pairing behavior and reproduction in hyalella azteca as sensitive endpoints for detecting long-term...
TRANSCRIPT

Pe
SMa
b
c
a
ARR1A
KADPPP
1
osf1Sttemnpab
L
0h
Aquatic Toxicology 144– 145 (2013) 59– 65
Contents lists available at ScienceDirect
Aquatic Toxicology
jou rn al hom ep age: www.elsev ier .com/ locate /aquatox
airing behavior and reproduction in Hyalella azteca as sensitivendpoints for detecting long-term consequences of pesticide pulses
igne Pedersena, Annemette Palmqvista, Pernille Thorbekb,ick Hamerb, Valery Forbesc,∗
Department of Environmental, Social and Spatial Change, Roskilde University, DenmarkSyngenta Jealott’s Hill, Bracknell, United KingdomSchool of Biological Sciences, University of Nebraska Lincoln, USA
r t i c l e i n f o
rticle history:eceived 18 June 2013eceived in revised form9 September 2013ccepted 25 September 2013
eywords:mphipodelayed toxicityermethrinulse exposure
a b s t r a c t
The aim of the present study was to examine acute and delayed effects of pulse exposure of the pyrethroidpesticide, permethrin, on precopulatory pairs of Hyalella azteca. Pairs of H. azteca were exposed to a single1 h pulse of different nominal concentrations of permethrin: 0, 0.3, 0.9 or 2.7 �g/L. During exposure, pair-ing behavior was observed, and during a 56 day post-exposure period the treatments were monitoredfor pairing behavior, survival and reproductive output. All permethrin-exposed pairs separated withinminutes during exposure and shortly thereafter became immobile; however they regained mobility aftertransfer to clean water. The time to re-form pairs was significantly longer in all tested concentrationscompared to the control, although all surviving pairs re-formed within the 56 day test period. Neverthe-less not all pairs exposed to 0.9 and 2.7 �g/L reproduced. Furthermore the numbers of juveniles producedby pairs exposed to 0.9 and 2.7 �g/L, but not 0.3 �g/L, were lower throughout the entire post-exposure
yrethroids period compared to the control groups, and the total numbers of juveniles produced during 56 days weresignificantly lower in organisms exposed to 0.9 and 2.7 �g/L, but not 0.3 �g/L, compared to the controlgroups. The long-term effects of short-term exposure on reproductive behavior of pairs could potentiallyhave consequences for the population dynamics of H. azteca. However, since individual-level responsescan both overestimate and underestimate effects at the population level, appropriate population modelsare needed to reduce the uncertainty in extrapolating between these levels of biological organization.
. Introduction
Non-target aquatic organisms are likely to be exposed to pulsesf pesticides following spray drift, accidental spills, drain flow andurface run-off after rainfall. These pulses may extend anywhererom a few minutes to several hours (Kreutzweiser and Wood,991; Liess et al., 1999; Rawn et al., 1982; Reinert et al., 2002).tandard acute ecotoxicity tests to assess the risks of pesticideso non-target organisms typically use juvenile organisms. In addi-ion, there is limited chronic toxicity testing looking at sublethalffects such as reproduction. Both standard acute and chronic testethods use constant exposure concentrations and such tests are
ot designed to assess the effects of intermittent exposure or the
otential for delayed effects. Here we explore the extent to whichbrief pulse of pesticide exposure has long-term effects on pairingehavior and reproductive output.
∗ Corresponding author at: School of Biological Sciences, University of Nebraska-incoln, 348 Manter Hall, Lincoln, NE 68588-0118, USA. Tel.: +1 4024726676.
E-mail address: [email protected] (V. Forbes).
166-445X/$ – see front matter © 2013 Published by Elsevier B.V.ttp://dx.doi.org/10.1016/j.aquatox.2013.09.027
© 2013 Published by Elsevier B.V.
We have chosen permethrin as a representative pyrethroidinsecticide since it is widely used and is found to be highly toxic toaquatic organisms (US EPA, 2000). Permethrin has been detectedin the aquatic environment (CCME, 2006; Starner et al., 2008), andapproximately 0.5–1.5 h after aerial applications surface water con-centrations ranged from 0.070 to 7.2 �g permethrin/L (Helson et al.,1993; Kreutzweiser and Wood, 1991; Lizotte et al., 2012a, b). USEPA estimated a worst case environmental concentration rangingfrom 0.2 to 5.32 �g permethrin/L depending on the crop (US EPA,2009).
We have chosen the freshwater amphipod, Hyalella azteca, as atest species since it is widely distributed and provides an importantpart of the food source for many fish, birds and large invertebratesthroughout North America (De March, 1981). Also, H. azteca hasbeen extensively used in aquatic toxicity tests, and its life cycle iswell described.
H. azteca is iteroparous and reproduces sexually. Reproduction
starts with amplexus in which the larger adult male grasps thefemale from the dorsal side with its gnathopods. The precopula-tory pair is able to swim and feed together. After 1 to 7 days inamplexus the pair separates briefly while the female sheds her old
6 xicolog
epesuS
aHpc(ot1Pvoi
ettsp
2
2
iawsifiawflsa7www
2
temr
eagt
ct2T
0 S. Pedersen et al. / Aquatic To
xoskeleton. Thereafter the two organisms reunite for a shorteriod, and copulation occurs. After copulation the female releasesggs into the marsupium where they are fertilized and the off-pring develop and hatch. The offspring are kept in the marsupiumntil the next molt (Cooper, 1965; Environment Canada, 1997;trong, 1973; US EPA, 2000).
Swimming in amplexus plays a key role in the reproductive cyclend is therefore essential for successful reproduction in H. azteca.owever, very few studies have investigated effects of pesticides onairing behavior of H. azteca. Previous studies have found that pre-opulatory pairing in H. azteca was impaired by pesticide exposureBlockwell et al., 1998; Pandey et al., 2011), and disruption of pairsf other amphipod species due to environmental changes includinghe presence of toxicants has been described in the literature (Davis,978; Linden, 1976; Malbouisson et al., 1994; Pascoe et al., 1994).airs of Gammarus pulex exposed for 1 h to the pyrethroid esfen-alerate separated during exposure, and subsequent reproductiveutput was significantly reduced during a 14 day monitoring periodn clean water (Cold and Forbes, 2004).
The specific aims of the present study were to examine sublethalffects of a 1 h pulse exposure of different concentrations of perme-hrin on precopulatory pairs of H. azteca and to determine whetherhere were any acute or delayed effects on pairing behavior or sub-equent reproductive output over a 56 day period following theulse exposure.
. Materials and methods
.1. Culturing conditions
H. azteca were obtained from Ghent University and maintainedn our laboratory at Roskilde University since February 2009. H.zteca were cultured in a temperature-controlled room at 23 ◦Cith a photoperiod of 16 h of light and 8 h of dark and a light inten-
ity of 500 ± 50 lux. Cultures were maintained in 63 L glass aquarian which water quality was controlled by means of a biologicallter containing mussel shells, stones and sand. The water in thequaria was a mixture of tap water from Roskilde University dilutedith deionized water (2:1) which was further mixed with artificial
reshwater (Borgmann, 1996) in the ratio of 2:1 (Pedersen, unpub-ished data). A screening study of nine different mixtures of waterhowed that this combination was the most favorable for survivalnd reproduction of H. azteca (Pedersen, unpublished data). pH was.8 and hardness was 221 measured as mg/L CaCO3. Three times aeek approximately 1.4 g rabbit chow (Chrisco, Koege, Denmark)as added to each aquarium, and every second week cotton gauzeas added as a substrate.
.2. Experimental procedures
The experiment was conducted under the same light andemperature conditions as the cultures (described above). Thexperiment lasted for 56 days, and measured endpoints wereobility, survival, pair separation, pair re-formation, time to first
eproduction and reproductive output.Each treatment consisted of 6 replicates (1 pair of H. azteca in
ach) per concentration. Each of the replicate pairs was exposed in 400 mL glass beaker containing 150 mL of test water and 5 × 5 cmauze as a substrate. The test water used in the experiments washe same as used for the cultures.
Pairs used in the experiment were randomly selected from a
ulture of 10 week old H. azteca. The pairs were pulse-exposedo different nominal concentrations of permethrin: 0, 0.3, 0.9 and.7 �g/L. The pulse lasted for 1 h to simulate a realistic run-off event.here were two control groups. Control group 1 (C1) was pulsey 144– 145 (2013) 59– 65
exposed in control water and treated identically to exposed orga-nisms to ensure that any differences among treatments were notdue to differences in handling and that any possible separation wasnot caused by transferring the pairs. Control group 2 (C2) was pulseexposed in control water, and after the pulse the pair was gentlyseparated with a pipette. This control group was included to enablecomparison of the time to pair re-formation with pairs exposed topermethrin.
After exposure, the pairs were transferred to clean water to berinsed of residual pesticide and subsequently transferred to anotherbeaker with clean water and cotton gauze as a substrate. Threetimes a week, 50 mL of the water was removed and gently replacedwith fresh aerated water, after which each pair was fed with 3 mgof homogenized rabbit chow diluted in water.
Each beaker was monitored for separation of pairs, pair re-formation and mobility with an interval of 1 min for the first 10 minand thereafter every 15 min until the end of the pulse exposure,once immediately after transfer to clean water and then at 2, 3, 6,20, and 24 h from the start of the experiment. From day 1 to 4 thebeakers were monitored twice a day and then daily until day 9, afterwhich they were monitored approximately three times a week. Thebeakers were also monitored for survival and reproduction. Everysecond week the juveniles produced by a pair were counted andremoved from the beakers. For each monitoring day, only thosereplicates with a surviving pair were included in the calculation ofreproductive output.
H. azteca were defined as immobile when lying at the bottomof the beakers, with no observed swimming behavior after gentledisturbance. Mortality was recorded when animals were partly orcompletely decomposed.
2.3. Preparation of test solutions
Permethrin was provided by Syngenta Ltd., as a mixture of cisand trans isomers (chemical purity of 98%) (Sigma–Aldrich, Fluka:45614). A stock solution of permethrin was made in acetone toprepare the test solutions. The same amount of acetone, 0.01%of the total volume, was added to each treatment including thetwo controls. A screening experiment was made with two controlgroups, one with and one without acetone. We found no signif-icant difference in behavior or survival between the two controlgroups and therefore used only acetone control groups in the exper-iment.
2.4. Water quality
Temperature, pH, dissolved oxygen level, hardness, alkalinityand ammonium were measured weekly. The pH was measuredwith a pH electrode (HANNA Instruments), dissolved oxygen levelwith a handheld dissolved oxygen meter (YSI 550 DO), water hard-ness with a Total Hardness Test (Aquamerck, No. 1.08020.0001),and ammonium with a JBL test kit. Water quality met the US EPAacceptability criteria (US EPA, 2000) such that ammonium concen-trations never exceeded 0.05 mg/L, and the oxygen concentrationswere above 6.18 mg/L in all treatments.
2.5. Permethrin extraction and analysis
Methods were based on Hladik et al. (2009) with modification.Strata-X 33� Solid Phase Extraction (SPE) cartridges (30 �g/3 mL)(Phenomenex) were conditioned with 2 × 1 mL of dichloromethaneand 2 × 1 mL of deionized water. Phenanthrene D10 and chrysene
D12 were used as internal and recovery standards, respectively.The 200 mL water samples were passed through a SPE cartridgewith a maximum flow rate of 10 mL min−1. The cartridges wereeluted with 2 × 1.5 mL of dichloromethane, collected in a 12 mL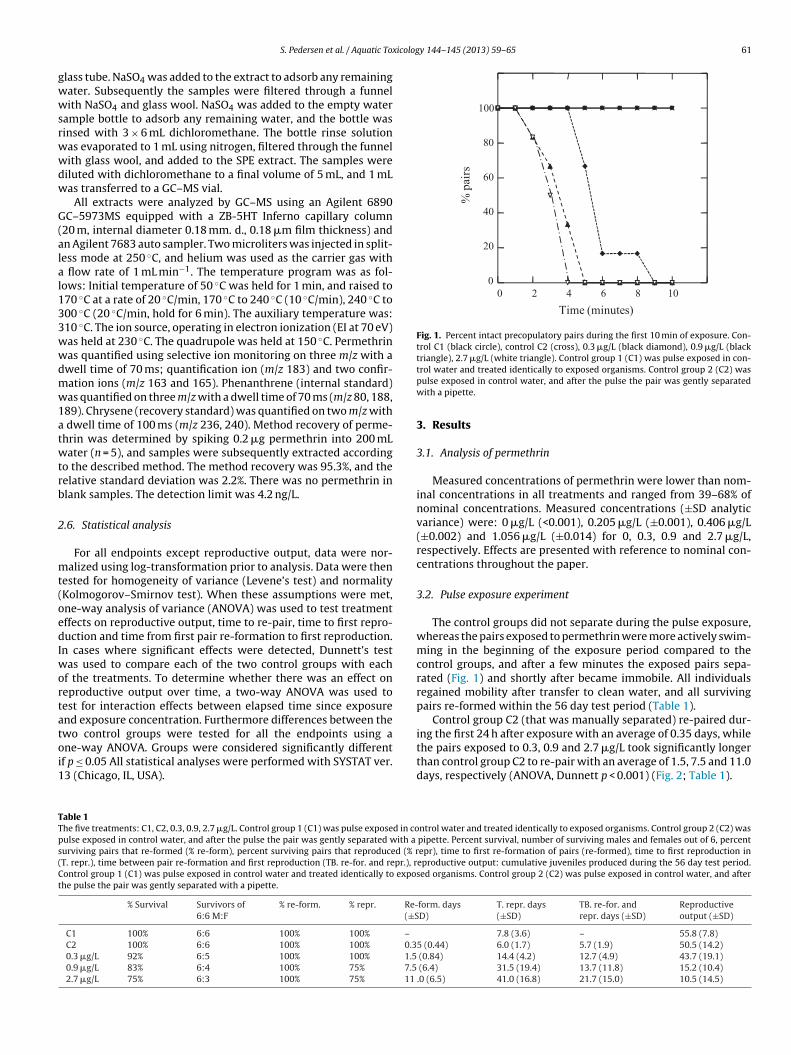
xicology 144– 145 (2013) 59– 65 61
gwwsrwwdw
G(alal133wwdmw1atwtrb
2
mt(oedIwortatoi1
0 2 4 6 8 10Time (minut es)
0
20
40
60
80
100
% p
airs
Fig. 1. Percent intact precopulatory pairs during the first 10 min of exposure. Con-trol C1 (black circle), control C2 (cross), 0.3 �g/L (black diamond), 0.9 �g/L (blacktriangle), 2.7 �g/L (white triangle). Control group 1 (C1) was pulse exposed in con-
TTps(Ct
S. Pedersen et al. / Aquatic To
lass tube. NaSO4 was added to the extract to adsorb any remainingater. Subsequently the samples were filtered through a funnelith NaSO4 and glass wool. NaSO4 was added to the empty water
ample bottle to adsorb any remaining water, and the bottle wasinsed with 3 × 6 mL dichloromethane. The bottle rinse solutionas evaporated to 1 mL using nitrogen, filtered through the funnelith glass wool, and added to the SPE extract. The samples wereiluted with dichloromethane to a final volume of 5 mL, and 1 mLas transferred to a GC–MS vial.
All extracts were analyzed by GC–MS using an Agilent 6890C–5973MS equipped with a ZB-5HT Inferno capillary column
20 m, internal diameter 0.18 mm. d., 0.18 �m film thickness) andn Agilent 7683 auto sampler. Two microliters was injected in split-ess mode at 250 ◦C, and helium was used as the carrier gas with
flow rate of 1 mL min−1. The temperature program was as fol-ows: Initial temperature of 50 ◦C was held for 1 min, and raised to70 ◦C at a rate of 20 ◦C/min, 170 ◦C to 240 ◦C (10 ◦C/min), 240 ◦C to00 ◦C (20 ◦C/min, hold for 6 min). The auxiliary temperature was:10 ◦C. The ion source, operating in electron ionization (EI at 70 eV)as held at 230 ◦C. The quadrupole was held at 150 ◦C. Permethrinas quantified using selective ion monitoring on three m/z with awell time of 70 ms; quantification ion (m/z 183) and two confir-ation ions (m/z 163 and 165). Phenanthrene (internal standard)as quantified on three m/z with a dwell time of 70 ms (m/z 80, 188,
89). Chrysene (recovery standard) was quantified on two m/z with dwell time of 100 ms (m/z 236, 240). Method recovery of perme-hrin was determined by spiking 0.2 �g permethrin into 200 mLater (n = 5), and samples were subsequently extracted according
o the described method. The method recovery was 95.3%, and theelative standard deviation was 2.2%. There was no permethrin inlank samples. The detection limit was 4.2 ng/L.
.6. Statistical analysis
For all endpoints except reproductive output, data were nor-alized using log-transformation prior to analysis. Data were then
ested for homogeneity of variance (Levene’s test) and normalityKolmogorov–Smirnov test). When these assumptions were met,ne-way analysis of variance (ANOVA) was used to test treatmentffects on reproductive output, time to re-pair, time to first repro-uction and time from first pair re-formation to first reproduction.
n cases where significant effects were detected, Dunnett’s testas used to compare each of the two control groups with each
f the treatments. To determine whether there was an effect oneproductive output over time, a two-way ANOVA was used toest for interaction effects between elapsed time since exposurend exposure concentration. Furthermore differences between the
wo control groups were tested for all the endpoints using ane-way ANOVA. Groups were considered significantly differentf p ≤ 0.05 All statistical analyses were performed with SYSTAT ver.3 (Chicago, IL, USA).able 1he five treatments: C1, C2, 0.3, 0.9, 2.7 �g/L. Control group 1 (C1) was pulse exposed in coulse exposed in control water, and after the pulse the pair was gently separated with a
urviving pairs that re-formed (% re-form), percent surviving pairs that reproduced (% rT. repr.), time between pair re-formation and first reproduction (TB. re-for. and repr.), rontrol group 1 (C1) was pulse exposed in control water and treated identically to expohe pulse the pair was gently separated with a pipette.
% Survival Survivors of6:6 M:F
% re-form. % repr. Re-(±S
C1 100% 6:6 100% 100% –
C2 100% 6:6 100% 100% 0.30.3 �g/L 92% 6:5 100% 100% 1.50.9 �g/L 83% 6:4 100% 75% 7.52.7 �g/L 75% 6:3 100% 75% 11
trol water and treated identically to exposed organisms. Control group 2 (C2) waspulse exposed in control water, and after the pulse the pair was gently separatedwith a pipette.
3. Results
3.1. Analysis of permethrin
Measured concentrations of permethrin were lower than nom-inal concentrations in all treatments and ranged from 39–68% ofnominal concentrations. Measured concentrations (±SD analyticvariance) were: 0 �g/L (<0.001), 0.205 �g/L (±0.001), 0.406 �g/L(±0.002) and 1.056 �g/L (±0.014) for 0, 0.3, 0.9 and 2.7 �g/L,respectively. Effects are presented with reference to nominal con-centrations throughout the paper.
3.2. Pulse exposure experiment
The control groups did not separate during the pulse exposure,whereas the pairs exposed to permethrin were more actively swim-ming in the beginning of the exposure period compared to thecontrol groups, and after a few minutes the exposed pairs sepa-rated (Fig. 1) and shortly after became immobile. All individualsregained mobility after transfer to clean water, and all survivingpairs re-formed within the 56 day test period (Table 1).
Control group C2 (that was manually separated) re-paired dur-
ing the first 24 h after exposure with an average of 0.35 days, whilethe pairs exposed to 0.3, 0.9 and 2.7 �g/L took significantly longerthan control group C2 to re-pair with an average of 1.5, 7.5 and 11.0days, respectively (ANOVA, Dunnett p < 0.001) (Fig. 2; Table 1).ntrol water and treated identically to exposed organisms. Control group 2 (C2) waspipette. Percent survival, number of surviving males and females out of 6, percentepr), time to first re-formation of pairs (re-formed), time to first reproduction ineproductive output: cumulative juveniles produced during the 56 day test period.sed organisms. Control group 2 (C2) was pulse exposed in control water, and after
form. daysD)
T. repr. days(±SD)
TB. re-for. andrepr. days (±SD)
Reproductiveoutput (±SD)
7.8 (3.6) – 55.8 (7.8)5 (0.44) 6.0 (1.7) 5.7 (1.9) 50.5 (14.2)
(0.84) 14.4 (4.2) 12.7 (4.9) 43.7 (19.1) (6.4) 31.5 (19.4) 13.7 (11.8) 15.2 (10.4).0 (6.5) 41.0 (16.8) 21.7 (15.0) 10.5 (14.5)
62 S. Pedersen et al. / Aquatic Toxicology 144– 145 (2013) 59– 65
0 2 4 6 141210 8Time (days)
0
20
40
60
80
100C
umul
ativ
e % o
f pai
rs th
at re
-form
ed
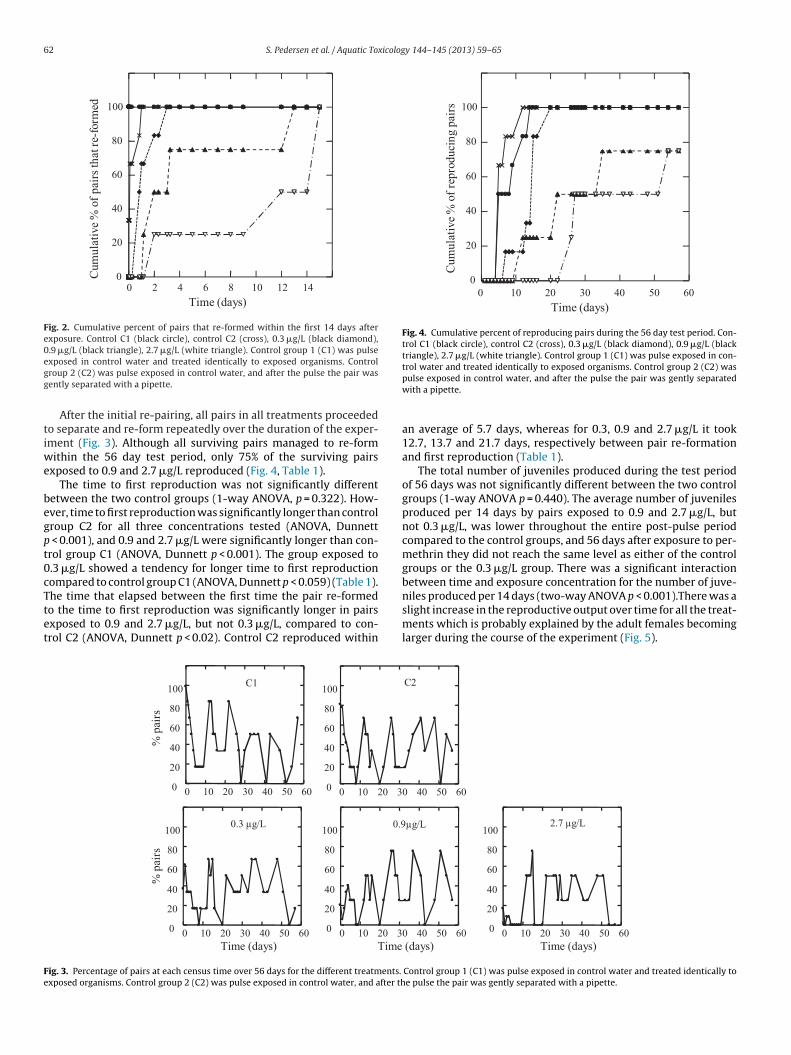
Fig. 2. Cumulative percent of pairs that re-formed within the first 14 days afterexposure. Control C1 (black circle), control C2 (cross), 0.3 �g/L (black diamond),0.9 �g/L (black triangle), 2.7 �g/L (white triangle). Control group 1 (C1) was pulseegg
tiwe
begpt0cTtet
0 10 20 30 40 50 60Time (days)
0
20
40
60
80
100
Cum
ulat
ive %
of r
epro
duci
ng p
airs
Fig. 4. Cumulative percent of reproducing pairs during the 56 day test period. Con-trol C1 (black circle), control C2 (cross), 0.3 �g/L (black diamond), 0.9 �g/L (blacktriangle), 2.7 �g/L (white triangle). Control group 1 (C1) was pulse exposed in con-trol water and treated identically to exposed organisms. Control group 2 (C2) was
Fe
xposed in control water and treated identically to exposed organisms. Controlroup 2 (C2) was pulse exposed in control water, and after the pulse the pair wasently separated with a pipette.
After the initial re-pairing, all pairs in all treatments proceededo separate and re-form repeatedly over the duration of the exper-ment (Fig. 3). Although all surviving pairs managed to re-form
ithin the 56 day test period, only 75% of the surviving pairsxposed to 0.9 and 2.7 �g/L reproduced (Fig. 4, Table 1).
The time to first reproduction was not significantly differentetween the two control groups (1-way ANOVA, p = 0.322). How-ver, time to first reproduction was significantly longer than controlroup C2 for all three concentrations tested (ANOVA, Dunnett
< 0.001), and 0.9 and 2.7 �g/L were significantly longer than con-rol group C1 (ANOVA, Dunnett p < 0.001). The group exposed to.3 �g/L showed a tendency for longer time to first reproductionompared to control group C1 (ANOVA, Dunnett p < 0.059) (Table 1).
he time that elapsed between the first time the pair re-formedo the time to first reproduction was significantly longer in pairsxposed to 0.9 and 2.7 �g/L, but not 0.3 �g/L, compared to con-rol C2 (ANOVA, Dunnett p < 0.02). Control C2 reproduced withinC1
60504030201000
20
40
60
80
100
% p
airs
0.9
320100Time
0
20
40
60
80
100
3201000
20
40
60
80
100
0.3 µg/L
6050403020100Time (da ys)
0
20
40
60
80
100
% p
airs
ig. 3. Percentage of pairs at each census time over 56 days for the different treatments.
xposed organisms. Control group 2 (C2) was pulse exposed in control water, and after th
pulse exposed in control water, and after the pulse the pair was gently separatedwith a pipette.
an average of 5.7 days, whereas for 0.3, 0.9 and 2.7 �g/L it took12.7, 13.7 and 21.7 days, respectively between pair re-formationand first reproduction (Table 1).
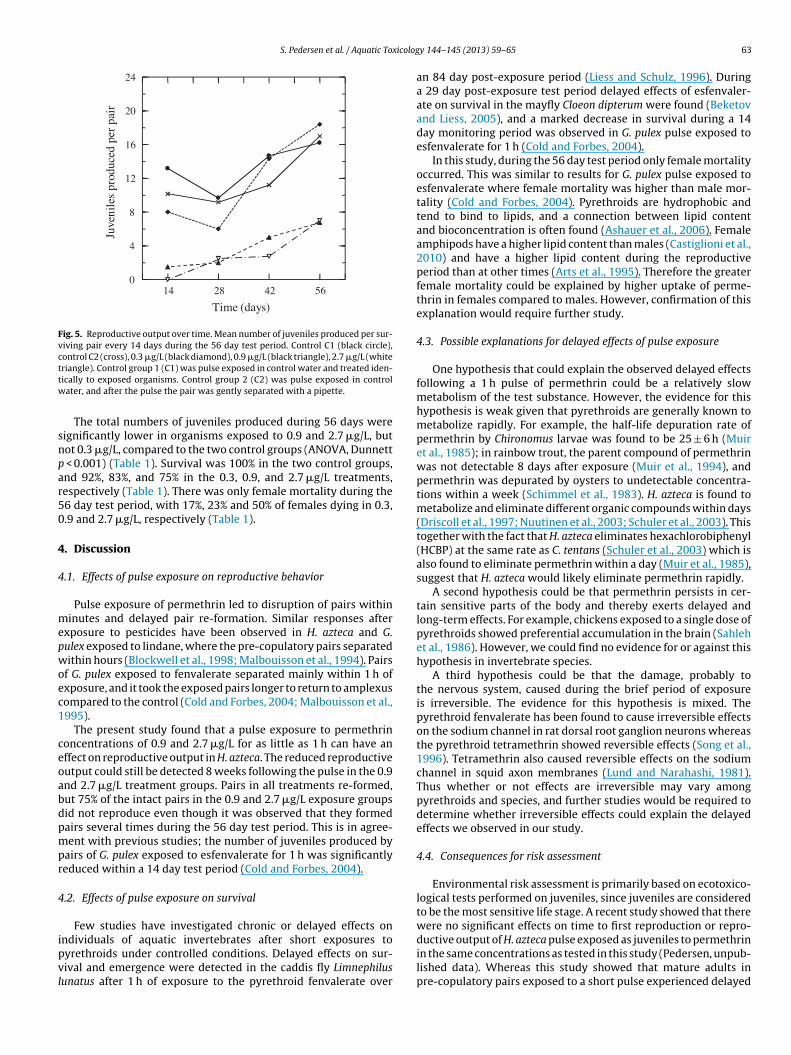
The total number of juveniles produced during the test periodof 56 days was not significantly different between the two controlgroups (1-way ANOVA p = 0.440). The average number of juvenilesproduced per 14 days by pairs exposed to 0.9 and 2.7 �g/L, butnot 0.3 �g/L, was lower throughout the entire post-pulse periodcompared to the control groups, and 56 days after exposure to per-methrin they did not reach the same level as either of the controlgroups or the 0.3 �g/L group. There was a significant interactionbetween time and exposure concentration for the number of juve-niles produced per 14 days (two-way ANOVA p < 0.001).There was a
slight increase in the reproductive output over time for all the treat-ments which is probably explained by the adult females becominglarger during the course of the experiment (Fig. 5).2.7 µg/L
6050403020100Time (da ys)
0
20
40
60
80
100µg/L
6050400 (da ys)
C2
6050400
Control group 1 (C1) was pulse exposed in control water and treated identically toe pulse the pair was gently separated with a pipette.
S. Pedersen et al. / Aquatic Toxicolog
Time (days)
14 28 42 560
4
8
12
16
20
24
Juve
nile
s pr
oduc
ed p
er p
air
Fig. 5. Reproductive output over time. Mean number of juveniles produced per sur-viving pair every 14 days during the 56 day test period. Control C1 (black circle),control C2 (cross), 0.3 �g/L (black diamond), 0.9 �g/L (black triangle), 2.7 �g/L (whitettw
snpar50
4
4
mepwoec1
ceoabdpmpr
4
ipvl
riangle). Control group 1 (C1) was pulse exposed in control water and treated iden-ically to exposed organisms. Control group 2 (C2) was pulse exposed in controlater, and after the pulse the pair was gently separated with a pipette.
The total numbers of juveniles produced during 56 days wereignificantly lower in organisms exposed to 0.9 and 2.7 �g/L, butot 0.3 �g/L, compared to the two control groups (ANOVA, Dunnett
< 0.001) (Table 1). Survival was 100% in the two control groups,nd 92%, 83%, and 75% in the 0.3, 0.9, and 2.7 �g/L treatments,espectively (Table 1). There was only female mortality during the6 day test period, with 17%, 23% and 50% of females dying in 0.3,.9 and 2.7 �g/L, respectively (Table 1).
. Discussion
.1. Effects of pulse exposure on reproductive behavior
Pulse exposure of permethrin led to disruption of pairs withininutes and delayed pair re-formation. Similar responses after
xposure to pesticides have been observed in H. azteca and G.ulex exposed to lindane, where the pre-copulatory pairs separatedithin hours (Blockwell et al., 1998; Malbouisson et al., 1994). Pairs
f G. pulex exposed to fenvalerate separated mainly within 1 h ofxposure, and it took the exposed pairs longer to return to amplexusompared to the control (Cold and Forbes, 2004; Malbouisson et al.,995).
The present study found that a pulse exposure to permethrinoncentrations of 0.9 and 2.7 �g/L for as little as 1 h can have anffect on reproductive output in H. azteca. The reduced reproductiveutput could still be detected 8 weeks following the pulse in the 0.9nd 2.7 �g/L treatment groups. Pairs in all treatments re-formed,ut 75% of the intact pairs in the 0.9 and 2.7 �g/L exposure groupsid not reproduce even though it was observed that they formedairs several times during the 56 day test period. This is in agree-ent with previous studies; the number of juveniles produced by
airs of G. pulex exposed to esfenvalerate for 1 h was significantlyeduced within a 14 day test period (Cold and Forbes, 2004).
.2. Effects of pulse exposure on survival
Few studies have investigated chronic or delayed effects on
ndividuals of aquatic invertebrates after short exposures toyrethroids under controlled conditions. Delayed effects on sur-ival and emergence were detected in the caddis fly Limnephilusunatus after 1 h of exposure to the pyrethroid fenvalerate overy 144– 145 (2013) 59– 65 63
an 84 day post-exposure period (Liess and Schulz, 1996). Duringa 29 day post-exposure test period delayed effects of esfenvaler-ate on survival in the mayfly Cloeon dipterum were found (Beketovand Liess, 2005), and a marked decrease in survival during a 14day monitoring period was observed in G. pulex pulse exposed toesfenvalerate for 1 h (Cold and Forbes, 2004).
In this study, during the 56 day test period only female mortalityoccurred. This was similar to results for G. pulex pulse exposed toesfenvalerate where female mortality was higher than male mor-tality (Cold and Forbes, 2004). Pyrethroids are hydrophobic andtend to bind to lipids, and a connection between lipid contentand bioconcentration is often found (Ashauer et al., 2006). Femaleamphipods have a higher lipid content than males (Castiglioni et al.,2010) and have a higher lipid content during the reproductiveperiod than at other times (Arts et al., 1995). Therefore the greaterfemale mortality could be explained by higher uptake of perme-thrin in females compared to males. However, confirmation of thisexplanation would require further study.
4.3. Possible explanations for delayed effects of pulse exposure
One hypothesis that could explain the observed delayed effectsfollowing a 1 h pulse of permethrin could be a relatively slowmetabolism of the test substance. However, the evidence for thishypothesis is weak given that pyrethroids are generally known tometabolize rapidly. For example, the half-life depuration rate ofpermethrin by Chironomus larvae was found to be 25 ± 6 h (Muiret al., 1985); in rainbow trout, the parent compound of permethrinwas not detectable 8 days after exposure (Muir et al., 1994), andpermethrin was depurated by oysters to undetectable concentra-tions within a week (Schimmel et al., 1983). H. azteca is found tometabolize and eliminate different organic compounds within days(Driscoll et al., 1997; Nuutinen et al., 2003; Schuler et al., 2003). Thistogether with the fact that H. azteca eliminates hexachlorobiphenyl(HCBP) at the same rate as C. tentans (Schuler et al., 2003) which isalso found to eliminate permethrin within a day (Muir et al., 1985),suggest that H. azteca would likely eliminate permethrin rapidly.
A second hypothesis could be that permethrin persists in cer-tain sensitive parts of the body and thereby exerts delayed andlong-term effects. For example, chickens exposed to a single dose ofpyrethroids showed preferential accumulation in the brain (Sahlehet al., 1986). However, we could find no evidence for or against thishypothesis in invertebrate species.
A third hypothesis could be that the damage, probably tothe nervous system, caused during the brief period of exposureis irreversible. The evidence for this hypothesis is mixed. Thepyrethroid fenvalerate has been found to cause irreversible effectson the sodium channel in rat dorsal root ganglion neurons whereasthe pyrethroid tetramethrin showed reversible effects (Song et al.,1996). Tetramethrin also caused reversible effects on the sodiumchannel in squid axon membranes (Lund and Narahashi, 1981).Thus whether or not effects are irreversible may vary amongpyrethroids and species, and further studies would be required todetermine whether irreversible effects could explain the delayedeffects we observed in our study.
4.4. Consequences for risk assessment
Environmental risk assessment is primarily based on ecotoxico-logical tests performed on juveniles, since juveniles are consideredto be the most sensitive life stage. A recent study showed that therewere no significant effects on time to first reproduction or repro-
ductive output of H. azteca pulse exposed as juveniles to permethrinin the same concentrations as tested in this study (Pedersen, unpub-lished data). Whereas this study showed that mature adults inpre-copulatory pairs exposed to a short pulse experienced delayed
6 xicolog
eeTtsbcsmslts
budrouoaafba2ftcdsd
A
Atal
R
A
A
B
B
B
C
C
C
C
D
4 S. Pedersen et al. / Aquatic To
ffects in time to first reproduction and reduced reproduction sev-ral weeks after the exposure even though they seemed to recover.his suggests that adults in pre-copulatory pairs are more sensi-ive toward pesticide exposure than juveniles. This difference inensitivity of juveniles and adult pre-copulatory pairs would note captured in current risk assessment lower tier screening pro-edures where juveniles are often used. Reproductive endpoints,uch as pre-copulatory pairing, could be included in the risk assess-ent process with chronic and higher-tier studies. However, this
tudy highlights that it is important to consider all aspects of theife cycle, and that some behavioral aspects, which could be impor-ant, may not be covered by extrapolation from other life stages orpecies.
The long-term effects of short-term exposure on reproductiveehavior of pairs could potentially have consequences for the pop-lation dynamics of H. azteca. Reduced reproduction for at least 56ays can especially affect temperate species for which the maineproductive period is seasonally dependent, and the numbers ofverwintering individuals are important for the next season’s pop-lation. Relatively little is known about the impact on populationsf non-target aquatic amphipods when different life-history traitsre affected by pesticides. Such information is essential as longs protection goals are based at the population level and testsor risk assessment measure effects at the individual level. It haseen shown that individual-level responses can both overestimatend underestimate effects at the population level (Forbes et al.,001; Hayashi et al., 2008; Hanson and Stark, 2012) and there-ore that appropriate population models are needed to reducehe uncertainty in extrapolating between these levels of biologi-al organization. The present results will be used in future work toevelop an individual-based model of H. azteca to explore the con-equences of impairments in reproductive behavior for populationynamics under different exposure and effect scenarios.
cknowledgements
This study was financed by Roskilde University, the Danishgency for Science, Technology & Innovation and Syngenta Ltd. We
hank Anne-Grete Winding, Klara Jensen, Anja Holden Damsholtnd Linus Malmquist for technical assistance and guidance duringaboratory work.
eferences
rts, M., Ferguson, M., Glozier, N., Robarts, R., Donald, D., 1995. Spatial and temporalvariability in lipid dynamics of common amphipods – assessing the potentialfor uptake of lipophilic contaminants. Ecotoxicology 4, 91–113.
shauer, R., Boxall, A., Brown, C., 2006. Uptake and elimination of chlorpyrifos andpentachlorophenol into the freshwater amphipod Gammarus pulex. Arch. Envi-ron. Contam. Toxicol. 51, 542–548.
eketov, M.A., Liess, M., 2005. Acute contamination with esfenvalerate and food lim-itation: chronic effects on the mayfly, Cloeon dipterum. Environ. Toxicol. Chem.24, 1281–1286.
lockwell, S.J., Maund, S.J., Pascoe, D., 1998. The acute toxicity of lindane to Hyalellaazteca and the development of a sublethal bioassay based on precopulatoryguarding behavior. Arch. Environ. Contam. Toxicol. 35, 432–440.
orgmann, U., 1996. Systematic analysis of aqueous ion requirements of Hyalellaazteca: a standard artificial medium including the essential bromide ion. Arch.Environ. Contam. Toxicol. 30, 356–363.
astiglioni, D.d.S., Dutra, B.K., Cahansky, A., Rodriguez, E., Oliveira, G.T., Bond-Buckup, G., 2010. Variations in biochemical composition and lipoperoxidationlevels of Hyalella bonariensis maintained in laboratory with different diets. Anim.Biol. 60, 349–360.
CME, 2006. Canadian Water Quality Guidelines: Permethrin. Scientific SupportingDocument. Canadian Council of Ministers of the Environment, Winnipeg.
old, A., Forbes, V.E., 2004. Consequences of a short pulse of pesticide exposure forsurvival and reproduction of Gammarus pulex. Aquat. Toxicol. 67, 287–299.
ooper, W.E., 1965. Dynamics and production of a natural population of fresh-wateramphipod, Hyalella azteca. Ecol. Monogr. 35, 377–394.
avis, J., 1978. Disruption of precopulatory behavior in amphipod Anisogammaruspugettensis upon exposure to bleached kraft pulp mill effluent. Water Res. 12,273–275.
y 144– 145 (2013) 59– 65
De March, 1981. Hyalella azteca (Saussure). In: Lawerence, S.G. (Ed.), Manual for theCulture of Selected Freshwater Invertebrates, 54. Can Spec Publ Fish Aquat Sci,pp. 61–77.
Driscoll, S., Landrum, P., Tigue, E., 1997. Accumulation and toxicokinetics of fluoran-thene in water-only exposures with freshwater amphipods. Environ. Toxicol.Chem. 16, 754–761.
Environment Canada, 1997. Biological test method: test for survival and growthin sediment using the freshwater amphipod Hyalella azteca. December EPS1/RM/33.
Forbes, V.E., Calow, P., Sibly, R.M., 2001. Are current species extrapolation mod-els a good basis for ecological risk assessment? Environ. Toxicol. Chem. 20,442–447.
Hanson, N., Stark, J.D., 2012. Comparison of population level and individual levelendpoints to evaluate ecological risk of chemicals. Environ. Sci. Technol. 46,5590–5598.
Hayashi, T.I., Kamo, M., Tanaka, Y., 2008. Population level ecological effectassessment: estimating the effect of toxic chemicals on density-dependentpopulations. Ecol. Res. 24, 945–954.
Helson, B., Payne, N., Sundaram, K., 1993. Impact assessment of spray drift fromsilvicultural aerial applications of permethrin on aquatic invertebrates usingmosquito bioassays. Environ. Toxicol. Chem. 12, 1635–1642.
Hladik, M.L., Smalling, K.L., Kuivila, M., 2009. Methods of analysis—determination ofpyrethroid insecticides in water and sediment using gas chromatography, Tech-niques and methods 5-C2, USGS, U.S. Department of the Interior U.S. GeologicalSurvey.
Kreutzweiser, D.P., Wood, G.A., 1991. Permethrin treatments in Canadian forests.Part 3: environmental fate and distribution in streams. Pestic. Sci. 33,35–46.
Liess, M., Schulz, R., 1996. Chronic effects of short-term contamination with thepyrethroid insecticide fenvalerate on the caddisfly Limnephilus lunatus. Hydro-biologia 324, 99–106.
Liess, M., Schulz, R., Liess, M.H., Rother, B., Kreuzig, R., 1999. Determination ofinsecticide contamination in agricultural headwater streams. Water Res. 33,239–247.
Linden, O., 1976. Effects of oil on the amphipod Gammarus oceanicus. Environ. Pollut.10, 239–250.
Lizotte, R.E., Shields, F.D., Murdock, J.N., Kröger, R., Knight, S.S., 2012a. Mitigatingagrichemicals from an artificial runoff event using a managed riverine wetland.Sci. Total Environ. 427/428, 373–381.
Lizotte Jr., R.E., Shields Jr., F.D., Murdock, J.N., Knight, S.S., 2012b. Responses ofHyalella azteca and phytoplankton to a simulated agricultural runoff event ina managed backwater wetland. Chemosphere 87, 684–691.
Lund, A., Narahashi, T., 1981. Kinetics of sodium-channel modification by the insec-ticide tetramethrin in squid axon membranes. J. Pharmacol. Exp. Ther. 219,464–473.
Malbouisson, J.F.C., Young, T.W.K., Bark, A.W., 1995. Use of feeding rate andre-pairing of precopulatory Gammarus pulex to assess toxicity of gamma-hexachlorocyclohexane (lindane). Chemosphere 30, 1573–1583.
Malbouisson, J.F.C., Young, T.W.K., Bark, A.W., 1994. Disruption of precopula in Gam-marus pulex as a result of brief exposure to gamma-hexachlorocyclohexane(lindane). Chemosphere 28, 2011–2020.
Muir, D.C.G., Hobden, B.R., Servos, M.R., 1994. Bioconcentration of pyrethroid insec-ticides and DDT by rainbow-trout – uptake, depuration, and effect of dissolvedorganic-carbon. Aquat. Toxicol. 29, 223–240.
Muir, D.C.G., Rawn, G.P., Townsend, B.E., Lockhart, W.L., Greenhalgh, R., 1985. Bio-concentration of cypermethrin, deltamethrin, fenvalerate and permethrin byChironomus tentans larvae in sediment and water. Environ. Toxicol. Chem. 4,51–61.
Nuutinen, S., Landrum, P., Schuler, L., Kukkonen, J., Lydy, M., 2003. Toxicokineticsof organic contaminants in Hyalella azteca. Arch. Environ. Contam. Toxicol. 44,467–475.
Pandey, R.B., Adams, G.L., Warren, L.W., 2011. Survival and precopulatory guardingbehavior of Hyalella azteca (Amphipoda) exposed to nitrate in the presence ofatrazine. Environ. Toxicol. Chem. 30, 1170–1177.
Pascoe, D., Kedwards, T.J., Maund, S.J., Muthi, E., Taylor, E.J., 1994. Laboratory andfield-evaluation of a behavioral bioassay – the Gammarus-Pulex (L) precopulaseparation (Gapps) test. Water Res. 28, 369–372.
Rawn, G.P., Webster, G.R.B., Muir, D.C.G., 1982. Fate of permethrin in model outdoorponds. J. Environ. Sci. Health B 17, 463–486.
Reinert, K.H., Giddings, J.M., Judd, L., 2002. Effects analysis of time-varying orrepeated exposures in aquatic ecological risk assessment of agrochemicals. Envi-ron. Toxicol. Chem. 21, 1977–1992.
Sahleh, M., Ibrahim, N., Soliman, N., Elsheimy, M., 1986. Persistance and distributionof cypermethrin, deltamethrin, and fenvalerate in laying chickens. J. Agric. FoodChem. 34, 895–898.
Schimmel, S.C., Garnas, R.L., Patrick, J.M., Moore, J.C., 1983. Acute toxicity, bioconcen-tration, and persistence of ac-222,705, benthiocarb, chlorpyrifos, fenvalerate,methyl parathion, and permethrin in the estuarine environment. J. Agric. FoodChem. 31, 104–113.
Schuler, L., Wheeler, M., Bailer, A., Lydy, M., 2003. Toxicokinetics of sediment-sorbedbenzo[a]pyrene and hexachlorobiphenyl using the freshwater invertebrates
Hyalella azteca, Chironomus tentans, and Lumbriculus variegatus. Environ. Toxicol.Chem. 22, 439–449.Song, J., Nagata, K., Tatebayashi, H., Narahashi, T., 1996. Interactions of tetramethrin,fenvalerate and DDT at the sodium channel in rat dorsal root ganglion neurons.Brain Res. 708, 29–37.

xicolog
S
S
S. Pedersen et al. / Aquatic To
tarner, K., White, J., Spurlock, F., Kelley, K., 2008. Assessment of pyrethroid con-tamination of streams in high-use agricultural regions of California. In: Gan, J.,
Spurlock, F., Hendley, P., Weston, D.P. (Eds.), Synthetic Pyrethroids Occurrenceand Behavior in Aquatic Environments. American Chemical Society, pp. 72–83(Chapter 4).trong, D.R., 1973. Amphipod amplexus, the significance of ecotypic variation. Ecol-ogy 54, 1383–1388.
y 144– 145 (2013) 59– 65 65
US EPA, 2009. Reregistration Eligibility Decision (RED) for Permethrin. 738-R-09-306.
US EPA, 2000. Methods for measuring the toxicity and bioaccumulation of sediment-associated contaminants with freshwater invertebrates. U.S. EnvironmentalProtection Agency. Duluth, Minnesota 55804. Office of Science and Technology.Office of Water U.S. Environmental Protection Agency Washington, D.C, seconded.