nociceptor excitation by thermal sensitization--a hypothesis
TRANSCRIPT

J. Sandkiihler, B. Bromm and GE Gebhart (Ed%) Progress in Brain Research, Vol. 129 0 2000 Elsevier Science B.V. All rights reserved
CHAPTER 3
Nociceptor excitation by thermal sensitization - a hypothesis
Peter W. Reeh L* and Gcibor Pethij 2
’ Institute of Physiology and Experimental Pathophysiolog): UniversitZitsstl: 17, O-91054 Erlangen, Germany 2 Institute of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Pets, Szigeti ~4.12, H-7624 Pets, Hungary
Introduction
Sensitivity to noxious heat of the skin and oral cavity appears to be a useful protective mechanism. How- ever, many deep tissues including skeletal muscle, dura, testis and colon, are also reported to be in- nervated by polymodal, mechano-heat-sensitive pri- mary afferents (Kumazawa et al., 1991; Bove and Moskowitz, 1997; Kumazawa and Mizumura, 1997; Su and Gebhart, 1998). Fig. 1 originates from a single-fiber recording in the rabbit phrenic nerve, in vitro, and the recorded unit formed a mechanore- ceptive field in the epineurium of the nerve and the joined mediastinal pleura. Radiant heat was very effective in inducing a sharp rise of the discharge frequency from close to 40°C on, and a very steep stimulus-response function precisely encoded the tissue temperature up to the peak of 46°C. An- other surprising deep tissue with graded responses to noxious heat is the peripheral nerve (Fig. 2). Axons of the desheathed sciatic nerve have recently been shown to release calcitonin gene-related peptide (CGRP) in response to heat. The threshold and slope of the stimulus-response function exhibits typical nociceptive magnitudes (Sauer et al., 1999). Thus,
*Corresponding author: P.W. Reeh, Institute of Physi- ology and Experimental Pathophysiology, Universit%str. 17, D-91054 Erlangen, Germany. Tel.: +49 (9131) 852- 2228; Fax: +49 (913 1) 852-2497; E-mail: [email protected]
sensitivity to noxious heat is not restricted to sensory nerve endings.
What may be the biological purpose of a sensory capacity to signal core temperatures that are more than sufficient to kill immediately? We believe that heat sensitivity and the transduction mechanisms in- volved subserve a second function, namely to sense inflammation and attacks by naturally occurring irri- tants. This is achieved by intracellular mechanisms in the nociceptive terminal that lower the heat thresh- old rapidly and profoundly, so that body or tissue temperature can become a gating force of nociceptor excitation and pain. Of course, this is a hypothesis meant to stimulate experimental investigation, but, at first glance, it explains the immediate pain relief that results from cooling injured, inflamed or chemically irritated tissue.
From routine clinical practice, it is well known that pain associated with acute inflammatory states, such as appendicitis or dental pulpitis and with in- jury such as surgical wounds or distorsions, can efficiently be alleviated by local cooling. In the hu- man skin, pain induced by infiltration with a buffer solution of low pH or by topical capsaicin appli- cation was abolished by cooling of the skin (Steen and Reeh, 1993; Kilo et al., 1995). The frequency of action potentials recorded from the rat saphenous nerve in response to intradermahy injected capsaicin was rapidly reduced by ice cooling of the inner- vated skin area (Szolcsanyi, 1977). Also, in the rat saphenous nerve, excitation of C-mechano-heat-sen- sitive (C-MH) nociceptors by topical mustard oil was

40
.
II 5 8 c 1’1 * 8 II s 8 - ‘I 1 c (I”‘b ‘IS b’Im 8 7 b m I’# ‘I 0 10 20 30 40s
Fig. 1. A mechano-heat-sensitive C-fiber in the rabbit pleura- phrenic nerve preparation, in vitro. Radiant heat stimulation evoked a log-linear increase in discharge rate (each dot repre- senting one spike) in many C-fibers of this preparation, me- chanical (von Frey) thresholds were widely scattered between 1 and 64 mN. The regression line was calculated through seconds 17-25 during the active heating phase and extrapolated to cut the l/s-line; by that, a heat threshold (7’) could be estimated (Sauer. Weidner and Reeh, unpublished observation).
blocked by placing ice on the receptive fields (Reeh et al., 1986).
Room temperature as heat stimulus
Discovery of heat-evoked ionic currents and of the heat-activated capsaicin receptor (VRl) and its ho- mologue (VRL-1) in sensory neurons of the rat dorsal root ganglion (DRG) has shed important new light on the transduction mechanisms by which heat, inflammatory mediators and chemical irritants may excite nociceptors and contribute to pain (Cesare and McNaughton, 1996; Caterina et al., 1997, 1999). Substances that are able to lower the heat threshold in DRG cells or in VRl-transfected cells are puta- tive candidates, in the light of the above hypothesis, to exert their excitatory effect on nociceptors by a detour through heat transduction pathways.
Indeed, bradykinin (BK) and phorbol ester, which both can activate (see also Malmberg, 2000, this volume) protein kinase C (PKC; Burgess et al., 1989; Dray et al., 1992; McGuirk and Dolphin, 1992), were shown to sensitize DRG cells to heat, decreasing their heat thresholds from above 40 to be- low 30°C within seconds (Cesare and McNaughton, 1996). In these experiments BK by itself induced a small transient inward current, whereas phorbol ester did not, which was taken as an argument for a PKC- and, thus, sensitization-independent mech- anism of BK-induced excitation. However, the first description of BK-induced currents in DRG cells al- ready had established that staurosporine, a blocker of protein kinases, including PKC, prevents this BK effect (Burgess et al., 1989). Thus, the sensitizing and the excitatory mechanism seem to share the same key enzyme, suggesting that they may be iden- tical. Whether a sensitizing agent is strong enough to induce a depolarizing current by itself depends on whether the instantaneous heat threshold is lower or higher, respectively, than the ambient temperature, which is not well defined by ‘room temperature’ as a starting point in most patch-clamp studies. Differ- ences in ambient temperature may explain why BK regularly induced excitatory inward currents in rat DRG cells in some laboratories, but rarely in others (Burgess et al., 1989; Nicol and Cui, 1994; Kress et al., 1997; Vyklicky et al., 1998).
When action potentials were recorded in deep tis- sues (at body temperature), BK excited almost all nociceptive C-fibers, the prevalence of BK sensi- tivity being 92% in cat knee joint (Kanaka et al., 1985), 93% in dog testis (Kumazawa and Mizu- mura, 1980), 87% in dog muscle (Kumazawa and Mizumura, 1997), 83% in cat cardiovascular system (Baker et al., 1980), and 73% in cat gastrointesti- nal organs (Longhurst et al., 1984). In the skin of the limbs (at 32°C and similar temperatures), only about 40-50% of the C-MH fibers are excited by BK in the cat (Beck and Handwerker, 1974), rat (Lang et al., 1990) and monkey (Khan et al., 1992). In the skin, however, the threshold to noxious heat was able to drop from 45 to 30-31°C after sensiti- zation, by topical capsaicin in this case, as revealed in psychophysical experiments (Szolcsanyi, 1977). Thus, the differences in chemical responsiveness be- tween skin and deep tissues may result from their

41
,
5 10 15 20 25 30 min
Fig. 2. Stimulated release of calcitonin-gene related peptide (CGRP, measured by enzyme-immunoassay) from isolated stretches of desheathed rat sciatic nerve (n = 6-16); the noxious temperatures caused significant enhancements of the CGRP content in eluates which were entirely or to 90% (at 52°C) dependent on external calcium. The log-linear stimulus response curve (insert) of the peak release demonstrates a high thermal coefficient Qlo = 15 (Sauer, Bove and Reeh, unpublished observation).
higher temperature rather than from more sensitive nociceptors.
In a recent patch-clamp study of rat DRG cells, the ambient temperature was precisely maintained at 22-24°C and capsaicin and BK as well as pH 6.1 were found to lower the threshold of heat-activated currents to the respective bath temperature (Vyklicky et al., 1999). Actually, if heat stimulation was super- imposed on capsaicin superfusion at a low concen- tration, the heat-activated current developed directly out of a smaller chemically induced inward current without any apparent threshold, suggesting that both currents are finally gated by thermodynamic forces and passing through channels which are controlled by temperature in the first place (Fig. 3). In this view “capsaicin is not an agonist per se, but func- tions as a modulatory agent, lowering the channel’s response threshold to the ubiquitous actions of heat.” This concept has explicitly been proposed, based on a study demonstrating the polymodality of VRl expressed in HEK 293 cells or Xenupus oocytes (Tominaga et al., 1998). In this paper, low pH was introduced as a third activator of VR 1, after capsaicin and heat, and a mutual interaction between these al- gogenic stimuli was shown: the higher the proton concentration, the lower was the threshold to heat stimulation and vice versa; the higher the ambient temperature, the lower was the proton concentra-
tion needed to evoke a sustained excitatory inward current. This ‘communicating tube’ system only de- veloped in cells transfected with VRl, and heat thresholds close to room temperature were found at pH 6.3.
The cellular and molecular findings are sufficient reason to admit low pH, as in inflammatory tissue acidosis, to our hypothesis after having incorporated capsaicin and phorbol esters as exogenous irritants, and BK as an inflammatory mediator.
Temperature coefficients and protein stability
Heat-activated currents as well as nociceptor dis- charges present with very steep stimulus-response curves with Qlo (temperature coefficient over a 10°C temperature range) values well beyond 10 (see Fig. 1; Vyklicky et al., 1999). The same holds true for noxious heat-induced CGRP release from pe- ripheral axons (Sauer et al., 1999). These tempera- ture coefficients are far higher than those describing transport or enzymatic reactions and most voltage- gated ion currents (except for the deactivation kinet- ics of a special Cl- channel from Torpedo showing a Qlo of 40, Pusch et al., 1997). Vyklicky et al. (1999) recently translated the high Qlo values into ‘net free energies of stabilization’ that act to determine the conformation of a hypothetical thermosensitive
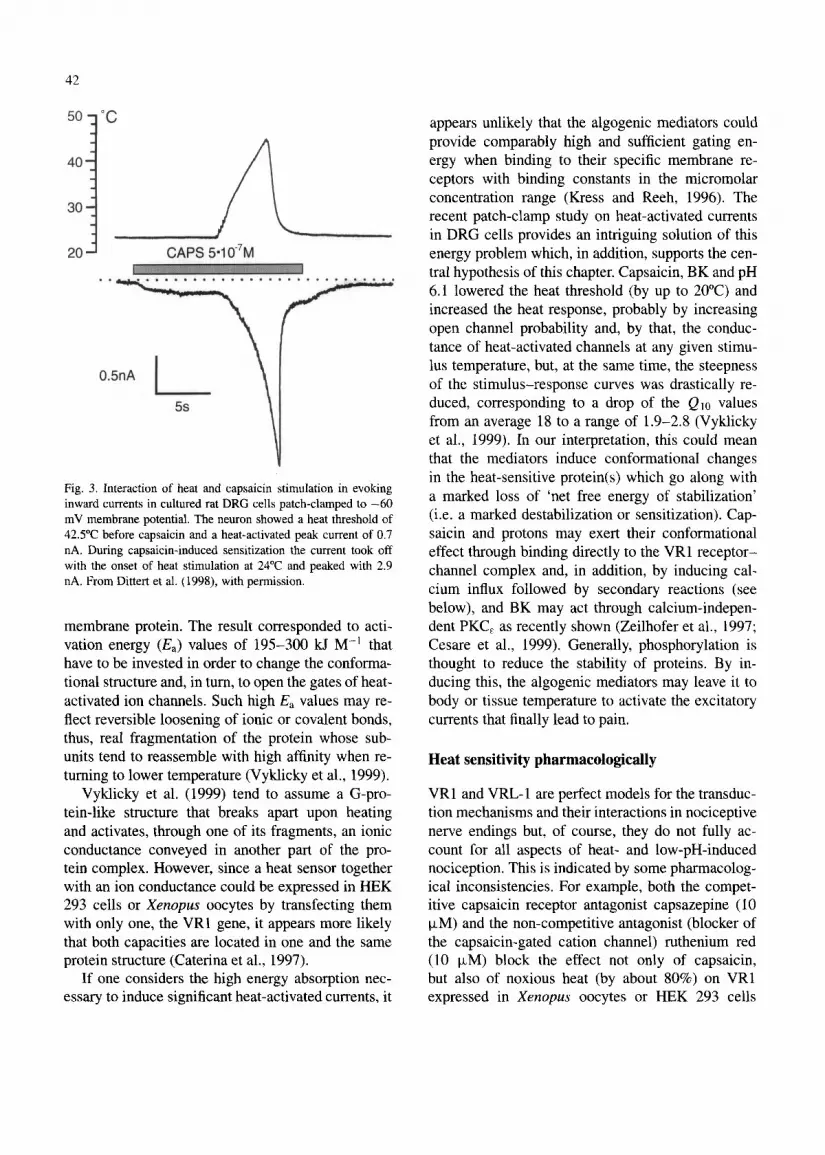
42
Fig. 3. Interaction of heat and capsaicin stimulation in evoking inward currents in cultured rat DRG cells patch-clamped to -60 mV membrane potential. The neuron showed a heat threshold of 42S’C before capsaicin and a heat-activated peak current of 0.7 nA. During capsaicin-induced sensitization the current took off with the onset of heat stimulation at 24°C and peaked with 2.9 nA. From Dittert et al. (1998), with permission.
membrane protein. The result corresponded to acti- vation energy (E,) values of 195-300 kJ M-’ that have to be invested in order to change the conforma- tional structure and, in turn, to open the gates of heat- activated ion channels. Such high E, values may re- flect reversible loosening of ionic or covalent bonds, thus, real fragmentation of the protein whose sub- units tend to reassemble with high affinity when re- turning to lower temperature (Vyklicky et al., 1999).
Vyklicky et al. (1999) tend to assume a G-pro- tein-like structure that breaks apart upon heating and activates, through one of its fragments, an ionic conductance conveyed in another part of the pro- tein complex. However, since a heat sensor together with an ion conductance could be expressed in HEK 293 cells or Xenopus oocytes by transfecting them with only one, the VRl gene, it appears more likely that both capacities are located in one and the same protein structure (Caterina et al., 1997).
If one considers the high energy absorption nec- essary to induce significant heat-activated currents, it
appears unlikely that the algogenic mediators could provide comparably high and sufficient gating en- ergy when binding to their specific membrane re- ceptors with binding constants in the micromolar concentration range (Kress and Reeh, 1996). The recent patch-clamp study on heat-activated currents in DRG cells provides an intriguing solution of this energy problem which, in addition, supports the cen- tral hypothesis of this chapter. Capsaicin, BK and pH 6.1 lowered the heat threshold (by up to 20°C) and increased the heat response, probably by increasing open channel probability and, by that, the conduc- tance of heat-activated channels at any given stimu- lus temperature, but, at the same time, the steepness of the stimulus-response curves was drastically re- duced, corresponding to a drop of the Qio values from an average 18 to a range of 1.9-2.8 (Vyklicky et al., 1999). In our interpretation, this could mean that the mediators induce conformational changes in the heat-sensitive protein(s) which go along with a marked loss of ‘net free energy of stabilization’ (i.e. a marked destabilization or sensitization). Cap- saicin and protons may exert their conformational effect through binding directly to the VRl receptor- channel complex and, in addition, by inducing cal- cium influx followed by secondary reactions (see below), and BK may act through calcium-indepen- dent PKC, as recently shown (Zeilhofer et al., 1997; Cesare et al., 1999). Generally, phosphorylation is thought to reduce the stability of proteins. By in- ducing this, the algogenic mediators may leave it to body or tissue temperature to activate the excitatory currents that finally lead to pain.
Heat sensitivity pharmacologically
VRl and VRL- 1 are perfect models for the transduc- tion mechanisms and their interactions in nociceptive nerve endings but, of course, they do not fully ac- count for all aspects of heat- and low-pH-induced nociception. This is indicated by some pharmacolog- ical inconsistencies. For example, both the compet- itive capsaicin receptor antagonist capsazepine (10 FM) and the non-competitive antagonist (blocker of the capsaicin-gated cation channel) ruthenium red (10 FM) block the effect not only of capsaicin, but also of noxious heat (by about 80%) on VRl expressed in Xenopus oocytes or HEK 293 cells
43
(Caterina et al., 1997; Tominaga et al., 1998). In the latter system, capsazepine also substantially inhibits the membrane current induced by low pH (Tominaga et al., 1998). On the other hand, only ruthenium red, and not capsazepine has a suppressive effect on the sustained pH-induced current in cultured DRG neu- rons (Zeilhofer et al., 1996; Vyklicky et al., 1998). In cat cornea1 nociceptors, excitation by carbon diox- ide (low pH) is not blocked by capsazepine (Chen et al., 1997). Also, both blockers are only partially effective in reducing (by about 30%) heat-activated currents in DRG cells (Kirschstein et al., 1999). A more extensive comparison of channel properties and pharmacological profiles has recently been published (Kress and Zeilhofer, 1999).
The pharmacological discrepancies regarding heat sensitivities have partly been resolved with the iden- tification and cloning of a capsaicin receptor homo- logue, VRL-1, which is expressed in rat DRG cells, shows a very high threshold (around 52°C) for nox- ious heat and is only blocked by ruthenium red and not by capsazepine (Caterina et al., 1999).
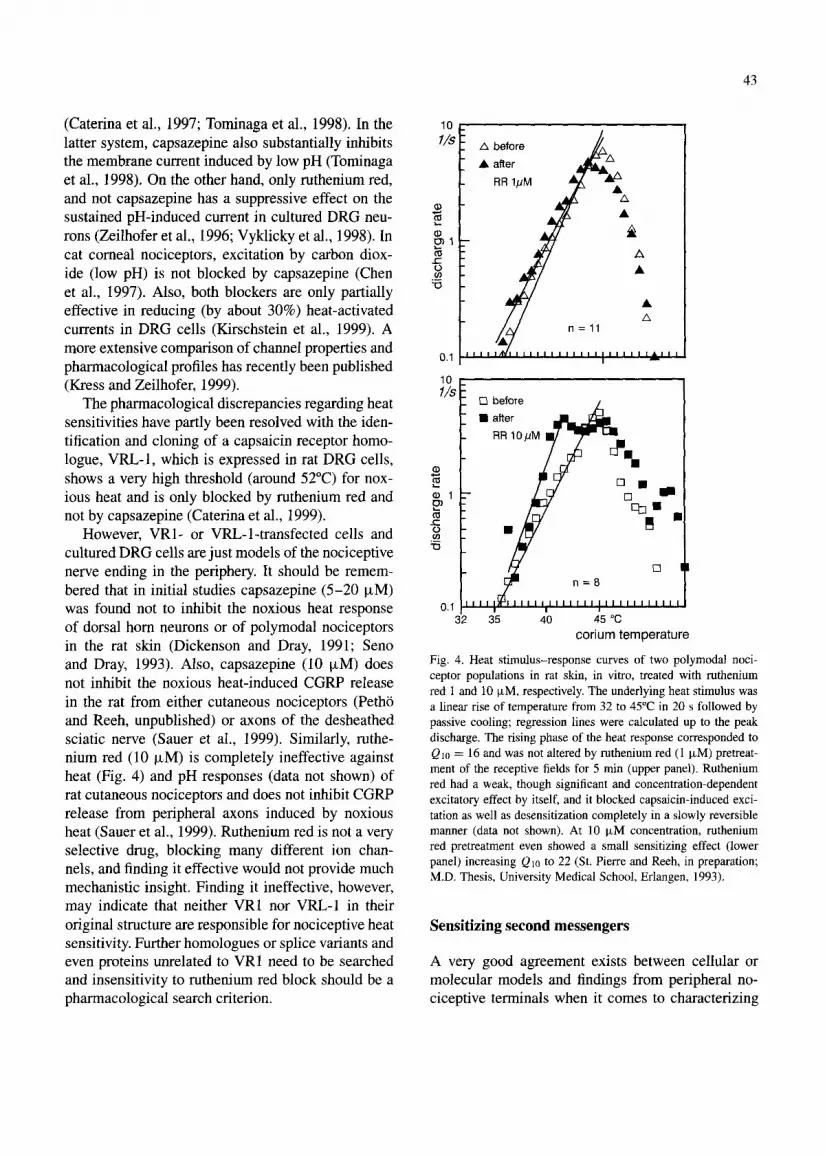
However, VRl- or VRL-1-transfected cells and cultured DRG cells are just models of the nociceptive nerve ending in the periphery. It should be remem- bered that in initial studies capsazepine (5-20 pM) was found not to inhibit the noxious heat response of dorsal horn neurons or of polymodal nociceptors in the rat skin (Dickenson and Dray, 1991; Seno and Dray, 1993). Also, capsazepine (10 pM) does not inhibit the noxious heat-induced CGRP release in the rat from either cutaneous nociceptors (Petho and Reeh, unpublished) or axons of the desheathed sciatic nerve (Sauer et al., 1999). Similarly, ruthe- nium red (10 FM) is completely ineffective against heat (Fig. 4) and pH responses (data not shown) of rat cutaneous nociceptors and does not inhibit CGRP release from peripheral axons induced by noxious heat (Sauer et al., 1999). Ruthenium red is not a very selective drug, blocking many different ion chan- nels, and finding it effective would not provide much mechanistic insight. Finding it ineffective, however, may indicate that neither VRI nor WI-1 in their original structure are responsible for nociceptive heat sensitivity. Further homologues or splice variants and even proteins unrelated to VRl need to be searched and insensitivity to ruthenium red block should be a pharmacological search criterion.
1
corium temperature
Fig. 4. Heat stimulus-response curves of two polymodal noci- ceptor populations in rat skin, in vitro, treated with ruthenium red 1 and 10 FM, respectively. The underlying heat stimulus was a linear rise of temperature from 32 to 45°C in 20 s followed by passive cooling; regression lines were calculated up to the peak discharge. The rising phase of the heat response corresponded to Qta = 16 and was not altered by ruthenium red (1 KM) pretreat- ment of the receptive fields for 5 min (upper panel). Ruthenium red had a weak, though significant and concentration-dependent excitatory effect by itself, and it blocked capsaicin-induced exci- tation as well as desensitization completely in a slowly reversible manner (data not shown). At 10 pM concentration, ruthenium red pretreatment even showed a small sensitizing effect (lower panel) increasing Qtc to 22 (St. Pierre and Reeh, in preparation; M.D. Thesis, University Medical School, Erlangen, 1993).
Sensitizing second messengers
A very good agreement exists between cellular or molecular models and findings from peripheral no- ciceptive terminals when it comes to characterizing

44
the essential role of calcium influx, which seems to be a sufficient, though not exclusive mechanism for induction of sensitization to heat. In the skin-nerve preparation, in vitro, loading nerve endings with sur- plus calcium-buffering capacity (using BAPTA-AM) consistently prevented the heat sensitization that fol- lowed application of either capsaicin, pH 6.1 or 5.4, ionomycin (a calcium ionophore) or intracellu- lar UV-photolysis of ‘caged calcium’ (NITR-S/AM; Gunther et al., 1999; Kress and Gunther, 1999).
In the same studies, three of the sensitizing stim- uli, capsaicin, low pH and ionomycin, were applied to patch-clamped DRG cells loaded with the calcium fluorochrome Fura-2, and a marked sensitization of heat-activated currents occurred which paralleled flu- orometric changes in calcium content with respect to both time course and magnitude (Gunther et al., 1999; Kress and Gunther, 1999). If calcium was removed from the extracellular solution around the cultured neurons, the capsaicin- and heat-activated currents were essentially unchanged, but calcium influx and sensitization to heat were abolished (Gun- ther et al., 1999). This is consistent with the results described above using BAPTA-AM and, together, the findings may provide further indication that VRl is not directly responsible for basic heat sensitivity or for sensitization of nociceptors to heat. Moreover, in capsaicin-induced heat sensitization, VRI seems to play the crucial role of a ligand-gated calcium channel that allows calcium to enter the nerve end- ings and exert the sensitizing effect through other heat-activated ion channels.
The BAPTA-AM loading method did not prevent BK-induced sensitization of primary afferents to heat, which lends further support to a calcium-independent, most likely PKC,-mediated mechanism (Cesare et al., 1999; Kessler et al., 1999). Activation of PKC may also be the mechanism by which histamine, through Hi receptors, can induce sensitization to heat of tes- ticular nociceptors (Koda et al., 1996).
With respect to the sensitizing role of intracel- lular calcium, it cannot yet be decided whether it acts through calcium-activated isoenzymes of PKC, through calcium-calmodulin kinases or through ac- tivating adenylyl cyclase isoenzymes, increasing cyclic AMP (CAMP) content and stimulating pro- tein kinase A (PKA) (see also Malmberg, 2000, this volume, for the role of protein kinase subtypes).
A contribution of the latter cascade is indicated by studies on cultured DRG neurons and by the fact that high concentrations of prostaglandin Ez (PGE2) can induce sensitization of testicular nociceptors to heat (Mizumura et al., 1993; Cui and Nicol, 1995). In the knee joint preparation, close-arterial injection of PGE:! can even induce weak excitatory effects (Schaible and Schmidt, 1988; Birrell and McQueen, 1993), a finding supported by effects of PGE2 in- jection into receptive fields of human itch receptors (C-fibers) recorded microneurographically (Schmelz et al., 1998). PGEz is thought to act by elevat- ing CAMP content, and stable membrane-permeant analogs of CAMP (but not cGMP) induce heat sen- sitization of cutaneous nociceptors (Kress et al., 1996). Thus, the intracellular pathways to sensitiza- tion seem to converge in protein phosphorylation by different protein kinases, acting on the putative heat- activated channels and counter-balanced by protein phosphatases whose cellular control mechanisms are largely unknown (Kress and Zeilhofer, 1999).
Alternative or complementary theories of inflam- matory sensitization do not aim at the process of sensory transduction, but at the conversion of gen- erator potentials into sequences of action potentials (see also Cummins et al., 2000, this volume, for the role of sodium channels). Patch-clamp work done on cultured sensory neurons offered two mod- ulatory mechanisms: shortening of the after-hyper- polarization of the action potentials, owing to BK receptor-mediated prostaglandin formation, and in- crease of tetrodotoxin-resistant voltage-gated Na+ currents due to simultaneous activation of PKC and PKA, for example by PGE2 (Weinrich et al., 1995; Gold et al., 1998). Both mechanisms could cooper- ate in increasing the gain of the conversion and, by that, the heat-induced discharge of nociceptive nerve fibers. However, neither mechanism can differentiate between heat- and mechanically evoked generator potentials and, thus, cannot account for the high se- lectivity of inflammatory sensitization to heat, and not to mechanical stimulation, in polymodal C-fibers (e.g. Kenins, 1984; Kocher et al., 1987; Kessler et al., 1992). Nonetheless, if as in other work PKA was activated in an assumed isolated manner by applica- tion of CAMP analogs, the result was sensitization to both heat and mechanical stimulation in cutaneous nociceptors (Kress et al., 1996).
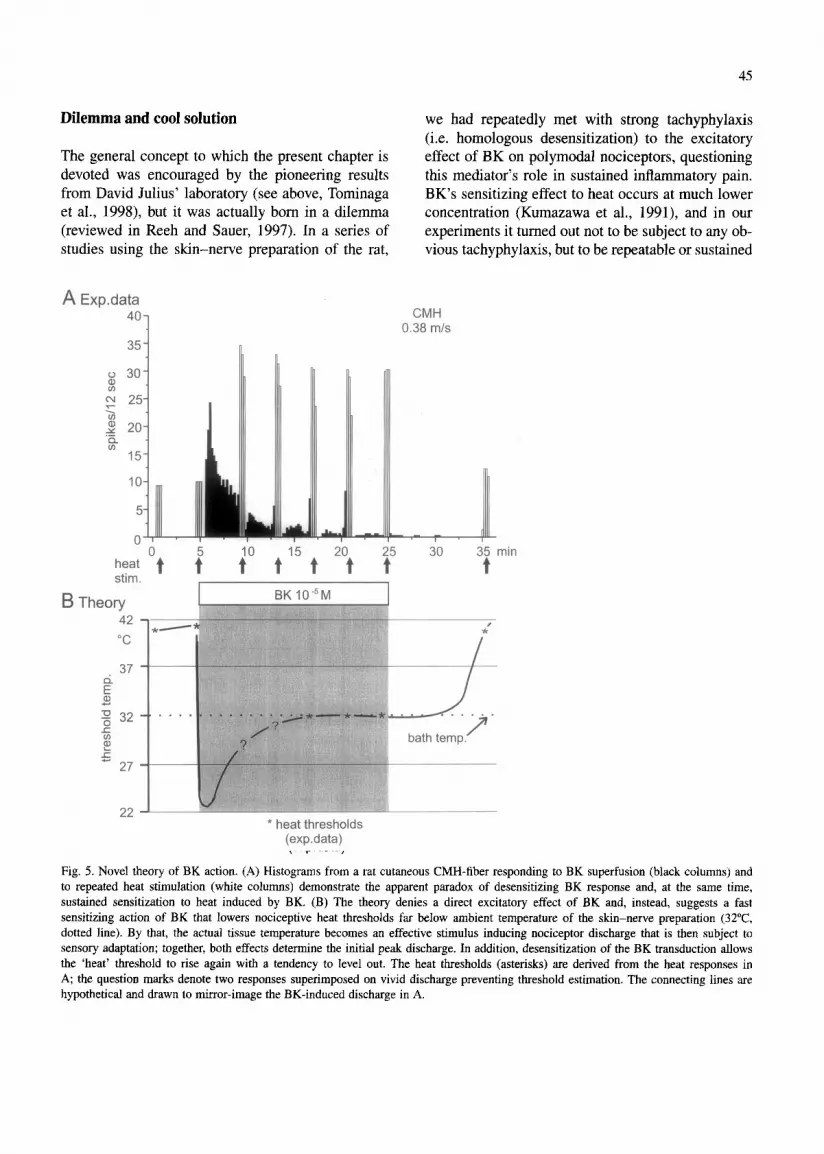
45
Dilemma and cool solution we had repeatedly met with strong tachyphylaxis (i.e. homologous desensitization) to the excitatory
The general concept to which the present chapter is effect of BK on polymodal nociceptors, questioning devoted was encouraged by the pioneering results this mediator’s role in sustained inflammatory pain. from David Julius’ laboratory (see above, Tominaga BK’s sensitizing effect to heat occurs at much lower et al., 1998), but it was actually born in a dilemma concentration (Kumazawa et al., 1991), and in our (reviewed in Reeh and Sauer, 1997). In a series of experiments it turned out not to be subject to any ob- studies using the skin-nerve preparation of the rat, vious tachyphylaxis, but to be repeatable or sustained
A Exp.data 40 CMH
0.38 m/s 35
0 30 8 RJ 25 c $ 2-g 20
In 15
10
5
0 35 min be., Ot t” ;” t I5 t ‘“t ‘t” 3o t
* heat thresholds (exp.data)
Fig. 5. Novel theory of BK action. (A) Histograms from a rat cutaneous CMH-fiber responding to BK superfusion (black columns) and to repeated heat stimulation (white columns) demonstrate the apparent paradox of desensitizing BK response and, at the same time, sustained sensitization to heat induced by BK. (B) The theory denies a direct excitatory effect of BK and, instead, suggests a fast sensitizing action of BK that lowers nociceptive heat thresholds far below ambient temperature of the skin-nerve preparation (32”C, dotted line). By that, the actual tissue temperature becomes an effective stimulus inducing nociceptor discharge that is then subject to sensory adaptation; together, both effects determine the initial peak discharge. In addition, desensitization of the BK transduction allows the ‘heat’ threshold to rise again with a tendency to level out. The heat thresholds (asterisks) are derived from the heat responses in A; the question marks denote two responses superimposed on vivid discharge preventing threshold estimation. The connecting lines are hypothetical and drawn to mirror-image the BK-induced discharge in A.

46
for as long as BK was present (Fig. 5A). This was in accordance with previous psychophysical work indi- cating that the thermal hyperalgesic effects of BK are much less prone to tachyphylaxis and more sustained than the algesic effects (Manning et al., 1991). In our experiments, the sensitizing effect of BK extended to a much larger subpopulation of polymodal affer- ents than the excitatory effect, and it even recruited a proportion of high-threshold C-mechanoreceptors (C-HTM) that reversibly became heat-sensitive af- ter BK superfusion. The sensitization to heat was much more pronounced in those units that were ex- cited by BK than in the ones not excited. The latter result is in accordance with a previous study that showed a strong correlation between the magnitude of BK-induced excitation and the degree of sensiti- zation to heat in testicular nociceptors (Kumazawa et al., 1991). Both findings are consistent with our theory that excitation by BK is the extreme of sensi- tization.
In a further study using subtype-selective BK re- ceptor antagonists, half of the C-MH and C-HTM fibers not excited by BK were sensitized to heat, and in half of these cases the sensitization could not be prevented by HOE140, the B2 receptor antagonist; the effects of BK were blocked by des-Arg9-[Leu*]-BK, the Bi receptor antagonist. The Bi receptor does not exhibit homologous desensi- tization, which would fit the lack of tachyphylaxis of the sensitizing BK effect. However, in the vast majority of fibers, the B2 receptor was found respon- sible for both sensitization and excitation, if present. Here the dilemma arose. If tachyphylaxis to BK is due to desensitization of the established transduction pathway (through PKC) and includes internalization of the B2 receptor (Pizard et al., 1999), how could such a mechanism possibly spare the BK-induced nociceptor sensitization to heat?
The only way out was to assume that the real magnitude of the sensitizing effect was much larger, at first, and declined to what we could detect when the excitatory effect of BK was fading (Fig. 5B). The peak of the sensitizing effect and its real time course may be hidden behind the apparent excitatory phase during superfusion with BK. To uncover this period, we had to get rid of the excitatory effect, remembering that moderate cooling had previously been shown to abolish ongoing nociceptor discharge
l
0 5 10 15 20 25 30
BK dM 45-j *--
p 40
0 35
li\
o-e-0
g30 '
/ . . /+. l ,* - - * - . . . . . . .
I. kf 25 25
20 , I , I , I , I , , 1 , 0 5 10 15 20 25 30
BK 1CfM
*2oi I,
0 5 10 15 20 25 30
time (min)
Fig. 6. Effect of BK on heat thresholds of four different C-poly- modal nociceptors in the cooled rat skin, in vitro. The skin- nerve preparation was cooled to 18-20°C before BK superfusion to prevent the apparent excitatory effect; the dotted lines repre- sent the normal bath temperature of 32°C. Radiant heat stimuli were ramp-shaped (19-4S’C, 40 s) but discontinued shortly after exceeding threshold in order to allow for frequent determinations (Petho and Reeh, unpublished observation).
in inflamed rat skin as well as pain in humans after cutaneous capsaicin application or acid infusion (Szolcsanyi, 1977; Reeh et al., 1986; Steen and Reeh, 1993; Kilo et al., 1995).
Fig. 6 shows the outcome of this consideration. In this ongoing study, the isolated skin-nerve prepa- ration is cooled to 18-20°C before BK or other

47
algogens are applied, and this has so far prevented all BK-, capsaicin- and most of the pH-induced dis- charge. Then, ramp-shaped heat stimuli are applied at short intervals and discontinued as soon as a few spikes are recorded to permit response threshold determination (temperature at second spike). The fig- ure gives examples from four individual polymodal C-fibers and shows the fast and dramatic sensitizing effect of a moderate concentration of BK. In several fibers, response threshold dropped by 20°C and more within less than 1 min; the first threshold determined during BK was always the lowest. It was followed by a partial increase in threshold (recovery toward the pre-BK threshold), thus revealing the previously missing decline of the sensitizing effect (‘desensiti- zation’) of BK and attaining a sustained level after a while. Rapid reversibility of the sensitization within a few minutes during wash-out of BK had previously been demonstrated and was not the subject of the present study (Koltzenburg et al., 1992).
If one considers (Fig. 6) the temporal profile of the thresholds staying below the normal temperature of the organ bath, it is easy to imagine that this nadir actually reflects the apparent excitatory effect of BK which normally occurs at higher ambient tem- peratures. Repeated short applications of BK would presumably induce less and less of the peak sensitiz- ing effect due to desensitization of BK transduction and, by that, the mirror image of sensitization, the apparent excitatory effect, would present with the well-known tachyphylaxis.
We speculate that the early and fast peak sensi- tization of nociceptors is mediated mainly by PKC because the sensitization of the heat response of DRG neurons by BK is equally fast, mimicked by the phorbol ester PMA and reversed by the PKC inhibitor staurosporine (Cesare and McNaughton, 1996). In a subsequent study, BK-induced translo- cation of PKC, into the plasma membrane of DRG neurons was clearly shown (Cesare et al., 1999). The plateau of sensitization indicated in Fig. 6 and, in particular, the short-lasting after-effect following wash-out of BK (which was the subject of the previ- ous investigations into BK-induced nociceptor sen- sitization) may progressively depend on secondary prostaglandin production. This is supported by an ongoing study showing that the active S(+) but not the inactive R(-) enantiomer of the cyclooxygenase
inhibitor flurbiprofen abolished heat sensitization in- duced by sustained (5 min) BK exposure, and this effect could partially be overcome by exogenously applied PGE2 (Petho et al., 1999). The proposed role of both PKC and prostaglandins in BK-induced heat sensitization is in accordance with the fact that BK, via B2 as well as B t receptors, can couple not only to phospholipase C/PKC (see above), but also to phos- pholipase AZ (Pizard et al., 1999), which initiates prostaglandin formation in the nerve ending itself or in the surrounding tissue. Supporting this view, BK-induced release of PGE2 from isolated rat skin has recently been measured (using an ELISA) and found sustained (over 15 min) at a high level after an initial phase of desensitization (Petho, Izydorczyk and Reeh, unpublished observation).
During the BK experiments with the cooled skin- nerve preparation, we occasionally applied complete heat stimuli, linearly increasing the temperature from 19 to 45°C in 40 s, to determine the pattern of re- sponse of sensitized fibers to heat. The result was the usual log-linear increase in the discharge rate, with a considerable number of spikes below the normal organ bath temperature of 32°C. The slopes of the stimulus-response curves were abnormally flat with Qia values around 3, just as predicted by Vyklicky et al. (1999) based on their work on heat-activated currents in DRG cells using capsaicin, BK and low pH as sensitizing agents at controlled room tem- perature (see above). In ongoing studies, with the cooled skin-nerve preparation, capsaicin had a sim- ilar sensitizing effect as BK, but, as expected, with a much stronger decline and desensitization after an initial peak effect. Low pH effects on heat thresholds were slow to develop, less profound, but very well sustained, again as expected from the sustained exci- tatory effect of this stimulus on nociceptors (Steen et al., 1992).
Conclusions
There is growing evidence that several inflammatory mediators, including BK (possibly PGE2 and his- tamine) and low pH as well as the sensitizing model agent capsaicin, act on nociceptors, at least partly, by lowering the threshold of their heat transduc- tion mechanisms so profoundly that body or lower tissue temperatures become a driving force of ex-

48
citation and pain. This unifying theory is attractive for pharmaceutical research and development be- cause diverse and multiple nociceptive mechanisms are converging onto one novel target, heat-activated ion channels, which, however, await final molecular identification.
Acknowledgements
The authors’ work was supported by Deutsche Forschungsgemeinschaft (SFB 353) European Union and Sander-Stiftung. G.P.‘s fellowship was granted by the Humboldt-Stiftung. Alexandra Derow made valuable contributions to the most recent data and Iwona Izydorczyk provided expert technical help.
References
Baker, D.G., Coleridge, H.M., Coleridge, J.G.G. and Nerdrum, T. (1980) Search for a cardiac nociceptor: stimulation by bradykinin of sympathetic afferent nerve endings in the heart of the cat. .I. Physiol., 306: 519-536.
Beck, PW. and Handwerker, H.O. (1974) Bradykinin and sero- tonin effects on various types of cutaneous nerve fibers. Pjfiigers Arch. EM J. Physiol., 314: 209-222.
Birrell, G.J. and McQueen, D.S. (1993) The effects of capsaicin, bradykinin, PGEz and cicaprost on the discharge of articular sensory receptors in vitro. Brain Res., 611: 103-107.
Bove, G.M. and Moskowitz, M.A. (1997) Primary afferent neu- rons innervating guinea pig dura. J. Neurophysiol., 77: 299- 308.
Burgess, G.M., Mullaney, I., McNeil, M., Dunn, P.M. and Rang, H.P. (1989) Second messengers involved in the mechanism of action of bradykinin in sensory neurons in culture. J. Neurosci., 9: 3314-3325.
Caterina, J.M., Schumacher, M.A., Tominaga, M., Rosen, T.A., Levine, J.D. and Julius, D. (1997) The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 389: 816-824.
Caterina, M.J., Rosen, T.A., Tominaga, M., Brake, A.J. and Julius, D. (1999) A caps&in-receptor homologue with a high threshold for noxious heat. Nature, 398: 436-441.
Chen, X., Belmonte, C. and Rang, H.P. (1997) Capsaicin and carbon dioxide act by distinct mechanisms on sensory nerve terminals in the cat cornea. Pain, 70: 23-29.
Cesare, P, Dekker, L.V., Sardini, A., Parker, P.J. and Mc- Naughton, P.A. (1999) Specific involvement of PKC, in sensi- tization of the neuronal response to painful heat. Neuron, 23: 611-624.
Cesare, P. and McNaughton, P. (1996) A novel heat-acti- vated current in nociceptive neurons and its sensitization by bradykinin. Proc. Natl. Acad. Sci. USA, 93: 15435-15439.
Cui, M. and Nicol, G.D. (1995) Cyclic AMP mediates the prostaglandin Ez-induced potentiation of bradykinin excitation in rat sensory neurons. Neuroscience, 2: 459-466.
Cummins, T.R., Dib-Haji, S.D., Black, J.A. and Waxman, S.G. (2000) Sodium channels and the molecular pathophysiology of pain. In: J. Sandktihler, B. Bromm and G.F. Gebhart (Eds.), Nervous System Plasticity and Chronic Pain, Progress in Brain Research, Vol. 129, Elsevier Science, Amsterdam, pp. 3-19.
Dickenson, A.H. and Dray, A. (1991) Selective antagonism of capsaicin by capsazepine: evidence for a spinal receptor site in capsaicin-induced antinociception. BE J. Pharmacol., 104: 1045-1049.
Dittert, I., Vlachova, V., Knotkova, H., Vitaskova, Z., Vykhcky, L., Kress, M. and Reeh, P.W. (1998) A technique for fast application of heated solutions of different composition to cultured neurons. J. Neurosci. Methods, 82: 195-201.
Dray, A., Patel, LA., Perkins, M.N. and Rueff, A. (1992) Bradykinin-induced activation of nociceptors: receptor and mechanistic studies on the neonatal rat spinal cord-tail prepa- ration in vitro. BI: J. Pharmacol., 107: 1129-l 134.
Gold, MS., Levine, J.D. and Correa, A.M. (1998) Modulation of TTX-R/N, by PKC and PKA and their role in PGEz-induced sensitization of rat sensory neurons in vitro. J. Neurosci., 18: 10345-10355.
Gunther, S., Reeh, P.W. and Kress, M. (1999) Rises in [Ca’+li mediate capsaicin- and proton-induced heat sensitization of rat primary nociceptive neurons. Eur: J. Neumsci., 1 I: 3143- 3150.
Kanaka, R., Schaible, H.G. and Schmidt, R.F. (1985) Activation of fine articular afferent units by bradykinin. Brain Res., 327: 81-90.
Kenins, P. (1982) Responses of single nerve fibres to capsaicin applied to the skin. Neurosci. Lett., 29: 83-88.
Kessler, W., Kirchhoff, C., Reeh, P.W. and Handwerker, H.O. (1992) Excitation of cutaneous afferent nerve endings in vitro by a combination of inflammatory mediators and conditioning effect of substance P. Exp. Bruin Res., 91: 467-476.
Kessler, F., Habelt, C., Averbeck, B., Reeh, P.W. and Kress, M. (1999) Heat-induced release of CGRP from isolated rat skin and effects of bradykinin and the protein kinase C activator PMA. Pain, 83: 289-295.
Khan, A.A., Raja, S.N., Manning, D.C., Campbell, J.N. and Meyer, R.A. (1992) The effects of bradykinin and sequence- related analogs on the response properties of cutaneous noci- ceptors in monkeys. Somatosens. Mot. Res., 9: 97-106.
Kilo, S., Forster, C., Geisslinger, G., Brune, K. and Handwerker, H.O. (1995) Inflammatory models of cutaneous hyperalgesia are sensitive to effects of ibuprofen in man. Pain, 62: 187- 193.
Kirschstein, T., Greffrath, W., Bttsselberg, D. and Treede, R.D. (1999) Inhibition of rapid heat responses in nociceptive pri- mary sensory neurons of rats by vanilloid receptor antagonists. J. Neurophysiol., 82: 2853-2860.
Kocher, L., Anton, F., Reeh, PW. and Handwerker, H.O. (1987) The effect of carrageenan-induced inflammation on the sensi- tivity of unmyelinated skin nociceptors in the rat. Pain, 29: 363-373.

49
Koda, H., Minagawa, M., S&Hong, L., Mizumura, K. and Ku- mazawa, T. (1996) Hl receptor mediated excitation and facil- itation of the heat response by histamine in canine visceral polymodal receptors studied in vitro. J. Neurophysiol., 76: 1396-1404.
Koltzenburg, M., Kress, M. and Reeh, P.W. (1992) The nocicep- tor sensitization by bradykinin does not depend on sympathetic neurones. Neuroscience, 46: 465-413.
Kress, M. and Gunther, S. (1999) Role of [Ca’+]i in the ATP- induced sensitization process of rat nociceptive neurons. J. Neurophysiol., 81: 2612-2619.
Kress, M. and Reeh, P.W. (1996) Transduction mechanisms in nociceptors - chemical excitation and sensitization in nocicep- tors. In: F. Cervero and C. Belmonte (Eds.), Neurobiology of Nociceptors, Oxford University Press, Oxford, pp. 258-297.
Kress, M. and Zeilhofer, H.U. (1999) Capsaicin, protons and heat: new excitement about nociceptors. Trends Pharmacol. Sci., 20: 112-118.
Kress, M., Rod], J. and Reeh, PW. (1996) Stable analogs of cyclic AMP but not cyclic GMP sensitize unmyelinated pri- mary afferents in rat skin to mechanical and heat stimulation but not to inflammatory mediators, in vitro. Neuroscience, 74: 609-617.
Kress, M., Reeh, PW. and Vyklicky, L. (1997) An interaction of inflammatory mediators and protons in small diameter dorsal root ganglion neurons. Neutosci. Lett., 224: 37-40.
Kumazawa, T. and Mizumura, K. (1980) Chemical responses of polymodal receptors of the scrotal contents in dogs. J. Physiol., 299: 219-232.
Kumazawa, T. and Mizumura, K. (1997) Thin-fibre receptors responding to mechanical, chemical, and thermal stimulation in the skeletal muscle of the dog. J. Physiol., 273: 179-194.
Kumazawa, T., Mizumura, K., Minagawa, M. and Tsujii, Y. (1991) Sensitizing effects of bradykinin on the heat responses of the visceral nociceptor. J Neurophysiol., 66: 18 19-1824.
Lang, E., Novak, P., Reeh, P.W. and Handwerker, H.O. (1990) Chemosensitivity of fine afferents from rat skin in vitro. J. Neurophysiol., 63: 887-901.
Longhurst, J.C., Kaufman, D.P., Ordway, G.A. and Mush, T.I. (1984) Effects of bradykinin and capsaicin on endings of afferent fibres from abdominal visceral organs. Am. J. Physiol. Regul. Integr Comp. Physiol., 247: R552-R559.
Malmberg, A. (2000) Protein kinase subtypes involved in in- jury-induced nociception. In: J. Sandkiihler, B. Bromm and G.F. Gebhart (Eds.), Nervous System Plasticity and Chronic Pain, Progress in Brain Research, Vol. 129. Elsevier Science, Amsterdam, pp. 51-59.
Manning, D.C., Raja, S.N., Meyer, R.A. and Campbell, J.N. (1991) Pain and hyperalgesia after intradermal injection of bradykinin in humans. Clin. Pharmacol. Ther, 50: 721-729.
MC&irk, S.M. and Dolphin, A.C. (1992) G-protein mediation in nociceptive signal transduction: an investigation into the excitatory action of bradykinin in a subpopulation of cultured rat sensory neurons. Neuroscience, 49: 117-128.
Mizumura, K., Minagawa, M., Tsujii, Y. and Kumazawa, T. (1993) Prostaglandin Ez-induced sensitization of the heat re-
sponse of canine visceral polymodal receptors in vitro. Neu- rosci. Len., 161: 117-119.
Nicol, G.D. and Cui, M. (1994) Enhancement by prostaglandin Ez of bradykinin activation of embryonic rat sensory neurones. J Physiol., 480: 485-492.
Petho, G., Derow, A. and Reeh, P.W. (1999) Cyclooxygenase products are involved in the bradykinin-induced sensitization to heat of rat cutaneous nociceptors, in vitro. Pfliigers Arch. Eur J. Physiol., 437: R134.
Pizard, A., Blaukat, A., Muller-Esterl, W., Alhenc-Gelas, F. and Rajerison, R.M. (1999) Bradykinin-induced internalization of the human B2 receptor requires phosphorylation of three serine and two threonine residues at its carboxyl tail. J. Biol. Chem., 18: 12738-12747.
Pusch, M., Ludewig, U. and Jentsch, T.J. (1997) Temperature dependence of fast and slow gating relaxations of ClC-0 chloride channels. J. Gen. Physiol., 109: 105-l 16.
Reeh, P.W. and Sauer, S. (1997) Chronic aspects in peripheral nociception. In: Jensen et al. (Eds.), Proceedings of the 8th World Congress on Pain, IASP Press, Seattle, pp. 115-13 1.
Reeh, P.W., Kocher, L. and Jung, S. (1986) Does neurogenic inflammation alter the sensitivity of unmyelinated nociceptors in the rat?. Brain Rex, 384: 42-50.
Sauer, SK., Bove, G.hl. and Reeh, PW. (1999) Heat-induced CGRP release from rat sciatic nerve: partial block by capsaicin antagonists. Pfliigers Arch. Eur J. Physiol., 437: R54.
Schaible, H.G. and Schmidt, R.F. (1988) Excitation and sen- sitization of fine articular afferents from cat’s knee joint by prostaglandin El, J. Physiol., 403: 9 I- 104.
Schmelz, M., Schmidt, R., Weidner, C., Torebjork, H.E. and Handwerker, H.O. (1998) Chemical responsiveness of mechanosensitive and -insensitive C nociceptors in human skin. Sot. Neurosci. Abstr, 24: 383.
Seno, N. and Dray, A. (1993) Capsaicin-induced activation of fine afferent fibres from rat skin in vitro. Neuroscience, 55: 563-569.
Steen, K.H. and Reeh, P.W. (1993) Sustained graded pain and hyperalgesia from harmless experimental tissue acidosis in human skin. Neutosci. Lett., 154: 113-l 16.
Steen, K.H., Reeh, PW., Anton, F. and Handwerker, H.O. (1992) Protons selectively induce lasting excitation and sensitization to mechanical stimulation of nociceptors in rat skin, in vitro. J. Neurosci., 12: 86-95.
Su, X. and Gebhart, G.F. (1998) Mechanoseusitive pelvic nerve afferent fibers innervating the colon of the rat are polymodal in character. J. Neurophysiol., 80: 2632-2644.
Szolcsanyi, J. (1977) A pharmacological approach to elucidation of the role of different nerve fibres and receptor endings in mediation of pain. J. Physiol. (Paris), 73: 251-259.
Tominaga, M., Caterina, M.J., Malmberg, A.B., Rosen, T.A., Gilbert, H., Skinner, K., Raumann, B.E., Basbaum, A.I. and Julius, D. (1998) The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron, 2 1: 53 l-543.
Vyklicky, L., Knotkova-Urbancova, H., Vitaskova, Z., Vlachova, V., Kress, M. and Reeh, P.W. (1998) Inflammatory mediators at acidic pH activate capsaicin receptors in cultured sensory neurons from newborn rat. J. Neurophysiol., 79: 670-676.

SO
Vyklicky, L., Vlachova, V., Vitaskova, Z., Dittert, I., Kabat, M. Zeilhofer, H.U., Reeh, P.W., Swandulla, D. and Kress, M. (1996) and Orkand, R.K. (1999) Temperature coefficient of membrane Ca2+ permeability of the sustained proton-induced cation cur- currents induced by noxious heat in sensory neurones in the rent in adult rat dorsal root ganglion neurons. J. Neurophysiol., rat. .I. Physiol., 517: 181-192. 76: 2834-2840.
Weinrich, D., Koschorke, GM., Undem, B.J. and Taylor, G.E. Zeilhofer, H.U., Kress, M. and Swandulla, D. (1997) Fractional (1995) Prevention of the excitatory actions of bradykinin by Ca2+ currents through capsaicin- and proton-activated ion inhibition of PC& formation in nodose neurones of the guinea- channels in rat dorsal root ganglion neurones. J. Physiol., 503: pig. J. Physiol., 483: 135-746. 67-78.