no support for socio-physiological suppression effect on metabolism of paired white mice (mus sp.)
TRANSCRIPT

Introduction
Small mammals often huddle together in groups. Thereasons for huddling behaviour may be diverse.However, theoretical models based on the geometryof exposed surface areas (e.g. Canals, Rosenmann &Bozinovic 1989, 1997), and direct measurements ofthe energy expended by animals grouped together inhuddles (Andrews & Belknap 1986; Bryant & Hails
1975), suggest that an inevitable consequence ofhuddling may be that the energy expenditures of theindividual animals in the huddle are reduced whencompared with solitary individuals. This energysaving may itself be a selective advantage favouringthe evolution of huddling behaviour in certain circum-stances (Sealander 1952; Fedyk 1971; Gebczynska &Gebczynski 1971; Baudinette 1972; Karasov 1983;Springer, Gregory & Barrett 1981).
FunctionalEcology 199913,373–382
© 1999 BritishEcological Society
ORIGINAL ARTICLE OA 000 EN
No support for socio-physiological suppression effect onmetabolism of paired white mice (Mus sp.)
J. R. SPEAKMAN and F. P. ROSSIAberdeen Centre for Energy Regulation and Obesity, Department of Zoology, University of Aberdeen, AberdeenAB24 2TZ, UK
Summary
1. Three experiments were performed on white mice (strain MF1) to establish if therewas social or physiological suppression of metabolism, mediated by elevated ambientlevels of CO2, when animals were in close contact. In the first two experiments themetabolism of two mice were each measured solitarily and paired together in the samechamber, both with and without a partition that allowed visual, auditory and olfactorycommunication but restricted physical contact.2. Comparisons were made between the sum of the two solitary measurements, andthe observed measurements of paired individuals. In the first experiment air flow waslow and ambient CO2 was always at levels predicted to produce suppression (> 0·2%),independent of whether the animals were measured alone or paired.3. The paired mice had greater metabolism than expected from the sum of theirsolitary metabolic rates whether the partition was present or absent. This increasewas due to increased activity of the animals when in pairs. This experimentindicated that when CO2 levels were maintained in the range 0·2–0·5% there was nosocial suppression of metabolism.4. In experiment 2 metabolic rates were measured of solitary mice exposed tosubthreshold levels of CO2 (≈ 0·12%) and paired mice that experienced levels abovethe supposed threshold (≈ 0·24%). Again the observed metabolism of the paired miceexceeded the sum of their metabolic rates when solitary. The increase was alsoattributable to greater activity of the animals in pairs.5. This experiment indicated no social suppression effect when CO2 varied in therange 0·12–0·24%. Moreover there was no significant difference between the oxygenconsumptions of mice in experiment 2 compared with experiment 1, indicating thatindependent of the social context there was no suppressive effect of increases in CO2
concentration over the range 0·12–0·5% CO2.6. Finally, in experiment 3, when two mice were placed in sequential separatechambers (allowing no social contact), their summed metabolism did not differ fromthe sum of their solitary measurements when a CO2 absorber was placed in the airflowlinking the chambers. When the CO2 absorber was absent the observed metabolism ofthe pair exceeded the sum of the solitary measurements.7. Overall, across all three experiments no evidence was found for either a social or aphysiological suppression of metabolism mediated via CO2 (0·12–0·5%).
Key-words: Bioenergetics, energy expenditure, huddling, oxygen consumption
Functional Ecology(1999)13,373–382
373

The physical basis for the reduction in energyexpenditure of huddled animals has been the focus ofsome debate. On one hand the effect may be due toreductions in the mean individual exposed surfacearea of huddled vs solitary animals (Contreras 1984;Vickery & Millar 1984; Canals et al. 1989).Alternatively huddles of animals may elevate the localtemperatures more effectively than solitary animals,meaning the local temperature experienced by thehuddle is greater than that experienced by solitaryindividuals (Andrews, Phillips & Makihara 1987;Hayes, Speakman & Racey 1992a). Hayes et al.(1992a) partitioned these effects, controlling for dif-ferences in activity between huddled and solitaryShort-Tailed Field Voles (Microtus agrestis), andfound that approximately half the observed savingcould be attributed to each effect. Canals et al. (1997)suggested the majority of the effect was accounted forby reduced surface area.
Apart from the physical effects of huddling onenergy expenditure there may also be direct biologicaleffects of animals upon each other when they are inclose proximity. Thus two animals huddling togethermay have a calming effect on each other and mutuallyreduce each other’s metabolic rates. For example,Martin, Fiorentini & Connors (1980) found that singlewhite mice (Mussp.) and gerbils (Meriones unguicu-latus) had greater metabolic rates than huddled trios inthe same chamber, or of the same three animals sepa-rated by partitions to prevent huddling. In direct con-trast to these findings, however, Contreras (1984)found no suppressive effect on metabolism when triosof mice and gerbils were measured together but sepa-rated by partitions, indicating no social effect onmetabolic rate.
In part the different results found by Martin et al.(1980) and Contreras (1984) may reflect subtle differ-ences in experimental design and the fact that in somecircumstances there may be more direct physiologicaleffects of one animal on the metabolism of its imme-diate neighbours. Herreid & Schlenker (1980) foundthat if a mouse was placed in a completely separatechamber upstream of a second mouse, the metabolismof the second mouse was suppressed for the durationthat the two chambers were connected. These experi-ments led them to suggest that an airborne factor wasresponsible for suppressing the metabolism of the sec-ond mouse.
In a second paper, Schlenker & Herreid (1981) per-formed some further manipulations using the samesystem to try to establish the nature of the airbornefactor. The conclusion of this work was that the sup-pressing factor was elevated levels (0·2–0·8%) ofCO2 in the second chamber owing to exhaled CO2
from the first mouse. In further experiments,Schlenker, Carlson & Herreid (1981) demonstratedthat the effect was still present in anosmic mice, indi-cating that the effect was mediated by CO2 and notsome other olfactory cue.
Observations of socio-physiological effects ofsmall mammals on each other have several potentialimplications for observations of energy savings whenanimals come together in huddles. In particular, atleast part of the observed saving may be due to socialand physiological effects rather than any physicaleffect due to reduced surface area or local heating.Whether animal metabolic rates are reduced simplyby the presence of conspecifics, or whether suppres-sion is additionally mediated physiologically by lowambient levels of CO2 is an important question thathas ramifications not only for our understanding ofwhy energy savings might occur in huddled animals,but also for respirometry measurements of energyexpenditure in general. Measurements of resting andbasal metabolic rates, for instance, are always madefor animals that are solitary, and, if an open flow sys-tem is employed, also in the CO2-rich atmosphere of arespirometry chamber. If there is a ‘calming’ effectwhen other individuals are present that is absent if theanimal is alone, this may have ramifications for theinterpretation of the practical significance of BMRand RMR measurements. In particular, are solitaryanimals in the wild in the ‘calm’ state, or in the samestate as that pertaining in measurement protocols forBMR and RMR? Alternatively, does the artificiallyhigh level of CO2 in a respirometry chamber lead tosuppressed metabolism relative to that normally expe-rienced by animals in the wild? These potential effectsmay contribute to the reported discrepancies betweentime and energy budget calculations of daily energyexpenditures of animals, and more direct measure-ments based on the doubly labelled water technique(Weathers et al. 1984; Nagy 1989). In the currentpaper we report a series of experiments in which weaimed to assess the effects of social and physiologicalsuppression on the metabolism of white mice.
Methods
GENERAL DETAILS
We performed experiments on white mice (StrainMF1) housed separately in plastic cages (30 cm×15 cm×12 cm) at 20 °C, 12L:12D, with water andfood (Special Diet Services CRM; BP Nutrition Ltd,Wytham, Essex, UK) available ad libitum. All theexperiments involved standard respirometry mea-surements of oxygen consumption using a systemthat has been previously described (Speakman &Racey 1988; Hayes et al. 1992a,b). In experiments 1and 2 a single Perspex respirometry chamber measur-ing 25 × 25 × 12 cm was used, with sawdust in thebase to absorb urine. The chamber could be dividedinto two sections by inserting a Perspex sheet downthe centre. The divide had 16 5-mm holes drilled intoit. There was consequently a free passage of air acrossit, and also, as it was Perspex, the animals could seeeach other through it although they could not make
374J. R. Speakman& F. P. Rossi
© 1999 BritishEcological Society,Functional Ecology,13, 373–382

physical contact. A temperature probe was insertednear the air outlet. While in the chamber the micewere not allowed to eat or drink but were not fooddeprived prior to entry. All the measurements weremade at a thermoneutral temperature of 30 °C (lowercritical temperature in these mice is 28 °C).Measurements wre made at thermoneutrality toeliminate any physical thermal effects of huddling inthe paired mice. Any observed suppression ofmetabolism could thus be attributed to social or phys-iological factors. In experiment 3 two separate cham-bers linked in series were used. Barometric pressurereadings were taken at the start of measurements andthe temperature inside the flowmeter monitoredthroughout the run. All oxygen consumption esti-mates were corrected to STP.
In all three experiments observations were made onsingle sex groups of five individuals. It had originallybeen planned to examine the role of factors such asgender on the extent of any socio-physiological sup-pression. However, it was found that whenever twomice of opposite sex were placed together they almostalways immediately fought or copulated – both ofwhich compromised the estimation of a restingmetabolic rate. In addition once females became preg-nant they changed their metabolism over time, thusalso compromising the experimental design whichinvolved sequential pairings of individuals over aperiod of days. After these observations the designwas restricted to single sex groups of females. Theanimals in each group of five were exhaustivelypaired together with all other members of their group(n = 10 pairings). In experiments 1 and 2 for each pair-ing measurements were made of metabolic rate (oxy-gen consumption) in four different conditions: (1)Mouse A alone; (2) Mouse B alone; (3) Mice A and Bboth in the chamber together with no divide; andfinally (4) Mice A and B both in the chamber but sepa-rated by the divide. As patterns of circadian and ultra-dian rhythms in metabolism have been observed in anumber of small laboratory animals (Lehmann 1976;Kleinknecht, Erkert & Nelson 1985; Stupfel et al.1987) including mice (Mount & Willmott 1967) theorder of the conditions was randomized. In conditions(1) and (2) the partition was present so that the animalwas allowed to stay only in one half of the respirome-try chamber; the side of the chamber used in each casewas randomly decided. In condition (4) allocation ofthe animals to each side of the divide was randomized.Between each condition the chamber was cleanedthoroughly and the sawdust changed.
Measurements were made in each condition for90 min. The first 30 min was considered a settlingperiod. Behavioural recordings were made in the last60 min, after which the animals were taken out of thechamber and the analysers allowed to stabilize atambient levels for 30 min before the start of the nextmeasurement. During the 60 min of behaviour record-ing the mice were observed continuously and their
behaviour logged at the end of each minute. Behaviourwas divided into three classes: resting, grooming andgeneral activity. When two mice were placed togetherin a chamber, without the partition, allogroomingwould occur; this was included in the grooming cate-gory. General activity included walking, digging andchewing the sawdust, investigating the chamber and,when the mice were placed together, any social inter-actions that might occur apart from allogrooming. Thelocations of each individual, and thus their distancesapart, were also recorded each minute. When the ani-mals were together in the unpartitioned chamber, oneindividual was marked by touching it briefly on theback with a water-soluble marker pen, prior to the startof the measurements. This had no apparent effect onbehaviour (no difference between marked andunmarked individuals in their time budgets).
EXPERIMENT 1: LOW FLOW RATE
In this experiment 10 mice were used. They were ran-domly divided into two groups of five and exhaus-tively paired within their own group. Each pairing wasused for 1 day’s measurements. The air flow used inthis experiment averaged 602 ml min–1. The flow wasnot varied within each condition, and thus the ambientCO2 levels were greater during the period when twomice were in the chamber compared with only singlemice. CO2 content was not monitored directlythroughout these experiments as a CO2 analyser wasnot available at the time the measurements weremade. In a separate group of the same strain of micefeeding on the same diet the respiratory quotient(RQ = CO2 production divided by O2 consumption)was measured using in-line CO2 and oxygen analy-sers. Steady-state RQ averaged 0·953 (SD = 0·04,range 0·91–0·99, n = 7). Thus for this strain of micefeeding on this diet the CO2 concentrations in thechambers would be ≈ 95% of the observed percentagereduction in oxygen concentration.
Given our determinations of RQ and the flow rateused in this experiment the CO2 content of the cham-ber was on average 0·22% when single mice were pre-sent and 0·44% when two mice were present. ThusCO2 content was always higher than 0·2%, and alwaysin the range shown previously by Schlenker &Herreid (1981) to produce suppression. Since at ther-moneutrality the possibility of any physical benefitsof sociality can be discounted, any suppression ofmetabolism in the paired conditions could thus beattributed to a social ‘calming’ effect, and any differ-ence between conditions (3) and (4) might point tohow this effect was mediated.
Two quantifications of metabolic rate were made.First, the mean oxygen consumption was measuredover the same 60 min for which behavioural observa-tions were made. Second, the RMR was measured asthe lowest consecutive 5 min of oxygen consumptionthroughout the entire run.
375Socio-physiologicaleffect onmetabolism
© 1999 BritishEcological Society,Functional Ecology,13, 373–382

EXPERIMENT 2: HIGH FLOW RATE
The protocol for the second experiment was identicalto that of the first, but a new group of 10 mice wasused, and the average flow of air though the systemwas increased to 1113 ml min–1. The increase in airflow between this experiment and the previous onewas estimated to be enough to eliminate any possibleself-inhibition of metabolic rate for the solitary mice.In the solitary conditions the ambient CO2 level wasestimated to be 0·12% but in the paired conditions itwas estimated to average 0·24%. These levels there-fore crossed the boundary at which Schlenker &Herreid (1981) suggested that suppression becomesevident (0·2%).
It was anticipated that if suppression due to CO2
occurs this would be detectable in two ways. Firstthere would be a reduction in the metabolism of thepaired individuals relative to the sum of their solitarymetabolic rates, the extent of which would be inde-pendent of the presence or absence of the partition.Second, the metabolic rates of the mice in this experi-ment would be expected to be higher than themetabolic rates of mice in the first experiment underidentical social conditions.
EXPERIMENT 3: SEPARATE CHAMBERS LINKED IN
SERIES
In the third experiment two separate cylindrical cham-bers (A and B) were used, each consisting of a Perspextube (30 cm long, 7 cm in diameter) closed at eitherend by a rubber bung. In each chamber a perforatedplastic floor (6·5 cm× 25 cm) was fitted to keep theanimals separate from their faeces and urine. Air inletand outlet were at opposite ends of the chamber and atemperature probe was inserted near the outlet. Thechambers were connected in series and the tube con-necting them could be substituted with a Perspex tubefilled with sodium hydroxide pellets and silica gel toremove the CO2 produced by the mouse in the firstchamber and prevent it from reaching the mouse in thesecond chamber.
In this experiment a further 10 mice were used, andthey were divided into two groups as in experiments 1and 2, but the conditions they were exposed to weredifferent:
1. Mouse in chamber A (upstream), with CO2
absorber between chambers.2. Mouse in chamber B (downstream), with CO2
absorber between chambers.3. Mice in chambers A and B, with CO2 absorber
between chambers.4. Mice in chambers A and B, without CO2 absorber.
The rationale for this protocol was to investigate thephysiological effect of CO2 alone, taking out anyeffects of social factors by keeping the two micecompletely isolated with respect to social contact.
Each mouse was randomly assigned to either chamberA or B, and was placed only in that chamber for thatday’s recordings. As in previous experiments, theorder of conditions was randomized. The same defini-tions were used for mean oxygen consumption andRMR as employed in the previous two experiments.
STATISTICS
In all three experiments a comparison was madebetween the summed oxygen consumption of twomice measured solitarily (generally termed the pre-dicted summed oxygen consumption) and that mea-sured when they were together. The animals wereassigned to groups of five individuals which wereexhaustively paired together (giving a sample size of10 comparisons). Since in all three experiments therewere two groups of five individuals, there was a totalof 20 comparisons between the summed predictionand the observed oxygen consumption of paired indi-viduals for each experiment. The means were com-pared across these 20 comparisons using pairedt-tests. The null hypothesis in each case was that therewould be no difference between the summed mea-surements and the measurements of the animalstogether. Although a priori we were interested in sup-pression, two-tailed tests were made because our nullhypothesis was that there was no effect, and we didnot want to ignore the possibility that paired micemight mutually stimulate each other’s metabolic rates.As several repeated tests were made within eachexperiment, the significance levels were adjusted forthe number of comparisons being made using theBonferoni correction. Cited significance levels (0.05and 0.01) include the correction for repeated testing.
A more refined prediction was also generated whichcontrolled for changes in the activity of the animalsbetween the separated and paired conditions. To dothis the measurements of oxygen consumption andsimultaneous records of the behaviour of solitary indi-viduals were used. Body mass and percentage of thetime spent resting were entered as predictors of theoxygen consumption in a regression analysis. Theexpenditure of each individual in the paired conditionwas then predicted by substituting the known massand percentage time spent resting for that individualinto the equation, and summing across the two indi-viduals. The derived prediction was tested in the sameway as the simpler prediction which did not take intoaccount changes in the behaviour, i.e. using a paired t-test. The predictions were made both with, and with-out, arcsine square root transformations of thepercentages. This transformation made no differenceto the significance of the effects.
In addition to paired comparisons within each exper-iment the oxygen consumptions of mice betweenexperiments 1 and 2 were also compared. As differentmice had been employed in these experimentsmetabolic rates in each condition were compared by
376J. R. Speakman& F. P. Rossi
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
two-sample t-tests. For the solitary comparisons sin-gle observations of each individual from each groupwere chosen to avoid pseudo-replication. Again thenull hypothesis was that there was no difference andtwo-tailed tests were employed.
The significance of behavioural changes betweenconditions was explored using a χ2-test. The behaviourof the two individuals when solitary was combined toderive an expectation, and this was compared with theactual observed behaviours when the animals weretogether. All tests were performed using the MINITAB
statistics package (Ryan, Joiner & Ryan 1985).
Results
EXPERIMENTS 1 AND 2: LOW AND HIGH FLOW RATES
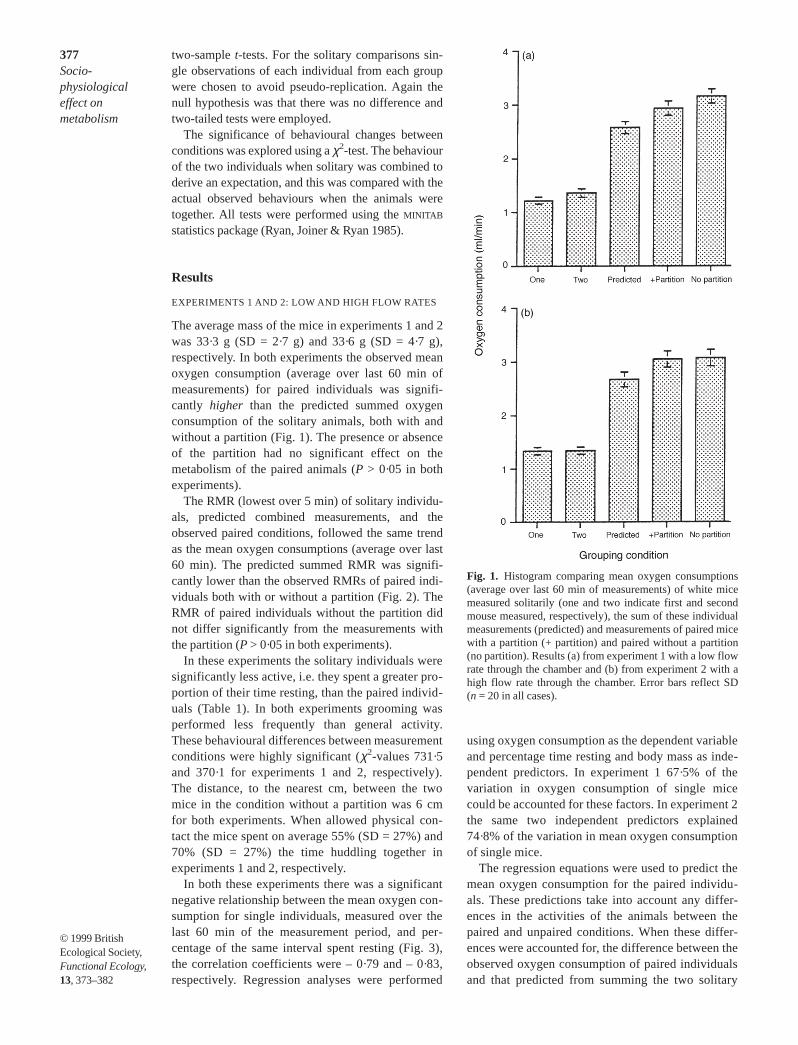
The average mass of the mice in experiments 1 and 2was 33·3 g (SD = 2·7 g) and 33·6 g (SD = 4·7 g),respectively. In both experiments the observed meanoxygen consumption (average over last 60 min ofmeasurements) for paired individuals was signifi-cantly higher than the predicted summed oxygenconsumption of the solitary animals, both with andwithout a partition (Fig. 1). The presence or absenceof the partition had no significant effect on themetabolism of the paired animals (P > 0·05 in bothexperiments).
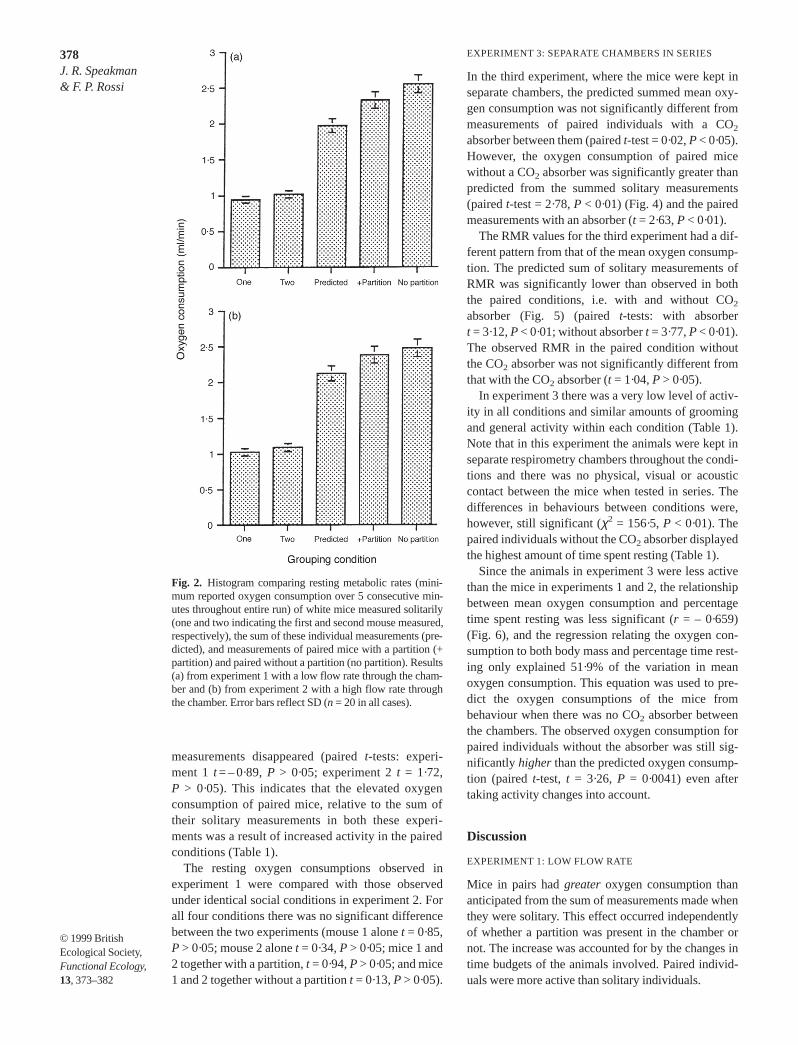
The RMR (lowest over 5 min) of solitary individu-als, predicted combined measurements, and theobserved paired conditions, followed the same trendas the mean oxygen consumptions (average over last60 min). The predicted summed RMR was signifi-cantly lower than the observed RMRs of paired indi-viduals both with or without a partition (Fig. 2). TheRMR of paired individuals without the partition didnot differ significantly from the measurements withthe partition (P > 0·05 in both experiments).
In these experiments the solitary individuals weresignificantly less active, i.e. they spent a greater pro-portion of their time resting, than the paired individ-uals (Table 1). In both experiments grooming wasperformed less frequently than general activity.These behavioural differences between measurementconditions were highly significant (χ2-values 731·5and 370·1 for experiments 1 and 2, respectively).The distance, to the nearest cm, between the twomice in the condition without a partition was 6 cmfor both experiments. When allowed physical con-tact the mice spent on average 55% (SD = 27%) and70% (SD = 27%) the time huddling together inexperiments 1 and 2, respectively.
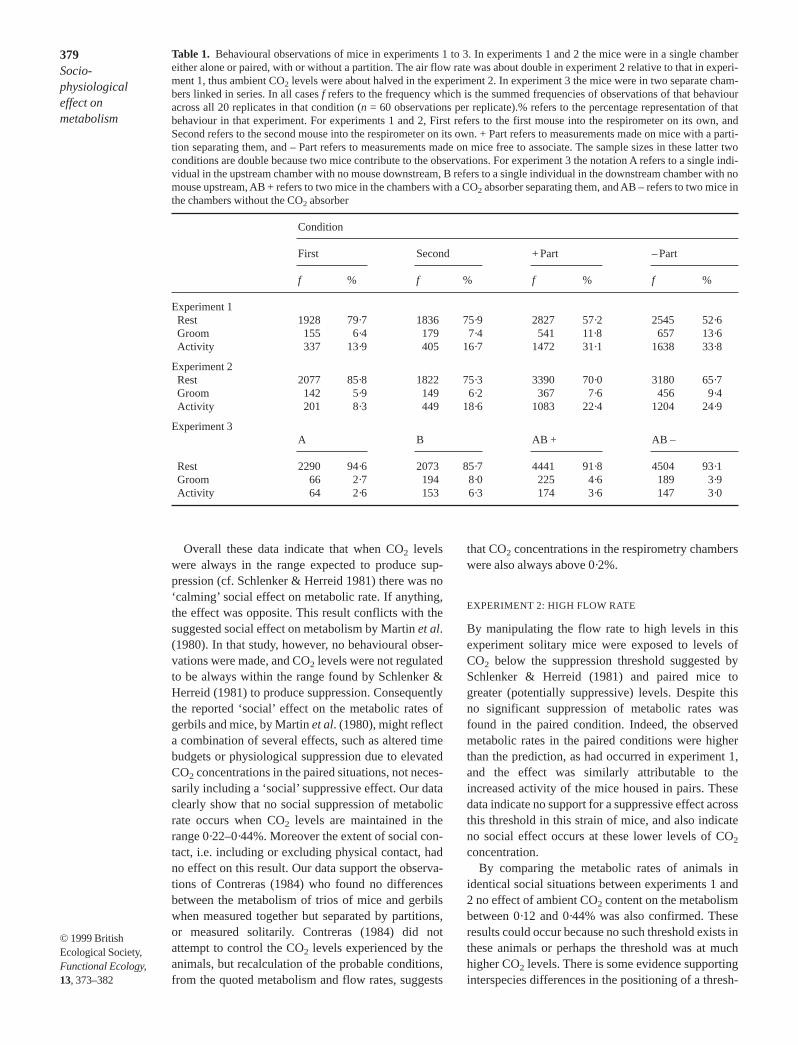
In both these experiments there was a significantnegative relationship between the mean oxygen con-sumption for single individuals, measured over thelast 60 min of the measurement period, and per-centage of the same interval spent resting (Fig. 3),the correlation coefficients were – 0·79 and – 0·83,respectively. Regression analyses were performed
using oxygen consumption as the dependent variableand percentage time resting and body mass as inde-pendent predictors. In experiment 1 67·5% of thevariation in oxygen consumption of single micecould be accounted for these factors. In experiment 2the same two independent predictors explained74·8% of the variation in mean oxygen consumptionof single mice.
The regression equations were used to predict themean oxygen consumption for the paired individu-als. These predictions take into account any differ-ences in the activities of the animals between thepaired and unpaired conditions. When these differ-ences were accounted for, the difference between theobserved oxygen consumption of paired individualsand that predicted from summing the two solitary
377Socio-physiologicaleffect onmetabolism
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
Fig. 1. Histogram comparing mean oxygen consumptions(average over last 60 min of measurements) of white micemeasured solitarily (one and two indicate first and secondmouse measured, respectively), the sum of these individualmeasurements (predicted) and measurements of paired micewith a partition (+ partition) and paired without a partition(no partition). Results (a) from experiment 1 with a low flowrate through the chamber and (b) from experiment 2 with ahigh flow rate through the chamber. Error bars reflect SD(n = 20 in all cases).
measurements disappeared (paired t-tests: experi-ment 1 t = – 0·89, P > 0·05; experiment 2 t = 1·72,P > 0·05). This indicates that the elevated oxygenconsumption of paired mice, relative to the sum oftheir solitary measurements in both these experi-ments was a result of increased activity in the pairedconditions (Table 1).
The resting oxygen consumptions observed inexperiment 1 were compared with those observedunder identical social conditions in experiment 2. Forall four conditions there was no significant differencebetween the two experiments (mouse 1 alone t = 0·85,P > 0·05; mouse 2 alone t = 0·34, P > 0·05; mice 1 and2 together with a partition, t = 0·94, P> 0·05; and mice1 and 2 together without a partition t = 0·13, P> 0·05).
EXPERIMENT 3: SEPARATE CHAMBERS IN SERIES
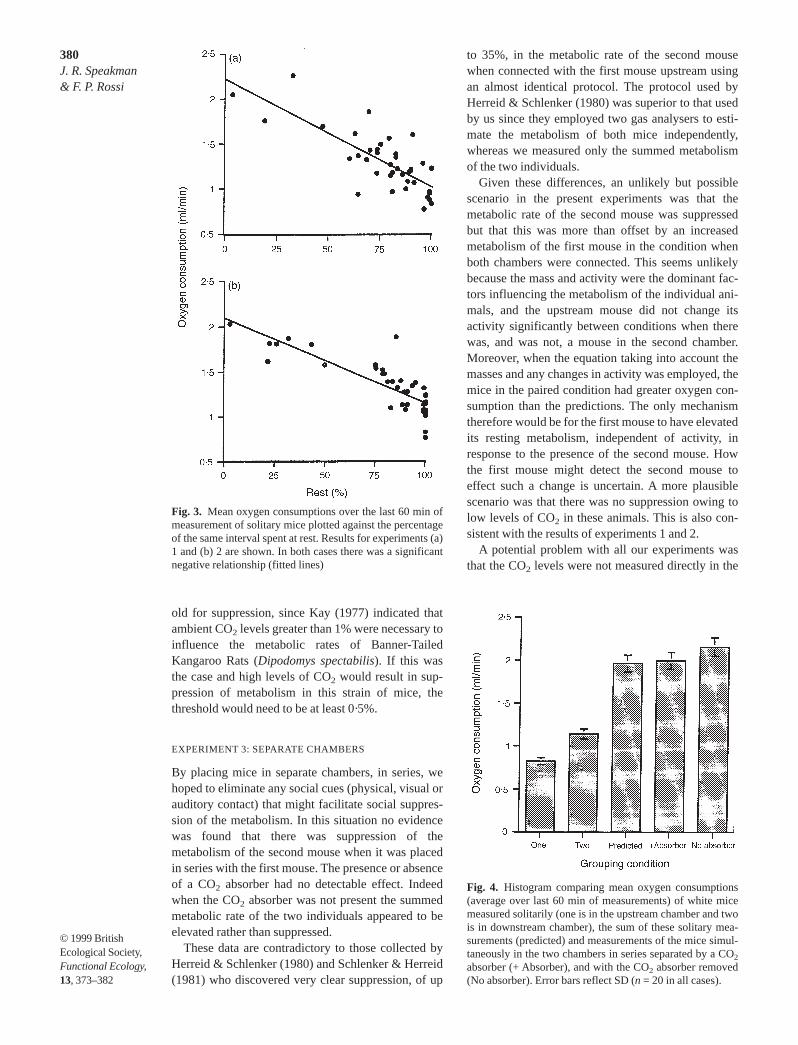
In the third experiment, where the mice were kept inseparate chambers, the predicted summed mean oxy-gen consumption was not significantly different frommeasurements of paired individuals with a CO2
absorber between them (paired t-test = 0·02, P< 0·05).However, the oxygen consumption of paired micewithout a CO2 absorber was significantly greater thanpredicted from the summed solitary measurements(paired t-test = 2·78, P < 0·01) (Fig. 4) and the pairedmeasurements with an absorber (t = 2·63, P < 0·01).
The RMR values for the third experiment had a dif-ferent pattern from that of the mean oxygen consump-tion. The predicted sum of solitary measurements ofRMR was significantly lower than observed in boththe paired conditions, i.e. with and without CO2
absorber (Fig. 5) (paired t-tests: with absorbert = 3·12, P< 0·01; without absorber t = 3·77, P< 0·01).The observed RMR in the paired condition withoutthe CO2 absorber was not significantly different fromthat with the CO2 absorber (t = 1·04, P > 0·05).
In experiment 3 there was a very low level of activ-ity in all conditions and similar amounts of groomingand general activity within each condition (Table 1).Note that in this experiment the animals were kept inseparate respirometry chambers throughout the condi-tions and there was no physical, visual or acousticcontact between the mice when tested in series. Thedifferences in behaviours between conditions were,however, still significant (χ2 = 156·5, P < 0·01). Thepaired individuals without the CO2 absorber displayedthe highest amount of time spent resting (Table 1).
Since the animals in experiment 3 were less activethan the mice in experiments 1 and 2, the relationshipbetween mean oxygen consumption and percentagetime spent resting was less significant (r = – 0·659)(Fig. 6), and the regression relating the oxygen con-sumption to both body mass and percentage time rest-ing only explained 51·9% of the variation in meanoxygen consumption. This equation was used to pre-dict the oxygen consumptions of the mice frombehaviour when there was no CO2 absorber betweenthe chambers. The observed oxygen consumption forpaired individuals without the absorber was still sig-nificantly higherthan the predicted oxygen consump-tion (paired t-test, t = 3·26, P = 0·0041) even aftertaking activity changes into account.
Discussion
EXPERIMENT 1: LOW FLOW RATE
Mice in pairs had greater oxygen consumption thananticipated from the sum of measurements made whenthey were solitary. This effect occurred independentlyof whether a partition was present in the chamber ornot. The increase was accounted for by the changes intime budgets of the animals involved. Paired individ-uals were more active than solitary individuals.
378J. R. Speakman& F. P. Rossi
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
Fig. 2. Histogram comparing resting metabolic rates (mini-mum reported oxygen consumption over 5 consecutive min-utes throughout entire run) of white mice measured solitarily(one and two indicating the first and second mouse measured,respectively), the sum of these individual measurements (pre-dicted), and measurements of paired mice with a partition (+partition) and paired without a partition (no partition). Results(a) from experiment 1 with a low flow rate through the cham-ber and (b) from experiment 2 with a high flow rate throughthe chamber. Error bars reflect SD (n = 20 in all cases).
Overall these data indicate that when CO2 levelswere always in the range expected to produce sup-pression (cf. Schlenker & Herreid 1981) there was no‘calming’ social effect on metabolic rate. If anything,the effect was opposite. This result conflicts with thesuggested social effect on metabolism by Martin et al.(1980). In that study, however, no behavioural obser-vations were made, and CO2 levels were not regulatedto be always within the range found by Schlenker &Herreid (1981) to produce suppression. Consequentlythe reported ‘social’ effect on the metabolic rates ofgerbils and mice, by Martin et al. (1980), might reflecta combination of several effects, such as altered timebudgets or physiological suppression due to elevatedCO2 concentrations in the paired situations, not neces-sarily including a ‘social’ suppressive effect. Our dataclearly show that no social suppression of metabolicrate occurs when CO2 levels are maintained in therange 0·22–0·44%. Moreover the extent of social con-tact, i.e. including or excluding physical contact, hadno effect on this result. Our data support the observa-tions of Contreras (1984) who found no differencesbetween the metabolism of trios of mice and gerbilswhen measured together but separated by partitions,or measured solitarily. Contreras (1984) did notattempt to control the CO2 levels experienced by theanimals, but recalculation of the probable conditions,from the quoted metabolism and flow rates, suggests
that CO2 concentrations in the respirometry chamberswere also always above 0·2%.
EXPERIMENT 2: HIGH FLOW RATE
By manipulating the flow rate to high levels in thisexperiment solitary mice were exposed to levels ofCO2 below the suppression threshold suggested bySchlenker & Herreid (1981) and paired mice togreater (potentially suppressive) levels. Despite thisno significant suppression of metabolic rates wasfound in the paired condition. Indeed, the observedmetabolic rates in the paired conditions were higherthan the prediction, as had occurred in experiment 1,and the effect was similarly attributable to theincreased activity of the mice housed in pairs. Thesedata indicate no support for a suppressive effect acrossthis threshold in this strain of mice, and also indicateno social effect occurs at these lower levels of CO2
concentration.By comparing the metabolic rates of animals in
identical social situations between experiments 1 and2 no effect of ambient CO2 content on the metabolismbetween 0·12 and 0·44% was also confirmed. Theseresults could occur because no such threshold exists inthese animals or perhaps the threshold was at muchhigher CO2 levels. There is some evidence supportinginterspecies differences in the positioning of a thresh-
379Socio-physiologicaleffect onmetabolism
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
Table 1. Behavioural observations of mice in experiments 1 to 3. In experiments 1 and 2 the mice were in a single chambereither alone or paired, with or without a partition. The air flow rate was about double in experiment 2 relative to that in experi-ment 1, thus ambient CO2 levels were about halved in the experiment 2. In experiment 3 the mice were in two separate cham-bers linked in series. In all cases f refers to the frequency which is the summed frequencies of observations of that behaviouracross all 20 replicates in that condition (n = 60 observations per replicate).% refers to the percentage representation of thatbehaviour in that experiment. For experiments 1 and 2, First refers to the first mouse into the respirometer on its own, andSecond refers to the second mouse into the respirometer on its own. + Part refers to measurements made on mice with a parti-tion separating them, and – Part refers to measurements made on mice free to associate. The sample sizes in these latter twoconditions are double because two mice contribute to the observations. For experiment 3 the notation A refers to a single indi-vidual in the upstream chamber with no mouse downstream, B refers to a single individual in the downstream chamber with nomouse upstream, AB + refers to two mice in the chambers with a CO2 absorber separating them, and AB – refers to two mice inthe chambers without the CO2 absorber
Condition
First Second + Part – Part
f % f % f % f %
Experiment 1Rest 1928 79·7 1836 75·9 2827 57·2 2545 52·6Groom 155 6·4 179 7·4 541 11·8 657 13·6Activity 337 13·9 405 16·7 1472 31·1 1638 33·8
Experiment 2Rest 2077 85·8 1822 75·3 3390 70·0 3180 65·7Groom 142 5·9 149 6·2 367 7·6 456 9·4Activity 201 8·3 449 18·6 1083 22·4 1204 24·9
Experiment 3A B AB + AB –
Rest 2290 94·6 2073 85·7 4441 91·8 4504 93·1Groom 66 2·7 194 8·0 225 4·6 189 3·9Activity 64 2·6 153 6·3 174 3·6 147 3·0
old for suppression, since Kay (1977) indicated thatambient CO2 levels greater than 1% were necessary toinfluence the metabolic rates of Banner-TailedKangaroo Rats (Dipodomys spectabilis). If this wasthe case and high levels of CO2 would result in sup-pression of metabolism in this strain of mice, thethreshold would need to be at least 0·5%.
EXPERIMENT 3: SEPARATE CHAMBERS
By placing mice in separate chambers, in series, wehoped to eliminate any social cues (physical, visual orauditory contact) that might facilitate social suppres-sion of the metabolism. In this situation no evidencewas found that there was suppression of themetabolism of the second mouse when it was placedin series with the first mouse. The presence or absenceof a CO2 absorber had no detectable effect. Indeedwhen the CO2 absorber was not present the summedmetabolic rate of the two individuals appeared to beelevated rather than suppressed.
These data are contradictory to those collected byHerreid & Schlenker (1980) and Schlenker & Herreid(1981) who discovered very clear suppression, of up
to 35%, in the metabolic rate of the second mousewhen connected with the first mouse upstream usingan almost identical protocol. The protocol used byHerreid & Schlenker (1980) was superior to that usedby us since they employed two gas analysers to esti-mate the metabolism of both mice independently,whereas we measured only the summed metabolismof the two individuals.
Given these differences, an unlikely but possiblescenario in the present experiments was that themetabolic rate of the second mouse was suppressedbut that this was more than offset by an increasedmetabolism of the first mouse in the condition whenboth chambers were connected. This seems unlikelybecause the mass and activity were the dominant fac-tors influencing the metabolism of the individual ani-mals, and the upstream mouse did not change itsactivity significantly between conditions when therewas, and was not, a mouse in the second chamber.Moreover, when the equation taking into account themasses and any changes in activity was employed, themice in the paired condition had greater oxygen con-sumption than the predictions. The only mechanismtherefore would be for the first mouse to have elevatedits resting metabolism, independent of activity, inresponse to the presence of the second mouse. Howthe first mouse might detect the second mouse toeffect such a change is uncertain. A more plausiblescenario was that there was no suppression owing tolow levels of CO2 in these animals. This is also con-sistent with the results of experiments 1 and 2.
A potential problem with all our experiments wasthat the CO2 levels were not measured directly in the
380J. R. Speakman& F. P. Rossi
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
Fig. 3. Mean oxygen consumptions over the last 60 min ofmeasurement of solitary mice plotted against the percentageof the same interval spent at rest. Results for experiments (a)1 and (b) 2 are shown. In both cases there was a significantnegative relationship (fitted lines)
Fig. 4. Histogram comparing mean oxygen consumptions(average over last 60 min of measurements) of white micemeasured solitarily (one is in the upstream chamber and twois in downstream chamber), the sum of these solitary mea-surements (predicted) and measurements of the mice simul-taneously in the two chambers in series separated by a CO2
absorber (+ Absorber), and with the CO2 absorber removed(No absorber). Error bars reflect SD (n = 20 in all cases).

chambers directly but were inferred from a combina-tion of the decline in oxygen percentage levels and theRQ observed in a separate group of animals. In a dif-ferent series of RQ measurements made 10 years pre-viously, using the same strain of mice but a differentcommercial diet, lower RQs of around 0·8 wereobserved (J. R. Speakman, unpublished data). If theRQ in the present experiments had been only 0·8 thiswould change the interpretation of individual experi-ments. In experiment 1, solitary mice would have beenbelow the supposed threshold eliciting suppression
(0·2%) but paired mice would have been above thethreshold. In experiment 2 the mice would havealways been below the threshold. In both cases theabsence of any effect of pairing on metabolic ratesstill suggests that neither social factors nor low levelsof ambient CO2 are suppressive factors. Our interpre-tations are therefore robust to the inferred RQ.
The most likely explanation for the discrepancybetween the current results and those of Herreid &Schlenker (1980) and Schlenker & Herreid (1981) isthat the effect detected by them was specific to thestrain of mice they studied (RR mice). Unfortunately,we could not obtain this strain of mice to test this pos-sibility. Another difference between our experimentsand some of their protocols was that our mice werealways housed alone but in some of their experimentsthe mice were housed together. The extent to whichprior sociality affects the extent of suppression andhow widespread the effect they detected is remainsopen to further investigation.
Conclusions
Across all these experiments, in which the social con-ditions and the CO2 environment were manipulated,no evidence was found for any significant suppressionof the metabolic rates of individuals in pairs, comparedwith when they were solitary. Where differences inoxygen consumption did occur they were in the direc-tion opposite to that expected from suppression, andwere linked to differences in time budgets of the ani-mals between the different conditions. Removing theeffects of the differences in time budgets generallyremoved these effects. We find no support for thenotions of social or physiological suppression effectson the metabolic rates of these animals.
Acknowledgements
We are grateful to Prof C.F. Herreid for searching forand providing details of suppliers for the RR mice.This work was supported by NERC grant GR3/5155.
References
Andrews, R.V. & Belknap, R.W. (1986) Bioenergetic bene-fits of huddling by deer mice (Peromyscus maniculatus).Comparative Biochemistry and Physiology85A,775–778.
Andrews, R.V., Phillips, D. & Makihara, D. (1987)Metabolic and thermoregulatory consequences of socialbehaviours between Microtus townsendii. ComparativeBiochemistry and Physiology87A, 345–348.
Baudinette, R.V. (1972) The impact of social aggregation onthe respiratory physiology of Australian hopping mice.Comparative Biochemistry and Physiology41A, 35–38.
Bryant, D.M. & Hails, C.J. (1975) Mechanisms of heat con-servation in the litters of mice (Mus musculus L.).Comparative Biochemistry and Physiology50A, 99–104.
Canals, M., Rosenmann, M. & Bozinovic, F. (1989)Energetics and geometry of huddling in small mammals.Journal of Theoretical Biology141,181–189.
381Socio-physiologicaleffect onmetabolism
© 1999 BritishEcological Society,Functional Ecology,13, 373–382
Fig. 5. Histogram comparing resting metabolic rates (mini-mum reported oxygen consumption over 5 consecutive min-utes throughout entire run) of white mice measured solitarily(one is in upstream chamber and two is in downstreamchamber), the sum of these solitary measurements (pre-dicted) and measurements of the mice simultaneously in thetwo chambers in series separated by a CO2 absorber (+Absorber), and with the CO2 absorber removed (Noabsorber). Error bars reflect SD (n = 20 in all cases).
Fig. 6. Mean oxygen consumption over the last 60 min ofmeasurement of solitary mice plotted against the percentageof the same interval spent at rest for animals in experiment 3.There was a significant negative relationship (fitted line).

Canals, M., Rosenmann, M. & Bozinovic, F. (1997)Geometric aspects of the energetic effectiveness of hud-dling in small mammals.Acta Theriologica42,321–328.
Contreras, L.C. (1984) Bioenergetics of huddling: test of apsycho-physiological hypothesis.Journal of Mammalogy65,256–262.
Fedyk, A. (1971) Social thermoregulation in Apodemus flav-icollis (Melchior, 1834).Acta Theriologica16,221–229.
Gebczynska, Z. & Gebczynski, M. (1971) Insulating proper-ties of the nest: social temperature regulation inClethrionomys glareolus(Schreber).Annales ZoologicaFennici8, 104–108.
Hayes, J.P., Speakman, J.R. & Racey, P.A. (1992a) The con-tributions of local heating and reducing exposed surfacearea to the energetic benefits of huddling by short-tailedfield voles (Microtus agrestis). Physiological Zoology65,742–762.
Hayes, J.P., Speakman, J.R. & Racey, P.A. (1992b)Sampling bias in respirometry.Physiological Zoology65,604–619.
Herreid, C.F. & Schlenker, E.H. (1980) Energetics of mice instable and unstable social conditions: evidence of an air-borne factor affecting metabolism.Animal Behaviour28,20–28.
Karasov, W.H. (1983) Wintertime energy conservation byhuddling in antelope ground squirrels (Ammospermophilusleucurus). Journal of Mammalogy64,341–345.
Kay, F.R. (1977) Environmental physiology of the banner-tailed kangaroo rat – II. Influences of the burrow environ-ment on metabolism and water loss.ComparativeBiochemistry and Physiology57A, 471–477.
Kleinknecht, S., Erkert, H.G. & Nelson, J.E. (1985)Circadian and ultradian rhythms of activity and O2-con-sumption in three nocturnal Marsupialian species:Petaurus breviceps, Phalangeridae; Dasyuroides byrnei,Dasyuridae; Monodelphis domestica Didelphidae.Zeitschrift für Saugertierkunde50,321–329.
Lehmann, U. (1976) Short-term and circadian rhythms in thebehavior of the vole,Microtus agrestis(L.). Oecologia23,185–199.
Martin, R.A., Fiorentini, M. & Connors, F. (1980) Social
facilitation of reduced oxygen consumption in Musmusculusand Meriones unguiculatus. Comparative Bio-chemistry and Physiology65A,519–522.
Mount, L.E. & Willmott, J.V. (1967) The relation betweenspontaneous activity, metabolic rate and the 24 hour cyclein mice at different environmental temperatures.Journalof Physiology190,371–380.
Nagy, K.A. (1989) Field bioenergetics: accuracy of modelsand methods.Physiological Zoology62,237–252.
Ryan, B.F., Joiner, B.L. & Ryan, T.A. Jr (1985) MinitabHandbook. PWS-Kent Publishing Company, Boston, MA.
Schlenker, E.H. & Herreid, C.F. (1981) The effect of low lev-els of carbon dioxide on metabolism of Mus musculus.Comparative Biochemistry and Physiology68A,673–676.
Schlenker, E.H., Carlson, D. & Herreid, C.F. (1981)Metabolic contribution of CO2 to the ‘group effect’.American Journal of Physiology241,R264–R266.
Sealander, J.A. Jr (1952) The relationship of nest protectionand huddling to survival of Peromyscusat low tempera-ture.Ecology33,63–71.
Speakman, J.R. & Racey, P.A. (1988) Validation of the dou-bly labeled water technique in small insectivorous nats bycomparison with indirect calorimetry.PhysiologicalZoology61,514–526.
Springer, S.D., Gregory, P.A. & Barrett, G.W. (1981)Importance of social grouping on bioenergetics of thegolden mouse, Ochrotomys nuttalli. Journal ofMammalogy62,628–630.
Stupfel, M., Gourlet, V., Court, L., Mestries, J., Perramon, A.& Merat, P. (1987) Periodic analysis of ultradian(40 min < tau < 24 h) respiratory variations in laboratoryvertebrates of various circadian activities.Chronobiologia14,365–375.
Vickery, W.L. & Millar, J.S. (1984) The energetics ofhuddling by endotherms.Oikos43,88–93.
Weathers, W.W., Buttemer, W.A., Hayworth, A.M. & Nagy,K.A. (1984) An evaluation of time-energy budget estimatesof daily energy expenditure in birds.Auk101,459–472.
Received 10 November 1997; revised 2 October 1998;accepted 15 October 1998
382J. R. Speakman& F. P. Rossi
© 1999 BritishEcological Society,Functional Ecology,13, 373–382