to KCa channels in smooth muscle cells from rat cerebral arteries. J
Gen Physiol 113: 229-238, 1999.
35. Perez JF and Sanderson MJ. The frequency of calcium oscillations induced by
5-HT, ACH, and KCl determine the contraction of smooth muscle cells of
intrapulmonary bronchioles. J Gen Physiol 125: 535-553, 2005.
36. Petkov GV, Bonev AD, Heppner TJ, Brenner R, Aldrich RW, and Nelson
MT. Beta1-subunit of the Ca2+-activated K+ channel regulates contractile activity of
mouse urinary bladder smooth muscle. J Physiol 537: 443-452., 2001.
37. Pluger S, Faulhaber J, Furstenau M, Lohn M, Waldschutz R, Gollasch M,
Haller H, Luft FC, Ehmke H, and Pongs O. Mice with disrupted BK channel beta1
Page 23 of 36

Semenov, et al. 24
subunit gene feature abnormal Ca(2+) spark/STOC coupling and elevated blood pressure.
Circulation Research 87: E53-60, 2000.
38. Ramanathan K, Michael TH, Jiang GJ, Hiel H, and Fuchs PA. A molecular
mechanism for electrical tuning of cochlear hair cells. Science 283: 215-217, 1999.
39. Sausbier M, Arntz C, Bucurenciu I, Zhao H, Zhou XB, Sausbier U, Feil S,
Kamm S, Essin K, Sailer CA, Abdullah U, Krippeit-Drews P, Feil R, Hofmann F,
Knaus HG, Kenyon C, Shipston MJ, Storm JF, Neuhuber W, Korth M, Schubert R,
Gollasch M, and Ruth P. Elevated blood pressure linked to primary hyperaldosteronism
and impaired vasodilation in BK channel-deficient mice. Circulation 112: 60-68, 2005.
40. Snetkov VA and Ward JP. Ion currents in smooth muscle cells from human
small bronchioles: presence of an inward rectifier K+ current and three types of large
conductance K+ channel. Exp Physiol 84: 835-8460, 1999.
41. Somlyo AV and Somlyo AP. Electromechanical and pharmacomechanical
coupling in vascular smooth muscle. J Pharmacol Exp Ther 159: 129-145, 1968.
42. Tanaka Y, Meera P, Song M, Knaus HG, and Toro L. Molecular constituents
of maxi KCa channels in human coronary smooth muscle: predominant alpha + beta
subunit complexes. J Physiol (Lond) 502: 545-557, 1997.
43. Tao L, Huang Y, and Bourreau JP. Control of the mode of excitation-
contraction coupling by Ca(2+) stores in bovine trachealis muscle. American Journal of
Physiology - Lung Cellular & Molecular Physiology 279: L722-732, 2000.
44. Vogalis F, Vincent T, Qureshi I, Schmalz F, Ward MW, Sanders KM, and
Horowitz B. Cloning and expression of the large-conductance Ca(2+)-activated K+
channel from colonic smooth muscle. Am J Physiol 271: G629-639, 1996.
Page 24 of 36

Semenov, et al. 25
45. Wallner M, Meera P, Ottolia M, Kaczorowski GJ, Latorre R, Garcia ML,
Stefani E, and Toro L. Characterization of and modulation by a beta-subunit of a human
maxi KCa channel cloned from myometrium. Receptors Channels 3: 185-199, 1995.
46. Weiger TM, Holmqvist MH, Levitan IB, Clark FT, Sprague S, Huang WJ,
Ge P, Wang C, Lawson D, Jurman ME, Glucksmann MA, Silos-Santiago I,
DiStefano PS, and Curtis R. A novel nervous system beta subunit that downregulates
human large conductance calcium-dependent potassium channels. J Neurosci 20: 3563-
3570, 2000.
47. Weiss RL and Weiss GB. Relationship between 45Ca movements, different
calcium components and responses to acetylcholine and potassium in tracheal smooth
muscle. Journal of Biological Chemistry 261: 10264-10270, 1986.
48. Yagi Y, Kuwahara M, and Tsubone H. Ca2+-activated K+ channel blockers
induce PKC modulated oscillatory contractions in guinea pig trachea. Comp Biochem
Physiol C Toxicol Pharmacol 131: 511-519, 2002.
49. Yagi Y, Kuwahara M, and Tsubone H. ChTX induces oscillatory contraction in
guinea pig trachea: role of cyclooxygenase-2 and PGE2. Am J Physiol Lung Cell Mol
Physiol 284: L1045-1054, 2003.
50. ZhuGe R, Fogarty KE, Tuft RA, and Walsh JV, Jr. Spontaneous Transient
Outward Currents Arise from Microdomains Where BK Channels Are Exposed to a
Mean Ca(2+) Concentration on the Order of 10 microM during a Ca(2+) Spark. J Gen
Physiol 120: 15-28, 2002.
FIGURE LEGENDS
Page 25 of 36

Semenov, et al. 26
Figure 1. The BK Channel β1 subunit is expressed in airway smooth muscle tissues.
Expression is assayed from a β-galactosidase reporter gene targeted to the BK channel
beta1 locus in mice (KO tissues). A. Filleted whole-mount KO trachea showing staining
in trachea smooth muscle. B. Wild type control. C. Frozen section of trachea in KO. D.
Frozen section of trachea in wild type tissue E. Magnification of trachea showing staining
in the smooth muscle layer. Arrows indicate smooth muscle layers.
Figure 2. Knockout of the β1 subunit reduces BK channel opening probability. A.
Single channel recording of BK channels from wild type and knockout trachea smooth
muscle cells assayed in excised patch, inside/out configuration (Internal solution was 7
µM buffered calcium). B. Summary data showing Po and C. open dwell times for wild
type (WT, open column) and knockout (KO, shaded column). Scale bar indicates 1
second for –40 mV traces, and 0.1 second for +40 mV traces. N= 6 WT, 8 KO. Heights
of columns and vertical lines represent means ± sem. Single asterisk indicate difference
of P < 0.05.
Figure 3. Spontaneous transient outward currents in isolated tracheal smooth
muscle cells. A. Examples of spontaneous transient BK current in wild type and
knockout airway smooth muscle cells. B. Summary of STOC frequency. C. Summary of
STOC amplitude. D. Summary of STOC area. Data is for N= 6-8 cells for WT and β1
KO. At –40 mV KOs did not show any STOCs in 7/7 cells whereas WT showed STOCS
in 3/8 cells. Data are mean ± sem. Triple asterisk represent P < 0.001, double asterisk P
difference < 0.01 and single asterisk represent P < 0.05.
Page 26 of 36

Semenov, et al. 27
Figure 4. Carbachol-induced calcium transients in wild type and knockout tracheal
smooth muscle cells. A. Example of Fura-2 measured calcium transients from wild type
(open circles) and knockout (filled circles) tracheal smooth muscle cells. Left inset shows
an enlargement of resting calcium that is not baseline subtracted (Note left and right
insets Y-scale is in nM). Large trace and right inset knockout traces are baseline
subtracted to adjust knockout to the same baseline as wild type. Components of calcium
are labeled A, resting calcium, B transient phase, C early sustained phase, D late
sustained phase. B. Example of Fura-2 measured calcium transients from knockout
tracheal smooth muscle cells that has oscillatory calcium changes. C. Summary data
comparing average values for different components in wild type (open columns) and
knockout (gray columns). Significant difference is seen at resting calcium (P = 0.0004,
N= 28 WT, 24 KO), during the sustained component D (P = 0.0062, N = 14 WT, 12 KO).
Sustained calcium components C and D were measured from 15-17 seconds, and 77-80
seconds, respectively, after addition of carbachol. Cells with oscillatory calcium
transients were excluded from these analysis.
Figure 5. Effect of nifedipine on carbachol-induced calcium transients. Example
traces of carbachol induced calcium transients before (open circle) and after nifedipine
block of voltage-gated calcium channels (closed circles) for A, Wild type cell, and B, β1
knockout cell. C. Summary data comparing average values for different components in
wild type (open boxes) and knockout (gray boxes) during calcium channel block with
nifedipine. No significant difference is seen at any component. D. Summary of relative
Page 27 of 36

Semenov, et al. 28
change of different components calculated as calcium after nifedipine subtracted from
before nifedipine (Δ[Ca2+]). Results are from paired experiments. Asterisk indicates
significance of P = 0.025. N = 13 WT, 11 KO.
Figure 6. Effect of removing calcium on carbachol-induced calcium transients.
Example traces of carbachol induced calcium transients before (open circle) and after
addition of 0 calcium (closed circles), 1 mM EGTA Krebs solution for A, Wild type cell,
and B, β1 knockout cell. C. Summary data comparing average values for different
components in wild type (open column) and knockout (gray column) during cholinergic
evoked calcium in 0 external calcium. No significant difference is seen at any
component. D. Summary of relative change of different components following
extracellular calcium removal. Asterisk indicates significance of P = 0.028. N = 14 WT,
12 KO.
Figure 7. β1 knockouts have greater cholinergic-evoked contraction of trachea. A.
Example of carbachol evoked contraction before and after 1 µM nifedipine. Contraction
traces are normalized to high potassium induced contractions. B. Cumulative dose
response curve to carbachol. Last dose (3x10-5 carbachol) is repeated as previous dose
with 1 µM nifedipine). P < 0.05 for 2x10-7 to 1x10-5 carbachol. N = 14 wild type, 14
knockout. Effect of BK channel block with paxilline (5 µM) in wild type, C, Example of
constriction to 1 µM carbachol without (dark trace) and with 5 µM paxilline block of BK
channels in WT and D, β1 knockout tracheas. E. Summary data for C and D.
Page 28 of 36

Semenov, et al. 29
Page 29 of 36
A
B C D E
Figure 1
Page 30 of 36
WT KO-40 mV
+40 mV
20 pA0.1 s1 s
oc
oc
A
Figure 2
Ope
n D
wel
l Tim
e (m
s)
0
0.5
1.0
1.5
2.0
2.5
WT KO
-40 mV-40 mV
0
0.01
0.02
0.03
WT KO 0
0.2
0.4
0.6
0.8
1.0+40 mV
WT KO
Ope
n P
roba
bilit
y
B C
**
01
23
456
7+40 mV
WT KO
Page 31 of 36
100 pA10 s
KO (α alone)
WT (α +β1)
-20 mV
0 mV
0 mV +20 mV
A
WT KO
Figure 3
0
5
10
15
20
25
30
35
-40 -20 0 +20Holding Potential (mV)
ST
OC
Am
plitu
de (
pA)
C
* ****
0
100
200
300
400
500
600
-40 -20 0 +20Holding Potential (mV)
ST
OC
Fre
quen
cy (
ST
OC
/min
)
B
***
**
**
0
100
200
300
400
500
-40 -20 0 +20Holding Potential (mV)
ST
OC
Are
a (
pA-m
s)
D
*
1.0 s100 pA
-20 mV-40 mV
-40 mV
Page 32 of 36
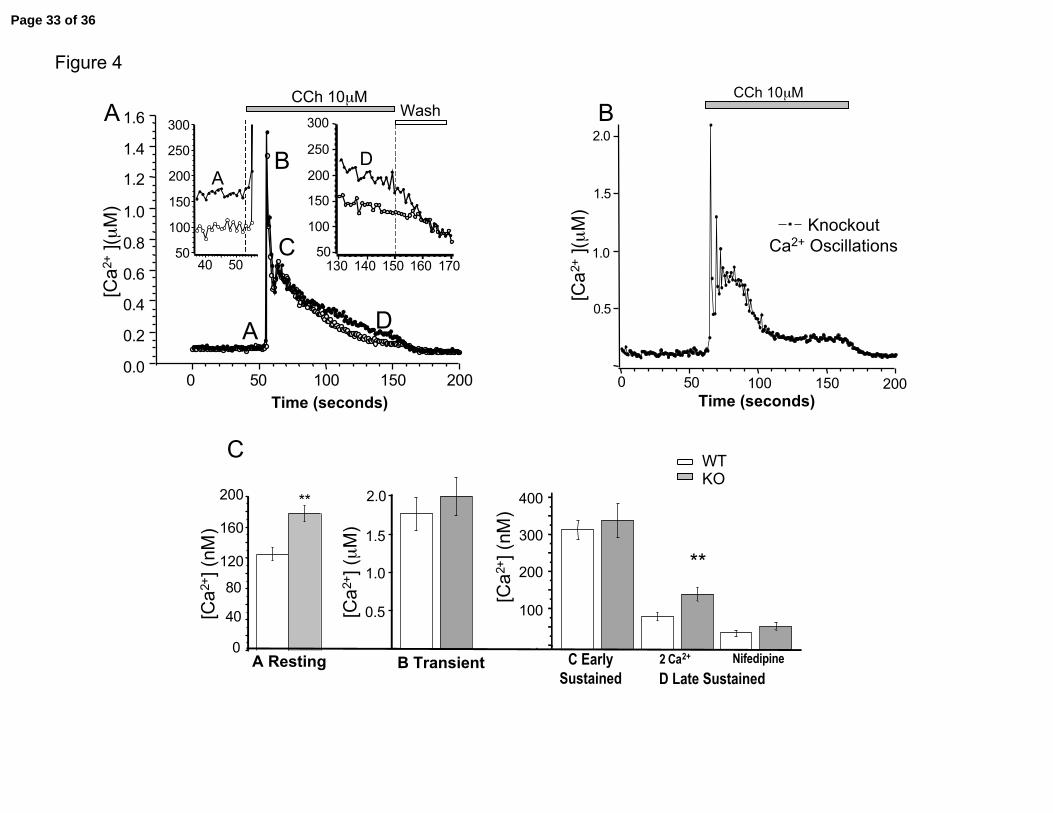
Figure 4
0
40
80
120
160
200 **
A Resting
[Ca
2+] (
nM)
WT KO
0.5
**
C Early Sustained
2 Ca2+B Transient
C
[Ca
2+] (
µM
)
100
200
300
400
1.0
1.5
2.0
[Ca
2+] (
nM)
Nifedipine
D Late Sustained
CCh 10µMCCh 10µM
0 50 100 150 200
1.0
2.0
1.5
0.5
KnockoutCa2+ Oscillations
B
Time (seconds)
[Ca
2+ ](
µM
)
200150100500
Wash300
250
200
150
100
50170160150140130
Time (seconds)
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
[C
a2+
](µ
M)
A300
250
200
150
100
50
5040
AD
A
B
C
D
Page 33 of 36
WT KO
[Ca
2+] (
nM)
0.5
C Early Sustained
D Late Sustained
B Transient
[Ca
2+] (
µM
)
100
200
300
400
1.0
1.5
2.0
-0.2
-0.4
-0.6
-0.8
-1.0
Δ[C
a2+
] (µ
M)
B Transient
-50
-100
-150
-200Δ
[Ca
2+] (
nM)
C Early Sustained
D Late Sustained
*
C D
Figure 5
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
200150100500
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
200150100500
A[C
a2+
](µ
M)
[Ca
2+ ](
µM
)
B
Time (seconds)Time (seconds)
WT 10µM CCh WT 10µM CCh + Nifedipine
KO 10µM CCh KO 10µM CCh + Nifedipine
Page 34 of 36
WT KO
[Ca
2+] (
nM)
C Early Sustained
D Late Sustained
B Transient
[Ca
2+] (
µM
)
100
200
300
400
-0.2
-0.6
-1.0
-1.4
Δ[C
a2+
] (µ
M)
B Transient
-40
-80
Δ[C
a2+
] (nM
)C Early
Sustained D Late
Sustained
**
C D
Figure 6
0.2
0.4
0.6
0.8
1.0
A B
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
200150100500
[Ca
2+ ](
µM
)
Time (seconds)
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
200150100500
[Ca
2+ ](
µM
)
Time (seconds)
WT 10µM CCh WT 10µM CCh + 0 Ca2+
KO 10µM CCh KO 10µM CCh + 0 Ca2+
-120
Page 35 of 36
-4-5-6-7-80
50
100
150
200
250
log [Carbachol] (M)
WTKO
+Nifedipine
KO
WT
0 10 20 30 40 50 60 70
200
150
100
50
0
Time (min)
A B1 µM CCh
1 µM CCh+ 1 µM Nifedipine
KO
WTWTKO
Figure 7%
K+
con
stric
tion 200
150
100
50
020151050
CCh 1µM
WT
WT+PX
Time (min)
CCh 1µM200
150
100
50
020151050
KO
KO+PX
Time (min)
200
150
100
50
0WT WT+PX KO KO+PX
C D E*
% K
+ c
onst
rictio
n
% K
+ c
onst
rictio
n
% K
+ c
onst
rictio
n
Page 36 of 36