optimization of the enzymatic hydrolysis of lupin ( lupinus ) proteins for producing ace-inhibitory...
TRANSCRIPT

Optimization of the Enzymatic Hydrolysis of Lupin (Lupinus) Proteinsfor Producing ACE-Inhibitory PeptidesGiovanna Boschin,*,† Graziana Maria Scigliuolo,‡ Donatella Resta,‡ and Anna Arnoldi†,‡
†Department of Pharmaceutical Sciences, University of Milan, via Mangiagalli 25, 20133 Milano, Italy‡HPF Nutraceutics s.r.l., via Balzaretti 9, 20133 Milano, Italy
ABSTRACT: Recently, the enzymatic hydrolysis of Lupinus albus and Lupinus angustifolius proteins with pepsin was showed toproduce peptides able to inhibit the angiotensin-converting enzyme (ACE). The objective of the present work was to testdifferent hydrolytic enzymes and to investigate three lupin species (L. albus, L. angustifolius, Lupinus luteus) with the final goal ofselecting the best enzyme/species combination for an efficient production of ACE-inhibitory peptide mixtures. Pepsin gavepeptides with the best IC50 values (mean value on three species 186 ± 10 μg/mL), followed by pepsin + trypsin (198 ± 16 μg/mL), chymotrypsin (213 ± 83 μg/mL), trypsin (405 ± 54 μg/mL), corolase PP (497 ± 32 μg/mL), umamizyme (865 ± 230μg/mL), and flavourzyme (922 ± 91 μg/mL). The three species showed similar activity scales, but after pepsin + trypsin andchymotrypsin treatments, L. luteus peptide mixtures resulted to be significantly the most active. This investigation indicates thatlupin proteins may be a valuable source of ACE-inhibitory peptides, which may explain the activity observed in experimental andclinical studies and foresee the application of lupin proteins into functional foods or dietary supplements.
KEYWORDS: enzymatic hydrolysis, functional foods, hypertension, lupin, nutraceutics
■ INTRODUCTION
Lupin seed represents a promising source of innovative foodingredients because it has a good protein content (34−43% ofdry matter, competitive with soybean), an acceptablecomposition of essential amino acids,1 and an interestingcontent of some other relevant nutrients such as tocopherolsand unsaturated fatty acids.2,3 Moreover, the former toxico-logical issue of lupin seeds, i.e., the presence of quinolizidinealkaloids (QAs), has been solved by the selection of low-alkaloid cultivars.4
Recent literature indicates that lupin consumption mayprovide some useful health benefits such as the reduction ofsystemic hypertension. The hypotensive effect of lupin proteinhas been assessed in vivo in Goto−Kakizaki rats, which develophypertension when fed with a high-salt diet.5 The rats were fedwith a 6% NaCl diet containing lupin protein isolates (20%weight/weight) for two weeks. At the end of the study, thesystolic blood pressure (SBP) was 18.6 mmHg lower than inthe control group.5 In an open study on subjects with moderatehypertension, the daily consumption of 35 g of lupin protein ina model beverage reduced the SBP by 9.5 mmHg (from 137.1to 127.6 mmHg) after 1 month and 9.0 mmHg (from 137.1 to128.1 mmHg) after 3 months, whereas it lowered the diastolicblood pressure by 3.0 mmHg (from 81.5 to 78.52 mmHg) andby 4.4 mmHg (from 81.50 to 77.14 mmHg), respectively.6
Finally, two long-term clinical studies showed that theconsumption of bread enriched with lupin flour producedvery small, but statistically significant, decreases of bloodpressure with respect to the control bread in normotensiveoverweight or obese subjects.7,8
Hypertension is usually treated with drugs, for example, withangiotensin I converting enzyme (ACE; EC 3.4.15.1)inhibitors.9 They inhibit ACE, an enzyme that plays animportant role in regulating blood pressure in the renin−
angiotensin system, because it catalyzes the conversion of thebiologically inactive angiotensin I to the potent vasoconstrictorangiotensin II and inactivates the potent vasodilator bradyki-nin.10
Nonpharmacologic measures, such as changes in lifestyle anduse of dietary supplements and functional foods, areencouraged in subjects with mild hypertension to avoid ordelay the use of synthetic drugs that may produce side effects.Specific dietary ingredients are the ACE-inhibitory peptidesproduced by hydrolyzing the proteins from different animal orplant foods, in particular, milk, soy, pea, and otherlegumes.11−15
In a previous paper, we have compared the ACE-inhibitoryactivity of samples obtained by hydrolyzing the proteins fromdifferent legume seeds with pepsin and have shown that lupin isthe most active, together with soybean.16 The generic termlupin actually indicates the main domesticated and cultivatedspecies: Lupinus albus (white lupin), Lupinus angustifolius(narrow-leaf lupin), and Lupinus luteus (yellow lupin).The objectives of the present investigation were to test
different enzymes for preparing peptide mixtures and toinvestigate three lupin species, L. albus, L. angustifolius, and L.luteus, as substrates. The final goal is selecting the best species/enzyme combinations for an efficient production of ACE-inhibitory peptide mixtures, which may be possibly used ininnovative functional foods or dietary supplements. Althoughnumerous papers have reported the ACE-inhibitory activity ofpeptides from different foods, lupin is still an unexplored
Received: September 3, 2013Revised: January 23, 2014Accepted: January 31, 2014Published: January 31, 2014
Article
pubs.acs.org/JAFC
© 2014 American Chemical Society 1846 dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−1851

substrate, also considering the clinical and experimental datareported above.
■ MATERIALS AND METHODSChemicals. The commercial sources of proteases were as follows:
pepsin (from porcine gastric mucosa; EC 3.4.23.1; commercial codeP6887), trypsin (from bovine pancreas; EC 3.4.21.4; commercial codeT1426), chymotrypsin (from bovine pancreas; EC 3.4.21.1;commercial code C4129), and flavourzyme (from Aspergillus oryzae;EC 232.752.2; commercial code P6110) from Sigma-Aldrich (St.Louis, MO, USA), corolase PP (from porcine pancreas; EC 3.4.21.4;commercial code 89082) from AB Enzymes (Darmstadt, Germany),and umamizyme (from Aspergillus oryzae; EC not specified; declaredactivity of 74.3 U/g) from Amano Enzyme Inc. (Nagoya, Japan).Amicon Ultra-0.5 filters (centrifuge tubes) were bought from Millipore(Billerica, MA, USA). HPLC-grade water was prepared with a Milli-Qpurification system (Millipore). Bio-Safe Coomassie and PrecisionPlus Protein Standards Dual Color marker for SDS-PAGE were fromBiorad (Biorad Laboratories Inc., Hercules, CA). All other chemicals(reagents and solvents) were from Sigma-Aldrich.Sampling. Lupin seeds of the species L. albus (cultivar Ares) were
provided by Terrena (Matrigne-Ferchaud, France), L. angustifolius(cultivar Boregine) by Fraunhofer institute IVV (Freising, Germany),and L. luteus (cultivar Mister) by Prof. Biagina Chiofalo (University ofMessina, Messina, Italy).TPE Preparation. The total protein extracts (TPEs) of the lupin
seeds were obtained as previously reported17 and stored at −20 °C.Briefly, proteins were extracted from defatted flour with 100 mM Tris-HCl/0.5 M NaCl buffer, pH 8.2, for 2 h at 4 °C. The solid residue waseliminated by centrifugation at 6500g for 20 min at 4 °C, and thesupernatant was dialyzed against 100 mM Tris-HCl buffer, pH 8.2, for24 h at 4 °C. The protein content was assessed according to Bradford,using bovine serum albumin (BSA) as standard.18
Enzymatic Hydrolysis. TPEs were dissolved in Tris-HCl buffer100 mM, pH 8. The pH value were adjusted to the optimal hydrolysisconditions for each enzyme with the addition of 1 M NaOH or 1 MHCl, if necessary. The enzymes were dissolved in appropriatesolutions: 30 mM NaCl for pepsin, and 1 mM HCl for trypsin,chymotrypsin, and corolase PP. Umamizyme was added as a powder tothe TPEs, whereas flavourzyme was a ready-to-use solution.Both enzyme solutions and the umamizyme powder were directly
added to the TPE solutions. Reaction mixtures were incubated at afixed temperature for different reaction times. For the two-stagedigestion with pepsin + trypsin, TPEs were initially incubated withpepsin at pH 2 for 4 h, the pH was adjusted to 8 with 1 M NaOH, andtrypsin was added to the reaction mixture.Finally, pepsin was inactivated by adjusting the pH to 7 with 1 M
NaOH, whereas all other enzymes were inactivated by heating thesolutions at 100 °C for 10 min. Then the solutions were cooled andstored at −20 °C.SDS-PAGE. Drawings were taken at various intervals and analyzed
by SDS-PAGE. They were separated on 15% SDS-polyacrylamide gelusing Mini Protean 3 Cell (Biorad Laboratories Inc., Hercules, CA).Electrophoresis was performed at a constant voltage of 80 V forstacking and 120 V for separation. Gels were stained with Bio-SafeCoomassie (Biorad), scanned with Versa Doc 3000 (Biorad), andanalyzed with Quantity One 4.6.8 Software (Biorad).Ultrafiltration. After the hydrolysis, all low-molecular-weight
peptides were separated from intact enzymes, proteins, and high-molecular-weight polypeptides by ultrafiltration through 3000 Dacutoff centrifuge filters (Amicon Ultra-0.5, Millipore, Billerica, MA,USA) at 12000g for 30 min at 4 °C. The permeates were then storedat −20 °C.Determination of Peptide Concentration. The peptide
concentration in the hydrolyzed samples was measured according toliterature methods19,20 based on chelating the peptide bonds by Cu(II)in alkaline media and monitoring the change of absorbance at 330 nm.In brief, a solution of X μL peptide mixture, (500 − X) μL of water,500 μL of 6% (w/w) NaOH in water, and 50 μL of active reagent
(containing 0.6 M sodium citrate, 0.9 M sodium carbonate, and 0.07M copper sulfate, 2.4 M NaOH, pH 10.6) was prepared. The reactionmixture was carefully mixed, incubated for 15 min at 20 °C, and thenthe absorbance was measured at 330 nm. A sterile solution of peptonefrom casein at 10 mg/mL in water was used as standard for thecalibration curve; the assay is linear in the range 100−1000 μg ofpeptides in cuvette.
ACE-Inhibitory Activity Assay. After hydrolysis and filtration, allsamples were tested for determining their ACE-inhibitory activity, asrecently reported.16 Briefly, 100 μL of 2.5 mM hippuryl-histidyl-leucine (HHL) in buffer 1 (100 mM Tris-HCOOH, 300 mM NaClpH 8.3) was mixed with 30 μL of sample in buffer 1 at differentconcentrations. Usually, six concentrations were used for each sample,and each solution was tested twice. Samples were preincubated at 37°C for 15 min, then 15 μL of ACE solution, corresponding to 3 mU ofenzyme in buffer 2 (100 mM Tris-HCOOH, 300 nM NaCl, 10 μMZnCl2, pH 8.3), were added; samples were incubated for 60 min at 37°C. The reaction was stopped with 125 μL of 0.1 M HCl. The aqueoussolution was extracted twice with 600 μL of ethyl acetate; the solventwas evaporated at 95 °C and the residue was dissolved in 500 μL ofbuffer 1 and then analyzed by HPLC.
HPLC analyses were performed with a HPLC 1200 series equippedwith an autosampler (Agilent Technologies, Santa Clara, US) with aLichrospher 100, C18 column (4.6 mm × 250 mm, 5 μm; Grace, Italy)using water and acetonitrile as solvent and following the gradient: 0min 5% acetonitrile, 10 min 60% acetonitrile, 12 min 60% acetonitrile,15 min 5% acetonitrile. Injection volume was 10 μL, wavelength 228nm, flow 0.5 mL/min. The evaluation of ACE inhibition was based onthe comparison between the concentrations of HA (hippuric acid) inthe presence or absence of an estimated inhibitor. The phenomenon ofautolysis of HHL to give HA was evaluated by a reaction blank, i.e., asample with the higher evaluated inhibitor concentration and withoutthe enzyme. The percentage of ACE inhibition was computedconsidering the area of HA peak with the following formula:
=−−
×⎡⎣⎢
⎤⎦⎥
A AA A
% ACE inhibition( )( )
100IB N
IB RB
where AIB is the area of HA in inhibitor blank (IB) sample (i.e., samplewith enzyme but without any estimated inhibitor), AN is the area ofHA in the n samples containing different amounts of the estimatedinhibitor (in our case the hydrolyzate), and ARB is the area of HA inthe reaction blank (RB) sample (i.e., sample without enzyme and withthe estimated inhibitor at the highest concentration).
The percentages of ACE inhibition were plotted vs the peptideconcentrations. When the maximum ACE-inhibition concentrationwas over 50%, the percentages of ACE-inhibition were plotted vs thelog10 peptide concentrations obtaining a sigmoid curve. This curvepermitted to calculate the inhibitory concentration 50% (IC50), i.e., theconcentration needed to observe a 50% inhibition of ACE activity. TheIC50 values were obtained testing independently each sample threetimes and are expressed as mean value ± standard deviation. The mostactive peptides have the lowest IC50 values.
Statistical Analysis. Statistical analyses were performed withStatgraphics Plus (version 2.1 for Windows). The data were evaluatedusing one-way analysis of variance followed by Fisher’s least significantdifference (LSD) procedure; values with different letters aresignificantly different for p < 0.05.
■ RESULTS AND DISCUSSION
In a preliminary part of the present investigation, we tested thetotal protein extract from the seeds of L. albus beforehydrolysis. As expected, it resulted to be inactive as ACE-inhibitor, according to literature.13,21 In fact, owing to thedimension of the active site of the enzyme, the interaction maytake place only when bioactive peptides are released from theparent protein by enzymatic or hydrolytic reactions, such asdigestion, or industrial processing, such as fermentation.
Journal of Agricultural and Food Chemistry Article
dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−18511847

Preparation of Enzymatic Hydrolyzates. The first partof the experimental work was devoted to establish the bestreaction conditions for each proteolytic enzyme. Drawings weretaken at various intervals and analyzed by SDS-PAGE in orderto evaluate the hydrolysis efficiency, changing the followingparameters: enzyme/substrate (E/S) ratio, pH, temperature,and reaction time. In the case of pepsin, the best pH was 2, thetemperature 37 °C, the E/S ratio 1/100, and 18 hours werenecessary to complete the reaction. Figure 1a shows the SDS-
PAGEs of the drawings of the hydrolysis of L. albus (A) and L.luteus (L) proteins collected immediately after the enzymeaddition (A0 and L0), after 1 h (A1h and L1h), and after 18 h(A18h and L18h).In general, the optimal conditions for using trypsin are 37 °C
and pH 7−8; in our experience, the best pH value forhydrolyzing lupin proteins is 8. With an E/S ratio of 1/100, thedigestion was complete after 18 h. The same conditions wereused also for chymotrypsin. The double digestion with thesequence pepsin + trypsin was performed treating with pepsinat pH 2 for 4 h and then, after changing the pH value to 8,adding trypsin and incubating for 24 h. Figure 2b shows theSDS-PAGE of the two-step digestion.For corolase PP digestion, better results were obtained by
changing the E/S ratio to 1/50. For flavourzyme andumamizyme, several changes were performed; the reactiontime was extended to 24 h and the E/S ratio was changed to 1/20 because both 1/50 and 1/100 ratios were ineffective. Inaddition, the incubation temperatures were increased to 45 and60 °C, respectively. Table 1 reports the final conditions used forproducing the samples used in the following experimentations.
After the digestion, the samples were ultrafiltered in order toseparate low-molecular-weight peptides from high-molecularones and intact enzymes.To select the best filtration conditions for maximizing the
ACE-inhibitory activity, the digested mixture obtained byhydrolyzing L. albus protein with pepsin was ultrafiltered eitherwith 10000 or 3000 Da cutoff membranes. Figure 2 shows twoACE inhibition curves obtained by plotting the peptideconcentration vs the percentage ACE-inhibition for the sampleswith molecular weight (MW) lower than 3000 or 10000 Da,respectively. Whereas the latter sample did not reach the 50%ACE-inhibition even at the highest tested concentration (1110μg/mL), the former, having molecular weight less than 3000Da, was much more active, showing a 87% ACE-inhibition atthe concentration of 893 μg/mL. A possible explanation is thatthe sample having MW lower than 10000 Da containsnumerous inactive peptides (with MW in the range 3000 and10000 Da), which decrease the concentration of the activepeptides. Low MW peptides, usually under 3000 Da, are knownas ACE-inhibitors in vivo because they can be absorbed at gutlevel.21 After these results, all subsequent experimentation wasperformed on hydrolyzed mixtures filtered through a 3000 Dacutoff membrane.
Evaluation of ACE-Inhibitory Activity. The ACE-inhibitory activity assays were performed after having equalizedthe peptide concentrations of all samples. The peptideconcentration was determined by using a literature colorimetricassay optimized for peptides.19,20 In our opinion, the evaluationof the peptide concentrations and their equalization beforeperforming the ACE-inhibitory test are of outmost importancein order to get a reliable comparison of their activities.The results of the ACE inhibition assays (maximum
percentage of ACE inhibition and IC50 value) of the peptidemixtures from L. albus, L. angustifolius, and L. luteus arereported in Tables 2, 3, and 4, respectively.
Figure 1. SDS-PAGE obtained after digestion of: a) Lupinus albus (A)and Lupinus luteus (L) proteins with pepsin, at 37 °C with enzyme/substrate (E/S) ratio 1/100; drawings collected immediately afterenzyme addition (A0 and L0), after 1 h (A1h and L1h), and after 18 h(A18h and L18h). b) Lupinus angustifolius proteins digested with pepsin+ trypsin drawings collected immediately after pepsin addition (P0),after 1 h (P1h), after 4 h (P4h), immediately after trypsin addition(Tro), after 1 h (Tr1h), after 24 h (overnight, Tro/n). M is the marker.
Figure 2. Total protein extract of Lupinus albus seed digested withpepsin: comparison between the ACE inhibition curves obtainedfiltering with 10000 and 3000 Da cutoff membranes.
Table 1. Optimized Parameters for the Hydrolysis of theLupin Total Protein Extract: Enzyme, pH, Temperature(°C), Enzyme/Substrate Ratio (E/S ratio), and Time (h)
enzyme pH temperature (°C) E/S ratio (w/w) time (h)
pepsin 2 37 1/100 18trypsin 8 37 1/100 18chymotrypsin 8 37 1/100 18corolase PP 8 37 1/50 18umamizyme 7 45 1/20 24flavourzyme 7 60 1/20 24
Journal of Agricultural and Food Chemistry Article
dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−18511848
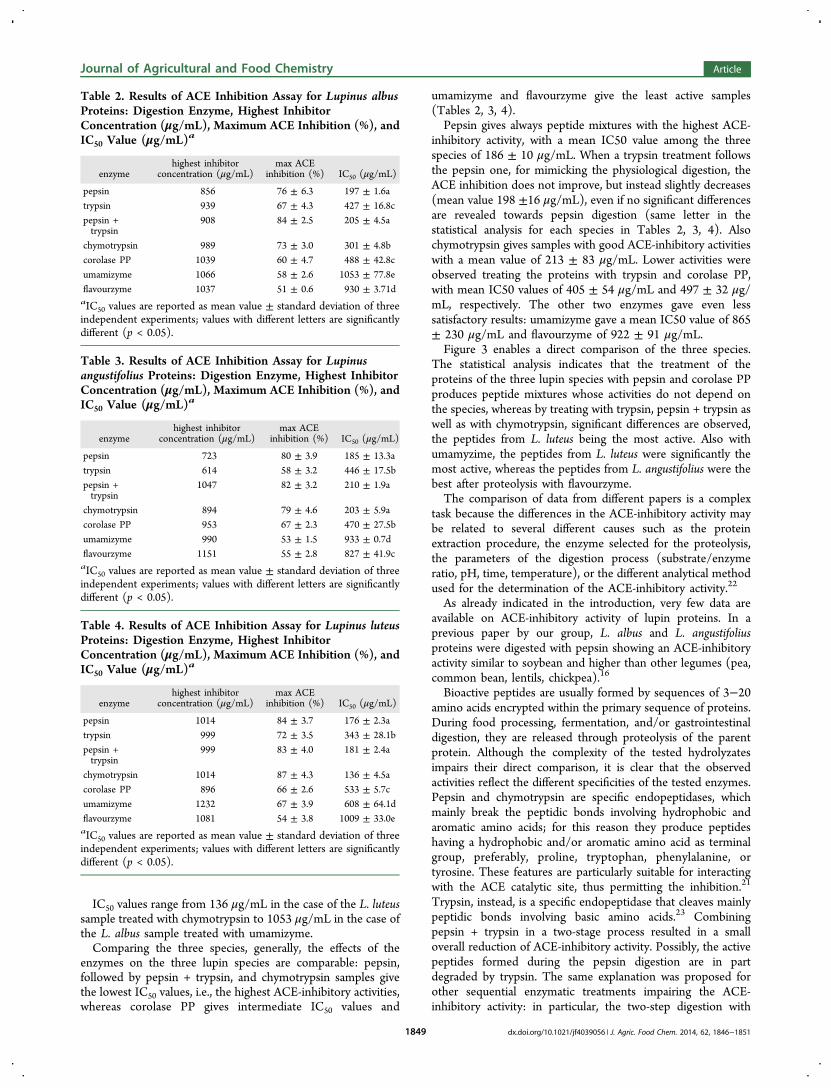
IC50 values range from 136 μg/mL in the case of the L. luteussample treated with chymotrypsin to 1053 μg/mL in the case ofthe L. albus sample treated with umamizyme.Comparing the three species, generally, the effects of the
enzymes on the three lupin species are comparable: pepsin,followed by pepsin + trypsin, and chymotrypsin samples givethe lowest IC50 values, i.e., the highest ACE-inhibitory activities,whereas corolase PP gives intermediate IC50 values and
umamizyme and flavourzyme give the least active samples(Tables 2, 3, 4).Pepsin gives always peptide mixtures with the highest ACE-
inhibitory activity, with a mean IC50 value among the threespecies of 186 ± 10 μg/mL. When a trypsin treatment followsthe pepsin one, for mimicking the physiological digestion, theACE inhibition does not improve, but instead slightly decreases(mean value 198 ±16 μg/mL), even if no significant differencesare revealed towards pepsin digestion (same letter in thestatistical analysis for each species in Tables 2, 3, 4). Alsochymotrypsin gives samples with good ACE-inhibitory activitieswith a mean value of 213 ± 83 μg/mL. Lower activities wereobserved treating the proteins with trypsin and corolase PP,with mean IC50 values of 405 ± 54 μg/mL and 497 ± 32 μg/mL, respectively. The other two enzymes gave even lesssatisfactory results: umamizyme gave a mean IC50 value of 865± 230 μg/mL and flavourzyme of 922 ± 91 μg/mL.Figure 3 enables a direct comparison of the three species.
The statistical analysis indicates that the treatment of theproteins of the three lupin species with pepsin and corolase PPproduces peptide mixtures whose activities do not depend onthe species, whereas by treating with trypsin, pepsin + trypsin aswell as with chymotrypsin, significant differences are observed,the peptides from L. luteus being the most active. Also withumamyzime, the peptides from L. luteus were significantly themost active, whereas the peptides from L. angustifolius were thebest after proteolysis with flavourzyme.The comparison of data from different papers is a complex
task because the differences in the ACE-inhibitory activity maybe related to several different causes such as the proteinextraction procedure, the enzyme selected for the proteolysis,the parameters of the digestion process (substrate/enzymeratio, pH, time, temperature), or the different analytical methodused for the determination of the ACE-inhibitory activity.22
As already indicated in the introduction, very few data areavailable on ACE-inhibitory activity of lupin proteins. In aprevious paper by our group, L. albus and L. angustifoliusproteins were digested with pepsin showing an ACE-inhibitoryactivity similar to soybean and higher than other legumes (pea,common bean, lentils, chickpea).16
Bioactive peptides are usually formed by sequences of 3−20amino acids encrypted within the primary sequence of proteins.During food processing, fermentation, and/or gastrointestinaldigestion, they are released through proteolysis of the parentprotein. Although the complexity of the tested hydrolyzatesimpairs their direct comparison, it is clear that the observedactivities reflect the different specificities of the tested enzymes.Pepsin and chymotrypsin are specific endopeptidases, whichmainly break the peptidic bonds involving hydrophobic andaromatic amino acids; for this reason they produce peptideshaving a hydrophobic and/or aromatic amino acid as terminalgroup, preferably, proline, tryptophan, phenylalanine, ortyrosine. These features are particularly suitable for interactingwith the ACE catalytic site, thus permitting the inhibition.21
Trypsin, instead, is a specific endopeptidase that cleaves mainlypeptidic bonds involving basic amino acids.23 Combiningpepsin + trypsin in a two-stage process resulted in a smalloverall reduction of ACE-inhibitory activity. Possibly, the activepeptides formed during the pepsin digestion are in partdegraded by trypsin. The same explanation was proposed forother sequential enzymatic treatments impairing the ACE-inhibitory activity: in particular, the two-step digestion with
Table 2. Results of ACE Inhibition Assay for Lupinus albusProteins: Digestion Enzyme, Highest InhibitorConcentration (μg/mL), Maximum ACE Inhibition (%), andIC50 Value (μg/mL)a
enzymehighest inhibitor
concentration (μg/mL)max ACE
inhibition (%) IC50 (μg/mL)
pepsin 856 76 ± 6.3 197 ± 1.6atrypsin 939 67 ± 4.3 427 ± 16.8cpepsin +trypsin
908 84 ± 2.5 205 ± 4.5a
chymotrypsin 989 73 ± 3.0 301 ± 4.8bcorolase PP 1039 60 ± 4.7 488 ± 42.8cumamizyme 1066 58 ± 2.6 1053 ± 77.8eflavourzyme 1037 51 ± 0.6 930 ± 3.71daIC50 values are reported as mean value ± standard deviation of threeindependent experiments; values with different letters are significantlydifferent (p < 0.05).
Table 3. Results of ACE Inhibition Assay for Lupinusangustifolius Proteins: Digestion Enzyme, Highest InhibitorConcentration (μg/mL), Maximum ACE Inhibition (%), andIC50 Value (μg/mL)a
enzymehighest inhibitor
concentration (μg/mL)max ACE
inhibition (%) IC50 (μg/mL)
pepsin 723 80 ± 3.9 185 ± 13.3atrypsin 614 58 ± 3.2 446 ± 17.5bpepsin +trypsin
1047 82 ± 3.2 210 ± 1.9a
chymotrypsin 894 79 ± 4.6 203 ± 5.9acorolase PP 953 67 ± 2.3 470 ± 27.5bumamizyme 990 53 ± 1.5 933 ± 0.7dflavourzyme 1151 55 ± 2.8 827 ± 41.9caIC50 values are reported as mean value ± standard deviation of threeindependent experiments; values with different letters are significantlydifferent (p < 0.05).
Table 4. Results of ACE Inhibition Assay for Lupinus luteusProteins: Digestion Enzyme, Highest InhibitorConcentration (μg/mL), Maximum ACE Inhibition (%), andIC50 Value (μg/mL)a
enzymehighest inhibitor
concentration (μg/mL)max ACE
inhibition (%) IC50 (μg/mL)
pepsin 1014 84 ± 3.7 176 ± 2.3atrypsin 999 72 ± 3.5 343 ± 28.1bpepsin +trypsin
999 83 ± 4.0 181 ± 2.4a
chymotrypsin 1014 87 ± 4.3 136 ± 4.5acorolase PP 896 66 ± 2.6 533 ± 5.7cumamizyme 1232 67 ± 3.9 608 ± 64.1dflavourzyme 1081 54 ± 3.8 1009 ± 33.0eaIC50 values are reported as mean value ± standard deviation of threeindependent experiments; values with different letters are significantlydifferent (p < 0.05).
Journal of Agricultural and Food Chemistry Article
dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−18511849

alcalase and flavourzyme performed on common bean proteinand chicken protein isolates.24,25
Corolase PP, a patented enzymatic complex used to simulatethe in vitro protein digestion in the intestinal tract, consists of amixture of proteolytic enzymes from pig pancreas, containingtrypsin and chymotrypsin, together with other aminopeptidasesand carboxypeptidases. Its moderate ACE-inhibitory activitymay be related to the absence of pepsin that seems to be thebest enzyme to produce lupin peptides with good ACE-inhibitory activity. Obviously, this enzyme may give goodresults with other proteins; for example, satisfactory resultswere obtained on sea cucumber proteins digested with pepsinand subsequently with corolase PP.26
Flavourzyme and umamizyme are “generally recognized assafe” (GRAS) enzymes used in the food industry. They arefood-grade fungal protease/peptidase complexes produced bysubmerged fermentation of a strain of Aspergillus oryzae,exhibiting both endoprotease and exopeptidase activities. Theywere tested with the final aim of a large-scale production ofhydrolyzates to be used as food supplements production butdid not provided useful results. In the literature, flavourzymedigestion was applied to cowpea proteins, giving peptides withIC50 values in the large range 0.04−170.6 μg/mL and torapeseed protein producing peptides with an ACE-inhibitoryactivity of about 50%.27,28 Moreover a double digestion withalcalase/flavourzyme on two different chickpea varieties gavepeptide mixtures with IC50 values of 316 and 228 μg/mL, whileon yellow pea proteins gave an IC50 value of 412 μg/mL.22
Umamizyme was used to digest canola meal proteins, givingpeptide mixtures with low ACE-inhibitory activity.29 Unfortu-nately, with lupin proteins, they give lower ACE-inhibitoryactivity than the other tested enzymes.In conclusion, this investigation confirms that lupin seed may
be a valuable source of ACE-inhibitory peptides, possiblyexplaining the activity observed in experimental and clinicalstudies.5−8 This may foresee their future applications into eitherfunctional foods or dietary supplements.
Of course, this is only a first step in the study of ACE-inhibitory activity of lupin peptides because other studies arenecessary to determine their stability and bioavailability. In fact,an open question is their capability to survive to thegastrointestinal digestion and to be absorbed in order toreach the bloodstream in their active form. Indeed, theliterature shows that peptides from other food sources (forexample milk) are systemically distributed.21,30 The problemcould be overcome also by using new emerging technologies,such as micro- or nanoencapsulation, that may offer feasiblesolutions for improving the stability of peptides in various foodproducts and during digestion.21,30 The bioavailability of ACE-inhibitory peptides may also be increased by cross-linking thetarget peptide to protein transduction domains or by means ofspecific peptide carriers able to deliver biologically activepeptides into cells. Moreover, peptide permeation may beachieved by chemical enhancers and surfactant-like agents.31
Finally, further studies are needed to determine thecomposition of active peptide mixtures, to isolate thepeptide(s) responsibles for ACE-inhibitory activity, and toconfirm their activity through in vitro and in vivo studies.
■ AUTHOR INFORMATION
Corresponding Author*Phone: +39-02-50318210. Fax: +39-02-50318202. E-mail:[email protected].
NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTS
We are grateful to Mariantonietta Loredana Mesce for herprecious help in experimental work, Biagina Chiofalo for L.luteus seeds, the Fraunhofer Institute IVV for L. angustifoliusseeds, Terrena for L. albus seeds, AB enzymes for the gift ofcorolase PP, Giovanna Speranza for the gift of umamizyme and
Figure 3. Comparison of mean values of ACE-inhibition IC50 values (expressed in μg/mL) of peptides mixtures obtained from Lupinus albus,Lupinus angustifolius, and Lupinus luteus proteins digested with pepsin, trypsin, pepsin + trypsin, chymotrypsin, corolase PP, umamizyme, andflavourzyme. Values with different letters are significantly different (p < 0.05).
Journal of Agricultural and Food Chemistry Article
dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−18511850

flavourzyme, and Andrea Meregaglia for enzymatic digestionsadvices.
■ ABBREVIATIONS USED
ACE, angiotensin-converting enzyme; BSA, bovine serumalbumin; E/S, enzyme/substrate ratio; HA, hippuric Acid;HHL, hippuryl-histidyl-leucine; HL, histidyl-leucine; IC50,inhibitory concentration 50%; MW, molecular weight; PP,from porcine pancreas; SDS-PAGE, sodium dodecyl sulfate−polyacrylamide gel electrophoresis; TPE, total protein extract
■ REFERENCES(1) Sujak, A.; Kotlarz, A.; Strobel, W. Compositional and nutritionalevaluation of several lupin seeds. Food Chem. 2006, 98, 711−719.(2) Boschin, G.; Arnoldi, A. Legumes are valuable sources oftocopherols. Food Chem. 2011, 127, 1199−1203.(3) Boschin, G.; D’Agostina, A.; Annicchiarico, P.; Arnoldi, A. Effectof genotype and environment on fatty acid composition of Lupinusalbus L. seed. Food Chem. 2008, 108, 600−606.(4) Boschin, G.; Annicchiarico, P.; Resta, D.; D’Agostina, A.; Arnoldi,A. Quinolizidine alkaloids in seeds of lupin genotypes of differentorigins. J. Agric. Food Chem. 2008, 56, 3657−3663.(5) Pilvi, T. K.; Jauhiainen, T.; Cheng, Z. J.; Mervaala, E. M.;Vapaatalo, H.; Korpela, R. Lupin protein attenuates the developmentof hypertension and normalises the vascular function of NaCl-loadedGoto−Kakizaki rats. J. Physiol. Pharmacol. 2006, 57, 167−176.(6) Novicka, G.; Klosiewicz-Latoszek, L.; Sirtori, C. R.; Arnoldi, A.;Naruszewicz, M. Lupin proteins in the treatment of hypercholester-olemia. Atheroscler. Suppl. 2006, 7, 477.(7) Belski, R.; Mori, T. A.; Puddey, I. B.; Sipsas, S.; Woodman, R. J.;Ackland, T. R.; Beilin, L. J.; Dove, E. R.; Carlyon, N. B.; Jayaseena, V.;Hodgson, J. M. Effects of lupin-enriched foods on body compositionand cardiovascular disease risk factors: a 12-month randomizedcontrolled weight loss trial. Int. J. Obes. 2011, 35, 810−819.(8) Lee, Y. P.; Mori, T. A.; Puddey, I. B.; Sipsas, S.; Ackland, T. R.;Beilin, L. J.; Hodgson, J. M. Effects of lupin kernel flour-enriched breadon blood pressure: a controlled intervention study. Am. J. Clin. Nutr.2009, 89, 766−772.(9) Aronow, W. S. Treatment of systemic hypertension. Am. J.Cardiovasc. Dis. 2012, 2, 160−170.(10) Yang, H.-Y. T.; Erdos, E. G.; Levin, Y. Dipeptidylcarboxypeptidase that converts angiotensin I and inactivatesbradykinin. Biochim. Biophys. Acta, Protein Struct. 1970, 214, 374−376.(11) Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T.Antihypertensive effect of sour milk and peptides isolated from itthat are inhibitors to angiotensin I-converting enzyme. J. Dairy Sci.1995, 78, 1253−1257.(12) Wu, J.; Ding, X. Characterization of inhibition and stability ofsoy-protein-derived angiotensin I-converting enzyme inhibitorypeptides. Food Res. Int. 2002, 35, 367−375.(13) Aluko, R. E. Determination of nutritional and bioactiveproperties of peptides in enzymatic pea, chickpea, and mung beanprotein hydrolysates. J. AOAC Int. 2008, 91, 947−956.(14) Guang, C.; Phillips, R. D. Plant food-derived angiotensin Iconverting enzyme inhibitory peptides. J. Agric. Food Chem. 2009, 57,5113−5120.(15) Roy, F.; Boye, J. I.; Simpson, B. K. Bioactive proteins andpeptides in pulse crops: pea, chickpea and lentil. Food Res. Int. 2010,43, 432−442.(16) Boschin, G.; Scigliuolo, G. M.; Resta, D.; Arnoldi, A. ACE-inhibitory activity of enzymatic protein hydrolysates from lupin andother legumes. Food Chem. 2014, 145, 34−40.(17) Brambilla, F.; Resta, D.; Isak, I.; Zanotti, M.; Arnoldi, A. A label-free internal standard method for the differential analysis of bioactivelupin proteins using nano HPLC-chip coupled with ion trap massspectrometry. Proteomics 2009, 9, 272−286.
(18) Bradford, M. M. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing the principleof protein−dye binding. Anal. Biochem. 1976, 72, 248−254.(19) Levashov, P. A.; Sutherland, D. S.; Besenbacher, F.; Shipovskov,S. A robust method of determination of high concentrations ofpeptides and proteins. Anal. Biochem. 2009, 395, 111−112.(20) Goa, J. A micro biuret method for protein determination.Determination of total protein in cerebrospinal fluid. Scand. J. Clin.Lab. Invest. 1953, 5, 218−222.(21) Vermeirssen, V.; van Camp, J.; Verstraete, W. Bioavailability ofangiotensin I converting enzyme inhibitory peptides. Br. J. Nutr. 2004,92, 357−366.(22) Roberts, P. R.; Burney, J. D.; Black, K. W.; Zaloga, G. P. Effectof chain length on absorption of biologically active peptides from thegastrointestinal tract. Digestion 1999, 60, 332−337.(23) Rui, X.; Boye, J. I.; Simpson, B. K.; Prasher, S. O. Angiotensin I-converting enzyme inhibitory properties of Phaseolus vulgaris beanhydrolysates: effects of different thermal and enzymatic digestiontreatments. Food Res. Int. 2012, 49, 739−746.(24) Pedroche, J.; Yust, M. M.; Giron-Calle, J.; Alaiz, M.; Millan, F.;Vioque, J. Utilisation of chickpea protein isolates for production ofpeptides with angiotensin I-converting enzyme (ACE)-inhibitoryactivity. J. Sci. Food Agric. 2002, 82, 960−965.(25) Barbana, C.; Boye, J. I. Angiotensin I-converting enzymeinhibitory activity of chickpea and pea protein hydrolysates. Food Res.Int. 2010, 43, 1642−1649.(26) Perez-Vega, J. A.; Olivera-Castillo, L.; Gomez-Ruiz, J. A.;Hernandez-Ledesma, B. Release of multifunctional peptides bygastrointestinal digestion of sea cucumber (Isostichopus badionotus).J. Funct. Foods 2013, 5, 869−877.(27) Segura Campos, M. R.; Chel Guerrero, L. A.; Betancur Ancona,D. A. Angiotensin-I converting enzyme inhibitory and antioxidantactivities of peptide fractions extracted by ultrafiltration of cowpeaVigna unguiculata hydrolysates. J. Sci. Food Agric. 2010, 90, 2512−2518.(28) He, R.; Alashi, A.; Malomo, S. A.; Girgih, A. T.; Chao, D.; Ju, X.;Aluko, R. E. Antihypertensive and free radical scavenging properties ofenzymatic rapeseed protein hydrolysates. Food Chem. 2013, 141, 153−159.(29) Wu, J.; Aluko, R. E.; Muir, A. D. Production of angiotensin I-converting enzyme inhibitory peptides from defatted canola meal.Biores. Technol. 2009, 100, 5283−5287.(30) Korhonen, H. Milk-derived bioactive peptides: from science toapplications. J. Funct. Foods 2009, 1, 177−187.(31) Hong, F.; Ming, L.; Yi, S.; Zhanxia, L.; Yongquan, W.; Chi, L.The antihypertensive effect of peptides: a novel alternative to drugs?Peptides 2008, 29, 1062−1071.
Journal of Agricultural and Food Chemistry Article
dx.doi.org/10.1021/jf4039056 | J. Agric. Food Chem. 2014, 62, 1846−18511851