ontogeny of kainate receptor gene expression in the developing rat midbrain and striatum
TRANSCRIPT

Molecular Brain Research 104 (2002) 1–10www.elsevier.com/ locate/bres
Research report
O ntogeny of kainate receptor gene expression in the developing ratmidbrain and striatum
b a,d , a a*Vanessa Lilliu , Carla Perrone-Capano , Roberto Pernas-Alonso , Ramon Diaz Trelles ,a,c b aG. Luca Colucci d’Amato , Alessandro Zuddas , Umberto di Porzio
aInternational Institute of Genetics and Biophysics, CNR, Naples, ItalybChild Neurology and Psychiatry, Department of Neuroscience, University of Cagliari, Italy
cCentro di Endocrinologia e Oncologia Sperimentale ‘‘ G. Salvatore’’, CNR, Naples, ItalydUniversity of Catanzaro ‘‘ Magna Grecia’’, Department of Pharmacobiology, Roccelletta di Borgia (CZ), Italy
Accepted 8 March 2002
Abstract
Kainate (KA) receptors are a family of ionotropic glutamate receptors, which mediate the excitatory synaptic transmission in variousareas of the mammalian CNS. We have studied the expression pattern of the genes encoding for KA receptor subunits (Glur5-1, Glur5-2,Glur6, Glur7, KA1 and KA2) in rat prenatal (E), postnatal and adult ventral mesencephalon (MES) and striatum (STR) and in fetalmidbrain primary cultures. Each receptor subunit shows a unique area- and temporal-expression pattern. In MES the onset of both Glur5subunits is delayed when compared to the other subunits. In addition, most of the transcripts for KA subunits gradually increase duringembryonic development and show a slight decrease during the first postnatal week. Differently, Glur6 and KA2 mRNAs show a sharpincrease at E14.5 and decrease thereafter, reaching the lowest levels during late embryonic and postnatal development. In the STR, thegene expression of all KA subunit mRNAs is higher during embryonic development than after birth, except KA1 transcripts, that show apeak at P5. In embryonic MES primary cultures, Glur5-2, Glur6 and KA2 mRNAs are higher at the beginning of the culture whencompared to older cultures, while the other subunit mRNAs do not show significant variation throughout the days in vitro. Thus, all theKA receptor subunit transcripts appear independently regulated during MES and STR development, probably contributing to theestablishment of the fine tuning of the excitatory circuits reciprocally established between these CNS areas. 2002 Elsevier ScienceB.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Excitatory amino acid receptors: structure, function and expression
Keywords: Basal ganglia; Midbrain; Gene expression; Primary cultures; Dopamine
1 . Introduction velopmental plasticity processes. On the other hand,overactivation of Glurs may initiate a cascade of events,
L-Glutamate acts as an excitatory neurotransmitter at which produce excitotoxicity, neuronal injury and death.many synapses in the mammalian central nervous system Glurs are distributed in two main distinct classes, iono-(CNS). Interaction ofL-glutamate with postsynaptic re- tropic (iGlurs) and metabotropic. The iGlurs are mul-ceptors (Glurs) is responsible for a wide variety of brain timeric proteins that are further subdivided, on the basis ofphysiological and pathophysiological responses [34]. Acti- their selective susceptibility to different agonists, into:vation of Glurs regulates most of the fast synaptic trans- N-methyl-D-aspartate (NMDA), a-amino-3-hydroxy-5-mission and is involved in memory formation and de- methyl-4-isoxazole propionic acid (AMPA) and kainate
(KA) receptors [13]. Although synaptic functions ofNMDA and AMPA receptors have been well understood,
*Corresponding author. IIGB CNR Via P. Castellino 111, 80135until recently the physiological roles of kainate receptorsNaples, Italy. Tel.:139-81-6132-362/363; fax:139-81-6132-352.have been elusive [8,16]. The recent applications of newE-mail addresses: [email protected](C. Perrone-Capano),http: / /
www.iigb.na.cnr.it / (C. Perrone-Capano). drugs and genetic knockout technology are providing
0169-328X/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.PI I : S0169-328X( 02 )00196-1

2 V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10
information on the role of KA receptors as major players activity of both ventral midbrain and striatal neuronsin excitatory synaptic transmission [26]. Compared to [6,17,18].AMPA receptors, KA receptors elicit a slow kinetic and a To compare AMPA [27] and KA receptor ontogeneticsmall peak amplitude of the excitatory postsynaptic cur- expression and to understand how KA receptor subunitsrent, suggesting that they transmit information about the are regulated during embryonic and postnatal developmentglobal afferent firing rate of glutamatergic fibers acting as in MES and STR, we have characterized, by RT-PCRintegrator detectors [16,23]. In addition, KA receptors have techniques, the pattern of expression of the known KAbeen proposed to modulate synaptic transmission through a subunit mRNAs during rat ontogeny. For Glur5, we havepresynaptic action that reduces glutamate release on ex- analyzed both the Glur5 mRNA splice variants, Glur5-1citatory synapses [28] and enhances GABA release on and Glur5-2, which differ for the presence or the absence,interneurons [10,28] Moreover, there is increasing evi- respectively, of an exon encoding 15 amino acids in thedence that KA receptors are implicated in both short- and N-terminus [19,3]. KA receptor gene expression has alsolong-term plasticity in the hippocampus [9,20] and in been analyzed during embryonic MES development inplastic responses of dorsal horn neurons involved in primary cultures.nociception [22,24,29]. Kainate receptors are encoded bytwo gene subfamilies: the first encoding Glur5–Glur7subtypes and the second encoding KA1 and KA2, which 2 . Materials and methodsdiffer in their ligand affinity: low- (Glur5, Glur6, Glur7)and high-affinity (KA1, KA2). KA receptors diversity is 2 .1. Animals and dissectionsfurther enhanced by subunit variants due to mRNA alter-native splicing [19,39,3] and mRNA editing processes Timed pregnant Sprague–Dawley rats (Charles Riverwhich regulates KA receptor channels conductance and Breeding Labs., Milan, Italy) were sacrificed in accordancecalcium permeability [8,13,24]. It has been established that with Society for Neuroscience guidelines and Italian law.only Glur5–Glur7 form functional homomeric channels Embryonic age (E) was determined by considering the dayreceptors [13,39] but also can exhibit promiscuous coas- of insemination (as confirmed by vaginal plug) as day E0.sembly [11]; KA1 and KA2 associate with Glur5–7 Embryos from 11.5 to 20 days of gestation were quicklysubunits to form heteromeric receptors with distinct phar- removed and placed in phosphate buffered saline (PBS)macological properties [32,39]. The precise composition of without calcium and magnesium and supplemented with 33most naive kainate receptors is not known with certainty, mM glucose. The ventral mesencephalon and the striatumhowever, expression patterns for the different subunits from various prenatal and postnatal ages were carefullyshow distinct, but overlapping distributions [43], suggest- dissected under sterile conditions and processed for RNAing the existence of possible combinations of native and protein isolation or for cell cultures.heteromeric receptors.
KA receptors are abundant throughout the entire nervous2 .2. Cell culturessystem [13,26]. However, temporal regulation of KAreceptor subunit gene expression and their expression Cells were dissociated from E13 rat ventral MES andprofiles during neurogenesis is still poorly understood. cultured as previously described [15]. Briefly, the tissuesTheir expression and function differs during embryogene- were dissected from embryos and dissociated using me-sis and in adult age [1,43,44]. KA1 is ubiquitous in the chanical trituration with a fire polished Pasteur pipette indeveloping mouse brain but restricted to few areas in the Earle’s balanced salt solution (EBSS, Sigma, St. Louis,adult [21]; transient hyperpolarization (inhibitory func- MO, USA) supplemented with 26 mM sodium bicarbonate,tion?) of KA receptors has been described in the rodent 1 mM cysteine, 0.5 mM EDTA, pH 7.4, 1 mg/ml bovinemesencephalon between embryonic day (E) 13 and E15, serum albumin (BSA, Sigma), 1 mg/ml ovomucoidwhen part of the mesencepahlic neurons start to respond (Sigma) and 0.01% pancreatic deoxyribonuclease (Sigma),canonically, with depolarization [14]. KA receptors are centrifuged 10 min at 800g, resuspended in platinghighly expressed in the basal ganglia and mesencephalon, medium and counted. Cells were plated at a density of
2suggesting their involvement in the control of the central 125 000/cm (12 multiwell plates, Costar, Milan, Italy), indopaminergic transmission [4,7,42,44]. The dopaminergic dishes coated with 15mg/ml of poly-D-Lysine (Sigma).neurons of the substantia nigra and ventral tegmental area Cells were grown in minimum essential medium andin the MES receive glutamatergic innervation, which plays nutrient mixture F12 (1:1, Gibco-BRL, Milan, Italy)an important role in modulating dopaminergic neurotrans- supplemented with glucose to reach 33 mM, 7.5% fetalmission and plasticity [25,31]. In turn, the striatum re- bovine serum (HyClone Labs., Milan, Italy) and 5% heatceives an extensive afferent glutamatergic input from inactivated horse serum (HyClone); 10 mM cytosineb-D-diverse regions of the cerebral cortex. Glutamatergic arabinofuranoside (Sigma) was added to the cultures afterafferents play a crucial role in controlling neuron basal 4 days in vitro to inhibit non-neuronal cell proliferation.

V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10 3
Cultures were always carried in triplicate sister samples for extension step (728C for 7 min). The number of cycleseach experimental point analyzed and were processed was experimentally chosen in order to fall into theseparately. exponential phase of the amplification reaction. The for-
ward and reverse primers (Genset, Paris, France) used are2 .3. RT-PCR analyses listed in Table 1, together with the length of amplified
fragments. Forward and reverse primers used for Glur5RNA was extracted from pools (5–18 embryos) of were designed in order to amplify across a splicing site;
embryonic rat midbrain or striatum of the same age or thus they give two amplified fragments corresponding tofrom single adult tissues, using the Tri-Reagent isolation the two molecular forms of Glur5 mRNA Glur5-1 andsystem (Molecular Research Center, Cincinnati, OH, USA) Glur5-2, differing for the presence or the absence of anand following the manufacturer’s instructions. The analy- insert of 40 bp [19,3]. Nonreverse-transcribed RNA tem-ses were always carried in triplicate obtained from differ- plates and mock controls were always run in PCR re-ent pools or tissue samples for each experimental point actions and never gave amplification products. Each RT-analyzed and were processed separately. The yield and the PCR from any given sample was assayed in at least threeintegrity of RNA were determined by spectrophotometrical independent reactions. Identity of each amplified fragmentmeasurement ofA and agarose gel electrophoresis, (Glur5, Glur6, Glur7, KA1 and KA2) was confirmed by260
respectively. RT-PCR analyses were as previously de- digestion with restriction enzymes. Identity of Glur5-1 andscribed [35]. Briefly, total RNA was treated with RQ1 Glur5-2 PCR products was assessed by sequence analysis.DNase (Promega, Florence, Italy) to prevent false results The co-amplification procedure allows to compare theby DNA contamination; the enzyme was removed by three expression of kainate Glur subunits and overcome theconsecutive treatments with 6ml of Strataclean resin inherent variations present within any individual PCR by(Stratagene, La Jolla, CA, USA), following the manufac- normalizing to HPRT mRNA level [35].
32turer’s instructions. A 2-mg amount of RNA were reverse The P-labeled amplified products were separated bytranscribed using random hexanucleotides as primers (6 electrophoresis in 1.5–2% agarose gel, dried and exposedmM) and 13 U of avian myeloblastoma virus (AMV) to a PhosphorImager screen. Quantitation was achieved byreverse transcriptase (RT; Promega). Of the reverse tran- integrating the volume areas of each fragment obtainedscribed cDNA 1/20 was amplified in a 25-ml reaction from scanning the screens with a PhosphorImager ap-mixture containing Taq DNA polymerase buffer (Phar- paratus (Molecular Dynamics, Sunnyvale, CA, USA),macia), 0.2 mM dNTPs, 0.4mM each primer, 1 U Taq equipped with anIMAGEQUANT software. The ratio between
32DNA polymerase (Pharmacia) and 1mCi [ P]dCTP (3000 the yield of each amplified product and that of co-am-Ci /mmol, Amersham, Milan, Italy). plified HPRT allowed a relative estimate of the mRNA
Different sets of primer pairs were used in the same levels [35]. Triplicate samples allowed statistical analysisreaction tube to co-amplify cDNA, together with primers (see below). This technique allows quantitative comparisonfor the hypoxantine-phosphoribosyl transferase (HPRT), a of transcripts amplified with the same primer pairs inconstantly expressed gene during CNS development, used different samples. However it is not suitable to compareas internal standard [37,41]. After a first denaturing step at values of amplified products for different primers since95 8C for 5 min, PCR amplification was performed for 27 their efficiency of annealing to target sequences can vary;cycles organized as follows: 958C for 0.5 min; 568C (for as a consequence, the intensity of different amplifiedGlur5, Glur6 and KA2) or 578C (Glur7 and KA1) for 0.5 fragments does not reflect true differences in templatemin; 728C for 0.5 min, and was followed by a final abundance [35].
Table 1aSequence of primers used in RT-PCR experiments
Primer (59-39) Forward (59-39) Reverse Size (bp) of PCRname products
HPRT CCTGCTGGATTACATTAAAGCACTG CCTGAAGTACTCATTATAGTCAAGG 370Glur5-1 AGAGGCTGGTGGTGGTTGACT CGCCCTTTCTCGTTGAACTGC 265Glur5-2 as Glur5-1 as Glur5-1 220Glur6 CAGGGTTCAGGATACTAAAT CCCATAGAGTGGTTTGTCAG 536Glur7 TTCCGCAAGTCTGATAGGAC GCTGAATCTGGCAATGACG 417KA1 CCAACTCCAGAGATTCACC TGGCATGGATGATGATAGT 272KA2 GGACGACCAGACCGTGTGT TGGCATCGATGATGATGGT 566a Forward and reverse primers used for Glur5 were designed in order to amplify across a splicing site; thus they give two amplified fragmentscorresponding to the two molecular forms of Glur5 mRNA Glur5-1 and Glur52, differing for the presence or the absence of an insert of 40 bp.

4 V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10
2 .4. Statistical analysis 3 .1.1. Glur5-1 mRNAIn the ventral MES, Glur5-1 mRNA was undetectable at
`Analysis of variance was used to compare data (Scheffe E11.5 when transcripts encoding for other KA subunitsF-test). were already observed (such as Glur6, KA1 and KA2).
Glur5-1transcripts appeared at E13 and gradually increaseduntil E17. After this stage, Glur5-1 mRNA level started togradually decrease, reaching a value in the adult slightly
3 . Results higher than at E13 (Fig. 1A).In the STR, Glur5-1 mRNA levels gradually decreased
3 .1. In vivo pattern of Glur subunits gene expression from the embryonic period to the postnatal life, reachingduring MES and STR ontogeny their lowest values two week after birth and in the adult
(Fig. 2A).By using a semiquantitative RT-PCR reaction assay, we Collectively, Glur5-1 gene expression was higher in
examined relative changes in the levels of Glur5-1, Glur5- MES than in STR.2, Glur6, Glur7, KA1 and KA2 transcripts in rat MES andSTR during neurogenesis starting from E11.5 (MES) and 3 .1.2. Glur5-2 mRNAE15 (STR) until the adult age. As Glur5-1, Glur5-2 mRNA in MES was undetectable at
Fig. 1. Pattern of gene expression of Glur subunits during ontogeny of the rat mesencephalon. The diagrams show the relative quantitation (mean6S.E.) ofthe Glur5-1 (A), Glur5-2 (B), Glur6 (C), Glur7 (D), KA1 (E) and KA2 (F) amplified products compared to that of the hypoxanthine phosphoribosyltransferase (HPRT, internal standard). Data are expressed as ratio Glur /HPRT at various embryonic (E) and postnatal (P) ages and in the adult (Ad). Thegels in the insets show an example of the coamplified products of Glurs and HPRT at any given age studied. The size of the amplified products is shown on
`the left in base pairs (bp). *,P,0.05 when compared to the E13 value (ANOVA, ScheffeF-test).

V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10 5
Fig. 2. Pattern of gene expression of Glur subunit mRNAs during ontogeny of the rat striatum. The diagrams show the relative quantitation (mean6S.E.) ofGlur5-1 (A), Glur5-2 (B), Glur6 (C), Glur7 (D), KA1 (E) and KA2 (F) amplified products compared to that of the hypoxanthine phosphoribosyltransferase (HPRT, internal standard). Data are expressed as ratio Glur /HPRT at various embryonic (E) and postnatal (P) ages and in the adult (Ad). Thegels in the insets show an example of the products of Glurs and HPRT at any given age studied. The size of the amplified products is shown on the left in
`base pairs (bp). *,P,0.05 when compared to the E13 value (ANOVA, ScheffeF-test).
the first time point analyzed (E11.5) and appeared at E13. adult, with a transient and statistically significant decreaseIts expression profile appeared rather irregular but on at P15, when it reached its lowest value (Fig. 2B). Onaverage it did not show significant variation throughout average, the level of Glur5-2 gene expression was similarembryonic development and adult age when compared to in MES and in STR.E13 (Fig. 1B).
In the STR Glur5-2 mRNA showed a rather constant 3 .1.3. Glur6 mRNAlevel of expression from the first time point analyzed until Contrary to Glur5-1 and Glur5-2 transcripts, Glur6

6 V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10
mRNA was already detectable in the ventral MES at The overall KA receptor gene expression in both MESE11.5, although at a very low level. Its expression profile and STR is summarized in Table 2 and is compared to thatshowed a sharp and transient peak between E13 and E14.5, of the AMPA Glurs, previously described [27]. The patternwhen it reached a value about 100-fold higher than at in the MES is different among the five KA Glurs, andE11.5. Glur6 transcript levels rapidly decreased at E15 and shows a sustained expression in the late embryonic andthereafter remain quite stable throughout development early postnatal ages. The only exception is Glur6, which(Fig. 1C). shows a different pattern, with a peak during early
In the STR, Glur6 mRNA showed a level of expression development and a modest expression thereafter. AMPAslightly higher in the embryonic age when compared to the Glurs gene expression appears on average sustained duringpostnatal period, although differences among most of the all embryonic and postnatal time points.time points analyzed were not statistically significant. A In the STR, on the other hand, all KA Glurs arestatistically significant decrease was observed when E15 expressed at higher levels during the embryonic period andvalues were compared to P15 values (Fig. 2C). the first days after birth, similarly to the AMPA Glurs gene
expression.3 .1.4. Glur7 mRNA
In rat MES, Glur7 mRNA was undetectable at E11.5 3 .2. Pattern of Glur subunits gene expression in MESand appeared starting from E13. Its level increased at leastprimary culturesthree-fold between E13 and E15, when it reached aplateau, which was maintained until P3. Glur7 transcript The relative levels of all KA receptor subunit mRNAslevels decreased thereafter until adult life, with a sharp and were evaluated in primary cultures generated from E13transient peak at P7 (Fig. 1D). MES.
In the STR, Glur7 mRNA showed a level of expression On average, the pattern of mRNA levels of Glur 5-1,at least three-fold higher during all the embryonic stages Glur7 and KA1 did not show significant variation duringwhen compared to the postnatal period (Fig. 2D). the 2 weeks of culture (Fig. 3A, D and E, respectively). On
the contrary, Glur5-2 mRNA level significantly decreased3 .1.5. KA1 mRNA only between the first and the last day of the culture (Fig.
In MES, KA1 transcripts appeared at quite low levels at 3B). Glur6 and KA2 transcripts showed a similar pattern inE11.5. Their levels increased reaching a peak between E16 vitro, being both higher during the first day in vitro andand E17 and remained slightly lower thereafter until adult showing a 50% decrease during the following 2 weeks ofage (Fig. 1E). culture (Fig. 3C and F, respectively).
In the STR, KA1 mRNA levels gradually increasedduring prenatal and early postnatal development, reachinga three-fold significant increase at P5 when compared to 4 . Discussionthe first time point analyzed (E15). After P5, KA1 geneexpression gradually decreased until the adult age, reach- A full understanding of the mechanisms and pathwaysing E15 values (Fig. 2E). Collectively, KA1 gene expres- necessary for the maturation of excitatory glutamatergicsion was higher in STR than in MES. synapses still remains elusive. The development of excitat-
ory circuitry is associated with the onset of the various3 .1.6. KA2 mRNA Glur subtypes. KA receptors represent a family of ionot-
In MES, KA2 mRNA was detectable from the first time ropic Glurs with still poorly understood functions. Recentpoint analyzed (E11.5), although at low values. Its levels evidence suggests that they mediate both post- and pre-rapidly increased, with a significant peak at E14.5; at this synaptic signaling in the mammalian CNS, and are in-stage, its relative values were more than 30-fold higher volved in brain plasticity [9,20]. Developmental andthan at E11.5. After E14.5, KA2 mRNA levels decreased activity-dependent changes in the functional synapticreaching at the end of embryonic life a value corre- expression of these receptors are subject to tight cellularsponding to 60% of the peak value. After birth, KA2 gene controls, including transcriptional and translational regula-expression was constant until adult life (Fig. 1F). tion. Altogether up- and downregulation of the various KA
In the STR, KA2 mRNA levels were high from the first receptor subunits occur not only throughout development,time point analyzed (E15) and during all the time points of but also following seizures, repetitive activation of affer-the embryonic development. KA2 transcripts decreased ents, or chronic administration of a variety of drugs [33].slightly in the first postnatal week and the decrease was It is well known that KA receptor subunits are abun-more robust after the first postnatal week. KA2 gene dantly distributed in the ventral mesencephalon and theexpression was almost three-fold lower in the adult STR, striatum [4,7,42,44], where they play a crucial role inwhen compared to fetal development (Fig. 2F). controlling basal activity and plasticity of both ventral
Altogether, KA2 gene expression was at least four-fold midbrain and striatal neurons which form important func-higher in STR than in MES, at all the time points analyzed. tional circuits in the mammalian CNS [36]. The aim of this
V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10 7
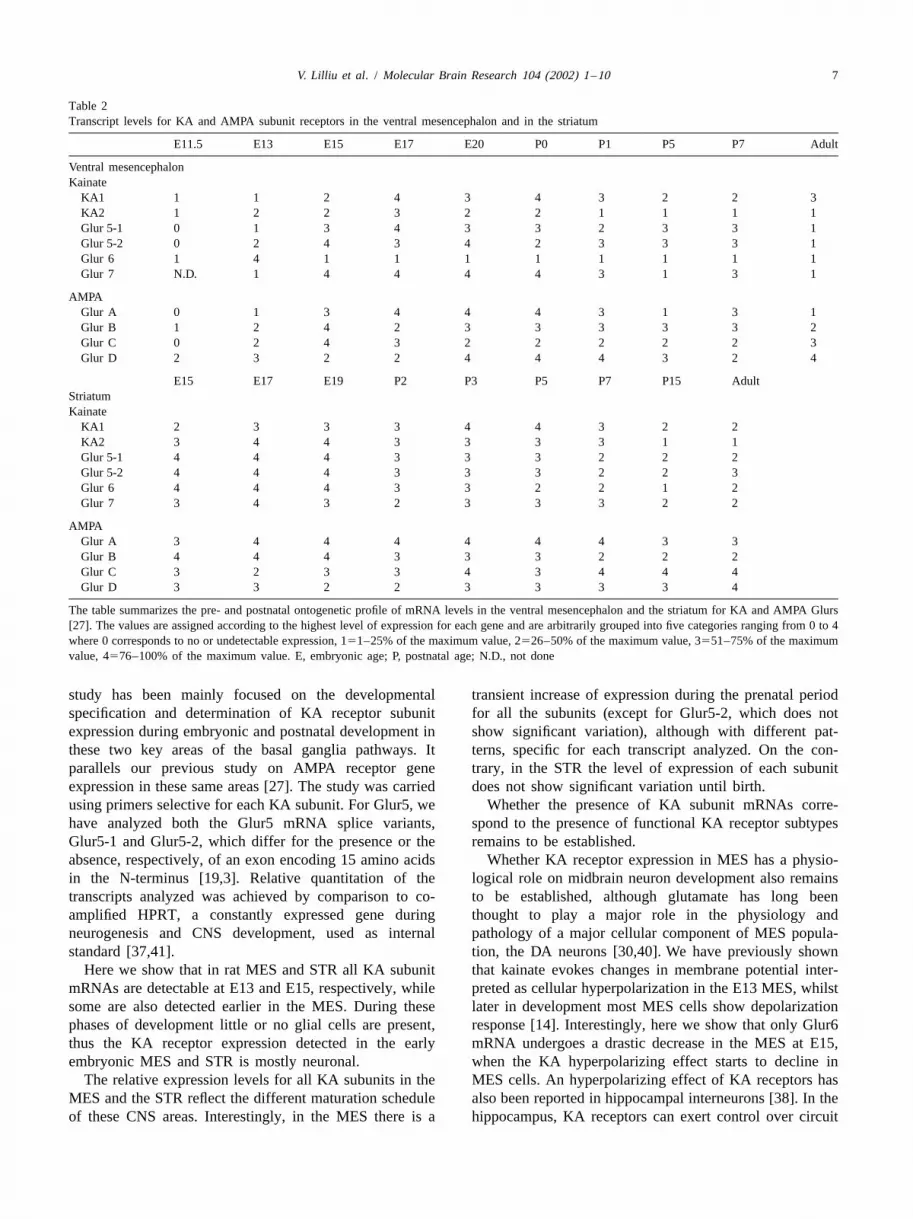
Table 2Transcript levels for KA and AMPA subunit receptors in the ventral mesencephalon and in the striatum
E11.5 E13 E15 E17 E20 P0 P1 P5 P7 Adult
Ventral mesencephalonKainate
KA1 1 1 2 4 3 4 3 2 2 3KA2 1 2 2 3 2 2 1 1 1 1Glur 5-1 0 1 3 4 3 3 2 3 3 1Glur 5-2 0 2 4 3 4 2 3 3 3 1Glur 6 1 4 1 1 1 1 1 1 1 1Glur 7 N.D. 1 4 4 4 4 3 1 3 1
AMPAGlur A 0 1 3 4 4 4 3 1 3 1Glur B 1 2 4 2 3 3 3 3 3 2Glur C 0 2 4 3 2 2 2 2 2 3Glur D 2 3 2 2 4 4 4 3 2 4
E15 E17 E19 P2 P3 P5 P7 P15 AdultStriatumKainate
KA1 2 3 3 3 4 4 3 2 2KA2 3 4 4 3 3 3 3 1 1Glur 5-1 4 4 4 3 3 3 2 2 2Glur 5-2 4 4 4 3 3 3 2 2 3Glur 6 4 4 4 3 3 2 2 1 2Glur 7 3 4 3 2 3 3 3 2 2
AMPAGlur A 3 4 4 4 4 4 4 3 3Glur B 4 4 4 3 3 3 2 2 2Glur C 3 2 3 3 4 3 4 4 4Glur D 3 3 2 2 3 3 3 3 4
The table summarizes the pre- and postnatal ontogenetic profile of mRNA levels in the ventral mesencephalon and the striatum for KA and AMPA Glurs[27]. The values are assigned according to the highest level of expression for each gene and are arbitrarily grouped into five categories ranging from 0to 4where 0 corresponds to no or undetectable expression, 151–25% of the maximum value, 2526–50% of the maximum value, 3551–75% of the maximumvalue, 4576–100% of the maximum value. E, embryonic age; P, postnatal age; N.D., not done
study has been mainly focused on the developmental transient increase of expression during the prenatal periodspecification and determination of KA receptor subunit for all the subunits (except for Glur5-2, which does notexpression during embryonic and postnatal development in show significant variation), although with different pat-these two key areas of the basal ganglia pathways. It terns, specific for each transcript analyzed. On the con-parallels our previous study on AMPA receptor gene trary, in the STR the level of expression of each subunitexpression in these same areas [27]. The study was carried does not show significant variation until birth.using primers selective for each KA subunit. For Glur5, we Whether the presence of KA subunit mRNAs corre-have analyzed both the Glur5 mRNA splice variants, spond to the presence of functional KA receptor subtypesGlur5-1 and Glur5-2, which differ for the presence or the remains to be established.absence, respectively, of an exon encoding 15 amino acids Whether KA receptor expression in MES has a physio-in the N-terminus [19,3]. Relative quantitation of the logical role on midbrain neuron development also remainstranscripts analyzed was achieved by comparison to co- to be established, although glutamate has long beenamplified HPRT, a constantly expressed gene during thought to play a major role in the physiology andneurogenesis and CNS development, used as internal pathology of a major cellular component of MES popula-standard [37,41]. tion, the DA neurons [30,40]. We have previously shown
Here we show that in rat MES and STR all KA subunit that kainate evokes changes in membrane potential inter-mRNAs are detectable at E13 and E15, respectively, while preted as cellular hyperpolarization in the E13 MES, whilstsome are also detected earlier in the MES. During these later in development most MES cells show depolarizationphases of development little or no glial cells are present, response [14]. Interestingly, here we show that only Glur6thus the KA receptor expression detected in the early mRNA undergoes a drastic decrease in the MES at E15,embryonic MES and STR is mostly neuronal. when the KA hyperpolarizing effect starts to decline in
The relative expression levels for all KA subunits in the MES cells. An hyperpolarizing effect of KA receptors hasMES and the STR reflect the different maturation schedule also been reported in hippocampal interneurons [38]. In theof these CNS areas. Interestingly, in the MES there is a hippocampus, KA receptors can exert control over circuit

8 V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10
Fig. 3. Pattern of gene expression of Glur subunits during in vitro ontogeny in rat mesencephalon primary cultures. The diagrams show the relativequantitation (mean6S.E.) of the Glur5-1 (A), Glur5-2 (B), Glur6 (C), Glur7 (D), KA1 (E) and KA2 (F) amplified products compared to that of thehypoxanthine phosphoribosyl transferase (HPRT, internal standard). Data are expressed as ratio Glur /HPRT at various days in vitro (D.i.v.). Cultures wereestablished from E13 ventral mesencephalon and carried in the presence of serum. The gels in the insets show an example of the coamplified products ofGlurs and HPRT at any given D.i.v. analyzed. The size of the amplified products is shown on the left in base pairs (bp). *,P,0.05 when compared to the
`D.i.v. 1 value (ANOVA, ScheffeF-test).
excitability by two parallel mechanisms: upregulation of the expression profile in vivo. In fact, most KA subunitsexcitatory activity through Glur6 and upregulation of gene expression in MES primary cultures show littleinhibitory activity through Glur5 [16]. They also have well difference between the majority of samples at variousdescribed epileptogenic activity, however recent observa- D.i.v. Only GluR6 and KA2 mRNAs show a peak oftions suggest that kainate receptors can also exert a expression at the first D.i.v., which perfectly match thosesurprising antiepileptogenic effect, mediated through observed in MES in vivo at E14.GABA neurotransmission [2,26]. Thus, excitatory and In conclusion, our results show that KA receptorinhibitory activities of KA receptors appear correlated with subunits display distinct developmental expression patternsubunit expression of Glur5 and Glur6 in the hippocampus in MES and STR, and that are all downregulated over[5]. Thus, it is possible that also the fine control of development, suggesting a functional emergence of differ-nigro-striatal circuit excitability could be accomplished by ent glutamate receptor activation within these areas. Inmodulating the relative levels of Glur6 and Glur5 in MES addition, our report also shows that the expression of KAand in STR. Indeed both Glur5 spliced forms are different- and AMPA receptor subunits [27] is differently regulatedly regulated in MES and STR during embryonic develop- during development and in the adult MES and STR (Tablement. 2). This observation supports the idea that KA and AMPA
Unlike AMPA receptor genes [27], the KA receptor gene receptor subtypes have distinct functions, as suggested byexpression profile in MES in vitro did not always mimic their lack of colocalization at individual synapses [12].

V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10 9
Dopamine synthesis precedes dopamine uptake in embryonic ratThe knowledge of the expression pattern of KA receptormesencephalic neurons, J. Neurochem. 56 (1991) 392–399.subunits during MES and STR ontogeny provides a
[16] M. Frerking, R.A. Nicoll, Synaptic kainate receptors, Curr. Opin.framework to analyze developmental disorders specifically Neurobiol. 10 (2001) 342–351.associated with excitatory circuits of the basal ganglia. [17] M. Garcia-Munoz, S.J. Young, P.M. Groves, Terminal excitability of
the corticostriatal pathway. I. Regulation by dopamine receptorstimulation, Brain Res. 551 (1991) 195–206.
[18] M. Garcia-Munoz, S.J. Young, P.M. Groves, Terminal excitability ofthe corticostriatal pathway. II. Regulation by glutamate receptorA cknowledgementsstimulation, Brain Res. 551 (1991) 207–215.
[19] P. Gregor, B.F. O’Hara, X. Yang, G.R. Uhl, Expression and novelCNR Biotechnology Target Project, CNR Progetto subunit isoforms of glutamate receptor genes Glur5 and Glur6,
Strategico Neurodegenerazione, MURST/MIUR Cofin. NeuroReport 4 (1993) 1343–1346.[20] J.E. Huettner, Kainate receptors: knocking out plasticity, Trends2001 (N. 2001052911 002), and Grant MURST cluster 02
] Neurosci. 24 (2001) 365–366.to IIGB, have supported this work. We thank B. Esposito,[21] K. Kask, J. Jerecic, D. Zamanillo, J. Wilbertz, R. Sprengel, P.H.
T. Vespa and R. Vito for technical help. Seeburg, Developmental profile of kainate receptor subunit KA1revealed by Cre expression in YAC transgenic mice, Brain Res. 876(2000) 55–61.
[22] G.A. Kerchner, T.J. Wilding, P. Li, M. Zhuo, J.E. Huettner,Presynaptic kainate receptors regulate spinal sensory transmission, J.R eferencesNeurosci. 21 (2001) 59–66.
[23] P. Konig, A.K. Engel, W. Singer, Integrator or coincidence detector?[1] S. Bahn, B.Volk, W. Wisden, Kainate receptor gene expression in the The role of the cortical neuron revisited, Trends Neurosci. 19 (1996)
developing rat brain, J. Neurosci. 9 (1994) 5525–5547. 415–416.[2] Y. Ben-Ari, R. Cossart, Kainate, a double agent that generates [24] C.J. Lee, H. Kong, M.C. Manzini, C. Albuquerque, M.V. Chao, A.B.
seizures: two decades of progress, Trends Neurosci. 23 (2000) MacDermott, Kainate receptors expressed by a subpopulation of21580–587. developing nociceptors rapidly switch from high to low Ca
[3] B. Bettler, J. Boulter, I. Hermans-Borgmeyer, A. O’Shea-Greenfield, permeability, J. Neurosci. 21 (2001) 4572–4581.E.S. Deneris, C. Moll, U. Borgmeyer, M. Hollmann, S. Heinemann, [25] M. Legault, R.A. Wise, Novelty-evoked elevations of nucleusCloning of a novel glutamate receptor subunit, Glur5: expression in accumbens dopamine: dependence on impulse flow from the ventralthe nervous system during development, Neuron 5 (1990) 583–595. subiculum and glutamatergic neurotransmission in the ventral
[4] S. Bischoff, J. Barhanin, B. Bettler, C. Mulle, S. Heinemann, Spatial tegmental area, Eur. J. Neurosci. 13 (2001) 819–828.distribution of kainate receptor subunit mRNA in the mouse basal [26] J. Lerma, A.V. Paternain, A. Rodriguez-Moreno, J.C. Lopez-Garcia,ganglia and ventral mesencephalon, J. Comp. Neurol. 379 (1997) Molecular physiology of kainate receptors, Physiol. Rev. 81 (2001)541–562. 971–998.
[5] I. Bureau, S. Bischoff, S.F. Heinemann, C. Mulle, Kainate receptor- [27] V. Lilliu, R. Pernas-Alonso, R. Diaz-Trelles, A. Zuddas, U. dimediated responses in the CA1 field of wild-type and Glur6- Porzio, C. Perrone-Capano, Ontogeny of AMPA receptor genedeficient mice, J. Neurosci. 15 (1999) 653–663. expression in the developing rat midbrain and striatum, Mol. Brain
[6] P. Calabresi, A. Pisani, N.B. Mercuri, G. Bernardi, The corticostriat- Res. 96 (2001) 133–141.al projection: from synaptic plasticity to dysfunctions of the basal [28] A.B. MacDermott, L.W. Role, S.A. Siegelbaum, Presynaptic ionot-ganglia, Trends Neurosci. 19 (1996) 19–24. ropic receptors and the control of transmitter release, Annu. Rev.
[7] K. Chergui, A. Bouron, E. Normand, C. Mulle, Functional Glur6 Neurosci. 22 (1999) 443–485.kainate receptors in the striatum: indirect downregulation of synaptic [29] A.B. MacDermott, Glutamate and GABA. A painful combination,transmission, J. Neurosci. 20 (2000) 2175–2182. Neuron 32 (2001) 376–378.
[8] R. Chittajallu, S.P. Braithwaite, V.R. Clarke, J.M. Henley, Kainate [30] J.H. Meador-Woodruff, D.J. Healy, Glutamate receptor expression inreceptors: subunits, synaptic localization and function, Trends schizophrenic brain, Brain Res. Rev. 31 (2000) 288–294.Pharmacol. Sci. 20 (1999) 27–35. [31] L.T. Meltzer, C.L. Christoffersen, K.A. Serpa, Modulation of
[9] A. Contractor, G. Swanson, S.F. Heinemann, Kainate receptors are dopamine neuronal activity by glutamate receptor subtypes, Neuro-involved in short- and long-term plasticity at mossy fiber synapses sci. Biobehav. Rev. 21 (1997) 511–518.in the hippocampus, Neuron 29 (2001) 209–216. [32] C. Mulle, A. Sailer, G.T. Swanson, C. Brana, S. O’Gorman, B.
[10] R. Cossart, R. Tyzio, C. Dinocourt, M. Esclapez, J.C. Hirsch, Y. Bettler, S.F. Heinemann, Subunit composition of kainate receptorsBen-Ari, C. Bernard, Presynaptic kainate receptors that enhance the in hippocampal interneurons, Neuron 28 (2000) 475–484.release of GABA on CA1 hippocampal interneurons, Neuron 29 [33] S.J. Myers, R. Dingledine, K. Borges, Genetic regulation of(2001) 497–508. glutamate receptor ion channels, Annu. Rev. Pharmacol. Toxicol. 39
[11] C. Cui, M.L. Mayer, Heteromeric kainate receptors formed by the (1999) 221–241.coassembly of Glur5, Glur6, and Glur7, J. Neurosci. 19 (1999) [34] S. Ozawa, H. Kamiya, K. Tsuzuki, Glutamate receptors in the8281–8291. mammalian central nervous system, Prog. Neurobiol. 54 (1998)
[12] S.H. DeVries, E.A. Schwartz, Kainate receptors mediate synaptic 581–618.transmission between cones and ‘Off’ bipolar cells in a mammalian [35] R. Pernas-Alonso, F. Morelli, U. di Porzio, C. Perrone Capano,retina, Nature 397 (1999) 157–160. Multiplex semiquantitative reverse transcriptase-PCR of low abun-
[13] R. Dingledine, K. Borges, D. Bowie, S.F. Traynelis, The glutamate dance neuronal mRNAs, Brain Res. Protoc. 4 (1999) 395–406.receptor ion channels, Pharmacol. Rev. 51 (1999) 7–61. [36] C. Perrone-Capano, U. di Porzio, Genetic and epigenetic control of
[14] U. di Porzio, S.V. Smith, E.A. Novotny, F. Morelli, J.L. Barker, Two midbrain dopaminergic neuron development, Int. J. Dev. Biol. 44functionally different glutamate receptors of the kainate subtype in (2000) 679–687.embryonic rat mesencephalic cells, Exp. Neurol. 120 (1993) 202– [37] C. Perrone-Capano, A. Tino, U. di Porzio, Target cells modulate213. dopamine transporter gene expression during brain development,
[15] M.L. Fiszman, A. Zuddas, M.I. Masana, J.L. Barker, U. di Porzio, NeuroReport 5 (1994) 1145–1148.

10 V. Lilliu et al. / Molecular Brain Research 104 (2002) 1–10
[38] A. Rodriguez-Moreno, J.C. Lopez-Garcia, J. Lerma, Two popula- exposure: a comparative polymerase chain reaction study, Mol.tions of kainate receptors with separate signaling mechanisms in Pharmacol. 43 (1993) 694–701.hippocampal interneurons, Proc. Natl. Acad. Sci. USA 97 (2000) [42] F.I. Tarazi, A. Campbell, S.K. Yeghiayan, R.J. Baldessarini, Locali-1293–1298. zation of ionotropic glutamate receptors in caudate-putamen and
[39] H.H. Schiffer, G.T. Swanson, S.F. Heinemann, Rat Glur7 and a nucleus accumbens septi of rat brain: comparison of NMDA,carboxy-terminal splice variant, Glur7b, are functional kainate AMPA, and kainate receptors, Synapse 30 (1998) 227–235.receptor subunits with a low sensitivity to glutamate, Neuron 19 [43] T.J. Wilding, J.E. Huettner, Functional diversity and developmental(1997) 1141–1146. changes in rat neuronal kainate receptors, J. Physiol. 532 (2001)
[40] M.S. Starr, S. Kaur, Stimulation of basal andL-DOPA-induced 411–421.motor activity by glutamate antagonists in animal models of [44] U. Wullner, D.G. Standaert, C.M. Testa, J.B. Penney, A.B. Young,Parkinson’s disease, Neurosci. Biobehav. Rev. 21 (1997) 437–446. Differential expression of kainate receptors in the basal ganglia of
[41] M. Steel, N.J. Buckley, Differential regulation of muscarinic re- the developing and adult rat brain, Brain Res. 768 (1997) 215–223.ceptor mRNA levels in neuroblastoma cells by chronic agonist