methylotrophic methanogens in the water column of an upwelling zone with a strong oxygen gradient...
TRANSCRIPT

http://wwwsoc.nii.ac.jp/jsme2/
Microbes Environ. Vol. 22, No. 3, 268–278, 2007
Methylotrophic Methanogens in the Water Column of an Upwelling Zone
with a Strong Oxygen Gradient Off Central Chile
HéCTOR A. LEVIPAN1,2, RENATO A. QUIñONES
1,3*, HANS E. JOHANSSON4, and HOMERO URRUTIA2
1 Centro de Investigación Oceanográfica en el Pacífico Sur-Oriental (COPAS), Universidad de Concepción,
Casilla 160C, Concepción, Chile2 Departamento de Microbiología, Facultad de Ciencias Biológicas, Universidad de Concepción, Casilla 160C,
Concepción, Chile3 Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de
Concepción, Casilla 160C, Concepción, Chile4 Biosearch Technologies, Inc., 81 Digital Drive, Novato, CA 94949, USA
(Received March 18, 2007—Accepted May 30, 2007)
The growing biogeochemical and economic importance of biogenic methane contained in gas hydrates neces-
sitates a better understanding of the microorganisms involved in the last phase of organic matter degradation.
Here, the distribution and relative abundance of methylotrophic methane-producing Archaea (mMPA) were stud-
ied in an upwelling area off central Chile by rRNA dot blot hybridization. The mMPA were detected mostly dur-
ing active upwelling periods, and were more abundant in the deeper layer of the water column (>50 m), where
they represented ~10% of the prokaryote rRNA extractable from seawater samples in the oxygen minimum zone
(OMZ). Significant correlations were found between the concentration of rRNA from mMPA and (i) nitrate con-
centration (r=−0.54, p=0.0392) and (ii) temperature (r=−0.51, p=0.0267). Enrichment experiments with water
samples from the OMZ were carried out to evaluate the cellular viability of mMPA. These experiments showed
that some of these Archaea remain viable in the planktonic environment although not essentially associated with
fecal pellets or any type of compact macroaggregate. The results suggest that mMPA in the water column come
mostly from sediment and that a fraction correspond to psychrophilic varieties of facultative methylotrophs.
Key words: Methylotrophic methane-producing Archaea, Methanogenesis, Upwelling, Oxygen minimum zone, 16S-rRNA
The dynamics of the main greenhouse gasses such as car-
bon dioxide and methane is highly relevant to global warm-
ing and hence a main research area for the international sci-
entific community16,18). Methane biogenesis occurs under
strict anaerobic conditions and has been mainly studied in
marine, estuary, and salt marsh sediments50,52,63,82) and, less
frequently, in planktonic samples33,43,51). Although anoxic
marine sediments represent an important source of methane,
the contribution of marine ecosystems corresponds only 2%
of the annual methane flow to the atmosphere60).
Methane represents a source of carbon and energy for an
important trophic group of aerobic bacteria known as
methanotrophs23). Moreover, the anaerobic oxidation of
methane (AOM) occurs in different marine ecosystems,
including euxinic water columns25,59,61,80). Uncultured meth-
anothrophic Archaea46,53) catalyze this process through the
formation of cell aggregates with sulfate-reducing bacteria
(SRB), although this syntrophic relationship is not necessar-
ily a requirement for AOM76,80). Recently, it has been
observed that AOM was coupled to denitrification57).
Methane cannot be biogenerated in sulfate-rich environ-
ments because, under high sulfate concentrations, SRB out-
compete methanogens in the use of some carbon sources
such as acetate8,82). Nevertheless, in a recent study using
estuarine sediment samples it was found that, depending on
the temperature, methane production becomes the most
important terminal electron accepting process in some
* Corresponding author. E-mail address: [email protected]; Tel.:
+56–41–204520; Fax: +56–41–225400.

Marine Methylotrophic Methanogens 269
anaerobic environments32). In the upper water column of the
oceans, methane can be found oversaturated with respect to
the atmosphere12,75,81) as a result of emissions from anoxic
bottom sediment55,64) and/or from dissociated methane
hydrates and active seeps45). Different mechanisms are also
responsible for the potential or in situ production of meth-
ane in the oxygenated water column: (a) anaerobic micro-
niches in animal guts, e.g., zooplankton and fishes33,51,77); (b)
methane production in copepod fecal pellets43); and (c)
methane release during zooplankton grazing on
phytoplankton14). In the majority of these studies, methano-
genic activity was stimulated in planktonic samples of con-
centrated particulate material obtained from sediment traps,
or from several liters of seawater concentrated by vacuum
filtration. Nevertheless, few studies have reported plank-
tonic methanogenic activity detected in small volumes of
unconcentrated seawater67).
The methane hydrate deposits located off central Chile
(35° S–45° S) contain about 3.2×1013 m3 of gas stored in
subsurface sediments, which is roughly 3% of the world
total49). The methane from these deposits is in large part bio-
genic, derived from the degradation of the organic material
that falls to the sediments24). The range of substrates used by
methanogens is limited, and some are constrained to only
one or two. However, methylated compounds are important
methane precursors in sediments because methane produc-
tion is not outcompeted by sulfate reduction when it is
derived from non-competitive substrates such as methy-
lamine, methanethiol, dimethylsulfide, and methanol32,52,79,82).
Therefore, given the growing geochemical and economic
importance of methane stores, a better understanding is nec-
essary of the ecology of the microorganisms responsible for
the last stage in the decomposition of organic material. Con-
sidering that methanogenic Archaea are non-spore-formers,
the existence of oxygen-tolerant strains67,84) through, e.g.,
the presence of catalase activity, superoxide dismutases,
and superoxide reductase19,39), could allow the survival of
free-living methanogens in marine planktonic environments
subjected to low oxygen levels. In this context, the objec-
tives of this study were to determine the presence and abun-
dance of methylotrophic methane-producing Archaea
(mMPA) in the water column of a coastal upwelling area off
central Chile and to evaluate the cellular viability of this
group in the oxygen minimum zone (OMZ) of this ecosys-
tem.
Materials and Methods
Site sampling and hydrographic measurements
Water samples were collected at five depths from a sta-
tion located off central Chile (36°30' S–73°07' W; Fig. 1)
onboard the R/V Kay-Kay (University of Concepción). The
station is located approximately 18 nautical miles from the
coast and has an approximate total depth of 90 meters. This
upwelling ecosystem is located within the Humboldt Cur-
rent System (HCS; Fig. 1). The HCS contains one of the
three most important oxygen minimum zones of the global
ocean40). These conditions are generated by high oxygen
consumption due to the decomposition of organic matter
and slow rates of water renewal by ventilation48). The OMZ
is an important physical barrier to aerobic respiration15),
making oxygen levels a critical factor for the pelagic
biota4,20,21,30,47). The sampling area is characterized by
upwelling and a semi-permanent subsurface OMZ related to
the Equatorial Subsurface Waters (ESSW) that are trans-
ported poleward by the Gunther Current5,68) (Fig. 1). A
strong oxygen gradient is generally clearly detectable from
approximately 15 m from the surface with low oxygen con-
ditions (<0.5 mL O2/L) reaching the bottom.
Samples of 7 L and 50 mL were taken directly from
Niskin bottles and kept in carboys pre-washed with 10 N
HCl and sterile Falcon tubes, respectively. The 7 L samples
were used for quantitative assays of Euryarchaea and
mMPA relative to prokaryotic domains by dot blot hybrid-
ization, whereas the 50 mL samples were preserved onboard
Fig. 1. Study area and sampling station indicated by an asterisk.

LEVIPAN et al.270
with pre-filtered (0.2 µm) formaldehyde to a final concen-
tration of 2% (v/v). These samples were used for estimating
the total number of prokaryote cells under an epifluores-
cence microscope (Zeiss Axioskop 2 plus, Carl Zeiss, Got-
tingen, Germany) using 4',6-diamidino-2-phenylindole
dihydrochloride (DAPI, final concentration 72 µM) and the
method described by Porter and Feig56). In addition, water
samples were taken at a depth of 80 m with Niskin bottles
and the seawater was poured gently (to avoid the generation
of bubbles) down the side of polycarbonate bottles (1 L,
Nalgene) until the bottles were filled completely. The bot-
tles were previously autoclaved (at 121°C for 20 min) or
pre-washed with 10 N HCl. The entire process was con-
ducted within an inflatable glove bag (AtmosBag, Sigma-
Aldrich, St. Louis, USA) under an atmosphere saturated
with N2 so that the samples were kept in a hypoxic environ-
ment the whole time. The samples were stored onboard in
the dark between 4–6°C until arrival at the laboratory and
were processed within 6 h after their collection at the ocean-
ographic station.
Water samples were also taken for analyses of: phos-
phate, silicate, nitrite, nitrate, ammonium, dissolved oxy-
gen, and chlorophyll. Temperature, salinity, and oxygen
profiles were obtained with a CTDO (model SBE-19, Sea-
Bird Electronics Inc., Bellevue, Washington, USA). Mea-
surements of PO4, SiO4, NO2-N, and NO3-N were made
according to the method described by Strickland and
Parsons73) using an Alpkem Flow-Solution IV autoanalyzer
(OI Analytical, College Station, Texas, USA). Measure-
ments of NH4-N were carried out following the method
described by Holmes et al.26), using a Turner Designs fluo-
rometer (model 10-AU, Sunnyvale, California, USA).
Moreover, oxygen measurements taken with the CTDO
were corrected through the determination of dissolved oxy-
gen in discreet samples that were processed using the Win-
kler method9). The standing stock of phytoplankton, mea-
sured as a concentration of chlorophyll-a (chl-a), was
determined according to the method described by Holm-
Hansen et al.27).
Enrichment medium and methane assay
The mMPA community was enriched directly from water
samples taken under hypoxic conditions as described above.
For this, sterile anaerobe flasks provided with a reinforced
tapered neck were prepared containing a cocktail of concen-
trated sterile solutions of vitamins and trace minerals to a
final concentration of 1% (v/v)83); resazurin, 0.001 g/L (as a
redox indicator); methylamine-HCl, 2.5 g/L; methanol
(final concentration 0.5% v/v) as a catalytic additional sub-
strate; and NH4Cl, 0.5 g/L (nitrogen source). Additionally,
concentrated solutions of sodium molybdate (5 mM final
concentration) and sodium 2-mercaptoethanesulfonate
(MESNA, final concentration 0.017% w/v) were included
in the cocktail as inhibitors of sulfate-reducing bacteria.
Quickly, seawater previously filtered through 25 µm was
added to each glass flask to give a final volume of 225 mL
(70% of the total volume of the anaerobe flask) and the pH
was immediately adjusted to 7.5 or 7.85. Then, with the
sterile conditions maintained, the anaerobe flasks were
gassed with filtered N2 for 3 to 5 minutes and hermetically
sealed with a conical butyl rubber stopper. Finally, a sterile
solution of cysteine-sulfide was injected to a final concen-
tration of 0.03% (w/v) as a reducing agent. Batches were
cultured in the dark and incubated in waterbaths at 30°C
with a pH of 7.5, and at 12°C with a pH of 7.85. The hypusi-
nation-inhibitor N 1-guanyl-1,7-diaminoheptane (GC7) was
used as a general inhibitor of Archaea in negative
controls29); hypusination is an essential archaeal and eukary-
otic posttranslational modification3,22,54). Jansson et al.29)
have found that GC7, whose target is the enzyme deoxyhy-
pusine synthase (DHS), is highly efficient in specifically
inhibiting the growth of Archaea presumably by blocking
protein synthesis. In our experiments, GC7 was used at time-
zero at a final concentration of 200 µM; then this dose was
given every three days during the first week and, after that,
every 4 days until the end of the experiments.
The volume of biogas generated by the cultures was
Fig. 2. Setup of the mounted system for determining methane pro-
duction by natural Archaea communities. Samples for the chro-
matographic analysis of gases were taken directly from the head-
space of item 7.

Marine Methylotrophic Methanogens 271
determined by the method of liquid displacement (Fig. 2)
adapted from the methodology described by Isa et al.28). The
concentration of methane in the biogas was determined
using a gas chromatographer (Hewlett-Packard, model
5890, Palo Alto, California, USA) equipped with a packed
column (1.83 m×3.18 mm; Porapak Q, 80/100) and a ther-
mal conductivity detector (TCD). The carrier gas was high-
purity helium-air and the column oven was programmed to
start with an initial temperature of 60°C for 0.5 min, and
then to increase to 125°C at 25°C/min. The injector and
TCD were set to 125°C. Standards of methane at 0.1, 1, and
5% were prepared for calibration. The biogas volume,
methane concentration, and DAPI counts were monitored
over time in cultures prepared in duplicate.
Dot blot hybridizations of 16S-rRNA
RNase-inactivation was achieved by treating all solutions
with diethylpyrocarbonate (DEPC; Sigma Chemical Co., St
Louis, USA) as previously described62). Seven liters of sea-
water were pre-filtered through 25 µm and concentrated by
vacuum filtration (<10 cm Hg) using filters of cellulose
ester (pore size 0.22 µm; GSWP04700, Millipore corp.,
Bedford, Massachusetts, USA). Subsequently, the microor-
ganisms retained on the filters were resuspended in pre-fil-
tered (0.22 µm; GSWP04700, Millipore corp.) seawater
containing Tween 20 (final concentration 0.05% v/v) using
a vortex and an ultrasonic waterbath at 4°C for 7–10 min-
utes. These suspensions were harvested by centrifuging at
11,500×g for 15 min at 4°C. The pellets were washed once
with phosphate-buffered saline (PBS: 120 mM NaCl and
2.7 mM KCl in 10 mM phosphate buffer, pH 7.6) and fro-
zen at −80°C until they were processed. The processing
began with the pellets thawed at room temperature, and then
the rRNA was extracted by combining enzymatic and
chemical (detergent) cell lysis according to the method
described by Summers74) with the following modifications:
15 µL of mutanolysin (5,000 U/mL, Sigma) was added in
each reaction to obtain a final concentration of 150 U/mL
and lysozyme (Sigma) was used at a final concentration of 1
mg/mL.
The rRNA extracts were loaded onto nitrocellulose mem-
branes for nucleic acids (Hybond-N; Amersham Bio-
Sciences, Buckinghamshire, United Kingdom) using a dot
blotting apparatus (Bio-Rad, Hercules, California, USA) as
previously described41). Hybridization was carried out at
44°C according to the protocol of Raskin et al.58) using 5'
end digoxigenin-labeled oligodeoxynucleotide probes
(Thermo Electron Corp., Thermo Biosciences GmbH, Ulm,
Germany). The hybridization temperature was optimized
with respect to the concentration of NaCl and to the percent-
age of formamide present in the hybridization cocktail with
the following control strains: Escherichia coli DH5α, Sta-
phylococcus aureus ATCC 29213, Pseudomonas aerugi-
nosa ATCC 27853, and Photobacterium phosphoreum
CECT 4172. The probes EUB3382) (5'-GCT GCC TCC
CGT AGG AGT-3') and ARCH91572) (5'-GTG CTC CCC
CGC CAA TTC CT-3') were used to determine total
prokaryotic rRNA. The probes EURY4987) and MS141458)
were used to determine Euryarchaea and mMPA, respec-
tively; and their respective sequences (5' to 3') are CTT
GCC CRG CCC TT and CTC ACC CAT ACC TCA CTC
GGG. The bacterial strains described above were used as
negative controls for all archaeal probes. The probes were
detected by an enzymatic reaction between alkaline phos-
phatase (conjugated to an anti-digoxigin antibody) and a
chemiluminescent substrate (CSPD) following the manufac-
turer’s recommendations (Roche Diagnostics GmbH, Man-
nheim, Germany). The chemiluminescent signals on the
membranes were transferred to fluorescence-sensitive films
(ECL, Amersham), digitized on a grey scale, and processed
for the quantification of rRNA using a rRNA-standard and
Quantity One software (Bio-Rad, version 4.2) as previously
described41). Euryarchaea and mMPA abundances were
finally calculated relative to total prokaryotic rRNA as
inferred from Massana et al.44).
Results and Discussion
Detection and abundance of methylotrophic
methane-producing Archaea (mMPA)
Methylotrophic methanogens Methanosarcinaceae-rela-
tives (i.e. belonging to the genera Methanosarcina, Metha-
nococcoides, Methanolobus, and Methanohalophilus58))
were detected in the water column almost exclusively in
active upwelling periods (austral spring-summer). During
these periods, the lower limit of the oxycline, thermocline,
and nutricline was observed at about 30 m (Fig. 3). Below
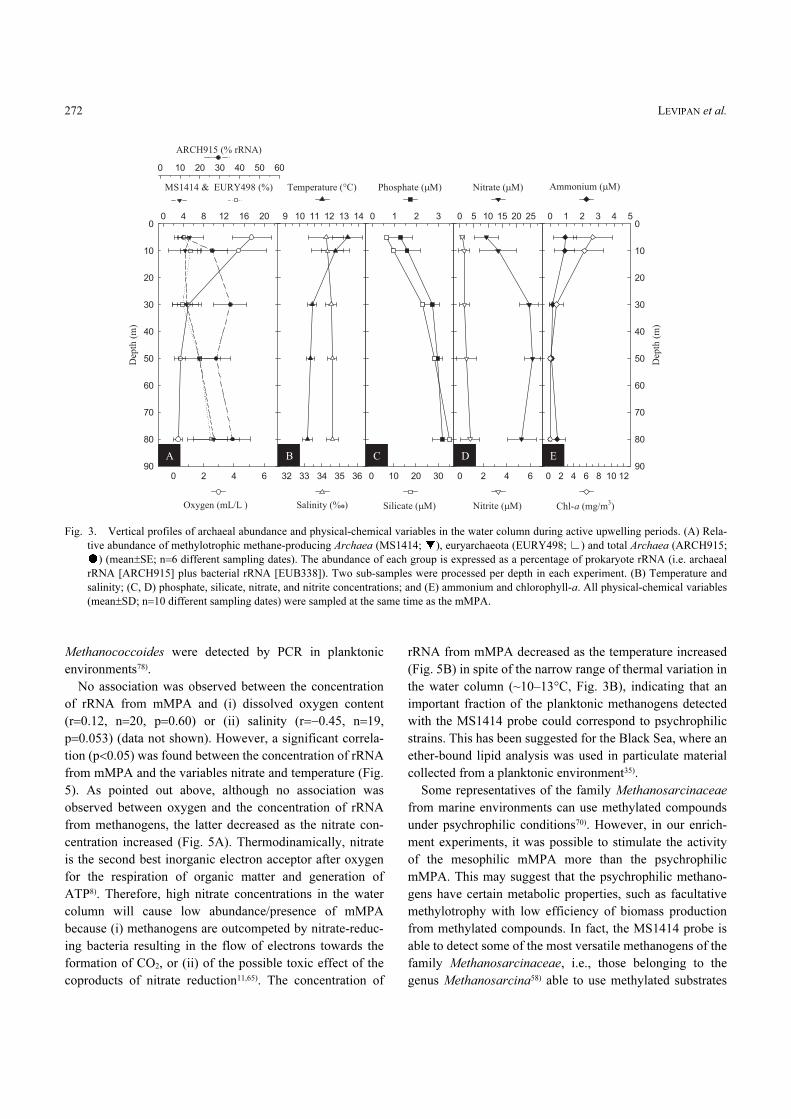
this depth, in the OMZ, the average relative abundance of
mMPA began to increase toward the bottom representing a
significant percentage (i.e. ~10%) of the total prokaryote
rRNA, and of both archaeal and euryarchaeal rRNAs (Fig.
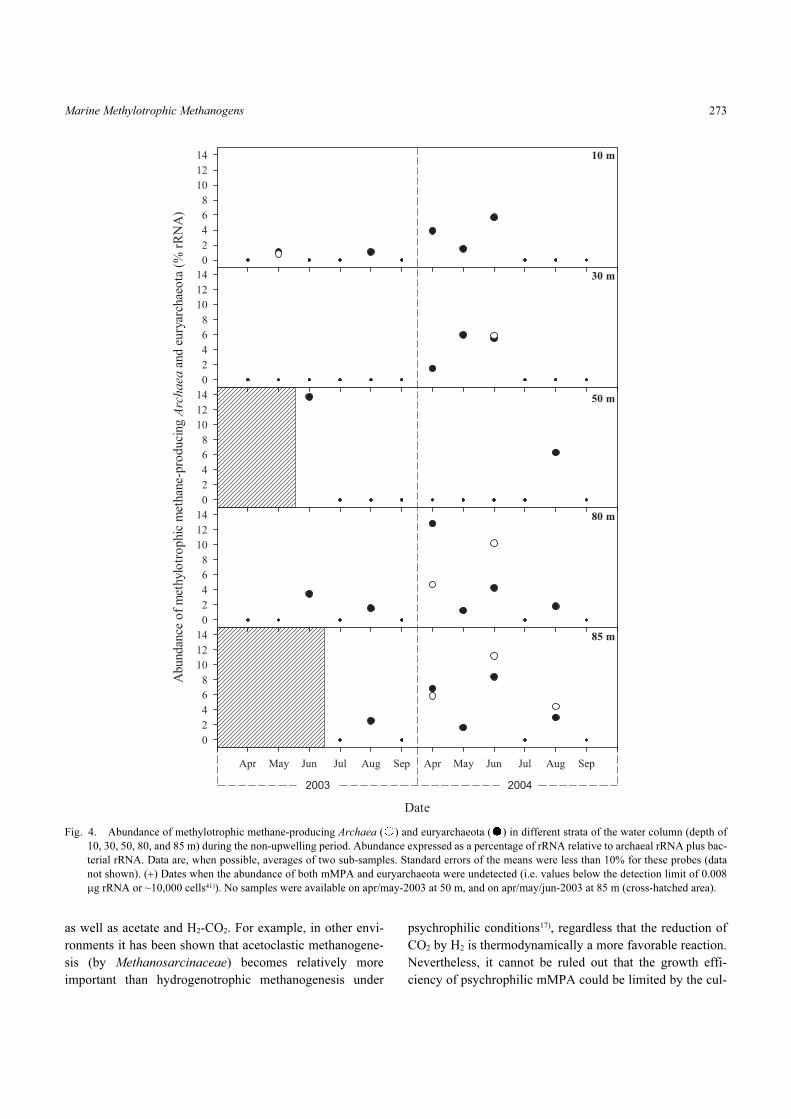
3A). The detection of mMPA and euryarchaeota was rare
during the non-upwelling periods (austral autumn-winter),
and for the former, occurred mainly in the near-bottom layer
between 80 and 85 m where it represented, on average,
8.5% of the prokaryote rRNA (Fig. 4). This is consistent
with previous findings in other marine ecosystems where
obligate-methylotroph methanogens belonging to the genus
LEVIPAN et al.272
Methanococcoides were detected by PCR in planktonic
environments78).
No association was observed between the concentration
of rRNA from mMPA and (i) dissolved oxygen content
(r=0.12, n=20, p=0.60) or (ii) salinity (r=−0.45, n=19,
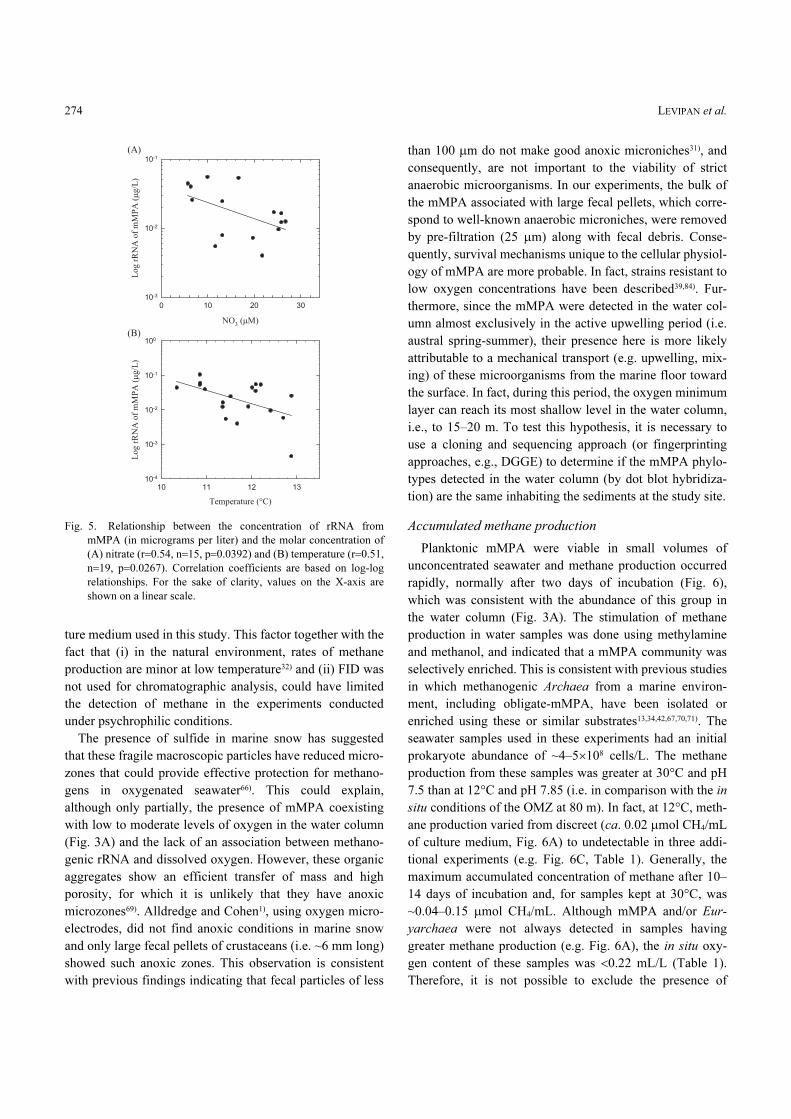
p=0.053) (data not shown). However, a significant correla-
tion (p<0.05) was found between the concentration of rRNA
from mMPA and the variables nitrate and temperature (Fig.
5). As pointed out above, although no association was
observed between oxygen and the concentration of rRNA
from methanogens, the latter decreased as the nitrate con-
centration increased (Fig. 5A). Thermodinamically, nitrate
is the second best inorganic electron acceptor after oxygen
for the respiration of organic matter and generation of
ATP8). Therefore, high nitrate concentrations in the water
column will cause low abundance/presence of mMPA
because (i) methanogens are outcompeted by nitrate-reduc-
ing bacteria resulting in the flow of electrons towards the
formation of CO2, or (ii) of the possible toxic effect of the
coproducts of nitrate reduction11,65). The concentration of
rRNA from mMPA decreased as the temperature increased
(Fig. 5B) in spite of the narrow range of thermal variation in
the water column (~10–13°C, Fig. 3B), indicating that an
important fraction of the planktonic methanogens detected
with the MS1414 probe could correspond to psychrophilic
strains. This has been suggested for the Black Sea, where an
ether-bound lipid analysis was used in particulate material
collected from a planktonic environment35).
Some representatives of the family Methanosarcinaceae
from marine environments can use methylated compounds
under psychrophilic conditions70). However, in our enrich-
ment experiments, it was possible to stimulate the activity
of the mesophilic mMPA more than the psychrophilic
mMPA. This may suggest that the psychrophilic methano-
gens have certain metabolic properties, such as facultative
methylotrophy with low efficiency of biomass production
from methylated compounds. In fact, the MS1414 probe is
able to detect some of the most versatile methanogens of the
family Methanosarcinaceae, i.e., those belonging to the
genus Methanosarcina58) able to use methylated substrates
Fig. 3. Vertical profiles of archaeal abundance and physical-chemical variables in the water column during active upwelling periods. (A) Rela-
tive abundance of methylotrophic methane-producing Archaea (MS1414; ), euryarchaeota (EURY498; ) and total Archaea (ARCH915;
) (mean±SE; n=6 different sampling dates). The abundance of each group is expressed as a percentage of prokaryote rRNA (i.e. archaeal
rRNA [ARCH915] plus bacterial rRNA [EUB338]). Two sub-samples were processed per depth in each experiment. (B) Temperature and
salinity; (C, D) phosphate, silicate, nitrate, and nitrite concentrations; and (E) ammonium and chlorophyll-a. All physical-chemical variables
(mean±SD; n=10 different sampling dates) were sampled at the same time as the mMPA.
Marine Methylotrophic Methanogens 273
as well as acetate and H2-CO2. For example, in other envi-
ronments it has been shown that acetoclastic methanogene-
sis (by Methanosarcinaceae) becomes relatively more
important than hydrogenotrophic methanogenesis under
psychrophilic conditions17), regardless that the reduction of
CO2 by H2 is thermodynamically a more favorable reaction.
Nevertheless, it cannot be ruled out that the growth effi-
ciency of psychrophilic mMPA could be limited by the cul-
Fig. 4. Abundance of methylotrophic methane-producing Archaea ( ) and euryarchaeota ( ) in different strata of the water column (depth of
10, 30, 50, 80, and 85 m) during the non-upwelling period. Abundance expressed as a percentage of rRNA relative to archaeal rRNA plus bac-
terial rRNA. Data are, when possible, averages of two sub-samples. Standard errors of the means were less than 10% for these probes (data
not shown). (+) Dates when the abundance of both mMPA and euryarchaeota were undetected (i.e. values below the detection limit of 0.008
µg rRNA or ~10,000 cells41)). No samples were available on apr/may-2003 at 50 m, and on apr/may/jun-2003 at 85 m (cross-hatched area).
LEVIPAN et al.274
ture medium used in this study. This factor together with the
fact that (i) in the natural environment, rates of methane
production are minor at low temperature32) and (ii) FID was
not used for chromatographic analysis, could have limited
the detection of methane in the experiments conducted
under psychrophilic conditions.
The presence of sulfide in marine snow has suggested
that these fragile macroscopic particles have reduced micro-
zones that could provide effective protection for methano-
gens in oxygenated seawater66). This could explain,
although only partially, the presence of mMPA coexisting
with low to moderate levels of oxygen in the water column
(Fig. 3A) and the lack of an association between methano-
genic rRNA and dissolved oxygen. However, these organic
aggregates show an efficient transfer of mass and high
porosity, for which it is unlikely that they have anoxic
microzones69). Alldredge and Cohen1), using oxygen micro-
electrodes, did not find anoxic conditions in marine snow
and only large fecal pellets of crustaceans (i.e. ~6 mm long)
showed such anoxic zones. This observation is consistent
with previous findings indicating that fecal particles of less
than 100 µm do not make good anoxic microniches31), and
consequently, are not important to the viability of strict
anaerobic microorganisms. In our experiments, the bulk of
the mMPA associated with large fecal pellets, which corre-
spond to well-known anaerobic microniches, were removed
by pre-filtration (25 µm) along with fecal debris. Conse-
quently, survival mechanisms unique to the cellular physiol-
ogy of mMPA are more probable. In fact, strains resistant to
low oxygen concentrations have been described39,84). Fur-
thermore, since the mMPA were detected in the water col-
umn almost exclusively in the active upwelling period (i.e.
austral spring-summer), their presence here is more likely
attributable to a mechanical transport (e.g. upwelling, mix-
ing) of these microorganisms from the marine floor toward
the surface. In fact, during this period, the oxygen minimum
layer can reach its most shallow level in the water column,
i.e., to 15–20 m. To test this hypothesis, it is necessary to
use a cloning and sequencing approach (or fingerprinting
approaches, e.g., DGGE) to determine if the mMPA phylo-
types detected in the water column (by dot blot hybridiza-
tion) are the same inhabiting the sediments at the study site.
Accumulated methane production
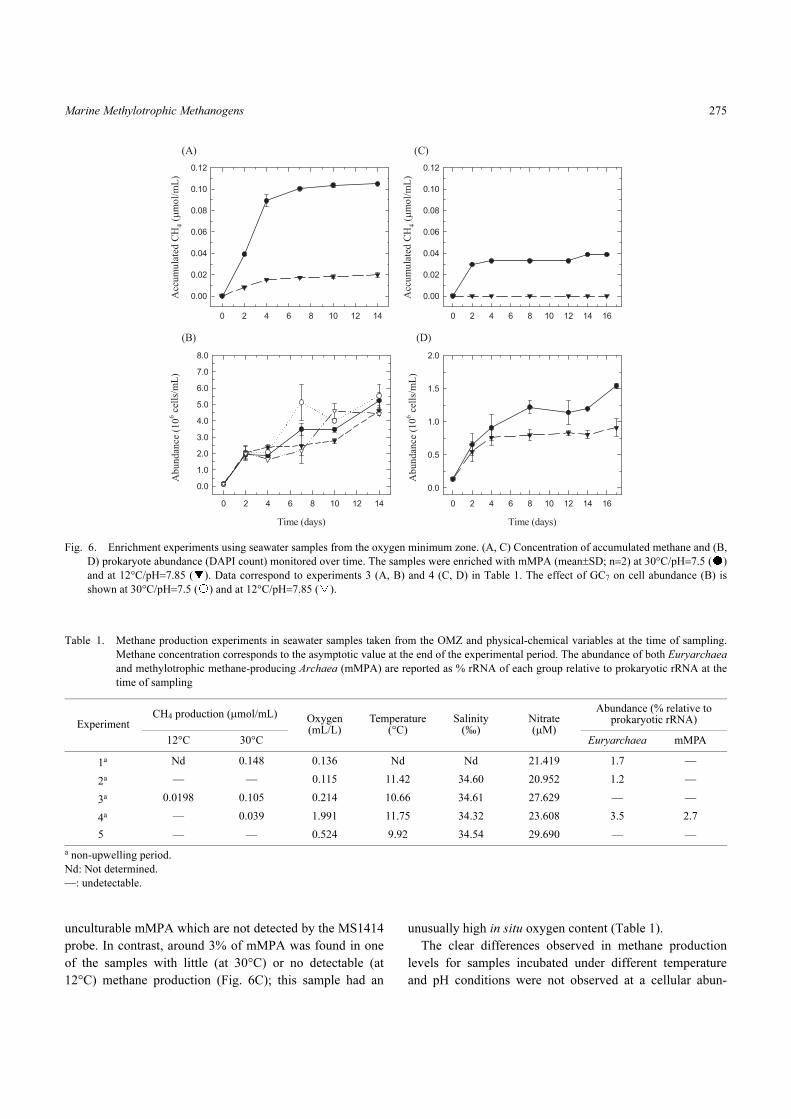
Planktonic mMPA were viable in small volumes of
unconcentrated seawater and methane production occurred
rapidly, normally after two days of incubation (Fig. 6),
which was consistent with the abundance of this group in
the water column (Fig. 3A). The stimulation of methane
production in water samples was done using methylamine
and methanol, and indicated that a mMPA community was
selectively enriched. This is consistent with previous studies
in which methanogenic Archaea from a marine environ-
ment, including obligate-mMPA, have been isolated or
enriched using these or similar substrates13,34,42,67,70,71). The
seawater samples used in these experiments had an initial
prokaryote abundance of ~4–5×108 cells/L. The methane
production from these samples was greater at 30°C and pH
7.5 than at 12°C and pH 7.85 (i.e. in comparison with the in
situ conditions of the OMZ at 80 m). In fact, at 12°C, meth-
ane production varied from discreet (ca. 0.02 µmol CH4/mL
of culture medium, Fig. 6A) to undetectable in three addi-
tional experiments (e.g. Fig. 6C, Table 1). Generally, the
maximum accumulated concentration of methane after 10–
14 days of incubation and, for samples kept at 30°C, was
~0.04–0.15 µmol CH4/mL. Although mMPA and/or Eur-
yarchaea were not always detected in samples having
greater methane production (e.g. Fig. 6A), the in situ oxy-
gen content of these samples was <0.22 mL/L (Table 1).
Therefore, it is not possible to exclude the presence of
Fig. 5. Relationship between the concentration of rRNA from
mMPA (in micrograms per liter) and the molar concentration of
(A) nitrate (r=0.54, n=15, p=0.0392) and (B) temperature (r=0.51,
n=19, p=0.0267). Correlation coefficients are based on log-log
relationships. For the sake of clarity, values on the X-axis are
shown on a linear scale.
Marine Methylotrophic Methanogens 275
unculturable mMPA which are not detected by the MS1414
probe. In contrast, around 3% of mMPA was found in one
of the samples with little (at 30°C) or no detectable (at
12°C) methane production (Fig. 6C); this sample had an
unusually high in situ oxygen content (Table 1).
The clear differences observed in methane production
levels for samples incubated under different temperature
and pH conditions were not observed at a cellular abun-
Fig. 6. Enrichment experiments using seawater samples from the oxygen minimum zone. (A, C) Concentration of accumulated methane and (B,
D) prokaryote abundance (DAPI count) monitored over time. The samples were enriched with mMPA (mean±SD; n=2) at 30°C/pH=7.5 ( )
and at 12°C/pH=7.85 ( ). Data correspond to experiments 3 (A, B) and 4 (C, D) in Table 1. The effect of GC7 on cell abundance (B) is
shown at 30°C/pH=7.5 ( ) and at 12°C/pH=7.85 ( ).
Table 1. Methane production experiments in seawater samples taken from the OMZ and physical-chemical variables at the time of sampling.
Methane concentration corresponds to the asymptotic value at the end of the experimental period. The abundance of both Euryarchaea
and methylotrophic methane-producing Archaea (mMPA) are reported as % rRNA of each group relative to prokaryotic rRNA at the
time of sampling
ExperimentCH4 production (µmol/mL) Oxygen
(mL/L)Temperature
(°C)Salinity
(‰)Nitrate (µM)
Abundance (% relative to prokaryotic rRNA)
12°C 30°C Euryarchaea mMPA
1a Nd 0.148 0.136 Nd Nd 21.419 1.7 —
2a — — 0.115 11.42 34.60 20.952 1.2 —
3a 0.0198 0.105 0.214 10.66 34.61 27.629 — —
4a — 0.039 1.991 11.75 34.32 23.608 3.5 2.7
5 — — 0.524 9.92 34.54 29.690 — —
a non-upwelling period.
Nd: Not determined.
—: undetectable.

LEVIPAN et al.276
dance level (Fig. 6). Nevertheless, a higher level of methane
production, independent of the incubation conditions, was
observed in samples that reached greater prokaryote abun-
dance in time. Moreover, we did not detect any methane
production in cultures with GC7 regardless of the incubation
period which confirms that this hypusination-inhibitor can
be used as an effective inhibitor of archaeal natural commu-
nities. Hypusination occurs in a two-step reaction catalyzed
by the enzymes DHS and deoxyhypusine hydroxylase, with
hypusine the product of the second reaction. Protein synthe-
sis initiation factor 5A (IF5A) is the only protein in eukary-
otes and Archaea known to contain hypusine3). Bacteria, on
the other hand, (i) do not have IF5A (i.e. the substrate of
DHS) but have a distant homologue, elongation factor P
(EFP)37), and (ii) only a few bacterial species have been
found to contain genes related to the DHS-like homosper-
midine synthase (HSS) genes, apparently without any func-
tional importance6). It is also unclear whether the presence
of DHS/HSS-like genes in Bacteria improves the perfor-
mance of individual cells, or only represents sequences tran-
siently imported36). Lastly, neither deoxyhypusine nor hypu-
sine has been found in Bacteria, explaining why they are
refractory to GC7. In this study, no significant differences in
the DAPI counts were found in cultures with versus without
GC7 (Fig. 6B). Consequently, the presence and growth of
non-archaeal methylotrophs (i.e. microorganisms not tar-
geted by GC7) in cultures with GC7 were the most likely
cause of the turbidity observed in the experiments con-
ducted in the presence of the inhibitor. In addition, none of
the samples showed a lag-period in prokaryote abundance
or in methane production.
Assuming a similar rRNA concentration for each of the
cells counted with DAPI in planktonic samples, the percent-
ages of 16S rRNA found for mMPA were expressed as cel-
lular abundance. Based on the total methane production
observed in our experiments, we estimated a maximum in
situ potential production of about 0.1 pmol CH4/cell/d.
It is well-known that methanogens can live in a wide
variety of environments (for review, see Chaban et al.10) and
Lange et al.38)). However, to the best of our knowledge, this
is the first study assessing the spatial variability and abun-
dance of methanogens in a coastal area with a shallow oxy-
gen minimum, comparing upwelling and non-upwelling
conditions. We have shown that methylotrophic methano-
gens are important in the water column off central Chile
during active upwelling periods (austral spring-summer),
especially within the OMZ, and that some of these microor-
ganisms remain viable in this oxygen-depleted planktonic
environment.
Acknowledgements
This research was funded by the COPAS Center
(FONDAP Nr. 150100007, CONICYT, Chile) and by FON-
DECYT grant N° 1000373 (CONICYT, Chile). We are
grateful to Ariel Pacheco, Gerdhard Jessen, and Karol
Espejo for their field support. We thank the captain and
crew of the R/V Kay-Kay for their valuable assistance and
support.
References
1) Alldredge, A.L., and Y. Cohen. 1987. Can microscale chemical
patches persist in the sea? Microelectrode study of marine snow,
fecal pellets. Science. 235:689–691.
2) Amann, R.I., B.J. Binder, R.J. Olson, S.W. Chisholm, R.
Devereux, and D.A. Stahl. 1990. Combination of 16S rRNA-
targeted oligonucleotide probes with flow cytometry for analyz-
ing mixed microbial populations. Appl. Environ. Microbiol.
56:1919–1925.
3) Bartig, D., H. Schümann, and F. Klink. 1990. The unique post-
translational modification leading to deoxyhypusine or hypusine
is a general feature of the archaebacterial kingdom. Syst. Appl.
Microbiol. 13:112–116.
4) Boyd, C.M., S.L. Smith, and T.J. Cowles. 1980. Grazing patterns
of copepods in the upwelling system off Peru. Limnol. Oceanogr.
25:583–596.
5) Brandhorst, W. 1971. Condiciones oceanográficas estivales
frente a la costa de Chile. Rev. Biol. Mar. 14:45–84.
6) Brochier, C., P. López-García, and D. Moreira. 2004. Horizontal
gene transfer and archaeal origin of deoxyhypusine synthase
homologous genes in bacteria. Gene. 330:169–176.
7) Burggraf, S., T. Mayer, R. Amann, S. Schadhauser, C.R. Woese,
and K.O. Stetter. 1994. Identifying members of the domain
Archaea with rRNA-targeted oligonucleotide probes. Appl. Envi-
ron. Microbiol. 60:3112–3119.
8) Capone, D.G., and R.P. Kiene. 1988. Comparison of microbial
dynamics in marine and freshwater sediments: contrasts in anaer-
obic carbon catabolism. Limnol. Oceanogr. 33 (Part 2):725–749.
9) Carpenter, J.H. 1965. The Chesapeake bay institute technique for
the winkler dissolved oxygen method. Limnol. Oceanogr.
10:141–143.
10) Chaban, B., S.Y.M. Ng, and K.F. Jarrell. 2006. Archaeal habitats-
from the extreme to the ordinary. Can. J. Microbiol. 52:73–116.
11) Chidthaisong, A., and R. Conrad. 2000. Turnover of glucose and
acetate coupled to reduction of nitrate, ferric iron and sulfate and
to methanogenesis in anoxic rice field soil. FEMS Microbiol.
Ecol. 31:73–86.
12) Conrad, R., and W. Seiler. 1988. Methane and hydrogen in sea-
water (Atlantic Ocean). Deep-Sea Res. 35:1903–1917.
13) Cynar, F.J., and A.A. Yayanos. 1991. Enrichment and character-
ization of a methanogenic bacterium from the oxic upper layer of
the ocean. Curr. Microbiol. 23:89–96.
14) de Angelis, M.A., and C. Lee. 1994. Methane production during
zooplankton grazing on marine phytoplankton. Limnol. Ocean-
ogr. 39:1298–1308.
15) Eissler, Y., and R.A. Quiñones. 1999. Microplanktonic respira-
tion off northern Chile during El Niño 1997–1998. J. Plank. Res.

Marine Methylotrophic Methanogens 277
21:2263–2283.
16) Falkowsky, P., R.J. Scholes, E. Boyle, J. Canadell, D. Canfield, J.
Elser, N. Gruber, K. Hibbard, P. Högberg, S. Linder, F.T. Mack-
enzie, I.I.I.B. Moore, T. Pedersen, Y. Rosenthal, S. Seitzinger, V.
Smetacek, and W. Steffen. 2000. The global carbon cycle: a test
of our knowledge of earth as a system. Science. 290:291–296.
17) Fey, A., and R. Conrad. 2000. Effect of temperature on carbon
and electron flow and on the archaeal community in methano-
genic rice field soil. Appl. Environ. Microbiol. 66:4790–4797.
18) Frankenberg, C., J.F. Meirink, M. van Weele, U. Platt, and T.
Wagner. 2005. Assessing methane emissions from global space-
borne observations. Science. 308:1010–1014.
19) Galagan, J.E., C. Nusbaum, and A. Roy. 2002. The genome of M.
acetivorans reveals extensive metabolic and physiological diver-
sity. Gen. Res. 12:532–542.
20) González, R.R., and R.A. Quiñones. 2000. Pyruvate oxidoreduc-
tases involved in glycolytic anaerobic metabolism of polychaetes
from the continental shelf off central-south Chile. Est. Coast.
Shelf Sci. 51:507–519.
21) González, R.R., and R.A. Quiñones. 2002. LDH activity in
Euphausia mucronata and Calanus chilensis: implications for
vertical migration behavior. J. Plank. Res. 24:1349–1356.
22) Gordon, E.D., R. Mora, S.C. Meredith, C. Lee, and S.L.
Lindquist. 1987. Eukaryotic initiation factor 4D, the hypusine-
containing protein, is conserved among eukaryotes. J. Biol.
Chem. 262:16585–16589.
23) Hanson, R.S., and T.E. Hanson. 1996. Methanotrophic bacteria.
Microb. Rev. 60:439–471.
24) Haq, B.U. 1999. Methane in the deep blue sea. Science. 285:543–
544.
25) Hinrichs, K.U., R.E. Summons, V. Orphan, S.P. Sylva, and J.M.
Hayes. 2000. Molecular and isotopic analysis of anaerobic meth-
ane-oxidizing communities in marine sediments. Org. Geochem.
31:1685–1701.
26) Holmes, R.M., A. Aminot, R. Kérouel, B.A. Hooker, and B.J.
Peterson. 1999. A simple and precise method for measuring
ammonium in marine and freshwater ecosystems. Can. J. Fish
Aquat. Sci. 56:1801–1808.
27) Holm-Hansen, O., C.J. Lorenzen, R.W. Holmes, and J.D.H.
Strickland. 1965. Fluorometric determination of chlorophyll. J.
Cons. Int. Explor. Mer. 30:3–15.
28) Isa, M.H., I.H. Farooqi, and R.H. Siddiqi. 1993. Methanogenic
activity test for study of anaerobic processes. Indian J. Environ.
Health. 35:1–8.
29) Jansson, B.P.M., L. Malandrin, and H.E. Johansson. 2000. Cell
cycle arrest in Archaea by the hypusination inhibitor N1-guanyl-
1,7-diaminoheptane. J. Bacteriol. 182:1158–1161.
30) Jarre, A., P. Muck, and D. Pauly. 1991. Two approaches for mod-
elling fish stock interactions in the Peruvian upwelling ecosys-
tem. ICES Mar. Sci. Symp. 193:171–184.
31) Jørgensen, B.B. 1977. Bacterial sulfate reduction within reduced
microniches of oxidized marine sediments. Mar. Biol. 41:7–17.
32) Kaku, N., A. Ueki, K. Ueki, and K. Watanabe. 2005. Methano-
genesis as an important terminal electron accepting process in
estuarine sediment at the mouth of Orikasa River. Microbes Envi-
ron. 20:41–52.
33) Karl, D.M., and B.D. Tilbrook. 1994. Production and transport of
methane in oceanic particulate organic matter. Nature. 368:732–
734.
34) Kiene, R.P., R.S. Oremland, A. Catena, L.G. Miller, and D.G.
Capone. 1986. Metabolism of reduced methylated sulfur com-
pounds by anaerobic sediments and a pure culture of an estuarine
methanogen. Appl. Environ. Microbiol. 52:1037–1045.
35) King, L.L., T.K. Pease, and S.G. Wakeham. 1998. Archaea in
Black Sea water column particulate matter and sediments-evi-
dence from ether lipid derivatives. Org. Geochem. 28:677–688.
36) Kurland, C.G., B. Canback, and O.G. Berg. 2003. Horizontal
gene transfer: A critical view. Proc. Natl. Acad. Sci. USA.
100:9658–9662.
37) Kyrpides, N.C., and C.R. Woese. 1998. Universally conserved
translation initiation factors. Proc. Natl. Acad. Sci. USA. 95:224–
228.
38) Lange, M., P. Westermann, and B.K. Ahring. 2005. Archaea in
protozoa and metazoan. Appl. Microbiol. Biotechnol. 66:465–
474.
39) Leadbetter, J.R., and J.A. Breznak. 1996. Physiological ecology
of Methanobrevibacter cuticularis sp. nov. and Methanobrevi-
bacter curvatus sp. nov., isolated from the hindgut of the termite
Reticulitermes flavipes. Appl. Environ. Microbiol. 62:3620–
3631.
40) Levin, L.A. 2003. Oxygen minimum zone benthos: adaptation
and community response to hypoxia. Oceanogr. Mar. Biol. Ann.
Rev. 41:1–45.
41) Levipan, H.A., E. Aspe, and H. Urrutia. 2004. Molecular analysis
of the community structure of nitrifying bacteria in a continuous-
flow bioreactor. Environ. Technol. 25:261–272.
42) Lyimo, T.J., A. Pol, H.J.M. Op den Camp, H.R. Harhangi, and
G.D. Vogels. 2000. Methanosarcina semesiae sp. nov., a dimetth-
ylsulfide-utilizing methanogen from mangrove sediment. Int. J.
Syst. Evol. Microbiol. 50:171–178.
43) Marty, D.G. 1993. Methanogenic bacteria in seawater. Limnol.
Oceanogr. 38:452–456.
44) Massana, R., L.T. Taylor, A.E. Murray, K.Y. Wu, W.H. Jeffrey,
and E.F. DeLong. 1998. Vertical distribution and temporal varia-
tion of marine planktonic archaea in the Gerlache Strait, Antarc-
tica, during early spring. Limnol. Oceanogr. 43:607–617.
45) McGinnis, D.F., A. Wüest, C.J. Schubert, L. Klauser, A. Lorke,
and R. Kipfer. 2005. Upward flux of methane in the Black Sea:
Does it reach the atmosphere?, p. 423–429. In J.H.W. Lee, and
K.M. Lam (ed.), Environmental hydraulics and sustainable water
management. Taylor & Francis Group, London.
46) Michaelis, W., R. Seifert, K. Nauhaus, T. Treude, V. Thiel, M.
Blumenberg, K. Knittel, A. Gieseke, K. Peterknecht, T. Pape, A.
Boetius, R. Amann, B.B. Jorgensen, F. Widdel, J. Peckmann,
N.V. Pimenov, and M.B. Gulin. 2002. Microbial reefs in the
black sea fueled by anaerobic oxidation of methane. Science.
297:1013–1015.
47) Morales, C.E., M. Braun, H. Reyes, J.L. Blanco, and A.G.
Davies. 1996. Anchovy larval distribution in the coastal zone off
northern Chile: the effect of low dissolved oxygen concentrations
and of a cold-warm sequence (1990–95). Invest. Mar. 24:77–96.
48) Morales, C.E., S. Hormazabal, and J. Blanco. 1999. Interannual
variability in the mesoscale distribution of the depth of the upper
boundary of the oxygen minimum layer off northern Chile (18°–
24° S): implications for the pelagic system and biogeochemical
cycling. J. Mar. Res. 57:909–932.
49) Morales, E. 2003. Methane hydrates in the Chilean continental
margin. Electron. J. Biotechnol. 6:80-84. Available online from:
http://www.ejbiotechnology.info/content/vol6/issue2/issues/1/
index.html.

LEVIPAN et al.278
50) Oremland, R.S. 1975. Methane production in shallow-water,
tropical marine sediments. Appl. Environ. Microbiol. 30:602–
608.
51) Oremland, R.S. 1979. Methanogenic activity in plankton samples
and fish intestines: a mechanism for in situ methanogenesis in
oceanic surface waters. Limnol. Oceanogr. 24:1136–1141.
52) Oremland, R.S., and S. Polcin. 1982. Methanogenesis and sulfate
reduction: competitive and noncompetitive substrates in estuarine
sediments. Appl. Environ. Microbiol. 44:1270–1276.
53) Orphan, V.J., H.H. Christopher, K.-U. Hinrichs, K.D. McKeegan,
and E.F. DeLong. 2001. Methane-consuming archaea revealed by
directly coupled isotopic and phylogenetic analysis. Science.
293:484–487.
54) Park, M.H., Y.B. Lee, and Y.A. Joe. 1997. Hypusine is essential
for eukaryotic cell proliferation. Biol. Signals. 6:115–123.
55) Piker, L., R. Schmaljohann, and J.F. Imhoff. 1998. Dissimilatory
sulfate reduction and methane production in Gotland deep sedi-
ments (Baltic Sea) during a transition period from oxic to anoxic
bottom water (1993–1996). Aquat. Microb. Ecol. 14:183–193.
56) Porter, K.G., and Y.S. Feig. 1980. The use of DAPI for identify-
ing and counting aquatic microflora. Limnol. Oceanogr. 25:943–
948.
57) Raghoebarsing, A.A., A. Pol, K.T. van de Pas-Schoonen, A.J.P.
Smolders, K.F. Ettwig, W.I.C. Rijpstra, S. Schouten, J.S.
Sinninghe Damsté, H.J.M. Op den Camp, M.S.M. Jetten, and M.
Strous. 2006. A microbial consortium couples anaerobic methane
oxidation to denitrification. Nature. 440:918–921.
58) Raskin, L., J.M. Stromley, B.E. Rittmann, and B.E. Stahl. 1994.
Group-specific 16S rRNA hybridization probes to describe natu-
ral communities of methanogens. Appl. Environ. Microbiol.
60:1232–1240.
59) Reeburgh, W.S. 1976. Methane consumption in Cariaco Trench
water and sediments. Earth Planet. Sci. Lett. 28:337–344.
60) Reeburgh, W.S. 1996. “Soft spots” in the global methane budget,
p. 334–342. In M.E. Lidstrom, and F.R. Tabita (ed.), Microbial
growth on C1 compounds. Kluwer Academic Publishers, Nor-
well, MA.
61) Reeburgh, W.S., B. Ward, S.C. Whalen, K.A. Sandbeck, K.A.
Kilpatrick, and L.J. Kerkhof. 1991. Black sea methane geochem-
istry. Deep-Sea Res. 38:S1189–S1210.
62) Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular
cloning: a laboratory manual. 2nd ed. Cold spring harbor labora-
tory press, New York.
63) Sansone, F.J., and C.S. Martens. 1981. Methane production from
acetate and associate methane fluxes from anoxic coastal sedi-
ments. Science. 211:707–709.
64) Schmaljohann, R. 1996. Methane dynamics in the sediment and
water column of Kiel harbour (Baltic Sea). Mar. Ecol. Prog. Ser.
131:263–273.
65) Scholten, J.C.M., and A.J.M. Stams. 1995. The effect of sulfate
and nitrate on methane formation in a freshwater sediment.
Anton. Leeuwenhoek. 68:309–315.
66) Shanks, A.L., and M.L. Reeder. 1993. Reducing microzones and
sulfide production in marine snow. Mar. Ecol. Prog. Ser. 96:43–
47.
67) Sieburth, J.Mc.N. 1993. C1 bacteria in the water column of Ches-
apeake Bay USA. I. Distribution of sub-populations of O2-toler-
ant, obligately anaerobic, methylotrophic methanogens that occur
in microniches reduced by their bacterial consorts. Mar. Ecol.
Prog. Ser. 95:67–80.
68) Silva, N. 1983. Masas de agua y circulación en la región del norte
de Chile, latitudes 18° S–32° S (Operación oceanográfica Mar
Chile XI- ERFEN II). Cien. Tec. Mar, CONA. 7:47–84.
69) Simon, M., H.-P. Grossart, B. Schweitzer, and H. Ploug. 2002.
Microbial ecology of organic aggregates in aquatic ecosystems.
Aquat. Microb. Ecol. 28:175–211.
70) Singh, N., M.M. Kendall, Y. Liu, and D.R. Boone. 2005. Isola-
tion and characterization of methylotrophic methanogens from
anoxic marine sediments in Skan Bay, Alaska: description of
Methanococcoides alaskense sp. nov., and emended description
of Methanosarcina baltica. Int. J. Syst. Evol. Microbiol.
55:2531–2538.
71) Sowers, K.R., and J.G. Ferry. 1983. Isolation and characterization
of a methylotrophic marine methanogen, Methanococcoides
methylutens gen. nov., sp. nov. Appl. Environ. Microbiol.
45:684–690.
72) Stahl, D.A., and R.I. Amann. 1991. Development and application
of nucleic acid probes in bacterial systematics, p. 205–248. In E.
Stackebrandt, and M. Goodfellow, (ed.), Sequencing and hybrid-
ization techniques in bacterial systematics. John Wiley Sons,
Chichester, England.
73) Strickland, J.D., and T.R. Parsons. 1972. A Practical Handbook
of seawater analysis. Bull. Fish. Res. Bd. Can. 167:310.
74) Summers, W.C. 1970. A simple method for extraction of RNA
from E. coli utilizing diethylpyrocarbonate. Anal. Biochem.
33:459–463.
75) Tsurushima, N., S. Watanabe, and S. Tsunogai. 1996. Methane in
the East China sea water. J. Oceanogr. 52:221–233.
76) Valentine, D.L. 2002. Biogeochemistry and microbial ecology of
methane oxidation in anoxic environments: a review. Anton.
Leeuwenhoek. 81:271–282.
77) van der Maarel, M.J.E.C., R.R.E. Artz, R. Haanstra, and L.J.
Forney. 1998. Association of marine archaea with the digestive
tracts of two marine fish species. Appl. Environ. Microbiol.
64:2894–2898.
78) van der Maarel, M.J.E.C., W. Sprenger, R. Haanstra, and L.J.
Forney. 1999. Detection of methanogenic archaea in seawater
particles and the digestive tract of a marine fish species. FEMS
Microbiol. Lett. 173:189–194.
79) Visscher, P.T., L.K. Baumgartner, D.H. Buckley, D.R. Rogers,
M.E. Hogan, C.D. Raleigh, K.A. Turk, and D.J. Des Marais.
2003. Dimethyl sulphide and methanethiol formation in microbial
mats: potential pathways for biogenic signatures. Environ. Micro-
biol. 5:296–308.
80) Wakeham, S.G., C.M. Lewis, E.C. Hopmans, S. Schouten, and
J.S. Sinninghe Damsté. 2003. Archaea mediate anaerobic oxida-
tion of methane in deep euxinic waters of the Black Sea.
Geochim. Cosmochim. Acta. 67:1359–1374.
81) Watanabe, S., N. Higashitani, N. Tsurushima, and S. Tsunogai.
1995. Methane in the western North Pacific. J. Oceanogr. 51:39–
60.
82) Winfrey, M.R., and D.M. Ward. 1983. Substrates for sulfate
reduction and methane production in intertidal sediments. Appl.
Environ. Microbiol. 45:193–199.
83) Wolin, E., N. Wolin, and R. Wolfe. 1963. Formation of methane
by bacterial extracts. J. Biol. Chem. 238:2882–2886.
84) Zehnder, A.J.B., and K. Wuhrmann. 1977. Physiology of a Meth-
anobacterium strain AZ. Arch. Microbiol. 111:199–205.