cocaine sensitization models an anhedonia-like condition in rats
TRANSCRIPT

Cocaine sensitization models an anhedonia-likecondition in rats
Simona Scheggi, Giovanna Marchese, Silvia Grappi, Maria Elena Secci,Maria Graziella De Montis and Carla Gambarana
Department of Neuroscience, Pharmacology Unit, University of Siena, Siena, Italy
Abstract
Anhedonia is a core symptom of depression that also characterizes substance abuse-related mood
disorders, in particular those secondary to stimulant abuse. This study investigated the long-lasting
condition of cocaine sensitization as an inducing condition for anhedonia in rats. Cortical-mesolimbic
dopamine plays a central role in assessing the incentive value of a stimulus and an increased dopamine
output in these areas after a novel palatable meal seems to correlate with the ability to acquire an instru-
mental behaviour aimed at earning it again. This dopaminergic response is associated with consistent
modifications in the phosphorylation pattern of some cAMP-dependent protein kinase (PKA) substrates
and it is mediated by dopamine D1 receptor stimulation. Thus, since behavioural cocaine sensitization is
characterized by tonically increased levels of phospho-Thr75 DARPP-32 that is a potent PKA inhibitor, we
hypothesized that cocaine-sensitized rats might reveal deficits in palatable food responding. Indeed, non-
food-deprived cocaine-sensitized rats showed no interest in palatable food, no dopaminergic response
after a palatable meal in terms of increased dopamine output and DARPP-32 phosphorylation changes,
and no ability to acquire a palatable food-sustained instrumental behaviour. Repeated administration of
an established antidepressant compound, imipramine, corrected these deficits and reinstated the dopa-
minergic response in the cortico-mesolimbic areas to control values. Thus, the behavioural modifications
observed in cocaine-sensitized rats satisfy some requirements for an experimental model of anhedonia
since they are induced by repeated cocaine administration (aetiological validity), they mimic an
anhedonia-like symptom (construct validity), and are reversed by the administration of imipramine
(predictive validity).
Received 7 December 2009 ; Reviewed 1 January 2010 ; Revised 12 April 2010 ; Accepted 20 April 2010 ;
First published online 2 June 2010
Key words : DARPP-32, dopamine, imipramine, instrumental learning, microdialysis.
Introduction
Non food-deprived rats can be attracted by and con-sume small amounts of a palatable food and, if theavailability of this food is made contingent on theemission of a specific action, they show the ability toacquire an appetitive behaviour instrumental inearning that food. This ability is probably dependenton the emotional value of the palatable food, anda correlation has been observed between the dopa-minergic response, in terms of increased dopamineoutput measured by microdialysis in the medial
prefrontal cortex (mPFC) and in the shell portion ofthe nucleus accumbens (NAcS), after consuming thatfood for the first time and the ability to acquire theappetitive behaviour (Gambarana et al. 2003). Thus,chronically stressed rats that show no increase indopamine output in the mesolimbic areas after con-suming the food do not learn how to earn it(Gambarana et al. 2003). The palatable food-eliciteddopaminergic response is associated with consist-ent modifications in the phosphorylation pattern ofsome cAMP-dependent protein kinase (PKA) sub-strates (Rauggi et al. 2005b), in particular DARPP-32.Dopamine, through D1 receptor-mediated activationof PKA, phosphorylates DARPP-32 at threonine-34(Thr34) and thereby converts DARPP-32 into a potentinhibitor of protein phosphatase-1 (Hemmings et al.1984), while phosphorylation at Thr75 converts
Address for correspondence : Dr C. Gambarana, Department of
Neuroscience, Pharmacology Unit, University of Siena, Via Moro,
4, 53100 – Siena, Italy.
Tel. : +39 0577234116 Fax : +39 0577234208
Email : [email protected]
International Journal of Neuropsychopharmacology (2011), 14, 333–346. f CINP 2010doi:10.1017/S1461145710000532
ARTICLE

DARPP-32 into an inhibitor of PKA (Bibb et al. 1999).Thus, DARPP-32 can act either as a phosphataseinhibitor or as a kinase inhibitor, depending on itsphosphorylation state. The increased dopamine out-put after a palatable meal is accompanied by an earlyincrease in phospho-Thr34 DARPP-32, phospho-GluR1
and phospho-NR1 levels, paralleled by a decreasein phospho-Thr75 DARPP-32 levels, followed by op-posite modifications 2–3 h after the meal ; SCH 23390,a selective dopamine D1 receptor antagonist, admin-istered shortly after the meal, prevents the occurrenceof the entire sequence (Danielli et al. 2010 ; Rauggiet al. 2005b).
Cocaine-sensitized (CS) rats show a tonic increase inphospho-Thr75 DARPP-32 and a decrease in phospho-Thr34 DARPP-32, phospho-GluR1 and phospho-NR1
levels in the mPFC and NAc (Scheggi et al. 2007).Cocaine sensitization and Thr75 DARPP-32 phos-phorylation are strictly correlated, since no cocainebehavioural sensitization develops in mice carrying apoint mutation with a non-phosphorylatable residue(alanine) at the 75 position of DARPP-32 (Zachariouet al. 2006). Moreover, in sensitized rats the increase indopamine output induced by acute cocaine adminis-tration is not associated with a dopamine D1 receptor-mediated increase in phospho-Thr34 DARPP-32levels (Scheggi et al. 2007). In other words, CS ratscan be considered a model of tonically reduceddopamine D1 receptor-mediated transmission incortical-mesolimbic areas. Thus, since dopamine D1
receptor-mediated transmission seems to play a rel-evant role in response to pleasurable stimuli, wehypothesized that in non-food-deprived CS rats thedopaminergic response to palatable food may bealtered and that this may translate into an anhedonia-like condition. To test this hypothesis, we evaluatedthe response of non-food-deprived CS rats to a palat-able food, vanilla sugar (VS), in terms of hedonic tastereactivity, dopamine output (measured by micro-dialysis) and dopamine D1 receptor-dependent sig-nalling in the NAcS. Moreover, the ability of CS ratsto acquire an instrumental behaviour – based on thereinforcing properties of VS pellets, the earning ofwhich is made contingent on the choice of one ofthe two divergent arms of a Y-maze (VS sustainedappetitive behaviour, VAB) (Ghiglieri et al. 1997) –was investigated. Since sensitized rats failed toshow behavioural and dopaminergic responses toVS consumption and to acquire VAB, we investi-gated whether repeated administration of imipramine(Imi), a classical antidepressant compound, wouldrestore their response to VS and VAB learning com-petence.
Materials and methods
Animals
Experiments were performed on male Sprague–Dawley rats (Charles River, Italy) that were allowedat least 1 wk habituation at the animal colony andweighed 200–225 g when the experimental proceduresbegan. Animals were housed (n=5 per cage) in anenvironment maintained at a constant temperatureand humidity with free access to food and water. A12-h reversed light/dark cycle (lights on 19:00 hours)was used. Experiments were performed from 09:00to 17 :00 hours under a red light and controlled noiseconditions in a testing room separate from and ad-jacent to the main animal room. The procedures usedwere in accordance with the European legislationon the use and care of laboratory animals (EECCouncil Directive 86/609) and were approved by theUniversity of Siena Ethics Committee.
Induction of cocaine sensitization
Sensitization was induced by administering cocaine(10 mg/kg.d i.p.) for 8 d as previously described(Scheggi et al. 2007). Rats were individually placed inlocomotor activity cages for 35 min after cocaine (orsaline) administration to strengthen environmentalcontingencies and to favour the development of sensi-tization. After a 10-d washout, rats were tested fortheir response to a cocaine challenge (5 mg/kg i.p.) :locomotor activity and stereotypies were recordedfor 30 min (the first 5 min were not taken into account).Locomotor activity was determined in motilitycages that detected horizontal activity and rearings(Imetronic, France). Stereotypies were scored byexperimenters blind to the experimental conditionsaccording to a published rating scale (Gambaranaet al. 1998 ; Scheggi et al. 2007). A cumulative scorereflecting the intensity of stereotypies during the30-min observation period was assigned to each rat.
VS pellet and VS solution preparation
VS pellets were prepared daily : standard food pellets(Harlan Italy, Italy) were crushed by mortar andpestle and the fragments were dampened with waterand rolled in powdered vanilla sugar (ZuccheroVanigliato, Italy) to obtain pellets weighing approxi-mately 150 mg. The 20% VS solution was prepareddaily by dissolving powdered vanilla sugar in water.
Induction of VAB
A dark Plexiglas Y-maze (15r40r20 cm for each ofthe three arms) was used; a VS pellet was placed in a
334 S. Scheggi et al.

small tray at the end of one of the two divergent arms.Sessions of 10 trials with 15-min intervals betweenthemwere administered daily for a total of 10 sessions,as previously described (Ghiglieri et al. 1997). Eitherthe right or the left arm was designated correct,balanced among animals. If the empty armwas chosen(incorrect trial), the rat was returned to its cage for15 min before the next trial. When the baited arm waschosen (correct trial) the rat was allowed to consumethe VS pellet and then returned to its cage for 15 minbefore the next trial. A trial was defined as incompletewhen the rat did not reach the end of one of the twodiverging arms. In each session, the variables recordedwere: number of correct, incorrect, incomplete trials,and number of VS pellets consumed.
Taste reactivity test
Twenty-four hours before the test, anaesthetizedrats (pentobarbital 50 mg/kg, scopolamine 0.4 mg/kgi.p.) were implanted with an oral cannula as pre-viously described (Gambarana et al. 2003) to allowinfusion of VS solution into the mouth. On test day,each animal received a 15-s infusion of VS solutionand patterns of a!ective taste reactivity were scoredduring the infusion and the 30-s post-infusion period.Hedonic reactions included lateral and rhythmictongue protrusions (Berridge & Robinson, 1998) thatwere counted as individual events and assigned ascore of 1.
Microdialysis procedure
Anaesthetized rats (pentobarbital 50 mg/kg, scopol-amine 0.4 mg/kg i.p.) were placed in a stereotaxicinstrument and a concentric vertical probe waslowered into the NAcS (AP +1.7 mm, L ¡1.2 mmfrom bregma, V x8.0 mm from skull surface) accord-ing to Paxinos & Watson (1998), and as previouslydescribed (Danielli et al. 2010 ; Rauggi et al. 2005b) [seealso Supplementary Material (available online)]. Aftersurgery, rats had 24 h recovery before the beginningof microdialysis. Water and standard food pelletswere available at this time and up to the end of theexperiment.
Immunoblotting
Rats were sacrificed, their heads were briefly im-mersed (3–5 s) in liquid nitrogen and the brainsrapidly removed and cut into 1-mm slices using anice-cold metal brain matrix (ASI Instruments Inc.,USA). The mPFC and NAcS were dissected out fromthe slices corresponding to plates 7–9 and 10–12,
respectively, of the rat brain atlas (Paxinos & Watson,1998). Tissues were flash-frozen in liquid nitrogenand stored at x80 xC until assayed. Immunoblottingwas performed as previously described (Danielliet al. 2010; Scheggi et al. 2007) and detailed in theSupplementary Material (available online). In theregions studied, the total amount of DARPP-32 wasunmodified in the di!erent experimental groupscompared to the control group (see Figs 6 and 7).
Drugs
Imi and cocaine were dissolved in deionized/distilledwater and injected at a volume of 1 ml/kg rat bodyweight ; rats in the control groups received the samevolume of saline. Pentobarbital was dissolved in amixture of 12% ethanol, 38% propylene glycol and50% deionized/distilled water (v/v) containingscopolamine, and was injected at a volume of 4 ml/kgbody weight. Pentobarbital was purchased fromSigmaChemical Co. (USA) and cocaine fromMacfarlanSmith Limited (UK). All other drugs and chemicalswere purchased from commercial sources.
Statistical analysis
Statistical analyses were performed on commerciallyavailable software (Prism 4.0a, GraphPad SoftwareInc., USA). Data from the microdialysis and VABexperiments were analysed using a two-way, mixedfactorial, repeated-measures ANOVA (rm-ANOVA)with treatment as the between-subject variable andtime or session as the within-subject variable. Westernblot data in each experimental group were analysedusing one-way ANOVA. Post-hoc analysis was per-formed by Dunnett’s or Bonferroni’s tests, whenp<0.05. The unpaired t test was used to compare twoexperimental groups.
Experimental design
In each experiment, rats underwent the sensitizationprotocol, or received saline [1 ml/kg.d i.p., for 8 d,repeated saline (RS)]. After a 10-d washout, all cocaine-treated and a group of five RS rats were challengedwith cocaine (5 mg/kg i.p.) to assess sensitization.In each experiment, cocaine-treated rats that showeda sensitized response [i.e. stereotypy scores, loco-motor activity counts and rearings significantly higherthan the RS group: cocaine-sensitized (CS)] and cocaine-naive RS rats were used. All experiments began 10 dafter the sensitization test (i.e. 21 d after the end ofthe sensitization protocol). Since preliminary experi-ments indicated that CS rats had little interest in
Cocaine sensitization and anhedonia 335

VS pellets, in order to study the acute response to VSconsumption (microdialysis and immunoblotting ex-periments) rats were surgically implanted with oralcannulas for VS solution delivery.
Expt 1. Behavioural and neurochemical responsesto palatable food in CS rats
Expt 1A. Taste reactivity pattern and dopaminergic outputin the NAcS in response to VS consumption in controland CS rats
RS (n=5) and CS (n=8) rats were implanted with oralcannulas and microdialysis probes ; 24 h later theyunderwent microdialysis and were administered VSsolution : behavioural responses were recorded andmicrodialysis samples were collected.
Expt 1B. Modifications in the phosphorylation levelsof DARPP-32 in the NAcS after VS consumption incontrol and CS rats
RS and CS rats naive to VS (n=25 in each treatmentgroup) received 1 wk habituation (individuallyhoused in a clean cage for 1 h in the testing room), thenthey were implanted with oral cannulas and 24 h laterwere sacrificed at di!erent times after administrationof VS solution (30 min, 2, 3 and 4 h) ; groups of RS andCS rats received water (time-0 groups) (n=5 in eachtime group).
Expt 1C. VAB acquisition in CS rats
RS and CS rats were exposed to the Y-maze for VABacquisition (n=11 in each group).
Expt 2. E!ects of Imi treatment on theanhedonia-like condition of CS rats
This experiment was performed in order to study thee!ects of a classical antidepressant compound onthe anhedonia-like condition observed in expt 1.
Expt 2A. Dopaminergic output in the NAcS in responseto VS consumption in control and CS rats treated ornot with Imi
RS rats received saline (1 ml/kg) and CS rats receivedsaline (1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for17 d. On day 17, RS, CS and CS+Imi rats were im-planted with oral cannulas and microdialysis probes ;24 h later, 14 h after the last treatment, they underwentmicrodialysis (n=5–6 in each group). In a separateexperiment, RS rats received saline (1 ml/kg, n=5)or Imi (5 mg/kg, n=6) i.p. twice a day for 17 days,they were then implanted with oral cannulas and
microdialysis probes and underwent microdialysis24 h later.
Expt 2B. E!ect of repeated Imi treatment on the abilityof CS rats to acquire VAB
RS rats received saline (1 ml/kg) and CS rats receivedsaline (1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for7 d before beginning and during the 10-d exposureto the Y-maze (n=8 in each group). Since previousresults demonstrated that Imi treatment does notmodify the ability of control rats to acquire VAB(Gambarana et al. 2001), a group of RS+Imi rats wasnot included. Animals were sacrificed 24 h after thelast session, brains were removed and the NAcS andmPFC were excised for immunoblotting (expt 2D).
Expt 2C. Taste reactivity pattern in response to VSconsumption in control and CS rats treated or not with Imi
CS and RS rats received saline (1 ml/kg) or Imi(5 mg/kg) i.p. twice a day. After 17 d of treatment, ratswere implanted with oral cannulas ; 24 h later (14–16 hafter the last treatment) they were administered VSsolution and tongue protrusions were counted (n=8in each group). The expression of sensitization wasthen examined in the RS, CS+saline and CS+Imigroups with a 5 mg/kg cocaine challenge, 7 d after thelast Imi or saline administration.
Expt 2D. Modifications in the DARPP-32 phosphorylationpattern and Cdk5 levels in the NAcS and mPFC ofcontrol and CS rats treated or not with Imi and exposedor not to VAB
This experiment was performed to analyse whetherImi treatment would restore to control values theDARPP-32 phosphorylation pattern and Cdk5 levelsin the NAcS and mPFC that are modified in CSrats (Scheggi et al. 2004, 2007). Moreover, the possiblee!ect of Y-maze exposure on these variables was alsoexamined (rats from expt 2B). CS and RS rats receivedsaline (1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for17 d before the experiment. Fourteen hours afterthe last treatment, RS, RS+Imi, CS and CS+Imi ratswere sacrificed, their brains were removed and theNAcS and mPFC were excised for immunoblotting(n=5–6 in each group).
Results
All cocaine-treated rats developed behavioural sensi-tization (Supplementary Table S1, available online),and all experiments began 10 d after the sensitizationtest.
336 S. Scheggi et al.
Expt 1. Behavioural and neurochemical responses topalatable food in CS rats
Expt 1A. Taste reactivity pattern and dopaminergic outputin the NAcS in response to VS consumption in controland CS rats
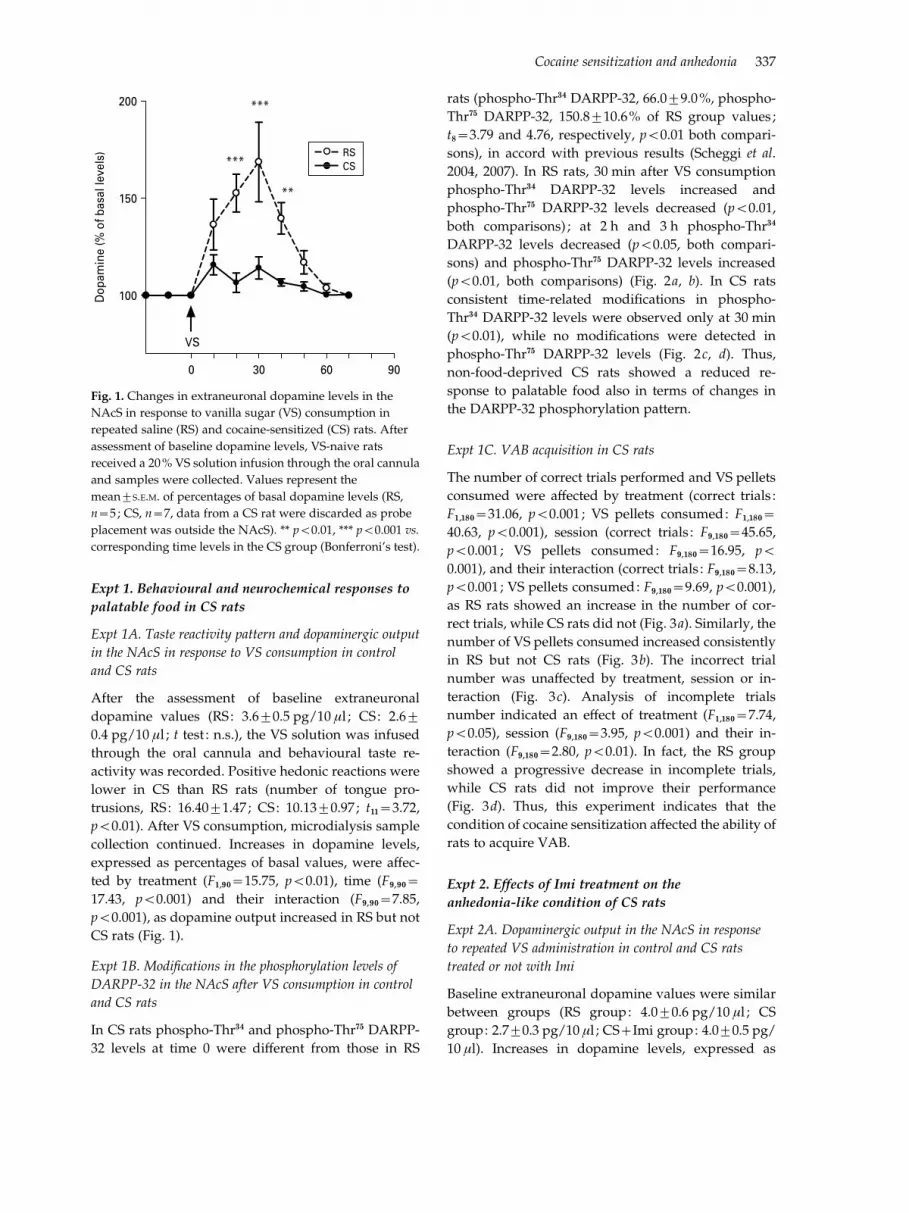
After the assessment of baseline extraneuronaldopamine values (RS: 3.6¡0.5 pg/10 ml ; CS: 2.6¡0.4 pg/10 ml ; t test : n.s.), the VS solution was infusedthrough the oral cannula and behavioural taste re-activity was recorded. Positive hedonic reactions werelower in CS than RS rats (number of tongue pro-trusions, RS: 16.40¡1.47 ; CS: 10.13¡0.97 ; t11=3.72,p<0.01). After VS consumption, microdialysis samplecollection continued. Increases in dopamine levels,expressed as percentages of basal values, were a!ec-ted by treatment (F1,90=15.75, p<0.01), time (F9,90=17.43, p<0.001) and their interaction (F9,90=7.85,p<0.001), as dopamine output increased in RS but notCS rats (Fig. 1).
Expt 1B. Modifications in the phosphorylation levels ofDARPP-32 in the NAcS after VS consumption in controland CS rats
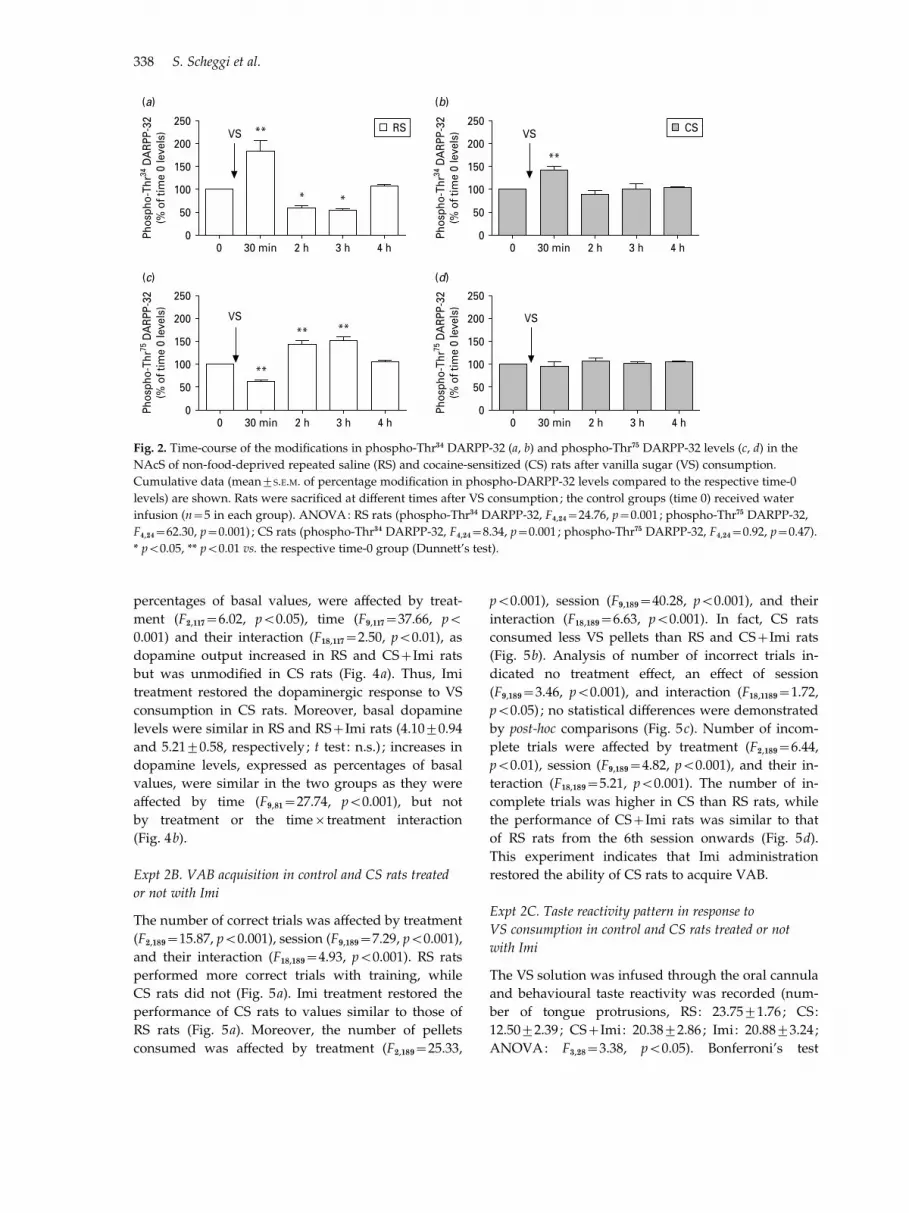
In CS rats phospho-Thr34 and phospho-Thr75 DARPP-32 levels at time 0 were di!erent from those in RS
rats (phospho-Thr34 DARPP-32, 66.0¡9.0%, phospho-Thr75 DARPP-32, 150.8¡10.6% of RS group values ;t8=3.79 and 4.76, respectively, p<0.01 both compari-sons), in accord with previous results (Scheggi et al.2004, 2007). In RS rats, 30 min after VS consumptionphospho-Thr34 DARPP-32 levels increased andphospho-Thr75 DARPP-32 levels decreased (p<0.01,both comparisons) ; at 2 h and 3 h phospho-Thr34
DARPP-32 levels decreased (p<0.05, both compari-sons) and phospho-Thr75 DARPP-32 levels increased(p<0.01, both comparisons) (Fig. 2a, b). In CS ratsconsistent time-related modifications in phospho-Thr34 DARPP-32 levels were observed only at 30 min(p<0.01), while no modifications were detected inphospho-Thr75 DARPP-32 levels (Fig. 2c, d). Thus,non-food-deprived CS rats showed a reduced re-sponse to palatable food also in terms of changes inthe DARPP-32 phosphorylation pattern.
Expt 1C. VAB acquisition in CS rats
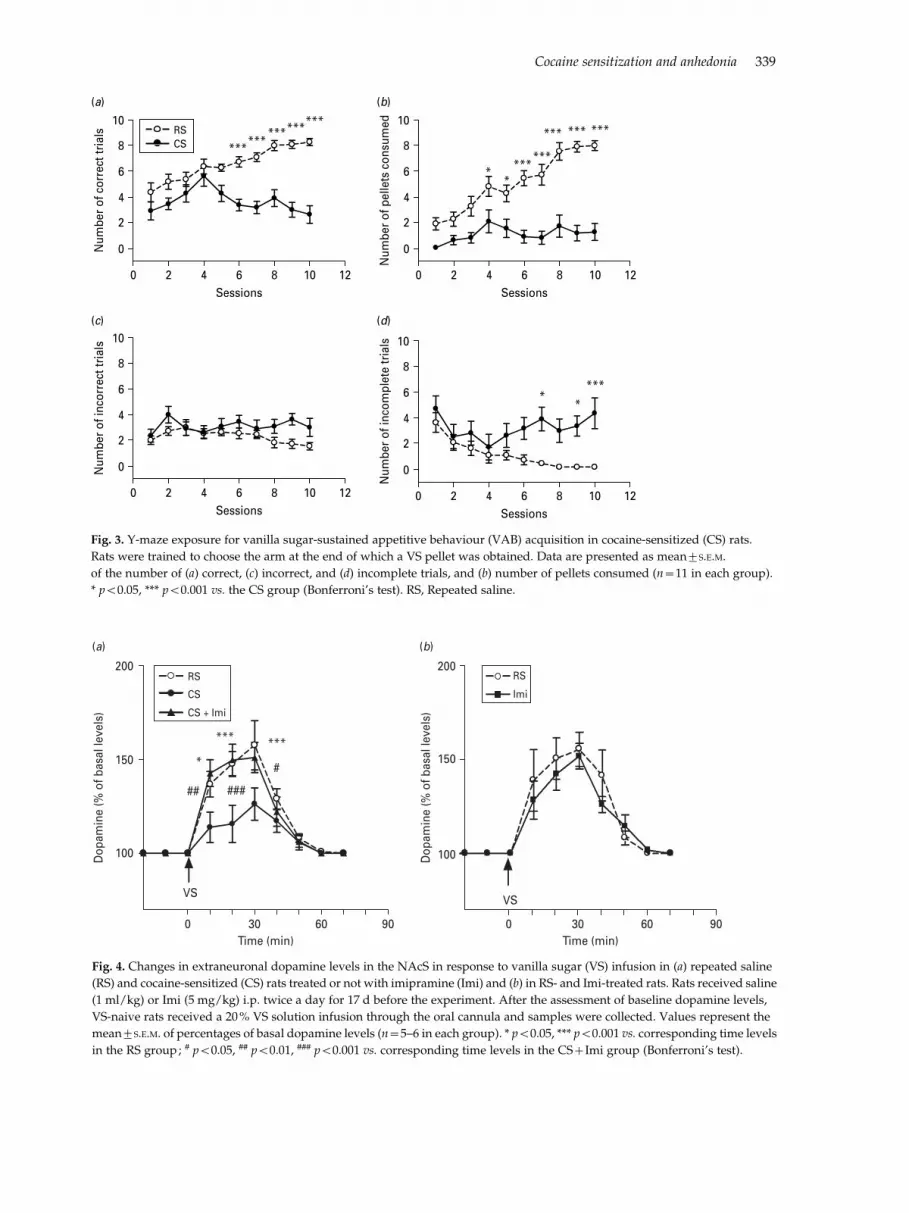
The number of correct trials performed and VS pelletsconsumed were a!ected by treatment (correct trials :F1,180=31.06, p<0.001 ; VS pellets consumed: F1,180=40.63, p<0.001), session (correct trials : F9,180=45.65,p<0.001 ; VS pellets consumed: F9,180=16.95, p<0.001), and their interaction (correct trials : F9,180=8.13,p<0.001 ; VS pellets consumed: F9,180=9.69, p<0.001),as RS rats showed an increase in the number of cor-rect trials, while CS rats did not (Fig. 3a). Similarly, thenumber of VS pellets consumed increased consistentlyin RS but not CS rats (Fig. 3b). The incorrect trialnumber was una!ected by treatment, session or in-teraction (Fig. 3c). Analysis of incomplete trialsnumber indicated an e!ect of treatment (F1,180=7.74,p<0.05), session (F9,180=3.95, p<0.001) and their in-teraction (F9,180=2.80, p<0.01). In fact, the RS groupshowed a progressive decrease in incomplete trials,while CS rats did not improve their performance(Fig. 3d). Thus, this experiment indicates that thecondition of cocaine sensitization a!ected the ability ofrats to acquire VAB.
Expt 2. E!ects of Imi treatment on theanhedonia-like condition of CS rats
Expt 2A. Dopaminergic output in the NAcS in responseto repeated VS administration in control and CS ratstreated or not with Imi
Baseline extraneuronal dopamine values were similarbetween groups (RS group: 4.0¡0.6 pg/10 ml ; CSgroup: 2.7¡0.3 pg/10 ml ; CS+Imi group: 4.0¡0.5 pg/10 ml). Increases in dopamine levels, expressed as
200
150
100Dop
amin
e (%
of b
asal
leve
ls)
VS
0 30 60 90
***
***
**
RSCS
Fig. 1. Changes in extraneuronal dopamine levels in the
NAcS in response to vanilla sugar (VS) consumption in
repeated saline (RS) and cocaine-sensitized (CS) rats. After
assessment of baseline dopamine levels, VS-naive rats
received a 20% VS solution infusion through the oral cannula
and samples were collected. Values represent the
mean¡S.E.M. of percentages of basal dopamine levels (RS,
n=5 ; CS, n=7, data from a CS rat were discarded as probe
placement was outside the NAcS). ** p<0.01, *** p<0.001 vs.
corresponding time levels in the CS group (Bonferroni’s test).
Cocaine sensitization and anhedonia 337
percentages of basal values, were a!ected by treat-ment (F2,117=6.02, p<0.05), time (F9,117=37.66, p<0.001) and their interaction (F18,117=2.50, p<0.01), asdopamine output increased in RS and CS+Imi ratsbut was unmodified in CS rats (Fig. 4a). Thus, Imitreatment restored the dopaminergic response to VSconsumption in CS rats. Moreover, basal dopaminelevels were similar in RS and RS+Imi rats (4.10¡0.94and 5.21¡0.58, respectively ; t test : n.s.) ; increases indopamine levels, expressed as percentages of basalvalues, were similar in the two groups as they werea!ected by time (F9,81=27.74, p<0.001), but notby treatment or the timertreatment interaction(Fig. 4b).
Expt 2B. VAB acquisition in control and CS rats treatedor not with Imi
The number of correct trials was a!ected by treatment(F2,189=15.87, p<0.001), session (F9,189=7.29, p<0.001),and their interaction (F18,189=4.93, p<0.001). RS ratsperformed more correct trials with training, whileCS rats did not (Fig. 5a). Imi treatment restored theperformance of CS rats to values similar to those ofRS rats (Fig. 5a). Moreover, the number of pelletsconsumed was a!ected by treatment (F2,189=25.33,
p<0.001), session (F9,189=40.28, p<0.001), and theirinteraction (F18,189=6.63, p<0.001). In fact, CS ratsconsumed less VS pellets than RS and CS+Imi rats(Fig. 5b). Analysis of number of incorrect trials in-dicated no treatment e!ect, an e!ect of session(F9,189=3.46, p<0.001), and interaction (F18,1189=1.72,p<0.05) ; no statistical di!erences were demonstratedby post-hoc comparisons (Fig. 5c). Number of incom-plete trials were a!ected by treatment (F2,189=6.44,p<0.01), session (F9,189=4.82, p<0.001), and their in-teraction (F18,189=5.21, p<0.001). The number of in-complete trials was higher in CS than RS rats, whilethe performance of CS+Imi rats was similar to thatof RS rats from the 6th session onwards (Fig. 5d).This experiment indicates that Imi administrationrestored the ability of CS rats to acquire VAB.
Expt 2C. Taste reactivity pattern in response toVS consumption in control and CS rats treated or notwith Imi
The VS solution was infused through the oral cannulaand behavioural taste reactivity was recorded (num-ber of tongue protrusions, RS : 23.75¡1.76 ; CS:12.50¡2.39 ; CS+Imi : 20.38¡2.86 ; Imi : 20.88¡3.24 ;ANOVA: F3,28=3.38, p<0.05). Bonferroni’s test
250
200
150
100
50
0Phos
pho-
Thr34
DA
RPP-
32(%
of t
ime
0 le
vels
)
0 30 min 2 h 3 h 4 h
250
200
150
100
50
0Phos
pho-
Thr34
DA
RPP-
32(%
of t
ime
0 le
vels
)
0 30 min 2 h 3 h 4 h
VS
250
200
150
100
50
0Phos
pho-
Thr75
DA
RPP-
32(%
of t
ime
0 le
vels
)
0 30 min 2 h 3 h 4 h
250
200
150
100
50
0Phos
pho-
Thr75
DA
RPP-
32(%
of t
ime
0 le
vels
)
0 30 min 2 h 3 h 4 h
VS
VS **
**
* *
VS
**
** **
(a) (b)
(c) (d)
RS CS
Fig. 2. Time-course of the modifications in phospho-Thr34 DARPP-32 (a, b) and phospho-Thr75 DARPP-32 levels (c, d) in the
NAcS of non-food-deprived repeated saline (RS) and cocaine-sensitized (CS) rats after vanilla sugar (VS) consumption.
Cumulative data (mean¡S.E.M. of percentage modification in phospho-DARPP-32 levels compared to the respective time-0
levels) are shown. Rats were sacrificed at di!erent times after VS consumption ; the control groups (time 0) received water
infusion (n=5 in each group). ANOVA: RS rats (phospho-Thr34 DARPP-32, F4,24=24.76, p=0.001 ; phospho-Thr75 DARPP-32,
F4,24=62.30, p=0.001) ; CS rats (phospho-Thr34 DARPP-32, F4,24=8.34, p=0.001 ; phospho-Thr75 DARPP-32, F4,24=0.92, p=0.47).
* p<0.05, ** p<0.01 vs. the respective time-0 group (Dunnett’s test).
338 S. Scheggi et al.
200
150
100Dop
amin
e (%
of b
asal
leve
ls)
VS
0 30 60 90Time (min)
200
150
100Dop
amin
e (%
of b
asal
leve
ls)
VS
0 30 60 90Time (min)
*** ***
#
#####
*
RS
CS
CS + Imi
RS
Imi
(a) (b)
Fig. 4. Changes in extraneuronal dopamine levels in the NAcS in response to vanilla sugar (VS) infusion in (a) repeated saline
(RS) and cocaine-sensitized (CS) rats treated or not with imipramine (Imi) and (b) in RS- and Imi-treated rats. Rats received saline
(1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for 17 d before the experiment. After the assessment of baseline dopamine levels,
VS-naive rats received a 20% VS solution infusion through the oral cannula and samples were collected. Values represent the
mean¡S.E.M. of percentages of basal dopamine levels (n=5–6 in each group). * p<0.05, *** p<0.001 vs. corresponding time levels
in the RS group; # p<0.05, ## p<0.01, ### p<0.001 vs. corresponding time levels in the CS+Imi group (Bonferroni’s test).
10
8
6
4
2
0Num
ber o
f cor
rect
tria
ls
0 2 4 6 8 10 12Sessions
10
8
6
4
2
0
Num
ber o
f pel
lets
con
sum
ed
0 2 4 6 8 10 12Sessions
10
8
6
4
2
0Num
ber o
f inc
orre
ct tr
ials
0 2 4 6 8 10 12Sessions
10
8
6
4
2
0
Num
ber o
f inc
ompl
ete
tria
ls
0 2 4 6 8 10 12Sessions
***
**
***
*********
*******
*
************
(a) (b)
(c) (d)
RSCS
Fig. 3. Y-maze exposure for vanilla sugar-sustained appetitive behaviour (VAB) acquisition in cocaine-sensitized (CS) rats.
Rats were trained to choose the arm at the end of which a VS pellet was obtained. Data are presented as mean¡S.E.M.
of the number of (a) correct, (c) incorrect, and (d) incomplete trials, and (b) number of pellets consumed (n=11 in each group).
* p<0.05, *** p<0.001 vs. the CS group (Bonferroni’s test). RS, Repeated saline.
Cocaine sensitization and anhedonia 339
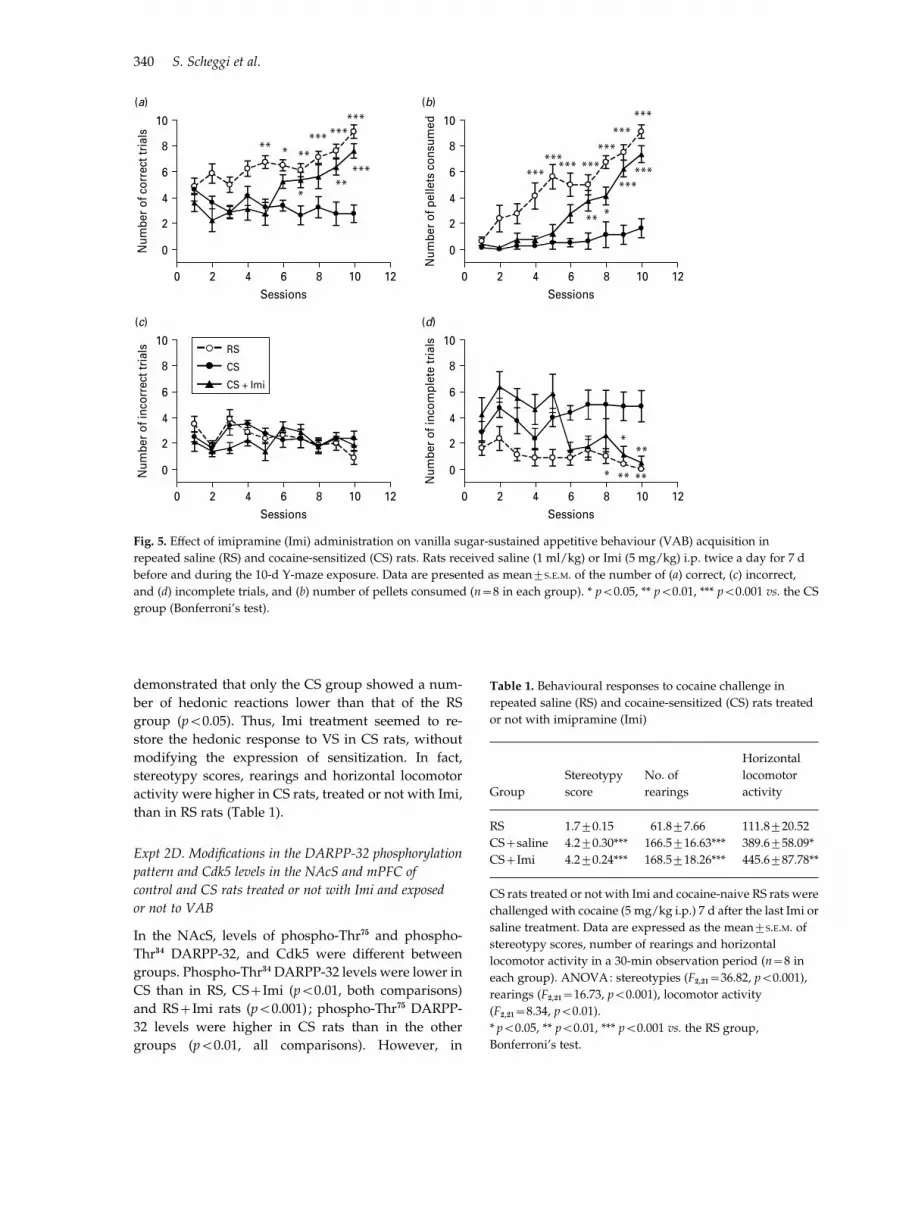
demonstrated that only the CS group showed a num-ber of hedonic reactions lower than that of the RSgroup (p<0.05). Thus, Imi treatment seemed to re-store the hedonic response to VS in CS rats, withoutmodifying the expression of sensitization. In fact,stereotypy scores, rearings and horizontal locomotoractivity were higher in CS rats, treated or not with Imi,than in RS rats (Table 1).
Expt 2D. Modifications in the DARPP-32 phosphorylationpattern and Cdk5 levels in the NAcS and mPFC ofcontrol and CS rats treated or not with Imi and exposedor not to VAB
In the NAcS, levels of phospho-Thr75 and phospho-Thr34 DARPP-32, and Cdk5 were di!erent betweengroups. Phospho-Thr34 DARPP-32 levels were lower inCS than in RS, CS+Imi (p<0.01, both comparisons)and RS+Imi rats (p<0.001) ; phospho-Thr75 DARPP-32 levels were higher in CS rats than in the othergroups (p<0.01, all comparisons). However, in
10
8
6
4
2
0Num
ber o
f cor
rect
tria
ls
0 2 4 6 8 10 12Sessions
(a)
10
8
6
4
2
0Num
ber o
f inc
orre
ct tr
ials
0 2 4 6 8 10 12Sessions
(c)
10
8
6
4
2
0
Num
ber o
f pel
lets
con
sum
ed
0 2 4 6 8 10 12Sessions
(b)
10
8
6
4
2
0
Num
ber o
f inc
ompl
ete
tria
ls
0 2 4 6 8 10 12Sessions
(d)
RS
CS
CS + Imi
******
***
********
***
******
******
************
***
** *
***
*****
Fig. 5. E!ect of imipramine (Imi) administration on vanilla sugar-sustained appetitive behaviour (VAB) acquisition in
repeated saline (RS) and cocaine-sensitized (CS) rats. Rats received saline (1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for 7 d
before and during the 10-d Y-maze exposure. Data are presented as mean¡S.E.M. of the number of (a) correct, (c) incorrect,
and (d) incomplete trials, and (b) number of pellets consumed (n=8 in each group). * p<0.05, ** p<0.01, *** p<0.001 vs. the CS
group (Bonferroni’s test).
Table 1. Behavioural responses to cocaine challenge in
repeated saline (RS) and cocaine-sensitized (CS) rats treated
or not with imipramine (Imi)
Group
Stereotypy
score
No. of
rearings
Horizontal
locomotor
activity
RS 1.7¡0.15 61.8¡7.66 111.8¡20.52
CS+saline 4.2¡0.30*** 166.5¡16.63*** 389.6¡58.09*
CS+Imi 4.2¡0.24*** 168.5¡18.26*** 445.6¡87.78**
CS rats treated or not with Imi and cocaine-naive RS rats were
challenged with cocaine (5 mg/kg i.p.) 7 d after the last Imi or
saline treatment. Data are expressed as the mean¡S.E.M. of
stereotypy scores, number of rearings and horizontal
locomotor activity in a 30-min observation period (n=8 in
each group). ANOVA: stereotypies (F2,21=36.82, p<0.001),
rearings (F2,21=16.73, p<0.001), locomotor activity
(F2,21=8.34, p<0.01).
* p<0.05, ** p<0.01, *** p<0.001 vs. the RS group,
Bonferroni’s test.
340 S. Scheggi et al.
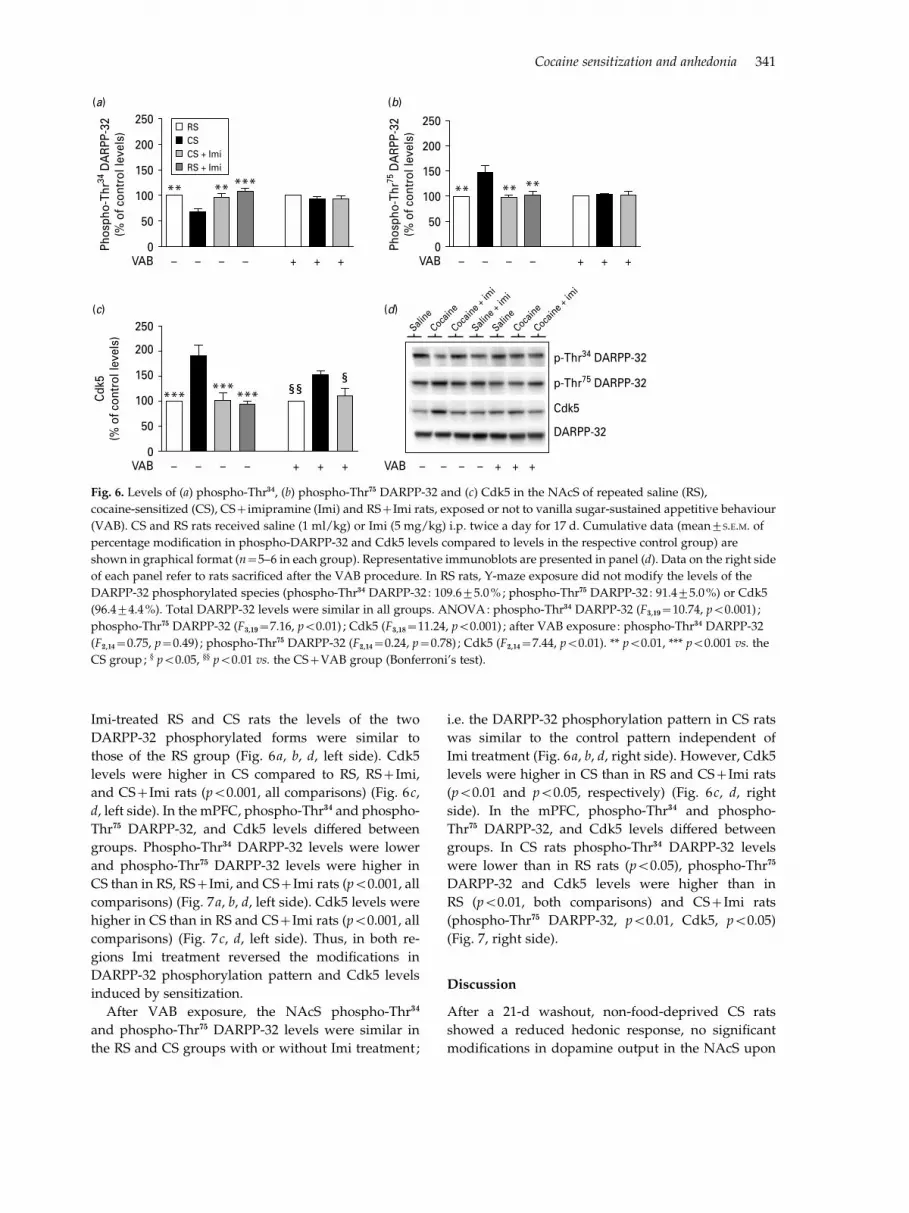
Imi-treated RS and CS rats the levels of the twoDARPP-32 phosphorylated forms were similar tothose of the RS group (Fig. 6a, b, d, left side). Cdk5levels were higher in CS compared to RS, RS+Imi,and CS+Imi rats (p<0.001, all comparisons) (Fig. 6c,d, left side). In the mPFC, phospho-Thr34 and phospho-Thr75 DARPP-32, and Cdk5 levels di!ered betweengroups. Phospho-Thr34 DARPP-32 levels were lowerand phospho-Thr75 DARPP-32 levels were higher inCS than in RS, RS+Imi, and CS+Imi rats (p<0.001, allcomparisons) (Fig. 7a, b, d, left side). Cdk5 levels werehigher in CS than in RS and CS+Imi rats (p<0.001, allcomparisons) (Fig. 7c, d, left side). Thus, in both re-gions Imi treatment reversed the modifications inDARPP-32 phosphorylation pattern and Cdk5 levelsinduced by sensitization.
After VAB exposure, the NAcS phospho-Thr34
and phospho-Thr75 DARPP-32 levels were similar inthe RS and CS groups with or without Imi treatment ;
i.e. the DARPP-32 phosphorylation pattern in CS ratswas similar to the control pattern independent ofImi treatment (Fig. 6a, b, d, right side). However, Cdk5levels were higher in CS than in RS and CS+Imi rats(p<0.01 and p<0.05, respectively) (Fig. 6c, d, rightside). In the mPFC, phospho-Thr34 and phospho-Thr75 DARPP-32, and Cdk5 levels di!ered betweengroups. In CS rats phospho-Thr34 DARPP-32 levelswere lower than in RS rats (p<0.05), phospho-Thr75
DARPP-32 and Cdk5 levels were higher than inRS (p<0.01, both comparisons) and CS+Imi rats(phospho-Thr75 DARPP-32, p<0.01, Cdk5, p<0.05)(Fig. 7, right side).
Discussion
After a 21-d washout, non-food-deprived CS ratsshowed a reduced hedonic response, no significantmodifications in dopamine output in the NAcS upon
250
200
150
100
50
0Phos
pho-
Thr34
DA
RPP-
32(%
of c
ontr
ol le
vels
)(a)
VAB – – – – + + +
250
200
150
100
50
0
Cdk5
(% o
f con
trol
leve
ls)
(c) (d)
VAB – – – – + + + VAB – – – – + + +
250
200
150
100
50
0Phos
pho-
Thr75
DA
RPP-
32(%
of c
ontr
ol le
vels
)
(b)
VAB – – – – + + +
Saline
Cocaine
Cocaine +
imi
Saline +
imi
Saline
Cocaine
Cocaine +
imi
p-Thr34 DARPP-32
p-Thr75 DARPP-32
DARPP-32
Cdk5
** ** ** ** **
****** ***
***
§§§
RSCSCS + ImiRS + Imi
Fig. 6. Levels of (a) phospho-Thr34, (b) phospho-Thr75 DARPP-32 and (c) Cdk5 in the NAcS of repeated saline (RS),
cocaine-sensitized (CS), CS+imipramine (Imi) and RS+Imi rats, exposed or not to vanilla sugar-sustained appetitive behaviour
(VAB). CS and RS rats received saline (1 ml/kg) or Imi (5 mg/kg) i.p. twice a day for 17 d. Cumulative data (mean¡S.E.M. of
percentage modification in phospho-DARPP-32 and Cdk5 levels compared to levels in the respective control group) are
shown in graphical format (n=5–6 in each group). Representative immunoblots are presented in panel (d). Data on the right side
of each panel refer to rats sacrificed after the VAB procedure. In RS rats, Y-maze exposure did not modify the levels of the
DARPP-32 phosphorylated species (phospho-Thr34 DARPP-32 : 109.6¡5.0%; phospho-Thr75 DARPP-32 : 91.4¡5.0%) or Cdk5
(96.4¡4.4%). Total DARPP-32 levels were similar in all groups. ANOVA: phospho-Thr34 DARPP-32 (F3,19=10.74, p<0.001) ;
phospho-Thr75 DARPP-32 (F3,19=7.16, p<0.01) ; Cdk5 (F3,18=11.24, p<0.001) ; after VAB exposure : phospho-Thr34 DARPP-32
(F2,14=0.75, p=0.49) ; phospho-Thr75 DARPP-32 (F2,14=0.24, p=0.78) ; Cdk5 (F2,14=7.44, p<0.01). ** p<0.01, *** p<0.001 vs. the
CS group; · p<0.05, ·· p<0.01 vs. the CS+VAB group (Bonferroni’s test).
Cocaine sensitization and anhedonia 341

VS ingestion and were unable to acquire VAB. Theyinitially explored the Y-maze apparatus but the fre-quency of correct choices did not increase and thenumber of incomplete choices remained higher thanthat of control animals. A non-food-deprived animaleats a palatable food because it is attracted by its odourand taste and the NAcS dopaminergic responsesignals the emotional value of the consumed food(Di Chiara, 2002). In non-food-deprived animals theemotional value of a palatable food has a prevalenthedonic component and VAB acquisition is prob-ably reinforced only by the hedonic value of VS.Consequently, a condition of reduced interest forand hedonic response to palatable food, associatedwith the lack of ability to acquire VAB, reasonablyreproduces the symptoms of ‘ loss of pleasure’ and‘lack of reactivity to usually pleasurable stimuli ’ thatdefine anhedonia as a melancholic feature in a major
depressive episode according to DSM-IV-TR criteria(APA, 2000). Thus, cocaine sensitization leads tobehavioural modifications that conform to face val-idity criteria for an experimental anhedonia model(McKinney & Bunney, 1969; Willner, 1995). Non-food-deprivation is a critical condition in this modeland it is then di"cult to compare it to models that arenot based on this condition or that use non-foodrewards (Fiorino & Phillips, 1999 ; Harmer & Phillips,1999). Since anhedonia also characterizes substanceabuse-related mood disorders, in particular centralstimulants abuse (APA, 2000), the condition of sensi-tization induced by repeated cocaine administrationalso conforms to construct validity requirements.The fact that cocaine sensitization-induced anhedoniawas observed after a 21-d washout and lasted un-modified for several weeks excludes possible commonmechanisms with the depressive-like symptoms
200
150
100
50
0Phos
pho-
Thr34
DA
RPP-
32(%
of c
ontr
ol le
vels
)
(a)
VAB – – – – + + +
(d)
VAB – – – – + + +
Saline
Cocaine
Cocaine +
imi
Saline +
imi
Saline
Cocaine
Cocaine +
imi
p-Thr34 DARPP-32
p-Thr75 DARPP-32
DARPP-32
Cdk5
200
150
100
50
0
Cdk5
(% o
f con
trol
leve
ls)
(c)
VAB – – – – + + +
200
150
100
50
0Phos
pho-
Thr75
DA
RPP-
32(%
of c
ontr
ol le
vels
)
(b)
VAB – – – – + + +
RSCSCS + ImiRS + Imi
*** *** *** *** *** *** §§ §§§
*** *** ***§§ §
Fig. 7. Levels of (a) phospho-Thr34, (b) phospho-Thr75 DARPP-32 and (c) Cdk5 in the mPFC of repeated saline (RS),
cocaine-sensitized (CS), CS+imipramine (Imi) and RS+Imi rats, exposed or not to vanilla sugar-sustained appetitive
behaviour (VAB). Cumulative data (mean¡S.E.M. of percentage modification in phospho-DARPP-32 and Cdk5 levels
compared to levels in the respective control group) are shown in graphical format (n=5–6 in each group). Representative
immunoblots are presented in panel (d). Data on the right side of each panel refer to rats sacrificed after the VAB procedure.
In RS rats, Y-maze exposure did not modify the levels of the DARPP-32 phosphorylated species (phospho-Thr34 DARPP-32 :
108.8¡5.4%, phospho-Thr75 DARPP-32 : 101.2¡8.7%) or Cdk5 (95.0¡4.3%). Total DARPP-32 levels were similar in all groups.
ANOVA: phospho-Thr34 DARPP-32 (F3,23=15.04, p<0.001) ; phospho-Thr75 DARPP-32 (F3,23=39.24, p<0.001) ; Cdk5
(F3,19=31.01, p<0.001) ; after VAB exposure : phospho-Thr34 DARPP-32 (F2,17=4.76, p<0.05) ; phospho-Thr75 DARPP-32
(F2,17=10.06, p<0.01) ; Cdk5 (F2,13=7.93, p<0.01). *** p<0.001 vs. the CS group; · p<0.05, ·· p<0.01 vs. the CS+VAB group
(Bonferroni’s test).
342 S. Scheggi et al.

described during acute psychostimulant withdrawal(Barr & Markou, 2005). Antidepressant drugs are,by definition, e!ective in treating depression and at-tenuating or reversing its symptoms. Imi adminis-tration did not modify the dopaminergic responseto VS consumption in control rats and reinstated thisresponse and the ability to acquire VAB in CS rats.The expression of locomotor sensitization was re-evaluated 7 d after Imi treatment, since long-termexposure to this antidepressant induces a transientincrease in motor response to central stimulants(Spyraki & Fibiger, 1981). Interestingly, the expressionof sensitization was una!ected by Imi treatment, asCS rats showed similar responses to cocaine challengeirrespective of Imi or saline treatment. This means theanhedonia-like condition appears to be independentof behavioural sensitization. Therefore, the conditionof sensitization conforms to predictive validity re-quirements, although this data should be confirmedby using other antidepressant treatments. The aim ofthis study was to model anhedonia in rats, and thebehavioural and neurochemical parameters measuredwere used to better define the model. A standardizedsensitization paradigm was utilized (Scheggi et al.2007), since the development and expression of thebehavioural and neurochemical modifications thatcharacterize sensitization to central stimulants is in-fluenced by the experimental conditions (Carey &Damianopulos, 2006 ; Carey et al. 2005 ; Gambaranaet al. 1998 ; Kalivas et al. 1992 ; Klebaur et al. 2002; Liet al. 2004; Uslaner et al. 2001). Sensitization is alsoobserved upon cocaine self-administration, a con-dition in which the underlying neurobiologicalmechanisms result from the pharmacological e!ectsof the drug and the cognitive processes associatedwith self-administration (Robinson & Berridge, 1993,2003 ; Tzschentke & Schmidt, 2003 ; Wolf, 1998). Ratsself-administering cocaine for prolonged periods oftime show an elevated intracranial self-stimulationthreshold during cocaine withdrawal, compared topre-drug baseline. The magnitude and duration ofthis anhedonic-like state is proportional to the amountof cocaine consumed during each binge (Markou &Koob, 1991) and after repeated 6-h binges it may lastfor at least 8 d, but possible correlations with behav-ioural sensitization have not been explored (Ahmedet al. 2002). Thus, the development of an anhedonia-like condition cannot be generalized from our exper-imental conditions regarding what may be observedin rats exposed to di!erent sensitization proceduresor repeatedly self-administering cocaine.
The proposed anhedonia-like condition is not justa behaviour-basedmodel, but includes neurobiological
correlates (reduced response to palatable food con-sumption in terms of dopamine output and dopamineD1 receptor-dependent signalling in the NAcS) that areresponsive to Imi treatment and that probably under-pin the behavioural phenotype. Indeed, a reducedemotional value of palatable food in CS ratswas hypothesized on the basis of some neurochemicaladaptations that characterize cocaine sensitizationin rats ; in particular, the tonic increase in phospho-Thr75 DARPP-32 levels, accompanied by a reduceddopamine D1 receptor responsiveness in the NAc, thatlasts as long as the sensitization state (2–3 months)(Scheggi et al. 2007). In agreement with previous data(Scheggi et al. 2004, 2007), CS rats showed increasedCdk5 and phospho-Thr75 DARPP-32 levels and de-creased phospho-Thr34 DARPP-32 levels in the mPFCand NAcS. Imi treatment not only restored in CS ratsthe NAcS dopamine output in response to VS con-sumption, but it also reinstated to control values theCdk5, phospho-Thr75 and phospho-Thr34 DARPP-32levels in the mPFC and NAcS.
VAB exposure modified some of the neurobio-logical variables examined, selectively in the NAcSof CS rats. In particular, after Y-maze exposure theDARPP-32 phosphorylation pattern was similar tocontrol values in the NAcS of CS rats, while Cdk5levels were still elevated. Thus, environmental en-richment reinstated some of the altered neurochemicalparameters, although it did not a!ect the core deficit.Cdk5 is the kinase responsible for Thr75 DARPP-32phosphorylation (Bibb et al. 1999) and its increasedexpression, even in the presence of phospho-Thr75
DARPP-32 levels similar to control values, probablyimplies an incomplete functional recovery. On theother hand, in the mPFC of CS rats the DARPP-32phosphorylation pattern was still altered and Cdk5levels increased. The tonically increased phospho-Thr75 DARPP-32 levels may interfere with dopamine-and/or glutamate-dependent neurotransmission and,since the mPFC plays a central role in reinforcement-guided action selection (Seamans et al. 2008), an ad-ditional deficit in associative learning in CS rats cannotbe excluded.
Dopamine neurons fire in a slow irregular fashion,resulting in tonic dopamine release, but in responseto salient environmental stimuli they fire in bursts,which leads to phasic increases in dopamine release(Overton & Clark, 1997). Transient increases in dopa-mine release highlight salient environmental signalswhile suppressing irrelevant signals, helping an ani-mal to pay attention to the environment and respondappropriately (Nicola, 2000). When the cortical andmesolimbic dopaminergic transmission is absent or
Cocaine sensitization and anhedonia 343

reduced, animals appear to be hyporeactive and mostenvironmental stimuli go unnoticed (Palmiter, 2007).Accordingly, although the hedonic response to palat-able food is not modified by impaired mesolimbicdopaminergic transmission (Berridge et al. 1989) and isalso present in chronically decerebrate rats (Grill &Norgren, 1978), in CS rats the hedonic response toVS consumption was reduced. Indeed, the hedonicquality of a particular food is not constant as it maybe modified by learning or by modifications in theanimal’s internal state (Sewards, 2004). Since reducedmesolimbic dopaminergic transmission attenuatesthe motivation to work for food rewards (Salamoneet al. 2005), it has been proposed that dopamine opti-mizes responding performance to relevant stimuli,and theories to explain this role have been expressedin terms of incentive motivation (Crow, 1973), appeti-tive phases of motivated behaviour (Fibiger & Phillips,1986), expectancy of motivational targets (Panksepp,1986), sensorimotor arousal (Salamone et al. 2005),incentive salience (Berridge, 2007), and/or incentivearousal (Di Chiara, 2002). The present results onlysupport the observations that decreased dopaminergicresponsiveness impairs an animal’s reactivity to sig-nificant environmental stimuli.
The VAB procedure is based on the reinforcingproperties of VS pellets, the earning of which is con-tingent on the choice of a Y-maze arm. VS emotionalvalue appeared to be reduced in CS rats that showeddecreased hedonic taste reactivity and reduced dopa-minergic response in the NAcS to its consumption.Repeated exposure to unavoidable stress disruptsthe ability to acquire VAB (Ghiglieri et al. 1997)and we have used this procedure as a model ofanhedonia for studying the e!ects of antidepressantcompounds (Gambarana et al. 2001; Rauggi et al.2005a). However, the chronic stress protocol is arather invasive procedure and the present datasuggest that cocaine sensitization may for some pur-poses be a valid alternative to chronic unavoidablestress-induced anhedonia. Moreover, the cocainesensitization-induced anhedonia lasts for a periodsu"cient to allow for long-term antidepressant treat-ments or behavioural manipulations. Finally, cocaine-induced sensitization is easily obtained and it can bestudied as a condition independent of the addictiveproperties of cocaine.
Note
Supplementary material accompanies this paper onthe Journal’s website (http://journals.cambridge.org/pnp).
Acknowledgments
The helpful discussions and critical comments onthe manuscript by Professor Alessandro Tagliamonteare gratefully acknowledged. The authors thank Ms.Colleen Pisaneschi for language editing of the manu-script.
Statement of Interest
None.
References
Ahmed SH, Kenny PJ, Koob GF, Markou A (2002).
Neurobiological evidence for hedonic allostasis
associated with escalating cocaine use. Nature Neuroscience
5, 625–626.APA (2000). Diagnostic and Statistical Manual of Mental
Disorders, Text Revision (4th edn). Washington, DC:
American Psychiatric Association.Barr AM, Markou A (2005). Psychostimulant withdrawal
as an inducing condition in animal models of
depression. Neuroscience and Biobehavioral Reviews 29,
675–706.Berridge KC (2007). The debate over dopamine’s role in
reward : the case for incentive salience. Psychopharmacology
191, 391–431.Berridge KC, Robinson TE (1998). What is the role of
dopamine in reward: hedonic impact, reward
learning, or incentive salience? Brain Research Review 28,
309–369.Berridge KC, Venier IL, Robinson TE (1989). Taste
reactivity analysis of 6-hydroxydopamine-induced
aphagia : implications for arousal and anhedonia
hypotheses of dopamine function. Behavioral Neuroscience
103, 36–45.Bibb JA, Snyder GL, Nishi A, Yan Z, et al. (1999).
Phosphorylation of DARPP-32 by Cdk5 modulates
dopamine signalling in neurons. Nature 402, 669–671.Carey RJ, Damianopulos EN (2006). Cocaine conditioning
and sensitization: the habituation factor. Pharmacology,
Biochemistry and Behavior 84, 128–133.Carey RJ, DePalma G, Damianopoulos E (2005). Acute
and chronic cocaine behavioral e!ects in novel vs.
familiar environments : open-field familiarity di!erentiates
cocaine locomotor stimulant e!ects from cocaine
emotional behavioral e!ects. Behavioral Brain Research 158,
321–330.Crow TJ (1973). Catecholamine-containing neurones and
electrical selfstimulation. 2. A theoretical interpretation
and some psychiatric implications. Psychological Medicine 3,
66–73.Danielli B, Scheggi S, Grappi S, Marchese G, et al. (2010).
Modifications in DARPP-32 phosphorylation pattern
after repeated palatable food consumption undergo
344 S. Scheggi et al.

rapid habituation in the nucleus accumbens shell
of non food-deprived rats. Journal of Neurochemistry 112,
531–541.Di Chiara G (2002). Nucleus accumbens shell and core
dopamine : di!erential role in behavior and addiction.
Behavioral Brain Research 137, 75–114.Fibiger HC, Phillips AG (1986). Reward, motivation,
cognition : psychobiology of mesotelencephalic systems.
In : Bloom FE (Ed.), Handbook of Physiology – The Nervous
System (pp. 647–675). Bethesda: American Physiological
Society.Fiorino DF, Phillips AG (1999). Facilitation of sexual
behavior and enhanced dopamine e#ux in the nucleus
accumbens of male rats after d-amphetamine-induced
behavioral sensitization. Journal of Neuroscience 19,
456–463.Gambarana C, Ghiglieri O, De Montis MG, Tagliamonte A
(1998). Under continuous dizocilpine infusion an
N-methyl-D-aspartate receptor independent form of
cocaine sensitization develops in rats. Behavioural
Pharmacology 9, 61–68.Gambarana C, Masi F, Leggio B, Grappi S, et al. (2003).
Acquisition of a palatable-food-sustained appetitive
behavior in satiated rats is dependent on the dopaminergic
response to this food in limbic areas. Neuroscience 121,
179–187.Gambarana C, Scheggi S, Tagliamonte A, Tolu P, et al.
(2001). Animal models for the study of antidepressant
activity. Brain Research Protocols 7, 11–20.Ghiglieri O, Gambarana C, Scheggi S, Tagliamonte A, et al.
(1997). Palatable food induces an appetitive behaviour
in satiated rats which can be inhibited by chronic stress.
Behavioural Pharmacology 8, 619–628.Grill HJ, Norgren R (1978). The taste reactivity test. II.
Mimetic responses to gustatory stimuli in chronic
thalamic and chronic decerebrate rats. Brain Research 143,
281–297.Harmer CJ, Phillips GD (1999). Enhanced conditioned
inhibition following repeated pretreatment with
d–amphetamine. Psychopharmacology 142, 120–131.Hemmings Jr. HC, Greengard P, Tung HY, Cohen P (1984).
DARPP-32, a dopamine-regulated neuronal
phosphoprotein, is a potent inhibitor of protein
phosphatase-1. Nature 310, 503–505.Kalivas PW, Striplin CD, Steketee JD, Klitenick MA, et al.
(1992). Cellular mechanisms of behavioral sensitization
to drugs of abuse. Annals of the New York Academy of Science
654, 128–135.Klebaur JE, Ostrander MM, Norton CS, Watson SJ, et al.
(2002). The ability of amphetamine to evoke arc (Arg 3.1)
mRNA expression in the caudate, nucleus accumbens and
neocortex is modulated by environmental context. Brain
Research 930, 30–36.Li Y, Acerbo MJ, Robinson TE (2004). The induction
of behavioural sensitization is associated with
cocaine-induced structural plasticity in the core (but not
shell) of the nucleus accumbens. European Journal of
Neuroscience 20, 1647–1654.
Markou A, Koob GF (1991). Postcocaine anhedonia.
An animal model of cocaine withdrawal.
Neuropsychopharmacology 4, 17–26.McKinney Jr. WT, Bunney Jr. WE (1969). Animal model
of depression. I. Review of evidence : implications for
research. Archives of General Psychiatry 21, 240–248.Nicola SM (2000). Dopaminergic modulation of neuronal
excitability in the striatum and nucleus accumbens. Annual
Review of Neuroscience 23, 185–215.Overton PG, Clark D (1997). Burst firing in midbrain
dopaminergic neurons. Brain Research Reviews 25, 312–334.Palmiter RD (2007). Is dopamine a physiologically relevant
mediator of feeding behavior? Trends in Neurosciences 8,
375–381.Panksepp J (1986). The neurochemistry of behavior. Annual
Review of Psychology 37, 77–107.Paxinos G, Watson C (1998). The Rat Brain in Stereotaxic
Coordinates. New York : Academic Press.Rauggi R, Cassanelli A, Raone A, Tagliamonte A, et al.
(2005a). Study of mirtazapine antidepressant e!ects
in rats. International Journal of Neuropsychopharmacology 8,
369–379.Rauggi R, Scheggi S, Cassanelli A, De Montis MG, et al.
(2005b). The mesolimbic dopaminergic response to novel
palatable food consumption increases dopamine-D1
receptor-mediated signalling with complex modifications
of the DARPP-32 phosphorylation pattern. Journal of
Neurochemistry 92, 867–877.Robinson TE, Berridge KC (1993). The neural basis of drug
craving : an incentive-sensitization theory of addiction.
Brain Research Reviews 18, 247–291.Robinson TE, Berridge KC (2003). Addiction. Annual Review
of Psychology 54, 25–53.Salamone JD, Correa M, Mingote SM, Weber SM (2005).
Beyond the reward hypothesis : alternative functions of
nucleus accumbens dopamine. Current Opinion in
Pharmacology 5, 34–41.Scheggi S, Raone A, De Montis MG, Tagliamonte A, et al.
(2007). Behavioral expression of cocaine sensitization in
rats is accompanied by a distinct pattern of modifications
in the PKA/DARPP-32 signaling pathway. Journal of
Neurochemistry 103, 1168–1183.Scheggi S, Rauggi R, Gambarana C, Tagliamonte A, et al.
(2004). DARPP-32 phosphorylation pattern in cocaine and
morphine sensitized rats. Journal of Neurochemistry 90,
792–799.Seamans JK, Lapish CC, Durstewitz D (2008). Comparing
the prefrontal cortex of rats and primates : insights
from electrophysiology. Neurotoxicity Research 14, 249–262.Sewards TV (2004). Dual separate pathways for sensory
and hedonic aspects of taste. Brain Research Bulletin 62,
271–283.Spyraki C, Fibiger HC (1981). Behavioural evidence for
supersensitivity of postsynaptic dopamine receptors
in the mesolimbic system after chronic administration of
desipramine. European Journal of Pharmacology 74, 195–206.Tzschentke TM, Schmidt WJ (2003). Glutamatergic
mechanisms in addiction. Molecular Psychiatry 8, 373–382.
Cocaine sensitization and anhedonia 345

Uslaner J, Badiani A, Day HE, Watson SJ, et al. (2001).
Environmental context modulates the ability of cocaine
amphetamine to induce c-fos mRNA expression in the
neocortex, caudate nucleus and nucleus accumbens. Brain
Research 920, 106–116.Willner P (1995). Animal models of depression : validity
and applications. Advances in Biochemical
Psychopharmacology 49, 19–41.
Wolf ME (1998). The role of excitatory amino acids in
behavioral sensitization to psychomotor stimulants.
Progress in Neurobiology 54, 679–720.Zachariou V, Sgambato-Faure V, Sasaki T,
Svenningsson P, et al. (2006). Phosphorylation of
DARPP-32 at threonine-34 is required for cocaine action.
Neuropsychopharmacology 31, 555–562.
346 S. Scheggi et al.