nad + auxotrophy is bacteriocidal for the tubercle bacilli
TRANSCRIPT

NAD+ Auxotrophy is Bacteriocidal for the Tubercle Bacilli
Catherine Vilchèze, Brian Weinrick, Ka-Wing Wong, Bing Chen, and William R. Jacobs Jr.*Howard Hughes Medical Institute, Department of Microbiology and Immunology, Albert EinsteinCollege of Medicine, Bronx, NY 10461, USA
SUMMARYThe human tubercle bacillus Mycobacterium tuberculosis can synthesize NAD+ using the de novobiosynthesis pathway or the salvage pathway. The salvage pathway of the bovine tubercle bacillusM. bovis was reported defective due to a mutation in the nicotinamidase PncA. This defectprevents nicotinic acid secretion, which is the basis for the niacin test that clinically distinguishesM. bovis from M. tuberculosis. Surprisingly, we found that the NAD+ de novo biosynthesispathway (nadABC) can be deleted from M. bovis, demonstrating a functioning salvage pathway.M. bovis ΔnadABC fails to grow in mice, whereas M. tuberculosis ΔnadABC grows normally inmice, suggesting that M. tuberculosis can acquire nicotinamide from its host. The introduction ofM. tuberculosis pncA into M. bovis ΔnadABC is sufficient to fully restore growth in a mouse,proving that the functional salvage pathway enables nicotinamide acquisition by the tuberclebacilli. This study demonstrates that NAD+ starvation is a cidal event in the tubercle bacilli andconfirms that enzymes common to the de novo and salvage pathways may be good drug targets.
Keywordsbovis; tuberculosis; NAD+; starvation; pncA
INTRODUCTIONThe rapid spread of tuberculosis (TB) strains that are multi-drug- and extensively drug-resistant has rendered crucial the need to develop new drugs to fight TB, a disease caused bythe bacillus Mycobacterium tuberculosis. Novel drug targets have to be identified in order todiscover drugs that will be effective against these drug-resistant TB strains.
Nicotinamide adenine dinucleotide (NAD+) is an essential molecule implicated in hundredsof biological reactions. NAD+ is a hydride acceptor in redox reactions and a substrate inreactions involving ADP-ribose (Khan et al., 2007). The enzymes implicated in theeukaryotic NAD+ biosynthesis have been shown to be good drug targets for several diseases,such as autoimmune diseases, cancer, cardiac diseases, and multiple sclerosis (Khan et al.,2007; Sauve, 2008). The de novo NAD+ biosynthesis pathway differs between eukaryotesand prokaryotes. The eukaryotes synthesize NAD+ using the kyurenate pathway, whichrequires eight enzymes to obtain NAD+ from tryptophan. In prokaryotes, the de novobiosynthesis pathway synthesizes NAD+ from aspartate using five enzymes (Fig. 1A). Thefirst step is the oxidation of the amino group of L-aspartate to an imine by the aspartateoxidase NadB. The quinolinic acid synthase NadA then condenses the resultingiminoaspartic acid with dihydroxyacetone phosphate to form quinolinic acid.
* Corresponding author. Phone: (718) 678-1075, Fax: (718) 678-1085, [email protected] .Online supplemental material. Tables S1 to S4 represent the genes up- or down-regulated 2-fold or more by NAm starvation in NAmauxotrophs of M. tuberculosis and M. bovis.
NIH Public AccessAuthor ManuscriptMol Microbiol. Author manuscript; available in PMC 2011 April 1.
Published in final edited form as:Mol Microbiol. 2010 April ; 76(2): 365–377. doi:10.1111/j.1365-2958.2010.07099.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Monodecarboxylation of quinolinic acid and conversion to the nucleotide form by thequinolinate phosphoribosyl transferase NadC yields nicotinic acid mononucleotide(NAMN). The NAMN adenyl transferase NadD catalyzes the adenylation of NAMN tonicotinic acid adenine dinucleotide (NAAD), which is amidated by the deamido-NAD ligaseNadE to form NAD+. The last 2 enzymes of the de novo pathway, NadD and NadE, are alsocommon to the NAD+ salvage or recycling pathway. The salvage of NAD+ can occur via thedegradation of NAD+ to nicotinamide (NAm) by the NAD+ glycohydrolase (Gholson,1966), followed by the hydrolysis of NAm to nicotinic acid (NAc) by the nicotinamidasePncA, and then the transfer of a phosphoribosyl group to NAc by PncB to give NAMN, theproduct of NadC. NAMN is the convergence point of the salvage and the de novo pathways(Fig. 1A). The synthesis of NAD+ from NAc is also referred to as the Preiss-Handlerpathway. Another branch of the salvage pathway found in bacteria is the formation ofnicotinamide mononucleotide (NMN), either by cleavage of the adenyl monophosphate(AMP) group of NAD+ or by the transfer of a phosphoribosyl unit to NAm. NMN can thenbe either degraded to NAm via glycohydrolase or retransformed into NAD+ by transfer of anadenyl group. Mycobacteria can synthesize NAD+ by using either the de novo biosynthesispathway or the salvage pathway (Begley et al., 2001; Foster and Moat, 1980).
Due to the presence of the NAD+ salvage pathway, the only enzymes in the NAD+
biosynthesis pathway that could be considered as drug targets are NadD and NadE (Gerdeset al., 2002). Inhibitors of the NadD enzyme of Escherichia coli and Bacillus anthracis havebeen identified that have no activity against the corresponding human enzyme (Sorci et al.,2009). NadE inhibitors with good antibacterial activities have been developed againstBacillus subtilis and Staphylococcus aureus (Velu et al., 2003; Velu et al., 2007). Some ofthese NadE inhibitors were tested against M. tuberculosis and exhibited bacteriocidalactivity but also high cytotoxicity (Boshoff et al., 2008). In this study, Boshoff et al.demonstrated that deletion of the first two genes of the NAD+ biosynthesis pathway, nadAand nadB, had no effect on the growth of M. tuberculosis in vivo (Boshoff et al., 2008). Thisresult was attributed to the ability of M. tuberculosis to scavenge NAm or NAc from the hostand use the salvage pathway to maintain its redox balance. Indeed, it has been reported thatmammalian tissues contain large amounts of NAm (Qin et al., 2006; Yang and Sauve,2006). Interestingly, the pathogen responsible for the bovine TB, M. bovis, is known to havea defective salvage pathway due to a mutation in the pncA gene, a single-base pair change(c169g) resulting in the amino acid change His57Asp (Scorpio and Zhang, 1996). Thisdefect is the basis of the resistance of M. bovis to the TB drug pyrazinamide (PZA) as wellas of the negative niacin test, which is one of the tests employed clinically to differentiate M.tuberculosis from M. bovis infections in patients (Konno, 1956; Konno et al., 1957; Konnoet al., 1958). The niacin production test is based on the accumulation of niacin in M.tuberculosis culture fluids, which does not occur with M. bovis. This was first observed byPope and Smith in 1946 (Pope and Smith, 1946), and later was attributed to a higher level ofNAD+ glycohydrolase activity in M. tuberculosis than in M. bovis (Kasarov and Moat,1972). Kasarov and Moat also demonstrated that the source of NAc in M. tuberculosis wasNAD+ (Kasarov and Moat, 1972). To test the role of the salvage pathway in the survival invivo of mycobacteria lacking the ability to synthesize NAD+ de novo, we attempted thedeletion of the NAD+ de novo pathway (nadABC) in M. bovis as well as in M. tuberculosis.
Here we report the surprising result that deletion of the de novo NAD+ biosynthetic genes ispossible in M. bovis despite the presumed lack of a functioning salvage pathway and that, incontrast to M. tuberculosis, NAm starvation is a cidal event in M. bovis ΔnadABC. Thisleads to the conclusion that a combination of drugs targeting both the de novo and thesalvage pathways could also efficiently kill M. tuberculosis.
Vilchèze et al. Page 2
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

RESULTSNAD+ de novo genes can be deleted in M. bovis, demonstrating a functional salvagepathway
The three genes specific to the de novo NAD+ biosynthesis pathway, nadA, nadB, and nadC,are in an operon in M. tuberculosis and in M. bovis. This 3490 bp operon is quite similar inM. tuberculosis and in M. bovis, with only a 1 bp difference in the nadC gene (a190g),which causes an N64D amino acid change. The specialized transduction system (Bardarov etal., 2002) was used to replace the nadABC operon in both M. tuberculosis CDC1551 and M.bovis Ravenel by a hygromycin resistance gene (hyg) (Fig. 1B). As the left- and right-flanking regions of the nadABC operon are quite similar with 2 bp differences that do notcause any amino acid changes, the same knockout phage was used to perform thetransductions in both M. tuberculosis and M. bovis.
Since the salvage pathway is functional in M. tuberculosis (Boshoff et al., 2008), we platedour transductions on NAm-containing plates to allow for growth of the M. tuberculosisΔnadABC transductants. The transductions with M. bovis were plated on plates containingeither NAm or NAc. We did not expect to obtain transductants on NAm-containing platessince the nicotinamidase PncA has been shown to be defective in M. bovis (Scorpio andZhang, 1996). Surprisingly, M. bovis ΔnadABC transductants were obtained on NAm-containing plates at a low frequency (<10−8). Southern analyses on two M. tuberculosisΔnadABC transductants and two M. bovis ΔnadABC transductants isolated on NAm-containing plates confirmed that the nadABC genes had been replaced with the hyg gene(Fig. 1B). These transductants were used for the rest of the study. To ensure that the M.bovis ΔnadABC transductants had not acquired compensatory mutations in pncA, the geneand promoter region of pncA in M. bovis ΔnadABC were sequenced and compared to thesequence of wt M. bovis. We found no other mutation than the described pncA mutation(c169g) in M. bovis ΔnadABC. The M. bovis ΔnadABC deletion mutants, like the M.tuberculosis ΔnadABC deletion mutants, were found to be NAm auxotrophs, suggesting thatthe nicotinamidase and the Preiss-Handler pathway are active in M. bovis. To confirm thatM. bovis ΔnadABC (mc24957) can utilize NAm to synthesize NAD+, we added radioactiveNAm to a culture of M. bovis ΔnadABC and analyzed the radioactive metabolites by HPLC.M. bovis ΔnadABC was able to synthesize NAD+ from NAm. The predominant metabolitesobtained were NAD+, NADH, and NADP+, with no trace of radioactive NAm (Fig. 2). Wetherefore conclude that the salvage pathway is indeed functional in M. bovis.
M. bovis ΔnadABC requires more NAm for growth than M. tuberculosis ΔnadABCMycobacteria have genes for both the de novo pathway and the salvage pathway tosynthesize NAD+ (Fig. 1A). Following deletion of the nadABC genes from the de novopathway, we observed that M. bovis ΔnadABC could synthesize NAD+ from NAm. Toestablish whether M. bovis ΔnadABC was as efficient as M. tuberculosis ΔnadABC in usingNAm, we determined the minimum amount of NAm required for the growth of M. bovisΔnadABC in liquid cultures. The strains were grown in media containing 20 mg/l of NAm,spun down, and washed extensively with PBS-tyloxapol to remove any trace of NAm. Thestrains were then inoculated in media containing different amounts of NAm (from 0.01 mg/lto 20 mg/l). No significant difference in growth rate was observed when the ΔnadABCmutants were grown in the presence of NAm at concentrations ranging from 1 - 20 mg/l butthe M. tuberculosis ΔnadABC and M. bovis ΔnadABC strains showed a difference in theirability to utilize low concentrations of NAm (<1 mg/l) to grow (Fig. 3A). M. bovisΔnadABC needed more than 10 times the amount of NAm to maintain growth than did M.tuberculosis ΔnadABC. This reflects the known mutation in the M. bovis pncA gene(Scorpio and Zhang, 1996). Recently, this mutation was shown to result in a 99% loss of
Vilchèze et al. Page 3
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

pyrazinamidase/nicotinamidase activity (Zhang et al., 2008). In contrast, no growthdifferences between M. bovis ΔnadABC and M. tuberculosis ΔnadABC were observed whenthe mutants were supplemented with NAc. The smallest amount of NAc that allowed for thegrowth of the ΔnadABC strains was 0.1 mg/l (0.8 μM). Complementation of M. bovisΔnadABC with the M. bovis nadABC operon cloned downstream of the hsp60 promoter ofan integrative E. coli mycobacterial shuttle plasmid (Stover et al., 1991) resulted in a strainthat grew without any supplement and at a rate similar to that of wild-type M. bovis Ravenel(Fig. 3A).
NAD+ starvation is a cidal event in M. bovis ΔnadABCStarvation for different nutrients can be bacteriostatic (such as the leucine auxotroph, F. C.Bange and W. R. Jacobs, Jr., unpublished) or bacteriocidal (Gordhan et al., 2002). Todetermine if NAD+ starvation was a static or a cidal event in the ΔnadABC strains, wemeasured colony forming units (CFUs) over a period of 3 weeks while the strains werestarved for NAm (Fig. 3B). NAD+ starvation of the M. bovis ΔnadABC strain was a cidalevent, resulting in a 5 - 6 log drop in CFUs after 3 weeks. In contrast, no significant loss inviability was recorded for M. tuberculosis ΔnadABC for the first 2 weeks of starvation. Onlyin the following week did we observe a slight decrease in CFUs (1 log). The difference inthe death kinetics between M. tuberculosis ΔnadABC and M. bovis ΔnadABC during NAmstarvation could be due to the ability of M. tuberculosis to uptake secreted NAc from themedia, which could then be re-used to synthesize NAD+ via the Preiss-Handler pathway(Fig. 1A). The niacin test for M. bovis ΔnadABC is negative, indicating that the strain mightnot produce and secrete NAc at a level sufficient for re-utilization. To test this hypothesis,we mixed cultures of M. tuberculosis ΔnadABC and M. bovis ΔnadABC pMV361 andstarved them for NAm. The starved cultures were plated on plates containing or notkanamycin (the marker in the empty vector pMV361) to allow for the distinction betweenM. tuberculosis ΔnadABC and M. bovis ΔnadABC. As observed with M. tuberculosisΔnadABC, the starvation of M. bovis ΔnadABC pMV361 mixed with M. tuberculosisΔnadABC was bacteriostatic (no loss or gain in CFUs) for the first 2 weeks while starvationof M. bovis ΔnadABC pMV361 was cidal. This result suggests that the difference in thedeath rates between the two mutants arises from the processing of internal NAD+ pools.
Transformation with the pncA gene from M. tuberculosis reverses the bacteriocidalphenotype of M. bovis ΔnadABC upon NAm starvation
The M. bovis ΔnadABC strain displays two phenotypes different from M. tuberculosisΔnadABC: increased requirements for NAm and bacteriocidal killing in its absence. To testif these phenotypes were solely caused by the known mutation in M. bovis pncA, the M.tuberculosis pncA gene was cloned into the replicative plasmid pMV261 under the hsp60promoter and transformed into M. bovis ΔnadABC.
The introduction of M. tuberculosis pncA into M. bovis ΔnadABC resulted in a strain thatwas able to grow on a lower amount of NAm than was the parent strain (Fig. 4A). The M.bovis ΔnadABC pMV261::pncATB requirement for NAm was similar to that of M.tuberculosis ΔnadABC. Addition of radioactive NAm to M. bovis ΔnadABCpMV261::pncATB culture led to the same formation of radioactive NAD+ and NADP+, withno trace of NAm or NAc as M. bovis ΔnadABC, M. bovis ΔnadABC complemented, and M.tuberculosis ΔnadABC (Fig. 2). As expected, the M. bovis ΔnadABC pMV261::pncATB
strain also regained one of the features that differentiate M. tuberculosis and M. bovis: apositive niacin test. Furthermore, in contrast with the parent strain M. bovis ΔnadABC, NAmstarvation of M. bovis ΔnadABC overexpressing M. tuberculosis pncA was not abacteriocidal event (Fig. 4B). We observed a slight increase in CFUs after 7 days of NAmstarvation followed by a very slow decrease in CFUs in the subsequent 2 weeks, comparable
Vilchèze et al. Page 4
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

to the NAm starvation of M. tuberculosis ΔnadABC. These results suggest that the defect inpncA is the key component in the bacteriocidal phenotype of M. bovis ΔnadABC upon NAmstarvation. Thus, inhibition of both the de novo and the salvage pathways has a dramaticeffect on the viability of the tubercle bacilli.
M. bovis generates more NAD+ than M. tuberculosisPrevious studies have shown that the inhibition of either the de novo NAD+ biosynthesispathway or the NAD+ salvage pathway has no remarkable effect on the viability of M.tuberculosis (Boshoff et al., 2008) while we have shown that simultaneous inhibition of bothtogether is a cidal event. To test the contribution of each pathway to the NAD+ pool, theconcentration of NAD+ and NADH in both wild-type and NAm-starved ΔnadABC strainswas measured.
The cellular concentrations of NAD+ and NADH nucleotides were first determined in theΔnadABC strains grown with 20 mg/l of NAm. Both the M. tuberculosis ΔnadABC and M.bovis ΔnadABC strains had NAD+ and NADH concentrations 60% lower than wild-typewhen they relied solely on the salvage pathway to produce NAD+ (Fig. 5A).Complementation of the ΔnadABC strains with the integrative plasmid pMV361::nadABCrestored NAD+ concentration to the wild-type levels in both strains (Fig. 5A). Interestingly,the NAD+ and NADH concentrations were also very different between wild type M.tuberculosis and M. bovis, with M. bovis containing about 65% more NAD+ than M.tuberculosis. The same ratio was also observed in the M. tuberculosis ΔnadABC and M.bovis ΔnadABC strains grown on NAm. In this case, these strains are using only the salvagepathway, yet the M. bovis ΔnadABC strain contains twice as much NAD+ as M. tuberculosisΔnadABC, despite an impaired salvage pathway. The difference in NAD+ cellular levelsbetween M. bovis and M. tuberculosis could be the result of the degradation by M.tuberculosis of “excess” NAD+ to NAc, which is then secreted. The NAD+ level in the M.bovis ΔnadABC pMV261::pncATB strain is similar to the M. bovis ΔnadABC strain (Fig.5A), yet the strain is capable of secreting NAc, as seen by its positive niacin test. Therefore,the difference in NAD+ levels between M. bovis and M. tuberculosis might reflect the defectof M. bovis in the degradation of NAD+ to NAm or NMN, as previously reported (Kasarovand Moat, 1972), rather than the defect in the nicotinamidase activity or be the result ofother physiological differences between M. tuberculosis and M. bovis.
As expected, upon NAm starvation, the levels of NAD+ in the M. bovis ΔnadABC straindecreased much more rapidly than did the NAD+ levels in M. tuberculosis ΔnadABC. Onaverage, after 2 days of NAm starvation, the M. bovis ΔnadABC and M. tuberculosisΔnadABC strains had lost 60% and 15% of their NAD+ levels, respectively (Fig. 5B). Thedecrease in the M. bovis ΔnadABC NAD+ levels became more severe as the starvationperiod increased, reaching 90% and 98% loss after 7 and 14 days, respectively. In contrast,NAm starvation had a much less severe effect on the NAD+ levels of the M. bovis ΔnadABCstrain overexpressing M. tuberculosis pncA. After 2 days of NAm starvation, a slightincrease in the NAD+ levels was measured followed by a slow decrease over time,correlating with the bacteriostasis observed in this strain upon NAm starvation. We concludethat the recycling of NAD+ by the salvage pathway prevents the cidal phenotype of M.tuberculosis ΔnadABC starvation.
Transformation with the pncA gene from M. tuberculosis reverses the defect of M. bovisΔnadABC in vivo growth
An M. tuberculosis mutant defective for the NAD+ de novo pathway was recently shown tohave no growth defect in vivo due to the ability of M. tuberculosis to salvage NAm or NAcfrom the host (Boshoff et al., 2008). With the dramatic difference in death curves observed
Vilchèze et al. Page 5
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

during NAD+ starvation between the M. bovis ΔnadABC and M. tuberculosis ΔnadABCstrains, we hypothesized that M. bovis ΔnadABC would be unable to grow in mice. To testthis hypothesis, low-dose aerosol infection of C57Bl/6 mice was performed with the M.bovis ΔnadABC and M. tuberculosis ΔnadABC strains, their parental strains, and thecomplemented strains. The growth of the organisms in the lungs, livers, and spleens wasfollowed by plating for CFU at different days post infection.
In the lungs of C57Bl/6 mice, the M. tuberculosis ΔnadABC mutant grew as well as did thewild-type M. tuberculosis and the complemented mutant (Fig. 6B). In striking contrast, thegrowth of M. bovis ΔnadABC was highly attenuated, compared to wild-type M. bovis (Fig.6A). Notably, after 3 weeks post-infection, there was more than a 3-log difference betweenthe growth of the wild-type M. bovis strain and the growth of the M. bovis ΔnadABC mutant.Although the strain kept on growing slowly, even after 17 weeks there was still a 2-logdifference between the mutant and the wild-type. Consistent with a lack of growth in thelungs, M. bovis ΔnadABC never grew in either the spleen or the liver, even after 17 weeks,suggesting that there was no hematogenous spread of organisms.
Puzzled by the growth of M. bovis ΔnadABC in the lungs of the C56Bl/6 mice, wehypothesized that the colonies growing were mutants of M. bovis ΔnadABC that had perhapsacquired another mutation in the pncA gene that allowed them to salvage and utilize NAmfrom the host, like M. tuberculosis is able to. Sequencing of the pncA gene and the promoterregion of the only three colonies isolated from the lungs at 3 weeks post-infection revealedno other mutations in the pncA region other than the parent strain mutation in pncA (c169g).Since there are at least ten putative amidases in the TB genome, we wondered whether theM. bovis ΔnadABC strains isolated in vivo had turned on another amidase to allow for thehydrolysis of NAm taken from the host. To test this hypothesis, the niacin test wasperformed on the colonies isolated from the lungs at 3 weeks, and we observed that all thecolonies tested were positive for this test, while the initial M. bovis ΔnadABC strain was not.
To assess the role of the salvage pathway in the survival of M. tuberculosis NAD+ de novomutants in vivo, we tested the in vivo growth of M. bovis ΔnadABC pMV261::pncATB (Fig.6C). Low-dose aerosol infection of C57Bl/6 mice was performed with M. bovis ΔnadABCpMV261::pncATB, M. bovis ΔnadABC, and M. bovis ΔnadABC complemented. CFUs weremeasured at days 1, 7, 21, and 56 following infection. Growth in the lungs (Fig. 6C), livers,and spleens of mice infected by M. bovis ΔnadABC pMV261::pncATB and by M. bovisΔnadABC complemented followed a similar kinetics, while the livers and spleens of miceinfected with M. bovis ΔnadABC were culture-negative and only one organism was isolatedfrom the lung of an M. bovis ΔnadABC-infected mouse at day 63.
DISCUSSIONThe impaired salvage pathway in M. bovis is one of the factors differentiating M.tuberculosis from M. bovis infection in patients. The known allele in M. bovis pncA (c169g,His57Asp), the gene encoding the pyrazinamidase/nicotinamidase that hydrolyzes the NAD+
degradation product NAm into NAc, is the basis for the negative niacin test and for theresistance to PZA (Scorpio and Zhang, 1996) observed in M. bovis. Given these well-established facts, we were rather surprised to obtain an M. bovis mutant in which the threegenes involved in the de novo NAD+ biosynthesis, nadA, nadB, and nadC, had been deleted.This successful deletion of the de novo NAD+ biosynthesis pathway in M. bovis highlightedinteresting differences from the previously reported deletion of nadA and nadB in M.tuberculosis (Boshoff et al., 2008) and from our own M. tuberculosis ΔnadABC knockout.M. bovis ΔnadABC requires 10 times more NAm to grow than does M. tuberculosisΔnadABC. This increased NAm requirement in M. bovis was a direct result of the pncA
Vilchèze et al. Page 6
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

mutation since introducing the M. tuberculosis pncA gene into the M. bovis ΔnadABCauxotroph restored the same NAm need for growth as did the M. tuberculosis auxotroph. Weconclude that M. bovis PncA must have low-level nicotinamidase activity. This is consistentwith previous observations that demonstrated a greatly reduced activity for the M. bovisnicotinamidase compared to M. tuberculosis (Konno et al., 1960; Zhang et al., 2008). Basedon our mouse studies, we can conclude that M. bovis does not typically utilize the salvagepathway for growth in mice since a deletion of the de novo pathway impairs growth. It isintriguing that numerous independent M. bovis strains have the defective pncA allele, whichwe and others have shown correlates with an impairment of the salvage pathway. It isnoteworthy that Shigella species, but not the related Escherichia coli species, all appear tohave mutations in the de novo NAD+ pathway (Prunier et al., 2007). While the selectivebasis for the maintenance of an allele that causes an impairment of the salvage pathwayremains unknown, we can speculate that it might play a role in the host specificity differencebetween M. tuberculosis and M. bovis. Further studies need to be performed to elucidate thebasis of host specificities, and the use of these mutants in the de novo pathways of M.tuberculosis and M. bovis should aid in such studies.
Another interesting question that arises is how to explain the resistance of M. bovis to PZA.Since M. bovis ΔnadABC can use NAm to grow, indicating that the hydrolysis of NAm toNAc does occur in M. bovis, we would predict that a similar hydrolysis of PZA topyrazinoic acid (POA), the active compound of PZA, could also happen in M. bovis, basedon the high degree of chemical similarity between NAm and PZA. Thus, one possibleexplanation is that the level of POA generated by M. bovis PncA is directly proportional tothe killing activity of PZA. Alternatively, the PZA resistance in M. bovis has also beenattributed to a lack of PZA uptake. However, since Raynaud et al. demonstrated that PZAand NAm use the same transport system (Raynaud et al., 1999), and since we have shownthat M. bovis ΔnadABC can use NAm to grow, we can reasonably hypothesize that there isno defect in the transport of NAm and that there is likely no defect in the transport of PZA inM. bovis. To distinguish between these two possibilities, the minimum inhibitoryconcentration (MIC) of PZA for the M. tuberculosis :pncA-overexpressing M. bovisΔnadABC strain and its parent strain was measured. The M. bovis ΔnadABC strainoverexpressing M. tuberculosis pncA was 100 times more sensitive to PZA (MIC = 3.1 mg/L) than its parent strain (MIC = 400 mg/L). Therefore, we conclude that the resistance of M.bovis to PZA is more likely due to a lower level of PncA activity than a transport problem.
Another notable difference between M. tuberculosis and M. bovis is the inability of the M.bovis auxotroph to utilize the NAm available in a mouse, the way M. tuberculosis ΔnadABCdoes. Most auxotrophs of M. tuberculosis and M. bovis fail to grow in mice (Hondalus et al.,2000; Jackson et al., 1999; Pavelka and Jacobs, 1999), indicating that the in vivoenvironment of the tubercle bacillus does not provide adequate levels of specific metabolitesor biosynthetic precursors. Other auxotrophs of M. tuberculosis, such as argF (Gordhan etal., 2002), proC, or trpD (Smith et al., 2001), failed to grow in the lungs of infectedimmunocompetent mice initially, but a few organisms could be isolated in these organs aftera few months, suggesting that the availability of specific metabolites could be alsodependent on the stage of infection. The growth of M. tuberculosis ΔnadABC in vivo can beattributed to the ability of M. tuberculosis to scavenge precursors for NAD+ biosynthesisfrom the host, as previously suggested (Bekierkunst and Artman, 1962; Boshoff et al., 2008;Windman and Bekierkunst, 1967). This suggests that the synthesizing of drugs that linkbacteriocidal moieties to NAm may be an effective drug development strategy by makingpoorly-absorbed compounds competent for uptake into M. tuberculosis cells in vivo.Previous studies had demonstrated that the NAD+ glycohydrolase was also impaired in M.bovis (Kasarov and Moat, 1972). The survival in vivo and the positive niacin test of the M.bovis ΔnadABC pMV261::pncATB strain suggest that: 1) only the defect in pncA is
Vilchèze et al. Page 7
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

responsible for the cidal effect of NAD+ starvation in M. bovis, and 2) the ability of thisstrain to secrete NAc does not correlate with a deficient NAD+ glycohydrolase.
The striking killing differences between M. bovis and M. tuberculosis upon NAD+ starvationled us to test whether M. bovis ΔnadABC infected and grew differently in macrophages thandid M. tuberculosis ΔnadABC or wild-type M. bovis. We found no significant differencesbetween these strains in their capacity to infect macrophages, and measured only a slightloss in CFU for M. bovis ΔnadABC after 3 days in macrophages. To identify differences ingene expression that might be responsible for the divergent phenotypes, we examined thetranscriptional response of M. bovis ΔnadABC and M. tuberculosis ΔnadABC to 7 days ofNAm starvation (Tables S1-S4). In a two-class SAM test to identify genes that aresignificantly differentially regulated by NAm starvation between M. tuberculosis and M.bovis, 983 were deemed significant, with many more positively significant (848 up-regulated in M. bovis, and down-regulated, absent, or not strongly induced in M.tuberculosis) than negatively significant (135 up-regulated in M. tuberculosis, down-regulated, absent, or not strongly induced in M. bovis). The preponderance of positivelysignificant genes is likely reflective of the severity of the phenotypic change in M. bovis.The transcriptional state of the dying M. bovis cells showed very limited similarity withvarious drug treatments or stress conditions (Boshoff et al., 2004). An interesting findingwas the broad similarity of the response of M. tuberculosis starved for NAm to that of M.tuberculosis starved for all nutrients, as reported (Betts et al., 2002). This similarity wasabsent from the M. bovis response to NAm starvation, suggesting that M. bovis is unable tomount an adaptive response that may protect M. tuberculosis. Part of this potential adaptiveresponse is the down-regulation of the nuo operon encoding the NADH dehydrogenase typeI, which oxidizes NADH to NAD+ and leads to the replenishment of the NAD+ pool. M.bovis up-regulates the nuo operon in an apparently futile attempt to restore NAD+ levels.Interestingly, the up-regulation of the nuo operon had also been observed in Escherichia colitreated with bacteriocidal drugs, leading to the hypothesis that cell death is ultimately causedby the rapid increase in NADH consumption leading to high levels of superoxide (Kohanskiet al., 2007).
The role of pncA in the cidal death of M. bovis ΔnadABC is crucial, since repairing the M.bovis salvage pathway by introducing the M. tuberculosis pncA gene results in loss of thebacteriocidal phenotype upon NAm starvation. In addition, the transformation of the M.bovis ΔnadABC with M. tuberculosis pncA also restores 3 other independent phenotypessimultaneously: 1) growth on lower concentration of NAm, 2) niacin secretion, and 3)reduction of NAD+ loss upon NAm starvation. Our study demonstrates that inhibiting boththe de novo and the salvage pathways simultaneously will lead to mycobacterial cell death.Another option is to design inhibitors that target the enzymes common to both pathways:NadD and NadE. Boshoff et al. recently showed that inhibitors of the NAD+ synthetaseNadE killed mycobacteria but were also cytotoxic (Boshoff et al., 2008). The mycobacterialNadE enzyme is more similar to the eukaryotic NadE enzyme than to other bacterial NadEenzymes. The amidation of NAAD to NAD+ performed by NadE occurs in two steps: 1)activation of the carboxylic group of NAAD with ATP by the addition of AMP and therelease of PPi followed by 2) amidation and the release of AMP. Two classes of NAD+
synthetases have been defined depending on the donor for the amidation reaction: ammonia-dependent NadE and glutamine-dependent NadE. Most bacteria have ammonia-dependentNadE but M. tuberculosis has a glutamine-dependent NadE, like the one found in eukaryotes(Magni et al., 2009). Designing NadE inhibitors that are specific for the mycobacterial NadEwith no activity against eukaryotic NadE might, therefore, be challenging.
Our data suggest that targeting the de novo pathway could also lead to new drug candidatesfor treating M. bovis infections. While zoonotic TB is extremely rare in humans in
Vilchèze et al. Page 8
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

economically developed nations as a result of the pasteurization of dairy products and thesystematic elimination of TB-infected cattle, this is not the case in the developing world(Grange, 2001). Inhibiting the NAD+ de novo pathway would also be valuable for thetreatment of patients infected with PZA-resistant M. tuberculosis, since 70-93% of PZA-resistant M. tuberculosis clinical strains carry mutations in the pncA gene or in its upstreamregion, leading to the loss of pyrazinamidase/nicotinamidase activity (Cheng et al., 2000;Louw et al., 2006; Singh et al., 2006). Resistance to PZA is especially high in patients withmultidrug-resistant and extensively drug-resistant TB, which renders their treatment unlikelyto succeed (Eker et al., 2008). Since none of the current anti-tuberculosis drugs, either first-,second-, or third-line, target NAD+ biosynthesis, developing new compounds that inhibitNAD+ biosynthesis might be a valuable endeavor to combat multidrug-resistant andextensively drug-resistant TB. We propose that simultaneously targeting both the salvageand the de novo NAD+ biosynthesis pathways represents a potentially effective way to treatinfection with tubercle bacilli.
EXPERIMENTAL PROCEDURESBacterial strains, plasmids, phages, and media
The M. bovis Ravenel and M. tuberculosis CDC1551 strains were obtained from laboratorystocks. The strains were grown in Middlebrook 7H9 medium (Difco, Sparks, MD)supplemented with 10% (v/v) OADC enrichment (Difco), 0.2% (v/v) glycerol, and 0.05%(v/v) tyloxapol. The solid media used was the same as that described above, with theaddition of 1.5% (w/v) agar. The plasmids pMV261 (Stover et al., 1991), pMV361 (Stoveret al., 1991), p004S (Hsu, K. & Jacobs Jr., W.R., unpublished), the shuttle cosmid pYUB412(Bardarov, S. & Jacobs Jr., W.R., unpublished), and the shuttle phasmid phAE159 (Kriakov,J. & Jacobs Jr., W.R., unpublished) were obtained from laboratory stocks. Hygromycin wasused at concentrations of 50 mg/l for mycobacteria and 150 mg/l for Escherichia coli.Kanamycin was used at concentrations of 20 mg/l for mycobacteria and 40 mg/l forEscherichia coli. Nicotinic acid (Sigma, St. Louis, MO) and nicotinamide (Sigma) stockswere prepared in sterile water and used at a concentration of 20 mg/l, unless otherwisestated. 14C-NAm was obtained from American Radiolabeled Chemicals (St. Louis, MO). Allother chemicals used in this study were obtained from Sigma.
Construction of the ΔnadABC strainsThe nadABC operons of M. tuberculosis and M. bovis were replaced by a hygromycincassette, using the specialized transduction system previously described (Bardarov et al.,2002). Briefly, a 1 kb region flanking the left and right sides of the nadABC operon werePCR-amplified from M. tuberculosis genomic DNA using the following primers (thecloning sites are underlined): nadAL1 (Van91I)TTTTTTTTCCATAAATTGGCGTGGCGCCTATCACAGGTTG, nadAL2 (Van91I)TTTTTTTTCCATTTCTTGGGTCGGCAGTCAGTTCATCCAC, nadCR1 (BstAP1)TTTTTTTTGCATAGATTGCCACACTCAGTGCGCGTGCTCG, nadCR2 (BstAP1)TTTTTTTTGCATCTTTTGCGCGTACTGATCGGCCCCGTTG. The PCR fragments werecut with the appropriate enzyme (site indicated above) and cloned into Van91I-cut p004S.The resulting cosmid was sequenced before digesting with pacI. The linearized cosmid wasligated to the pacI-cut shuttle phasmid phAE159 and the resulting phasmid was packaged invitro (GigapackII, Stratagene, La Jolla, CA). High-titer phage lysates were used to transduceM. tuberculosis CDC1551 and M. bovis Ravenel (Bardarov et al., 2002). The transductantswere checked for the deletion of the nadABC operon by Southern analysis (the genomicDNA of the transductants was cut with BamHI and probed with the left flank of the nadABCregion). The ΔnadABC strains were grown in Middlebrook 7H9 medium (see above)containing NAm (20 mg/l). For all the growth experiments (in vitro, in macrophages, and in
Vilchèze et al. Page 9
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

vivo) and the starvation experiments, the strains were washed 3-5 X with phosphate buffersaline (PBS) prior to the experiment.
Complementation of knockout strainsThe complementing plasmids were obtained by PCR amplification of the nadABC regionfrom M. tuberculosis H37Rv and M. bovis Ravenel genomic DNA, using the followingprimers: nadAF (EcoR1) TTGAATTCGCAATGACTGTGCTGAATCG, nadCR (HindIII)TTTAAGCTTCAGATCG GGTGCAGCATTTC. The PCR products were cloneddownstream of the hsp60 promoter of the E. coli-mycobacteria shuttle plasmid vectorpMV261 (replicative vector) and the integrative vector pMV361, using the cloning sitesindicated above, and sequenced. The plasmids were introduced into the knockout strains byelectroporation. The cultures (50 ml) were grown at 37°C in 500 ml roller bottles to logphase and spun down. The cell pellets were washed twice with a 10% glycerol aqueoussolution (50 ml) and resuspended in 0.5 ml of 10% glycerol aqueous solution. To the cellsuspension (0.17 ml) was added the appropriate complementing plasmid (2 μl). Afterelectroporation (2.5 V, 25 μFd, 1000 Ω), the cell suspension was added to 1 ml ofMiddlebrook media and incubated at 37°C for 24 hr before plating on kanamycin- andhygromycin-containing plates.
Analysis of 14C-NAm metabolitesCultures (10 ml), grown to early log phase (OD600nm ≈ 0.3), were washed 3 X with PBS,resuspended in media without NAm, and labeled with 14C-NAm (5 μCi) for 72 hr at 37°C.The cultures were spun down and washed three times with PBS. The cell pellets wereresuspended in 0.3 ml of water. Chloroform (0.3 ml) and glass beads (0.1 g) were added.The suspensions were lyzed using the Thermo Scientific FastPrep machine (45 sec, speed 6),spun down and the aqueous phase was filtered through a 0.22 μm filter. The aqueousfraction (50 μl) was injected in a Hewlett-Packard model HP1100 gradient chromatographequipped with an IN/US β-RAM model 2B flow-through radioisotope beta-gamma radiationdetector. Separation was achieved using an Alltech all-guard column (part no. 77082 and96080) coupled to a reverse-phase C18 column (4.6 by 150 mm; 3-mm column diameter;Alltima C18 [Alltech]). The flow rate was set at 1 ml/min and the wavelength at 260 nm.The mobile phase was 0.1M KH2PO4 - 6 mM tetrabutylammonium phosphate monobasic(pH = 5.75)/methanol. The samples were run using the elution system described previously(Sestini et al., 2000).
Determination of NADH and NAD+ cellular concentrationsCultures (30 ml) were grown at 37°C to log phase (OD600nm ≈ 1.0) in Middlebrook 7H9medium (see above) with NAm (20 mg/l) when necessary, prior to NAD+ and NADHextractions. For the starvation experiments, the strains (150 ml) were grown to early log-phase (OD600nm ≈ 0.5), spun down, washed 5 X with PBS, resuspended in Middlebrook7H9 medium (see above) without NAm, and incubated at 37°C for 2, 7, and 14 days, prior toNAD+ and NADH extractions. A sample (12 ml) of each culture was spun down, treatedwith 0.2M HCl (1 ml, NAD+ extraction) or 0.2M NaOH (1 ml, NADH extraction), andheated at 50°C for 10 min. The samples were cooled to 0°C and neutralized with 0.1MNaOH (1 ml, NAD+ extraction) or 0.1M HCl (1 ml, NADH extraction). After centrifugation,the supernatant was filtered twice through a 0.2 μm filter and analyzed. The concentration ofNAD+ (or NADH) was obtained by measuring spectrophotometrically the rate of 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide reduction by the yeast type IIalcohol dehydrogenase in the presence of phenazine ethosulfate at 570 nm (Leonardo et al.,1996; San et al., 2002).
Vilchèze et al. Page 10
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Aerosol infection of C57Bl/6 miceC57Bl/6 mice (National Cancer Institute, Bethesda, MD), 20 mice per group, were infectedvia the aerosol route using a 106 CFU/ml mycobacterial suspension in PBS containing0.05% tween 80 and 0.04% antifoam. Three mice from each group were sacrificed after 24hr of infection and lung homogenates were plated on Middlebrook 7H10 plates containingthe appropriate antibiotic to determine the initial infection dose. At 1, 3, 8, and 17 (for M.bovis strains only) weeks post-infection, 3 mice were sacrificed from each group todetermine the bacterial burden in the lung, spleen, and liver.
Determination of the minimum inhibitory concentration (MIC) of PZAM. bovis ΔnadABC and M. bovis ΔnadABC pMV261::pncATB strains were grown to anOD600nm ≈ 1.0 and diluted to 1 × 107 CFU/ml. 0.05 ml of the dilutions were used toinoculate 5 ml of Middlebrook 7H9 media supplemented as described above, acidified to pH= 6.0, and containing various concentrations of PZA (0, 1.56, 3.12, 6.25, 12.5, 25, 50, 100,200, 400, and 800 mg/L). The cultures were incubated while shaking at 37°C for 2 weeks.MIC was determined as the concentration of PZA that inhibited growth.
Macrophage infection and phagocytosis assayBone marrow-derived macrophages from C57Bl/6 were differentiated in 10% conditionedmedium from L cell for 7 days on glass coverslips. Before the infection, the M. bovis strains(wild-type, M. bovis ΔnadABC, and M. bovis ΔnadABC complemented) were washed 3 Xwith PBS. Differentiated macrophages were infected with the M. bovis strains for 4 hr at amultiplicity of infection (MOI) of 1. After the infection, cells were fixed in 2%paraformaldehyde and blocked in 10% goat serum. Extracellular mycobacteria were stainedby a rabbit polyclonal antibody against Mycobacterium species (Cygnus Technologies, Inc.,Southport, NC). Extracellular and intracellular mycobacteria were visualized using an anti-Mycobacterium antibody and their ability to auto-fluoresce after excitation at 390-410 nm.Cells were counted on a fluorescent microscope (100 X magnification). Typically, infectedcells had 20-40 bacteria. Infected cells were scored positive for phagocytosis when they had5 or fewer extracellular bacteria. The percentage of infected cells positive for phagocytosiswas determined from 50 cells in each of four coverslips for each strain. Growth inmacrophage was determined by infecting C57Bl/6 bone marrow-derived macrophages withM. bovis strains, as described above. Gentamycin (0.05mg/ml) was added to the DMEMmedia. After 4hr, 1, 2 and 3 days, infected macrophages were washed twice with PBS andlyzed with 0.05% SDS (0.2 ml). After a 5 min wait, the cells were pipetted up and down 10times. The lysates were diluted and plated on Middlebrook 7H10 plates (see above)containing NAm if necessary. The plates were incubated at 37°C for 4-5 weeks.
Microarray analysisTriplicate cultures (50 ml) of M. tuberculosis and M. bovis ΔnadABC mutants were grown in7H9 OADC glycerol 0.05% tween broth in 500 ml roller bottles to an OD600nm= 0.1 in aroller incubator at 37°C. The cells were washed 3 X in PBS and resuspended in 50 ml 7H9OADC glycerol 0.05% tween broth with or without 20mg/l NAm and returned to theincubator. After 7 days, cultures were harvested and the cell pellet was resuspended in 1 mlQiagen RNA Protect reagent (Qiagen, Germantown, MD) and incubated 4 hr at roomtemperature before storage at − 80°C. Cells were collected by centrifugation andresuspended in 1 ml buffer RLT from a Qiagen RNeasy kit. The suspension was transferredto Fast-Prep Blue Cap tubes and processed for 45 sec at speed 6 in a Fast-Prep apparatus(MP Bio, Irvine, CA). After a brief incubation on ice, the debris was spun down and thesupernatant (~750 μL) was removed to a fresh microcentrifuge tube. 500 μl absolute ethanolwas added and the samples were applied to RNeasy columns in two applications. RNA was
Vilchèze et al. Page 11
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

purified, as recommended by the Qiagen protocol. RNA yield, purity, and integrity werechecked on a Nanodrop spectrophotometer (Nanodrop Products, Wilmington, DE) and anAgilent Bioanalyzer 2100 microfluidic system (Agilent Technologies, Santa Clara, CA).Contaminating DNA was removed with Ambion TURBO DNA-free (Ambion, Austin, TX),according to the manufacturer’s instructions. cDNA probes were prepared as per PFGRCprotocol SOP#M007 (http://pfgrc.jcvi.org/index.php/microarray/protocols.html). Briefly, 2μg total RNA was used as template for reverse transcription with random hexamer primersand aminoallyl-dUTP in a 2:3 ratio to dTTP in the nucleotide mix. After overnightincubation at 42°C, the reaction was stopped, RNA was hydrolyzed, and unincorporatednucleotides and free amines were removed with the Qiagen MinElute kit as per protocol,with amine-free phosphate wash and elution buffers. cDNA concentration and purity weremeasured by Nanodrop. Cy3 and Cy5 dyes were coupled to the aminoallyl-cDNA, whichwas then purified using a MinElute kit. Probe concentration, purity, and dye incorporationwere measured by Nanodrop before probes were combined, speed-vac dried, and dissolvedin hybridization buffer. Probes were hybridized according to the PFGRC protocolSOP#M008 to 70-mer oligo DNA microarrays representing the complete M. tuberculosisgenome with 4 in-slide replicates (JCVI PFGRC M. tuberculosis v. 4), which had beenprehybridized. One of the replicates was dye-flipped. Slides were washed and dried, thenscanned on a GenePix 4000A scanner (Molecular Devices, Sunnyvale, CA). Images wereprocessed with the TM4 software suite (www.TM4.org). TIGR Spotfinder was used to gridand quantitate spots. TIGR MIDAS was used for Lowess normalization, standard deviationregularization, and in-slide replicate analysis, with all quality control flags on and one badchannel tolerance policy set to generous. Results were analyzed in MeV with SignificanceAnalysis of Microarrays (SAM) and hierarchical clustering algorithms. The array data havebeen deposited in the Gene Expression Omnibus at NCBI (www.ncbi.nlm.nih.gov/geo) withaccession number GSE18909.
Quantitative real-time PCR (qRT-PCR)The differential expression of a pair of genes from each set (Tables S1-S4) was verified byquantitative real-time PCR. The DNA-free RNA samples used for the microarrayexperiments (3 biological replicates) were reverse transcribed in 20 μL reactions containing~400 ng RNA, 3 μg random hexamers, and 500 μM final dNTPs. Reactions were incubatedfor 5 min at 65°C, and for 1 min on ice. The first strand buffer, 5 mM DTT, 1 μlRNaseOUT, and 1 μl Superscript III RT, was then added and left for 5 min at roomtemperature, followed by a 1 hr incubation at 50°C. For the real-time reaction, each primer(250 nM final concentration) and 7.5 μl of template reaction (1:20 dilution) in 25 μl volumewith Applied Biosystems Power SYBR Green PCR master mix (Applied Biosystems, FosterCity, CA) were used. Samples (8 μl) were loaded in triplicate in 384-well plates and run onan ABI 7900 HT quantitative thermocycler with the following program: 2 min at 50°C, 10min at 95°C, 40 cycles of 10 sec at 95°C, 20 sec at 60°C, and 30sec at 72°C, followed by adissociation stage of 15 sec at 95°C, 15 sec at 60°C, and 15 sec at 95°C to check specificityof the products. Threshold cycles were normalized to those for 16S rRNA.
qRT-PCR Primers: ctpF F: CAACACCTTGGCGGTGGTAA; ctpF R:GGCCTCTGCCTTGGATTCTTl; atpD F: GGTGTTCTCCAAGGGCATCT; atpD R:CCTCCGACAACTCGTTCGATA; lat F: ACGAGTTTGATGCACTGCTG; lat R:TAAGATTGCCACCCCATGTCl; rrs F: GCCGTAAACGGTGGGTACTAl; rrs R:TGCATGTCAAACCCAGGTAA; pknD F: CCGTTGATCTCGCCGTAGTCA; pknD R:CCGAGGACACCCGCAAGC; nuoH F: CGCTCAACGGCTTTCGCAACA; nuoH R:GGCGCTGGGCTGGAAGTTATT; katG F: GAAGCCGAACCCGAACGTCTT; katG R:AACAGCTGGCCCGACAAC; hspX F: TCCGCGCGTACCTCGTAG; hspX R:
Vilchèze et al. Page 12
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

GGCCACCACCCTTCCCGTTCA; esxI F: CGTTGGCCTGCTCGTAGATCA; esxI R:TGTTGACCGCGAGTGAC.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe are grateful to Mei Chen, John Kim, Weijun Liu, and Regy Lukose for technical assistance. We acknowledgesupport for this work from NIH grants AI43268 and AI026170.
REFERENCESBardarov S, Bardarov S Jr, Pavelka MS Jr, Sambandamurthy V, Larsen M, Tufariello J, Chan J,
Hatfull G, Jacobs WR Jr. Specialized transduction: an efficient method for generating marked andunmarked targeted gene disruptions in Mycobacterium tuberculosis, M. bovis BCG and M.smegmatis. Microbiology. 2002; 148:3007–3017. Jr. Jr. Jr. [PubMed: 12368434]
Begley TP, Kinsland C, Mehl RA, Osterman A, Dorrestein P. The biosynthesis of nicotinamideadenine dinucleotides in bacteria. Vitam. Horm. 2001; 61:103–119. [PubMed: 11153263]
Bekierkunst A, Artman M. Tissue metabolism in infection. DPNase activity, DPN levels, and DPN-linked dehydrogenases in tissues from normal and tuberculous mice. Am. Rev. Respir. Dis. 1962;86:832–838. [PubMed: 13970359]
Betts JC, Lukey PT, Robb LC, McAdam RA, Duncan K. Evaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and protein expression profiling. Mol Microbiol.2002; 43:717–731. [PubMed: 11929527]
Boshoff HI, Myers TG, Copp BR, McNeil MR, Wilson MA, Barry CE 3rd. The transcriptionalresponses of Mycobacterium tuberculosis to inhibitors of metabolism: novel insights into drugmechanisms of action. J Biol Chem. 2004; 279:40174–40184. [PubMed: 15247240]
Boshoff HI, Xu X, Tahlan K, Dowd CS, Pethe K, Camacho LR, Park TH, Yun CS, Schnappinger D,Ehrt S, Williams KJ, Barry CE 3rd. Biosynthesis and recycling of nicotinamide cofactors inMycobacterium tuberculosis. An essential role for NAD in nonreplicating bacilli. J. Biol. Chem.2008; 283:19329–19341. [PubMed: 18490451]
Cheng SJ, Thibert L, Sanchez T, Heifets L, Zhang Y. pncA mutations as a major mechanism ofpyrazinamide resistance in Mycobacterium tuberculosis: spread of a monoresistant strain in Quebec,Canada. Antimicrob Agents Chemother. 2000; 44:528–532. [PubMed: 10681313]
Eker B, Ortmann J, Migliori GB, Sotgiu G, Muetterlein R, Centis R, Hoffmann H, Kirsten D, SchabergT, Ruesch-Gerdes S, Lange C. Multidrug- and extensively drug-resistant tuberculosis, Germany.Emerg. Infect. Dis. 2008; 14:1700–1706. [PubMed: 18976552]
Foster JW, Moat AG. Nicotinamide adenine dinucleotide biosynthesis and pyridine nucleotide cyclemetabolism in microbial systems. Microbiol. Rev. 1980; 44:83–105. [PubMed: 6997723]
Gerdes SY, Scholle MD, D’Souza M, Bernal A, Baev MV, Farrell M, Kurnasov OV, Daugherty MD,Mseeh F, Polanuyer BM, Campbell JW, Anantha S, Shatalin KY, Chowdhury SA, Fonstein MY,Osterman AL. From genetic footprinting to antimicrobial drug targets: examples in cofactorbiosynthetic pathways. J Bacteriol. 2002; 184:4555–4572. [PubMed: 12142426]
Gholson RK. The pyridine nucleotide cycle. Nature. 1966; 212:933–935.Gordhan BG, Smith DA, Alderton H, McAdam RA, Bancroft GJ, Mizrahi V. Construction and
phenotypic characterization of an auxotrophic mutant of Mycobacterium tuberculosis defective inL-arginine biosynthesis. Infect. Immun. 2002; 70:3080–3084. [PubMed: 12011001]
Grange JM. Mycobacterium bovis infection in human beings. Tuberculosis (Edinb). 2001; 81:71–77.[PubMed: 11463226]
Hondalus MK, Bardarov S, Russell R, Chan J, Jacobs WR Jr. Bloom BR. Attenuation of andprotection induced by a leucine auxotroph of Mycobacterium tuberculosis. Infect. Immun. 2000;68:2888–2898. [PubMed: 10768986]
Vilchèze et al. Page 13
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jackson M, Phalen SW, Lagranderie M, Ensergueix D, Chavarot P, Marchal G, McMurray DN,Gicquel B, Guilhot C. Persistence and protective efficacy of a Mycobacterium tuberculosisauxotroph vaccine. Infect. Immun. 1999; 67:2867–2873. [PubMed: 10338493]
Kasarov LB, Moat AG. Metabolism of nicotinamide adenine dinucleotide in human and bovine strainsof Mycobacterium tuberculosis. J. Bacteriol. 1972; 110:600–603. [PubMed: 4336690]
Khan JA, Forouhar F, Tao X, Tong L. Nicotinamide adenine dinucleotide metabolism as an attractivetarget for drug discovery. Expert Opin Ther Targets. 2007; 11:695–705. [PubMed: 17465726]
Kohanski MA, Dwyer DJ, Hayete B, Lawrence CA, Collins JJ. A common mechanism of cellulardeath induced by bactericidal antibiotics. Cell. 2007; 130:797–810. [PubMed: 17803904]
Konno K. New chemical method to differentiate human-type tubercle bacilli from other mycobacteria.Science. 1956; 124:985. [PubMed: 13380414]
Konno K, Kurzmann R, Bird KT. The metabolism of nicotinic acid in Mycobacteria: a method fordifferentiating tubercle bacilli of human origin from other Mycobacteria. Am. Rev. Tuberc. 1957;75:529–537. [PubMed: 13411410]
Konno K, Kurzmann R, Bird KT, Sbarra A. Differentiation of human tubercle bacilli from atypicalacid-fast bacilli. II. Clinical application. Am. Rev. Tuberc. 1958; 77:675–680. [PubMed:13521263]
Konno K, Nagayama H, Oka S. Differentiation of bovine tubercle bacilli from other mycobacteria bythe determination of nicotinamidase activity. Am Rev Respir Dis. 1960; 81:550–554. [PubMed:14410868]
Leonardo MR, Dailly Y, Clark DP. Role of NAD in regulating the adhE gene of Escherichia coli. J.Bacteriol. 1996; 178:6013–6018. [PubMed: 8830700]
Louw GE, Warren RM, Donald PR, Murray MB, Bosman M, Van Helden PD, Young DB, Victor TC.Frequency and implications of pyrazinamide resistance in managing previously treatedtuberculosis patients. Int J Tuberc Lung Dis. 2006; 10:802–807. [PubMed: 16848344]
Magni G, Di Stefano M, Orsomando G, Raffaelli N, Ruggieri S. NAD(P) biosynthesis enzymes aspotential targets for selective drug design. Curr Med Chem. 2009; 16:1372–1390. [PubMed:19355893]
Pavelka MS Jr. Jacobs WR Jr. Comparison of the construction of unmarked deletion mutations inMycobacterium smegmatis, Mycobacterium bovis bacillus Calmette-Guerin, and Mycobacteriumtuberculosis H37Rv by allelic exchange. J. Bacteriol. 1999; 181:4780–4789. [PubMed: 10438745]
Pope H, Smith DT. Synthesis of B-complex vitamins by tubercle bacilli when grown in syntheticmedia. Amer. Rev. Tuberc. 1946; 54:559–563. [PubMed: 20286146]
Prunier AL, Schuch R, Fernandez RE, Mumy KL, Kohler H, McCormick BA, Maurelli AT. nadA andnadB of Shigella flexneri 5a are antivirulence loci responsible for the synthesis of quinolinate, asmall molecule inhibitor of Shigella pathogenicity. Microbiology. 2007; 153:2363–2372.[PubMed: 17600080]
Qin W, Yang T, Ho L, Zhao Z, Wang J, Chen L, Zhao W, Thiyagarajan M, MacGrogan D, RodgersJT, Puigserver P, Sadoshima J, Deng H, Pedrini S, Gandy S, Sauve AA, Pasinetti GM. NeuronalSIRT1 activation as a novel mechanism underlying the prevention of Alzheimer disease amyloidneuropathology by calorie restriction. J Biol Chem. 2006; 281:21745–21754. [PubMed:16751189]
Raynaud C, Laneelle MA, Senaratne RH, Draper P, Laneelle G, Daffe M. Mechanisms ofpyrazinamide resistance in mycobacteria: importance of lack of uptake in addition to lack ofpyrazinamidase activity. Microbiology. 1999; 145:1359–1367. [PubMed: 10411262]
San KY, Bennett GN, Berrios-Rivera SJ, Vadali RV, Yang YT, Horton E, Rudolph FB, Sariyar B,Blackwood K. Metabolic engineering through cofactor manipulation and its effects on metabolicflux redistribution in Escherichia coli. Metab. Eng. 2002; 4:182–192. [PubMed: 12009797]
Sauve A. NAD+ and vitamin B3: from metabolism to therapies. J. Pharmacol. Exp. Ther. 2008;324:883–893. [PubMed: 18165311]
Scorpio A, Zhang Y. Mutations in pncA, a gene encoding pyrazinamidase/nicotinamidase, causeresistance to the antituberculous drug pyrazinamide in tubercle bacillus. Nat. Med. 1996; 2:662–667. [PubMed: 8640557]
Vilchèze et al. Page 14
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Sestini S, Jacomelli G, Pescaglini M, Micheli V, Pompucci G. Enzyme activities leading to NADsynthesis in human lymphocytes. Arch Biochem Biophys. 2000; 379:277–282. [PubMed:10898945]
Singh P, Mishra AK, Malonia SK, Chauhan DS, Sharma VD, Venkatesan K, Katoch VM. The paradoxof pyrazinamide: an update on the molecular mechanisms of pyrazinamide resistance inMycobacteria. J Commun Dis. 2006; 38:288–298. [PubMed: 17373362]
Smith DA, Parish T, Stoker NG, Bancroft GJ. Characterization of auxotrophic mutants ofMycobacterium tuberculosis and their potential as vaccine candidates. Infect. Immun. 2001;69:1142–1150. [PubMed: 11160012]
Sorci L, Pan Y, Eyobo Y, Rodionova I, Huang N, Kurnasov O, Zhong S, MacKerell AD Jr. Zhang H,Osterman AL. Targeting NAD biosynthesis in bacterial pathogens: Structure-based developmentof inhibitors of nicotinate mononucleotide adenylyltransferase NadD. Chem Biol. 2009; 16:849–861. [PubMed: 19716475]
Stover CK, de la Cruz VF, Fuerst TR, Burlein JE, Benson LA, Bennett LT, Bansal GP, Young JF, LeeMH, Hatfull GF, et al. New use of BCG for recombinant vaccines. Nature. 1991; 351:456–460.[PubMed: 1904554]
Velu SE, Cristofoli WA, Garcia GJ, Brouillette CG, Pierson MC, Luan CH, DeLucas LJ, BrouilletteWJ. Tethered dimers as NAD synthetase inhibitors with antibacterial activity. J Med Chem. 2003;46:3371–3381. [PubMed: 12852767]
Velu SE, Mou L, Luan CH, Yang ZW, DeLucas LJ, Brouillette CG, Brouillette WJ. Antibacterialnicotinamide adenine dinucleotide synthetase inhibitors: amide- and ether-linked tethered dimerswith alpha-amino acid end groups. J Med Chem. 2007; 50:2612–2621. [PubMed: 17489580]
Windman I, Bekierkunst A. Oxidized and reduced forms of nicotinamide nucleotides in livers oftuberculous mice. Am. Rev. Respir. Dis. 1967; 96:296–298. [PubMed: 4378130]
Yang T, Sauve AA. NAD metabolism and sirtuins: metabolic regulation of protein deacetylation instress and toxicity. AAPS J. 2006; 8:E632–643. [PubMed: 17233528]
Zhang H, Deng JY, Bi LJ, Zhou YF, Zhang ZP, Zhang CG, Zhang Y, Zhang XE. Characterization ofMycobacterium tuberculosis nicotinamidase/pyrazinamidase. FEBS J. 2008; 275:753–762.[PubMed: 18201201]
Vilchèze et al. Page 15
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 1.Deletion of the NAD+ de novo pathway genes, nadABC, in M. bovis and M. tuberculosis. A.NAD+ biosynthetic and salvage pathways. NadA, NadB, and NadC enzymes are unique tothe NAD+ de novo biosynthesis pathway, while the two last enzymes, NadD and NadE, arecommon to both the de novo and the salvage NAD+ pathways. The salvage pathwaydecomposes NAD+ into nicotinamide (NAm) or NMN, or converts NAm from the media tonicotinic acid mononucleotide (NAMN) using 2 enzymes, PncA and PncB. The enzymesand intermediates involved in this study are indicated in bold. B.Southern analysis of M.bovis ΔnadABC and M. tuberculosis ΔnadABC transductants. The diagram shows thereplacement of the nadABC operon in M. bovis by a hygromycin resistance gene (hyg) usingspecialized transduction. Southern analysis of the genomic DNA from wt and ΔnadABCmutants, digested with BamH1, confirmed the replacement of the nadABC genes by hyg inM. bovis and M. tuberculosis.
Vilchèze et al. Page 16
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 2.Metabolization of NAm by M. bovis. M. bovis strains were labeled with 14C-NAm for 72 hr.The cell pellets were washed intensively and lyzed. The 14C-labeled metabolites extractedfrom the cell pellets were separated by reverse-phase HPLC and detected with a beta-gammaradiation detector. The metabolites were identified by comparison with chromatograms ofstandards. The position of the signal for NAm is indicated by an arrow. Comp stands forcomplemented.
Vilchèze et al. Page 17
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 3.Growth of M. bovis ΔnadABC and M. tuberculosis ΔnadABC in vitro. A. The strains weregrown with NAm (20 mg/l) to log phase, spun down, washed 5 X with PBS, resuspended inmedia without NAm, and inoculated into media containing either NAm at differentconcentrations or NAc at 20 mg/l. The complemented strains were grown without anysupplement. The cultures were incubated with shaking at 37°C for 3 weeks and growth wasfollowed by measuring OD600nm. B. The strains were grown, washed, diluted, andinoculated in media containing or not containing NAm (20 mg/l). At each time point,samples were taken, diluted, and plated onto Middlebrook 7H10 plates containing OADC,glycerol, and NAm (20 mg/l). The plates were incubated at 37°C for 4-5 weeks and colonieswere counted. The concentrations are in mg/l.
Vilchèze et al. Page 18
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 4.Growth of M. bovis ΔnadABC pMV261::pncATB in vitro. M. bovis ΔnadABC wastransformed with M. tuberculosis pncA cloned into the replicative plasmid pMV261. A. M.bovis ΔnadABC pMV261::pncATB and its parent strain M. bovis ΔnadABC were grown inNAm-containing media, washed 5 X in PBS, diluted, and inoculated in media containingNAm (20 and 0.1 mg/l). Growth was followed by measuring OD600nm at diverse timepoints. B. M. bovis ΔnadABC pMV261::pncATB and M. bovis ΔnadABC were grown andwashed as described above, diluted 1-50, and inoculated in media without NAm (20 mg/l).At each time, samples were taken, diluted, and plated onto Middlebrook 7H10 platescontaining NAm (20 mg/l). The plates were incubated at 37°C for 4-5 weeks and colonieswere counted.
Vilchèze et al. Page 19
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
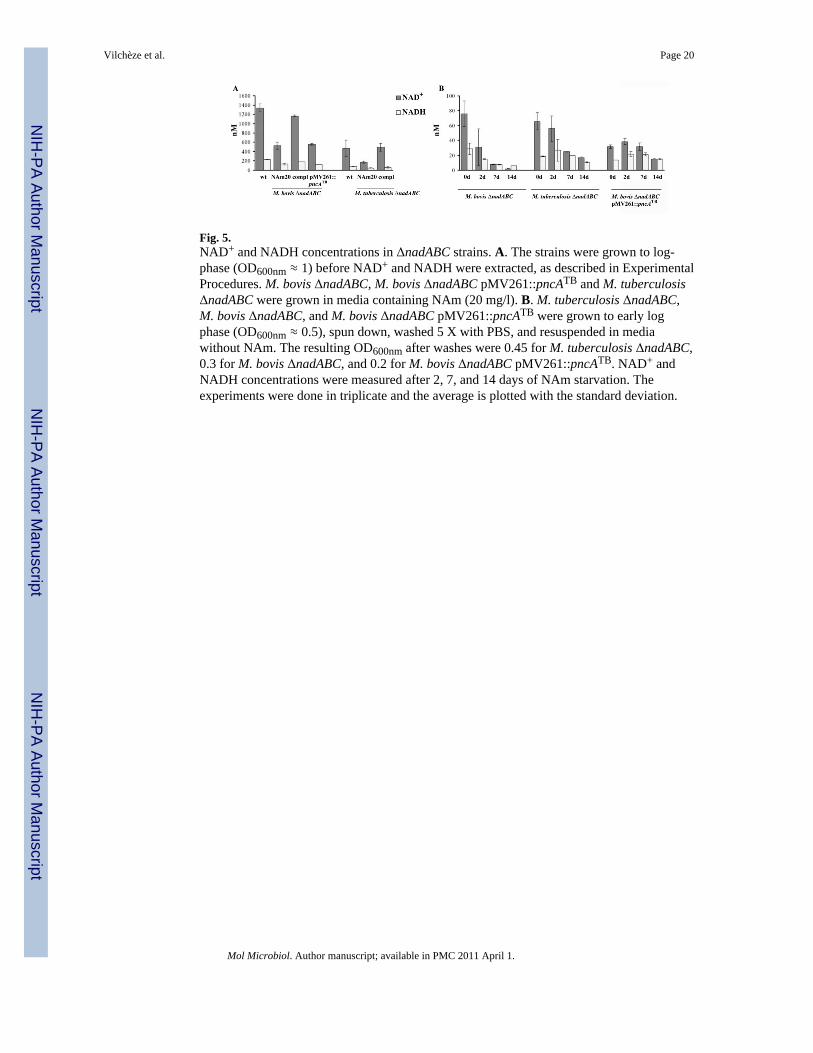
Fig. 5.NAD+ and NADH concentrations in ΔnadABC strains. A. The strains were grown to log-phase (OD600nm ≈ 1) before NAD+ and NADH were extracted, as described in ExperimentalProcedures. M. bovis ΔnadABC, M. bovis ΔnadABC pMV261::pncATB and M. tuberculosisΔnadABC were grown in media containing NAm (20 mg/l). B. M. tuberculosis ΔnadABC,M. bovis ΔnadABC, and M. bovis ΔnadABC pMV261::pncATB were grown to early logphase (OD600nm ≈ 0.5), spun down, washed 5 X with PBS, and resuspended in mediawithout NAm. The resulting OD600nm after washes were 0.45 for M. tuberculosis ΔnadABC,0.3 for M. bovis ΔnadABC, and 0.2 for M. bovis ΔnadABC pMV261::pncATB. NAD+ andNADH concentrations were measured after 2, 7, and 14 days of NAm starvation. Theexperiments were done in triplicate and the average is plotted with the standard deviation.
Vilchèze et al. Page 20
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 6.Growth in the lungs of immunocompetent C57Bl/6 mice, following low-dose aerosolinfection, of: A. M. bovis, M. bovis ΔnadABC and M. bovis ΔnadABC complemented; B. M.tuberculosis, M. tuberculosis ΔnadABC and M. tuberculosis ΔnadABC complemented; andC. M. bovis ΔnadABC, M. bovis ΔnadABC pMV261::pncATB, and M. bovis ΔnadABCcomplemented
Vilchèze et al. Page 21
Mol Microbiol. Author manuscript; available in PMC 2011 April 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript