marker-assisted selection in common beans and cassava
TRANSCRIPT

Chapter 7
marker-assisted selection in common beans and cassava
Mathew W. Blair, Martin A. Fregene, Steve E. Beebe and Hernán Ceballos

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish82
SummaryMarker-assisted selection (MAS) in common beans (Phaseolus vulgaris L.) and cassava(Manihot esculenta) is reviewed in relation to the breeding system of each crop and thebreeding goals of International Agricultural Research Centres (IARCs) and NationalAgriculturalResearchSystems (NARS).The importanceof eachcrop ishighlightedandexamplesofsuccessfuluseofmolecularmarkerswithinselectioncyclesandbreedingpro-grammesaregiven for each.Forcommonbeans, examples aregivenofgene tagging forseveral traits that are important forbeanbreeding for tropical environments andaspectsconsideredthatcontributetosuccessfulapplicationofMAS.Simpletraitsthataretaggedwith easy-to-use markers are discussed first as they were the first traits prioritized forbreeding at the International Center for Tropical Agriculture (CIAT) and with NARSpartners in Central America, Colombia and eastern Africa. The specific genes for MASselectionwerethebgm-1geneforbeangoldenyellowmosaicvirus(BGYMV)resistanceandthebc-3geneforbeancommonmosaicvirus(BCMV)resistance.MASwasefficientforreducingbreedingcostsunderbothcircumstancesaslandandlaboursavingsresultedfrom eliminating susceptible individuals. The use of markers for other simply inheritedtraitsinmarker-assistedbackcrossingandintrogressionacrossAndeanandMesoamericangenepools is suggested.ThepossibilityofusingMASforquantitative traitssuchas lowsoil phosphorus adaptation is also discussed as are the advantages and disadvantages ofMASinabreedingprogramme.Forcassava,theuseofmultipleflankingmarkersforselec-tionofadominantgene,CMD2forcassavamosaicvirus(CMV)resistanceatCIATandtheInternationalInstituteofTropicalAgriculture(IITA)aswellaswithNARSpartnersintheUnitedRepublicofTanzaniausingaparticipatoryplantbreedingschemearereviewed.MASforthesamegeneisimportantduringintrogressionofcassavagreenmite(CGM)andcassavabrownstreak(CBS)resistancefromawildrelative,M. esculentasubspp.flabelli-folia.Theuseofadvancedbackcrossingwithadditionalwildrelativesisproposedasawaytodiscovergenesforhighproteincontent,waxystarch,delayedpost-harvestphysiologicaldeterioration,andresistancetowhitefliesandhornworm.OtherpotentialtargetsofMASsuchasbetacaroteneanddrymattercontentaswellaslowercyanogenicpotentialaregiven.In addition, suggestions are made for the use of molecular markers to estimate averageheterozygosity during inbreeding of cassava and for the delineation of heterotic groupswithin the species. A final section describes the similarities and differences between theMASschemespresentedforthetwocrops.Differencesbetweenthespeciescanbeascribedpartiallytothebreedingandpropagationsystemsofcommonbeans(seedpropagated,self-pollinating) and cassava (clonally propagated, cross-pollinating). In addition, differencesin growth cycles, breeding methods, availability of genetic markers, access to selectionenvironmentsandtheaccompanyingopportunitiesforphenotypicselectioninfluencethedecisionsinbothcropsofwhenandhowtoapplyMAS.RecommendationsaremadeforapplyingMASinbreedingofbothcropsincludingcarefulprioritizationoftraits,markersystems,geneticstocks,scalingup,planningofcrossesandthebalancebetweenMASandphenotypicselection.

Chapter 7 – Marker-assisted selection in common beans and cassava 83
Common BeanS: importanCe and genetiCS Common beans (Phaseolus vulgaris L.)are the most important grain legume fordirect human consumption, especially inLatin America and eastern and southernAfrica.Theyareseed-propagated,truedip-loids (2n=22) andhavea relatively smallgenome(650Mb)(Broughtonet al.,2003).Originating in the Neotropics, commonbeans were domesticated in at least twomajor centres in Mesoamerica and theAndes (Gepts, 1988) and possibly in athird minor centre in the northern Andes(Islam et al., 2002). Wide DNA poly-morphism is expressed between the twomajor gene pools. Mesoamerican beanstypically have small to medium size seedsandcanbeclassed into fourraces thataredistinguished by randomly amplified pol-ymorphic DNA (RAPD) polymorphisms(Beebeet al.,2000).Andeanbeansusuallyhavemediumtolargeseeds,andlandraceshave been classed into three races basedon plant morphology and agro-ecologicaladaptation (Singh, Gepts and Debouck,1991).Thesecanbedifferentiatedbymic-rosatellites(M.Blair,unpublisheddata)butthegeneticdistanceamongAndeanracesisnarrower than that among Mesoamericanraces (Beebeet al., 2001).A largenumberof gene tagging studies have been con-ducted in common beans, predominantlywithRAPDmarkers, someofwhichhavebeen converted subsequently to sequencecharacterized amplified regions (SCARs;reviewed most recently by Miklas et al.,2006).
Beans display a wide range of growthhabits (Van Schoonhoven and Pastor-Corrales, 1987), from determinate bushtypes, to indeterminate upright or vinybush types, to vigorous climbers. Bushtypesarethemostwidelygrown,andarea
relativelyshortseasoncrop,maturinginaslittleas60days fromseeding ina tropicalclimateandyieldingfrom700to2000kg/haonaverage.Ontheotherhand,insmall-holder agriculture where land is scarce,labour-intensive, high-yielding climbingbeansenjoycontinuingorevenexpandingpopularity. Climbing beans can mature in100to120daysatmid-elevations,butcandelayaslongastenmonthsathighereleva-tionsandcanproducethehighestyieldsforthecrop,upto5000kg/ha.Thesefeatureshave significant implications for breedingprogrammes. In bush types it is possibleto obtain up to three cycles per year inthe field, or even four cycles in green-house conditions. Breeding bush beans isthus quite agile with regard to advance ofgenerations,althoughseedharvestofindi-vidual plants is sometimes limited. Withclimbingbeans,ontheotherhand,atbestitispossibletoobtaintwocyclesperyearwith field grown plants, while managingclimbingbeansinthegreenhouseislogisti-callydifficult.However,whilebushbeansproduce on average 20 to 50 seeds/plant,individual plants of climbing beans oftenproduceenoughseedstoplantseveralrows(100to150seeds).
Beans are self-pollinating and thusbreeding methods for autogamous cropsare employed. Pedigree selection or someadaptation thereof is most common, andboth recurrent (Muñoz et al., 2004) andadvanced(orinbred)backcrossing(SullivanandBliss,1983;Buendiaet al.,2003;Blair,Iriarte and Beebe, 2003) have been used.Recurrentselectionhasalsobeenemployed(Kelly and Adams, 1987; Beaver et al.,2003)but seldomina formal sensewithadefined population structure. Singh et al.(1998) suggested a system that they calledgamete selection in which individual F1plants of multiple parent crosses give rise

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish84
tofamilies.ThissystemtakesadvantageofthevariabilityamongF1plantsthat iscre-ated between segregating parental plants.The choice of breeding method and itsadaptation to specific circumstances, thegrowth cycle of the crop in relation todifferent planting seasons, the access toselection environments and the accom-panying opportunities for phenotypicselectionandtheeaseofimplementingthespecific markers to be used will all influ-ence the decisions about where and howMAS will be most cost effective and usedtobestadvantage.
maS in bean breeding: experiences of Ciat and narS Molecular markers have been sought forboth simple and complex traits in beans,withaneyetoeventualapplicationinMAS.TaggingofgenesandQTLincommonbeanand their application to MAS have beenreviewed previously (Kelly et al., 2003;Miklaset al.,2006).Inthepresentchapter,some of the aspects that contribute to thesuccessful use of MAS are considered ingreater detail, referring to examples takenfrombeanbreedinginthetropicsatCIATand within NARS. Simple and complextraitsarediscussedseparately,astheyrep-resenttwocontrastingsortsofexperience.
Simple traitsBean golden yellow mosaic virus resistanceBeangoldenyellowmosaicvirus(BGYMV)isawhitefly-transmittedGeminivirus,anda major production limitation of beans inthe mid-to-low altitude areas of CentralAmerica, Mexico and the Caribbean.Host resistance to the virus is the mostpractical means of control, and any newvarietyintheseproductionareasmustcarryresistance. Studieson inheritanceof resist-ance revealed a major gene denominated
bgm-1 in breeding line A429 (Blair andBeaver,1993)thatoriginatesintheMexican(Durango race) accession “Garrapato” orG2402.Minorgenes (Miklaset al., 2000c)aswellasadditionalrecessiveanddominantresistancegenesexist forthevirus(Miklaset al., 2006). In most production areaswhere BGYMV exists, it is necessary topyramidgenesforadequatediseasecontrol.Although lines developed in CIAT targettheseareas,BGYMVdoesnotexistatlevelsthat would permit selection under fieldconditions inPalmira,Colombia,atCIATheadquarters. Therefore, MAS was desir-abletoassurerecoveryofatleastthemostimportant resistance genes. MAS has alsobeenemployedinthePanamericanSchoolinZamorano,Honduras,asacomplementto field screening, to extend selection tositesandseasonswithlessdiseasepressure(J.C.Rosas,personalcommunication).
A co-dominant RAPD marker wasidentified for the bgm-1 gene (Urrea et al.,1996)thatwassubsequentlyconvertedto a SCAR marker named SR2 (CIAT,1997).TheDNAfragmentassociatedwithbgm-1 gene has only been observed inone genotype other than G2402 and itsderivatives, and thus the polymorphismhas been very useful for recognizing thepresence of the gene in different geneticbackgrounds.ThisSCARwasevaluatedonasmanyas7000plants inasinglesowing(CIAT,2001;2003).Theuniquenessofthemarker’s polymorphism and its reliabilityover laboratories, seasons and geneticbackgrounds have facilitated its wide use.Morerecently,asecondSCAR(SW12.700)was developed from the W12.700 RAPDfor a QTL located on linkage group b04(Miklas et al., 2000c), and this has alsobeen incorporated into the breedingprogramme of CIAT. The combination ofbgm-1andtheQTLisexpectedtoofferan
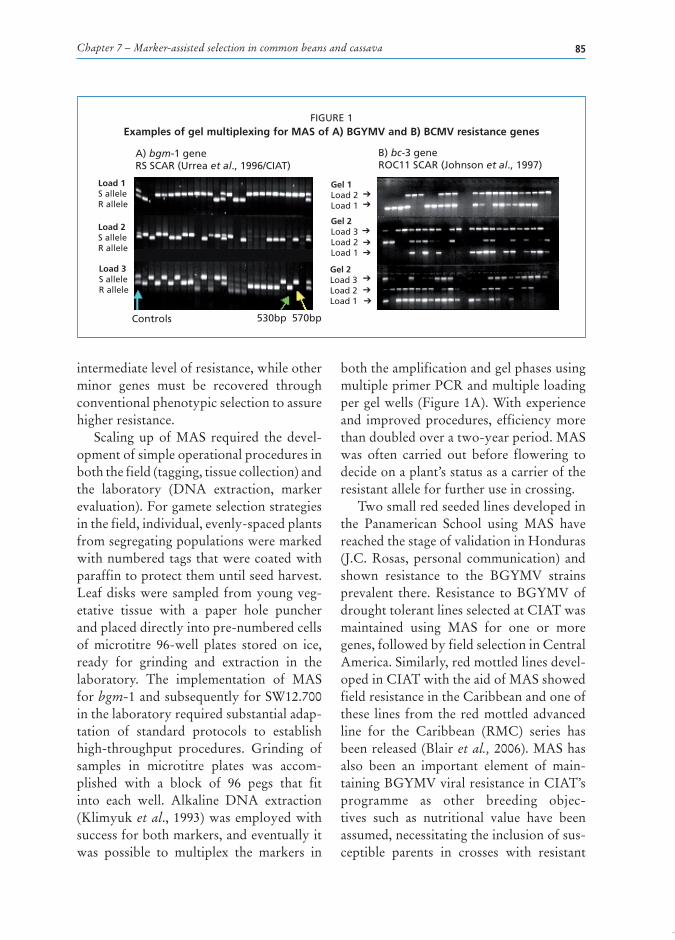
Chapter 7 – Marker-assisted selection in common beans and cassava 85
intermediatelevelofresistance,whileotherminor genes must be recovered throughconventionalphenotypicselectiontoassurehigherresistance.
Scaling up of MAS required the devel-opmentofsimpleoperationalproceduresinboththefield(tagging,tissuecollection)andthe laboratory (DNA extraction, markerevaluation).Forgameteselectionstrategiesinthefield,individual,evenly-spacedplantsfromsegregatingpopulationsweremarkedwithnumberedtagsthatwerecoatedwithparaffintoprotectthemuntilseedharvest.Leafdisksweresampled fromyoungveg-etative tissue with a paper hole puncherandplaceddirectlyintopre-numberedcellsof microtitre 96-well plates stored on ice,ready for grinding and extraction in thelaboratory. The implementation of MASforbgm-1andsubsequentlyforSW12.700inthelaboratoryrequiredsubstantialadap-tation of standard protocols to establishhigh-throughput procedures. Grinding ofsamples in microtitre plates was accom-plished with a block of 96 pegs that fitinto each well. Alkaline DNA extraction(Klimyuket al.,1993)wasemployedwithsuccessforbothmarkers,andeventuallyitwas possible to multiplex the markers in
boththeamplificationandgelphasesusingmultipleprimerPCRandmultipleloadingpergelwells(Figure1A).Withexperienceand improvedprocedures, efficiencymorethandoubledoveratwo-yearperiod.MASwas often carried out before flowering todecideonaplant’sstatusasacarrieroftheresistantalleleforfurtheruseincrossing.
Twosmallredseededlinesdevelopedinthe Panamerican School using MAS havereachedthestageofvalidationinHonduras(J.C. Rosas, personal communication) andshown resistance to the BGYMV strainsprevalent there. Resistance to BGYMV ofdroughttolerantlinesselectedatCIATwasmaintained using MAS for one or moregenes,followedbyfieldselectioninCentralAmerica.Similarly,redmottledlinesdevel-opedinCIATwiththeaidofMASshowedfieldresistanceintheCaribbeanandoneofthese lines fromthe redmottledadvancedline for the Caribbean (RMC) series hasbeenreleased(Blairet al.,2006).MAShasalso been an important element of main-tainingBGYMVviralresistanceinCIAT’sprogramme as other breeding objec-tives such as nutritional value have beenassumed,necessitatingtheinclusionofsus-ceptible parents in crosses with resistant
FiGURe 1examples of gel multiplexing for maS of a) Bgymv and B) BCmv resistance genes
A) bgm-1 geneRS SCAR (Urrea et al., 1996/CIAT)
B) bc-3 geneROC11 SCAR (Johnson et al., 1997)
Controls 530bp 570bp
Gel 1Load 2Load 1
Gel 2Load 3Load 2Load 1
Gel 2Load 3Load 2Load 1
��
�
��
�
�
�
Load 1S alleleR allele
Load 2S alleleR allele
Load 3S alleleR allele

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish86
lines.MASforthistraithasalsobeenprac-tisedattheUniversityofPuertoRicoandattheBiotechnologyInstituteofCuba.
Bean common mosaic virus and bean common mosaic necrotic virusBean common mosaic virus (BCMV) andthe relatednecrotic strains (bean commonmosaic necrotic virus [BCMNV]) areaphid-transmitted potyviruses that arefoundworldwideandareseed-bornefromseason to season. BCMNV resistance isvery important in Africa where necroticstrains are prevalent and has become arenewedpriorityforpartsoftheCaribbeanwhere necrotic strains have been discov-ered.BCMVisalsoendemicintheAndeanregionwhereitpersistsinfarmer-savedseedandlong-seasonclimbingbeans.Climbingbeansaregrowninbothintensive(trellised/staked monoculture) and extensive (inter-cropping with maize) farming systems. Inbothsystems theneed toprotect thecropfromeasily transmittedviraldiseases suchas BCMV or BCMNV is great; however,veryfewclimbingbeanshavebeenbredforresistancetoBCMV.AnumberofBCMV/BCMNVresistancegeneshavebeentaggedincludingthedominantIgene(withwhichthe necrotic strains interact to producenecrosis) and the recessive bc-3, bc-2 andbc-12 genes (Haley, Afanador and Kelly,1994; Melotto, Afanador and Kelly, 1996;Johnson et al., 1997;Miklaset al., 2000a).Thegenescanbedistinguishedbyinocula-tionwithdifferentviralisolates,andarangeof molecular marker tags are available foreach gene (reviewed in Kelly et al., 2003;Miklas et al., 2006). The dominant I genewasincorporatedintoawiderangeofsmallseededbushbeansatCIAT,whileresistantbush beans of the bush bean resistant toblack root (BRB) series carrying reces-sivegenesweredevelopedinthe1990sand
have been widely distributed as breedingparents. The need to reselect the recessivegenes with confidence from segregatingpopulationsmakesMASapriority.
CIATstartedacollaborativeprojectwiththe Colombian national bean programmebased at the Colombian AgriculturalResearch Corporation (CORPOICA) in2002to introgressBCMVresistancegenesfrom BRB lines into local landraces andimproved genotypes of Andean climbingbeans (CIAT,2002,2003,2004;Santanaet al.,2004).Duringthebreedingprogrammefor BCMV and over the course of fouryears, MAS was used extensively basedprimarily on the SCAR marker ROC11developed for the bc-3 gene (Johnson et al.,1997)andtheSCARmarkerSW13forthe I gene (Melotto, Afanador and Kelly, 1996) along with virus screening to con-firm the selection of resistant progeny.The programme was successful in movingbc-3resistanceintoabackgroundofcreammottled and red mottled seed types forbothhighlandareas(knownasCargamantocommercial class) as well as mid-altitudeareas through triple-, double- and back-crosses.Althoughvirusresistancewasalsoscreened phenotypically, the frequency ofescape, the complex interaction of mul-tiplegenesandtherecessivenatureofmostof these made MAS the best option forbreedingresistantvarietiesrapidly.Inaddi-tion, as climbing bean breeding is a moretime-consuming and expensive endeavourthanbushbeanbreedingduetothelongerseason, wider plant spacing and need forstakingmaterial,MASwasalsofoundtobeaveryeffectivemeasuretoreducebreedingcostsandsaveonbreedingnurseryspace.
TheimplementationofMASforBCMVwas based on a combination of the previ-ouslydevelopedSCARmarkerspreviouslymentioned and techniques developed at

Chapter 7 – Marker-assisted selection in common beans and cassava 87
CIATfor the selectionofBGYMVresist-ance as discussed previously. AlthoughmostBCMVandBCMNVresistancegeneshad been tagged with SCAR markers,implementation required efforts to vali-dateandscaleuptheuseofthemarkersinappliedbreedingprogrammes.GenotypingfortheROC11markerwascarriedoutonadvanced lines given that this marker isdominantandinrepulsionwiththeresist-anceallele.Inotherwords,theabsenceofabandwas indicativeof thepresenceof therecessive bc-3 allele and therefore it wasmoreappropriatetoevaluateafterfixationof the alleles to homozygosity throughmass or pedigree selection with singleplant selections in the F4 and F5 genera-tionwhensingleplantrowswereevaluatedfor the resistance gene marker. To deter-minewhethertheadvancedlinecontinuedto segregate for the gene, alkaline DNAextraction was conducted on leaf discscollectedfromfourleafletsfromfourindi-vidualplantsperlineusingahole-puncherrather than from a single plant per familyoradvanced line.Thepresenceorabsenceofpolymerasechainreaction(PCR)prod-uctswasevaluatedforeachgenotypebasedon scanned photographs or gel captureimageryofmultiplexedgels(Figure1B)topredictifthegenotypecontainedtheresist-anceorthesusceptibleallele.
Onceoptimizedforparentalgenotypes,MASwasconductedonalargenumberofprogenyrows.Forexample in2003,morethan4000advancedlineswereevaluatedfortheROC11markerforgenotypesgrownatthreesiteswithinColombia(CIAT-Darien,CIAT headquarters and CORPOICA-Rionegro).DNAwascollectedatallthreesites and shipped successfully to the labo-ratoryin96-wellplateformatasdiscussedabove.BoththeROC11andSW13markersweresinglecopySCARsthatdidnotpro-
duce extra bands and therefore were easyto multiplex. To facilitate the evaluationofmarkersonalargenumberofadvancedlines, usually within two to three weeks,andincreasetheefficiencyofMAS,severalinnovations were implemented: loading ofagarosegels(firstwithtwoandthenthreeloadings), increasingnumbersofwellspercomb(first30-wellandthen42-wellcombswereused),useof384-wellPCRplatesandmultipipetor loadingofgels.TheresultingsavingsdecreasedthetimetoPCRamplifyandloadagelbyapproximately50percentandincreasedthenumberofgenotypesrunpergelby225percent.
Therapidincreaseinefficiencyobtainedduring the application of the ROC11marker shows the advantages of testingnew markers in practical breeding pro-grammes.Theuseandadvantagesofthesemolecular markers has been presented atanOrganizationofAmericanStates-spon-sored course in Colombia given in 2002and a Rockefeller Foundation-sponsoredcourse inUgandagiven in2003.Basedonthis programme and the training courses,MAS for BCMV genes was initiated aspart of a recently approved AssociationforStrengtheningAgriculturalResearchinEastern and Central Africa (ASARECA)projectforthreecountriesineasternAfricaandtrainingofresearchersfromtheAndeanregion has allowed more breeding linesfromPerutobescreened(CIAT,2004).
other examples of maS for simply inherited traitsSeveral pathogens, especially fungalpathogens, have co-evolved with the beanhost, and present a population structure(Andean/MesoAmerican) that mimicsthe major gene pools of bean (Pastor-Corrales, Jara and Singh, 1998). This isthe case with Phaeoisariopsis griseola, the

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish88
causal agent of angular leaf spot (ALS),and Colletotrichum lindemutheanum,which induces anthracnose. In both cases,pathogenisolatestendtobemorevirulenton host genotypes of the same gene pool(AndeanorMesoamerican) and less soonhost genotypes from the contrary genepool. Resistance genes of utility to onehostgenepoolthustendtooriginateintheother gene pool and require introgressionfromonegenepooltotheother.MAShasgreat potential for introgression as DNApolymorphisms are maximized in widecrossesacrossgenepools,andmarkersareavailable for this purpose for both ALS(Carvalho et al., 1998; Sartorato et al.,1999; Nietsche et al., 2000; Ferreira et al.,2000;Mahukuet al.,2004)andanthracnose(Younget al.,1998;AwaleandKelly2001;VallejoandKelly,2001).
Other cases of wide crosses in whichMAS can be of use include those for theselectionofgenesforresistancetoastorageinsect, the Mexican bean weevil (Zabrotes subfasciatus[Boheman])derivedfromwildbeanaccessionsfromMexico.Selectionforresistance has also been achieved by anal-ysis for the active resistance agent, a seedprotein called arcelin, by either antibodyreactionorelectrophoresis,butMASissim-plerandmoreefficientthaneitheroftheseanalyses that require protein extraction.Evenwidercrossesof commonbeanwithPhaseolus acutifoliushaverecoveredresist-ance to common bacterial blight (causedby Xanthomonas axonopodis pv. phaseoli)(Muñozet al.,2004)andmarkershavealsobeen developed for these resistance genes(Junget al.,1997;Miklaset al.,2000b;Parket al., 1999; CIAT, unpublished data). Inthesecasesalso,thefactofdeployinggenesfromrelativelywidecrossesfavoursmain-taining a state of DNA polymorphism inrelationtothetargetgenotypes.
Complex multigenic traitsIn addition to the studies previously dis-cussed, several attempts have been carriedout to tag quantitative trait loci (QTL)forabioticstresstoleranceorinsectresist-ance in common bean, although most ofthese traits might better be described asoligogenic,asresultsusuallysuggestthatalimited number of loci (from three to six)areinvolvedintheirgeneticcontrol.
One example is tolerance to low soilphosphorus that was investigated in thelandrace G21212. Linkage group b08proved tobeespecially important toyieldunder low phosphorus, with as many asthree important and loosely linked QTL(Beebe,VelascoandPedraza,1999;Miklaset al.,2006).Interestingly,thesesameQTLwerelinkedtoQTLforresistancetoThrips palmiKarnyderivedfromthesamesource(Freiet al.,2005).Thisisapromisingcandi-dateforapplyingMASintheshorttermforabiotic stress tolerance, although anothernotableattemptwasalsomadefordroughttolerance breeding with MAS through ajoint programme between Michigan StateUniversity and the National Institutefor Forestry, Agriculture and LivestockResearch (INIFAP) in Mexico (Schneider,BrothersandKelly,1997).
In theory, a breeder would prefermarkers for low heritability quantitativetraits that are difficult to select throughphenotypicselection.However,ingeneral,markers for polygenic or oligogenic traitshavenotmovedintotheapplicationphase.The sameproblems thatmakephenotypicselectiondifficult apply in somedegree toMAS.Multipleminorgenes thatareoftenassociatedwithpoorheritabilityalsoimplythat it is difficult to identify QTL withhighlysignificanteffectsandthatmerittheinvestment of MAS. Furthermore, goodgenome coverage is usually necessary to

Chapter 7 – Marker-assisted selection in common beans and cassava 89
detect the QTL that explain the highestamount of genetic variability, and this hasbeen difficult to achieve in intragene poolcrossesincommonbeans.
However, genetic analysis by markershas been very useful for revealing theinheritanceofquantitativetraits,especiallyphysiologicaltraits,evenwhenthemarkersinvolved did not result in application inMAS.AnalysisofQTLwasappliedtoroottraits of bean as they relate to absorptionofphosphorusfromsoil(Liaoet al.,2004;Yan et al., 2005; Beebe et al., 2006). Thispermitted associating different physiolog-ical traits toPuptakeandestimating theirimportance in nutrient acquisition. Oncetraitsarebetterunderstood,thenanappro-priateselectionstrategycanbedevised,beit phenotypic or MAS. Thus, markers canbeusefultoabreedingprogrammebyelu-cidating basic plant mechanisms even iftheyarenotapplieddirectlyinselection.
Breeding schemes: adaptation to include maSThe eventual application of MAS requirescareful prioritization of traits and evenspecificgenesforwhichmarkersaretobesought, in light of the importance of thetraitandgenes,andoptionsforphenotypicselection. One should never assume thatMASisnecessarilysuperiortophenotypicselection,which forsometraitsmaybeaseffectiveandefficientastheuseofmolecularmarkers.However, ifagene issufficientlyimportant in a breeding programme todemand that advanced lines have such agene (as in thecaseof thebgm-1gene forvirusresistance inCentralAmerica), thereis probably some point in the selectionprocess at which MAS would be useful.Also, it is not necessary to select manygenes by MAS for it to be of great value.Forexample,ifasinglegeneissegregating
and 50percent of plants lack the gene inadvancedgenerations,aneffectiveselectionwould eliminate half the population andincrease the subsequent efficiency of thebreedingprogrammebyafactorof2.
Oncemarkersareavailable,akeyissueisdeterminingtherangeofparentalgenotypeswithinwhichamarkerispolymorphicandtherefore useful for selection. Markers ofgenesthatoriginatefromwidercrosses(e.g.fromdifferentraces,genepoolsorspecies)willhaveaprogressivelygreaterchanceofbeing polymorphic among a range of par-ents(Figure2)andthereforediagnosticforthegeneofinterest.Theexampleofbgm-1is againagoodcase inpoint as the resist-ance allele and the SR2 marker are bothuniquetotheDurangogenepoolandpol-ymorphic in combinations across otherMesoamericanracesaswellastheAndeangenepool.Incontrast,theROC11markerfor the bc-3 gene is only polymorphicacross gene pools and therefore not diag-nosticfortheresistantallele.
If a breeder has several potential par-entsamongwhichtochooseandthesearecomparable with regard to other traits,it might be preferable to eliminate thosethat carry a band that would be confusedwith the linked marker and would resultinfalsepositives.Conversely,ifmorethanone marker is available for a given gene,one might focus on those linked markersthatmaintainpolymorphisminthegreaternumber of combinations. In some com-binations it might be informative to useboth linked markers simultaneously, bothto discern recombinants and to confirmmarkers.
Several possible schemes for the intro-duction of MAS to different breedingschemes are represented in Figure 3. Abreeder must consider at what genera-tion in the breeding programme selection

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish90
by MAS will give the greatest cost/ben-efitratio.Thiswouldprobablybeearlyinthe breeding programme for the pedigreemethod or for gamete selection while itwouldbe later in theprogrammeforbulkmethodormassselection(Figure3).Inthecaseofearlygenerationselection,elimina-tionofplantswithoutthegene(s)willavoidunproductiveinvestment.
advantages and disadvantages of maSMAS provides real advantages where theconditions are not favourable for pheno-typicselection, forexample, in thecaseofBGYMV, which does not exist at epiphy-toticlevelsinCIAT.Indeed,bgm-1behavesasarecessivegene,sophenotypicselection
in early generations would be inefficientinrecoveringthegeneintheheterozygousstate.
The sameprinciplewould apply to therecessivebc-3gene,althoughthe lackofamarkerlinkedincouplingtothisgenehasbeenaseriousdrawbackandhaslimitedtheeffectiveness of MAS to advanced genera-tionswhenthegeneisfixedbyinbreeding.Inthiscase,earlygenerationselectionwithMAS would be limited to negative selec-tion against homozygous dominant andheterozygous plants, and this eliminatespotentiallyusefulallele-carryinggenotypes.Indeed, MAS is impossible in generationssuch as the F1 or BC1F1 to the suscep-tibleparentwhennohomozygousrecessiveplantsexistatall.
FiGURe 2potential of maS in crosses of varying genetic diversity
Low – due to poor polymorphism
High – due to good polymorphism
Narrowelite x elite
IntermediateInter-racial
Widewild/secondary/tertiary
gene pools
Method Purpose
Mass/Pedigree Elite crosses
Gamete/Recurrent Genepyramiding
Adv. / Rec. / Cong. IntrogressionBackcross
Width of crosses
Potential of MAS
MAS potential
Modified from Kelly, Schneider & Kolkman, 1999 and Singh, 1999

Chapter 7 – Marker-assisted selection in common beans and cassava 91
In other cases where phenotypic selec-tion methods are available, the advantageof MAS resides in its simplicity. This isthe case in the selection of arcelin, whichcanbeachievedthroughproteinextractionfollowedbyantibodydetectionorelectro-phoresis, but both of these are laboriouswhileMAScanbeappliedmorerapidlyandwith much greater throughput. Similarly,markers forcommonblightresistanceandanthracnose have the advantage of obvi-atingtheneedforfieldinoculationsthataresometimesineffectiveifenvironmentalcon-ditions are not favourable. The advantageof MAS is much greater if a single DNAextraction can serve for the evaluation ofseveral markers, as in the multiplexing ofbgm-1andSW12.700markers.
In spite of attempts to apply MAS tocomplex traits, examples of successfulapplication are still limited to relativelysimple traits. This is contrary to someprevious expectations that markers wouldbenefit mostly traits of low heritability.However, experience has shown that theability to manipulate even one important
genewithconfidencecanmakeabreedingprogramme more efficient, if that gene ishighlydesirableandvaluableforadvancedmaterials.
Meanwhile the disadvantages of MAScompared with phenotypic selection arebasedon effectiveness and cost considera-tions.TheeffectivenessofMAS is relativeto the ease of applying a given marker,its reliability and its level of linkage withthe gene of interest. Although molecularmarkers theoretically have a heritabilityof 1.0, variability among laboratories oramong runs within a laboratory makemarkerslessthan100percentreliable.Thisis especially true for RAPD markers forwhichbandamplification isdependentonDNAconcentrationandquality,annealingtemperature and thermocycling condi-tions, Taq polymerase concentration andthe relative proportion of various otheringredients to the PCR cocktail. In com-parison, SCAR markers are much morereliable and repeatable and therefore havehigher heritability than RAPD markers.Linkage distance between a marker for a
FiGURe 3application of co-dominant and dominant markers during the breeding cycle
in common beans
Mass selection F1 F 1:2 F1:3 F 1:4 F 1:5 F 5:6
Pedigree selection F1 F 1:2 F2:3 F 3:4 F 4:5 F 5:6 F 5:7
Backcrossing BC1 BC 2 BC 3 BC 4 BC 4 F 1:2
Gamete selection BC x F 1:2 BC x F1:3 BC x F 3:4
MC 1F 1:2 MC1F 1:2 MC 1F 2:3 Adv. line
Method Generation of crossing or inbreeding
F F 1:2 1:3 F 1:4 F 1:5 F 5:6
F F 1:2 2:3 F 3:4 F 4:5 F 5:6 F 5:7
BC BC BC F 1:2
BC F 1:2 1:3 BC F 3:4
MC F 1:2 F 1:2 MC F 2:3 Adv. line
note: asterisks indicate co-dominant marker, while open circles indicate dominant marker in repulsion and closed circles indicate dominant marker in coupling.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish92
gene of interest and the actual locus itselfalso affects the reliability of a marker. Inturn,thetypeofcross(wideversusnarrow)and parents involved (closely or distantlyrelated)affectthefrequencyofrecombina-tion around introgressed genes as well asthelevelofpolymorphismofthecrossandwhether the marker will present distinctalleles for the desirable and undesirablecharacter states. In this regard, there is atradeoff as MAS is most effective whenthere ishighpolymorphism in thecrossesbeing evaluated (Figure 2). However, thisispreciselythebreedingsituationinwhichgene introgression is most difficult, time-consuming and plagued by linkage drag,as is the case for interspecific or interspe-cific-derived crosses, hybridizations withwildorwild-derivedgenotypesandcrossesbetween the Andean and Mesoamericangene pools. This issue is being addressedin beans with the development and map-pingofmicrosatellitemarkers(Blairet al.,2003) that are much more polymorphicandusefulfordiagnosingtheinheritanceofgenomic segments in narrow crosses. Thefirst application of microsatellite markersforMAS in commonbeanswas the selec-tion of arcelin based bruchid resistanceusinggene-derivedsimplesequencerepeatsthatarediagnosticfortheintrogressionofalleles for resistance from wild beans intocultivated backgrounds (CIAT, 2004), butothersshouldalsoshowpromise.
In terms of cost considerations, therelative costs of MAS versus phenotypicselectionarerelativetoeachtraitandsitua-tion.ThewidelyheldperceptionthatMASisexpensiveisoftenduetotheingredientsandtimeusedtoprepareDNAextractionsand PCR reactions, although these costshave been reduced by innovations suchas the alkaline DNA extraction technique(Klimyuket al.,1993)thatobviatestheneed
fororganicsolventsorexpensiveenzymesinvolved in other mini-preparation tech-niques(AfanadorandHadley,1993).WhileexperiencedlabourwaspreviouslyrequiredforDNAextractionatCIATorinNARSbreeding programmes, the alkaline extrac-tionmethodallowsmostlaboratorystepstobecarriedoutevenbyuntrainedpersonnel.Furthermore, MAS costs can be reducedby miniaturization, especially in the PCRreaction(forexample,useof384-wellPCRplatesandsmallreactionvolumes)andre-useofingredients(forexampleplasticwareincludingpipettetipsandmicrotitreplatesaswellasagarosefromusedgels).Aspre-viously mentioned, multiplexing adds tothe efficiency and therefore reduces thedatapointcostsofMAS.
Currently, MAS with SCAR markersand alkaline extraction at CIAT cost lessthanUS$0.25perdatapoint.Thereforetheexpense of MAS is now not as impor-tant an issue as previously. In this regard,MASsometimeshastheadvantageofbeingimplemented in any generation and underbothfieldorgreenhouseconditions,whilephenotypic selection often requires a sep-arate planting and specialized labour forinoculation, agronomic management andevaluations or scoring. However, in thefinal analysis, the most efficient and costeffectivebreedingprogrammewillprobablybeonethatcombinesMASandphenotypicselection in some optimal combination. Itispreciselythechallengeofthebreedertodefinethatoptimalcombination.
OnelastdisadvantageofrelyingonMASis that it commits a breeder to a uniquegene(s)foragiventrait.Forexample,theremight be multiple genes or gene combi-nations for resistance to a disease, or fora physiological trait such as root struc-ture. To the extent that a breeder relieson MAS for selection, this excludes other

Chapter 7 – Marker-assisted selection in common beans and cassava 93
possiblegenesandtheuseofotherpoten-tially useful parents that do not share theDNApolymorphismthatisusedinMAS.On the other hand, phenotypic selectionwouldpermitrecognizingdifferentgeneticoptions for a desired phenotype. Thus,MAS is most useful when it is applied totrulyuniquegenes.
CaSSava: importanCe and genetiCSCassava is a perennial shrub but it isgenerallyharvestedasanannualcropat10–11months of age. Basically every part oftheplantcanbeutilized.Thestarchyrootsareavaluablesourceofenergyandcanbeboiled or processed in different ways forhuman consumption and different indus-trialpurposessuchasstarches,animalfeedor alcohol (Ceballos et al., 2006). Cassavastorage roots are not tubers and thereforecannotbeused forreproductivepurposes;stems are the common planting materials.Cassava foliage is not widely exploited inspite of its high nutritive value (Buitrago,1990; Babu and Chatterjee, 1999). Foliageconsumption by humans is relativelycommon in certain countries of Africa,AsiaandLatinAmerica.Theuseoffoliagefor animal feeding is generating increasedinterestinAsia.
Cassava can be propagated by eitherstem cuttings or botanical seed. However,theformeristhepracticemostwidelyusedbyfarmers formultiplicationandplantingpurposes.Propagationfromtrueseedoccursunder natural conditions and is commonin breeding programmes. Occasionallybotanical seed is also used in commer-cialpropagationschemes(Rajendranet al.,2000).
Cassavaismonoeciousandallogamous,with female flowers opening 10–14 daysbefore the male ones on the same branch.
Pollination can be done manually in acontrolled way to produce full-sib fami-lies or else in polycross nurseries whereopenpollinationtakesplaceand,therefore,half-sib families are produced. Self-pol-lination is feasible when using male andfemale flowers on different branches oron different plants of the same geno-types (Jennings and Iglesias, 2002). Someclonesflowerrelativelyearlyatfourorfivemonthsafterplantingwhereasothersonlydosoateighttotenmonthsafterplanting.Asa result, the timerequired for the seedto mature, the growing cycle of the cropandtheneedtoplantwiththearrivaloftherainstakeabouttwoyearsbetweenagivencrossbeingplannedandtherespectiveseedbecoming available. On average, betweenone and two seeds (out of the three pos-sible inthetrilocularfruit)perpollinationareobtained (Kawano,1980; JenningsandIglesias,2002).
Breeding objectivesProductivityplaysamajorroleinindustrialuses of cassava, whereas stability of pro-ductionisfundamentalinthemanyregionswherecassavaisthemainsubsistencecrop.Industrialusesofcassavarequirehighdrymattercontentasthemainqualitytraitforthe roots, whereas for human consump-tion the emphasis is on cooking quality,frequently even over productivity, as thedetermining trait. Stability of productionis associated with resistance or toleranceto major biotic and abiotic stresses, withtheemphasisvaryingwiththetargetenvi-ronment. Genetic resistance to the mostimportantdiseasesandpestsandthepreva-lentabioticstressescanbefoundincassavagermplasm (Hillocks and Wydra, 2002;Bellottiet al.,2002;Belloti,2002;Ceballoset al., 2004). Although cyanogenic glu-cosides are found in every tissue except

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish94
thecassavaseed,mostprocessingmethodsallowarapidreleaseandeliminationofthecyanide. Depending on the end use, highorlowcyanideclonesarepreferred.Otherrelevanttraitsfortherootsaredrymatter,proteinandcarotenoidcontent(Chávezet al.,2005).
Breeding schemesGenetic improvement of clonally propa-gated non-inbred crops such as cassavais made possible by the fact that a supe-rior genotype can be fixed at any stage inthe breeding scheme, even after a singlecross,theequivalentofanF1incommercialhybrids such as maize. Therefore, non-additivegeneactions includingdominanceandepitasisbecomeimportantcomponentsof the genetic variance to be manipulatedbythebreeder(Jaramilloet al.,2005;Calleet al.,2005;Perezet al.,2005a).Largeeffec-tivebreedingpopulationsizesarerequiredto retain favourable dominant alleles andepistaticlocicombination.
Asinmostcropbreedingactivities,cas-sava genetic improvement starts with theproductionofnewrecombinantgenotypesderivedfromselectedeliteclones.Scientificcassavabreedingbeganonlyafewdecadesago,andthedivergencebetweenlandraces
andimprovedgermplasmisnotaswideasin other crops. Therefore, accessions forgermplasmbankcollectionsfromdifferentresearch institutions play a more relevantroleincassavathaninothercropsthathavebeen scientifically bred for longer periodsof time. Parental lines are selected basedmainly on their performance per se andlittleprogresshasbeenmadetousegeneralcombining ability (Hallauer and MirandaFo, 1988) as a criterion forparental selec-tion.Sexualseedsobtainedbythedifferentcrossingschemesaregerminatedtoinitiatea new cycle of selection. The multiplica-tionrateofcassavaplantingmaterialislowasfivetotencuttingscanbeobtainedfromoneplant.This impliesa lengthyselectionprocess,andinfactittakesaboutsixyearsfrom the time the botanical seed is ger-minated until enough planting material isavailableformultilocationreplicatedtrials.
Table 1 illustrates a typical selectioncycleincassava.Itbeginswiththecrossingof elite clones and finishes when the fewclonessurvivingtheselectionprocessreachthe stage of regional trials across severallocations. It should be emphasized thatthere is some variation among the fewcassava-breedingprogrammes in theworldwith respect to the number of genotypes
table 1typical selection cycle in cassava beginning with the crossing of elite clones to the point when few clones surviving the selection process reach the stage of regional trials across several locations
year activity number plants per genotype
1-2 crosses among elite clones planned, nurseries planted and pollinations made
Up to 100 000
3 F1: evaluation of seedlings from botanical seeds. Strong selection for african cassava mosaic virus (acMV) in africa.
100 000a; 50 0000b; 50 000 c
1
4 clonal evaluation trial (cet) 20 000–30 000 a, b 700 c 6–8 (1 rep, 1 location)
5 Preliminary yield trial (PYt) 100 a; 300 b; 80 c 20–60 (3 reps, 1 location)
6 advanced yield trial (aYt) 25 a; 100 b; 20–25 c 100–500 (3 reps, 2–3 location)
7-9 Regional trials (Rt) 5-30 a, b, c 500-4 000 (3 reps, 3–4 locations)
Figures for cassava breeding at a iita (ibadan, nigeria); b ciat (cali, colombia) and c ciat and Rayong Field Research Station from Department of agriculture (thailand).Source: adapted from Jennings and iglesias, 2002.

Chapter 7 – Marker-assisted selection in common beans and cassava 95
and plants representing them through thedifferentstages.Table1alsoprovidesanideaoftheselectionpressuresgenerallyapplied.
Strong emphasis on highly heritabletraits (plant type, branching habits andreactiontodiseases,harvest indexanddrymattercontent) isappliedduringtheearlyphases of selection (F1 and CET), (Hahn,HowlandandTerry,1980;Hahn,TerryandLeuschner, 1980; Hershey, 1984; Kawano,2003;Ceballoset al.,2004).Asthenumberof plants representing each genotypeincreases, the weight of selection criteriashifts towards low heritability traits suchas root yield. The clones that show out-standingperformanceintheregionaltrialsare released as new varieties and, eventu-ally,incorporatedasparentsinthecrossingnurseries. With that the selection cycle isfinishedandanewonebegins.Thewholeprocess has the following characteristics(Ceballoset al.,2004):• theprocessisindeedphenotypicselection
becausenofamilydataareinvolved;• nodataarecollectedintheearlystages
of selection. Therefore, data regard-ing general combining ability effects(∼breedingvalue)arenotavailableforabetterselectionofparentalmaterials;
• there is no proper separation betweengeneral (GCA∼additive) and specific(SCA∼heterotic) combining abilityeffects. The outstanding performanceofselectedmaterialsis likelytodependonpositiveheteroticeffectsthatcannotbe transferred to theprogenies thataresexuallyderivedfromthem;
• noinbreedingisincorporatedpurposelyin the selection process. Therefore,large genetic loads are likely to remainhiddenincassavapopulationsandusefulrecessivetraitsaredifficulttodetect;
• several stages of selection are based onunreplicatedtrials.Alargeproportionof
genotypes iseliminatedwithoutproperevaluation.For the above-mentioned reasons, cas-
sava breeding is difficult, expensive andtoacertaindegreeinefficient(Perezet al.,2005a;Cachet al.,2005a,b).Kawanoet al.(1998)mentionthat,duringa14-yearperiodabout 372 000 genotypes derived from4130crosseswereevaluatedat theCIAT-RayongFieldCropResearchCenter.Onlythreegenotypesemergedfromtheselectionprocess to be released as official varieties.Similar experiences have been observedat the International Institute of TropicalAgriculture (IITA), CIAT-Colombia andBrazil. Therefore, the development andadaptation of molecular tools for cas-sava genetic improvement offer importantadvantages tomake theprocessmoreeffi-cientandeffective.
maS in cassava breedingCassavageneticimprovementcanbemademore efficient through the use of easilyassayable molecular genetic or DNAmarkers (MAS) that enable the preciseidentification of genotype without theconfounding effect of the environment,thereby increasing heritability. MAS canalsocontributetotheefficientreductionoflarge breeding populations at the seedlingstagebasedupon“minimumselectioncri-teria”.This isparticularly importantgiventhe lengthof thegrowingcycleofcassavaandtheexpenseinvolvedintheevaluationprocess.Therefore,apre-selectionattheF1phase (see Table 1) could greatly enhancethe efficiency of the CET experiments.Theselectionofprogeniesbasedongeneticvaluesderivedfrommolecularmarkerdatasubstantially increases the rate of geneticgain, especially if the number of cycles ofevaluation or generations can be reduced(Meuwissen,HayesandGoddard,2001).

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish96
AnotherapplicationofMAS incassavabreeding is reducing the length of timerequiredfortheintrogressionoftraitsfromwildrelatives.Wildrelativesareimportantsources of genes for pest and diseaseresistance incassava (Hahn,HowlandandTerry, 1980; Hahn, Terry and Leuschner,1980; Chavarriaga et al., 2004), but theneed to reduce or eliminate undesirabledonorgenomecontentandlinkagedragcanlengthen theprocess,making itunrealisticformostbreeders.SimulationsbyStamandZeven (1981) indicate that markers couldreduce linkage drag and would reducethe number of generations required inthe backcross scheme. Hospital, Chevaletand Mulsant (1992) corroborated this inachieving a reduction of two backcrossgenerations with the use of molecularmarker selection. Frisch, Bohn andMelchinger (1999), through a simulationstudy,foundthatuseofmolecularmarkersfortheintrogressionofasingletargetallelesaved two to four backcross generations.They inferred that MAS had the potentialtoreachthesamelevelofrecurrentparentgenome in generation BC3 as reached inBC7withoutmolecularmarkers.
The decision to employ DNA-basedmarkers in cassava breeding is primarilybasedon theheritabilityofa traitand theamount of genotypic variance explainedbythemarker.Therearemanyinstancesincassava breeding where h2 is low or zero.
Someexamplesare:• plant health traits where the pathogen
or pest pressure is absent or low, suchascassavamosaicdisease(CMD)intheNew World tropics or cassava greenmite(CGM)duringthewetseason;
• variableorerraticpestpressure,e.g.theCGM or diseases such as the cassavafrogskindisease(FSD);
• evaluationbaseduponasingleplant;
• variableexperimentalfieldsand/orpoormanagement resulting in large experi-mentalerrors;
• traits that are affected by the stage ofplant growth or the part of the organusedfortissueanalysis,e.g.cyanogenicpotential.In the above-mentioned instances,
having a marker(s) that explains a largeproportionofthegeneticvariancecanaccel-erateprogressinbreeding.Evenwhereh2ismoderateorhigh,selectionbymarkerscanbeadvantageous:• where different sources of genes exist
for the trait that are indistinguishableby phenotype alone and pyramidingis difficult and time consuming, e.g.for different sources of resistance to adiseaseorpest;
• where molecular tags that can be usedinexpensively and rapidly to identifydesirable genotypes early in thebreedingcycleexist,therebyeliminatingthe need to evaluate large numbers ofplantsphenotypically,andobviatingtheconfoundingeffectsoftheenvironment.Markers may permit the efficientelimination of undesirable genotypesat the seedling stage. For example, thenumber of genotypes at the seedlingstage canbe reducedby50percent if atrait is controlled by a single gene, orby 87.5percent if controlled by threegenes;
• for the introgression of useful genesfrom exotic germplasm into adaptedgenepools.MAScanbeusedtoidentifygenotypes that carry minimal amountsofflankingdonorparentgenomearoundthe gene of interest for faster back-crossing;
• for definition of heterotic pools in agroupofgermplasmaccessionsformoredirectedcrosses;

Chapter 7 – Marker-assisted selection in common beans and cassava 97
• fordefinitionofaverageheterozygosityintheselectionofpartially inbredlinesfortolerancetoinbreeding;
• for identification of the male parentin elite germplasm derived from poly-crosses by fingerprinting. This tool isalso useful for checking the identityof different genotypes to eliminateduplicationingermplasmcollections.Best results are achieved when MAS is
combined with phenotypic data as com-pared with either approach independently(Hospital, Chevalet and Mulsant, 1992).Phenotypic data would reduce the cost ofgenotyping especially if phenotypic eval-uation is conducted on early generations(GimelfarbandLande,1994).ThisnotonlyreducesthecostofMASbutalsoincreasesits efficiency. Some examples of MAS incassava breeding conducted at an interna-tionalcentreandnationalprogrammesaredescribedbelow.
Molecular MAS for CMD resistance at an IARCAn ideal target for MAS is breeding fordisease resistance in the absence of thepathogen. This is the case of CMD in theAmericas,wherethediseasedoesnotoccur.CMD is a viral disease first reported byWarburgin1894ineasternAfrica(quotedbyStoreyandNichols,1938).Severalvar-iants of the disease (East Africa cassavamosaicvirus[EACMV],SouthAfricacas-savamosaicvirus[SACMV],Indiancassavamosaic virus [ICMV]) have been reported(Swanson and Harrison, 1994) and areendemic in all cassava growing regions ofAfrica and southern India, where it is themost severe production constraint. Thewhite fly vector of CMD, Bemisia tabacibiotype A, does not colonize cassava intheNewWorldbutrecentlyanewbiotypeofB. tabaci,biotypeB(alsoreferredtoas
B. argentifolia), has become widespreadintheAmericasandhasawidehostrangeincluding cassava (Polston and Anderson,1997),increasingthepossibilitythatCMD,EACMV, SACMV, ICMV or a nativeAmericangeminiviruswillbecomeestab-lished on cassava in the neo-tropics. Thisis a frightening prospect for cassava pro-ductioninLatinAmerica,consideringthatmostLatinAmericancassavagermplasmisvery susceptible to CMD (Okogbenin et al.,1998).Thesusceptibilityofneo-tropicalgermplasmtoCMDalsolimitstheutiliza-tion of germplasm from the crop’s centreofdiversityintheneo-tropicsforthesekeycassava production regions. Breeding forresistancetoCMDinLatinAmerica,wherethediseasedoesnotexistandisunlikelytobeintroducedduetoverystrictquarantinecontrols,requiresthetoolsofMAS.
EvaluationsatIITAidentifiedanexcel-lent sourceof resistance toCMDinsomeNigerian landraces (A.G.O. Dixon 1989,unpublished data), namely TME3, TME7,TME5, TME8, TME14 and TME28. Thisresistance is effective against all knownstrains of the virus, including the viru-lentUgandanvariant(UgV)(Akanoet al.,2002;CIAT,2001).CIAT,incollaborationwith IITA in Ibadan, Nigeria, and withsupport from the Rockefeller Foundation,devel-oped several molecular markers forthis source of CMD resistance, revealedto be controlled by a single dominantgene designated as CMD2 (Akano et al.,2002).At least fivemarkers tightlyassoci-ated to CMD2 have been developed, theclosest being RME1 and NS158 at dis-tances of four and seven cM respectively.The dominant nature of CMD2 and its effectiveness against a wide spectrum ofviral strains makes its deployment veryappealing for protecting cassava againstthe actual or potential ravages of CMD

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish98
in both Africa and Latin America. CIATand IITA undertook a project to verifythe utility of these markers for MAS inbreeding CMD resistance by developingcrossesbetween thesourcesofTME3andsusceptiblevarieties.Atotalofsixfamilies,ranginginsizefrom36–840genotypes,anda total of 2490genotypes were used. Thecrossesweregenotypedwith twomarkersand also evaluated for CMD resistancein a high CMD pressure area in Nigeria.Results of the marker analysis and phe-notypic evaluation of CMD resistance inthe field revealed that the markers RME1and NS158 SSR were excellent predictiontools for CMD resistance in some crosses(apredictionaccuracyof70–80percent).Ina few families,however, themarkerswerenotpolymorphicbetweentheresistantandsusceptibleparentand,therefore,werenotuseful.Thishighlightstheneedtodevelopmanymarkersaroundageneofinterestina MAS programme and then to use thosemarkers to evaluate the parents and iden-tifythebestmarkersforthedifferentcrosscombinations.
EighteenprogeniesfromTME3carryingthe CMD2 marker were established fromembryo axes and imported to CIAT fromIITA.1Theywerecrossedextensivelytoeliteparents. Seeds harvested from the crossesweregerminatedin vitrofromembryoaxesaccordingtostandardprotocolsforcassava(Fregeneet al.,1997,CIAT,2002)toallowsharingtheCMDresistantgenotypeswithcollaborators in Africa and India. Eachplantlet was multiplied after three to fourweeks of growth to obtain three to fiveplants.Afteranotherfourweeks,leavesofall
1 Phytosanitary conditions for the exchangeof cassava germplasm between Africa and Asiaare very stringent, but appropriately indexed in vitro cultures of embryo axes are permitted forexperimentalpurposes.
plantswereremovedformolecularanalysisand the plants multiplied again to obtain10–20 plantlets. DNA isolation was by arapid mini preparation method developedforrice(Nobuyukiet al.,2000).TheDNAobtainedissufficientfor100reactionsandcan be held in the Costar plates for twomonthsat–20oCwithoutanydegradation.PCR amplification, polyacrylamide gelelectrophoresis (PAGE) or agarose gelanalysisofSSRmarkersNS158andRME1wereasdescribedbyMbaet al.(2001).Theversatilityof spreadsheetsmakes themtheappropriatesoftwaretohandle thediverseinformationgeneratedbyMAS.Gelimagesfrom the marker analysis were entereddirectly into a spreadsheet that containsinformation on the parents, tissue cultureand greenhouse records, and subsequentphenotypic evaluation of the progenies.After molecular analysis, genotypes thatcarry the marker allele associated withCMD2 were further multiplied to obtainat least 30 plants. Ten plants were sent tothe greenhouse for hardening and latertransferredtothebreedingprogrammeforevaluation. Five plants were kept in vitro,while15plantswereshippedtopartnersinIndiaandAfricaasshownintheflowchartforMAS(Figure4).
Todate,morethan50000progenyhavebeenevaluatedwithCMDlinkedmarkersand resistant lines shared with nationalprogrammes in India or Africa, and alsoincorporated into the breeding scheme atCIAT. The cost of a single marker datapointisUS$0.30and32000samplescanbeprocessedinayear.
MAS for CMD resistance at a NARS Although evaluation for CMD resistanceinsub-SaharanAfricaisrelativelyeasyandmostareashave sufficientdiseasepressureto permit moderate to high heritability

Chapter 7 – Marker-assisted selection in common beans and cassava 99
of resistance, overlapping outbreaks ofCGM,cassavabacterialblight (CBB), andCMDarecommon(LeggandOgwal,1998)and the need for modest-sized breedingpopulations make MAS for CMD resist-ance a powerful tool to accelerate cassavaimprovement even in Africa. A MAS andparticipatoryplantbreeding (PPB)projectwas initiated in 2003 with funding fromthe Rockefeller Foundation to improvethe resistance of local cassava varieties intheUnitedRepublicofTanzania toCMDand CGM and also to provide proof ofconcept for the use of MAS to acceleratecassavaimprovement.TheUnitedRepublicof Tanzania is the fourth largest producer
of cassava in Africa with average yieldsofabout8tonnes/ha(FAO,2001).This isbelowthecontinent’saverageof10tonnes/ha, and well below the average yield of14 tonnes/ha of Africa’s (and the world’s)largestproducer,Nigeria.
The low yield in the United Republicof Tanzania is caused by many factors,including the susceptibility of commonlygrownvarietiestomajordiseasesandpestssuchasCMDandthecassavabrownstreakdisease(CBSD).Theprojectcrossesfarmer-preferred germplasm, by agro-ecology, toimproved introductions that are resistanttoCMDandtoCGM.MarkersassociatedwithresistancetoCMDareusedtoreduce
FiGURe 4 Schematic representation of steps employed in breeding for resistance to the Cmd
in latin america cassava gene pools
the entire process from sexual seeds to tissue plants for shipment or transfer to the screen house takes approximately three months.
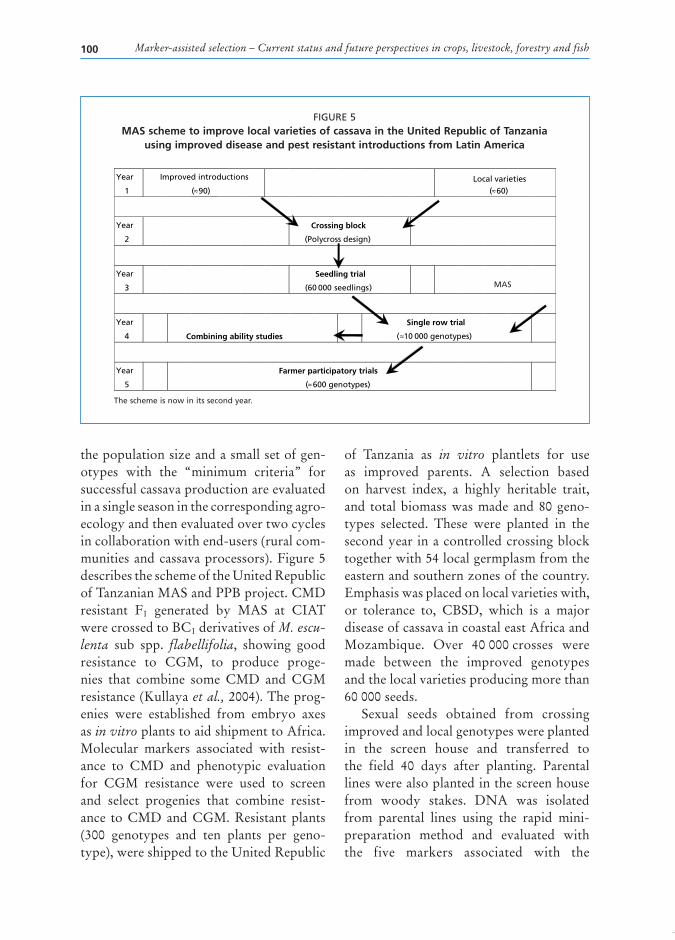
Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish100
the scheme is now in its second year.
thepopulationsizeandasmallsetofgen-otypes with the “minimum criteria” forsuccessfulcassavaproductionareevaluatedinasingleseasoninthecorrespondingagro-ecologyandthenevaluatedovertwocyclesincollaborationwithend-users(ruralcom-munitiesandcassavaprocessors).Figure5describestheschemeoftheUnitedRepublicofTanzanianMASandPPBproject.CMDresistant F1 generated by MAS at CIATwerecrossedtoBC1derivativesofM. escu-lenta sub spp. flabellifolia, showing goodresistance to CGM, to produce proge-nies that combine some CMD and CGMresistance(Kullayaet al.,2004).Theprog-enies were established from embryo axesasin vitroplantstoaidshipmenttoAfrica.Molecular markers associated with resist-ance to CMD and phenotypic evaluationfor CGM resistance were used to screenand select progenies that combine resist-ance to CMD and CGM. Resistant plants(300 genotypes and ten plants per geno-type),wereshippedtotheUnitedRepublic
of Tanzania as in vitro plantlets for useas improved parents. A selection basedon harvest index, a highly heritable trait,and totalbiomasswasmadeand80geno-types selected. These were planted in thesecondyear inacontrolledcrossingblocktogetherwith54localgermplasmfromtheeasternandsouthernzonesofthecountry.Emphasiswasplacedonlocalvarietieswith,or tolerance to, CBSD, which is a majordiseaseofcassavaincoastaleastAfricaandMozambique. Over 40000crosses weremade between the improved genotypesandthelocalvarietiesproducingmorethan60000seeds.
Sexual seeds obtained from crossingimprovedandlocalgenotypeswereplantedin the screen house and transferred tothe field 40 days after planting. Parentallineswerealsoplantedinthescreenhousefrom woody stakes. DNA was isolatedfrom parental lines using the rapid mini-preparation method and evaluated withthe five markers associated with the
FiGURe 5maS scheme to improve local varieties of cassava in the united republic of tanzania
using improved disease and pest resistant introductions from latin america
Year
1
Improved introductions
(≈90)
Year
2
Crossing block
(Polycross design)
Year
3
Seedling trial
(60 000 seedlings)
Year
4
Combining ability studies
Single row trial
(≈10 000 genotypes)
Year
5
Farmer participatory trials
(≈ 600 genotypes)
Local varieties(≈60)
MAS

Chapter 7 – Marker-assisted selection in common beans and cassava 101
CMD2 mediated resistance to CMD.Polymorphism in pair-wise combinationsof the parental lines was observed with atleast one of the five markers and will beused on the progeny. The phenotype oftheprogenywillbeevaluatedatthreeandsix months after planting for resistanceto CMD, CBSD and CGM. Markers arecurrentlybeingtestedforCGMresistanceand are being developed for resistance toCBSD; when their utility is confirmed,theywillalsobeusedonprogenies.
Using published broad sense herita-bility of 0.6 for CMD resistance (Hahn,Terry and Leuschner, 1980), it is expectedthat 24 000 symptomless genotypes willbe analysed with markers associated withresistancetoCMD.ThegainofMASwillbe the elimination of at least 38 400 (4800 x 8 plants) that would have been car-ried to the single row trial stage (eightplant-rowspergenotype),consideringthatbreeders traditionally select 20percent atthe seedling trial stage. This representsa reduction of about 4 ha at the CET. Ifmarkerscanbeusedtoselectforresistanceto CGM and CBSD, then an additionalnumber of genotypes can be eliminatedfromtheCETleadingtoevengreatersav-ings. Using MAS for CMD alone wouldreducethesizeoffieldtrialsby50percent.If additional second and third traits wereincluded,reductionscouldbeashighas75and 87.5percent, respectively. Perhaps themostimportantadvantage,however,comesfromtheincreasedgeneticgainarisingfromhigher heritabilities in these field evalua-tionswithfewergenotypes.
MAS for transferring useful traits from wild relatives of cassava into the cultivated gene poolWildManihot germplasmoffersawealthofusefulgenesforthecultivatedM. esculenta
species, but its use in regular breedingprogrammesisrestrictedbylinkagedragandthe long reproductive breeding cycle. Forexample, several accessions of M.esculentasub spp. flabellifolia, M. peruviana andM. tristis have high levels of proteins(Nichols,1947; Asiedu et al., 1992; CIAT,2004). Low amylose content starch (3–5percent) or waxy starch of relevance tothe cassava starch industry has also beenidentified in two wild relatives of cassava,namelyM. crassisepalaandM. chlorostricta. The only source of dramatically delayedpost-harvest physiological deterioration(PPD)hasbeenidentifiedinaninterspecifichybrid between cassava and M. walkerae.The M. walkerae parent was collectedin Mexico and held at the WashingtonUniversity, St. Louis, United States ofAmerica (Bertram, 1993). It was broughtto CIAT in 1998 in an attempt to useit in improving PPD. Furthermore, theonly source of resistance to the cassavahornworm and the most widely deployedsource of resistance to CMD wereidentified in fourth backcross generationprogenies of M. glaziovii (Jennings, 1976;Chavarriagaet al.,2004).Moderatetohighlevels of resistance to CGM, whitefliesandthecassavamealybughavebeenfoundin interspecific hybrids of M. esculentasub spp. flabellifolia. The delayed PPDtrait and resistance to the pests weresuccessfully transferred to F1 interspecifichybrids suggesting dominant or additivegeneactionofthegene(s)involved(CIAT,unpublisheddata).
Thelongreproductivecycleandlengthytime required to develop new cassavavarieties (10–15 years) often discouragestheuseofwildspeciesinmostconventionalcassava breeding programmes. However,theuseofmolecularmarkerstointrogressa single target region of the genome can

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish102
save between two to four backcrossgenerations (Frischet al., 1999). Indeed,it has been shown in several crops thatthe“tremendousgeneticpotential” lockedup in wild relatives can be released moreefficientlythroughtheaidofnewtoolsofmolecular genetic maps and the advancedbackcross QTL mapping scheme (ABC-QTL)(TanksleyandMcCouch,1997).
Forseveralyearsnowmolecularmarkertools and a modified ABC-QTL schemehave been tested in cassava at CIAT fortheintrogressionofusefulgenesfromwildrelatives.TheschemeentailsgeneratingBC1crosses and carrying out QTL mappingfollowedbyselectionofgenotypescarrying
thegenomeregionofinterestwithminimumsegments of the donor genome (Figure6).The modified ABC-QTL is currentlybeingusedatCIATtointrogressgenesforhighproteincontent,waxystarch,delayedPPD, and resistance to whiteflies and thehornworm. The most advanced of theseMAS projects is the introgression of highproteincontentfromclosewildrelativesofcassava.TwoBC1 familiesofbetween250and 300 progenies were developed fromtwo accessions of M. esculenta sub spp.flabellifolia OW284-1 and OW231-3, andtheimprovedcassavavarietyfromThailandRayong 60 (MTAI 8 in the germplasmcollection).TheBC1familieswereplanted
FiGURe 6modified advanced backcross qtl scheme in the introgression of useful traits from
wild relatives into cassava
To crossing block with elite or local varietiesTocrossingblockwitheliteorlocalvarieties

Chapter 7 – Marker-assisted selection in common beans and cassava 103
in a CET for evaluation of root proteincontentattenmonths.ThegrandparentallinesoftheBC1populationweregenotypedwithover800simplesequencerepeat(SSR)markersavailableforcassavaandabout300polymorphicmarkerswere identified.Thepolymorphicmarkersarebeingassayed intheprogeniesafterwhichQTLanalysiswillbeconductedusingthephenotypicproteinandmolecularmarkerdata.GenotypesthathaveQTLforproteinandaminimumofthedonor parent genome will be selected andusedforproducingtheBC2generation.
Forintrogressionofnaturallyoccurringmutant granule-bound starch synthase(GBSSI) for waxy starch in wild relatives,a more targeted approach was taken.Sequencingoftheglycosyltransferaseregionof theGBSSIgene fromthewild relativesand two cassava accessions identified foursingle nucleotide polymorphisms (SNPs)thatdifferentiatedthewildaccessionsfromcassava. Allele-specific molecular markersunique to these SNPs were developed forselection of these alleles in a breedingscheme.
Genetic crosses were made betweenM. chlorosticta accession CW14-11 andMTAI8, and the resulting F1 was back-crossed to MTAI8. The allele specificmarker will be used together with otheragronomictraits,particularlyperformance,to select for BC1 that carry the mutantGBSSallelesforself-pollinationtorecoverthewaxytrait.Theidentificationofnaturalmutantsinakeygeneanddevelopmentofmarkersrepresentaninnovativemoleculartool to accelerate the introgression offavourable alleles from wild relatives intocassava. Backcross derivatives have alsobeen developed from M. walkerae (MWal001)fordelayedpost-harvestphysiologicaldeterioration; from MNG11 (a BC4derivativeofM. glaziovii)forresistanceto
hornworm;andfromM. esculentasubspp.flabellifolia (FLA447-1) for resistance towhiteflies.Phenotypicandgeneticmappingof these backcross populations are inprogresstobefollowedbyidentificationofQTLandselectionofprogeniestogeneratethenextgeneration.MASwilllaterbeusedtocombinethesegenesintoprogenitorsforuseasparents inbreedingwhich, togetherwith low cost marker technologies, willbe distributed extensively to nationalprogrammes in Africa, Asia and LatinAmericatoproduceimprovedvarieties.
Marker-assisted estimation of average heterozygosity during inbreeding of cassavaA principal use of molecular markers byprivate sector breeding companies is toacceleratethedevelopmentofinbredlines.Cassava genotypes are heterozygous andvery little inbreeding has been practisedto date. However, inbred lines arebetter as parents as they do not have theconfoundingeffectofdominanceandcarrylower levels of genetic load (undesirablealleles).Speedofinbreedingdependsuponthe average heterozygosity of the originalparental lines, the homozygosity level ofthe selected genotypes at the end of theself-pollinating phase and the process ofselectionofprogeniestobeself-pollinated(Scotti et al., 2000). Basically in theinbreedingprocesstwoeventsgotogether:phenotypicallythereisadecreaseinvigour,whichiscorrelatedwiththeincreasedlevelsofhomozygosity.Whiletheaimistoselectvigorousplants(toleranttoinbreeding),intheprocessplantsmaybeselectedthatarelesshomozgygousthantheexpectedaveragefortheirgeneration.Itisexpectedthatthefirstfewcyclesofself-pollinationwillresultinamarkedreductionofvigour(inbreedingdepressionassociatedwiththegeneticloadoftheparentallines);therefore,selectionfor

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish104
tolerancetoinbreedingdepressionmustbeexerted. However, such selection is biasedby the differences in homozygosity levelsof segregating partially inbred genotypes.This highlights the need for a methodto measure the level of heterozygosityin these partially inbred individuals andto use this in a co-variance correction inthe selection of phenotypically vigorousgenotypes.Molecularmarkerscanbeusedto estimate the level of homozygosity ofa given plant, enabling selection of plantswithtruetolerancetoinbreeding.
Molecularmarkerscan identify regionsin thegenomethatareparticularlyrelatedtotheexpressionofheterosisandformeas-uringgeneticdistancesamonginbredlinestodirectcrosseswithhigherprobabilitiesofhigh heterosis. Co-dominant SSR markerson a genome-wide basis are suitable forthispurpose.Theeffectof self-pollinationonvigourandheterozygositywasanalysedin nine S1 families, heterozygosity beingestimatedintheS1familiesby100mappedSSRmarkersthatcoverover80percentofthecassavagenomeandplantvigourbydryroot yield and plant biomass. Results willassist in selecting the best performing andleastheterozygousplantsduringinbreedingby identifying superior partially inbredparental lines. Molecular markers couldalsobeusedtodelineateheteroticgroupsincassava. Genetic resources of cassava havebeencharacterizedattheregional(Fregeneet al., 2003) and global (Hurtado et al.,2005) levels. Highly differentiated groupsof accessions were observed particularlyamonggroupsofmaterialsfromGuatemalaandAfricaandtheymayrepresentheteroticpools. These groupings are being testedbased on molecular markers by geneticcrossing between and within the groupsas a first step to define heterotic patternsfor a more systematic improvement of
combining ability via recurrent reciprocalselection.
Other potential MAS targetsSeveralothertraits forwhichMAScanbeapplied to increase efficiency of breedinginclude:
Beta-caroteneCIATandanumberofpartnersareinvolvedinaprojecttoproducecassavavarietieswithhigherlevelsofβ-caroteneinyellowroots.Thisisonewayofcombatingthedeficiencyof this key micronutrient in areas wherecassavaisamajorstaple.Theexperimentalapproach to increasing cassava β-carotenecontentincludesconventionalbreedingandgenetictransformation.Thediscoveryofawidesegregationpatternof rootcolour intwoS1familiesfromtheColombianlandraceMCOL72(crosscodeAM273)andMTAI8 (AM 320) was the basis for moleculargenetic analysis of β-carotene content incassava. Three markers, SSRY251, NS980andSSRY330,werefoundtobeassociatedwithβ-carotene content. These are in thesame region of the genome and togetherexplain>80percentofphenotypicvariationfor β-carotene content in the populationusedforthisstudy.Thehomozygousstateofcertainallelesofthesemarkerstranslatesinto higherβ-carotene content, suggestingthatbreedingforthistraitcanbenefitfrommolecular markers to assist in combiningfavourable alleles in breeding populations.The work is continuing with the searchforadditionalfavourablealleles inyellow-rootedgermplasmtogivethebestpossiblephenotypicexpressionofthetrait.
Cyanogenic potential AcollaborativeprojectbetweentheSwedishUniversityofAgriculturalSciences(SLU),Uppsala, the Medical Biotechnology

Chapter 7 – Marker-assisted selection in common beans and cassava 105
Laboratories (MBL), Kampala, and CIAT,is aimed at the genetic mapping of CNPin cassava. An S1 family-AM 320, derivedfrom the bitter variety MTAI 8 is thebasis for the study. This family has beenevaluated for cyanogenic glucoside con-tent and has been genotyped with morethan200diversityarraytechnology(DarT)markers at CAMBIA, Australia, and 150SSR markers at CIAT. The discovery ofmolecular markers for CNP will providea tool to select efficiently for low cyano-genicpotential incassava.Alsoongoing isthegeneticmappingofthetwocytochromeP450genesCYP79D1andD2thatcatalysethe rate-limiting step of the biosynthesisof the cyanogenic glucosides, linamarin inthe S1 family AM 320. The group is alsolooking for an association with QTL forCNP.ItisexpectedthatmarkersassociatedwithCNPwillbe identified at the endofthestudy.
Dry matter contentFew key traits in cassava hold greaterpotential for increasing cost-effectivenessvia MAS than root dry matter content(DMC). This trait is usually measured attheendofthegrowthcycle.AnumberofgeneticandenvironmentaleffectsinfluenceDMC.Itisusuallyhighestbeforetheonsetof rains, but drops after the rains beginas the plant mobilizes starch from therootsforre-growthofleaves(Byrne,1984).Defoliation from pest and disease attackscan lower DMC. Breeding programmeshave been quite successful in improvingDMC, especially for industrial markets.The entry point for developing markersassociated with DMC was recent diallelexperiments (Jaramillo et al., 2005; Calleet al., 2005; Pérez et al., 2005a, b; Cachet al., 2005b). Diallels, in this case madeup of 90 families, are an ideal method
to identify genes controlling DMC thatare useful in many genetic backgrounds.Estimatesofgeneralandspecificcombiningability (SCA and GCA, respectively) formany traits of agronomic interest werecalculated,withemphasisonDMC.BasedonGCAestimates,parentswereselectedtogenerate larger-sized progenies for DMCmapping. Sizes of families in the originaldiallelexperimentwereabout30progenies,which israthersmall forgeneticmapping.Parallel to the development of mappingpopulations was the search for markersassociatedwithDMCusingtwoF1families,GM 312 and GM 313, selected from thediallelexperimenthavingparentswithhighGCAforDMC.
Initialmarkeranalysisusingbulkedseg-regantanalysisledtothediscoveryoftwomolecular genetic markers, SSRY160 andSSRY150, which explain about 30 and18percent,respectively,ofphenotypicvar-iance for DMC. These markers are beinganalysed on approximately 700 genotypesderivedfrom23crosseswithparentshavinghigh GCA for DMC in order to confirmtheir utility across genetic backgrounds.Parallel to this, larger families are beingdeveloped from selected parents for QTLmappingofDMC.
disadvantages of maSPerhapsthegreatestdisadvantageofMASisthe timeandfinancial investmentrequiredto develop markers that are widely appli-cable for traits of agronomic importance.Often a marker developed in one or afew related genotypes will not work forothergenotypes inabreedingschemedueto allelic effects. Furthermore, develop-ment of markers, particularly for QTL, iscomplicated by epistatic interactions andthe critical need for good quality phe-notypic data. Several ways around this

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish106
problem have been proposed, such as theuseofcandidategenesinvolvedinthetraitsdirectly as selectable markers without theneed for laborious gene tagging experi-ments. However, unravelling the geneticsand the development of markers for suchtraits is still many years down the road.Newmethodsofassociationmappingandlinkage disequilibrium mapping that relyuponnon-randomassociationofcandidategenesormarkersonahighresolutionmapwithaphenotypeofinterestinanon-struc-tured collection of genotypes have beenused extensively in human medicine toidentifygenesinvolvedindisease(CardonandBell,2001).Giventheenormousdiffi-cultiesofquantitativemapping inhumansand the success of association mapping,thesemethodshavealsobeenproposedasways around the problems in developingmarkersforlowheritabilitytraitsinplants(Gaut and Long, 2003). The developmentof (partially) inbredcassavagenetic stockswill certainly accelerate the application ofMAS for the genetic improvement of thecrop.
ConCluSionSGivenlimitedresources,furtherprioritiza-tionoftraitsisneededforthedevelopmentofmarkersiftheydonotalreadyexist.Toppriority should be given to MAS for themostimportantpestsanddiseasesprevalentintheregionforwhichdurablesourcesofresistancegenes exist.Priority shouldalsobe given to DMC as this is another traitthat, althoughhavingahighnarrowsenseheritabilityat the timeof evaluation (usu-ally after the onset of the rains to permitplantingimmediatelythereafter),issignifi-cantly affectedbynon-genetic factors andisnotashighlyheritable.Thereareseveralinitiatives to assist national programmesacquirenewmoleculartoolstoincreasethe
cost-effectiveness of breeding. Prominentamong these are the “molecular breedingcommunities of practice” project of theGeneration Challenge Programme (GCP,www.generationcp.org)andtheRockefellerFoundation-funded African MolecularMarkerNetwork(AMMANET,www.afri-cancrops.net/ammanet).Bothhavetrainingprogrammesonmolecularbreedingthatareopentonationalprogrammescientists.TheCIAT cassava project has also developeda Web-based database resource includingprotocols, populations, and markers forMASincassavathatcaneasilybeaccessedby national programmes (www.ciat.cgiar.org/mascas).
Cassava and common beans: contrasts Cassavaandbeansaresimilarwithrespectto themodest levelof research input theyhaveenjoyedoverthepastthreetofivedec-ades. Both have been part of the researchagenda of CIAT and of the CGIAR fornearly thirty years, and especially beanshave benefited from inputs from labo-ratories and programmes in the UnitedStates of America and, to a lesser degree,Europe.However,researchinvestmentsforhigh-scalegenomicsthroughmarkerdevel-opmentinthesecropshasbeenfarlessthanfor the “super crops” like maize, rice orsoybeanthatenjoyparticipationbythepri-vatesector,butaremorethanminororphancropswithlocalusageinthetropics.
Yet biologically, these two crops arewidely contrasting. Cassava is a perennialversus beans, which are short-seasonannuals, although climbing beans at highaltitudescanbesimilartocassavaingrowthcycle. Beans are an autogamous seed cropwhile cassava is an allogamous crop withvegetativepropagation.Accompanyingthislatter dichotomy are differences in geneaction.Beanspresent largelyadditivegene

Chapter 7 – Marker-assisted selection in common beans and cassava 107
action, while cassava expresses importantcomponents of dominance and epistaticaction. Finally, cassava as a clonal cropcan fix heterotic combinations, while alack of genetic male sterility or apomixissystems in common bean have curtailedthe development of a hybrid industry forthis seed crop even though heterosis isobserved.
In spite of their biological and otherdifferences, the results of several yearsexperience with MAS in beans and cas-savaaresurprisinglysimilar.Inbothcrops,MAS is being employed principally tobolster phenotypic selection for diseaseresistancegenes.Diseaseresistanceisoftengovernedbyrelativelyfewgenes,andphe-notypicdataareobtainedmoreeasily.Onthe other hand, MAS for more complextraits has yet to find ready application.While there are candidates for such traitsin both crops (root bulking in cassava;low phosphorus or drought tolerance inbeans), the complexity of these traits hasmadetheidentificationofreliablemarkersmore difficult and has delayed applica-tion. Obtaining reliable phenotypic dataforcomplextraitsisespeciallydifficultandis often the biggest bottleneck to eventualapplicationofMAS.Inthecaseofcassava,no inbred parents have been used to dateforthedevelopmentofmolecularmarkers,makingthegeneticanalysismoredifficult.
However,somedifferencesintheappli-cation of MAS for the two crops may benoted, arising from the form of repro-duction of each crop. The time frame toselect cassava clones through multiloca-tional trials is about six to seven years.During thisperiodandwitheachstep thenumberofgenotypesisreducedasaresultoftheselectionexerted,butthegenotypeofeachindividualcloneremainsstable.InthecaseofbeansfromtheF1untilstabilization
of pure lines there is an intense segrega-tionprocessintheearlygenerationswhichtapersoffinlatergenerations.InbothcropsMAScanbeusedintheearlystagesoftheselectionprocessbutwithdifferentobjec-tives. In cassava, MAS can help to selectearly on the clone that will ultimately bereleased, whereas in beans MAS is usedto “direct” the segregation process in themore desirable direction. Although mapswithsignificantsaturationareavailableforboth crops, these have been constructedoverseveralyears,employinggenotypes(inthecaseofbeans)fromdifferentgenepoolswithwidepolymorphism.Asmallpropor-tionofthesemarkers(often20–30percent)is polymorphic in other hybrid combina-tionsamongthegenotypeswithinthesamegenepoolorracethathavebeencreatedtotagaspecifictrait.Thus,genomecoverageisoftenstillnotoptimalforthehighqualityQTL analysis that is usually needed forcomplextraits.
reCommendationSCareful prioritization of traits, marker system and genetic stocks for maSThelimitedresourcesavailableforcassavaorbeanresearchrequireajudiciousallocationofefforts.Inthepast10–20yearstherehasbeen increased investment in molecularmarkerresearchinbothcrops.However,aconsiderableproportionofthatresearchwasdirectedatdemonstratingtheusefulnessofdifferenttechniques,e.g.RAPD,restrictionfragment length polymorphism (RFLP),amplified fragment length polymorphism(AFLP), etc. Over this period there hasbeen an ever-changing set of technologiesbut relatively little actual benefit derivedfrom their application. There is a trade-off between being on the cutting edgewiththenewesttechnologiesand“stickingit out” with an “outdated” technology

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish108
until some benefit is extracted from it.On the other hand, far too much efforthas been expended in the identification ofmarkers for traits without carrying thesethroughtoapplication.Oftengenetaggingisacomponentofashort-termproject,anddoesnotreceivethenecessaryfollowupinimplementation.Ineachcase, theessentialquestion is: what are the key genes foreachcrop?Andoncedefined,whichgenesmerittheinvestmenttodevelopmolecularmarkers? For investments in molecularmarker development to yield results, itis important that traits are chosen forwhichmolecularbreedinghasbothaclearadvantageoverfield-basedselectionandisfeasible in theshort tomediumterm.It isalso important that emphasis be given toselectingtherelevantcrosses,pedigreesandpopulationsinwhichtopractiseMAS,andto have in place appropriate phenotypingstrategies for the confirmation of MASresults. In this regard, the use of parentalsurveysofmanyofthegenotypesinvolvedin a given breeding programme is animportantfirststepinimplementingMAS.
Short- and long-term research related to maSThepresentresearchstructure that isnor-mallybasedonshort-termprojects,usuallyof three years’ duration, can seldom beexpectedtodeliverresultsofusablemarkersforcomplextraits.Suchshort-termprojectsthat seek to establish the basis for MASor to implement selection should limittheir objectives to simply inherited traits.On the other hand, longer-term fundingeither of a programmatic or successiveproject funding nature, must be obtainedto address more complex traits governedby QTL as these would normally requireat least two phases of three-year projects.Thefirstphasemightbeexpectedtoreveal
theinheritanceofagiventrait,establishingthelocationandnumbersofQTL,whileasecondphasewouldberequiredtovalidatetheseovermoreenvironmentsand to findmarkersthatarepolymorphicoverawidenumberofgenotypesandthereforewidelyuseful for breeding, as well as adapted torapid laboratory techniques. A medium-to long-term investment likewise impliescareful prioritization of such traits, withregardtopotentialimpactandtheeventualneedforMAS.Thesereflectionsarebasedupon presently available laboratory tech-niques,butastechniquesformoredetailedandwidespreadevaluationoflociandgen-otypes are developed (e.g. gene chips foranalysisofmultipleloci),conclusionscouldchangesignificantly.
Scaling-up technologiesAfterthedevelopmentofmolecularmarkersforatraitandtheirinitialimplementation,a period of scaling-up in use of the spe-cific markers is necessary. Sometimes thisinvolves changes to MAS protocols, inthe marker detection technique or in themarkers themselves. Marker re-design hasbeenacommonelementofscaling-upexer-cises and can involve something as simpleaschangingaPCRfragmentsizetoimple-mentingaSNPassayfortheactualsequencedifferences between alleles. Technologiesthat speed up the implementation processandlowerthecostsassociatedwithscaling-uparecrucialtothesuccessofMASandareoftenneglected.
development of markers that are useful in a large number of crossesOftenamarkerdeveloped foraparticulartraitinoneorafewrelatedgenotypeswillnot work for other genotypes with highvalueofthetraitduetodifferencesingeneoralleliceffects.Unravellingthegeneticsof

Chapter 7 – Marker-assisted selection in common beans and cassava 109
major traits of agronomic interest even ina subsetof eliteparentsused forbreedingis beyond the resources available for beanand cassava research. Association map-ping and linkage disequilibrium mapping,which rely upon non-random associationofcandidategenesormarkersonahighres-olution map with a phenotype of interestin a non-structured collection of geno-types,havebeenproposedasawayaroundthisproblem.Associationmappingcanbeusedtodiscovernewmarker-traitassocia-tions or to validate associations that werefound through conventional genetic map-ping. The GCP is facilitating associationmappingoftraitsofagronomicimportancein cassava and beans with the goal of dis-covering more useful markers for a widerrangeofgenotypes.
the need to strike a balance between maS and field-based selection Occasionally thequestion is raised:whichis better, MAS or conventional selec-tion? This very question betrays a falsedichotomythathindersprogress.Byitself,MAS is seldom an adequate selection tooland therefore must be combined withconventional phenotypic selection. Theobjectiveshouldbetodeveloptheoptimalbalance between conventional and molec-ular breeding, and the “best” balance willbeuniquetoeachsituation,crop,selectionscheme,environmentandopportunitiesfordifferentselectionmethods.Moreemphasisis needed on combined selection systems,ratherthanviewingMASasareplacementforphenotypicorfieldselection.
referenCeSAfanador, L.K. & Hadley, S.D. 1993.Adoptionofamini-prepDNAextractionmethodforRAPD
markeranalysisincommonbean.Bean Improv. Coop.35:10–11.Akano, A.O., Dixon, A.G.O., Mba, C., Barrera, E. & Fregene, M.2002.Geneticmappingofadomi-
nantgeneconferringresistancetocassavamosaicdisease.Theor. Appl. Genet.105:521–525.Asiedu, R., Hahn, S.K., Bai, K.V. & Dixon, A.G.O.1992.Introgressionofgenesfromwildrelatives
intocassava.InM.O.Akoroda&O.B.Arene,eds.Proc. 4th. Triennial Symp. Internat. Soc. Trop. Root Crops-Africa Branch, pp.89-91.Nigeria,ISTRC-AB/IDRC/CTA/IITA.
Awale, H.E. &. Kelly, J.D. 2001.DevelopmentofSCARmarkerslinkedtoCo-42geneincommonbean.Ann. Rep. Bean Improv. Coop.44:119–120.
Babu, L. & Chatterjee, S.R.1999.Proteincontentandaminoacidcompositionofcassavatubersandleaves.J. Root Crops.25:163–168.
Beaver, J.S., Rosas, J.C., Myers, J., Acosta, J., Kelly, J.D., Nchimbi-Msolla, S.M., Misangu, R., Bokosi, J., Temple, S., Aranud-Santana, E. & Coyne, D.P. 2003. Contributions of the bean/cowpea CRSP to cultivar and germplasm development in common bean. Field Crops Res. 82:87–102.
Beebe, S., Velasco, A. & Pedraza, F.1999.MarcajedegenespararendimientoencondicionesdealtoybajofósforoenlasaccesionesdefrijolG21212yBAT881.VI Reunião Nacional de Pesquisa de Feijão.Salvador,Brazil.
Beebe, S., Skroch, P.W., Tohme, J., Duque, M.C., Pedraza, F. & Nienhuis, J. 2000. Structure ofgeneticdiversityamongcommonbeanlandracesofMesoamericanoriginbasedoncorrespondenceanalysisofRAPD.Crop Sci.40:264–273.
Beebe, S., Rengifo, J., Gaitan, E., Duque, M.C. & Tohme, J.2001.DiversityandoriginofAndeanlandracesofcommonbean.Crop Sci.41:854–862.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish110
Beebe, S.E., Rojas, M., Yan, X., Blair, M.W., Pedraza, F., Muñoz, F., Tohme, J. & Lynch, J.P.2006.Quantitativetraitlociforrootarchitecturetraitscorrelatedwithphosphorusacquisitionincommonbean.Crop Sci.46:413–423.
Bellotti, A.C.2002.Arthropodpests.InR.J.Hillocks, J.M.Thresh&A.C.Bellotti, eds.Cassava: biology, production and utilization, pp.209-235.Wallingford,UK,CABIPublishing.
Bellotti, A.C., Arias B., Vargas, V.O., Reyes H.J.A. & Guerrero, J.M. 2002. Insectos y ácarosdañinos a la yuca y su control. In B. Ospina & H. Ceballos, eds. La yuca en el tercer milenio. Sistemas modernos de producción, procesamiento, utilización y comercialización, CIATPublicationNo.327,pp.160-203.Cali,Colombia.
Bertram, R.B. 1993. Application of molecular techniques resources of cassava (Manihot esculentaCrantz, Euphorfiabeae): interspecific evolutionary relationships and intraspecific characterization.UniversityofMaryland,USA.(Ph.D.thesis).
Blair, M.W. & Beaver, J.S.1993.InheritanceofbeangoldenmosaicresistancefrombeangenotypeA429.Ann. Rep. Bean Improv. Coop.36:143–144.
Blair, M.W., Iriarte, G. & Beebe, S.E.2003.QTLanalysisofanAndeanadvancedbackcrosspopula-tionforyieldtraitsderivedfromwildP. vulgaris.Ann. Rep. Bean Improv. Coop.46:7–9.
Blair, M.W., Pedraza, F., Buendía, H.F., Gaitán-Solís, E., Beebe, S.E., Gepts, P. & Tohme, J. 2003.Developmentofagenome-wideanchoredmicrosatellitemapforcommonbean(Phaseolus vulgarisL.)Theor. Appl. Genet.107:1362–1374.
Blair, M.W., Beaver, J.S., Nin, J.C., Prophete, E. & Singh, S.P. 2006.RegistrationofPR9745–232andRMC-3red-mottleddrybeangermplasmlineswithresistancetobeangoldenyellowmosaicvirus.Crop Sci. 46:1000–1002.
Broughton, W.J., Hernandez, G., Blair, M.W., Beebe, S.E., Gepts, P. & Vanderleyden, J.2003.Beans(Phaseolusspp.)–modelfoodlegumes.Plant & Soil252:55–128.
Buendia, H.F., Beebe, S.E., Blair, M.W., Teran, H. & Pedraza, F.2003.IdentificacióndemarcadoresmolecularesasociadosagenesderendimientoenunapoblaciónRC2F3.7defrijolcomúnPhaseolus vulgarisL.(DOR390xG19892).Fitotecnia Colombiana3:57–64.
Buitrago, A.J.A.1990.La yuca en la alimentación animal,p.446.Cali,Colombia,CIAT.Byrne, D.1984.Breedingcassava. Plant Breeding Rev.2:73–134.Cach, N.T., Lenis, J.I., Perez, J.C., Morante, N., Calle, F. & Ceballos, H.2005a.Inheritanceofrel-
evanttraitsincassava(Manihot esculentaCrantz)forsub-humidconditions.Plant Breed.125(2):177–182.
Cach, N.T., Perez, J.C., Lenis, J.I., Calle, F., Morante, N. & Ceballos, H.2005b.Epistasis in theexpressionof relevant traits in cassava (Manihot esculentaCrantz) for sub-humidconditions. J. Heredity96:586–592.
Calle, F., Perez, J.C. Gaitán, W., Morante, N., Ceballos, H., Llano, G. & Alvarez, E.2005.Diallelinheritanceofrelevanttraitsincassava(Manihot esculentaCrantz)adaptedtoacid-soilsavannas.Euphytica 144:177–186.
Cardon, L.R. & Bell, J.I.2001.Associationstudydesignforcomplexdisease.Nature Revs. Genet.2:91–99.
Carvalho, G.A., Paular, T.J., Alzate-Marin, A.L., Nietsche, S., Barros, E.G. & Moreira. M.A.1998.HerançadaresistênciadalinhagemAND277defeijoeiro-comunàraça63-23dePhaeoisariopsis griseola e identificação de marcador RAPD ligado ao gene de resistência. Fitopatol. Bras. 23:482–485.

Chapter 7 – Marker-assisted selection in common beans and cassava 111
Ceballos, H., Iglesias, C.A., Pérez, J.C. & Dixon, A.G.O. 2004.Cassavabreeding:opportunitiesandchallenges.Plant Mol. Biol.56:503–515.
Ceballos, H., Fregene, M., Lentini, Z.J., Sanchez, T., Puentes, Y.I., Perez, J.C., Rosero, A. & Tofiño, A.P. 2006.Developmentandidentificationofhigh-valuecassavaclones.Acta Hort.703:63–70.
Chavarriaga, P., Prieto, S., Herrera, C.J., López, D., Bellotti, A. & Tohme, J.2004.Screeningtrans-genics unveils apparent resistance to hornworm (E. ello) in the non-transgenic, African cassavaclone60444.In A. Alves&J.Tohme,eds.Addingvaluetoasmall-farmercrop.Proc. 6th. Internat. Scientific Mtg. Cassava Biotech. Network,BookofAbstracts,p.4.Cali,Colombia,CIAT.
Chávez, A.L., Sánchez, T., Jaramillo, G., Bedoya, J.M., Echeverry, J., Bolaños, E.A., Ceballos, H. & Iglesias, C.A.2005.Variationofqualitytraitsincassavarootsevaluatedinlandracesandimprovedclones.Euphytica 143:125–133.
CIAT (International Center for Tropical Agriculture).1997.Assessingandenhancingagrobiodi-versitythroughbiotechnology.Annual Report,1994-1997,pp.63–65.Cali,Colombia.
CIAT.2001.Beanimprovementforsustainableproductivity,inputuseefficiencyandpovertyallevia-tion.Annual Report,pp.37–38.Cali,Colombia.
CIAT.2002.Beanimprovementforsustainableproductivity,inputuseefficiencyandpovertyallevia-tion.Annual Report,pp.104–107.Cali,Colombia.
CIAT.2003.Beanimprovementforsustainableproductivity,inputuseefficiencyandpovertyallevia-tion..Annual Report,pp.32–33and67–75.Cali,Colombia.
CIAT.2004.Beanimprovementforsustainableproductivity,inputuseefficiencyandpovertyallevia-tion,Annual Report,pp.84and115–117.Cali,Colombia.
FAO.2001.Food Outlook.December1999.Rome.Ferreira, C.F., Borém, A., Carvalho, G.A., Nietsche, S., Paula, T.J., de Barros, E.G. & Moreira,
M.A. 2000. Inheritance of angular leaf spot resistance in common bean and identification of aRAPDmarkerlinkedtoaresistancegene.Crop Sci.40:1130–1133.
Fregene, M., Angel, F., Gomez, R., Rodriguez, F., Chavarriaga, P., Roca, W., Tohme, J., & Bonierbale, M.1997.Amoleculargeneticmapofcassava(Manihot esculenta Crantz).Theor. Appl. Genet.95:431–441.
Fregene, M., Suarez, M., Mkumbira, J., Kulembeka, H., Ndedya, E., Kulaya, A., Mitchel, S.,
Gullberg, U., Rosling, H., Dixon, A. & Kresovich, S.2003.Simplesequencerepeat(SSR)diver-sityofcassava(Manihot esculentaCrantz)landraces:geneticstructureinapredominantlyasexuallypropagatedCrop Theor. Appl. Genet.107:1083–1093.
Frei, A., Blair, M.W., Cardona, C., Beebe, S.E., Gu, H. & Dorn, S. 2005.Identificationofquantita-tivetraitlociforresistancetoThrips palmiKarnyincommonbean(Phaseolus vulgarisL.).Crop Sci.45:379–387.
Frisch, M., Bohn, M. & Melchinger, A.E. 1999. Comparison of selection strategies for marker-assistedbackcrossingofagene.Crop Sci.39:1295–1301.
Gaut, B.S. & Long, A.D.2003.Thelowdownonlinkagedisequilibrum.Plant Cell15:1502–1506.Gepts, P.1988.AmiddleAmericanandanAndeancommonbeangenepool.In P.Gepts,ed.Genetic
resources of Phaseolusbeans, pp.375-390.Dordrecht,Netherlands,KluwerAcademicPublishers.Gimelfarb, A. & Lande, R. 1994. Simulation of marker-assisted selection in hybrid populations.
Genetical Res.63:39–47.Hahn, S.K., Howland, A.K. & Terry, E.R. 1980.Correlatedresistancetocassavatomosaicandbac-
terialblightdiseases.Euphytica29:305–311.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish112
Hahn, S.K., Terry, E.R. & Leuschner, K.1980.Breedingcassavaforresistancetocassavamosaicdis-ease.Euphytica 29:673–683.
Haley, S.D., Afanador, L. & Kelly, J.D. 1994.IdentificationandapplicationofarandomamplifiedpolymorphicDNAmarkerfortheIgene(potyvirusresistance)incommonbean.Phytopathology84:157–160.
Hallauer, A.R. & Miranda Fo., J.B.1988.Quantitative genetics in maize breeding, second edition.USA,IowaStateUniversityPress.
Hershey, C.H.1984.Breedingcassavaforadaptationtostressconditions:developmentofamethod-ology,pp.303-314.Proc. 6th Internat. Symp. Soc. Trop. Root Crops, Lima,Peru.
Hillocks, R.J. & Wydra, K. 2002. Bacterial, fungal and nematode diseases. In R.J. Hillocks, J.M.Thresh&A.C.Bellotti,eds.Cassava: biology, production and utilization, pp.261–280.USAandUK,CABIPublishing.
Hospital, F. & Charcosset, M.1997.Marker-assistedintrogressionofquantitativeloci.Genetics147:1469–1485.
Hospital, F, Chevalet, C. & Mulsant, P.1992.Usingmarkers ingene introgressionbreedingpro-grams.Genetics132:1119–1210.
Hurtado L., P.X., Ospina, C., Marín, J., Buitrago R.Ch., Fregene, M.A., De Vicente, M.C., Wongtiem, P., Killian, A. & Wenzl, P. 2005. Comparación de dos marcadores moleculares:Microarreglos-microsatélites para el análisis de la diversidad estructural de yuca Manihot escu-lenta Crantz [resumen] [CD-ROM]. In Congreso Asociación Colombiana de Fitomejoramiento y Producción de Cultivos (9, 2005, Palmira, Colombia). [Resúmenes] [CD-ROM]. CorporaciónColombianadeInvestigaciónAgropecuaria(CORPOICA),Cali,CO.p.138.
Ikeda, N., Bautista, N.S., Yamada, T., Kamijima, O. & Ishii, T.2001.Ultra-simpleDNAextractionmethodformarker-assistedselectionusingmicrosatellitemarkersinrice.Plant Mol. Bio. Reporter19:27–32.
Islam, F.M.A., Basford, K.E., Redden, R.J., Gonzalez, A.V., Kroonenberg, P.M. & Beebe, S. 2002.Geneticvariabilityincultivatedcommonbeanbeyondthetwomajorgenepools.Genetic Resources and Crop Evolution49(3):271–283.
Jaramillo, G., Morante, N., Pérez, J.C., Calle, F., Ceballos, H., Arias, B. & Bellotti, A. C.2005.Diallelanalysisincassavaadaptedtothemidaltitudevalleysenvironments.Crop Sci.45:1058–1063.
Jennings, D.L. 1976. Breeding for resistance to African cassava mosaic. African cassava mosaic.Report Interdiscipl. WorkshopIDRC071e,pp.39–44.Muguga,Kenya.
Jennings, D.L. & Iglesias, C.2002.Breedingforcropimprovement,pp.149–166.InR.J.Hillocks,J.M. Thresh & A.C. Bellotti, eds. Cassava: biology, production and utilization. UK, CABIPublishing.
Johnson, W.C., Guzman, P., Mandala, D., Mkandawire, A.B.C., Temple, S., Gilbertson, R.L. & Gepts, P. 1997.Moleculartaggingofthebc-3geneforintrogressionintoAndeancommonbean.Crop Sci.37:248–254.
Jung, G., Skroch, P.W., Coyne, D.P., Nienhuis, J., Aranud-Santana E., Ariyarathne, H.M., Kaeppler, S.M. & Bassett, M. 1997. Molecular marker-based genetic analysis of tepary bean-derived common bacterial blight resistance in different development stages of common bean. J. Am.. Soc. Hort. Sci.122:329–337.
Kawano, K.1980.Cassava.InW.R.Fehr&H.H.Hadley,eds.Hybridization of crop plants,pp225-233.Madison,WI,USA,ASAandCSSA.

Chapter 7 – Marker-assisted selection in common beans and cassava 113
Kawano, K.2003.Thirtyyearsofcassavabreedingforproductivity-biologicalandsocialfactorsforsuccess.Crop Sci.43:1325–1335.
Kawano K., Narintaraporn, K., Narintaraporn, P., Sarakarn, S., Limsila, A., Limsila, J., Suparhan, D., Sarawat, V. & Watananonta, W.1998.Yieldimprovementinamultistagebreedingprogramforcassava.Crop Sci.38:325–332.
Kelly, J.D. & Adams, M.W. 1987.Phenotypicrecurrentselectioninideotypebreedingofpintobeans.Euphytica 36:69–80.
Kelly, J.D., Schneider, K.A. & Kolkman, J.M. 1999.Breeding to improveyield.InS.P.Singh,ed.Common bean improvement in the twenty-first century. Developments in plant breeding,Vol.7,pp.185–222.Dordrecht,Netherlands,KluwerAcademicPublishers.
Kelly, J.D, Gepts, P., Miklas, P.N. & Coyne, D.P.2003.TaggingandmappingofgenesandQTLandmolecular-markerassistedselectionfortraitsofeconomicimportanceinbeanandcowpea.Field Crops Res.82:135–154.
Klimyuk, V.I., Carroll, B.J., Thomas, C.M. & Jones, J.D.G.1993.Alkalitreatmentforrapidprepa-rationofplantmaterialforreliablePCRanalysis.Plant J.3:493–494.
Kullaya, A., Mtunda, K., Kulembeka, H., Ferguson, M., Marin, J., Ospina, C., Barrera, E., Jarvis, A., Morante, M., Ceballos, H., Tohme, J. & Fregene, M.2004.Molecularmarker-assistedandfarmer participatory improvement of cassava germplasm for farmer/market preferred traits inTanzania.6th.Internat. Scientific Mtg. Cassava Biotech. Network.Cali,Colombia,CIAT.
Legg, J.P. & Ogwal, S.1998.ChangesintheincidenceofAfricancassavamosaicvirusdiseaseandtheabundanceofitswhiteflyvectoralongsouth-northtransectsinUganda.J. Appl. Entom.122:169–178.
Liao, H., Yan, X., Rubio, G., Beebe, S.E., Blair, M.W. & Lynch, J.P.2004.Basalrootgravitropismandphosphorusacquisitionefficiencyincommonbean.Funct. Plant Biol.31:1–12.
Mahuku, G., Montoya, C., Henríquez, M.A., Jara, C., Terán, H. & Beebe, S.2004.InheritanceandcharacterizationofangularleafspotresistancegeneincommonbeanaccessionG10474andiden-tificationofanAFLPmarkerlinkedtotheresistancegene.Crop Sci.44:1817–1824.
Mba, R.E.C., Stephenson, P., Edwards, K., Melzer, S., Nkumbira, J., Gullberg, U., Apel, K., Gale, M., Tohme, J. & Fregene, M. 2001. Simple sequence repeat (SSR) markers survey of the cas-sava(Manihot esculentaCrantz)genome:towardsaSSR-basedmoleculargeneticmapofcassava. Theor. Appl. Genet.102:21–31.
Melotto, M., Afanador, L. & Kelly, J.D. 1996.DevelopmentofaSCARmarkerlinkedtotheIgeneincommonbean.Genome 39:1216–1219.
Meuwissen, T.H.E., Hayes, B.J. & Goddard, M.E. 2001. Prediction of total genetic value usinggenome-widedensemarkermaps.Genetics157:1819–1829.
Miklas, P.N., Larsen, R.C., Riley, R. & Kelly, J.D.2000a.Potentialmarker-assistedselectionforbc-12 resistance to bean common mosaic potyvirus in common bean. Euphytica 116(3):211–219.
Miklas, P.N., Smith, J.R., Riley, R., Grafton, K.F., Singh, S.P., Jung, G. & Coyne, D.P. 2000b.Marker-assistedbreedingforpyramidedresistancetocommonbacterialblightincommonbean.Ann. Rep. Bean Improv. Coop.43:39–40.
Miklas, P.N., Stone, V., Daly, M.J., Stavely, J.R., Steadman, J.R., Bassett, M.J., Delorme, R. & Beaver, J.S. 2000c. Bacterial, fungal, and viral disease resistance loci mapped in a recombinantinbredcommonbeanpopulation(‘Dorado’/XAN176).J. Am. Soc. Hort. Sci.125:476–481.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish114
Miklas, P.N., Kelly, J.D., Beebe, S.E. & Blair, M.W. 2006. Common bean breeding for resistanceagainstbioticandabioticstresses:fromclassicaltoMASbreeding.Euphytica147:105–131.
Muñoz, L.C., Blair, M.W., Duque, M.C., Roca, W. & Tohme, J.2004.Levelofintrogressionininter-specific(Phaseolus vulgarisxP. acutifolius)congruity-backcrosslines.Crop Sci.44:637–645.
Nichols, R.F.W.1947.Breedingcassavaforvirusresistance.East Afr. Agric. J.12:184–194.Nietsche, S., Borém, A., Carvalho, G.A., Rocha, C., Paula, T.J., de Barros, E.G. & Moreira, M.A.
2000. RAPD and SCAR markers linked to a gene coferring resistance to angular leaf spot incommonbean.J. Phytopathol.148:117–121.
Nobuyuki, I., Bautista, N.S., Yamada, T., Kamijima, O. & Ishi, T.2000.UltrasimpleDNAextrac-tion method for marker-assisted selection using microsatellite markers in rice. Plant Mol. Biol. Reporter18:1–6.
Okogbenin, E., Porto, M.C.M & Dixon, A.G.O. 1998.InfluenceofplantingseasononincidenceandseverityofAfricancassavamosaicdiseaseinthesubhumiczoneofNigeria.In I.J.Ekanayake,ed.Root crops and poverty alleviation,pp.338–392.Ibadan,Nigeria,IITA.
Park, S.O., Coyne, D.P., Mutlu, N., Jung, G. & Steadman, L.R. 1999. Confirmation of molec-ularmarkersandflowercolorassociatedwithQTLforresistancetocommonbacterialblightincommonbeans.J. Amer. Soc. Hort. Sci.124:519–526.
Pastor-Corrales M.A., Jara, C. & Singh, S.1998.Pathogenicvariationin,sourceof,andbreedingforresistancetoPhaeoisariopsis griseolacausingangularleafspotincommonbean.Euphytica103:161–171.
Pérez, J.C., Ceballos, H., Jaramillo, G., Morante, N., Calle, F., Arias, B. & Bellotti, A.C. 2005a.Epistasisincassava(Manihot esculentaCrantz)adaptedtomid-altitudevalleyenvironment.Crop Sci.45:1491–1496.
Pérez, J.C., Ceballos, H., Calle, F., Morante, N., Gaitán, W., Llano, G. & Alvarez, E. 2005b.Within-familygeneticvariationandepistasisincassava(Manihot esculentaCrantz)adaptedtotheacid-soilsenvironment. Euphytica145:77–85.
Polston, J.E. & Anderson, P.K. 1997.Theemergenceofwhiteflytransmittedgeminivirusesintomatointhewesternhemisphere.Plant Disease81:1357–1369.
Rajendran, P.G., Ravindran, C.S., Nair, S.G. & Nayar, T.V.R.2000.Truecassavaseeds(TCS)forrapid spread of the crop in non-traditional areas. Thiruvananthapuram, Kerala, India, CentralTuberCropsResearchInstitute.22pp.
Santana, G.E., Blair, M.W., Morales, F., Mahuku, G., Jara, C. & Castaño, M.2004.Usodetecnicasclasicasyavanzadasparaidentificargenotiposdefrijolresistentesaantracnosisymosaicocomún.Fitotecnia Colombiana4:44–54.
Sartorato, A., Nietsche, S., Barros, E.G. & Moreira, M.A. 1999.SCARmarkerlinkedtoangularleafspotresistancegeneincommonbean.Bean Improv. Coop.42:23–24.
Schneider, K.A., Brothers, M.E. & Kelly, J.D.1997.Marker-assistedselectiontoimprovedroughttoleranceincommonbean.Crop Sci.37:51–60.
Scotti, C., Pupilli, F., Salvi, S. & Arcioni, S.2000.Variation inVigotandinRFLP-estimatedhet-erozygositybyselfingtetraploidalfalfa:newperspectivesfortheuseofselfinginalfalfabreeding.Theor. Appl. Genet.101:120–125.
Shull, G.F.1952.Beginningsoftheheterosisconcept.InJ.W.Gowen,ed.Heterosis, pp.14–48. Ames,IA,USA,IowaStateCollegePress.

Chapter 7 – Marker-assisted selection in common beans and cassava 115
Singh, S.P. 1999. Integrated genetic improvement. In S.P. Singh, ed. Common bean improvement in the twenty-first century. Developments in plant breeding, Vol. 7, pp. 133–166. Dordrecht,Netherlands,KluwerAcademicPublishers.
Singh, S.P., Gepts, P. &. Debouck, D.G.1991.Racesofcommonbean(Phaseolus vulgaris,Fabaceae).Econ. Bot.45:379–396.
Singh, S.P., Cardona, C., Morales, F.J., Pastor-Corrales, M.A. & Voysest, O. 1998.Gameteselectionforuprightcariocabeanwithresistancetofivediseasesandaleafhopper.Crop Sci.38:666–672.
Stam, P., & Zeven, A.C.1981.Thetheoreticalproportionofdonorgenomeinnear-isogeniclinesofself-fertilizersbredbybackcrossing.Euphytica 30:227–238.
Storey, H.H. & Nichols, R.F.W.1938.Studiesonthemosaicdiseaseofcassava.Ann. Appl. Bio.25:709–806.
Sullivan, J.G. & Bliss, F.A.1983.Expressionofenhancedseedproteincontentininbredbackcrosslinesofcommonbean.J. Am. Soc. Hort. Sci.108:787–791.
Swanson, M.M. & Harrison, B.D.1994.Properties,relationshipsanddistributionofcassavamosaicgeminiviruses.Trop. Sci.34:15–25.
Tanksley, S.D. & McCouch, S.R. 1997.Seedbanksandmolecularmaps:Unlockinggeneticpotentialfromthewild.Science277:1063–1066.
Urrea, C.A., Miklas, P.N., Beaver, J.S. & Riley. R.H. 1996.Acodominantrandomlyamplifiedpoly-morphicDNA(RAPD)markerusefulforindirectselectionofBGMVresistanceincommonbean.J. Am. Soc. Hort. Sci.121:1035–1039.
Vallejo, V. & Kelly, J.D.2001.DevelopmentofaSCARmarkerlinkedtoCo-5geneincommonbean.Ann. Rept. Bean Improv. Coop.44:121–122.
Van Schoonhoven, A. & Pastor-Corrales, M.A. 1987.Standardsystemfortheevaluationofbeangermplasm.Cali,Colombia,CIAT.
Yan, X., Liao, H., Beebe, S.E., Blair, M.W. & Lynch, J.P. 2005.MolecularmappingofQTLsassoci-atedwithroothairsandacidexudationasrelatedtophosphorusuptakeincommonbean.Plant & Soil265:17–29.
Young, R.A., Melotto, M., Nodari, R.O. & Kelly, J.D. 1998.Markerassisteddissectionoftheoligo-genicanthracnoseresistanceinthecommonbeancultivarG2333.Theor. Appl. Genet.96:87–94.


Chapter 8
marker-assisted selection in maize: current status, potential, limitations
and perspectives from the private and public sectors
Michel Ragot and Michael Lee

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish118
SummaryMorethantwenty-fiveyearsaftertheadventofDNAmarkers,marker-assistedselection(MAS) has become a routine component of some private maize breeding programmes.LineconversionhasbeenoneofthemostproductiveapplicationsofMASinmaizebreed-ing,reducingtimetomarketandresultingincountlessnumbersofcommercialproducts.Recently,applicationsofMASforforwardbreedinghavebeenshowntoincreasesignifi-cantlytherateofgeneticgainwhencomparedwithconventionalbreeding.CostsassociatedwithMASarestillveryhigh.Furtherimprovementsinmarkertechnologies,datahandlingand analysis, phenotyping and nursery operations are needed to realize the full bene-fitsofMASforprivatemaizebreedingprogrammesandtoallowthetransferofprovenapproachesandprotocolstopublicbreedingprogrammesindevelopingcountries.

Chapter 8 – Marker-assisted selection in maize 119
introduCtionTheabilitytoidentifygeneticcomponentsof traits,particularlyquantitative traits, inMendelianfactors,andtomonitorordirecttheirchangesduringbreedingthroughtheuse of DNA-based markers has createdmuch enthusiasm. Claims were some-times made that marker-assisted selection(MAS) would rapidly replace phenotypicselectionanddramatically reduce the timerequired to develop commercial varieties(Mazur, 1995). At the turn of this cen-tury, phenotypic selection was still theapproach on which maize breeding pro-grammesmostlyreliedtodevelopnewandimproved cultivars while MAS had con-tributed to advances in introgression, orbackcrossbreeding(Ragotet al.,1995;Ho,McCouch and Smith, 2002; Ribaut, JiangandHoisington,2002;Morriset al.,2003).Overlyoptimistic statements andexagger-atedpromisesaboutthepowerofMAStoimprovecomplextraitscreatedexcessivelyhigh and largely unfulfilled hopes andprompted a wave of cautious and some-times pessimistic views (Melchinger, Utzand Schön, 1998; Young, 1999; GoodmanandCarson,2000;Bernardo,2001).
Recently, multinational corporationswith large maize breeding programmesreported the routine and successful use ofMAS (Johnson, 2004; Niebur et al., 2004;Eathington, 2005; Crosbie et al., 2006).Ratesofgeneticgaintwiceashighasthoseachieved through conventional breedingwerereportedforMASinmaize.AccountswerealsogivenofanumberofMAS-derivedsingle-cross(i.e.simple)hybridsbeingcur-rently on the market. Although too littleis known about the methods (e.g. breed-ingschemes,mathematicalalgorithms)andtools (e.g. marker technologies, computerprograms,databases)usedtodevelopthesehybrids, these results have raised confi-
dence in the ability of MAS to increasethe rate of genetic gain over what can beachieved through conventional breeding.As technologies evolve and marker geno-typesbecomelessexpensive,MASbecomesincreasinglywithinthereachofdevelopingcountries. Whenever necessary, transfer ofmethods or tools from private companiesto developing countries should be madepossible while preserving the commer-cial interests of the companies concerned,therebycontributingtoincreasingtherateofgeneticgainwhereitismostneeded.
Much has happened in maize breedingsinceStuberandMoll(1972)firstreportedthat selection forgrainyield inmaizehadresulted in changes in allele frequenciesat several isozyme loci throughout thegenome. In so doing, they essentially laidthe grounds for MAS in maize. Indeed,if phenotypic selection could produce achange in marker allele frequencies, thenwhy could deliberately altering markerallele frequencies at specific loci not pro-duce predictable phenotypic changes foroneorseveraltraits?
The objectives of this chapter are toprovide the scientific community anddecision-makers with information on thecurrent status of MAS in maize breed-ingprogrammes,includingthemajorstepsthat led to it, and to provide suggestionsto developing countries for deploying thetechnologyandmethodsinvolvedinaneffi-cient,cost-effectiveandrealisticmanner.
how haS maS Been uSed By the private SeCtor to improve the maize Crop?Applications of DNA markers in privatemaize breeding programmes startedin the 1980s with the identification ofDNA clones used to detect restrictionfragment length polymorphisms (RFLPs)

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish120
inthenucleargenome.Asdescribedbelow,the methods used to detect RFLPs wereincompatible with the magnitude, speedand efficiency of all but a few aspects ofselection in maize breeding programmes.Gradually, however, the methods used todetectDNApolymorphismsandtocreatemeaningfulinformationfromDNAmarkerandphenotypicdatasetshaveevolvedtothepoint where they are routine componentsofsomemaizebreedingprogrammesintheprivatesector.
Selection occurs at various stages inmaize breeding programmes. The firstopportunity arises when choosing inbredlinestomateasparentsofnewpopulations.In some programmes, all such inbreds aregenotyped systematically at DNA markerloci(SmithandSmith,1992).Ifthemarkerloci are sufficiently close on genetic orphysicalmapsthenreasonablygoodinfer-ences may be made about the inbred’shaplotype. Such information is used toestablish identity, resolve disagreementsrelatedtogermplasmownershipandacqui-sition,enforce laws intendedtoencouragegenetic diversity of the hybrids and avoidusinginbredsthatcontaintransgeneswhichmay violate regulatory considerations andrestrictions.Theseselectionpractices,whileadmittedly not conventional MAS, haveledto improvements inthemaizecropbyenabling more informed stewardship anddeployment of genetic resources and byproviding a degree of protection of intel-lectualpropertyandrelatedinvestmentsinmaizebreeding.
Unquestionably,themostpervasiveanddirect use of MAS in maize by the pri-vate sector has been with backcrossing oftransgenesintoeliteinbredlines,thedirectparents of the commercial hybrids (Ragotet al.,1995;Crosbieet al.,2006).Currently,the most widely deployed transgenes and
combinations thereof (i.e. gene stacks) arefor resistance to herbicides or insects (e.g.Ostrinia andDiabrotica).As thecommer-cial maize crop of any region, maturityzone, market or country is not yet uni-form or homogeneous for any transgene,maize breeders have elected to developnear-isogenicversions(transgenicandnon-transgenic)ofeliteinbredsandcommercialhybridsinordertosatisfycombinationsoflicensing agreements, agronomic practices,regulatory requirements, market demandsand product development schemes. Thishas required companies to have twoparallelmaizebreedingprogrammes,trans-genic and non-transgenic. In this manner,marker-assisted backcrossing (MABC) oftransgenes,andtoalesserdegree,ofnativegenesandquantitativetraitloci(QTL)forothertraits,hasexpeditedthedevelopmentofcommercialhybrids.
Morerecently,marker-assistedrecurrentselection (MARS) schemes and infrastruc-ture have been developed for “forwardbreeding” of native genes and QTL forrelatively complex traits such as diseaseresistance,abioticstresstoleranceandgrainyield(RibautandBetrán,1999;Ragotet al.,2000; Ribaut, Jiang and Hoisington, 2000;Eathington, 2005; Crosbie et al., 2006).Simulation studies suggested that MAScouldbeeffectiveforsuchtraitsundercer-tain conditions (Edwards and Page, 1994;GimelfarbandLande,1994),buttheinitialempirical attempts at such selection werenot successful (Stromberg, Dudley andRufener, 1994; Openshaw and Frascaroli,1997; Holland, 2004; Moreau, Charcossetand Gallais, 2004) except in the specialcase of sweetcorn (Edwards and Johnson,1994;YousefandJuvik,2001).Thesuccessreportedforsweetcornisduetothefactthatthegeneticbaseof sweetcorn isextremelynarrowrelativetodentorflintmaize;thus

Chapter 8 – Marker-assisted selection in maize 121
predicted gains and extrapolations acrosspopulationsaremorereliable.Also,pheno-typicanalysesofmanytraitsinasweetcornbreedingprogrammeareextremelyexpen-sive because they involve processing largevolumesofgrain;therefore,MASwouldberelatively inexpensive and effective undersuch circumstances. However, subsequentdevelopments in technology, refinementsin analytical methods and improvementsin experimental designs have been assem-bledintoaprocessthathasshownpromisefor some reference populations of dentmaize (Ragot et al., 2000; Johnson, 2004;Crosbie et al., 2006) as improvement ingrainyieldfromMASoftenexceededthatfrom non-MAS approaches. Presumably,such results will lead to the developmentofnewandsuperiorinbredlinesandcom-mercialhybridsinacost-effectivemanner.WhiletheimpactofsuchMAShasnotyetbeen fully realized in the maize crop, themethods have been employed to variousdegreesbyprogrammesintheprivatesec-torthathavethenecessaryinfrastructure.
ThepotentialforMAStocontributetoimprovements in the maize crop shouldincreaseinparallelwithourunderstandingof the relationships among genomes, theenvironment and phenotypes. Candidatetransgenes will be developed on a regu-lar basis and their contributions to maizeimprovement will be realized in the mostefficient manner with MAS. Likewise, theidentificationofcandidatenativegenesandtheir gene products and functions, andof other DNA sequences (e.g. miRNA,matrixattachmentandregulatoryregions),willimprovethepowerofmethodssuchasassociation mapping and genome scans toassess theirgenotypicvalue in thecontextof defined reference populations of sig-nificance to maize breeding (Thornsberryet al., 2001; Rafalski, 2002; Niebur et al.,
2004;Varshney,GranerandSorrels,2005).Beyond its use in MARS schemes, thisinformation might make it reasonable toreconsider ideas such as methods for pre-dictinghybridperformancethatmayhavebeen limited by the amount and type ofinformationandbythedesignoftheexper-iment when they were initially evaluated(Bernardo,1994).
methodology and deSign of Breeding programmeS Supported By maSAs expected, private sector maize pro-grammes focus entirely on inbred-hybridbreedingschemesintendedtodevelopeliteinbredlinesthatenabletheprofitablepro-duction of commercial F1 hybrids. To alarge extent, MAS breeding programmesuse the same designs and methods knownto maize breeders for decades and genericdescriptions of these have been published(HallauerandMiranda,1981;SpragueandDudley,1988;Bernardo,2002).WhenMASisincludedinthebreedingprogramme,thesignificant differences are, of course, theavailability of genotypic data at differentstages of selection and some knowledgeoftherelationshipsbetweenthegenotypicandphenotypicdata sets for the referencepopulation(s)inthetargetenvironment(s).
In contrast to conventional breedingschemes, the methods and design ofinfrastructure needed to support MAShave been the areas of greatest change.In order to utilize MAS, companieshad to make significant investments toassemble or modify various aspects ofinfrastructure such as methods to detectDNApolymorphism,manageinformation,or analyse and track samples, softwareto relate genotype with phenotype, andoff-season or continuous nurseries. Thesecomponentshadtobeintegratedwitheach

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish122
other and with breeding activities, whichmeant that scientists needed to learn howand when MAS provided a comparativeadvantageoverothermethods.
maS: enabling methods, tools and infrastructurePerhapsthecomponentofinfrastructureingreatestneedofdevelopmentwasrelatedtotheacquisitionofgenotypicdata(i.e.DNAmarkers).Although theconceptof associ-ating markers with quantitative traits wasnotnew(Sax,1923),thediscoveryreportedbyStuberandMoll(1972)wasverysignifi-cant.StuberandMoll(1972)describedforthefirsttimeassociationsbetweenmolecu-lar markers and quantitative traits whileprevious associations had been based onmorphological markers (Sax, 1923). Theadvantages of molecular over morpholog-ical markers soon became obvious anddetailed descriptions of these advantageswere published by Tanksley et al. (1989)andStuber(1992).
Twooftheseadvantagesareofparticularimportance.First,molecularmarkergeno-types can usually be obtained from anyplanttissue,evenfromyoungseedlingsorkernels, while morphological markers fre-quently require the observation of whole,matureplants.Selectioncanthereforeoccurearlier in the plant’s cycle when usingmolecular markers than when using mor-phologicalmarkers.Theabilitytoconductearly selection, possibly before flowering,canhave a tremendous impacton the rateof genetic gain of a breeding programmeandthereforeconstitutesaverysignificantadvantage of molecular over morphologi-calmarkers.
Second, molecular markers are neutralmarkers. They are not affected by envi-ronmental or growing conditions. Theyarenotaffectedbythegeneticbackground
either,nordo theyaffectphenotypes.Theexpressionofmorphologicaltraits,bycon-trast, can be dependent on environmentalorgrowingconditions.Inaddition,epista-sic interactions are often observed amongmorphological marker loci or betweenmorphologicalmarker lociandthegeneticbackground. These epistatic interactionspreventdistinguishingallgenotypesassoci-atedwithmorphologicalmarkersandfurtherlimitthenumberofmorphologicalmarkersthatcanbestudiedsimultaneously.
Although isozyme markers had manyadvantages over morphological markers,the lack of a sufficient number of poly-morphic loci limited their use for MAS(Goodmanet al.,1980).Nevertheless, iso-zyme markers are still used for qualitycontrolduringseedproduction.
RFLPs (Botsteinet al., 1980)arebasedonDNApolymorphismsdetectedthroughrestriction nuclease digestions followedby DNA blot hybridizations. The abun-dance and high level of polymorphism ofRFLPs, especially in maize, allowed theconstruction of extensive maize geneticmaps (Helentjaris et al., 1986; Burr et al.,1988; Hoisington, 1989; Coe et al., 1995;Daviset al.,1999)aswellastheidentifica-tionandmappingofmanyQTL.
Being robust, reproducible and co-dominant, RFLPs are perfectly suited forgenetic studies as well as for MAS appli-cations. Their two main disadvantagesare the large quantities of DNA required,andthedifficultytominiaturizeandauto-mate. Nevertheless, RFLPs were quicklyadoptedandrepresentedthemarkersystemofchoiceformanyplantspeciesincludingmaize throughout the 1980s and duringmuchofthe1990s.
The development of the polymerasechain reaction (PCR) (Saiki et al., 1988)turned out to be a major breakthrough in

Chapter 8 – Marker-assisted selection in maize 123
molecular marker technology. PCR-basedmarkersrequirelittleDNA,allowingsam-pling of young seedlings and very earlyselection and thereby optimization ofbreedingschemes.PCR-basedmarkerpro-tocolsareveryamenabletoautomationandminiaturizationandimprovementstopro-tocols resulted in considerable reductionsin both cost and time required to pro-ducedatapoints.ThefirsttwoPCR-basedmarkersystemswererandomamplifiedpol-ymorphic DNA (RAPDs), and amplifiedfragment length polymorphisms (AFLPs).Detailed descriptions and critical assess-mentsofthesetwosystemscanbefoundinWelshandMcClelland (1990),Williamset al. (1990), Penner et al. (1993), Ragot andHoisington (1993), Skroch and Nienhuis(1995)and Joneset al. (1997) forRAPDs,andinVoset al.(1995),Joneset al.(1997)and Castiglioni et al. (1999) for AFLPs.They are also described in other chaptersofthisbook.
Simplesequencerepeats(SSRs)ormic-rosatellites rapidly became the marker ofchoice inmaize,almostentirelydisplacingRFLPs and previously developed PCR-based marker systems. Polymorphism ofSSRs is due to variable numbers of shorttandem repeats, often two or three basepairs in length and usually flanked byunique regions (Tautz, 1989). SSRs arevery reproducible (Jones et al., 1997) andco-dominant(Shattuck-Eidenset al.,1990;SeniorandHeun,1993;Senioret al.,1996)and are therefore very suitable for maizeMASapplications.
Many additional variations of PCR-basedmarkersystemshavebeendevelopedand a thorough review can be found inMohanet al.(1997).
AlltheDNA-basedmarkersystemsdes-cribedtodatearegel-basedsystems,amajorconstraintforautomation.Singlenucleotide
polymorphisms (SNPs) (Lindblad-Toh et al., 2000) can be revealed in many waysincluding allele-specific PCR, primerextensionapproaches,orDNAchips,allofwhicharenotgel-based.SNPscangenerallybe scored as co-dominant markers, exceptin the case of insertion-deletion polymor-phisms.AlthoughallelicdiversityatSNPsisusuallylimitedtotwoalleles,thislimitationcanbeoffsetbytheabundanceofSNPsandtheanalysisofhaplotypes,combinationsofgenotypes at several neighbouring SNPs.Haplotype analyses increase informative-ness(Chinget al.,2002),althoughatsomeexpensebecausetwotofourSNPshavetobegenotypedwhereoneSSRsufficed.SNPgenotypingcanbehighlyminiaturizedandautomated, thereby reducing the cost andallowingtheproductionofverylargenum-bers of data points. With genetic mapscontainingseveralthousandmappedSNPs,thesehavebecomethemarkerofchoiceforprivatemaizeMASprogrammes.
DNA marker technology has been adynamic and often expensive componentofthe infrastructureneededforMAS.Forexample, one corporation indicated hav-ingspenttensofmillionsofUnitedStatesdollars to develop an automated systemfor detecting RAPDs, a technology thatwasneversuitedforMASinalargemaizebreeding programme. Later, another cor-poration spent an even greater amount ofmoney to acquire technology for matrix-assistedlaserdesorption/ionizationtimeofflight(MALDI-TOF)analysisofamplifiedDNA fragments. These technologies wereeitherrapidlyreplacedorneverused.Suchdecisions would have bankrupted mostnational maize programmes or a coupleof centres belonging to the ConsultativeGroup on International AgriculturalResearch (CGIAR). Fortunately, this areaof infrastructure has matured somewhat

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish124
and become more stable so that start-upand operating costs, while still high forsomeprogrammes,aremorepredictable.
Statisticalmethodsandrelatedsoftwarehave also been areas of significant devel-opment, especially for the detection anddescriptionofputativeQTL.QTL,whicharenothingmorethanassociationsbetweenmarkers and traits, were first describedusing simple association tests betweentrait values and marker genotypes (StuberandMoll,1972).Thesetestsconsidereachmarker locus independently and neitherrequirenortakeadvantageoftheexistenceof genetic maps. Statistical methods havebeendeveloped that takeadvantageof theexistence of genetic maps (see review byManly and Olson, 1999). These statisticalmethods,simple intervalmapping(LanderandBotstein,1989)andcompositeintervalmapping (Jansen, 1993;Zeng,1993, 1994),test the existence of associations betweenhypothetical marker genotypes and traitvaluesatseveralpointsinintervalsbetweenpairsofadjacentmarkerlocionthegeneticmap, allowing the positioning of QTL onthese genetic maps. All of the previousmethodsarebasedonsingleQTLmodels.Otherstatisticalmethodshavebeendevel-opedthatsimultaneouslytestthepresenceofseveralQTLinthegenome(Kao,ZengandTeasdale,1999).
Many software packages are availablefor QTL mapping and based on one orseveralofthestatisticalmethodsdevelopedtodate.Notwopackagesareexactlyalikeand all have specific strengths and weak-nesseswithrespecttoparticularsituations,makingitsometimesbeneficialtousemorethan one package to perform QTL map-pinganalyses.ThesoftwarepackagesmostcommonlyusedforQTLmappinginmaizeinclude QTL Cartographer (Basten, WeirandZeng,1994),MapQTL(vanOoijenand
Maliepaard,1996),andPLABQTL(UtzandMelchinger, 1996). All of these only han-dle bi-allelic populations, while MCQTL(Jourjon et al., 2005) also performs QTLmappinginmulti-allelicsituations,includ-ing bi-parental populations made fromsegregating parents, or sets of bi-parental,bi-allelicpopulations.
More recently, methods based onBayesian analysis (Jansen, Jannink andBeavis, 2003; Gelman et al., 2004) andassociation (Varshney,Graner andSorrels,2005)or in silicomapping (ParisseauxandBernardo, 2004) have been proposed asmore powerful and refined approaches toassess the relationships between genotypeand phenotype that are needed for MAS.Methods of Bayesian analysis should beless affected by the uncertainties of QTLeffects and locations and produce betterestimates of those parameters in MAS.Association mapping approaches are par-ticularly useful to validate the relevanceof genes and alleles in specific germplasmsuch as that used by maize breeders. In silicomappingtakesadvantageofthepedi-gree relationships among individuals tostructure the population used to establishmarker-trait associations. This approach,whichishighlycomplexduetothepopu-lation structure resulting from pedigreebreeding, is particularly appropriate formaize where data across many years andenvironments are available for large setsof related individuals. Certainly, as theannotationofgenomesgraduallyimproves,suchmethodswillbecommoncomponentsof breeding programmes. Currently, theapplications of methods such as associa-tionmappingforMASarehinderedbythefactthataverylowpercentageofthegenesin crop plants have a function assigned tothem on the basis of direct experimenta-tion.However,thisimpoverishedsituation

Chapter 8 – Marker-assisted selection in maize 125
is being enriched through a variety ofprojectsonfunctionalgenomics.
Insharpcontrast to themanymethodsandsoftwarepackagesdevelopedforQTLidentification and mapping, little has beenpublishedforMAS.Thispaucityof infor-mation on MAS tools most likely reflectsboththelowlevelofactivityinthepublicsector and the fully proprietary nature ofdevelopmentsintheprivatesector.
Inparallelwithadvancements inDNAtechnologyandstatisticalmethods,privatesectorprogrammeshaveenhancedthecapa-bilities and capacities of their continuousnurseries. Such nurseries have been usedfordecadesbyprogrammesinboththepri-vateandpublicsectors.InordertoconductMASto itsgreatestadvantage,continuousnurserieshadtobemanaged,equippedandstaffedinnewwayssothattheplantscom-plete their lifecycleasquicklyaspossibleandthatthegenotypicdata(andsometimessome phenotypic data) needed for MASmaybecollectedateachsexualgeneration.Three to four sexual generations per yearmaybecompletedatsuchnurseries.
Theseactivitiesandthecontinuouscol-lection of both genotypic and phenotypicdata in the target environment and theirintegrated analyses create huge data setsthat must be analysed quickly and relatedto other large extant data sets. Data man-agement and bioinformatics for breedershavethereforebecomecriticalcomponentsof the infrastructure needed to use MAS.Prior to the advent of MAS, some largeprivate breeding programmes had estab-lishedagroupofdedicateddatamanagerstoassistwith researchandmarketing, andwith the arrival of genomics and MAStheneed for suchdedicated specialistshasincreasedgreatly.
Once the basic infrastructure had beenestablished to complement the activi-
ties of maize breeders, programmes wereready to implement several basic aspectsofMAS;manyofwhich arederived fromwellestablishedmethodsandprinciplesofmaizebreeding.
maS-based breedingSelectionoccursatvariousstages inmaizebreeding programmes. The first opportu-nity for selection is the choice of inbredlines to mate as parents of new popula-tions.PriortotheadventofDNAmarkerdata, the selection of such parents wouldbe based on a combination of phenotypicassessments,pedigreeinformation,breedingrecordsandchance(HallauerandMiranda,1981;SpragueandDudley,1988).Insomeprogrammes today, all such inbreds aregenotyped systematically at DNA markerloci. Depending on the resources andobjectives, the degree of genotyping mayrange from a low density of marker loci(e.g. SNPs in candidate genes) to higherdensity whole genome scans (Varshney,GranerandSorrels,2005).Thesegenotypicdata, alone or integrated with phenotypicinformation, may reveal novel aspects ofmaize gene pools, heterotic groups, hap-lotypeevolution,genecontentandparentsused in MAS for specific target environ-ments(FuandDooner,2002;Nieburet al.,2004;Crosbieet al.,2006).Whenproperlyintegrated with phenotypic informationand functional genomics, genotypic dataof inbred lines should allow breeders tochoose parents that, when mated, shouldprovidepopulationsorgenepoolsenrichedfor the more desirable combinations offavourable alleles. Such a starting point isahugeadvantageinplantbreedingbecauseit increases the probability of selectingprogeny that are superior to the parentsandthatapproximateapredictedoptimumgenotype.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish126
MABC is certainly the form of MASwiththemostimmediateandobviousben-efits for maize breeding. MABC is usedforthreemainpurposes:selectionoftrans-genes (orofnativeDNAsequencesof themaize genome, whether genes or QTL),elimination of unwanted regions of thedonor-parent genome linked to the trans-gene and selection of unlinked regions ofthe recurrent-parent genome. With theexceptionofDNAmarkersandtransgenes,thesehavebeenthesamegoalsofbackcrossbreedingsincetheinceptionofthatmethoddecadesago(Fehr,1987).Ofcourse,DNAmarkers enable breeders to identify prog-eny that contain the desired recombinantchromosomesanddonor-parentgenomeinamoredirectmanner.Also,MABCfacili-tates the process of combining more thanone transgene in a given inbred line (e.g.“gene or trait stacking or pyramiding”).This reduces the number of generationsneededtoreachcertainstagesofabreedingprogrammeandreducesthetimeneededtoproducecommercialhybridsforthemarket.GenericMABCschemessuitableformaizebreedingprogrammeshavebeendescribedindetailforsinglegenes(Hospital,Chevaletand Mulsant, 1992; Ragot et al., 1995;Frisch,BohnandMelchinger,1999a,1999b;Frisch and Melchinger, 2001a; Hospital,2001;Ribaut,JiangandHoisington, 2002),for QTL (Hospital and Charcosset, 1997;Bouchez et al., 2002) and for gene stacks(FrischandMelchinger,2001b).Versionsofsuchschemeshavebeenusedinmaizebreed-ingprogrammesintheprivatesector,oftenat theircontinuousnurseries (Ragotet al.,1995).Mostrecently,MABChasalsobeenadopted as a tool to develop sets of near-isogeniclines(NILs)forgenomicsresearch(PelemanandvanderVoort,2003).
Theoreticalandsimulationstudieshavebeen conducted to identify the most effi-
cient MABC protocols. Parameters mostcommonly studied include the number ofindividuals genotyped at each generation,thenumberofmarkersused,relativeselec-tionpressureforrecombinationaroundthetargetlocusorglobalrecoveryofrecurrentparent genome and the number of indi-vidualsselectedatanygeneration.Optimalvalues for each of the above depend onthe objective of the MABC approach interms of quality (required level of recur-rent parent genome recovery), speed(fastestpossible conversionor setnumberof generations) and resources (unlimitedor limited). While the fastest and high-est quality MABC approaches have themost expensive protocols, less intensiveapproaches can result in significant timesavings and quality improvements whencompared with conventional backcrossingapproachesandatafractionofthecostofthemostexpensiveMABCprotocols.
Frisch, Bohn and Melchinger (1999b)showed that to minimize linkage dragaround the target locus (loci), selection ofrecombination events close to the targetlocus (loci) should be conducted in theearly backcross generations. Frisch andMelchinger (2001a) and Ribaut, Jiang andHoisington (2002) further demonstratedthat minimizing linkage drag around thetarget locus requires very large numbersof individuals (possibly hundreds) to begenotyped.HospitalandCharcosset(1997)proposed a selection scheme based onselecting a single individual to be back-crossed.Bycontrast,FrischandMelchinger(2001a) proposed selecting several indi-viduals and determining the family sizeof their backcross progeny based on theindividuals’ genotypes. By using varyingrather thanconstantnumbersof individu-als or markers at the different backcrossgenerations,itwasshownthatthenumber

Chapter 8 – Marker-assisted selection in maize 127
of marker data points required could bereducedand thus theefficiencyofMABCimproved(Hospital,ChevaletandMulsant,1992;Frisch,BohnandMelchinger,1999b).Several studies also showed that using alimited number of markers on non-car-rierchromosomeswassufficienttorecovermorethat95percentoftherecurrentparentgenomeinthreeorfewerbackcrossgener-ations (Hospital, Chevalet and Mulsant,1992;Visscher,HaleyandThompson,1996;ServinandHospital,2002).
Oneofthemostimportantlessonsfromthevarioustheoreticalandsimulationstud-ies of MABC is that the effects of thedifferent MABC parameters are not inde-pendent of each other. With maize, largebackcross populations can be generatedfromasingleplantwhenthatplantisusedasthemaleandrecurrentparentplantsareused as females. Marker systems in maizeare also such that very large amounts ofmarker data can be generated on plantsbefore flowering. Potential MABC pro-tocols are almost endless in maize andidentifying the most efficient is only pos-sibleonacase-by-casebasis.Forexample,whileachievingalmostcompleterecoveryoftherecurrentparent’sgenome isnecessaryforregisteringbackcross-derived linesandhybrids inmanyEuropeancountries,par-tialrecoverymightbesufficienttoimprovethe agronomic performance of varieties indevelopingcountries.TheoptimalMABCprotocolsforthesetwostrikinglydifferentobjectiveswillbeverydifferent.Protocolsfor the first objective will involve back-groundselectionandtheuseofbackgroundmarkersveryclosetothetargetlocus(loci).Protocols for the second objective mightinvolve markers for the target locus (loci)only,whilerelyingonsuccessivebackcrossgenerationstorecoveranadequateamountofrecurrentparentgenome.
SuccessfulexamplesofMABCinmaizeinclude backcrossing of transgenes (Ragotet al.,1995),andQTLforinsectresistance(Willcox et al., 2002), flowering matu-rity (Ragot et al., 2000; Bouchez et al.,2002) and grain yield (Ho, McCouch andSmith,2002).
Methods of “forward breeding” withDNAmarkershavealsobeenproposedandimplementedbymaizebreedingprogrammes.As with the pedigree-based methods ofmaizebreedingfavouredbytheprivatesec-tor,manyofthe“new”methodsthatutilizegeneticdata fromDNAmarkers integratedwithphenotypicdata are essentiallya formof recurrent selection, a method that hasbeen in use for several decades (HallauerandMiranda, 1981).Thekey advantagesofthe new versions of recurrent selection are,ofcourse,theavailabilityofgeneticdataforall progeny at each generation of selection,theintegrationofgenotypicandphenotypicdata,andtherapidcyclingofgenerationsofselection and information-directed matingsatcontinuousnurseries.
At least two distinct forms of forwardbreeding with MAS have been proposed:singlelarge-scaleMAS(SLS-MAS)(Ribautand Betrán, 1999) and MARS (EdwardsandJohnson,1994;Lee,1995;Stam,1995).A key difference between the methods isthat SLS-MAS employs DNA markers atonlyonegenerationandattemptstoretaingeneticvariation inregionsof thegenomeunlinked to the DNA markers, whileMARS uses markers at each generation,exhausting genetic variation in mostregions of the genome. Versions of bothSLS-MAS and MARS have been used bybreedingprogrammesintheprivatesector(Johnson,2004;Eathington,2005;Crosbieet al.,2006).
SLS-MAS is of particular interest inpedigreebreedingasitconsistsofscreening

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish128
and selecting individuals at a few lociat early generations, usually F2 or F3,(Eathington, Dudley and Rufener, 1997),usinglargepopulations(RibautandBetrán,1999). Individualsdisplayinghomozygousfavourablegenotypesatthelociofinterestareselectedandself-pollinatedwhileothersare discarded. Self-pollinated progeny ofthe selected plants then proceed normallythrough subsequent steps of pedigreebreeding. Screening large populations isnecessarytoensurethatgeneticdiversityismaintainedatregionsnotundergenotypicselection, thereby allowing furtherphenotypic selection to be conducted.Loci at which marker selection operatescan be QTL as described by Ribaut andBetrán (1999). SLS-MAS is thus limitedbyissuessuchastheprecisionoftheQTLparameters(position,effect),andrelevanceof the QTL across environments or genepools.SLS-MAScanalsobeconductedforgenes, eliminating many of the limitationspertaining to QTL. Although a powerfulapproachadoptedinseveralspecies(barley,soybean, sunflower, wheat) to enrichbreedingpopulationsatafewloci(Crosbieet al.,2006),SLS-MASdoesnotappeartohave been widely implemented in maizebreedingprogrammes.
MARS targets all traits of importancein a breeding programme and for whichgenetic information can be obtained.Genetic information is usually obtainedfrom QTL analyses performed on experi-mentalpopulationsandcomesintheformof maps of QTL with their correspond-ing effects. If the QTL mapping analysisisconductedbasedonabi-parentalpopu-lation, the sign of the effect at each QTLindicateswhichof the twoparents carriedthefavourablealleleatthatQTL.Asbothparentsoftencontribute favourablealleles,the idealgenotype is amosaicofchromo-
somalsegmentsfromthetwoparents.Thisassumesthatthegoalistoobtainindividualswithasmanyaccumulatedfavourablealle-lesaspossible,adifferentgoalfromthatofmarker-assistedpopulationimprovementasstudied elsewhere (Lande and Thompson,1990; Gimelfarb and Lande, 1994; Gallais,Dillmann and Hospital, 1997; Hospital,ChevaletandMulsant,1997;Knapp,1998;Moreau et al., 1998; Xie and Xu, 1998).Population improvement schemes aregenerally based on the random matingof selected individuals while the schemeproposedhereisbasedondirectedrecom-bination between specific individuals. Asreported by Stam (1995), the ideal geno-type, defined as the mosaic of favourablechromosomal segments from two parents,willusuallyneveroccurinanyFnpopula-tionofrealisticsize.Itis,however,possibleto design a breeding scheme to produceor approach this ideal genotype based onindividualsoftheexperimentalpopulation.Thisbreedingschemecouldinvolveseveralsuccessivegenerationsofcrossingindividu-als(Stam,1995;PelemanandvanderVoort,2003)andwouldthereforeconstitutewhatis referred to as MARS or genotype con-struction. This idea can be extended tosituations where favourable alleles comefrom more than two parents (Stam, 1995;PelemanandvanderVoort,2003).
Van Berloo and Stam (1998, 2001) andCharmetet al. (1999)developedcomputersimulations around this idea and assessedtherelative merits of marker-assistedgenotype construction over phenotypicselec-tion. MARS was simulated in anexperimental population where QTL hadbeen mapped. Index (genetic) values werecomputed for each individual based onits genotypes at QTL-flanking markers(van Berloo and Stam, 1998, 2001). Allsimulation studies of MARS found that

Chapter 8 – Marker-assisted selection in maize 129
it was generally superior to phenotypicselection in accumulating favourable alle-lesinoneindividual(vanBerlooandStam,1998, 2001; Charmet et al., 1999). MARSappeared to take better advantage of thegeneticdiversitypresentinthepopulationsto which it was applied than phenotypicselection. Simulation research conductedby van Berloo and Stam (2001) showedthat MARS was between 3 and almost20percent more efficient than phenotypicselection. The advantage of MARS overphenotypicselectionwasgreaterwhenthepopulation under selection was larger ormoreheterozygous(BC1sorF2svs.RILs,recombinant inbred lines, or DHs, dou-bled haploids). Although van Berloo andStam (2001) limited their simulations topopulationsofupto200individuals, theirresults seem to indicate that the relativeadvantage of marker-assisted over pheno-typic selection would keep increasing aspopulationsize increased.Thesamesimu-lation studies showed that the advantageof marker-assisted over phenotypic selec-tionwaslargerwhendominantQTLwereinvolved in the selection index, or whentrait heritability was low in the case ofselectionforasingle trait (vanBerlooandStam,1998,2001).Theselatterobservationsare of little relevance to most commer-cial maize breeding programmes, the goalof which is generally the development ofinbred lines improved for several traitsthat will be later combined into superiorhybrid varieties. They should, however,increase the appeal of MARS approachesfor breeding programmes aimed at devel-opingopen-pollinatedvarieties.
Simulations have also addressed theimpact of the amount and quality ofQTL information on selection efficiency.Simulation and empirical studies (Beavis,1994, 1999) showed that QTL mapping
experiments based on segregatingpopulations of less than 500 individualsgenerallyrevealedonlyasubsetofallQTLaffecting the complex traits segregating inthese populations. Quantitative trait lociinformation used in subsequent MARSwas therefore necessarily incomplete.Van Berloo and Stam (2001) showed thatthe relative advantage of MARS overphenotypic selection decreased rapidlywhen the fraction of the total genotypicvarianceexplainedbytheQTLincludedinthe selection index decreased. By contrast(vanBerlooandStam,1998;Charmetet al.,1999),theefficiencyofMARSseemstoberatherrobusttothewell-documented(Lee,1995)uncertaintyofQTLgeneticlocations.Theuseofgenotypicinformationatmarkersflanking the QTL possibly explains thisobservation.
The cost efficiency of MARS was alsoinvestigated through simulation (Moreauet al., 2000; Xie and Xu, 1998). Whensimulating selection for a single trait,Moreauet al.(2000)foundthat,irrespectiveof the heritability of the trait, MARS wasalwaysmorecostefficientthanphenotypicselectionifthecostofgenotypingwaslessthan that of evaluating one individual inone plot. When simulating simultaneousselection for multiple traits, Xie and Xu(1998) found that MARS was more costefficient than phenotypic selection if thecost of genotyping was less than that ofphenotyping one individual for all traits.These studies were based on a singlegeneration of MARS. Also, they did nottakeintoconsiderationanyfactorsbesidesgenotypingandphenotypingcosts,althoughfactorsinfluencingthelengthofaselectioncycle or the number of cycles that can becompletedinayearcanobviouslyaffecttherelativeeconomicmeritsofmarker-assistedandphenotypicselection.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish130
In contrast to the abundance of QTLmapping reports, very few accounts ofMARS experiments are found in theliterature.Moreau,Charcosset andGallais(2004) compared phenotypic, marker-only,andcombinedrecurrentselectionforgrain yield and grain moisture at harvestover several cycles and years in maize.Combined selection was based both onphenotypic and marker information whilemarker-onlyselectionwasbasedonmarkerinformation only. Both the marker-onlyand the combined selection methodsconstitute MARS approaches. Severalcombinations of these three methods ofselection were applied to the segregatingpopulation that served to map the QTLused in marker-based selection indices.Over the six years of the experiment,two cycles of phenotypic selection, twocyclesofcombinedselection,onecycleofcombinedselectionfollowedbytwocyclesofmarker-onlyselection,andonecycleofmarker-only selection were conducted inparallel. A reassessment of the positionsandeffectsofQTLwasconductedafterthefirstcycleforthethreeschemescontainingmultiple cycles. All MARS methods weremore efficient than phenotypic selectionto increase the frequency of favourablealleles at QTL. Nevertheless, Moreau,Charcosset and Gallais (2004) reportedno significant difference between marker-assisted and phenotypic selection on themultitrait performance index, although allMARSmethodsresultedingeneticgainforboth grain yield and grain moisture whilephenotypicselectionresultedingeneticgainforgrainyieldbutanunfavourableevolutionof grain moisture. This disappointingresult was tentatively explained by thehighheritabilityofthetraits,favourabletophenotypicselection,while thepercentageof total phenotypic variance explained by
theQTLdetectedforbothtraitswasonlyabout 50percent. One very encouragingresultofthisexperiment,althoughMoreau,Charcosset and Gallais (2004) failed topresent it as such, was that the first cycleof marker-only selection was as efficientas phenotypic or combined selection indelivering genetic gain. Two conclusionscan be drawn from this observation.First, the QTL identified in the initialexperimental population were in generalnot artefacts. Second, selection pressureappliedat theseQTL,andaimedat fixingalleles identified as favourable, resulted ina change in performance of the selectedpopulation in the desired direction whencomparedwiththeinitialpopulation.
A similar experiment, although basedsolely on marker-only recurrent selection,wasreportedbyOpenshawandFrascaroli(1997). They conducted MARS in maizesimultaneously for four traits, for each ofwhichabouttenQTLhadbeenidentified.They showed that genetic gain had beenachieved in the first cycle of MARS, butthat latercyclesdidnotresult inanygain.PossibleexplanationsgivenfortheseresultsincludeduncertaintiesaboutQTLparame-ters(locationandeffect),interactioneffects(epistasis, genetic x environment interac-tion),andthefactthatselectionwasbasedonsinglemarkersratherthanchromosomalsegments(OpenshawandFrascaroli,1997).
Recent communications from severalprivateMARSresearchprogrammes(Ragotet al., 2000; Eathington, 2005; Crosbieet al., 2006) revealed large-scale success-ful applications in maize. Accounts weregiven of commercial maize hybrids forwhich at least one of the parental lineswas derived through MARS. Eathington(2005) and Crosbie et al. (2006) reportedthat the rates of genetic gain achievedthrough MARS were about twice those

Chapter 8 – Marker-assisted selection in maize 131
of phenotypic selection in some referencepopulations. Marker-only recurrent selec-tion schemes have been implemented foravarietyoftraitsincludinggrainyieldandgrain moisture (Eathington, 2005), or abi-oticstresstolerance(Ragotet al.,2000),andmultiple traits are being targeted simulta-neously. Selection indices were apparentlybasedon10toprobablymorethan50loci,these being either QTL identified in theexperimentalpopulationwhereMARSwasbeing initiated, QTL identified in otherpopulations, or genes. Marker genotypesaregeneratedforallmarkersflankingQTLincluded in the selection indices (Ragotet al., 2000). Plants are genotyped at eachcycle and specific combinations of plantsare selected for crossing, as proposed byvanBerlooandStam(1998).Several,prob-ably three to four, cycles or MARS areconductedperyearusingcontinuousnurs-eries. In maize, early versions of suchschemeshavebeentestedandimplemented(Johnson,2004;Crosbieet al.,2006).
Resultsreportedintheserecentcommu-nicationsaboutprivateMARSexperiments(Ragotet al.,2000;Eathington,2005)areinsharpcontrasttothoseinearlierpublications(Openshaw and Frascaroli, 1997; Moreau,CharcossetandGallais,2004).Several fac-torscanexplainthesediscrepancies:• Size of the populations submitted to selec-
tion at each cycle. Given reports thatincreasing population size should resultin higher genetic gain through MARS(van Berloo and Stam, 2001,) it is likelythat populations submitted to selectionin private programmes are rather large,larger than the 160 and 300individualsreported respectively by Openshaw andFrascaroli (1997) and Moreau, Charcos-setandGallais(2004).
•Use of flanking versus single markers.The use of flanking markers for QTL
under selection allows better predictionof the genotype at the QTL than whenusingsinglemarkers.Whensinglemark-ers are used, recombination events thatoccurbetweenthemarkerandtheQTLleadtolossoflinkagebetweenthemark-er and the QTL much faster than whenflanking markers are used, thereby rap-idlyreducingthepredictivepowerofthesinglemarker.
•Early selection, pre-flowering.Theabilitytoselectplantsbeforefloweringensuresoptimalmatingschemesasthegenotypesofplantsbeingselfedorintercrossedarefully known. However, this is not thecase when selection cannot take placebeforefloweringandinvolvesintercross-ingselfedprogeniesofselectedplants,thegenotypes of which might have driftedsignificantly from those of their geno-typedparents.
•Number of generations per year. To theauthors’knowledge,noneofthesimula-tionorexperimentalstudiesofMARShasassessedtheeffectsofcyclelengthonitsefficiency despite its direct relationshipto the rate of genetic gain. In maize,cyclelengthcanbereducedthree-tosix-fold when using marker-only recurrentselection compared with phenotypicrecurrentselection.Consequently,marker-onlyrecurrentselectionwillbesuperiorto phenotypic selection as soon as thegenetic gain achieved through one cycleof MARS is, respectively, more than athirdorasixthofthatachievedthroughonecycleofphenotypicselection.Privatemaizebreedingprogrammeshaveaccessto off-season nurseries. Furthermore,they have often established efficientcontinuousnurserieswherethreetofourgenerations of maize can be grown peryear. The use of such nurseries allowsthem to carry MARS continuously, i.e.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish132
withuptofourcyclesperyear,whereasphenotypicrecurrentselectionis limitedtoonecycleperyearatmost.Theimpacton the rate of genetic gain of such animplementationofMARSmightbeverypositive even if MARS did not presentanyadvantageoverphenotypicselectiononaper-cyclebasis.
•Cost of marker data points.Largeprivatecompanieshavemadeconsiderableeffortsto reduce both the cost of marker datapointsandthecostofexperimentalfieldplots. The ratio of cost of marker datapointtocostofexperimentalfieldplotismostlikelylowerinlargeprivatebreedingprogrammesthaninmostpublicresearchlaboratoriesorsmallprivateprogrammes,potentially leading todifferentviewsontheeconomicefficiencyofMARS.Marker-based and phenotypic selection
can be mobilized in many different ways,with respect to each other, in marker-assisted breeding schemes. Marker andphenotypicinformationcanbeusedeithersimultaneously or sequentially. Selectionof parents for breeding populations canbe made using marker information alone,phenotypicinformationalone,oracombi-nationofeach.Selectionofindividualsinabackcrossprogrammecanbemadeon thesole basis of either marker or phenotypicinformation, or using both. Advancementof individuals in a line development pro-grammecanalsobemadeateachgenerationon the basis of either marker informationonly, phenotypic information only, or acombination of each. In order to maxi-mizetherateofgeneticgainitislikelythatMAS breeding schemes such as MABCand MARS will involve generations ofmarker-only selection conducted at con-tinuousnurseries.Theadventofimprovedmethods of producing doubled haploidswill certainly further influence the way
marker-basedandphenotypicselectionaremobilizedwithrespecttoeachother.
In spite of the development ofmarker-only selection and regardless ofthe underlying technology and breedingscheme,high-qualityphenotypingremainsvital and without substitute at severalstages; but it may become more focused.Phenotypic evaluation remains the ulti-matescreenbeforeanycultivarisreleased.MAS-derived lines and hybrids that meetphenotypic requirements are selected forfurtherevaluationandselectiononthebasisoftheirphenotypicvalue,whilethosethatdo not are discarded. Phenotypic evalua-tionisalsocriticaltoestablishmarker-traitassociationsorperformthecandidategenevalidations required to conduct MAS.Here, high quality phenotyping is neces-sary.Phenotypingprotocolswill thereforelikely be different from those commonlyusedforphenotypicselection.Experimentsmaybeconductedthatinvolveside-by-sidecomparisonsofdifferenttreatmentssuchaswater stressornitrogen fertilization levelstodissectcomplextraitsintotheircompo-nentsandfacilitatetheelucidationoftheirgeneticbasis.
Enhancements of such approachesto maize breeding will be based on theincorporation of improved methods ofproducing doubled haploid inbred lines,informationfromfunctionalgenomicsandbylearninghowtoincorporatefavourablenativegeneticvariationsystematicallyafterMAS has reduced the genetic variationin the original reference populations tounacceptablelevels.
advantageS and limitationS of maS in maize Breeding programmeSadvantages of maSFor private breeding programmes, MAShasofferedseveralattractivefeatures,most

Chapter 8 – Marker-assisted selection in maize 133
of which are related to time and resourceallocations.
MABCclearlyprovidestheinformationneeded to reduce the number of gener-ations of backcrossing, to combine (i.e.“stack”)transgenes,“native”genesorQTLinto one inbred or hybrid quickly, and tomaximize the recovery of the recurrentparent’s genome in the backcross-derivedprogeny. In several private breeding pro-grammes,MABChasenabled thenumberof backcrossing generations needed torecover 99percent of the recurrent parentgenome to be reduced from six to three,reducingthetimeneededtodevelopacon-vertedvarietybyoneyear (Crosbieet al.,2006;Ragotet al.,1995).AsalinederivedbyMABCcanbemadetobeverysimilarto the original non-converted line, mostof its attributes, including agronomic per-formance, can be assumed to be equal orsimilar to those of the original line. Onlylimited phenotyping is therefore neces-sarytoverifytheseassumptions,comparedwith the extensive multiyear phenotypingrequired when backcrossing is conductedwithoutmarkers.OneortwoyearscanbesavedwithMABCduringpost-conversionphenotypingwhencomparedwithconven-tionalbackcrossing, resulting inanoveralltime advantage of MABC over conven-tionalbackcrossingofuptothreeyears.
In many situations, the greatest advan-tagesandprofitsarerealizedbythosewhoarefirsttothemarketwiththeirproducts.Also, for reasons related to the practicesofseedproductionorlegalaspectsofcropregistration procedures, it may be quiteimportant to be able to produce near-isogenic versions of inbreds and hybrids;MABC provides such ability at a higherprobability.
BycontrastwithMABC,SLS-MASandMARSdonotnecessarilydecreasethetime
needed to develop inbred lines. The useof MARS might actually increase it. TheadvantageofSLS-MASandMARSresidesintheirabilitytoincreasetherateofgeneticgain (Eathington,2005),whichpotentiallyresults in higher performing lines andhybridsthancanbedevelopedthroughphe-notypicselectiononly.BothSLS-MASandMARS increase the frequency of favour-able alleles in the population of selectedindividuals. The difference between thetwo approaches is that SLS-MAS oper-ates on few loci while MARS operateson many. When SLS-MAS or MARS areused, the effective size of the populationon which selection operates is increased,either directly for SLS-MAS or indirectlythroughseveralconsecutivegenerationsforMARS when compared with phenotypicpedigree selection. This increase in effec-tivepopulationsizepermitstheapplicationof a greater selection intensity and henceproduces a higher genetic gain. SLS-MASand MARS can also be seen as pre-selec-tionstepsifconductedpriortophenotypicselectionandthereforeimprovethechancesof evaluatinggenotypeswithahigher fre-quencyoffavourableallelesphenotypicallybecausethetrulyundesirableportionofthepopulationmayhavebeeneliminatedpriorto phenotyping. Phenotypic selection cantherefore be conducted with higher selec-tion intensity than would be possible ifnopre-selectionhad takenplace, resultingpotentiallyinadditionalgeneticgain.
Alternatively, the resources used forphenotyping can be allocated differentlybased on whether individuals have beenpre-selected or not with MAS. MASschemesforforwardbreedingshouldenablebreedingprogrammestoreallocateorfocusresources forphenotypicevaluation inthetarget environment.For example, ifDNAmarkers are linked to genes for resistance

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish134
to a disease or insect then it should bepossibleinitiallytoselectresistantprogenyin the absence of the disease or insectby using the DNA data at continuousnurseries. The selected progeny couldthen be evaluated using relatively moreexpensivebioassayswiththepest(s) inthetargetenvironment.ThisshiftinresourcesisinherenttoMARSschemesforcomplextraits(EdwardsandJohnson,1994;Johnson,2004; Crosbie et al., 2006). By enrichingpopulationsthroughrapidcyclesofMARSat continuous nurseries, breeders shouldderiveahigherfrequencyofprogenywithfavourable alleles and haplotypes that arethen evaluated in the target environment.WithoutMARS,resourcesforevaluationinthetargetenvironmentwouldbedilutedbytheinclusionoftoomanyprogenywithanundesirablegeneticconstitution.
Concerns about reduced genetic diver-sityamongcommercialmaizehybridsanddepletionofgeneticdiversityingenepoolsused in breeding may be partially alle-viated by successful implementations ofMAS.MABCmayrevive interest inusingessentially untapped maize exotic germ-plasm as a source of favourable alleles forimprovement of elite varieties. Very smalland targeted chromosomal segments ofexoticorigincanbe introgressed intoeliteinbred lines with limited risk of carry-ing along undesirable characteristics. Suchan approach could be beneficial in maizealthough no accounts of its implementa-tion have been reported despite the manyyears as reports of its successful use intomato(Tanksleyet al.,1996;Bernacchiet al.,1998a,b;Robertet al.,2001),rice(Xiaoet al.,1998),andsoybean(Concibioet al.,2003).MARS,inturn,mayalsocontributetoincreasinggeneticdiversityamongcom-mercialmaizehybridsbecause,byfocusingonselectingspecificrecombinationevents,
it will result in the development of gen-uinely new genomic rearrangements. AsQTL identified in any experiment repre-sentonlyafractionofthelociresponsiblefor the phenotypes of complex traits, onecan assume that breeding programmes indifferent private companies will conductMARS based on their different geneticmodels and select for different genomicrearrangements. As a result, hybrids ofsimilar and high performance might bedeveloped that are based on different setsof favourable alleles at different loci, rep-resenting distinct “genetic solutions” andcontributing to increased genetic diversityinfarmers’fields.
AnindirectbutimportantadvantageofMAS and its underlying information andtechnologyrelates to intellectualproperty.Some maize breeding programmes havecreatedaformofwealththroughtheircol-lectionandknowledgeofmaizegermplasm.Significant investmentshavebeenmade inmaize breeding as exemplified by the bil-lions of United States dollars that wereusedtopurchaseafewprivateprogrammesbetween 1995 and 2005. Protecting andmaximizing returns on such investmentshave always been important but are nowofgreaterconcern.InformationfromMASshould be advantageous for addressingissuesconcerningownershipandderivationof germplasm, relatedness among germ-plasmandfortheformationofsomeclaimsinpatentsandsimilardocuments.
Perhaps one of the greatest advantagesof MAS is that, for the first time, maizebreeders have the means of learning someofthegeneticdetailsaboutgermplasmandtheresponsetoselection.Somemaizepro-grammes intheprivatesectorhavestartedthis process (Niebur et al., 2004). As realfunctionsbecomeassociatedwiththemanycandidategenesandotherDNAsequences,

Chapter 8 – Marker-assisted selection in maize 135
the opportunities for learning about andunderstanding the response to selectionwill increase dramatically. It may then bepossible to ameliorate some of the limita-tionsofMASandtrulybreedbydesign.
limitations of maSWhile not truly an inherent limitation ofthe methods involved, one unavoidablelimitationofMASisthecostofassemblingandintegratingthenecessaryinfrastructureandpersonnel.Thesecanbesubstantialandbeyond the means of many programmes.For such programmes, implementation ofMAS could lead to a delusional or unbal-anced reallocation of resources from vitalactivities such as high-quality phenotypicevaluationandselectioninthetargetenvi-ronment.Currently,onlythelargestmaizebreedingprogrammesinagivenmarketorregionhavethescaleofsalesanddiversityof products that can justify and supportMAS and withstand some of the financialburdensofestablishingandreplacingcom-ponents of the system (e.g.changes in themethodsandplatformsfordetectingDNApolymorphisms).
Some inherent limitations to MAS arerelated to the estimates of QTL positionand genetic effects and the rates of falsepositives and negatives. Confidence inter-vals for QTL are typically 10–15 cM; agenetic region that should not be a majorbarrier for implementing MAS althoughit could become a limitation to achievinggeneticgainbypreventingtheselectionofdesired recombination events. The adventofassociationmappingandagrowingpoolof candidate genes should provide someresources needed to minimize problemsrelatedtotheestimationofQTLposition.The genetic effects of QTL are overesti-mated for many reasons, some of whichare linked to experimental designs for
phenotyping or population developmentwhileothersareinherenttotheprocessofQTL detection (Lee, 1995; Beavis, 1998;Melchinger,UtzandSchön,1998;Holland,2004). In addition, genetic effects relatedto epistasis are either poorly estimated orignoredbyprogrammesintheprivatesec-tor (Holland, 2001; Crosbie et al., 2006).Such assessments of genetic effects willinflate predictions of genetic gain. Therelative merit of MAS will depend on thenature of predictions, actual results andcostsofalternativemethods.
A possible limitation of MAS withmaize is thestructureandcontentofvari-ous gene pools. Examples of maize genepools would include European flint anddent germplasm, United States dents andvarious heterotic groups within each oftheseandother largerpools.SurveyswithDNAmarkershaveestablisheddifferencesamong such groups of germplasm (SmithandSmith,1992;Nieburet al.,2004).Therelativelyallele-richmaizegenepoolscou-pled with genetic heterogeneity for manytraitswillhinder theability toextrapolateinformation about genotype-phenotyperelationshipsacrossgenepools.Suchtrans-fer of information is expected to be moresuccessful in relatively homogeneous andless diverse maize gene pools (e.g. sweet-corn or popcorn) and with self-pollinatedplant species (Lee,1995).Therehavebeenundocumented reports of a few alleles atQTLthathave relativelyuniversalgeneticeffects across a relatively broad range ofmaize populations and target environ-ments, but details of such genetic factorshave not been publicly disclosed (Crosbieet al.,2006).Moreresourceswillneedtobedevotedtodiscoveringwheregeneticinfor-mationcannotbeeasilyextrapolatedacrossgene pools or even populations within agenepoolcomparedwithsituationswhere

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish136
it could. Although this should not impactthe economic efficiency of MABC or for-ward breeding, it could affect the overallcostefficiencyofMAS.
Finally,theefficacyforMASinrelativelycomplex populations such as syntheticsand open-pollinated varieties (OPVs) hasnotbeen investigated.Comparedwith thebi-allelic populations used in the privatesector, suchpopulationsare likely tohavemorethantwoallelesatagivenlocus.Also,unlike the simple bi-allelic populations,allele frequency should be an importantcomponentofpredictionswithsuchpopu-lations. Therefore, there should be moregeneticeffectsandinteractionstoconsiderwhen making predictions based on MASwithOPVsandsynthetics.
Inthefuture,successfulimplementationofMASinmaizemayleadtomorefrequentproblemsrelatedtolimitedgeneticvariation.The emphasis of aggressive private sectormaize breeding programmes on crossesbetweenelite,relatedinbredlinestocreatesegregating source populations has led toconcerns about the depletion of geneticdiversityinsuchgenepoolsandtheabilityto enhance such gene pools with highquality genetic variation (Niebur et al.,2004). Such concerns, which existed priorto the deployment of DNA markers andMAS in maize, are likely to increase asMAS becomes more prevalent. If MAS inforwardbreedingschemesisaseffectiveasreported, then alleles and haplotypes mayapproach fixation more rapidly (Crosbieet al., 2006). At that point, breedingprogrammes will need to repeat theprocessofcalibratinggenotype-phenotyperelationships in a slightly different arrayof reference populations to start the nextmetacycleofMAS(Johnson,2004).
Thereismuchanticipationforthefutureof MAS as genic sequences become the
marker loci, functional information isdis-covered for themanycandidategenesandgeneproductsareassessedfortheirpoten-tial as useful sources of information inbreeding programmes (Varshney, Granerand Sorrels, 2005; Lee, 2006). Certainly,thesehugesetsofrawdatawillcontributeto progress. Eventually, other sources ofgenetic variation unrelated to the primaryDNAsequencesuchasDNAmethylationwillbeevaluatedfortheirinfluenceongen-otype-phenotype relationships. Currently,epigeneticvariationismostlyignoredfromthat assessment although it is well knownthat much of the maize genome may bemethylated (Kaeppler, 2004) and may bemore dynamic than predicted by currentgenetic models and mechanisms (Fu andDooner,2002).Also,theinfluencesofnon-codingsequences suchas small interferingRNA(siRNA),matrixattachment regionsand long-distance regulatory sequenceshave yet to be considered for their effectsongeneticvariationandestimatesofgeneticvaluesusedinMAS(Lee,2006).
Most of the early limitations of MAS,duetotheavailabilityorcostofgenotypicdata, have been overcome. However,the availability or cost of high-qualityphenotypic information is becomingone of the major limitations of MAS.During the past 20 years, developmentof new technologies and automation andminiaturization of laboratory procedureshave contributed to reducing the cost ofmarker data points as well as the timeneeded to produce them. Large-scalemarker laboratories produce marker datapoints at less than a tenth of the cost of20yearsago.Bycontrast,neithercostnorthe time required to produce phenotypicdata has changed much, if at all, in thesame timeframe. As the establishment ofmarker-trait associations and ultimately

Chapter 8 – Marker-assisted selection in maize 137
the success of MAS depends on access tohigh-quality phenotypic data, means willhave to be found to decrease the cost ofphenotypic informationwhilemaintainingor increasing its quality. Alternatively,a greater proportion of budgets needsto be devoted to collecting phenotypicinformation.
aChievementS of maize Breeding programmeS with maSInsomeimportantways,maizebreedinghasgraduallychangedsincethemid1990swiththeadventofgenomics.Geneticprincipleswere always an important component ofmodern maize breeding and now geneticinformationofvarioustypesisseepingintobreeding schemes. MAS is the connectionbetween the growing pool of geneticinformation and actual plant breeding.Establishingandenhancingthisconnectionhavebeenimportantachievements.
For the simplest breeding scenario,programmes in the private sector havedemonstratedthatMABCisaneffectiveandroutine method to backcross one or moretransgenesintoestablishedeliteinbredlines,the direct parents of commercial hybrids.Hybrids with effective combinationsof transgenes have been very successfulin the market. Consequently, MAS hasaccelerated the delivery of some productstothemarket;animportantachievementincompetitiveeconomies.
Programmesintheprivatesectorhavealsodemonstratedasufficientdegreeofefficacyof MAS methods to secure protection ofintellectual property in patents. Methods,ideas and linkage relationships have beenincluded in claims of patents or patentapplications related toMAS (e.g.US5492547 1996; US6 455 758B1 2002; US2005/0144664A1 2005; WO2005/000006A22005; WO2005/014858A2 2005), or the
establishment of marker-trait associations(e.g. US5746 023P 1998; US6368806B12002; US6 399 855B1 2002). Given themagnitudeoftheinvestmentsmadeinmaizebreeding by the private sector, receivingsuch a legal position may be a valuableachievementfortheownerofthepatent.
The efficacy of MAS for forwardbreeding of complex traits has yet to befirmly established. Positive results fromcalibration studies have been reported,but although accounts of MAS-derivedcommercial varieties have been made(Eathington, 2005), the impact on actualbreeding and the development of newcommercialhybridshasnotbeendisclosedto a significant extent (Johnson, 2004;Nieburet al.,2004;Crosbieet al.,2006).Atthispointintime,itisthereforetooearlytomakeadefinitiveanddatabasedassessmentofthisaspectofMAS.
The history and cost of the geneticgain achieved through MAS will certainlyvary among target environments. In someregions of the world, such as the centralUnited States, maize breeding achievedsteady genetic gains in grain yield forseveral consecutive decades prior tothe advent of MAS (Duvick, Smith andCooper, 2004). Nevertheless, the cost perunit gain has increased as more resourcesare needed for phenotypic evaluation inmore environments (Smith et al., 1999).However, the advent of applied genomicsandthediscoveryofmanygenesandgenefunctions,coupledwithMAS,couldreducethe dependence on costly phenotypicinformationforbreeding.Inregionswherebiotic and abiotic stress factors are moreimportantthaninthecentralUnitesStates,MASmaybeveryeffective.Ultimately,thevalueandachievementsofMASwilldependon the ecological and socio-economiccontextofthetargetenvironment.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish138
CollaBoration Between the private and puBliC SeCtorS in maS and maize improvementThe increased investments in maizebreeding, expected returns on investmentandconcernsregardingintellectualpropertyby the private sector have made it moredifficult for corporations to collaboratewith external parties of any kind. Suchfactorshindertheexchangeofinformationandmaterialthatiscommonincollaborativeprojects. Nevertheless, around the world,theprivate andpublic sectors stillmanagetocollaboratethroughvariousmechanismsand at different levels in the pursuit ofmaize improvement. Such collaborationinvolves interactions among multinationalcorporations, philanthropic foundations,national and subnational governments,universities and individuals. Majorcategories of collaboration include socialprogrammesandinstitutions,researchanddevelopment,andeducation.
In many regions of the world, privatesectormaizebreedingwouldnothavegrownwithout some critical social programmesand institutions. For example, legislationrelated to intellectual property, transferof capital and material, and regulatoryapproval of biotechnical innovations inmaize improvement have been importantcomponents of legal systems that haveencouraged financial investment in maizebreeding. The stability of these systemsand the rule of law have contributed tothe long-term gains in selection. Also,long-term crop subsidy programmes insomeregionshaveprovidedanelementofsecurity for investments inmaize researchand development by the private sector(Troyer,2004;Crosbieet al.,2006).Inthosesame regions, MAS has been deployedinitially andon the largest scale formaizebreeding.
With respect to research and develop-ment, there is a long history of effectivecollaboration between the public and pri-vatesectorsinmaizebreeding.Whilesuchinteraction continues in the era of MAS,thenatureofthecollaborationhaschangedwith the growth and development of thebreeding programmes in the private sec-tor. Initially, collaboration was absolutelyvital for the private sector because breed-ing programmes in the public sector wereimportant,orthesole,sourcesoftheinbredlinesuseddirectlybytheprivatesectortoproduce commercial hybrids or to sourcepopulationsfromwhicheliteinbredswerederived. Also, the inbred lines from thepublic sectorwereusuallyprovidedonanunrestricted basis and without paymentsofroyaltiesorlicensingfees.Publicbreed-ing programmes continue to develop eliteinbred lines, occasionally in collaborationwiththeprivatesector(e.g.theGermplasmEnhancement of Maize programme in theUnitedStates;Pollak,2003).However,thedirectimpactofcontemporarypublicgerm-plasm varies greatly among regions andgradually, in many regions of the world,theprivatesectorhasbecometheprimarysourceofelitemaizeinbredlinesandcom-mercialhybrids.
In addition to germplasm, most or allof thecritical concepts,methodsandbasictechnologieshave theirorigins in thepub-licsector(Nieburet al.,2004;Troyer,2004;Crosbie et al., 2006). The private sector,withitsuniqueabilitytoconcentratecapi-talthroughvariousmechanisms(e.g.profitsfromproductsorlicencefees,venturecapi-tal and financial markets), is in the bestposition to allocate resources quickly toassess, modify and apply new develop-mentsinMASandancillaryareasofmaizeimprovement across large geographicaland political regions of a market zone. As

Chapter 8 – Marker-assisted selection in maize 139
described in previous sections, cost-effec-tiveMASrequiresseveralcomponentsofanintegrated infrastructure, some features ofwhichhavehadarelativelyhighrateofren-ovation and replacement (e.g. methods ofdetectingDNApolymorphism),andthere-forerequiredsubstantialfinancialresources.Competing corporations and the potentialforprofitprovidethenecessarymotivationforsuchinvestments(Troyer,2004;Crosbieet al., 2006). To the authors’ knowledge,such financial mechanisms either do notexistorarelimitedinthepublicsector.
Collaboration between the public andprivate sectors in MAS for maize may bestrongest in basic genetics and genomeannotation. In order for MAS to reachits full potential, it may be necessary toacquire a much better understandingof gene function and products. For anyplant species, only a small percentage ofgenes and other DNA sequences have afunction defined through direct experi-mentation(Lee,2006).Discoveriesinplantgenefunctionwilloccurinmanylaborato-ries around the world and, ultimately, thedevelopment groups in the private sectorwill have the necessary concentration ofresourcesandsenseofpurposetoassemblethe relatively raw basic information intotools andproducts fromMAS.Themaizenuclear genome, with tens of thousandsofgenesandmanyother importantDNAsequences, is mostly a “black box” withrespect to understanding the role of thesein mediating phenotypes in response toenvironmentalcues.Suchunderstanding,apotentialkeytoMASandmaizeimprove-ment,canonlyoccurthroughinformalandformal collaboration between the publicand private sectors investigating a broadarrayofplantspecies.
Examples of collaborative researchbetween the public and private sectors
relevanttoMASinmaizeincludeattemptsto select for hybrid yield (Stromberg,Dudley and Rufener, 1994), QTLmapping and selecting of hybrid yield(Eathington, Dudley and Rufener, 1997)andgrainquality (Laurieet al., 2004), thedevelopment of the IBM population ofrecombinant inbred lines, and mappinggenomic regions that include the vgt1locus in maize (Lee et al., 2002; Salvi et al., 2002). National collaborative researchprogrammessuchasGénoplanteinFranceand GABI in Germany, as well as severalprojectswithintheEuropeanCommission-sponsored framework programmes, areadditional examples of such collaboration.Certainly, other collaborative projectsbetween the public and private sectorshave been conducted in maize MAS buttheir proprietary nature prevents publicdisclosure.
Future collaborative research activitiesin maize MAS could assume many forms.In most regions of the world, the privatesectorhastheobvioussuperiorityintermsof infrastructure needed for genotyping,phenotyping and data analysis. Theseresourcesaremostlydevoted to thedirectpursuitofproductsandprofits.Thatpursuitmayalsobethegreatestdisadvantageoftheprivate sector because such a focus limitsthe attention devoted to many interestingyet seemingly ancillary observations ofgenotype-phenotype relations in MAS.Some components of that infrastructurecould possibly be made accessible to thepublic sector as “in-kind” contributionsto collaborative or service-related projectsin regions that are unlikely to emerge asimportantmarketsfortheprivatesectororforphenotypesandgermplasmthatarenotofdirectinteresttotheprivatesector.
Education and training are alsoimportant areas in which the public and

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish140
privatesectorsshouldcollaborate.WiththeadventofMAS,therehasbeenanobviousneed for maize breeders in the privatesector to become familiar with all aspectsof the process, and the public sector hasdeveloped several new short courses andtraining sessions in MAS-related concepts(Nieburet al.,2004;Crosbieet al.,2006).
Such knowledge is now considered astandard component of recent graduatetraining.However,whilenewstudentsmayhave an adequate grasp of the theoreticalaspects of MAS, their lack of exposure totheprivatesector’sadvancedinfrastructurerepresents a gap in their education. Thissituationissimilartothatofstudentswitha new degree in engineering who joinadvancedengineeringanddesigngroupsinotherindustries:theprivatesector’scapacitytoconcentrateandfocuscapitaloftenleadsto advanced infrastructure that does notexistinthepublicsector.Insuchsituations,new students have to navigate a rathersteep learning curve before they becomeproductive members of their new group.To reduce the slope of the learning curve,theprivatesectorcouldprovideinternshipstograduate studentsor toprofessorswhoteachplantbreedingcourses.Itisunlikelythatthepublicsectorwillhavetheresourcestoduplicateorexceedsomefeaturesoftheinfrastructure that has been developed formaizeMASintheprivatesector.Therefore,forsomeaspectsofeducation,itwillbetoeveryone’s benefit to find ways to worktogether.
private SeCtor perSpeCtiveS on maS for maize improvementThe development of molecular markersin the 1980s provided the first tools todissect the genetic basis of traits andselect individuals based on their predictedgeneticvalue.Backintheseearlydays,the
availability of genetic information was alimiting factor. Today’s landscape is verydifferent as advances in applied genomicsand laboratory technology have providedthetoolstogenerategeneticinformationforall traits of interest. Gene similarities andsynteny across genomes mean that muchof the informationgeneratedonanyplantspecieshasrelevancetootherplantspecies.The speed at which genetic informationbecomesavailableneverceasestoincrease.Rather than itsavailability, it is theabilityto handle and utilize genetic informationthat is becoming the limiting factor forMAS. New and improved informationtechnology and bioinformatics capabilitiesthereforeneedtobedevelopedthatconnectthegrowingwealthofgenetic informationwith maize breeding programmes whereknowledgeaboutthegeneticbasisoftraitsandallelicvariationattheselociistranslatedintovarieties.
QTL and gene mapping will remainkey for the generation and use of geneticinformation. As sequencing of cerealgenomes including maize progresses,physical mapping of cloned genes willbecomeapowerfulalternativetostatisticalapproaches. Characterization of allelicdiversity at loci of interest can proceedfromanalysesofbi-parentalpopulationsorassociationstudies.AneffectivealternativeistheuseofsetsofNILs,orintrogressionline (IL) libraries (Peleman and van derVoort, 2003). As NILs developed arounda specific locus differ only by the alleleat this locus, and because most traits ofagronomicinterestinmaizearequantitative,phenotypicdifferencesamongsuchNILsareexpectedtoberathersmall.Highprecisionphenotypingwillnotonlyberequiredbutwill be critical for the evaluation of suchmaterial(PelemanandvanderVoort,2003).Privatecorporationshaverealizedtheneed

Chapter 8 – Marker-assisted selection in maize 141
for such high precision phenotyping ascanbe seen fromtheiractive recruitingoftrait-specific phenotyping scientists oftenlocatedintargetedareaswherethetraitofinterest can be more easily measured (e.g.positions dedicated to drought toleranceandlocatedinaridregionsoftheworld).
In order to further the implementationof MAS in breeding, increased numbersof marker data points will be required.Private corporations have established orare developing the capacity to producehundreds of millions of data points peryear in service laboratories, distinct fromresearch units. Besides, smaller “biotech”companiesaredevelopingtechnologiesthatcould reduce thecostofeachmarkerdatapoint to a mere few United States cents.Moving to marker systems that are notbasedongelsispermittingtheautomationofmostlaboratorysteps.Datapointsarebeingproducedaroundtheclockwithlaboratorytechnicians working in shifts. Here again,private companies are actively recruitinghighly qualified technology specialists aswell as laboratory managers whose role ismoretooptimizetherunningofproductionplants than dwell on the science. Beyondlaboratories, plant handling is becoming abottleneck to high-throughput protocols.High-throughput facilities have to beestablished and equipped at continuousnurserysitespotentiallytohandlemillionsofplantsperyear.
There is little doubt that the largestprivate maize breeding programmes areinvestingveryheavilyintheimplementationof MAS. Unless regulatory issues changedramatically, MABC will remain thepreferred means of delivering transgenestothemarket.FasterMABCprotocolswillalways represent a potential commercialadvantage inanareawherecompetition isfierceandaone-yearadvantagemaymean
muchonthemarket.MostrecentinvestmentshavebeendirectedatimplementingMARSin breeding. The size of the investmentin this approach seems to suggest thatprivatecorporationshavemoreinsightintoits benefits compared with conventionalbreeding than has been reported publicly.Genotype-driven breeding should alsoallow faster development of specializedvarietiesasthemaizemarketbecomesmoreandmorefragmentedbasedonend-useofthe harvest: animal feed (silage or grain),ethanol, dry or wet milling. Favourablealleles for traits of interest are likely tobe spread across more than two linestherefore requiring the assembly of allelesfrom many different sources in a singleinbred line. Proposals have been made toachieve such goals (Peleman and van derVoort 2003), although software tools todetermine the optimal breeding schemesare not yet available to generate these“ideal”genotypes.
Maize breeding is likely to changemore in the coming 10 or 20 years thanit has over the past 50. Developing newhybridsefficientlynowrequiresintegratingdata from many sources, sometimesbeyond maize, generating high-qualitygenotypic and phenotypic data neededfor the construction of “ideal” genotypes,and finally selecting phenotypically thebest individuals from populations ofmarker-assisted-derived materials. Manystakeholders beyond maize breeders nowtake an active part in the development ofnew varieties and therefore breeding willincreasingly become the responsibility ofgroupsofindividualswithcomplementaryskills than stand-alone breeders. Trainingof all to understand and challenge thecontribution of others will be critical tooperatingmultidisciplinarybreedingteamsefficiently.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish142
maS for maize improvement in developing CountrieSA rapid analysis of the implementationof MAS in private maize breeding pro-grammespointstothreeelementsasbeingof particular importance: availability ofhigh-quality phenotypic data, access tolow-costmolecularmarkerdatapointsandaccesstoreliablecontinuousnurseries.Theimportance of high-quality phenotypicanalyses has been clearly recognized bygroupsintheprivatesector(Nieburet al.,2004;Crosbieet al.,2006).Implementationof MAS in maize breeding requires largeamountsofmarkerdatapoints tobegen-erated. Private groups have spent mucheffort developing technologies and plat-formstoachievecost-efficientgenotyping.Simultaneously,highlyefficientcontinuousnurserieshavebeen established in tropicalenvironmentsorlocalgreenhouses.
By contrast, maize breeding for devel-oping countries is rather fragmented.National agricultural research institutionsand international centres of the CGIARsuchastheInternationalMaizeandWheatImprovement Center (CIMMYT) focusmuchoftheireffortsonpoorfarmersandunderserved regions. Private maize breed-ing programmes are also established ina number of developing countries. Dueto the large up-front costs of assemblinginfrastructure and personnel for genotyp-ing, it is unlikely that individual nationalmarker laboratories could produce datapointsinacost-efficientmanner.However,regional facilitiesservingtheneedsofsev-eral national programmes and supportedby local laboratories that could processsamples (processing samples could be aseasy as taking and air-drying them) andprovide information in a timely manner,would probably be very sustainable alter-natives. Such a regional molecular service
laboratory has been established recentlyin Nairobi, Kenya, in a joint effort bytwo CGIAR centres, CIMMYT and theInternational Livestock Research Institute(ILRI) and Kenya’s Agricultural ResearchInstitute (KARI), under the CanadianInternational Development Agency(CIDA)-funded Biosciences eastern andcentralAfrica(BecA)platform,toprovidetechnical access and training for Africanmaize breeders (Delmer, 2005). Such afacility could be an excellent componentof a comprehensive maize breeding effortif it is possible to establish and maintainhigh-qualitypersonnelandfacilitiesforallof the other aspects of maize breeding inkey target environments. However, with-outhigh-qualitycapabilitiesinphenotypicevaluationandselection,molecularlabora-torieswillbeworthless.Researchprojectsinvolving large-scale (transnational) phe-notypicevaluationsofkeygeneticmaterialandfocusedonspecifictraits(tolerancetobiotic or abiotic stresses) should providegenetic information that is both locallyrelevantandbroadlyapplicable(geograph-ically and in terms of germplasm). Suchprojectswouldalsospreadthecostofphe-notypingacrossallparticipantsbutwouldonly be successful with effective transna-tionalcoordination.
Private companies running MAS inmaize could contribute to its implemen-tation in developing countries in severalways.First,theycouldmakesomeoftheirgeneticinformationavailable,therebyadd-ing to that already available in the publicdomain. Much information is being gen-erated in the private sector on traits ofimportancetodevelopingcountriessuchasdiseaseresistance(e.g.greyleafspot,north-erncornleafblight,Fusariumstalkandearrots), drought tolerance and nitrogen useefficiency.Aftervalidationof its relevance

Chapter 8 – Marker-assisted selection in maize 143
tothegermplasmandenvironmentsoftar-getareas,thisgeneticinformationcouldbeused to select efficiently for specific traitsthrough MAS. Second, private companiescouldprovideaccesstosomeoftheirgeno-typing or nursery platforms. Genotypingsamples for MAS projects in developingcountries would not substantially disruptprivate companies’ own research if con-ducted in periods of lower activity, andwould provide these MAS projects withmarkerdatapointsforaslowacostaspos-sible. Third and probably most critically,private companies could train scientistsfrom developing countries on the princi-ples, mechanics and logistics of applyingandimplementingMASinmaize.Scientistsin private maize breeding groups havealready identifiedmanyof thepitfalls andovercomemanyofthehurdleslinkedwiththe implementation of MAS. Transfer ofthisknowledgetoscientistsfromdevelop-ingcountrieswouldhelp them immenselyto design marker-based breeding schemesadaptedtotheirsetsofconstraints.
Beyondtheircontributiontotheimple-mentationofMAS inmaize indevelopingcountries,privatecompaniescould,inverysimilar ways, contribute to MAS pro-grammesinotherspeciesofimportancetodevelopingcountriesbutremotefromtheircoreinterests.Syntenyandgeneconserva-tionacrossspeciesshouldallowsomeofthemaizegeneticinformationtobetransferabletootherspecies.Technologyplatformsandbreeding approaches developed for MASinmaizeshouldbegoodmodelsforothercrops and some might be directly usable.Mechanisms or organizations need to beput in place for these transfers of knowl-edgeandtechnologiestooccurfromprivatemaizeMASprogrammestoothercropsindeveloping countries. Private programmeswill likely not drive these transfers but
might be very willing to contribute or bedirectly involved in specific projects pro-videdadequateframeworksexist.
Public–private partnerships will needto be established to manage intellectualproperty issues related to the transfers ofinformation,materialortechnologiesfromprivate companies to developing coun-tries (Naylor et al., 2004). The AfricanAgricultural Technology Foundation(AATF) is one initiative that has beenestablishedtodealwithsuchissues.Severalprivate corporations with major invest-ments in MAS in maize have agreed toprovide access to germplasm and knowl-edge for African countries (Naylor et al.,2004;Delmer,2005).
As with the private sector in Europeand North America, it will be necessarytoprovideregularandeasyaccess toedu-cation and training in maize MAS as thephenotypes and population structures arelikelytodifferfromthoseencounteredbyprogrammes in the private sector in rela-tivelyhigh-inputproductionenvironments.Also, and incommonwith thechanges inthe private sector, some reorganization orrestructuringofpublicsectorprogrammesmaybewarrantedwiththeadventofmorespecializedrolesforsomepersonnel.
Understandingthegeneticbasisoftraitsandcloningandsequencingtheunderlyinggenes will not have an impact on poorfarmers unless translated into varietiesthrough breeding. Implementing MASrequires significant investments in bothpeople and infrastructures. Some of themost promising marker-based breedingschemes (e.g. MARS), take about as longas conventional breeding schemes todevelop improved varieties and thereforerequire long-term funding commitments.Funding of practical crop improvementhas declined for several years, particularly

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish144
in the international public sector (Knight,2003), and as a result investments havefavoured research at the expense ofpracticalapplications(Nayloret al.,2004).Whether current funding mechanisms
based on short-term (two to five years)grantsareadequate toallowmaizeoranyother breeding programmes in developingcountriestobenefitfromthemuchneededadvantagesofMASisquestionable.
referenCeSBasten, C.J., Weir, B.S. & Zeng, Z-B.1994.Zmap--aQTLcartographer.InC.Smith,J.S.Gavora,B.
Benkel,J.Chesnais,W.Fairfull,J.P.Gibson,B.W.Kennedy&E.B.Burnside,eds.Proc. 5th Wrld. Congr. Genet. Appl. Livest. Prodn., Vol.22, pp.65–66.Guelph,Ontario,Canada,UniversityofGuelph.
Beavis, W.D. 1994. The power and deceit of QTL experiments: lessons from comparative QTLstudies. Proc. 49th Ann. Corn and Sorghum Ind. Res. Conf.,pp.250–266.Washington,DC,USA,AmericanSeedTradeAssociation.
Beavis, W.D.1998.QTLanalyses:power,precisionandaccuracy.InH.Paterson,ed.Molecular dis-section of complex traits,pp.143–162.BocaRaton,FL,USA,CRCPress.
Beavis, W.D.1999.Integratingthecentraldogmawithplantbreeding.Abstr. Plant & Anim. Genome7:28.
Bernacchi, D., Beck-Bunn, T., Emmatty, D., Eshed, Y., Inai, S., Lopez, J., Petiard, V., Sayama, H., Uhlig, J., Zamir, D. & Tanksley, S.1998a.AdvancedbackcrossQTLanalysisoftomato.II.Evaluationofnear-isogeniclinescarryingsingle-donorintrogressionsfordesirablewildQTLalle-lesderivedfromLycopersicon hirsutumandL. pimpinellifolium.Theor. Appl. Genet.97:170–180.
Bernacchi, D., Beck-Bunn, T., Eshed, Y., Lopez, J., Petiard, V., Uhlig, J., Zamir, D. & Tanksley, S.1998b.AdvancedbackcrossQTLanalysisintomato.I.IdentificationofQTLsfortraitsofagro-nomicimportancefromLycopersicon hirsutum.Theor. Appl. Genet.97:381–397.
Bernardo, R.1994.Predictionofmaizesingle-crossperformanceusingRFLPsandinformationfromrelatedhybrids.Crop Sci.34:20–25.
Bernardo, R.2001.Whatifweknewallthegenesforaquantitativetraitinhybridcrops?Crop Sci.41:1–4.
Bernardo, R.2002.Breeding for quantitative traits in plants.Woodbury,MN,USA,StemmaPress.Botstein, D., White, R.L., Skolnick, M. & Davis, R.W.1980.Constructionofgeneticlinkagemapin
humanusingrestrictionfragmentlengthpolymorphisms.Am. J. Human Genet.32:314–331.Bouchez, A., Hospital, F., Causse, M., Gallais, A. & Charcosset, A. 2002.Marker-assistedintrogres-
sionoffavorableallelesatquantitativetraitlocibetweenmaizeelitelines.Genetics162:1945–1959.Burr, B., Burr, F.A., Thompson, K.H., Albertsen, M.C. & Stuber, C.W.1988.Genemappingwith
recombinantinbredsinmaize.Genetics 118:519–526.Castiglioni, P., Ajmone-Marsan, P., van Wijk, R. & Motto, M. 1999.AFLPmarkersinamolecularlinkage
mapofmaize:codominantscoringandlinkagegroupdistribution.Theor. Appl. Genet.99:425–431.Charmet, G., Robert, N., Perretant, M.R., Gay, G., Sourdille, P., Groos, C., Bernard, S. & Bernard,
M. 1999. Marker-assisted recurrent selection for cumulating additive and interactive QTL’s inrecombinantinbredlines.Theor. Appl. Genet.99:1143–1148.
Ching, A., Caldwell, K.S., Jung, M., Dolan, M., Smith, O.S., Tingey, S., Morgante, M. & Rafalski, A.2002.SNPfrequency,haplotypestructureandlinkagedisequilibriuminelitemaizeinbredlines.BMC Genetics3:19.

Chapter 8 – Marker-assisted selection in maize 145
Coe, E., Hancock, D., Kowalewski, S. & Polacco, M. 1995. Gene list and working maps. Maize Genet. Coop. Newsl.69:191–267.
Concibio, V.C., La Vallee, B., McLaird, P., Pineda, N., Meyer, J., Hummer, L., Yang, J., Wu, K. & Delannay, X.2003.IntrogressionofaquantitativetraitlocusforyieldfromGlycine sojaintocom-mercialsoybeancultivars.Theor. Appl. Genet.106:575–582.
Crosbie, T.M., Eathington, S.R., Johnson, G.R., Edwards, M., Reiter, R., Stark, S., Mohanty, R.G., Oyervides, M., Buehler, R.E., Walker, A.K., Dobert, R., Delannay, X., Pershing, J.C., Hall, M.A. & Lamkey, K.R.2006.Plantbreeding:past,present,andfuture.InK.R.Lamkey&M.Lee,eds.Plant Breeding: The Arnel R. Hallauer Internat. Symp.Ames,IA,USA.,BlackwellPublishing.
Davis, G.L., McMullen, M.D., Baysdorfer, C., Musket, T., Grant, D., Staebell, M., Xu, G., Polacco, M., Koster, L., Melia-Hancock, S., Houchins, K., Chao, S. & Coe Jr., E.H.1999.Amaizemapstandardwithsequencedcoremarkers,grassgenomereferencepointsand932expressedsequencetaggedsites(ESTs)ina1736-locusmap.Genetics152:1137–1172.
Delmer, D.P.2005.Agricultureinthedevelopingworld:connectinginnovationsinplantresearchtodownstreamapplications.Proc. Natl. Acad. Sci. USA102: 15739–15746.
Duvick, D.N., Smith, J.S.C. & Cooper, M.2004.Long-termselectioninacommercialhybridmaizebreedingprogram.InJ.Janick,ed.Plant Breeding Rev.24:109–152.
Eathington, S.R.2005.Practicalapplicationsofmoleculartechnologyinthedevelopmentofcom-mercial maize hybrids. Proc. 60th Ann. Corn and Sorghum Seed Res. Conf. Washington, D.C.,USA,AmericanSeedTradeAssociation.
Eathington, S.R., Dudley, J.W. & Rufener, G.K.1997.Usefulnessofmarker-QTLassociationsinearlygenerationselection.Crop Sci.37:1686–1693.
Edwards, M. & Johnson, L. 1994. RFLPs for rapid recurrent selection. Proc. Symp. Analysis of Molecular Marker Data, pp. 33–40. Corvallis, OR, USA, American Society of HorticulturalScienceandCropScienceSocietyofAmerica.
Edwards, M.D. & Page, N.J.1994.Evaluationofmarker-assistedselectionthroughcomputersimula-tion.Theor. Appl. Genet.88:376–382.
Fehr, W.R.1987.Principles of cultivar development.NewYork,USA,Macmillan.Frisch, M. & Melchinger, A.E.2001a.Marker-assistedbackcrossingforintrogressionofarecessive
gene.Crop Sci.41:1485–1494.Frisch, M. & Melchinger, A.E.2001b.Marker-assistedbackcrossingforsimultaneousintrogression
oftwogenes.Crop Sci.41:1716–1725.Frisch, M., Bohn, M. & Melchinger, A.E.1999a.Minimalsamplesizeandoptimalpositioningofflank-
ingmarkersinmarker-assistedbackcrossingfortransferofatargetgene.Crop Sci.39:967–975.Frisch, M., Bohn, M. & Melchinger, A.E. 1999b.Comparison of selection strategies for marker-
assistedbackcrossingofagene.Crop Sci.39:1295–1301.Fu, H. & Dooner, H.K.2002.Intraspecificviolationofgeneticco-linearityand its implications in
maize.Proc. Natl. Acad. Sci. USA99:9573–9578.Gallais, A., Dillmann, C. & Hospital, F.1997.Ananalyticalapproachofmarkerassistedselection
withselectiononmarkersonly.InR.Krajewski&Z.Kaczmarek,eds.Advances in biometrical genetics. Proc.10th. Mtg. EUCARPIA Section Biometrics in Plant Breeding,pp.111–116.Poznan,InstituteofPlantGenetics,PolishAcademyofSciences.
Gelman, A., Carlin, J.B., Stern, H.S. & Rubin, D.B.2004.Bayesian data analysis,2nd edition.BocaRaton,FL,USA,ChapmanandHall/CRC.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish146
Gimelfarb, A. & Lande, R. 1994. Simulation of marker-assisted selection in hybrid populations.Genet. Res.63:39–47.
Goodman, M.M. & Carson, M.L.2000.Realityvs.myth:cornbreeding,exotics,andgeneticengi-neering. Proc. 55th Ann. Corn and Sorghum Res. Conf., pp. 149–172. Washington, D.C., USA,AmericanSeedTradeAssociation.
Goodman, M.M., Stuber, C.W., Newton, K. & Weissinger, H.H.1980.Linkagerelationshipsof19enzymelociinmaize.Genetics 96:697–710.
Hallauer, A.R. & Miranda, J.B.1981.Quantitative genetics in maize breeding.Ames,IA,USA,IowaStateUniversityPress.
Helentjaris, T., Slocum, M., Wright, S., Schasfer, A. & Nienhuis, J.1986.Constructionofgeneticlinkagemapsinmaizeandtomatousingrestrictionfragmentlengthpolymorphisms.Theor. Appl. Genet.72:761–769.
Ho, J.C., McCouch, S.R. & Smith, M.E.2002.ImprovementofhybridyieldbyadvancedbackcrossQTLanalysisinelitemaize.Theor. Appl. Genet.105:440–448.
Hoisington, D.1989.Workinglinkagemaps.Maize Genet. Coop. Newsl.63:141–151.Holland, J.B.2001.Epistasisandplantbreeding.InJ.Janick,ed.Plant Breeding Rev.21:29–32.Holland, J.B. 2004. Implementation of molecular markers for quantitative traits in breeding pro-
grams– challenges and opportunities. In T. Fischer, N. Turner, J. Angus, L. McIntyre, M.Robertson,A.Borrell&D.Lloyd,eds.New directions for a diverse planet. Proc. 4th Internat. Crop Sci. Congr.,Brisbane,Australia(availableatwww.cropscience.org.au/icsc2004).
Hospital, F. 2001.Sizeofdonorchromosomesegments around introgressed loci and reductionoflinkagedraginmarker-assistedbackcrossprograms.Genetics 158:1363–1379.
Hospital, F. & Charcosset, A.1997.Marker-assistedintrogressionofquantitativetraitloci.Genetics 147:1469–1485.
Hospital, F., Chevalet, C. & Mulsant, P.1992.Usingmarkersingeneintrogressionbreedingpro-grams.Genetics132:1199–1210.
Jansen, R.C.1993.Intervalmappingofmultiplequantitativetraitlocibyusingmolecularmarkers.Theor. Appl. Genet.85:252–260.
Jansen, R.C., Jannink, J-L. & Beavis, W.D.2003.Mappingquantitativetraitlociinplantbreedingpopulations:useofparentalhaplotypesharing.Crop Sci.43:829–834.
Johnson, G.R.2004.Marker-assistedselection.InJ.Janick,ed. Plant Breeding Rev.24:293–310.Jones, C.J., Edwards, K.J., Castaglione, S., Winfield, M.O., Sala, F., van de Wiel, C., Bredemeijer,
G., Vosman, B., Matthes, M., Daly, A., Brettschneider, R., Bettini, P., Buiatti, M., Maestri, E., Malcevschi, A., Marmiroli, N., Aert, R., Volkaert, G., Rueda, J., Linacero, R., Vazquez, A. & Karp, A.1997.ReproducibilitytestingofRAPD,AFLPandSSRmarkersinplantsbyanetworkofEuropeanlaboratories.Mol. Breed.3:381–390.
Jourjon, M-F., Jasson, S., Marcel, J., Ngom, B. & Mangin, B. 2005.MCQTL:multi-allelicQTLmappinginmulti-crossdesign. Bioinformatics21:128–130.
Kao, C-H., Zeng, Z-B. & Teasdale, R. 1999.Multiple intervalmapping forquantitative trait loci.Genetics 152:1203–1216.
Kaeppler, S.2004.Biotechnology:newhorizons.InC.W.Smith,J.Betrán,&E.C.A.Runge,eds.Corn: origin, history, technology, and production,pp.399–425.Hoboken,NJ,USA,JohnWiley&Sons.
Knapp, S.J.1998.Marker-assistedselectionasastrategyfor increasing theprobabilityofselectingsuperiorgenotypes.Crop Sci.38:1164–1174.

Chapter 8 – Marker-assisted selection in maize 147
Knight, J.2003.Cropimprovement:adyingbreed.Nature421:568–570.Lande, R. & Thompson, R. 1990. Efficiency of marker-assisted selection in the improvement of
quantitativetraits.Genetics124:743–756.Lander, E.S. & Botstein, D.1989.MappingMendelianfactorsunderlyingquantitative traitsusing
RFLPlinkagemaps.Genetics121:185–199.Laurie, C.C., Chasalow, S.D., LeDeaux, J.R., McCarroll, R., Bush, D., Hauge, B., Lai, C., Clark,
D., Rocheford, T.R. &. Dudley, J.W.2004.Thegeneticarchitectureofresponsetolong-termarti-ficialselectionforoilconcentrationinthemaizekernel.Genetics168:2141–2155.
Lee, M.1995.DNAmarkersinplantbreedingprograms.Adv. Agron.55:265–344.Lee, M.2006.Thephenotypicandgenotypicerasofplantbreeding.InK.R.Lamkey&M.Lee,eds.
Plant Breeding: The Arnel R. Hallauer Internat. Symp.Ames,IA,USA,BlackwellPublishing.Lee, M., Sharopova, N., Beavis, W.D., Grant, D., Katt, M., Blair, D. & Hallauer, A.R. 2002.
Expanding thegeneticmapofmaizewith the intermatedB73xMo17 (IBM)population.Plant Mol. Biol.48:453–461.
Lindblad-Toh, K., Winchester, E., Daly, M., Wang, D., Hirschhorn, J.N., Laviolette, J.P., Ardie, K., Reich, D.E., Robinson, E., Sklar, P., Shah, N., Thomas, D., Fan, J.B., Gingeras, T., Warrington, J., Patil, N., Hudson, T.J. & Lander, E.S.2000.Large-scalediscoveryandgenotypingofsingle-nucleotidepolymorphismsinthemouse.Nature Genet.24:381–386.
Manly, K.F. & Olson, J.M. 1999. Overview of QTL mapping software and introduction to MapManagerQT.Mamm. Genome10:327–334.
Mazur, B.J. 1995. Commercializing the products of plant biotechnology. Trends Biotech. 13:319–323.
Melchinger, A.E., Utz, H.F. & Schön, C.C.1998.Quantitativetraitlocus(QTL)mappingusingdif-ferenttestersandindependentpopulationsamplesinmaizerevealslowpowerofQTLdetectionandlargebiasinestimatesofQTLeffects.Genetics149:383–403.
Mohan, M., Nair, S., Krishna, T.G., Yano, M., Bhatia, C.R. & Sasaki, T.1997.Genomemapping,molecularmarkersandmarker-assistedselectionincropplants.Mol. Breed.3:87–103.
Moreau, L., Charcosset, A. & Gallais, A.2004.Experimentalevaluationofseveralcyclesofmarker-assistedselectioninmaize.Euphytica137:111–118.
Moreau, L., Charcosset, A., Hospital, F. & Gallais, A.1998.Marker-assistedselectionefficiencyinpopulationsoffinitesize.Genetics 148:1353–1365.
Moreau, L., Lemarié, S., Charcosset, A. & Gallais, A.2000.Economicefficiencyofonecycleofmarker-assistedselection.Crop Sci.40:329–337.
Morris, M., Dreher, K., Ribaut, J-M. & Khairallah, M.2003.Moneymatters (II):costsofmaizeinbred line conversion schemes at CIMMYT using conventional and marker-assisted selection.Mol. Breed.11:235–247.
Naylor, R.L., Falcon, W.P., Goodman, R.M., Jahn, M.M., Sengooba, T., Tefera, H. & Nelson, R.J.2004.Biotechnology in thedevelopingworld: acase for increased investments inorphancrops.Food Policy29:15–44.
Niebur, W.S., Rafalski, J.A., Smith, O.S. & Cooper, M.2004.Applicationsofgenomicstechnolo-giestoenhancerateofgeneticprogressforyieldofmaizewithinacommercialbreedingprogram.InT.Fischer,N.Turner,J.Angus,L.McIntyre,M.Robertson,A.Borrell&D.Lloyd,eds.New directions for a diverse planet. Proc. 4th Internat. Crop Sci. Congr.,Brisbane,Australia.(availableatwww.cropscience.org.au/icsc2004).

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish148
Openshaw, S. & Frascaroli, E. 1997.QTLdetectionandmarker-assistedselectionforcomplextraitsinmaize.Proc. 52nd Ann. Corn and Sorghum Res. Conf., pp.44–53.Washington,DC,AmericanSeedTradeAssociation.
Parisseaux, B. & Bernardo, R.2004.In silicomappingofquantitativetraitlociinmaize.Theor. Appl. Genet.109:508–514.
Peleman, J.D. & van der Voort, J.R.2003.Breedingbydesign.Trends Plant Sci.7:330–334.Penner, G.A., Bush, A., Wise, R., Kim, W., Domier L., Kasha, K., Laroche, A., Scoles, G., Molner,
S.J. & Fedak, G.1993.ReproducibilityofrandomamplifiedpolymorphicDNA(RAPD)analysisamonglaboratories.PCR Meth. Appl.2:341–345.
Pollak, L.M. 2003.Thehistoryandsuccessofthepublic-privateprojectongermplasmenhancementofmaize(GEM).Adv. Agron.78:45–87.
Rafalski, A. 2002. Applications of single nucleotide polymorphisms in crop genetics. Curr. Opin. Plant Biol.5:94–100.
Ragot, M. & Hoisington, D.A.1993.Molecularmarkersforplantbreeding:comparisonsofRFLPandRAPDgenotypingcosts.Theor. Appl. Genet.86:975–984.
Ragot M., Biasiolli, M., Delbut, M.F., Dell’Orco, A., Malgarini L., Thevenin, P., Vernoy, J., Vivant, J., Zimmermann, R. & Gay, G. 1995.Marker-assistedbackcrossing: apractical example.InA.Bervillé&M.Tersac,eds.Les Colloques, No. 72, Techniques et utilisations des marqueurs molécu-laires,pp.45–56.Paris,France,INRA.
Ragot, M., Gay, G., Muller, J-P. & Durovray, J.2000.EfficientselectionfortheadaptationtotheenvironmentthroughQTLmappingandmanipulationinmaize.InJ-M.Ribaut&D.Poland,eds.Molecular approaches for the genetic improvement of cereals for stable production in water-limited environments,pp.128–130.México,D.F.,CIMMYT.
Ribaut, J-M. & Betrán, J.1999.Singlelarge-scalemarker-assistedselection(SLS-MAS).Mol. Breed.5:531–541.
Ribaut, J-M., Jiang, C. & Hoisington, D.2002.Efficiencyofagene introgressionexperimentbybackcrossing.Crop Sci.42:557–565.
Ribaut, J-M., Edmeades, G., Perotti, E. & Hoisington, D.2000.QTLanalyses,MASresults,andperspectivesfordrought-toleranceimprovementintropicalmaize.InJ-M.Ribaut&D.Poland,eds.Molecular approaches for the genetic improvement of cereals for stable production in water-lim-ited environments,pp.131–136.México,D.F.,CIMMYT.
Robert, V.J.M., West, M.A.L., Inai, S., Caines, A., Arntzen, L., Smith, J.K. & St-Clair, D.A.2001.Marker-assistedintrogressionofblackmoldresistanceQTLallelesfromwildLycopersicon chesma-niitocultivatedtomato(L. esculentum)andevaluationofQTLphenotypiceffects.Mol. Breed.8:217–233.
Saiki, R.K., Gelfand, D.H., Stoffel, S., Scharf, S., Higuchi, R.H., Horn, G.T., Mullis, K.B. & Erlich, H.A.1988.Primer-directedenzymaticamplificationofDNAwithathermostableDNApolymer-ase. Science239:487–491.
Salvi, S., Tuberosa, R., Chiapparino, E., Maccaferri, M., Veillet, S., van Beuningen, L., Isaac, P., Edwards, K. & Phillips, R.L.2002.TowardspositionalcloningofVgt1,aQTLcontrolling thetransitionfromthevegetativetothereproductivephaseinmaize.Plant Mol. Biol.48:601–613.
Sax, K. 1923.TheassociationofsizedifferenceswithseedcoatpatternandpigmentationinPhaseolus vulgaris.Genetics8:552–560.
Senior, M.L. & Heun, M.1993.Mappingmaizemicrosatellitesandpolymerasechainreactionconfir-

Chapter 8 – Marker-assisted selection in maize 149
mationofthetargetrepeatsusingaCTprimer.Genome 36:884–889.Senior, M.L., Chin, E.C.L., Lee, M., Smith, J.S.C. & Stuber, C.W. 1996. Simple sequence repeat
markersdevelopedfrommaizesequencesfoundintheGENEBANKdatabase:Mapconstruction.Crop Sci.36:1676–1683.
Servin, B. & Hospital, F. 2002.Optimalpositioningofmarkers to controlgeneticbackground inmarker-assistedbackcrossing.J. Hered.93:214–217.
Shattuck-Eidens, D.M., Bell, R.N., Neuhausen, S. & Helentjaris, T.1990.DNAsequencevaria-tionwithinmaizeandmelon:Observationsfrompolymerasechainreactionanddirectsequencing.Genetics 126:207–217.
Skroch, P. & Nienhuis, J.1995.ImpactofscoringerrorandreproducibilityofRAPDdataonRAPDbasedestimatesofgeneticdistance.Theor. Appl. Genet.91:1086–1091.
Smith, J.S.C. &. Smith, O.S.1992.Fingerprintingcropvarieties.Adv. Agron.47:85–140.Smith, O.S., Hoard, K., Shaw, F. & Shaw, R.1999.Predictionofsingle-crossperformance.InJ.G.
Coors&S.Pandey,eds.The genetics and exploitation of heterosis in crops,pp.277–285.Madison,WI,USA,AmericanSocietyofAgronomy.
Sprague, G.F. &. Dudley, J.W.1988.Corn and corn improvement,3rdedition.Madison,Wisconsin,USA,AmericanSocietyofAgronomy.
Stam, P. 1995. Marker-assisted breeding. In J.W. Van Ooijen & J. Jansen, eds. Biometrics in plant breeding: applications of molecular markers.Proc. 9th Mtg. EUCARPIA Section on Biometrics in Plant Breeding,pp.32–44.Wageningen,Netherlands,CPRO-DLO.
Stromberg, L.D., Dudley, J.W. & Rufener, G.K. 1994. Comparing conventional early generationselectionwithmolecularmarkerassistedselectioninmaize.Crop Sci.34:1221–1225.
Stuber, C.W. 1992. Biochemical and molecular markers in plant breeding. In J. Janick, ed. Plant Breeding Rev.pp37–61.
Stuber, C.W. & Moll, R.H.1972.Frequencychangesofisozymeallelesinaselectionexperimentforgrainyieldinmaize(Zea mays L.).Crop Sci.12:337–340.
Tanksley, S.D., Young, N.D., Patterson, A.H. & Bonierbale, M.W.1989.RFLPmappinginplantbreeding:Newtoolsforanoldscience.Biotech.7:257–264.
Tanksley, S.D., Grandillo, S., Fulton, T.M., Zamir, D., Eshed, Y., Petiard, V., Lopez, J. & Beck-Bunn, T.1996.AdvancedbackcrossQTLanalysis inacrossbetweenaneliteprocessing lineoftomatoanditswildrelativeL. pimpinellifolium.Theor. Appl. Genet.92:213–224.
Tautz, D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA.Nucl. Acids Res.17:6463–6471.
Thornsberry, J.M., Goodman, M.M., Doebley, J., Kresovich, S., Nielsen, D. & Buckler, E.S. 2001.Dwarf8polymorphismsassociatewithvariationinfloweringtime.Nature Genet.28:286–289.
Troyer, A.F.2004.Persistentandpopulargermplasminseventycenturiesofcornevolution.InC.W.Smith,J.Betrán&E.C.A.Runge,eds.Corn: origin, history, technology, and production, pp.133–232.Hoboken,NJ,USA,JohnWiley&Sons.
Utz, H.F. & Melchinger, A.E.1996.PLABQTL:aprogramforcompositeintervalmappingofQTL.J. Quant. Trait Loci2:1.
van Berloo, R. & Stam, P.1998.Marker-assistedselectioninautogamousRILpopulations:asimula-tionstudy.Theor. Appl. Genet.96:147–154.
van Berloo, R. & Stam, P.2001.Simultaneousmarker-assistedselectionformultipletraitsinautoga-mouscrops.Theor. Appl. Genet.102:1107–1112.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish150
van Ooijen, J.W. & Maliepaard, C.1996.MapQTLversion3.0:softwareforthecalculationofQTLpositionsongeneticmaps.Plant Genome IV,SanDiego,CA,USA(availableat www.intl-pag.org/4/abstracts/p316).
Varshney, R.K., Graner, A. & Sorrels, M.E.2005.Genomics-assistedbreeding forcrop improve-ment.Trends Plant Sci.10:621–630.
Visscher, P.M., Haley, C.S. & Thompson, R.1996.Marker-assistedintrogressioninbackcrossbreed-ingprograms.Genetics 144:1923–1932.
Vos P., Hogers, R., Bleeker, M., Reijans, M., van de Lee, T., Hornes, M., Frijters, A., Pot, J., Peleman, J., Kuiper, M. & Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting.Nucl. Acids Res.23:4407–4414.
Welsh, J. & McClelland, M.1990.FingerprintinggenomesusingPCRwitharbitraryprimers.Nucl. Acids Res.18:7213–7218.
Willcox, M.C., Kairallah, M.M., Bergvinson, D., Crossa, J., Deutsch, A., Edmeades, G.O., González-de-León, D., Jiang, C., Jewell, D.C., Mihm, A J., Williams, W.P. & Hoisington, D.2002.Selectionforresistancetosouthwesterncornborerusingmarker-assistedandconventionalbackcrossing.Crop Sci.42:1516–1528.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A. & Tingey, S.V. 1990. DNA poly-morphisms amplified by arbitrary primers are useful as genetic markers. Nucl. Acids Res. 18:6531–6535.
Xiao, J., Li, J., Grandillo, S., Nag Ahn, S., Yuan, L., Tanksley, S.D. & McCouch, S.R. 1998.Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon.Genetics 150:899–909.
Xie, C. & Xu, S.1998.Efficiencyofmultistagemarker-assistedselectionintheimprovementofmul-tiplequantitativetraits.Heredity 80:489–498.
Young, N.D. 1999. A cautiously optimistic view for marker-assisted breeding. Mol. Breed. 5:505–510.
Yousef, G.G. & Juvik, J.A. 2001. Comparison of phenotypic and marker-assisted selection formquantitativetraitsinsweetcorn.Crop Sci.41:645–655.
Zeng, Z-B.1993.Theoreticalbasisforseparationofmultiplelinkedgeneeffectsinmappingquantita-tivetraitloci.Proc. Natl. Acad. Sci. USA90:10972–10976.
Zeng, Z-B.1994.Precisionmappingofquantitativetraitloci.Genetics 136:1457–1468.

Chapter 9
molecular marker-assisted selection for resistance to pathogens
in tomato
Amalia Barone and Luigi Frusciante

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish152
SummarySincethe1980s,theuseofmolecularmarkershasbeensuggestedtoimprovetheefficiencyofreleasingresistantvarieties,thusovercomingdifficultiesmetwithclassicalbreeding.Fortomato,ahigh-densitymolecularmapisavailableinwhichmorethan40resistancegenesarelocalized.Markerslinkedtothesegenescanbeusedtospeedupgenetransferandpyra-miding.SuitablePCRmarkerstargetingresistancegeneswereconstructeddirectlyonthesequencesofresistancegenesoronrestrictionfragmentlengthpolymorphisms(RFLPs)tightlylinkedtothem,andusedtoselectresistantgenotypesinbackcrossschemes.Insomecases, theBC5generationwasreached,andgenotypes thatcumulated twohomozygousresistantgeneswerealsoobtained.Theseresultssupportedthefeasibilityofusingmarker-assistedselection(MAS)intomatoandreinforcingthepotentialofthisapproachforothergenes,which is todayalsodrivenbythedevelopmentofnewtechniquesand increasingknowledgeaboutthetomatogenome.

Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 153
introduCtionTomato (Solanum lycopersicum, formerlyLycopersicon esculentum)isoneofthemostwidelygrownvegetablecropsintheworld.It isusedasafreshvegetableandcanalsobe processed and canned as a paste, juicesauce,powderorasawhole.Worldvolumehas increased approximately 10percentsince1985,reflectingasubstantialincreaseindietaryuseofthetomato.Nutritionally,tomato is a significant source of vitaminsAandC.Furthermore,recentstudieshaveshowntheimportanceoflycopene,amajorcomponent of red tomatoes, which hasantioxidant properties and may help toprotect against cancer and heart disease(RaoandAgarwal,2000).
One of the main constraints to tomatocultivationisdamagecausedbypathogens,including viruses, bacteria, nematodes andfungi, which cause severe losses in pro-duction. The control of pathogen spreadmainlyinvolvesthreestrategies:husbandrypractices,applicationofagrochemicalsanduseof resistantvarieties.Husbandry tech-niquesgenerallyhelptorestrictthespreadofpathogensandtheirvectorsaswellastokeep plants healthy, thus allowing patho-genattacktobelimited.Chemicalcontrolgivesgoodresultsforsomepathogens,butpoor results against others, such as bacte-ria,andhaspracticallynoeffectonviruses.Moreover, reducing chemical treatmentslowers the health risks to farmers andconsumers. Therefore, in order to achievesustainable agriculture and obtain high-quality, safeandhealthyproducts, theuseofresistantvarietiesisoneoftheprincipaltoolstoreducepathogendamage.
Since the early part of the twentiethcentury, breeding for disease resistance hasbeen a major method for controlling plantdisease.Varieties that are resistantor toler-anttooneoranumberofspecificpathogens
are already available for many crops, andhybrids with multiple resistance to severalpathogens exist and are currently used invegetable production. In tomato, geneticcontrol of pathogens is a very useful prac-tice with most resistance being monogenicand dominant. Various sources of resist-ancehavebeenusedintraditionalbreedingprogrammes, and resistant breeding lines,varietiesandF1hybridshavebeendevelopedwith varying stability and levels of expres-sion(Table1)(Laterrot,1996;GardnerandShoemaker,1999;Scott,2005).
marker-aSSiSted Breeding for pathogen reSiStanCeAlthough conventional plant breedinghas had a significant impact on improv-ing tomato for resistance to importantdiseases, the time-consuming process of
table 1list of pathogen resistances present in tomato breeding lines, varieties and f1 hybrids obtained through conventional breeding
virus
beet curly top virus (bctV)tobacco mosaic virus (tMV)tomato mosaic virus (toMV)tomato yellow leaf curl virus (tYlcV)tomato spotted wilt virus (tSWV) Bacteria Corynebacterium michiganense
Pseudomonas solanacearum
Pseudomonas syringae pv. tomato
nematodes Meloidogyne spp.fungiAlternaria alternata f. sp. lycopersici
Alternaria solani
Cladosporium fulvum
Fusarium oxysporum f. sp. lycopersici
Fusarium oxysporum f. sp. radicis-lycopersici
Phytophthora infestans
Pyrenochaeta lycopersici
Stemphylium solani
Verticillium dahliae
Modified from laterrot (1996) and updated as reported in the text.
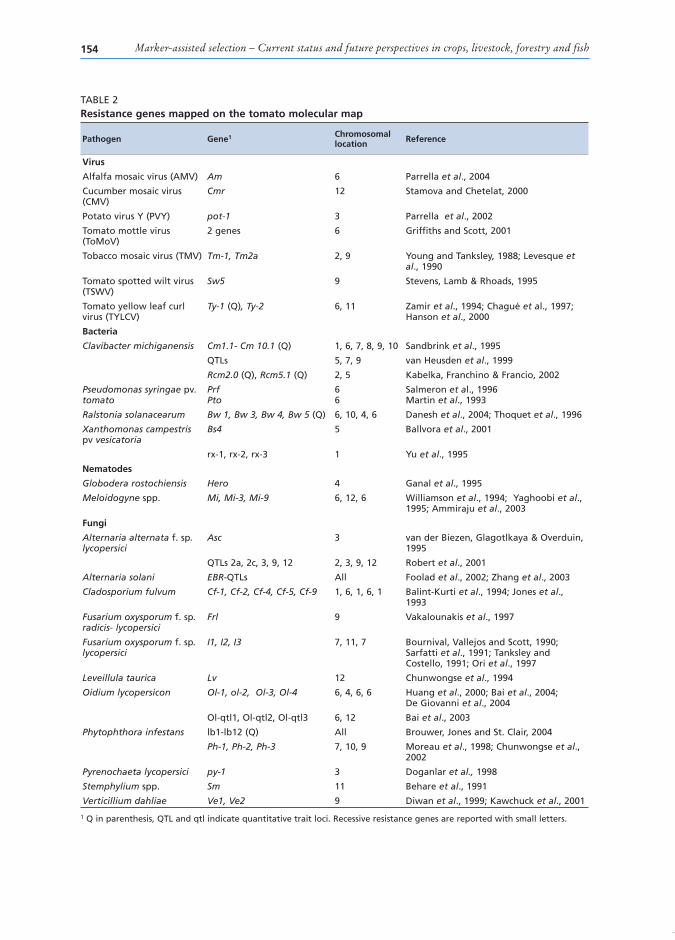
Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish154
table 2resistance genes mapped on the tomato molecular map
pathogen gene1 Chromosomal location reference
virus
alfalfa mosaic virus (aMV) Am 6 Parrella et al., 2004
cucumber mosaic virus (cMV)
Cmr 12 Stamova and chetelat, 2000
Potato virus Y (PVY) pot-1 3 Parrella et al., 2002
tomato mottle virus (toMoV)
2 genes 6 Griffiths and Scott, 2001
tobacco mosaic virus (tMV) Tm-1, Tm2a 2, 9 Young and tanksley, 1988; levesque et al., 1990
tomato spotted wilt virus (tSWV)
Sw5 9 Stevens, lamb & Rhoads, 1995
tomato yellow leaf curl virus (tYlcV)
Ty-1 (Q), Ty-2 6, 11 Zamir et al., 1994; chagué et al., 1997; Hanson et al., 2000
Bacteria
Clavibacter michiganensis Cm1.1- Cm 10.1 (Q) 1, 6, 7, 8, 9, 10 Sandbrink et al., 1995
Qtls 5, 7, 9 van Heusden et al., 1999
Rcm2.0 (Q), Rcm5.1 (Q) 2, 5 Kabelka, Franchino & Francio, 2002
Pseudomonas syringae pv. tomato
Prf Pto
6 6
Salmeron et al., 1996 Martin et al., 1993
Ralstonia solanacearum Bw 1, Bw 3, Bw 4, Bw 5 (Q) 6, 10, 4, 6 Danesh et al., 2004; thoquet et al., 1996
Xanthomonas campestris pv vesicatoria
Bs4 5 ballvora et al., 2001
rx-1, rx-2, rx-3 1 Yu et al., 1995
nematodes
Globodera rostochiensis Hero 4 Ganal et al., 1995
Meloidogyne spp. Mi, Mi-3, Mi-9 6, 12, 6 Williamson et al., 1994; Yaghoobi et al., 1995; ammiraju et al., 2003
fungi
Alternaria alternata f. sp. lycopersici
Asc 3 van der biezen, Glagotlkaya & overduin, 1995
Qtls 2a, 2c, 3, 9, 12 2, 3, 9, 12 Robert et al., 2001
Alternaria solani EBR-Qtls all Foolad et al., 2002; Zhang et al., 2003
Cladosporium fulvum Cf-1, Cf-2, Cf-4, Cf-5, Cf-9 1, 6, 1, 6, 1 balint-Kurti et al., 1994; Jones et al., 1993
Fusarium oxysporum f. sp. radicis- lycopersici
Frl 9 Vakalounakis et al., 1997
Fusarium oxysporum f. sp. lycopersici
I1, I2, I3 7, 11, 7 bournival, Vallejos and Scott, 1990; Sarfatti et al., 1991; tanksley and costello, 1991; ori et al., 1997
Leveillula taurica Lv 12 chunwongse et al., 1994
Oidium lycopersicon Ol-1, ol-2, Ol-3, Ol-4 6, 4, 6, 6 Huang et al., 2000; bai et al., 2004; De Giovanni et al., 2004
ol-qtl1, ol-qtl2, ol-qtl3 6, 12 bai et al., 2003
Phytophthora infestans lb1-lb12 (Q) all brouwer, Jones and St. clair, 2004
Ph-1, Ph-2, Ph-3 7, 10, 9 Moreau et al., 1998; chunwongse et al., 2002
Pyrenochaeta lycopersici py-1 3 Doganlar et al., 1998
Stemphylium spp. Sm 11 behare et al., 1991
Verticillium dahliae Ve1, Ve2 9 Diwan et al., 1999; Kawchuck et al., 2001
1 Q in parenthesis, Qtl and qtl indicate quantitative trait loci. Recessive resistance genes are reported with small letters.

Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 155
making crosses and backcrosses, and theselection of the desired resistant progeny,make it difficult to respond adequately tothe evolution of new virulent pathogens.Moreover,severalinterestingresistancesaredifficulttousebecausethediagnostictestsoftencannotbedevelopedduetothechal-lenge posed by inoculum production andmaintenance.Inaddition,wheresymptomsaredetectableonlyonadultplants and/orfruits, diagnostic tests can be particularlyexpensiveanddifficulttoperform.
Since the 1980s, the use of molecularmarkers has been suggested as a tool forbreeding many crops, including tomato.Inthelasttwodecades,molecularmarkershavebeenemployedtomapandtagmajorgenes and quantitative trait loci (QTL)involved in monogenic and polygenicresistance control, known respectively asverticalandhorizontal resistance.Todate,more than 40 genes (including many sin-gle genes and QTL) that confer resistancetoallmajorclassesofplantpathogenshavebeenmappedonthetomatomolecularmap(Table2) and/or cloned from Solanaceous species, as reportedbyGrube,Radwanskiand Jahn (2000). Since then, other resist-ance genes together with resistance geneanalogues (RGAs), which are structurallyrelated sequences based on the proteindomain shared among cloned R genes(Leisteret al.,1996),havebeenaddedtothemap. A molecular linkage map of tomatobasedonRGAshasalsobeenconstructedinwhich29RGAswerelocatedonnineofthe12tomatochromosomes(Fooladet al.,2002;Zhanget al.,2002).SeveralRGAlociwere found in clusters and their locationscoincided with those of several knowntomatoRgenesorQTL.Thismapprovidesabasisforfurtheridentifyingandmappinggenes and QTL for disease resistance andwillbeusefulforMAS.
In fact, independently of the type ofmarker used for selection, by making itpossibletofollowthegeneunderselectionthrough generations rather than waitingforphenotypicexpressionoftheresistancegene, markers tightly linked to resistancegenes can greatly aid disease resistanceprogrammes. In particular, genetic map-pingofdiseaseresistancegeneshasgreatlyimproved the efficiency of plant breedingandalsoledtoabetterunderstandingofthemolecularbasisofresistance.
DNAmarkertechnologyhasbeenusedincommercialplantbreedingprogrammessincetheearly1990s,andhasprovedhelp-ful for the rapid and efficient transfer ofuseful traits into agronomically desirablevarietiesandhybrids(Tanksleyet al.,1989;LefebvreandChèvre,1995).Markerslinkedto disease resistance loci can now be usedfor MAS programmes, thus also allowingseveralresistancegenestobecumulatedinthesamegenotype(“pyramiding”ofresist-ance genes), and they may be also usefulfor cloning and sequencing the genes. Intomato, several resistancegeneshavebeensequenced to date, among them Cf-2, Cf-4,Cf-5,Cf-9,Pto,Mi,I2,andSw5.TheseclonedRgenesnowprovidenewtoolsfortomatobreeders to improve the efficiencyofbreedingstrategies,viaMAS.AlthoughMASisstillnotusedroutinelyforimprov-ing disease resistance in many importantcrops (Michelmore,2003), it isbeingusedby seed companies forimproving simpletraitsintomato(FooladandSharma,2005).Furthermore, while the deep knowledgeof the tomato genome and the availabilityof a high-density molecular map for thisspecies (Pillen et al., 1996) should pro-vide further opportunities to acceleratebreeding throughMAS, the time-consum-ing and expensive process of developingmarkers associated with genes of inter-

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish156
est and the high cost of genotyping largepopulationshasandwill continue to limitthe use of MAS in most tomato breedingprogrammes.
The potential of MAS to speed up thebreedingoftomatousingmolecularmark-ers linked to various resistance genes hasbeen examined in the authors’ laboratory.The two main goals of the research wereto find the most suitable markers, andto test the feasibility of MAS for pyra-miding resistance genes in tomato “elite”lines selected for their good processingqualities.
StrategieS for gene tranSfer and pyramidingSix tomato genotypes carrying variousresistancegenes(Table3)werecrossedwithtomato“elite”linespreviouslyselectedforyield and quality but lacking resistancetraits.Eachresistantgenotypewascrossedinitially with each “elite” tomato line andvarious backcross schemes were then car-riedoutstartingfromdifferentF1hybrids.Ateachbackcrossgenerationthescreeningofresistantgenotypeswasperformedusingmolecularmarkers linkedtotheresistancegenes and DNA extracted from youngleaves at seedling stage. Only resistantplantswerethentransplantedandgrowninthegreenhouse.Atflowering,crossesweremade with the recurrent parent to obtainthesubsequentgenerations.
AstheefficiencyofMASdependsontheavailability of polymerase chain reaction(PCR)-basedmarkershighly linked to theresistance gene to be selected, for eachresistance gene the most suitable markersystemwas investigated.Forthispurpose,three different strategies were undertaken.The first involvedsearchingPCRmarkersalready available in the literature andverifying their usefulness on the geneticmaterial used. The second consisted ofdesigningPCRprimersfromthesequenceofclonedgenesreportedintheGeneBankdatabase of the National Center forBiotechnology Information (www.ncbi.nlm.nih.gov/Genbank), while the thirdinvolved designing PCR primers fromRFLPmarkers tightly linked to resistancegenes.Thislaststrategywasmadepossibleby theavailabilityof sequencesofvariousmapped tomato RFLPs in the SolGenesdatabase(www.sgn.cornell.edu).
Inmostcases,theresultswereobtainedusing cleaved amplified polymorphicsequences(CAPS;KonieczynandAusubel,1993),whichrequireonePCRreactionfol-lowedbyrestrictiondigestoftheamplifiedfragment.Inthreecases(markerslinkedtogenesMi,Sw5andTm2a),theprimersandenzymesusedwerethosereportedinthelit-erature(Williamsonet al.,1994;Folkertsmaet al.,1999;Sobiret al.,2000). In thecaseof gene py-1, the procedure reported inthe literature (Doganlar et al., 1998) was
table 3tomato genotypes used as resistant parents in the backcross breeding schemes. for each genotype resistant genes are reported
genotype resistance gene pathogen
Momor Frl, Tm2a, Ve Fusarium oxysporum f. sp. radicis-lycopersici, tMV, Verticillium dahliae
Motelle I2, Mi, Ve Fusarium oxysporum f. sp lycopersici, Meloidogyne spp., Verticillium dahliae
okitzu I2, Tm2a Fusarium oxysporum f. sp lycopersici, tMV
ontario Pto Pseudomonas syringae
Pyrella py-1 Pyrenochaeta lycopersici
Stevens Sw5 tSWV
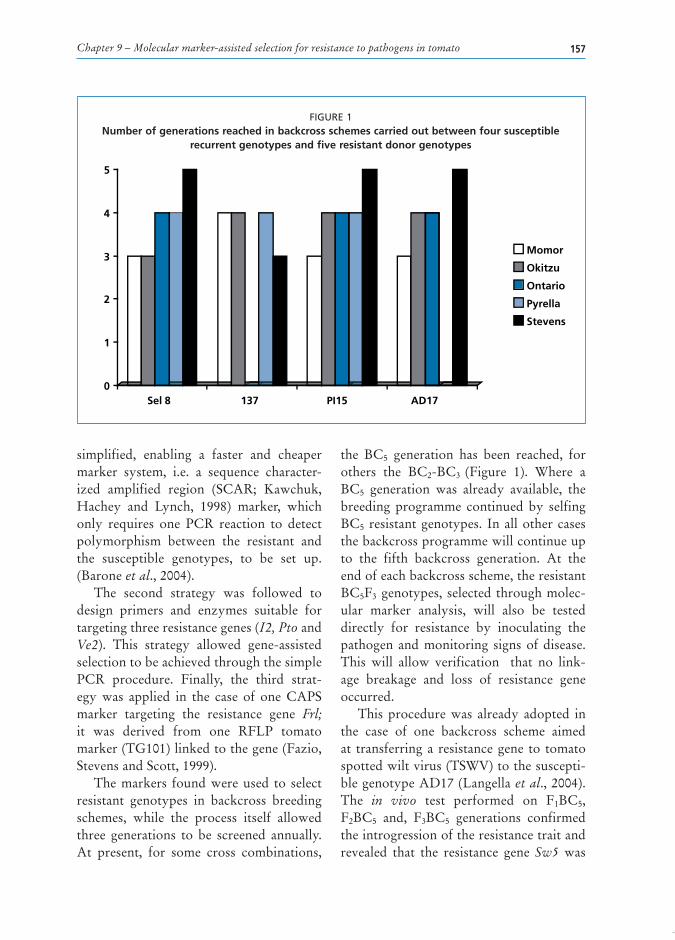
Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 157
simplified, enabling a faster and cheapermarker system, i.e. a sequence character-ized amplified region (SCAR; Kawchuk,Hachey and Lynch, 1998) marker, whichonly requires one PCR reaction to detectpolymorphism between the resistant andthe susceptible genotypes, to be set up.(Baroneet al.,2004).
The second strategy was followed todesign primers and enzymes suitable fortargetingthreeresistancegenes(I2, PtoandVe2). This strategy allowed gene-assistedselectiontobeachievedthroughthesimplePCR procedure. Finally, the third strat-egy was applied in the case of one CAPSmarker targeting the resistance gene Frl;it was derived from one RFLP tomatomarker(TG101)linkedtothegene(Fazio,StevensandScott,1999).
Themarkers foundwereused to selectresistant genotypes in backcross breedingschemes, while the process itself allowedthree generations to be screened annually.At present, for some cross combinations,
the BC5 generation has been reached, forothers the BC2-BC3 (Figure 1). Where aBC5 generation was already available, thebreeding programme continued by selfingBC5 resistantgenotypes. Inallothercasesthebackcrossprogrammewillcontinueupto the fifth backcross generation. At theendofeachbackcrossscheme,theresistantBC5F3genotypes, selectedthroughmolec-ular marker analysis, will also be testeddirectly for resistance by inoculating thepathogen and monitoring signs of disease.This will allow verification that no link-age breakage and loss of resistance geneoccurred.
This procedure was already adopted inthe case of one backcross scheme aimedat transferringa resistancegene to tomatospottedwiltvirus(TSWV)tothesuscepti-blegenotypeAD17(Langellaet al.,2004).The in vivo test performed on F1BC5,F2BC5 and, F3BC5 generations confirmedtheintrogressionoftheresistancetraitandrevealed that the resistance gene Sw5 was
0
1
2
3
4
5
Sel 8 137 PI15 AD17
Momor
Okitzu
Ontario
Pyrella
Stevens
FiGURe 1number of generations reached in backcross schemes carried out between four susceptible
recurrent genotypes and five resistant donor genotypes

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish158
fixedatthehomozygousstageattheF3BC5generation.
Finally,besidesthetransferofoneresist-ance gene to each susceptible genotype, acrossing scheme was undertaken to accu-mulatetwoorthreeresistancegenesinthesame genotype. In this case, the decisionwasmadetostopthebackcrossschemeattheBC3orBC4generationasbothparen-tallineswerecultivatedvarietiesandhencegenetically very similar, and therefore therecovery of the recurrent genome couldbesatisfactory.F1BC4hybridscarryingthesame genetic background in the recurrentparent have been intercrossed, followingthe breeding scheme shown in Figure 2.At the end of each F1BC4 x F1BC4 cross
and after selecting the genotypes carryingall theresistantallelesat theheterozygouslevel,oneortwoselfinggenerationswillbecarried out to fix all the resistant genes atthehomozygouslevel.
Thisstrategyhasalreadystartedinsomecasesandthefirsthomozygousmultiresist-ant genotypes have been obtained. AlsoavailablearetwoF2genotypesoutof52ana-lysedplants,obtainedbyintercrossingtheF1BC4 progeny from PI15 x Stevens withthe F1BC4 progeny from PI15 x Ontario(Table4).ThisF2generationexhibitedtwogenotypes carrying both resistant genesSw5 and Pto at the homozygous level aswellas29genotypescarryingbothgenesattheheterozygouslevel.
Theworkreportedhereontransferringresistance genes among tomato genotypesdemonstrates the usefulness of MAS forimproving traditional breeding strategies.The contribution of molecular markerslinkedtoresistancegeneswasveryefficientin reducing the time and space necessaryfor selection, enabling both early screen-ing for resistance and reduced numbers ofgenotypes to be transplanted. The mostchallenging work was the search for suit-able markers, which often required bothconsiderable time and financial resources.Different strategies were used successfullytofindthemostsuitablemarkerstoperformMASfortransferringeightresistancegenesintosuperiortomatogenotypes;suchstrat-egiescouldberepeatedintomatoformanyother genes due to advanced molecularknowledgeofthegenomeofthisspecies.
perSpeCtiveSThe availability of PCR-based markersformanyresistancegenesallowsMASforbiotic resistance in tomato to be appliedsuccessfully inany laboratorywithouttheneed for highly sophisticated techniques.
FiGURe 2 Breeding scheme used for the
pyramiding of two resistant homozygous genes (Sw5 and Pto) in the
susceptible genotype pi15
PI15 x Stevens Sw5/Sw5 PI15 x Ontario Pto/Pto
F1BC 1 Sw5/-
F1BC 2 Sw5/-
F1BC 3 Sw5/-
F1BC 4 Sw5/-
F1BC 1 Pto/-
F1BC 2 Pto/-
F1BC 3 Pto/-
F1BC 4 Pto/-X
F 1 (F 1BC 4 x F 1BC 4) Sw5/-, Pto/-
F 2 (F 1BC 4 x F 1BC 4) Sw5/Sw5, Pto/Pto
X
Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 159
Indeed, once a marker has been set up,its use on large populations for resistancescreening is then routine. Technical facili-tiesaretodayavailableforscreeningmanysamples simultaneously and also costs forequipmentaredecreasing. Inaddition, therapiddevelopmentofnewmoleculartech-niques, combined with the ever-increasingknowledge about the structure and func-tion of resistance genes (Hulbert et al.,2001),willhelp to identifynewmolecularmarkersforMAS,suchassinglenucleotidepolymorphisms (SNPs). Moreover, thanksto the International Solanaceae GenomeProject (SOL), sequencing of the tomatogenome is in progress, and in a few yearsthis will enhance information on resist-ance genes. This in turn will facilitate thedevelopment of molecular markers fromtranscribedregionsofthegenome,therebyallowinglarge-scalegene-assistedselection(GAS)tobeachieved.
Over the coming years, selection forpathogen resistance in tomato will beunderpinnedbyresearchaimedat:mappingother resistance genes for new pathogens;developingPCR-based functionalmarkers(AndersenandLudderstedt,2003);design-
ing the most suitable breeding schemes(Peleman and van der Voort, 2003), espe-cially for transferring QTL resistances;large-scale screening through automa-tion;allele-specificdiagnostics(Yanget al.,2004); and DNA microarrays (Borevitzet al., 2003). In effect, the combination ofnewknowledgeandnewtoolswillleadtochangesinthestrategiesusedforbreedingby exploiting the potential of integrating“omics” disciplines with plant physiologyand conventional plant breeding, a proc-ess that will drive the evolution of MASintogenomics-assistedbreeding(Morganteand Salamini, 2003; Varshney, Graner andSorrells,2005).
aCknowledgementSThe authors wish to thank Mr MarkWalters for editing the manuscript andAngelaCozzolino forher technicalassist-ance. This research was partially financedby the Ministry of Agriculture andForestry of Italy in the framework ofprojectPROMAR(PlantProtectionbytheUse of Molecular Markers). Contributionno.119fromtheDepartmentofSoil,PlantandEnvironmentalSciences.
table 4 resistant heterozygous plants obtained in some cross combinations realized for pyramiding of resistance genes
Cross pyramided genes
generation analysed plant (number)
resistant plant (number)
(137 x Momor) F1bc3 x (137 x ontario) F1bc3
Tm2a – Frl Ve – Pto F1 50 7
(aD17 x okitzu) F1bc4 x (aD17 x Stevens) F1bc4
Tm2a – Frl
Sw5F1 24 5
(aD17 x ontario) F1bc4 x (aD17 x Stevens) F1bc4
Pto – Sw5 F1 24 6
(Pi15 x Stevens) F1bc4 x (Pi15 x ontario) F1bc4
Sw5 - Pto F2 52 29
referenCeSAmmiraju, J.S.S., Veremis, J.C., Huang, X., Roberts, P.A. & Kaloshian, I. 2003.Theheat-stable
root-nematoderesistancegeneMi-9fromLycopersicon peruvianumislocalizedontheshortarmofchromosome6.Theor. Appl. Genet.106:478–484.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish160
Andersen, J.R. & Lubberstedt, T.2003.Functionalmarkersinplants.Trends Plant Sci.8:554–560.Bai, Y., Huang, C-C., van der Hulst, R., Meijer-Dekens, F., Bonnema, G. & Lindhout, P. 2003.
QTLs for tomato powdery mildew resistance (Oidium lycopersici) in Lycopersicon parviflorumG1.1601co-localizewithtwoqualitativepowderymildewresistancegenes.Mol. Plant - Microbe Interact.16:169–176.
Bai, Y., van der Hulst, R., Huang, C-C., Wei, L., Stam, P. & Lindhout, P. 2004.MappingOl-4,ageneconferringresistancetoOidium neolycopersiciandoriginatingfromLycopersicon peruvianumLA2172,requiresmulti-allelic,singlelocusmarkers.Theor. Appl. Genet. 109:1215–1223.
Balint-Kurti, P.J., Dixon, M.S., Jnes, D.A., Norcott, K.A. & Jones, J.D.G. 1994. RFLP linkageanalysisoftheCf-4andCf-9genesforresistancetoCladosporium fulvumintomato.Theor. Appl. Genet.88:691–700.
Ballvora, A., Pierre, M., van den Ackerveken, G., Chornack, S., Rossier, O., Ganal, M., Lahaye, T. & Bonas, U.2001Geneticmappingand functionalanalysisof the tomatoBs4 locusgovern-ingrecognitionoftheXanthomonas campestrispv.vesicatoriaAvrBs4protein.Mol. Plant-Microbe Interact.14:629–638.
Barone, A., Langella, R., Ercolano, M.R., Di Matteo, A. & Monti, L.2004Strategiesforpathogenresistancegenetransferintomatowiththeaidofmolecularmarkers.In S.G.Pandalai,ed.Recent research developments in genetics and breeding,pp.211–222.Kerala,India,ResearchSignpost.
Behare, J., Laterrot, H., Sarfatti, M. & Zamir, D.1991.RFLPmappingoftheStemphyliumresist-ancegeneintomato.Mol. Plant-Microbe Interact.4:489–492.
Borevitz, J.O., Liang, D., Plouffe, D., Chang, H-S., Zhu, T., Weigel, D., Berry, C.C., Winzeler, E. & Chory, J. 2003.Large-scaleidentificationofsingle-feaqturepolymorphismsincomplexgenomes.Genome Res.13:513–523.
Bournival, B.L., Vallejos, C.E. & Scott, J.W. 1990.Geneticanalysisofresistancestorace1and2ofFusarium oxysporum f. sp. lycopersici from the wild tomato Lycopersicon pennellii. Theor. Appl. Genet.79:641–645.
Brouwer, D.J., Jones, E.S. & St Clair, D.A. 2004. QTL analysis of quantitative resistance toPhytophthora infestans(lateblight)intomatoandcomparisonstopotato.Genome47:475–492.
Chagué, V., Mercier, J.C., Guénard, M., de Courcel, A.G.L. & Vedel, F. 1997. Identification ofRAPDmarkerslinkedtoalocusinvolvedinquantitativeresistancetoTYLCVintomatobybulkedsegregantanalysis.Theor. Appl. Genet.95:671–677.
Chunwongse, J., Bunn, T.B., Crossman, C., Jing, J. & Tanksley, S.D. 1994.Chromosomallocaliza-tionandmolecular-markertaggingofthepowderymildewresistancegene(Lv)intomato.Theor. Appl. Genet. 89:76–79.
Chunwongse, J., Chunwongse, C., Black, L. & Hanson, P. 2002. MolecularmappingofthePh-3geneforlateblightresistanceintomato.J. Hort. Sci. & Biotech. 77:281–286.
Danesh, D., Aarons, S., McGill, G.E. & Young, N.D.1994.Geneticdissectionofoligenicresistancetobacterialwiltintomato.Mol. Plant-Microbe Interact.7:464–471.
De Giovanni, C., Dell’Orco, P., Bruno, A., Ciccarese, F., Lotti, C. & Ricciardi, L. 2004.IdentificationofPCR-basedmarkers(RAPD,AFLP)linkedtoanovelpowderymildewresistancegene(ol-2)intomato.Plant Sci. 166:41–48.
Diwan, N., Fluhr, R., Eshed, Y., Zamir, D. & Tanksley, S.D.1999.MappingofVeintomato:ageneconferringresistancetobroad-spectrumpathogen,Verticillium dahliaerace1.Theor. Appl. Genet.98:315–319.

Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 161
Doganlar, S., Dodson, J., Gabor, B., Beck-Bunn, T., Crossman, C. & Tanksley, S.D.1998Molecularmappingof thepy-1gene for resistance to corky root rot (Pyranochaeta lycopersici) in tomato.Theor. Appl. Genet.97:784–788.
Fazio, G., Stevens, M.R., & Scott, J.W. 1999.IdentificationofRAPDmarkers linkedto fusariumcrownandrootrotresistance(Frl)intomato.Euphytica105:205–210.
Folkertsma, R.T., Spassova, M.I., Prins, M., Stevens, M.R., Hille, J. & Goldbach, R.W. 1999.Construction of a bacterial artificial chromosome (BAC) library of Lycopersicon esculentum cv.StevensanditsapplicationtophysicallymaptheSw5locus.Mol. Breed. 5:197–207.
Foolad, M.R. & Sharma, A. 2005. Molecular markers as selection tools in tomato breeding. Acta Hort.695:225–240.
Foolad, M.R., Zhang, L.P., Khan, A.A., Nino-Liu, D. & Lin, G.Y.2002.IdentificationofQTLsforearlyblight(Alternaria solani)resistanceintomatousingbackcrosspopulationsofaLycopersicon esculentumxL. hirsutumcross.Theor. Appl. Genet.104:945–958.
Ganal, M.W., Simon, R., Brommonschenkel, S., Arndt, M., Tanksley, S.D. & Kumar, A. 1995.Geneticmappingofawide spectrumnematoderesistancegene (Hero) againstGloboderarosto-chiensisintomato.Mol. Plant-Microbe Interact.8:886–891.
Gardner, R.G. & Shoemaker, P.B.1999.“MountainSupreme”earlyblight-resistanthybridtomatoanditsparentsNCEBR-3andNCEBR-4tomatobreedinglines.Hort. Sci.34:745–746.
Griffiths, P.D. & Scott, J.W.2001.InheritanceandlinkageoftomatomottlevirusresistancegenesderivedfromLycopersicon chilenseaccessionLA1932.J. Am. Soc. Hort. Sci.126:462–467.
Grube, R.C., Radwanski, E.R. & Jahn, M.2000.ComparativegeneticsofdiseaseresistancewithintheSolanaceaeGenetics155:873–887.
Hanson, P.M., Bernacchi, D., Green, S., Tanksley, S.D. & Muniy-Appa, V.2000.Mappingawildtomatointrogressionassociatedwithtomatoleafcurlvirusresistanceinacultivatedtomatoline.J. Am. Soc. Hort. Sci.125:15–20.
Huang, C.C., van de Putte, P.M., Haanstra-van der Meer, J.G., Mejer-Dekens, F. & Lindhout, P. 2000. Characterization and mapping of resistance to Oidium lycopersicum in two Lycopersicon hirsutum accessions:evidenceforcloselinkageoftwoOl-genesonchromosome6.Heredity85:511–520.
Hulbert, S.H., Webb, C.A., Smith, S.M. & Sun, Q.2001.Resistancegenecomplexes:evolutionandutilization.Ann. Rev. Phytopathol.39:285–312.
Jones, D.A., Dickinson, M.J., Balint-Kurt, P.J., Dixon, M.S. & Jones, J.D.G.1993.TwocomplexresistancelocirevealedintomatobyclassicalandRFLPmappingofCf-2,Cf-4,Cf-5,andCf-9genesforresistancetoCladosporium fulvum.Mol. Plant-Microbe Interact.6:348–357.
Kabelka, E., Franchino, B. & Francio, D.M. 2002. Two loci from Lycopersicon hirsutum LA407confer resistance to strains of Clavibacter michiganense subsp. michiganensis. Phytopath. 92:504–510.
Kawchuck, L.M., Hachey, J. & Lynch, D.R.1998.Developmentof sequencecharacterizedDNAmarkerslinkedtoadominantVerticilliumwiltresistancegeneintomato.Genome 41:91–95.
Kawchuk, L.M., Hachey, J., Lynch, D.R., Kulcsar, F., van Rooijen, G., Waterer, D.R., Robertson, A., Kokko, E., Byers, R., Howar, R.J., Fischer, R. & Prufer, D. 2001.TomatoVediseaseresistancegenesencodecellsurface-likereceptors.Proc. Nat. Acad. Sci. USA98:6511–6515.
Konieczyn, A. & Ausubel, F.M. 1993. A procedure for mapping Arabidopsis mutations using co-dominantecotype-specificPCR-basedmarkers.Plant J.4:403–410.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish162
Langella, R., Ercolano, M.R., Monti, L., Frusciante, L. & Barone, A. 2004 Molecular markerassistedtransferofresistancetoTSWVintomatoelitelines.J. Hort. Sci. & Biotech.79:806–810
Laterrot, H. 1996.Breedingstrategiesfordiseaseresistanceintomatoeswithemphasistothetrop-ics:currentstatusandresearchchallenges,pp.126–132.Proc.1st Internat. Symp. Trop. Tomato Dis.Recife,Brazil.
Lefebvre, V. & Chèvre, A.M.1995.Toolsformarkingplantdiseaseandpestresistancegenes:areview.Agronomie15:3–19.
Leister, D., Ballvora, A., Salamini, F. & Gebhardt, C.1996.PCR-basedapproachforisolatingpath-ogenresistancegenesfrompotatowithpotentialforwideapplicationinplants.Nature Genet.14:421–429.
Levesque, H., Vedel, F., Mathieu, C. & de Courcel, A.G.L. 1990.IdentificationofashortrDNAspacer sequence highly specific of a tomato line containing Tm-1 gene introgressed fromLycopersicon hirsutum. Theor. Appl. Genet.80:602–608.
Martin, G.B., Brommonschenkel, S., Chunwongse, J., Frary, A., Ganal, M.W., Spivey, R., Wu, T., Earle, E.D. & Tanksley, S.D. 1993.Map-basedcloningofaproteinkinasegeneconferringdiseaseresistanceintomato.Science262:1432–1436.
Michelmore, R.W.2003.Theimpactzone:genomicsandbreedingfordurablediseaseresistance.Curr. Opin. Plant Biol.6397–6404.
Moreau, P., Thoquet, P., Olivier J., Laterrot, H. & Grimsley, N.H.1998.GeneticmappingofPh-2,asinglelocuscontrollingpartialresistancetoPhytophthorainfestansintomato.Mol. Plant-Microbe Interact.11:259–268.
Morgante, M. & Salamini, F.2003.Fromplantgenomicstobreedingpractice.Curr. Opin. Biotechnol.14:214–219.
Ori, N., Eshed, Y., Paran, I., Prestino, G., Aviv, D., Tanksley, S.D., Zamir, D. & Fluhr, R. 1997.TheI2CfamilyfromthewiltdiseaseresistancelocusI2belongstothenucleotidebinding,leucine-richrepeatsuperfamilyofplantresistancegenes.Plant Cell9:521–532.
Parrella, G., Ruffel, S., Moretti, A., Morel, C., Palloix, A. & Caranta, C.2002.Recessiveresistancegenes against potyviruses are localized in colinear genomic regions of the tomato (Lycopersiconspp.)andpepper(Capsicumspp.)genomes.Theor. Appl. Genet.105:855–861.
Parrella, G., Moretti, A., Gognalons, P., Lesage, M-L., Marchoux, G., Gebre-Selassie, K. & Carata, C. 2004.TheAmgenecontrollingresistancetoalfalfamosaicvirusintomatoislocatedintheclus-terofdominantresistancegenesonchromosome6.Phytopath.94:345–350.
Peleman, J.D. & van der Voort, J.R. 2003.Breedingbydesign.Trends Plant Sci.8:330–334.Pillen, K., Pineda, O., Lewis, C.B. & Tanksley, S.D.1996.Statusofgenomemappingtools in the
taxonSolanaceae.In A.Paterson,ed.Genome mapping in plants,pp.281–308.Austin,TX,USA,Landes.
Rao, A.V. & Agarwal, S.2000.Roleofantioxidantlycopeneincancerandheartdisease.J. Am. Coll. Nutr.19:563–569.
Robert, V.J.M., West, M.A.L., Inai, S., Caines, A., Arntzen, L., Smith, J.K. & St. Clair, D.A. 2001.Marker-assistedintrogressionofblackmoldresistanceQTLallelesfromwildLycopersicon chees-manii tocultivatedtomato(L. esculentum)andevaluationofQTLphenotypiceffects.Mol. Breed.8:217–233.
Salmeron, J.M., Oldroyd, G.E.D., Rommens, C.M.T., Scofield, S.R., Kim, H.S., Lavello, D.T., Dahlbeck, D. & Staskawicz, B.J. 1996.TomatoPrfisamemberoftheleucine-richrepeatclassof

Chapter 9 – Molecular marker-assisted selection for resistance to pathogens in tomato 163
plantdiseaseresistancegenesandliesembeddedwithinthetomatoPtokinasegenecluster.Cell86:123–133.
Sandbrink, J.M., van-Ooijen, J.W., Purimahua, C.C., Vrielink, M., Verkerk, R., Zabel, P. & Lindhout, P.1995.LocalizationofgenesforbacterialcankerresistanceinLycopersicon peruvianumusingRFLPs.Theor. Appl. Genet.90:444–450.
Sarfatti, M., Abu-Abied, M., Katan, J. & Zamir, D. 1991. RFLP mapping of I1, a new locus intomatoconferringresistancetoFusarium oxysporumf.sp. lycopersicirace1.Theor. Appl. Genet.80:22–26.
Scott, J.W.2005.Perspectivesontomatodiseaseresistancebreeding:past,present,andfuture.Acta Hort.695:217–224.
Sobir, Ohmori T., Murata, M. & Motoyoshi, F.2000.MolecularcharacterizationoftheSCARmark-erstightlylinkedtotheTm-2locusofthegenusLycopersicon.Theor. Appl. Genet. 101:64–69.
Stamova, B.S. & Chetelat, R.T.2000. InheritanceandgeneticmappingofcucumbermosaicvirusresistanceintrogressedfromLycopersicon chilenseintotomato.Theor. Appl. Genet. 101:527–537.
Stevens, M.R., Lamb, E.M. & Rhoads, D.D. 1995.MappingtheSw-5locusfortomatospottedwildvirusresistanceintomatoesusingRAPDandRFLPanalyses.Theor. Appl. Genet. 90:451–456.
Tanksley, S.D. & Costello, W. 1991.ThesizeofL. pennellii chromosome7segmentcontainingtheI-3geneintomatobreedinglinesmeasuredbyRFLPprobing.Rep. Tomato Genet. Coop.41:60.
Tanksley, S.D., Young, N.D., Paterson, A.H. & Bonierbale, M.W. 1989. RFLP mapping in plantbreeding:newtoolsforanoldscience.BioTech.7:257–264.
Thoquet, P., Olivier, J., Sperisen, C., Rogowsky, P., Laterrot, H. & Grimsley, N.H. 1996.QuantitativetraitlocideterminingresistancetobacterialwiltintomatocultivarHawaii7996.Mol. Plant-Microbe Interact.9:826–836.
Vakalounakis, D.J., Laterrot, H., Moretti, A., Ligoxigakis, E.K. & Smardas, K. 1997. LinkagebetweenFr-1(Fusarium oxysporumf.sp.radici-lycopersicyresistance)andTm-2(tobaccomosaicvirusresistance-2)lociintomato(Lycopersicon esculentum).Ann. Appl. Biol.130:319–323.
van der Biezen, E.A., Glagotlkaya, T. & Overduin, B. 1995. Inheritance andgeneticmappingofresistancetoAlternaria alternataf.sp.lycopersiciinLycopersicon pennellii.Mol. Gen. Genet.247:453–461.
van Heusden, A.W., Koornneef, M., Voorrips, R.E., Bruggemann, W., Pet, G., Vrielink-van Ginkel, R., Chen, X. & Lindhout, P.1999.ThreeQTLsfromLycopersicon peruvianumconferahighlevelofresistancetoClavibacter michiganensissubsp.michiganensis.Theor. Appl. Genet.99:1068–1074.
Varshney, R.K., Graner, A. & Sorrells, M.E. 2005.Genomics-assistedbreedingforcropimprove-ment.Trends Plant Sci.10:621–630.
Williamson, V.M., Ho, J-Y., Wu, F.F., Miller, N. & Kaloshian, I.1994.APCR-basedmarkertightlylinkedtothenematoderesistance,Mi,intomato.Theor. Appl. Genet.87:757–763.
Yaghoobi, J., Kaloshian, I., Wen, Y. & Willimson, V.M.1995.MappingofanewnematoderesistancelocusinLycopersiconperuvianum.Theor. Appl. Genet.91:457–464.
Yang, W., Bai, X., Kabelka, E., Eaton, C., Kamoun, S., van der Knapp, E. & Francis, D. 2004.DiscoveryofsinglenucleotidepolymorphismsinLycopersicon esculetum bycomputeraidedanaly-sisofexpressedsequencetags.Mol. Breed. 14:21–34.
Young, N.D. & Tanksley, S.D.1988.UseofisogeniclinesandsimultaneousprobingtoidentifyDNAmarkerstightlylinkedtotheTm-2geneintomato.Genetics120:579–589.

Marker-assisted selection – Current status and future perspectives in crops, livestock, forestry and fish164
Yu, Y.G., Wang, J.F., Stall, R.E. & Allejos, C.E.1995.Genomiclocalizationoftomatogenesthatcon-trolahypersensitivityreactiontoXanthomonas campestrispvvesicatoria(Doidge)dye.Genetics141:675–682.
Zamir, D., Ekstein-Michelson, I., Zakay, U., Navot, N., Zeidan, M., Sarfatti, M., Eshed, Y., Harel, E., Pleban, T., van-Oss, H., Kedar, N., Rabinowitch, H.D. & Czosnek, H.1994.Mappingandintrogression of a tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Genet. 88:141–146.
Zhang, L.P., Khan, A., Nino-Liu, D. & Foolad, M.R.2002.Amolecularlinkagemapoftomatodis-playingchromosomal locationsof resistancegeneanalogsbasedonaLycopersiconesculentumxLycopersicon hirsutumcross.Genome45:133–146.
Zhang, L.P., Lin, G.Y., Nino-Liu, D. & Foolad, M.R. 2003.MappingQTLsconferringearlyblight(Alternaria solani)resistanceinaLycopersicon esculentumxL. hirsutumcrossbyselectivegenotyp-ing.Mol. Breed.12:3–19.