macroinvertebrates associated with chara in a tropical coastal lagoon (imboassica lagoon, rio de...
TRANSCRIPT

Hydrobiologia 457: 215–224, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
215
Macroinvertebrates associated with Chara in a tropical coastallagoon (Imboassica lagoon, Rio de Janeiro, Brazil)
Edelti Faria Albertoni1, Cleber Palma-Silva1∗ & Francisco de Assis Esteves2
1Fundação Universidade Federal do Rio Grande, Departamento de Ciências Morfo-Biologicas,Laboratorio de Ecologia, C.P. 474, CEP 96201-900, Rio Grande, RS, BrazilE-email: [email protected](∗Author for correspondence)2Universidade Federal do Rio de Janeiro, Departamento de Ecologia, Laboratorio de Limnologia,Rio de Janeiro, RJ, Brazil
Received 2 May 2000; in revised form 21 May 2001; accepted 1 June 2001
Key words: coastal lagoon, Chara, macroinvertebrates, community, associated fauna, water level
Abstract
Imboassica lagoon is an urban coastal lagoon located in the municipality of Macaé (RJ), which has been exposedto a process of artificial eutrophication through the inflow of untreated sewage, as well as artificial openings ofthe sandbar that separates it from the ocean, provoking drastic modifications in this ecosystem. The sampling forthe analysis of the community of macroinvertebrates associated with Charopyceae (Chara angolensis e C. fibrosa)were performed between October 1995 and October 1997, with a total of 9 samplings at two sampling stations:one located in the innermost area of the lagoon and another at the mouth of the main sewage channel, close to thesandbar. Throughout this period, four artificial sandbar-opening events were recorded, resulting in changes in thebiomass of the macroalgae and in the densities of the macroinvertebrate populations. Through the analysis of thedensity of organisms per square meter, and density of organisms per 100 g dry weight of Chara, it was establishedthat the biomass of the macroalgae has a direct influence on the density of the associated macrofauna. The greaterdevelopment of the periphytic community in the area close to the inflow of sewage allows for the developmentof a community of macroinvertebrates with high densities, composed mainly of scraping organisms, such as themollusc Heleobia australis (Hidrobiidae).
Introduction
Submersed macrophytes are found in a great variety ofhabitats, exhibiting variations in the composition andabundance of the associated macroinvertebrate com-munity according to the type of macrophyte (Hargeby,1990). These variations have been explained by differ-ences in the plant morphology, influencing the archi-tecture of the habitat and, as a consequence, the degreeof shelter and food availability.
Charophyceae are macroalgae often found in lakesand lagoons, usually achieving high values of biomass(Kairesalo et al., 1992). Due to their morphology, theyhave an extensive surface area, enabling the colon-ization of a great periphytic community, which is adietary resource for a rich community of invertebrates
and their predators (Glowacka et al., 1976; Kairesaloet al., op.cit.).
Studies focusing on the structure and function ofcommunities of macroinvertebrates associated withthese macroalgae were developed, mainly in temperateenvironments (Dvorak & Best, 1982; Hargeby, 1990;Mbahinzireki et al., 1991; Roca et al., 1993, amongothers), and are scarce in tropical systems.
Palma-Silva (1999) found that the Charophyceaecommunity is very developed in the Imboassica la-goon, and has great importance as a nutrient pool.This author shows that Chara spp. have high absortionrate of carbon, nitrogen and phosphorus, and help tomaintain the nutrient concentrations in the water at lowlevels, preventing excess phytoplanktonic growth.
This research aimed to identify and quantify the

216
groups of organisms associated with 2 species ofChara (C. angolensis and C. fibrosa) at Imboassicalagoon, focusing on the macroinvertebrate community,and comparing two sampling stations with differenttrophic and limnological characteristics.
Characterization of the study area
Imboassica is a coastal lagoon located at the muni-cipality of Macaé (22◦ 30′ S and 42◦ 00′ W), Riode Janeiro State, Brazil (Fig. 1). The regional cli-mate is subhumid, with a maximum mean temperatureof 29.9 ◦C in February (summer) and a minimummean temperature of 25.4 ◦C in July (winter). An-nual precipitation varies between 800 mm and 120 mm(RADAMBRASIL, 1983 ). It is a shallow ecosystem,with an area 3.26 m2 and a medium depth of 1.1 m(Panosso et al., 1998) and is separated from the seaby a sand barrier about 50 m wide. It is located in theurban zone and subject to several anthropic impacts,notably the dumping of ‘in natura’ domestic sewagethrough channels ending at the lagoon.
The surroundings are partially occupied by resid-ential areas and, sometimes, during the rainy season,inundation of these areas occurs (Branco et al., 1997).An artifical opening (by human action) of the sandbarrier then causes drastic drop in the water level,and sea water entrance. This artificial opening of thesandbar expose almost the entire lagoon’s bed, andresults in the death of the Chara stand, which redevel-ops after the lagoon recover its former water level(Palma-Silva, 1999). The water column recompositiondepends mainly on the pluviometric precipitation, andthe Chara beds redevelop in different speeds at differ-ent times of year. These modifications involve changein the biotic communities (Branco, 1998; Albertoni,1999; Albertoni et al., 1999; Palma-Silva et al., 2000).
Materials and methods
The samplings took place between October/1995, andOctober/1997, and were taken in Oct 95, Feb 96, Apr96, Nov 96, Mar 97, Apr 97, May 97, Jul 97 and Oct97. The periodicity of the samplings was determinedby the presence of Chara in the lagoon, which is underthe direct influence of the artificial breachings and therecomposition of the water level, what occurs in differ-ent speeds at different times of year. In the sampling
period, 4 breachings were registered (Nov 95; Apr 96;Nov 96; and Jan 97).
Two sampling stations were established: Station 1– located in the interior part of the lagoon (far fromthe ocean), and Station 2 – located closer to the oceanand close to the mouth of one of the main channels ofsewage dumping (Fig. 1). Samplings quadrats of 25× 25 cm, with an attached net (1 mm mesh), wereused. The species C. angolensis and C. fibrosa occurin mixed stands, and their morphology did not presentslarge differences, then macroinvertebrate communitywas analyzed jointly for both species.
The sampling sites were chosen randomly insidethe stand of Chara. Three replicates for associatedfauna and three different replicates for biomass estim-ating were sampled in each station, cutting all Charaat the sediment level. After sampling, the plants weretaken to the laboratory, fixed with 4% formaldehyde,and the macrofauna was separated with a stereoscopicmicroscope. For estimating the biomass of Chara,the plants were dried in an oven at 60 ◦C for themeasurement of dry weight.
The organisms were separated and identified to thelowest taxonomic level possible, according to Pennak(1978), Lehmkuhl (1979), and taxonomic specialists.For statistical analyses, categorization in to highertaxons (classes, orders and families) were used formost organisms, apart from the gastropod, Heleobiaaustralis.
The results are shown as density of organisms per100 g of dry weight of Chara. Mann–Whitney testswere used to compare densities between sampling sta-tions, considering all values, and regressions betweenChara biomass and macroinvertebrate density persquare meter were performed.
Several abiotic variables – salinity, measured witha salinometer-refractometer Meiji Techno RF-235;temperature, measured with a digital thermistor FAC-400; and depth of the water column, measured with amarked ruler – were collected on each sampling date.
Results
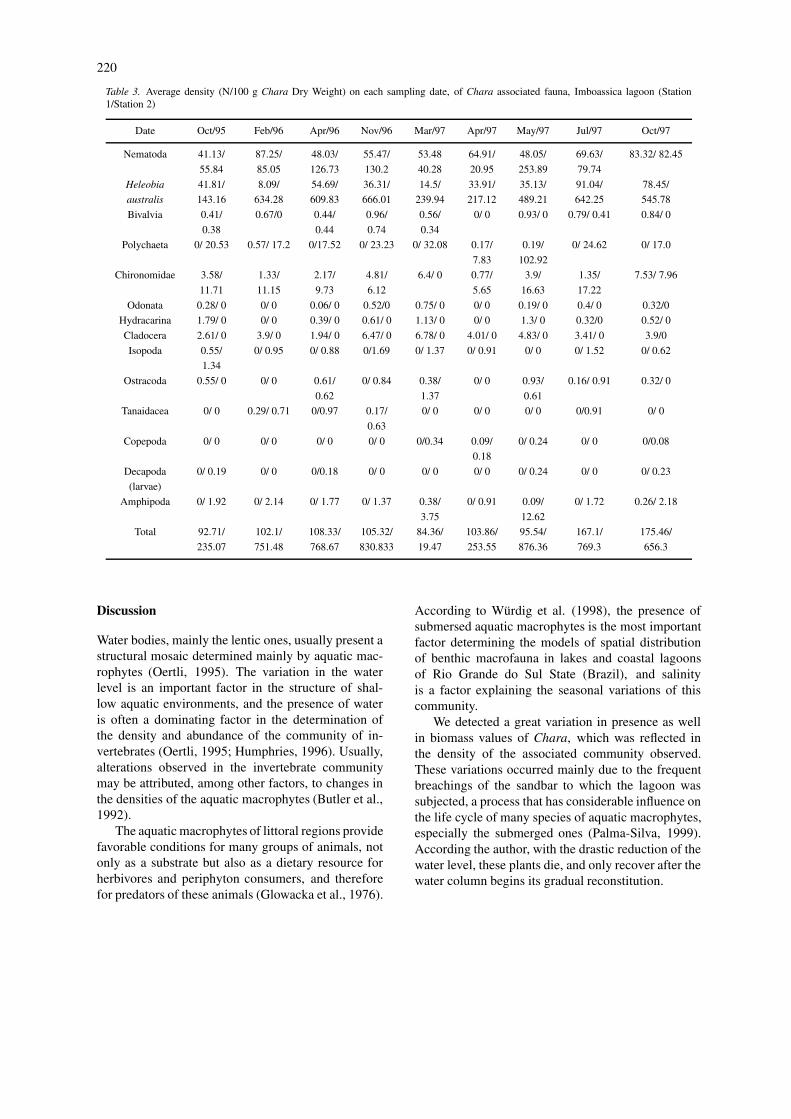
The taxa associated with Chara at Imboassica lagoonare shown in Table 1. The artificial breachings of thesandbar influence the abiotic characteristics of Im-boassica lagoon, and Table 2 shows this alterations,in a two year period. The nitrogen and phosphoruscontent show the nutrient levels at the two samplingsites, make evident the eutrophic characteristics close
217
Figure 1. Study area, with the location of the two sampling stations.
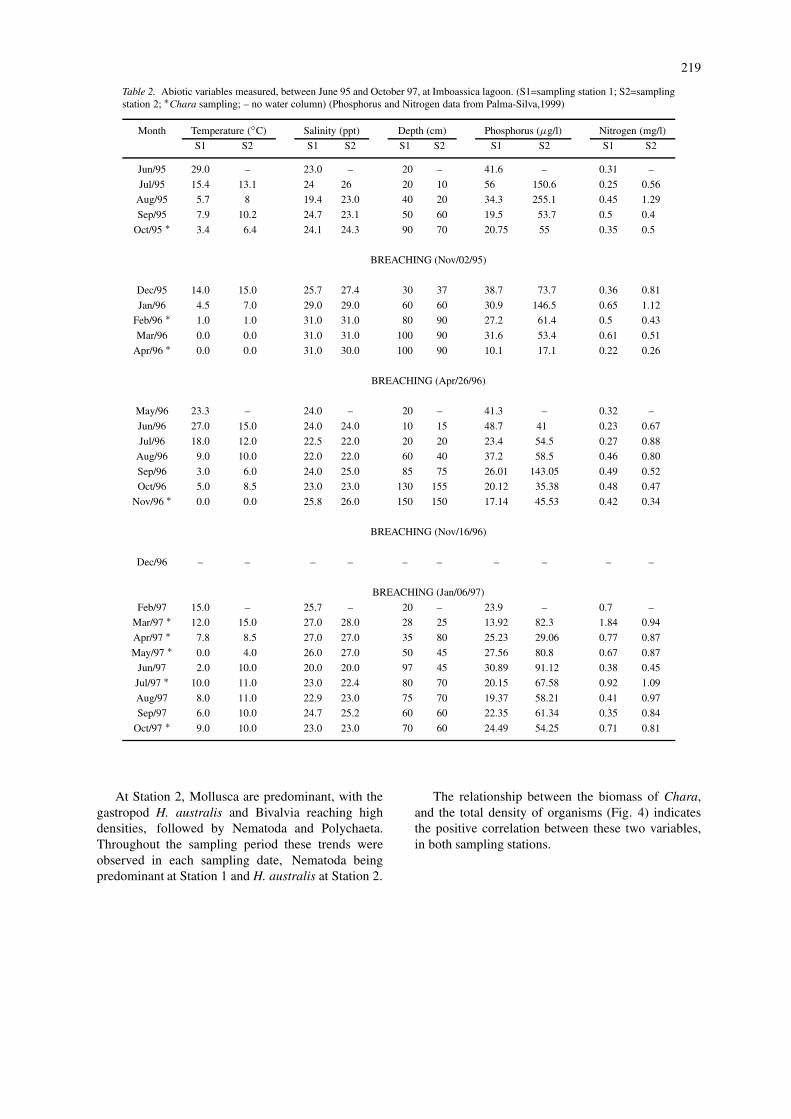
Figure 2. Biomass variation (g dry weight) of Chara at sampling station 1 (S1) and sampling Station 2 (S2), at Imboassica lagoon (data fromPalma-Silva, 1999). (arrows show sandbar openings).
to the Station 2. The variation in Chara biomass isshow in Figure 2, and the average values of organismsdensity are shown in Table 3.
The total number of invertebrates found in eachsampling station was compared and we found thatStation 2 can support significantly higher densit-ies of fauna associated with Chara (Mann–Whitney,p<0.001) (Fig. 3). Regarding the different animal
groups, most of them exhibited different densities ineach station (Mann–Whitney, p<0.01) (Table 4).
At Station 1, Nematoda are predominant, fol-lowed by the gastropod Heleobia australis, and severalother groups of organisms, notably insects (especiallyChironomidae and Odonata) and Cladocera (Ilyocryp-tus sp.)

218
Table 1. Taxa associated with Chara spp., between October 95 and October 97, Imboassicalagoon. (1=collected at sampling station 1; 2=collected at sampling station 2)
ALGAE:
Cyanophyceae => Microcystis sp., Oscillatoria sp. and Ankistrodesmus sp. (2)
Bacillariophyceae => sessile and filamentous (Melosira sp.) (2)
colonial Chlorophyceae (2)
Zygnemaphyceae => Spirogira sp. (2)
POLYCHAETA: Nereididae
Laeonereis cuvieri (2)
Polydora sp. (1; 2)
Paraprionospis pinnata (2)
MOLLUSCA:
Gastropoda => Hidrobiidae => Heleobia australis (1; 2)
Bivalvia (1; 2)
INSECTA:
Diptera: Chironomidae (1; 2)
Odonata (1)
CRUSTACEA:
Ostracoda: Darwinulidae and Cyprididae; (1; 2)
Amphipoda: Tallitridae => Hyalella azteca and Gammarus sp. (1; 2)
Isopoda: Sphaeromatidae => Dies fluminensis (1; 2)
Copepoda: Calanoida (2), Harpacticoida (1; 2) and Cyclopoida => Halicyclops sp. (2)
Tanaidacea (1; 2)
Cladocera => Ilyocryptus sp. (1)
Decapoda => Palaemonidae => Macrobrachium sp. (larvae) (2)
ARACHNIDAE:
Hydracarina (1)
Figure 3. Average density of associated fauna (all taxa) with Chara organisms per 100 g dry weight of Chara, Imboassica lagoon (arrows showsandbar openings).
219
Table 2. Abiotic variables measured, between June 95 and October 97, at Imboassica lagoon. (S1=sampling station 1; S2=samplingstation 2; ∗Chara sampling; – no water column) (Phosphorus and Nitrogen data from Palma-Silva,1999)
Month Temperature (◦C) Salinity (ppt) Depth (cm) Phosphorus (µg/l) Nitrogen (mg/l)
S1 S2 S1 S2 S1 S2 S1 S2 S1 S2
Jun/95 29.0 – 23.0 – 20 – 41.6 – 0.31 –
Jul/95 15.4 13.1 24 26 20 10 56 150.6 0.25 0.56
Aug/95 5.7 8 19.4 23.0 40 20 34.3 255.1 0.45 1.29
Sep/95 7.9 10.2 24.7 23.1 50 60 19.5 53.7 0.5 0.4
Oct/95 ∗ 3.4 6.4 24.1 24.3 90 70 20.75 55 0.35 0.5
BREACHING (Nov/02/95)
Dec/95 14.0 15.0 25.7 27.4 30 37 38.7 73.7 0.36 0.81
Jan/96 4.5 7.0 29.0 29.0 60 60 30.9 146.5 0.65 1.12
Feb/96 ∗ 1.0 1.0 31.0 31.0 80 90 27.2 61.4 0.5 0.43
Mar/96 0.0 0.0 31.0 31.0 100 90 31.6 53.4 0.61 0.51
Apr/96 ∗ 0.0 0.0 31.0 30.0 100 90 10.1 17.1 0.22 0.26
BREACHING (Apr/26/96)
May/96 23.3 – 24.0 – 20 – 41.3 – 0.32 –
Jun/96 27.0 15.0 24.0 24.0 10 15 48.7 41 0.23 0.67
Jul/96 18.0 12.0 22.5 22.0 20 20 23.4 54.5 0.27 0.88
Aug/96 9.0 10.0 22.0 22.0 60 40 37.2 58.5 0.46 0.80
Sep/96 3.0 6.0 24.0 25.0 85 75 26.01 143.05 0.49 0.52
Oct/96 5.0 8.5 23.0 23.0 130 155 20.12 35.38 0.48 0.47
Nov/96 ∗ 0.0 0.0 25.8 26.0 150 150 17.14 45.53 0.42 0.34
BREACHING (Nov/16/96)
Dec/96 – – – – – – – – – –
BREACHING (Jan/06/97)
Feb/97 15.0 – 25.7 – 20 – 23.9 – 0.7 –
Mar/97 ∗ 12.0 15.0 27.0 28.0 28 25 13.92 82.3 1.84 0.94
Apr/97 ∗ 7.8 8.5 27.0 27.0 35 80 25.23 29.06 0.77 0.87
May/97 ∗ 0.0 4.0 26.0 27.0 50 45 27.56 80.8 0.67 0.87
Jun/97 2.0 10.0 20.0 20.0 97 45 30.89 91.12 0.38 0.45
Jul/97 ∗ 10.0 11.0 23.0 22.4 80 70 20.15 67.58 0.92 1.09
Aug/97 8.0 11.0 22.9 23.0 75 70 19.37 58.21 0.41 0.97
Sep/97 6.0 10.0 24.7 25.2 60 60 22.35 61.34 0.35 0.84
Oct/97 ∗ 9.0 10.0 23.0 23.0 70 60 24.49 54.25 0.71 0.81
At Station 2, Mollusca are predominant, with thegastropod H. australis and Bivalvia reaching highdensities, followed by Nematoda and Polychaeta.Throughout the sampling period these trends wereobserved in each sampling date, Nematoda beingpredominant at Station 1 and H. australis at Station 2.
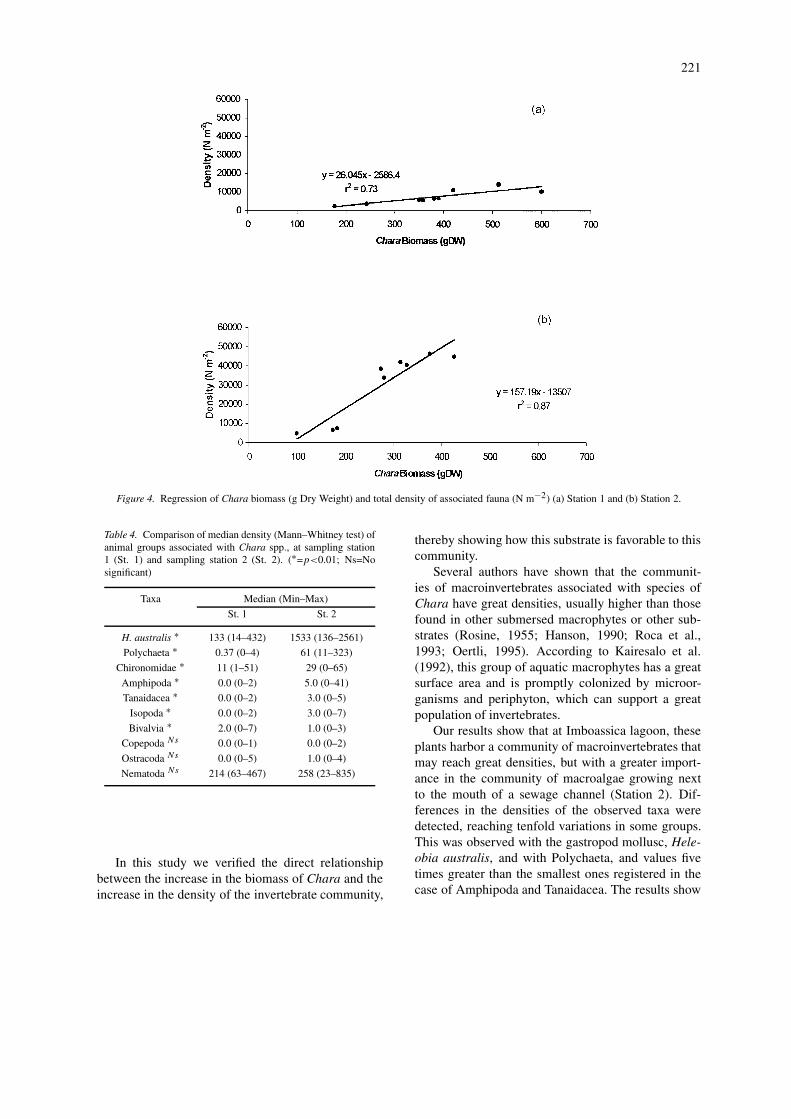
The relationship between the biomass of Chara,and the total density of organisms (Fig. 4) indicatesthe positive correlation between these two variables,in both sampling stations.
220
Table 3. Average density (N/100 g Chara Dry Weight) on each sampling date, of Chara associated fauna, Imboassica lagoon (Station1/Station 2)
Date Oct/95 Feb/96 Apr/96 Nov/96 Mar/97 Apr/97 May/97 Jul/97 Oct/97
Nematoda 41.13/ 87.25/ 48.03/ 55.47/ 53.48 64.91/ 48.05/ 69.63/ 83.32/ 82.45
55.84 85.05 126.73 130.2 40.28 20.95 253.89 79.74
Heleobia 41.81/ 8.09/ 54.69/ 36.31/ 14.5/ 33.91/ 35.13/ 91.04/ 78.45/
australis 143.16 634.28 609.83 666.01 239.94 217.12 489.21 642.25 545.78
Bivalvia 0.41/ 0.67/0 0.44/ 0.96/ 0.56/ 0/ 0 0.93/ 0 0.79/ 0.41 0.84/ 0
0.38 0.44 0.74 0.34
Polychaeta 0/ 20.53 0.57/ 17.2 0/17.52 0/ 23.23 0/ 32.08 0.17/ 0.19/ 0/ 24.62 0/ 17.0
7.83 102.92
Chironomidae 3.58/ 1.33/ 2.17/ 4.81/ 6.4/ 0 0.77/ 3.9/ 1.35/ 7.53/ 7.96
11.71 11.15 9.73 6.12 5.65 16.63 17.22
Odonata 0.28/ 0 0/ 0 0.06/ 0 0.52/0 0.75/ 0 0/ 0 0.19/ 0 0.4/ 0 0.32/0
Hydracarina 1.79/ 0 0/ 0 0.39/ 0 0.61/ 0 1.13/ 0 0/ 0 1.3/ 0 0.32/0 0.52/ 0
Cladocera 2.61/ 0 3.9/ 0 1.94/ 0 6.47/ 0 6.78/ 0 4.01/ 0 4.83/ 0 3.41/ 0 3.9/0
Isopoda 0.55/ 0/ 0.95 0/ 0.88 0/1.69 0/ 1.37 0/ 0.91 0/ 0 0/ 1.52 0/ 0.62
1.34
Ostracoda 0.55/ 0 0/ 0 0.61/ 0/ 0.84 0.38/ 0/ 0 0.93/ 0.16/ 0.91 0.32/ 0
0.62 1.37 0.61
Tanaidacea 0/ 0 0.29/ 0.71 0/0.97 0.17/ 0/ 0 0/ 0 0/ 0 0/0.91 0/ 0
0.63
Copepoda 0/ 0 0/ 0 0/ 0 0/ 0 0/0.34 0.09/ 0/ 0.24 0/ 0 0/0.08
0.18
Decapoda 0/ 0.19 0/ 0 0/0.18 0/ 0 0/ 0 0/ 0 0/ 0.24 0/ 0 0/ 0.23
(larvae)
Amphipoda 0/ 1.92 0/ 2.14 0/ 1.77 0/ 1.37 0.38/ 0/ 0.91 0.09/ 0/ 1.72 0.26/ 2.18
3.75 12.62
Total 92.71/ 102.1/ 108.33/ 105.32/ 84.36/ 103.86/ 95.54/ 167.1/ 175.46/
235.07 751.48 768.67 830.833 19.47 253.55 876.36 769.3 656.3
Discussion
Water bodies, mainly the lentic ones, usually present astructural mosaic determined mainly by aquatic mac-rophytes (Oertli, 1995). The variation in the waterlevel is an important factor in the structure of shal-low aquatic environments, and the presence of wateris often a dominating factor in the determination ofthe density and abundance of the community of in-vertebrates (Oertli, 1995; Humphries, 1996). Usually,alterations observed in the invertebrate communitymay be attributed, among other factors, to changes inthe densities of the aquatic macrophytes (Butler et al.,1992).
The aquatic macrophytes of littoral regions providefavorable conditions for many groups of animals, notonly as a substrate but also as a dietary resource forherbivores and periphyton consumers, and thereforefor predators of these animals (Glowacka et al., 1976).
According to Würdig et al. (1998), the presence ofsubmersed aquatic macrophytes is the most importantfactor determining the models of spatial distributionof benthic macrofauna in lakes and coastal lagoonsof Rio Grande do Sul State (Brazil), and salinityis a factor explaining the seasonal variations of thiscommunity.
We detected a great variation in presence as wellin biomass values of Chara, which was reflected inthe density of the associated community observed.These variations occurred mainly due to the frequentbreachings of the sandbar to which the lagoon wassubjected, a process that has considerable influence onthe life cycle of many species of aquatic macrophytes,especially the submerged ones (Palma-Silva, 1999).According the author, with the drastic reduction of thewater level, these plants die, and only recover after thewater column begins its gradual reconstitution.
221
Figure 4. Regression of Chara biomass (g Dry Weight) and total density of associated fauna (N m−2) (a) Station 1 and (b) Station 2.
Table 4. Comparison of median density (Mann–Whitney test) ofanimal groups associated with Chara spp., at sampling station1 (St. 1) and sampling station 2 (St. 2). (∗=p<0.01; Ns=Nosignificant)
Taxa Median (Min–Max)
St. 1 St. 2
H. australis ∗ 133 (14–432) 1533 (136–2561)
Polychaeta ∗ 0.37 (0–4) 61 (11–323)
Chironomidae ∗ 11 (1–51) 29 (0–65)
Amphipoda ∗ 0.0 (0–2) 5.0 (0–41)
Tanaidacea ∗ 0.0 (0–2) 3.0 (0–5)
Isopoda ∗ 0.0 (0–2) 3.0 (0–7)
Bivalvia ∗ 2.0 (0–7) 1.0 (0–3)
Copepoda Ns 0.0 (0–1) 0.0 (0–2)
Ostracoda Ns 0.0 (0–5) 1.0 (0–4)
Nematoda Ns 214 (63–467) 258 (23–835)
In this study we verified the direct relationshipbetween the increase in the biomass of Chara and theincrease in the density of the invertebrate community,
thereby showing how this substrate is favorable to thiscommunity.
Several authors have shown that the communit-ies of macroinvertebrates associated with species ofChara have great densities, usually higher than thosefound in other submersed macrophytes or other sub-strates (Rosine, 1955; Hanson, 1990; Roca et al.,1993; Oertli, 1995). According to Kairesalo et al.(1992), this group of aquatic macrophytes has a greatsurface area and is promptly colonized by microor-ganisms and periphyton, which can support a greatpopulation of invertebrates.
Our results show that at Imboassica lagoon, theseplants harbor a community of macroinvertebrates thatmay reach great densities, but with a greater import-ance in the community of macroalgae growing nextto the mouth of a sewage channel (Station 2). Dif-ferences in the densities of the observed taxa weredetected, reaching tenfold variations in some groups.This was observed with the gastropod mollusc, Hele-obia australis, and with Polychaeta, and values fivetimes greater than the smallest ones registered in thecase of Amphipoda and Tanaidacea. The results show

222
that the conditions of Station 2 are more amenablefor the great majority of animal groups, the exceptionbeing Bivalvia. This is an eutrophic region, allowingfor the growth of a more dense periphytic communityin comparison to Station 1, with the presence of co-lonial Cyanophyceae (Microcystis sp.), sessile andfilamentous diatoms (Melosira sp.), Chlorophyceaeand Zygnemaphyceae (Spirogyra sp.).
According to Higler (1981), Gastropoda and Bi-valvia are the main constituents of the community inChara. In this study, Heleobia australis showed thehighest values of density among the studied groups.Although abundant in Station 1, it reached its highestdensities at Station 2. In a region close to the Station2 of this study, Fernandes (1998) observed that thefilamentous Cyanophyceae occur in great abundancein some months of the year in the periphyton of Typhadomingensis. We suppose that this also happens in theperiphyton of Chara, which would favor the devel-opment of communities of the associated fauna withscraping and detritivore characteristics. Gonçalves etal. (1998a, b) found the greatest biomass of H. aus-tralis in the Imboassica lagoon close to the Station 2 ofthe present study, and attributed this to the greater foodavailability provided by the periphyton of T. domin-gensis and of other aquatic macrophytes where thisgastropod feeds. The favorable environment close tothe sewage channel is probably maintained by thecommunity of emergent aquatic macrophytes of thesite, which prevent the entrance of the total organicpollution (Lopes-Ferreira, 1998a, b), and so reducingthe negative effects on the structure of the community.
According to Callisto et al. (1998), H. australis isfavored by the presence of marine water in Imboas-sica lagoon, since it has eurihaline characteristics, asdoes Polychaeta. Both are more abundant in Station2, which has a more pronounced marine influencethan Station 1. Station 2 also had the highest dens-ities of Amphipoda, Tanaidacea and Isopoda, groupsusually found in coastal environments. Isopoda of theSphaeromatidae family are common in coastal aquaticenvironments of Brazil (Loyola & Silva, 1960). Wür-dig & Dornelles da Silva (1990) registered the pres-ence of Dies fluminensis in mesohaline coastal lagoonsof the Tramandaí lake system (RS, Brazil), found thisspecies in 80% of the samples of Chara sp. classifyingit as characteristically mesohaline.
The groups that did not exhibit statistically sig-nificant differences between the two stations wereNematoda, Copepoda and Ostracoda, found in similardensities at both.
Ostracods are common organisms in fresh wa-ter and mesohaline environments, and Würdig et al.(1990) found that the family Cyprididae is dominantin aquatic macrophytes, among them Chara sp. Ac-cording to Albertoni & Würdig (1996), the higherabundance and diversity of ostracods is related to thesubmersed aquatic macrophytes at a coastal lagoon inthe south of Brazil (Lagoa do Gentil – RS), and Dar-winulidae and Cyprididae are the dominant families,the same ones found at Imboassica lagoon.
Other organisms, such as the Cladocera Ilyocryp-tus sp., were only registered at Station 1, and areorganisms characteristically associated to submersedaquatic macrophytes and fresh water environments(Pennak, 1978). Hydracarina were also registered onlyin this station, in low densities.
Some aquatic insects such as nymphs of Odonatawere also observed only at this sampling station. Thelarvae of Chironomidae, on the other hand, had higherdensities at Station 2, even though they were con-stantly observed at Station 1. Chironomidae is, char-acteristically, a group constantly found in water bodieswith diverse characteristics, especially regarding theenvironmental variations.
According to Callisto et al. (1996), the higherabundance of larvae of Chironomidae associated tothe periphyton of Typha domingensis in Imboassicalagoon is related to the influence of the sewage, whichraises the nutrient concentrations close to the Station2. In an analysis at a specific level of this group somedifferences through the distribution on the salinity andnutrients gradient of the lagoon would probably befound.
Bivalvia were found in both sampling stations, butwith higher densities at Station 1. Larvae of Macro-brachium sp. had low densities, and therefore theseorganisms were considered as infrequent in aquaticmacrophytes, due to the planktonic characteristics ofits first larval stages. Copepoda were also registered atlow densities and, the exception for Harpacticoida, theothers had planktonic habit.
It was observed that the lagoon level of artificialeutrophication, at this moment, through the dump-ing of domestic sewage, stimulates the growth ofperiphytic microorganisms and algae, which providefood to the invertebrates, therefore explaining thehigher density found in the sampling station closest tothe mouth of the channel that dumps the sewage intothe lagoon.
The differential establishment of some groups re-lated to the different abiotic characteristics of the

223
sampling station shows that the salinity would prob-ably act as a factor in the establishment of the com-munities of invertebrates associated to Chara, with acommunity exhibiting fresh water characteristics farfrom the sea, at Station 1, and another with mesohalinecharacteristics in the area closest to the sandbar.
Conclusions
The processes of artificial breaching of the sand-bar have a direct influence on the biomass values ofChara, and so on the occurrence and density of thecommunity of macroinvertebrates associated with it.The habitat provided by Chara. is favorable to the de-velopment of several animal groups, especially Hele-obia australis (Mollusca), Polychaeta, Chironomidaeand Nematoda. The artificial enrichment caused by thedumping of sewage favors, up to this moment, thedevelopment of the periphytic community of Charain the area close to the mouth of the sewage channel,which provides food for several associated organisms,explaining the higher density of the macrofauna thatwas observed in this area. The entrance of sea waterwhen the sandbar is breaching may promote differ-ences in the groups that characterize the associatedfauna, with the separation of communities with meso-haline characteristics in the area closer to the sand-bar and fresh water community in the more interiorportion of the lagoon.
Acknowledgements
We thank PETROBRAS, CAPES and CNPq for thefinancial support; the several specialists for the iden-tification of: Dr Norma C. Bueno – Charophyceae;Drs Paulo Paiva and Christine Ruta – Polychaeta;Dr Paulina Maia Barbosa – Cladocera; MsC Bet-ina Koslowsky-Suzuki – Copepoda; Dr Sérgio deMelo – phytoplanktonic and periphytic algae, and theanonimous referee for the suggestions.
References
Albertoni, E. F. & N. L. Würdig, 1996. Comunidade de ostracodesassociada à macrófitas aquáticas na Lagoa do Gentil, Tramandaí,RS. Acta Limnol. Brasil 8: 103–114.
Albertoni, E. F., C. Palma-Silva & F. A. Esteves, 1999. Larvae andpost-larvae of Penaeidae and Palaemonidae in coastal lagoons ofthe north of Rio de Janeiro (Macaé, RJ). Rev. Brasil. Biol. 59:109–117.
Albertoni, E. F., 1999. Aspectos de Crescimento, Reprodução e Ali-mentação de Peneídeos e Paleomonídeos em Lagoas Costeiras doNorte do Estado do Rio de Janeiro. Tese, Doutorado, Programade Pós-Graduação em Ecologia, UFRJ. Rio de Janeiro. 200 pp.
Branco, C. W. C., 1998. Composição e aspectos ecológicos dascomunidades zooplanctônicas nas lagoas Imboassica, Cabiúnas eComprida. In Esteves, F. A. (ed.), Ecologia das Lagoas Costeirasdo Parque Nacional da Restinga de Jurubatiba e do Município deMacaé (RJ). NUPEM/UFRJ. Rio de Janeiro. 464 pp. Cap. 4.7:pp. 247–271.
Branco, C. W. C., T. Aguiaro, F. A. Esteves & E. P. Caramaschi,1997. Food sources of the teleost Eucinostomus argenteus in twocostal lagoons of Brazil. Stud. Neotrop. Fauna Environm. 32: 33–40.
Butler, R. S., E. J. Moyer, M. W. Hulon & V. P. Williams, 1992.Littoral zone invertebrate communities as affected by a habitatrestoration project on lake Tohopekaliga, Florida. J. Freshwat.Ecol. 7: 317–328.
Callisto, M., A. Serpa-Filho, S. J. Oliveira & F. A. Esteves,1996. Chironomid on leaves of the aquatic macrophyte Typhadomingensis in a coastal lagoon (Brazil). Stud. Neotrop. FaunaEnviron. 31: 50–52.
Callisto, M., J. F. Jr. Gonçalves, J. J. F. Leal & M. M. Petrucio,1998. Macroinvertebrados bentônicos nas lagoas Imboassica,Cabiúnas e Comprida. In Esteves, F. A. (ed.), Ecologia das La-goas Costeiras do Parque Nacional da Restinga de Jurubatiba edo Município de Macaé (RJ). NUPEM/UFRJ. Rio de Janeiro.464 pp. Cap. 4.9: pp. 283–298.
Dvorak, J. & E. P. H. Best, 1982. Macro-invertebrate communitiesassociated with the macrophytes of Lake Vechten: structural andfunctional relationships. Hydrobiologia 95: 115–126.
Fernandes, V. O., 1998. Variação temporal e espacial na composiçãoda comunidade perifítica na lagoa Imboassica. In Esteves, F.A. (ed.), Ecologia das Lagoas Costeiras do Parque Nacionalda Restinga de Jurubatiba e do Município de Macaé (RJ).NUPEM/UFRJ. Rio de Janeiro. 464 pp. Cap. 4.5: pp. 221–236.
Glowacka, I., G. J. Soszka & H. Soszka, 1976. Invertebratesassociated with macrophytes. In Pieczynska, E. (ed.), Selec-ted Problems of Lake Littoral Ecology. University of Warsaw,Warsaw: Chap. 6, pp. 97–122.
Gonçalves, J. F. Jr., J. J. L. Fonseca & M. Callisto, 1998a. Popu-lation dynamic of Heleobia australis (Gastropoda) in a coastallagoon (Rio de Janeiro, Brazil). Verh. int. Ver. Limnol. 26:2056–2057.
Gonçalves, J. F. Jr., M. Callisto & J. J. F. Leal, 1998b. Relaçãoentre a composição granulométrica do sedimento e as comunid-ades de macroinvertebrados bentônicos nas lagoas Imboassica,Cabiúnas e Comprida. In Esteves, F. A. (ed.), Ecologia das La-goas Costeiras do Parque Nacional da Restinga de Jurubatiba edo Município de Macaé (RJ). NUPEM/UFRJ. Rio de Janeiro.464 pp. Cap. 4.10. pp. 299–310.
Hanson, J. M., 1990. Macroinvertebrate size-distributions of twocontrasting freshwater macrophyte communities. Freshwat. Biol.24: 481–491.
Hargeby, A., 1990. Macrophyte associated invertebrates and theeffect of habitat permanence. Oikos 57: 338-346.
Higler, L. W. G., 1981. Bottom fauna and littoral vegetation faunain lake Maarsseveen. Hydrobiol. Bull. 15: 82–87.
Humphries, P., 1996. Aquatic macrophytes, macroinvertebrateassociations and water levels in a lowland Tasmanian river.Hydrobiologia 321: 219–233.
Kairesalo, T., G. Jónsson, K. Gunnarsson, C. Lindegaard & P. M.Jónasson, 1992. Metabolism and community dynamics within

224
Nitella opaca (Charophyceae) beds in Thingvallavatn. Oikos 64:241–256.
Lemhkuhl, D. M., 1979. How to Know the Aquatic Insects. ThePicture Key Nature Series, Wm C. Brown Company Publishers,Dubuque: 168 pp.
Lopes-Ferreira, C., 1998a. Redução das concentrações de nitrogênioe fósforo dos efluentes domésticos lançados na lagoa Imboassica,através de uma região colonizada por macrófitas aquáticas. InEsteves, F. A. (ed.), Ecologia das Lagoas Costeiras do ParqueNacional da Restinga de Jurubatiba e do Município de Macaé(RJ). NUPEM/UFRJ. Rio de Janeiro. 464 pp. Cap. 6.1: pp. 375–389.
Lopes-Ferreira, C., 1998b. A importância da região colonizada pormacrófitas aquáticas na mitigação da degradação sanitária da la-goa Imboassica. In Esteves, F. A. (ed.), Ecologia das LagoasCosteiras do Parque Nacional da Restinga de Jurubatiba e doMunicípio de Macaé (RJ). NUPEM/UFRJ. Rio de Janeiro. 464pp. Cap. 6.2: pp. 391–399.
Loyola e Silva, J., 1960. Sphaeromatidae do litoral brasileiro. B.Univ. Paraná, Zool. 4: 1–182.
Mbahinzireki, G., F. Uiblein & H. Winkler, 1991. Microhab-itat selection of ostracods in relation to predation and food.Hydrobiologia 222: 115–119.
Oertli, B., 1995. Spatial and temporal distribution of the zoobenthoscommunity in a woodland pond (Switzerland). Hydrobiologia300/301: 195–204.
Palma-Silva, C., 1999. Ecologia de Macrófitas Aquáticas em umaLagoa Costeira Sujeita a Impactos Antrópicos ( Lagoa Im-boassica, Macaé, RJ). Tese de Doutorado. Programa de Pós-Graduação em Ecologia. UFRJ: 153 pp.
Palma-Silva, C., E. F. Albertoni & F. A. Esteves, 2000. Eleocharismutata (L.) Roem. Et Schult. subject to drawdowns in a tropicalcoastal lagoon, State of Rio de Janeiro, Brazil. Plant Ecol. 148:157–164.
Panosso, R. F., J. L. Attayde & D. Muehe, 1998. Morfometria das la-
goas Imboassica, Cabiúnas, Comprida e Carapebus: Implicaçõespara seu funcionamento e manejo. In Esteves, F. A. (ed.), Eco-logia das Lagoas Costeiras do Parque Nacional da Restinga deJurubatiba e do Município de Macaé (RJ). NUPEM/UFRJ. Riode Janeiro. 464 pp. Cap 3.1: pp. 91–108.
Pennak, R. W., 1978. Freshwater Invertebrates of the United States.2nd edn. Willey Interscience publications, New York: 803 pp.
Porter, K. G., 1977. The plant-animal interface in freshwaterecosystems. Am. Sci. 65: 159–170.
Radambrasil, 1983. Levantamento de Recursos Naturais. FolhasSF.23/24, Rio de Janeiro/Vitória: geologia, geomorfologia, pe-dologia, vegetação e uso potencial da terra. Ministério de Minase Energia, Rio de Janeiro. 775 pp.
Roca, J. R., A. Baltanas & F. Uiblein, 1993. Adaptative responsesin Cypridopsis vidua (Crustacea: Ostracoda) to food and shelteroffered by a macrophyte (Chara fragilis). Hydrobiologia 262:127–131.
Rosine, W. N., 1955. The distribution of invertebrates on submergedaquatic plant surfaces in Muskee lake, Colorado. Ecology 36:308–314.
Würdig, N. L. & C. M. Dornelles da Silva, 1990. Distribuiçãoe abundância da comunidade de Isopoda na Lagoa do Gen-til, sistema lagunar de Tramandaí, RS. Acta Limnol. Brasil. 3:785–806.
Würdig, N. L., S. F. Freitas & I. da V. Fausto, 1990. Comunidade deostracodes associada ao bentos e macrófitas aquáticas da Lagoado Gentil, Tramandaí, Rio Grande do Sul. Acta Limnol. Brasil.3: 807–828.
Würdig, N. L., E. F. Albertoni, C. P. Ozorio, S. Wiedenbrüg &G. Rodrigues, 1998. The influence of environmental parametersin the structure of the benthic community in coastal lakes andlagoons of Rio Grande do Sul, Brazil. Verh. int. Ver. Limnol. 26:1514–1517.