late quaternary paleolimnology of onepoto maar, auckland, new zealand: implications for the drivers...
TRANSCRIPT

lable at ScienceDirect
Quaternary International 253 (2012) 18e31
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Late Quaternary paleolimnology of Onepoto maar, Auckland, New Zealand:Implications for the drivers of regional paleoclimate
Paul Augustinus a,*, Ursula Cochran b, Giri Kattel a,1, Donna D’Costa a, Phil Shane a
a School of Environment, University of Auckland, PB 92019, 23 Symonds Street, Auckland, New ZealandbGNS Science, PO Box 30-368, Lower Hutt, New Zealand
a r t i c l e i n f o
Article history:Available online 10 March 2011
* Corresponding author.E-mail address: [email protected] (P. A
1 Present address: Centre for Environmental ManagEngineering, University of Ballarat, Ballarat 3353, Aus
1040-6182/$ e see front matter � 2011 Elsevier Ltd adoi:10.1016/j.quaint.2011.02.028
a b s t r a c t
A high-resolution record of lacustrine environments spanning ca. 30e9 cal ka BP was obtained from One-poto maar, northern North Island, New Zealand. The multi-proxy record of environmental change is con-strained by tephrochronology and AMS 14C ages and provides evidence for episodes of rapid environmentalchange during the Last Glacial Coldest Period (LGCP: 28.5e18 cal ka BP) and Late GlacialeInterglacialTransition (LGIT) from northern New Zealand. The Onepoto pollen record indicates that the LGCPwas cold,dry and windy in the Auckland region with vegetation dominated by herbs and grasses in a beech forestmosaic. At the same time the diatom record indicates oligotrophic conditions with low lakes levels andturbulence whilst cladocerans indicate low water temperatures. The inference of cold, dry and windyconditions during the LGCP is supported by geochemical evidence for increased sediment influx, charcoaland CO2 limiting conditions for terrestrial macrophytes. Rapid climate amelioration and forest expansionafter ca.18 cal ka BP correspondswith reduced sediment influx, diatomand cladoceran-inferredhigher lakelevels indicating increasing moisture availability and temperature. Diatom flora indicates that an oligo-trophic, circumneutral lake was becoming established during the LGIT but conditions were still variable.Between ca.13.8 and 12.5 cal ka BP two brief drier and possibly cooler episodes are apparent (ca. 13.8e13.2and 13e12.5 cal ka BP) followed by awarm phase combinedwith generally stable high lake levels betweenca. 12.5 and 10.5 cal ka BP. Subsequently the lake water chemistry became more alkaline and eutrophic,possibly because sea level had risen high enough by this time to influence ground water chemistry.
� 2011 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
New Zealand has a large number of lakes that have the potentialto record signals of past environmental changes at both local andregional scales. Furthermore, such lakes should be responsive to anyregional reorganisation of atmospheric and oceanic circulationrelated to global glacialedeglacial climate shifts since New Zealandis located in the mid-latitudes of the Southern Hemisphere, strad-dling the subtropical convergence zone and exposed predominantlyto westerly winds (Pillans, 1990; Shulmeister et al., 2004).
Suitable high-resolution and well-dated lacustrine sedimentrecords of late glacial to early Holocene environments are rare fromthe southwest Pacific. Such records are important in the context ofthe present debate over the nature, distribution, timing and forc-ings of sustained climate perturbations such as the Last Glacial
ugustinus).ement, School of Science andtralia.
nd INQUA. All rights reserved.
Maximum (LGM) in New Zealand (e.g., Alloway et al., 2007;Newnham et al., 2007; Augustinus et al., in press) and resolutionof the debate regarding the persistence of the Antarctic ColdReversal and/or Younger Dryas in Southwest Pacific region(e.g.,Denton and Hendy, 1994; Singer et al., 1998; Alloway et al., 2007;Barrows et al., 2007). In the New Zealand context, the develop-ment of high-resolution paleoenvironmental records has beenhindered by poor chronology, the often fragmentary nature of thesequences and variable sensitivity of the paleoclimatic indicators(McGlone, 1995; Lowe et al., 2008). However, in the Aucklandregion there are several maar crater lakes that provide continuousand high-resolution paleoenvironmental records that span at leastthe late glacial to early Holocene within which are numerousknown age rhyolitic tephra that allow the development of a robustchronology (Molloy et al., 2009).
Furthermore, the Auckland region of northern New Zealand(Fig. 1) is important in a paleoenvironmental context because itforms an ecological boundary between warm far northern NorthIsland and cooler regions to the south with the southern limit ofclosed forest during the Last Glacial Maximum somewhere in the
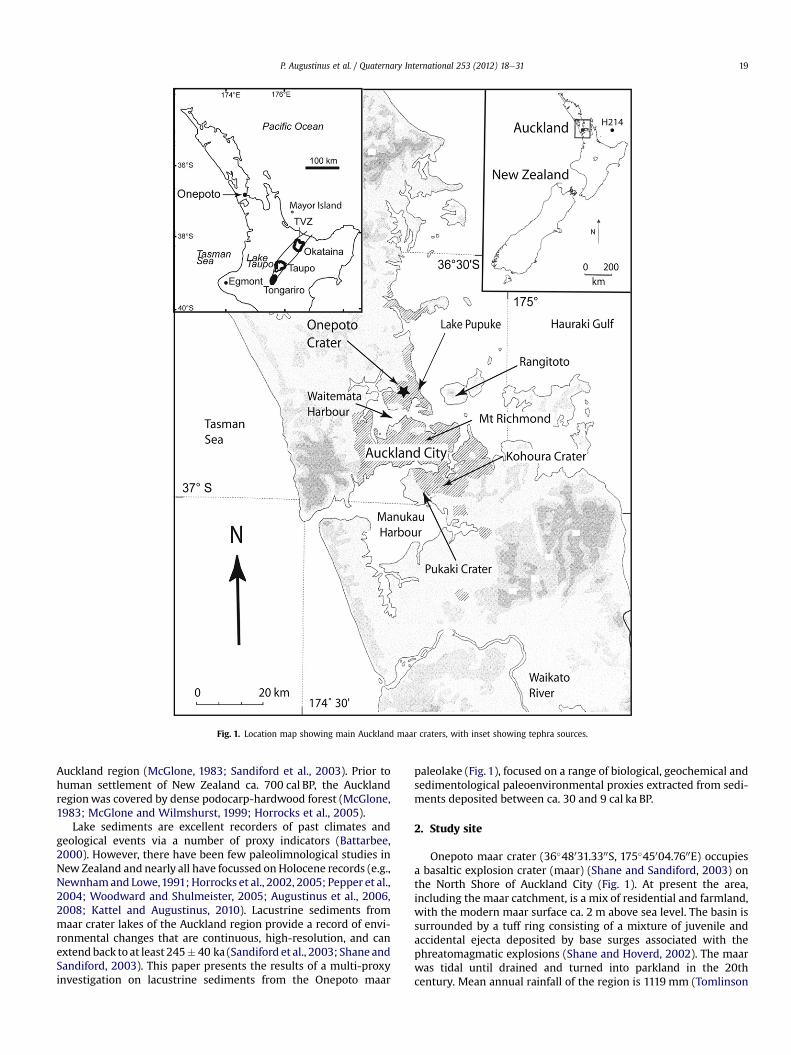
Fig. 1. Location map showing main Auckland maar craters, with inset showing tephra sources.
P. Augustinus et al. / Quaternary International 253 (2012) 18e31 19
Auckland region (McGlone, 1983; Sandiford et al., 2003). Prior tohuman settlement of New Zealand ca. 700 cal BP, the Aucklandregionwas covered by dense podocarp-hardwood forest (McGlone,1983; McGlone and Wilmshurst, 1999; Horrocks et al., 2005).
Lake sediments are excellent recorders of past climates andgeological events via a number of proxy indicators (Battarbee,2000). However, there have been few paleolimnological studies inNew Zealand and nearly all have focussed on Holocene records (e.g.,Newnhamand Lowe,1991; Horrocks et al., 2002, 2005; Pepper et al.,2004; Woodward and Shulmeister, 2005; Augustinus et al., 2006,2008; Kattel and Augustinus, 2010). Lacustrine sediments frommaar crater lakes of the Auckland region provide a record of envi-ronmental changes that are continuous, high-resolution, and canextendback to at least 245� 40 ka (Sandiford et al., 2003; Shane andSandiford, 2003). This paper presents the results of a multi-proxyinvestigation on lacustrine sediments from the Onepoto maar
paleolake (Fig. 1), focused on a range of biological, geochemical andsedimentological paleoenvironmental proxies extracted from sedi-ments deposited between ca. 30 and 9 cal ka BP.
2. Study site
Onepoto maar crater (36�48031.3300S, 175�45004.7600E) occupiesa basaltic explosion crater (maar) (Shane and Sandiford, 2003) onthe North Shore of Auckland City (Fig. 1). At present the area,including the maar catchment, is a mix of residential and farmland,with the modern maar surface ca. 2 m above sea level. The basin issurrounded by a tuff ring consisting of a mixture of juvenile andaccidental ejecta deposited by base surges associated with thephreatomagmatic explosions (Shane and Hoverd, 2002). The maarwas tidal until drained and turned into parkland in the 20thcentury. Mean annual rainfall of the region is 1119 mm (Tomlinson

P. Augustinus et al. / Quaternary International 253 (2012) 18e3120
and Sansom, 1994a), and is distributed evenly throughout the year.Mean daily air temperature is 15.6 �C and the average daily range is6.0 �C (Tomlinson and Sansom, 1994b).
3. Materials and methods
3.1. Core collection and sampling
A 60.5 m long core (Core On2) was obtained fromOnepoto maarcrater (Fig. 1) using a mobile drill rig. The basaltic lapilli at the baseof the core is attributed to the formative eruption and 39Are40Ardated to 245� 40 ka (Shane and Sandiford, 2003). Subsequent tothis event, the core provides a complete record of sedimentation inthe crater, which comprises ca. 24.5 m of organic-rich laminatedlacustrine sediments interbedded with distal rhyolitic, andesiticand local basaltic tephra (Shane and Hoverd, 2002; Molloy et al.,2009) overlain by ca. 36 m of marine sediments deposited subse-quent to the early Holocene breach of the crater wall commensu-rate with postglacial sea level rise ca. 8.5 cal ka BP (Hayward et al.,2002).
The focus is on the paleolimnological record contained in ca.270 cm of lacustrine sediments deposited immediately prior to themarineefreshwater transition here (Fig. 2). The sediments weresampled at intervals ranging from 10 to 2 cm depending on theproxy employed. All sediment and sample depths are given asdepths below the marineefreshwater sediment interface (definedas 0 m) in core On2.
3.2. Core chronology
The core chronology was established using chemical fingerprinting of well-dated rhyolitic tephra from the Taupo VolcanicZone (TVZ) (Shane and Hoverd, 2002; Molloy et al., 2009; Table 1,Fig. 2) and AMS 14C dating (Augustinus et al., in press). All of therhyolites are calc-alkaline and havemajor oxide abundances typicalof TVZ deposits (e.g. Shane, 2000) with glass composition, miner-alogy and stratigraphy allowing most rhyolite tephra erupted afterthe Rotoehu tephra (ca. 50 ka BP) to be matched with known TVZevents (Molloy et al., 2009). Accelerator mass spectrometry (AMS)14C age determinations were carried out on both bulk sedimentsamples and pollen concentrates. The analyses were undertaken onthe tandem accelerators at the Australian Nuclear Science andTechnology Organisation (ANSTO), Sydney, Australia, and the RafterRadiocarbon Laboratory, Wellington, New Zealand.
3.3. Inorganic sediment properties
Grain size analysis was undertaken at ca. 2.5 cm intervals. Thesamples were treated with H2O2 to remove organic matter,dispersed with sodium hexametaphosphate and sonified beforemeasurement with a Malvern Mastersizer 2000 laser particle sizer.Grain size is given here as the percentage greater than 32 mm andused as an indicator of clastic sediment influx to the lake as this sizefraction was shown by Augustinus et al. (in press) to match withother sediment influx proxies.
Trace and major element concentrations were measured usingX-ray fluorescence on a Siemens SRS3000 in the Department ofGeology, University of Auckland. Sub-samples for XRF were ashedat 930 �C, ground, homogenised and fused in glass beads preparedwith 12:22 lithium tetraborate to lithium metaborate XRF flux ina ratio of 2 g of ignited sample to 6 g of flux. The calibration uses 34international standards and uses concentrations based on variablealphas as the matrix correction method using Brucker SPECTRA-plus software (V1.51). Samples were collected at 2.5 cm intervalswith Ti, Al, and Ni used here as indicators of phases of clastic
sediment influx to the lake, and Fe/Mn is used as an indicatorof redox conditions (Lucchini et al., 2003). Elemental values aregiven as percentage for major elements (Al, Ti, Fe, Mn) and ppm fortrace Ni.
3.4. Organic matter geochemistry
Total organic carbon (TOC), total nitrogen (TN), total sulphur(TS) and bulk organic matter d13C and d15N analysis of sedimentorganic matter first involved acidification with 10% HCl to removeany carbonate, after which samples were washed three times withde-ionised water, freeze-dried and crushed to a powder. The TSanalyses were undertaken using a LECO TruSpec CNS analyser inthe School of Environment, University of Auckland, whilst TOC andTN analyses were undertaken at the Isotope Laboratory at theUniversity of Waikato, Hamilton, New Zealand, using a Dumaselemental analyser (Europa Scientific ANCA-SL) interfaced to anisotope mass spectrometer (Europa Scientific 20-20 Stable IsotopeAnalyser) for d13C and d15N measurement.
The relative proportion of sedimentary organic matter derivedfrom terrestrial plants and in situ algal productivity can be identi-fied from the difference between the TOC/TN ratios, with increasedinflux of terrestrial plant macrophyte-derived organic matterproducing TOC/TN ratios >20, whilst algae typically have TOC/TNratios between 4 and 10 (Meyers, 1994; Meyers and Lallier-Vergès,1999). Increasing TOC is often used an indicator of increase interrestrial biomass and higher productivity (e.g., Meyers andLallier-Vergès, 1999; Street-Perrott et al., 2004), whilst high TN isan indicator of proteinaceous material sourced from phyto-plankton. TS indicates the formation of sulphides associated withenhanced anoxic conditions below the lake hypolimnion.
Several factors influence d13C values in terrestrial plant-derivedorganic matter: (1) partial pressure of atmospheric CO2 (pCO2),with low atmospheric pCO2 during the LGCP driving d13C enrich-ment due to reduced discrimination against the heavier isotopeduring the photosynthetic process; (2) climatic stresses such asfluctuations in water supply/dryness and wind stress on terrestrialplants; (3) proportion of organic matter from different sourceorganisms; and (4) post-depositional d13C changes (Street-Perrottet al., 2004). Since there are no native C4 plants in New Zealand,d13C enrichment due to climate change-induced shifts from plantsusing the C3eC4 photosynthetic pathways did not occur, thus d13Cchanges in the sediment organic matter need to be viewed asa response to changes in (1) and (2). d15N is an indicator of organicmatter sources as dissolved inorganic nitrate has a mean isotopicratio of 7e10&, whereas atmospheric molecular nitrogen hasa d15N of ca. 0& (Meyers and Lallier-Vergès, 1999).
Biogenic silica (BGS) content of lake sediments is commonlyused as an indicator of algal productivity (e.g., Prokopenko et al.,2001; Augustinus et al., 2008) and it was estimated using themethod of Mortlock and Froelich (1989), with Si concentrationmeasurement by ICP-OES at GNS Science, Wairakei, New Zealand.
3.5. Biological proxies
Diatom analysis was carried out on sub-samples of approxi-mately 1 cm3 taken at 5e10 cm intervals from 265 cm to 1 cmdepth in Onepoto core On2. Diatom samples were prepared foranalysis by oxidation with dilute hydrogen peroxide (10%) ina water bath at 70 �C for 2 h, or until obvious organic residue hadbeen removed. Samples were then rinsed by repeated washingwith distilled water. Sub-samples of processed sediment weremounted using Naphrax�.
At least 300 diatom valves were counted from each sample usinga Zeiss compoundmicroscope at�600magnification (oil immersion)

Fig. 2. Photographic log of Onepoto core On2 between 10 cm above and 270 cm depth below the marineefreshwater transition showing marker tephra and selected AMS 14C ages.
P. Augustinus et al. / Quaternary International 253 (2012) 18e31 21
withdifferential interference contrast. TaxonomywasbasedonFoged(1979), Hustedt (1985), Krammer and Lange-Bertalot (1986, 1988,1991a, 1991b, 2000), Round et al. (1990) and Sonneman et al.(2000). Diatom species abundances are expressed as percentages,
Table 1Rhyolitic tephra and their ages from Onepoto maar crater.
Tephra name Depth (cm) Age (14C yr)a Age (cal yr BP)c Age (cal yr )d
Rotoma 25 8530� 10 9505� 25 9523� 11Opepe 37 9050� 40 10,075� 155 10,217� 28Waiohau 95 11,850� 60 13,635� 165 13,689� 95Rotorua 114 13,080� 50 15,425� 325 15,824� 332Rerewhaakaitu 142 14,700� 110 17,625� 425 17,913� 241Okareka 192 18,323� 56b 21,800� 500b 21,862� 183Te Rere 218 21,100� 320 25,271� 779 25,256� 446Kawakawa 233 22,590� 230 27,097� 957 27,282� 373Poihipi 253 23,470� 170 28,181� 383 28,254� 196Okaia 263 24,600� 160b 30,092� 340b 29,435� 249
a Ages from Lowe et al. (2008) (1� s range, 68.2%).b Ages from Molloy et al. (2009).c Ages from Lowe et al. (2008) (2� s range, 95.4%).d AgescalibratedusingOxCal 4.1using INTCal09 (Reimeretal., 2009) after correction
for Southern Hemisphere offset (�40� 20 yr; Hogg et al., 2009) (2� s range, 95.4%).
calculated using total numbers of valves recorded for each sample.Concentrations are expressed as valves per gram of sediment afterdividing the above by the dry weight of the initial sample used.Specieswere grouped together according to their habitat preferencesas derived from Round et al. (1990) and Van Dam et al. (1994).
The method of cladoceran sample preparation and analysisreferred to here is detailed in Kattel and Augustinus (2010) andinvolved the analysis of 48 samples collected at 2e10 cm spacingbetween 0 and 270 cm depth in Onepoto core On2. The closerspacing was used over the Last GlacialeInterglacial Transition inorder to identify rapid limnological changes that would be missedby the coarser resolution analysis. Given their limited taxonomicprecision, identification of the New Zealand chydorid cladoceranswas carried out only to the genus level. However, cladoceransregarded as cosmopolitan species were identified to the specieslevel using Frey (1958, 1959, 1986), Chapman and Lewis (1976),Whiteside et al. (1978) and Korhola and Rautio (2001). Totalcladoceran ephippiawere also recorded because they are one of thepotential indicators of past climatic events.
Samples for pollen analysis were collected from the 270 cm longcore On2 at 2.5 cm intervals, except between 0 and 30 cm depth,

P. Augustinus et al. / Quaternary International 253 (2012) 18e3122
upto the marineelacustrine transition (defined as 0 m; Fig. 2). Thesamples were prepared by the standard acetylation and hydro-fluoric acid method and bleached (Moore et al., 1991). The pollensum was at least 250 grains, excluding Leptospermum type, herba-ceous wetland plants and ferns. Percentage pollen diagram wasprepared using the TILIA program (E. Grimm, Illinois StateMuseum). Charcoal identified was all microscopic and quantifiedby counting all particles larger than 5 mm encountered over 100fields of view at �200 magnification. Charcoal encountered ismicroscopic.
4. Results
4.1. Stratigraphy
The composite sediment log from the Onepoto paleolake corehighlights the major sedimentary facies identified (Fig. 2). Threedistinct types of sediment were encountered in the core: lakesediment, tephra, and estuarine sediment. The estuarine sedimentsare massive, fossiliferous clayey silts with generally homogenouscolour, structure and grain size (Fig. 2). Lacustrine sedimentcomprises of generally well-laminated muds with variable colourand degree of lamination development. Here, the focus is on theupper 270 cm of the sequence below the marineefreshwatertransition (0 m).
4.2. Chronology and sedimentation rates
Ten key rhyolitic tephra bedswere encountered in the 0e270 cmdepth section of the Onepoto core and are shown in Table 1 andFig. 2. The tephra ages are given in Table 1 with the 14C ages derivedfrom Lowe et al. (2008) and Molloy et al. (2009). Nine AMS 14C ageswere obtained from the core on a mix of bulk carbon and pollenconcentrates (Table 2). Both tephra and AMS 14C ages were cali-brated with OxCal 4.1 using the latest IntCal09 calibration (Reimeret al., 2009) after correction for Southern Hemisphere offsetof �40� 20 BP (Hogg et al., 2009) to construct the age model.
Sedimentation rates were extrapolated to unit boundariesdefined by known age tephra to obtain age estimates for transitionsbetween major inferred paleoenvironmental shifts using a simple
Table 2AMS radiocarbon ages from Onepoto maar paleolake sediments
CorrectedDepth (cm)b
Laboratorycode
d13C % Modern 14C age BPc Calendarage yr BPd
1a NZA16604 �28.59 36.32� 0.20 8085� 45 9009� 8951a NZA16002 �28.8 27.4� 0.23 10,336� 70 12,192� 14461 OZH871 �28.5 28.76� 0.36 10,010� 110 11,562� 20071a NZA16000 �29.48 22.54� 0.22 11,919� 80 13,766� 10871 NZA16243 �25.34 24.04� 0.10 11,398� 70 13,267� 7382a NZA16605 �27.79 24.97� 0.16 11,096� 55 12,975� 98132a NZA16001 �28.16 17.43� 0.20 13,983� 100 17,066� 155167 NZA16002 �27.33 12.50� 0.15 16,653� 85 19,799� 173191.5 NZA16791 �22.94 10.18� 0.13 18,300� 100 21,847� 198196 OZH872 �23.5 10.26� 0.15 18,290� 120 21,842� 207266 OZH876 �28.2 4.69� 0.12 24,570� 210 29,671� 295
Wk¼Waikato Radiocarbon Dating Laboratory, Hamilton, New Zealand; NZA¼Rafter Radiocarbon Dating Laboratory, Lower Hutt, New Zealand; OZH/K¼Australian Nuclear Science and Technology Organisation, Sydney, Australia.Ages determined from the bulk organic fraction after standard acid/alkali/acidtreatment except where indicated.
a Ages determined from pollen concentrates.b All depths are below the marineefreshwater sediment interface.c 1� s range, 68.2%.d Ages calibrated using OxCal 4.1.1 using IntCal09 (Reimer et al., 2009) after
correction for Southern Hemisphere offset (�40� 20 yr; Hogg et al., 2009) (2� s
range, 95.4%).
linear extrapolation between age markers. The details of the agemodel are presented in Augustinus et al. (in press) and will not bediscussed further here.
4.3. Grain size
Sediment grain size is variable and displays trends that closelymirror other erosion indicators (Fig. 3). Percentage of sedimentgreater than 32 mm was chosen for this purpose as the fractioncoarser than 32 mm was considered to reflect the influx of terrige-neous detritus to the maar lake (Augustinus et al., in press). Fig. 3indicates significant changes in grain size up core with sedimentfiner than 32 mm size dominating below the Kawakawa tephra(233 cm depth), above which the percentage of sediment coarserthan 32 mm increases sharply with four distinct peaks between theKawakawa and Rerewhakaaitu tephra (142 cm depth). Above thisdepth there is a sharp reduction in percentage >32 mm withoscillation around a mean of ca. 15%. Above 50 cm depth,percentage >32 mm increases slightly up to the marineefreshwatersediment transition (Fig. 3).
4.4. Inorganic geochemistry
Sediment Al, Ti and Ni content variation reflects influx ofterrestrial sediment with clear trends apparent in each of theelements down core (Fig. 3). Al and Ti concentrations displaysimilar variation from 270 to 136 cm depth whereby they increaseimmediately above 270 cm before reducing sharply at 245 cmdepth (Fig. 4). Al and Ti percentages remain high and oscillate abouta mean value of 19 and 1.5% respectively until 151 cm depth abovewhich a steady reduction occurs until 140 cm. In contrast, Niconcentration displays a sharp increase at 242 cm depth abovewhich they rise to a peak at 240 cm (Fig. 3). Between 242 and136 cm depth Ni displays greater variability than Al and Ti withseveral peaks and troughs before reducing to low levels above140 cm depth (Fig. 3).
Al, Ti and Niall display low but variable concentrations from 140up to 60 cm above which a sharp reduction in all three occurs andnear constant values are maintained until 48 cm depth (Fig. 3).Above 42 cm Al and Ti% increase to a peak between 40 and 30 cmdepth before reducing to low levels up to the marineefreshwatertransition.
Fe/Mn ratio is generally low and variable but with a broad peakat 225e200 cm depth and a further peak at 94e82 cm beforereturning to low Fe/Mn between 60 and 50 cm depth. Subse-quently, the ratio increases to a peak of 900 at 30 cm depth abovewhich it declines to 110 just below the marineefreshwater transi-tion (Fig. 3).
BGS percentages are high at 270 cm depth above which theyreduce to a minimum at 245e230 cm (Fig. 4). Above 220 cm theconcentrations increase and oscillate around ca. 5% except forminima between 190 and 210 cm. Above ca. 140 cm depth BGSvaries and increases to a peak of 14% at 100 cm depth, above whichBGS concentrations drop sharply before increasing to 6e7%between 70 and 40 cm. BGS reduces sharply above 40 cm to ca. 1.5%before increasing to ca. 6% up to the marineefreshwater transitions(Fig. 4).
4.5. Organic matter geochemistry
Bulk sediment total organic carbon (TOC), total nitrogen (TN),total sulphur (TS), TOC/TN (Fig. 3) and d13C content (Fig. 4) eachdisplay significant variability down core. Sediment TOC displayslow but variable percentages between ca. 270 and 145 cm depth.TOC% increases steadily up core with a sharp increase at 122 cm

Fig. 3. Multi-proxy record showing index and elemental geochemical trends in Onepoto maar paleolake spanning ca. 30e9 cal ka BP. Horizontal cross-hatched boxes labelled (1)and (2) represent inferred erosion phases (sediment influx), whilst (3) represent inferred warm, moist phase with reduced sediment influx to the lake basin. (1) also represents theduration of the LGCP in the Onepoto record.
P. Augustinus et al. / Quaternary International 253 (2012) 18e31 23
after which it increases until 90 cm depth. Between 85 and 75 cmdepth a marked drop in TOC occurs and at 70 cm depth there isa further sharp reduction in TOC abovewhich near constant level ofca. 8% is maintained before a further sharp increase at 42 cm depthup to the marineefreshwater transition (Fig. 3).
The TOC/TN ratio for the section of core between 270 and 0 cmranges from 38 to 10.5 (Fig. 3). The ratio is at a minimum between240 and 230 cm depth, increasing up core to >20 with furtherminima at 180 and 150 cm respectively. A subsequent peak inTOC/TN occurs at 122 cm above which it remains at >20 untilbetween 70 and 40 cm depth with a minimum of 12.2 at 50 cm. The
Fig. 4. Multi-proxy record showing trends in a range of organic matter derived and biopercentage is smoothed with a 2 point running mean. Cross-hatched box labelled (1) represe(4) represents the same sustained warm and moist period labelled (3) in Fig. 3.
TOC/TN ratio subsequently increases again to values mostly>20 upto the marineefreshwater transition. TS is uniformly low between270 and 140 cm depth above which TS% rises gradually to ca. 1% at115 cm, a percentage around which it oscillates until it increasesabruptly above 40 cm depth with a peak of 3%.
Bulk d13C enrichment commences immediately above the Poi-hipi tephra (253 cm depth) and attains a peak enrichmentof�22.5& at 225 cm depth. Marked d13C depletion to�27& occursimmediately above the Te Rere tephra (218 cm depth) beforefurther d13C enrichment between 220 and 145 cm depth (Fig. 4).A sharp drop to more depleted d13C above 145 cm is maintained
logical proxies from Onepoto maar spanning ca. 30e9 cal ka BP. The Ascarina pollennts the LGCP, whilst (2) and (3) represent inferred cooler and drier phases respectively.

P. Augustinus et al. / Quaternary International 253 (2012) 18e3124
until 82 cm depth above which a further brief phase of d13Cenrichment occurs before further depletion and a final phase ofd13C enrichment between 70 and 45 cm depth. d13C remainsdepleted up to the marineefreshwater transition.
d15N is relatively steady from 270 to 80 cm depth at ca.2e3&(Fig. 3). d15N subsequently attains an enrichment peak of4.8& before reducing sharply to almost 0& at 75 cm depth. Abovethis depth d15N increases toþ2.2& before reducing sharply to 0.5&at 20 cm depth. d15N enrichment characterises the remainder of thecore up the to the marineefreshwater transition (Fig. 3).
4.6. Diatom assemblages
Diatoms are well preserved in the Onepoto paleolake sedimentand occur in moderate concentrations (millions of valves per gram)in most samples. Over 70 species were identified in this sequencebut on average only 14 different species were counted from eachsample. The diatom assemblages at most depths, except for those inthe upper 40 cm, are dominated by Discostella stelligera andStaurosirella pinnata sensu lato (Fig. 5). D. stelligera is widespreadand abundant in many New Zealand lakes (Reid, 2005). It isa freshwater, planktonic diatom that prefers circumneutral, oligo-trophic waters (Sonneman et al., 2000).
S. pinnata sensu lato is used here to refer to several species of thefamily Fragilariaceae. S. pinnata appears to be the most abundant ofthese species but Punctastriata ovalis, Staurosira construens andStaurosira elliptica are also present. These species are difficult todifferentiate using light microscopy (Williams and Round, 1987).Furthermore, small species of the family Fragilariaceae (such asS. pinnata, P. ovalis, S. construens and S. elliptica) are commonworldwide and are frequently reported to co-occur so that their
Fig. 5. Diatom record from Onepoto maar between ca. 30 and 9 cal ka BP showing percenta summary plot of different habitat types according to individual species preferences).
environmental preferences are likely to be similar (Haworth, 1976).Therefore, for the purposes of this study, not every taxon of thefamily Fragilariaceaewas enumerated individually. S. pinnata sensulato has been found in a diverse range of environments, but basedon a number of studies cited in Appendix it is considered reason-able to associate this taxonwith shallow/turbulent, slightly alkalinewaters in moderately cold conditions.
4.6.1. Zone 1 (265e250 cm)This zone is characterised by a peak in diatom concentration and
a peak in the abundance of D. stelligera at 255 cm depth(Fig. 5). Thetychoplanktic diatoms Pseudostaurosira brevistriata and S. pinnataare also significant in this zone whilst benthic diatoms are a minorcomponent of the diatom sum.
4.6.2. Zone 2 (250e140 cm)This zone is defined by lower diatom concentrations than in
zones 1 and 3; a dominance of S. pinnata sensu lato (60e90%); andthe presence of the aerophilic diatom Diatomella balfouriana(Fig. 5). D. stelligera percentages are relatively low except between220 and 170 cm depth with a peak in abundance at 184 cm (Fig. 5).The only significant benthic diatom is Planothidium frequentissi-mum, which appears between 210 and 170 cm depth. The aerophilicdiatom Diatomella balouriana is a not able component of the diatomsum between 220 and 170 cm depth, coeval with the zone of highD. stelligera and P. frequentissimum percentages (Fig. 5).
4.6.3. Zone 3 (140e40 cm)This zone is characterised by containing the highest concentra-
tions of diatoms in the sequence and the highest proportions ofD. stelligera (Fig. 5). However, both the diatom concentration and
age abundance of major species (those occurring at >5% in one or more samples, and

P. Augustinus et al. / Quaternary International 253 (2012) 18e31 25
proportions of D. stelligera are variable within the zone and thepercentage of S. pinnata is relatively high. Towards the topof the zone(40e60 cm depth), proportions of S. pinnata decrease below 30%whilst D. stelligera dominates the diatom sum and diatom concen-trations reach their peaks at 42 and 56.5 cm depth. Benthic diatomsonlymake a significant contribution to the diatom sum at the base ofthe zone and aerophilic diatoms are virtually absent (Fig. 5).
4.6.4. Zone 4 (40e0 cm)Diatom concentrations are lowest in zone 4 (Fig. 4) and it is
dominated by Aulacoseira granulata, P. brevistriata and S. pinnata(Fig. 5). A. granulata has almost completely replaced D. stelligeraabove 30 cm depth and Cyclotella meneghiniana becomes a signifi-cant componentof thediatomsumand increases in abundance to thetop of the sequence. This zone is also marked by an increase in thebenthic diatoms Achnanthidium, Cymbella deliculata and Cymbellagracilis between 40 and 20 cm depth commensurate with reductionin S. pinnata and low planktonic diatom percentages (Fig. 5).
The details of the Onepoto pollen and cladoceran recordsbetween 0 and 270 cm depth have been discussed in Augustinuset al. (in press) and Kattel and Augustinus (2010) respectively.Consequently the trends in these taxa up core will not be discussedfurther here although summary pollen and cladoceran diagramsare given in Figs. 6 and 7.
5. Discussion
The Last Glacial Coldest Period (LGCP) to early Holocene pale-olimnological record from Onepoto maar paleolake has been sub-divided into four phases that will be discussed with respect to thesynchronous changes in many of the proxies for environmentalchange examined here.
5.1. Pre-LGCP (ca. 30e28.5 cal ka BP)
This period appears to represent a shallow, oligotrophic, cir-cumneutral lake as indicated by high diatom concentrations and
Fig. 6. Percentage composition of main elements of the pollen record from Onepoto maar bon sum of dryland taxa.
moderate percentages of the planktic diatom D. stelligera (Figs. 4and 5). However, the high percentages of S. pinnata suggest thatlimnological conditions were fluctuating enough to allow S. pinnatato remain competitive. The pollen record from Onepoto maarpaleolake indicates that the Auckland region landscape at this timewas largely mixed forest with Fuscopora, Nothofagus menziesii, andLibocedrus dominant within a shrubland-grassland environmentdominated by Halocarpus (Fig. 6). The presence of Dacrydiumcupressinum and Prumnopitys taxifolia indicates that conditionswere cool and wet as P. taxifolia is the most frost tolerant of the tallpodocarp taxa (Sandiford et al., 2003). Increased wetness is sup-ported by the absence of charcoal from the record at this time(Fig. 3).
The cladoceran record over this period is similar to that duringthe LGCP although at ca. 30 cal ka BP littoral cladocera dominateand ephippia concentrations are high, with the latter indicative ofextreme environmental stress and steep climatic gradients whichinduced the chydorid cladocerans to undergo gamogenesis (Katteland Augustinus, 2010). The increasing importance of cold-tolerantBosmina meridionalis and the large Alona sp., Alona affinis and Alonaquadrangularis type at this time confirm that lake waters were cool,whilst low cladoceran L:P ratios suggest constant lake levels at thistime (Figs. 4 and 7).
5.2. LGCP (28.5e18 cal ka BP)
The pollen record indicates that the Auckland region during thisperiod was dominated by patchy beech forest with Fuscopora andN. menziesii within a shrubland-grassland environment dominatedby Halocarpus and Poaceae(Fig. 6). The duration of the LGCP in theAuckland region is represented byD. cupressinum pollen whichmaintains low levels between ca. 28.5 and 18 cal ka BP (Figs. 4and 6). Tree ferns (Cyathea dealbata) were rare, and, whencombined with the presence of N. menziesii support a model oftemperature reduction of at least 4 �C during the LGCP in theAuckland region (Sandiford et al., 2003). Microscopic charcoal iscommon between ca. 28.5 and 18.5 cal ka BP (Fig. 3) andmost likely
etween ca. 30 and 9 cal ka BP. The pollen diagram is represented as a percentage based

Fig. 7. Percentage composition of major cladoceran taxa in Onepoto maar between ca. 30 and 9 cal ka BP. Included are the main planktonic and littoral taxa, littoral/planktonic (L:P)ratio and concentration of ephippia (counts/g of dry sediment).
P. Augustinus et al. / Quaternary International 253 (2012) 18e3126
reflects burning associated with drier and windier climate duringthe LGCP. However, it is possible that the charcoal influx reflectsthe erosion of the tuff ring soils with reworking of the char-coal, although inferred phases of erosion during the Late Gla-cialeInterglacial Transition (see Section 5.3) do not display a coevalincrease in charcoal so that a primary burning signal is thepreferred interpretation for the charcoal peaks during the LGCP.
Swamp vegetation expanded during the LGCP at Onepoto(Figs. 6 and 8) with the expansion of wetland vegetation probablyreflecting lake level reduction and subsequent expansion of areasthat could be colonised, rather than an increase in lake level since inthis scenario the steep crater rim would reduce the available areafor colonisation. Conditions must have been favourable for S. pin-nata during this period as it dominates the diatom taxa. Thisimplies relatively shallow and/or turbulent waters, moderately coldand very variable conditions (Vos and de Wolf, 1988; Weckströmand Korhola, 2001; Schmidt et al., 2004). This interpretation issupported by a reduction in planktic diatoms, although the increasein D. stelligera between ca. 25.5 and 20 cal ka BP (Figs. 4 and 5) mayindicate an increase in lake levels over this period.
The occurrence of the soil diatom D. balfouriana between ca. 23and 20 cal ka BP (Fig. 5) indicates either an increase in wetland/moist soil environments around the lake and/or an increase in thetransport of aerophilic diatoms to the central lake basin during thisperiod. It is likely that both of these changes occurred as the pollenrecord suggests an expansion of wetland areas around the lake(Fig. 6) and increased sediment influx and charcoal input are alsorecorded at this time (Figs. 3 and 8). Increased sediment influx tothe lake also explains the lower concentrations of diatoms observedin this zone as increased input of allochthonous sediment would
have lowered the diatom to sediment ratio even if diatomproductivity remained constant (Fig. 4). At this time Onepoto maarlake was probably relatively shallow, cold and turbulent, with anincreased transport of allochthonous soil diatoms into the lake.
The diatom-inferred model of low lake level and cool lake waterduring this period is supported by the dominance of two large cold-tolerant littoral cladocerans, A. affinis and A. quadrangularis whilstmost other littoral chydorid assemblages were low (Kattel andAugustinus, 2010). Although the low ratio of littoral/planktonic(L:P) assemblages suggests increased water depth and open waterareas which would have favoured planktonic Bosmina assemblages(Fig. 4), many other factors may also have influenced changes in L:Pratios so that reliable paleohydrological inferences are not possibleusing this ratio alone. For example, strong seasonality, along withvariations in nutrient dynamics, lake water chemistry, predationand competition often play a prominent role in changes in clado-ceran composition and diversity (e.g., Meijering, 1983; Whitesideand Swindoll, 1988). The production of large numbers of ephippia(reproductive eggs) by chydorid cladocerans is coeval with thetermination of the LGCP at ca. 18 cal ka BP and may reflect cladoc-eran response to environmental stress at this time (Kattel andAugustinus, 2010).
The inferred climatic deterioration during the LGCP, increasedallochthonous sediment influx and cool, relatively shallow lakewaters represented by the biological record is supported by d13C,grain size, sediment elemental geochemistry, TOC/TN, diatomconcentrations and BGS (Figs. 3, 4 and 8) whereby the pollen anddiatom-inferred cool LGCP is contemporaneous with d13C enrich-ment and increased clastic sediment influx. The TOC/TN ratio duringthe LGCP usually exceeds 20 (Fig. 3) indicating a predominantly

Fig. 8. Summary diagrams showing typical differences in selected proxies for lake and catchment during: (a) Last Glacial Coldest Period (ca. 28.8e18 cal ka BP), and (b) LateGlacialeInterglacial Transition to early Holocene (ca. 16e9 cal ka BP). Note that the latter period is one of significant variability in many of the proxies and does not represent thesituation throughout this time.
P. Augustinus et al. / Quaternary International 253 (2012) 18e31 27
terrestrial macrophyte source for the organic matter (Meyers andLallier-Vergès, 1999). The terrestrial macrophyte-derived d13Csignal in the lacustrine organicmatter during this periodwas largelydriven by superimposed changes in atmospheric pCO2 and climat-ically induced water stress, not changes in algal productivity.Consequently, d13C enrichment coevalwith the cooling, burning andclastic sediment influx peaks probably reflects increased stress onthe terrestrial plants that dominate the organic matter flux to thelake basin.
The pollen-inferred reduction of forest and expansion of herbsand grasses probably reduced ground cover and allow increasederosion of the crater rim soils and exposed tuff ring. This inter-pretation supported by the higher Ti and Al contents of the sedi-ment during the LGCP (Fig. 3). These elements represent variationin clastic sediment influx as they are exclusively related to theterrigenous component of the sediment as they represent the mainsilicate phases (Nesbitt and Wilson, 1992; Hill et al., 2000; Lucchiniet al., 2003). Ti and Al start to increase at ca. 29.5 cal ka BP, ca. 1e1.5ka prior to the inferred commencement of the LGCP (Fig. 4). Thesubsequent sharp increase in Ni concentrations in the sediments atca. 27.5 cal ka BP reflects deep erosion of the tuff ring soils anderosion of the tuff ring itself (Augustinus et al., in press). The Ni isprobably derived from olivine (Nesbitt and Wilson, 1992) withmuch of the Ni being lost during early stage of chemical weatheringof the newly exposed basalts and tuff ring. The trends in Ti, Al andNi suggest sustained erosion, but with reduced sediment flux afterca. 18 cal ka BP (Fig. 3).
The Fe/Mn ratio is low throughout the LGCP (Figs 3 and 8)suggesting reducing conditions and both Fe and Mn mobilityassociated with reduced lake level. This interpretation is supportedby low TS% during the LGCP indicating more aerobic conditions dueto increased mixing in the water column.
5.3. Late GlacialeInterglacial Transition (ca. 17.9e10.5 cal ka BP)
The postglacial amelioration of climate apparent in the multi-proxy paleoenvironmental analysis of the Onepoto maar lakesediment record is best reflected in the expansion of tall trees,especially D. cupressinum, P. taxifolia and Podocarpus, plus theexpansion of tree ferns, Cyathea dealbata (Fig. 6). The warmingtrend is also indicated by the sharp decline in Libocedrus at ca.18 cal ka BP commensuratewith a reduction in importance of shruband herb pollen, especially Coprosma, Halocarpus and Poaceae(Fig. 6).
Over this period the highest diatom concentrations wereencountered and the highest proportions of D. stelligera (Fig. 4). Thesharp increase in planktic diatoms coeval with termination of theLGCP ca. 18 cal ka BP suggests warming lake water and increasedwater depth as does the commensurate reduction in S. pinnata(Fig. 5). However, the marked variability in diatom concentrationand the alternating proportions of D. stelligera and S. pinnatabetween 17 and 12 cal ka BP suggests unstable lake conditions andvariability in lake level occurred throughout this transitionalperiod.
The distinct change from cold to warm-water preferring littoralcladoceran assemblages indicates that the end of the LGCP shouldhave been abrupt (Kattel and Augustinus, 2010). Nevertheless, thewarm preferring littoral cladocerans were not able to colonise thelake immediately and the continued dominance of B. meridionalisindicates a variable response of the cold-preferring littoralcladocerans and oligotrophic planktonic cladocerans to the rapidlychanging lake environment (Kattel and Augustinus, 2010).
From ca. 14 to 12.5 cal ka BP there was a significant reduction inconcentration of B. meridionalis and an increase in abundance oflittoral chydorid cladocerans such as A. guttata, A. nana and A. excisa

P. Augustinus et al. / Quaternary International 253 (2012) 18e3128
and an increase in the cladoceran littoral:planktonic ratio (Fig. 7)suggesting shallowing of the lake and cooler lake water. The lowabundance of B. meridionalis during this period may be a responseto rapid climate-driven changes in lacustrine conditions (Kattel andAugustinus, 2010).
The increase in TOC/TN after the termination of the LGCP indi-cates increasing dominance of terrestrial organic matter influx overin situ aquatic organic matter production in the lake at that time.There is no indication of erosion events in the other proxies, nor isthere an apparent reduction in aquatic productivity suggesting thatthis represents an increased contribution of terrestrial organicmatter to the overall organic matter flux to the lake base. The peakin BGS% at ca. 14.5 cal ka BP is coeval with reduction of the TOC/TNratio indicating an increased aquatic algal contribution to theorganic matter flux in the lake at that time, although the terrestrialorganic matter flux continued to increase and remained thedominant organic matter component.
After 18 cal ka BP multi-proxy record indicates rapid forestexpansion, slope stability, increasedmoisture availability and risingtemperatures. Increasing moisture availability is supported bypodocarp forest expansion (Fig. 6) and the trend to depleted bulkorganic matter d13C values immediately after ca. 18 cal ka BP (Figs. 4and 8). This depletion most likely reflects increased moistureavailability and reduced wind stress on the terrestrial plantmacrophytes so they are able to discriminate against the heavierisotope of carbon during the photosynthetic process (Augustinuset al., in press). Wetland plants are virtually absent at this time(Fig. 6) and probably indicates the maintenance of high lake levelswith catchment and slope stability indicated by generally relativelylow and stable Ti, Al, and Ni concentrations combined withincreasing terrestrial organic matter influx to the lake.
The steady reduction in pollen of drought and frost tolerantP. taxifolia after ca. 11.8 cal ka BP (Fig. 6) may indicate a moreequable climate. Furthermore, the increase in Ascarina at ca.12.2 cal ka BP and its peak at ca. 11 cal ka BP (Figs. 4 and 6),combined with the decline in P. taxifolia and increase in the treeferns, supports a model of increasing precipitation and reduction infrost frequency after ca. 12.2 cal ka BP. Annual temperatures wereprobably generally similar to today with milder and less windyconditions.
During this late glacial period of overall forest stability in One-poto maar lake brief oscillations in several of the proxies areevident. An episode of increased sediment flux between ca. 13.8and 13.2 cal ka BPwas inferred from the coeval increase in Ti, Al andNi content of the sediment (Fig. 4), although a marked decrease inBGS% and Dacrydium pollen% occurs over this period (Fig. 4) sug-gesting cooler and possibly drier conditions with increased erosion.Lowering of lake level at this time is indicated by reduction inplanktic diatom D. stelligera commensurate with low abundance ofthe planktic cladocera Bosmina and peak in cladoceran L:P ratio(Fig. 4). Hence relatively dry and cool conditions are inferred duringthis period which is near coeval with the Late Glacial Reversal (LCR)apparent in many New Zealand terrestrial paleoclimate records(Alloway et al., 2007).
A subsequent increase in sediment influx between ca. 13 and12.4 cal ka BP is inferred from increased Ni, Al and Ti content of thesediment (Fig. 3). The TOC/TN ratio is >20 over this interval indi-cating a dominantly terrestrial macrophyte source for the organicmatter so that the observed d13C enrichment at this time (Fig. 4)may relate to a brief period of dryness-induced stress on the plants(Augustinus et al., in press). A decrease in lake level at this timeinferred from reduction in planktic diatoms, low levels of Bosminaand high cladoceran L:P ratios supports this interpretation, and thereduced biomass between ca.13 and 12.4 cal ka BP indicated by BGSand TOC reduction suggests cooler conditions.
A marked and rapid change in environment at ca. 12.2 cal ka BPis indicated by the sharp reduction in terrestrial organic matterinflux coeval with changes in TOC/TN to less than 20, d13C enri-chment, increased biogenic silica% and sharp increase inD. cupressinum and Ascarina pollen (Fig. 4). The high biogenic silicaconcentrations reflect increase in algal productivity in the lake andis supported by lower TOC/TN values of 16e12 indicating a mixedalgal-terrestrial organic matter source (Meyers, 1994), whilst theincrease in D. cupressinum and Ascarina pollen reflects increasinglymild, moist conditions. The 12.2e11.5 cal ka BP section of theinferred warmer phase displays reduction in the planktic diatomD. stelligera commensurate with a sharp drop in B. meridionalis andincrease in cladoceran L:P ratio (Fig. 4). Combined, these changessupport a significant lowering of lake level at this time.
The sharp reduction in diatom S. pinnata between ca. 11.5 and10.2 cal ka BP suggests that conditions became more stable and/orwater depth increased. Lake water is inferred to be relatively warmand the scarcity of aerophilic species indicates little allochthonousinput to the lake. Reduced erosion in the catchment is supported byreduction in sediment influx over this period, whilst warmerconditions are indicated by high proportion of frost-intolerantAscarina and increased autochthonous biological sedimentationreflected in high BGS%, enriched d13C indicative of high algalproductivity and reduction in TOC/TN to 16e12 over this period(Figs. 3 and 4). The dominance of the planktic diatom D. stelligeraafter 11.5 cal ka BP is matched by a marked increase in B. mer-idionalis and reduction of the cladoceran L:P ratio suggestinga sharp increase in lake level at this time. The observation that thischange is not reflected in an increase in erosion indicates closedcanopy podocarp forest and slope stability.
d15N attains maximum enrichment at ca. 12.2 cal ka BP afterwhich it decreases rapidly to a minimum at ca. 11.5 cal ka BP (Figs. 4and 8) suggesting a direct link with the inferred phase of maximumwarmth, change in lake level and a change in the source signal ofthe organic matter. The changes in the organic d15N values areprobably associated with depletion of dissolved N in the lakebetween 12.2 and 11.5 cal ka BP due to reduced in-wash of soilnutrients and reflect biological draw down of the limited pool ofdissolved N. The depleted d15N values are probably a response toa period of extremely stable stratification in a deep lake withproduction of lake derived organic matter dominated by N-fixingcyanobacteria.
5.4. Early Holocene (ca. 10.5e9 cal ka BP)
TOC increases sharply at this time, indicating increased influx ofterrigenous organic matter with this interpretation supported bythe coeval increase in TOC/TN ratio to >20 (Fig. 3). The marked d13Cdepletion and reduction in biogenic silica supports this model,though a sharp increase in Ti and Al concentrations indicates thatan increase in erosion of the crater rim soils also occurs at ca.10.5 cal ka BP (Figs. 3 and 4). This increase in erosion and influx ofclastic sediment is supported by the increase in grain size at thistime. After ca. 9.5 cal ka BP, erosion reduces and both aquatic andterrestrial biomass increase indicating the maintenance of warm,moist conditions with catchment slope stability and closed canopyforest which continues up to the level of the marineefreshwatersediment transition at ca. 9 cal ka BP.
A reduction in lake level immediately after ca. 10.5 ka BP issuggested by the increase in benthic diatoms and tychoplankticP. brevistriata (Fig. 5), indicating a shallow/turbulent littoral zoneexisted at this time. Bosmina and cladoceran L:P ratios remainedhigh until ca. 10 cal ka BP indicating that the controls on these taxaare complex and cannot be simply interpreted in terms of lake leveland water temperature. The replacement of D. stelligera by

P. Augustinus et al. / Quaternary International 253 (2012) 18e31 29
A. granulata after ca. 10 cal ka BP indicates that the lake became lessacidic and more eutrophic at this time. As this is close to thetransition to estuarine conditions it is likely that the changes inaquatic taxa encountered here are driven by postglacial sea levelhaving risen high enough to have influenced the chemistry ofground water inflow to the lake.
The increase in TS and Fe/Mn after 10.5 cal ka BP suggestsincreasingly stable lake stratification with production of sulphidesand reducing conditions in the stable anoxic hypolimnion (Fig. 8).This interpretation is supported by the depleted d15N valuesalthough enrichment after ca. 9.5 cal ka BP suggests a change in theorganic matter source such as a reduced contribution fromN-fixingbacteria.
6. Summary and conclusions
Although paleoecological records have been published previ-ously for the Auckland region spanning the LGCP to early Holocene,these reconstructions rely on pollen to unambiguously reflectclimate change (e.g., Sandiford et al., 2003; Newnham et al., 2007).Consequently the present multi-proxy paleolimnological study ofOnepoto maar paleolake provides an important contribution to theunderstanding of the controls on lake environments over theperiod 30e9 cal ka BPin northern New Zealand. In particular, itappears that during the Last Glacial Cold Period in the Aucklandregion, cool conditions deduced from the pollen-inferred vegeta-tion assemblage are coeval with cladoceran and diatom-inferredlower lake levels suggesting cooler and drier and probably windierclimate. These climatic inferences are supported by the sedimentelemental and organic matter geochemistry confirming increasedcatchment erosion and sediment influx accompanied by increaseddryness and stress on terrestrial macrophytes. The LGCP lastedfrom ca. 28.5e18 cal ka BP and confirms previous suggestions(Suggate and Almond, 2005; Alloway et al., 2007; Newnham et al.,2007) that the Last Glacial Maximum, at least in the New Zealandregion of the southwest Pacific, commenced several ka earlier andlasted longer than the Last Glacial Maximum in the marine d18Orecord (e.g., Mix et al., 2001).
Rapid warming accompanied by increase in biomass andbroadleaf forest expansion at ca. 18 cal ka BP is matched bya marked increase in planktic diatoms and cladoceran indicators ofwarmer lake water and increasing lake levels. The trend of inferredwarming and increased moisture availability during the Late Gla-cialeInterglacial Transition is complex and punctuated by twoshort phases of drier and possibly cooler climate between ca.13.8e13.2 and ca. 13.0e12.4 cal ka BP. These episodes overlap withthe timing of the Antarctic Cold Reversal (ACR) and reflect thecooling apparent in the deglacial paleoceanographic histories of theBay of Plenty in core H214 (Samson et al., 2005), core MD97-2121from the south east of the North Island (Carter et al., 2008) andseveral of the New Zealand terrestrial records (e.g., Turney et al.,2003; Williams et al., 2005; Hajdas et al., 2006; Alloway et al.,2007; Vandergoes et al., 2008) at this time. In the northwestSouth Island speleothem records (Williams et al., 2005) suggestthat the late glacial climate reversal commenced slightly later thanthe start of the ACR and continued beyond the start of the YoungerDryas Chronozone (YDC).
Following the cooler and drier phases in the Onepoto record,between ca. 12.2 and 10.5 cal ka BP a complex but generally warmand frost-free period of high lake productivity and generally highlake level occurred which overlaps with the timing of the YDC aswell as the Early Holocene Thermal Maximumwhich is recognisedin many New Zealand paleoenvironmental records between 11.5and 10.5 cal ka BP (Alloway et al., 2007). The evidence for maximumwarmth and high lake levels during the YDC at Onepoto, combined
with observation of drier and possibly cooler phases during theACR, contribute to the growing body of evidence for an asynchro-nous see-saw response to formation of North Atlantic Deep Waterbetween the hemispheres evident in the mid and high latitudes ofthe Southern Hemisphere (Blunier et al., 1998; Blunier and Brook,2001; Knorr and Lohmann, 2003; EPICA Community Members,2006).
The initiation of the LGCP in Auckland region is contempora-neous with the post-AIM3 cooling in the EPICA Drønning MaudLand (EDML) ice core record, as well as the commencement of post-LGCP warming in EDML at ca. 18 ka BP, demonstrating a strongteleconnection between the southern mid-latitudes and Antarctica,at least in this sector of the Southern Hemisphere. Hence the phaserelationship between the Auckland maar lake and EDML recordsand their out-of-phase relationship with the Greenland ice coreDansgaard-Oeschger Events provides further support for inter-hemispheric asynchrony in rapid climate change.
Acknowledgements
We would like to thank the University of Auckland ResearchCommittee (grants 3604411 and 3602487), Australian Institute ofNuclear Science and Engineering (grant 05/004) and The RoyalSociety of New Zealand Marsden Fund (grant UOA0517) for finan-cial support for this project.
Appendix. Ecological preferences of S. pinnata sensu lato
Current autecological preference information for small Staur-osirella and Staurosira species is not usually very specific. There isa fair amount of confusion in the literature as a result of thesespecies having broad environmental tolerances, being difficult toidentify under light microscope and having undergone severalchanges in taxonomic nomenclature (e.g., Williams and Round,1987). Outlined below are the studies that were used to gainenvironmental information from the presence of S. pinnata sensulato in the cores.
Haworth (1976) describes small species of the family Fragi-lariaceae as “pioneers” because they are the first taxa to occurafter glaciation in many sequences from the northern hemi-sphere regardless of lake type. She also suggests they thriveunder conditions of continuous environmental change. Alongaltitudinal transects of alpine lakes these species are oftenabundant at the higher altitude, colder end of the spectrum(Weckström and Korhola, 2001; Schmidt et al., 2004). They cantolerate a wide range of trophic states and generally prefer waterwith a pH >7 (Van Dam et al., 1994). In modern samples fromNew Zealand they have been found in very shallow water aroundlake margins (Hill, 1998; Cochran, 2002). In terms of habitat, theyare classified as ‘tychoplanktic’ because they are generallyconsidered to live attached to sediment grains but often end upin the plankton in shallow and/or turbulent environments (Vosand de Wolf, 1988).
Schmidt et al. (2004) identified morphotypes of S. pinnata and ofStaurosira aff. venter that had a significant response to summerwatertemperature and found their optima to be between 9 and 10 �C.However, the range in mean summer temperature encompassed bytheir dataset was 1.7e12.4 �C and both taxa had monotonicallyincreasing response curves (Schmidt et al., 2004). Therefore, it ispossible thatwith a dataset extending intowarmerwaters, optima ofthese taxa would be found to be higher. Weckström and Korhola(2001) derived summer air temperature optima for several ofthese species from lakes in Arctic Lapland. Their dataset covereda temperature range of 8e15 �C and derived optima for Fragilariabrevistriata, Fragilaria pinnata and Fragilaria construens var. venter of

P. Augustinus et al. / Quaternary International 253 (2012) 18e3130
between 11 and 12 �C. Although neither of these studies covera temperature gradient likely to encompass the spectrum experi-enced by Onepoto Maar lake over the last 30,000 years, they doindicate that small species of Fragilariaceae occur abundantly inlakes with relatively cold mean summer temperatures.
Consequently, it is reasonable to associate S. pinnata sensu latowith shallow/turbulent, slightly alkaline waters in moderately coldand variable conditions. Although S. pinnata can tolerate eutrophicconditions, the presence of D. stelligera throughout much of thesequence suggests that waters were oligotrophic and close to cir-cumneutral for much of this time. The co-existence of these taxawas probably possible because they occupy different habitats withS. pinnata sensu lato living attached to sediment grains or floatingin the littoral zone (Vos and de Wolf, 1988) and D. stelligera in thewater column of the lake (Sonneman et al., 2000).
References
Alloway, B.V., Lowe, D.J., Barrell, D.J.A., Newnham, R.M., Almond, P.C.,Augustinus, P.C., Bertler, N.A.N., Litchfield, N.I., McGlone, M.S., Shulmeister, J.,Vandergoes, M.J., Williams, P.W., 2007. Towards a climate event stratigraphy forNew Zealand over the past 30,000 years (NZ-INTIMATE project). Journal ofQuaternary Science 22, 9e35.
Augustinus, P.C., D’Costa, D., Deng, Y.-B., Hägg, J. and Shane, P. A multi-proxy recordof changing environments from ca 30,000 to 9,000 cal yr BP Onepoto maarpaleolake, Auckland, New Zealand. Journal of Quaternary Science, in press,doi:10.1002/jqs.1463.
Augustinus, P.C., Reid, M., Andersson, S., Deng, Y., Horrocks, M., 2006. Biological andgeochemical record of anthropogenic impacts in recent sediments from LakePupuke, Auckland City, New Zealand. Journal of Paleolimnology 35, 789e805.
Augustinus, P., Bleakley, N., Deng, Y., Shane, P., Cochran, U., 2008. Rapid change inearly Holocene environments inferred from Lake Pupuke, Auckland City, NewZealand. Journal of Quaternary Science 23, 435e447.
Barrows, T.T., Lehman, S.J., Fifield, L.K., De Deckker, P., 2007. Absence of cooling inNew Zealand and the adjacent ocean during the Younger Dryas Chronozone.Science 318, 86e89.
Battarbee, R.W., 2000. Palaeolimnological approaches to climatic change, withspecial regard to the biological record. Quaternary Science Reviews 19,107e124.
Blunier, T., Brook, E.J., 2001. Timing of millennial-scale climate change in Antarcticaand Greenland during the last glacial period. Science 291, 109e112.
Blunier, T., et al., 1998. Asynchrony of Antarctic and Greenland climate changeduring the last glacial period. Nature 394, 739e743.
Carter, L., Manighetti, B., Ganssen, G., Northcote, L., 2008. Southwest Pacific modu-lation of abrupt climate change during theAntarctic ColdReversal-YoungerDryas.Palaeogeography, Palaeoclimatology, Palaeoecology 260, 284e298.
Chapman, M.A., Lewis, M.H., 1976. An Introduction to the Freshwater Crustacea ofNew Zealand. William Collin (New Zealand) Ltd.
Cochran, U.A., 2002. Detection of large Holocene earthquakes in the sedimentaryrecord of Wellington, New Zealand, using diatom analysis. Unpublished PhDThesis, Victoria University of Wellington.
Denton, G.H., Hendy, C.H., 1994. Younger Dryas age advance of Franz Josef Glacier inthe Southern Alps, New Zealand. Science 264, 1434e1437.
EPICA Community Members, 2006. One-to-one coupling of glacial climate vari-ability in Greenland and Antarctica. Nature 444, 195e198.
Foged, N., 1979. Diatoms in New Zealand, the North Island. Bibliotheca Phycologica47, 1e224.
Frey, D.G., 1958. The late-glacial cladoceran fauna of a small lake? Archiv fürHydrobiologie 54, 209e275.
Frey, D.G., 1959. The taxonomic and phylogenetic significance of the headpores ofthe chydoridae (Cladocera). Internationale Revue der Gesamten Hydrobiologie44, 27e50.
Frey, D.G., 1986. Cladocera analysis. In: Berglund, B.E. (Ed.), Handbook of HolocenePalaeoecology and Palaeohydrology. John Wiley & Sons Ltd., pp. 667e692.
Hajdas, I., Lowe, D.J., Newnham, R.M., Bonani, G., 2006. Timing of the late-glacialclimate reversal in the Southern Hemisphere using a high resolution radiocarbonchronology for Kaipo bog, New Zealand. Quaternary Research 65, 340e345.
Haworth, E.Y., 1976. Two late-glacial (late Devensian) diatom assemblage profilesfrom Northern Scotland. New Phytologist 77, 227e256.
Hayward, B.W., Grenfell, H.R., Sandiford, A., Shane, P.A.R., Morley, M.S., Alloway, B.V.,2002. Foraminiferal and molluscan evidence for the Holocene marine history oftwo breached maar lakes, Auckland, New Zealand. New Zealand Journal ofGeology and Geophysics 45, 467e480.
Hill, I.G., Worden, R.H., Meighan, I.G., 2000. Geochemical evolution of a palae-olaterite: the Interbasaltic Formation, Northern Ireland. Chemical Geology 166,65e84.
Hill, N.L., 1998. Palaeoseismicity and neotectonics of the eastern Awatere Fault,Marlborough, New Zealand. Unpublished BSc (Hons) Thesis, Victoria Universityof Wellington.
Hogg, A., Bronk Ramsey, C., Turney, C., Palmer, J., 2009. Bayesian evaluation of theSouthern Hemisphere radiocarbon offset during the Holocene. Radiocarbon 51,1165e1176.
Horrocks, M., Augustinus, P.C., Deng, Y., Shane, P., Andersson, S., 2005. Holocenevegetation, environment and tephra recorded from Lake Pupuke, Auckland,New Zealand. New Zealand Journal of Geology and Geophysics 48, 85e94.
Horrocks, M., Deng, Y., Nichol, S.L., Shane, P.A., Ogden, J., 2002. A palaeo-environ-mental record of natural and human change from the Auckland Isthmus, NewZealand, during the Late Holocene. Journal of the Royal Society of New Zealand32, 337e353.
Hustedt, F., 1985. The Pennate Diatoms. Koeltz Scientific Books, Hirschberg.Kattel, G.R., Augustinus, P.C., 2010. Cladoceran-inferred environmental change in an
Auckland maar lake, Lake Onepoto. New Zealand Journal of Geology andGeophysics 53, 30e41.
Knorr, G., Lohmann, G., 2003. Southern Ocean origin for the resumption of Atlanticthermohaline circulation during deglaciation. Nature 424, 532e536.
Korhola, A.A., Rautio, M., 2001. Cladocera and other branchipod crustaceans. In:Smol, J.P., Birks, H.J.B., Last, W.M. (Eds.), Zoological Indicators. Tracking Envi-ronmental Change Using Lake Sediments, vol. 4. Kluwer Academic Publishers,Dordrecht, The Netherlands 240 p.
Krammer, K., Lange-Bertalot, H., 1986. Süsswasserflora von Mitteleuropa. Bacillar-iophyceae i Teil Naviculaceae. Gustav Fischer Verlag, Stuttgart. 876 p..
Krammer, K., Lange-Bertalot, H., 1988. Süsswasserflora von Mitteleuropa. Bacillar-iophyceae ii Teil Bacillariaceae, Epithemiaceae, Surirellaceae. Gustav FischerVerlag, Stuttgart. 610 p.
Krammer, K., Lange-Bertalot, H., 1991a. Süsswasserflora von Mitteleuropa. Bacil-lariophyceae iii Teil Centrales, Fragilariaceae, Eunotiaceae. Gustav Fischer Ver-lag, Stuttgart. 576 p.
Krammer, K., Lange-Bertalot, H., 1991b. Süsswasserflora von Mitteleuropa. Bacil-lariophyceae iv Teil Achnanthaceae. Gustav Fischer Verlag, Stuttgart. 437 p..
Krammer, K., Lange-Bertalot, H., 2000. Bacillariophyceae: English and Frenchtranslation of the keys. In: Susswasserflora von Mitteleuropa, 2/5. SpektrumAkademischer Verlag, Heidelberg. 311 p.
Lowe, D.J., Shane, P.A.R., Alloway, B.V., Newnham, R.M., 2008. Fingerprints and agemodels for widespread New Zealand tephra marker beds erupted since 30,000years ago: a framework for NZ-INTIMATE. Quaternary Science Reviews 27,95e126.
Lucchini, F., Dinelli, E., Calanchi, N., 2003. Chemostratigraphy of Lago Albano sedi-ments (Central Italy): geochemical evidence of palaeoenvironmental changes inlate Quaternary. Journal of Paleolimnology 29, 109e122.
McGlone, M.S., 1983. Polynesian deforestation of New Zealand: a preliminarysynthesis. Archaeology in Oceania 18, 1e10.
McGlone, M.S., 1995. Lateglacial landscape and vegetation change and the YoungerDryas climatic oscillation in New Zealand. Quaternary Science Reviews 14,867e881.
McGlone, M.S., Wilmshurst., J.M., 1999. Dating initial Maori impact in New Zealand.Quaternary International 59, 17e26.
Meijering, M.P.D., 1983. On the occurrence of “Arctic” Cladocera with specialreference to those along the Strait of Belle Isle (Quebec, Labrador, NewFoundland). Internationale Revue der Gesamten Hydrobiologie 68, 885e893.
Meyers, P.A., 1994. Preservation of source identification of sedimentary organicmatter during and after deposition. Chemical Geology 144, 289e302.
Meyers, P.A., Lallier-Vergès, E., 1999. Lacustrine sedimentary organic matter recordsof Late Quaternary paleoclimates. Journal of Paleolimnology 21, 345e372.
Mix, A.C., Bard, E., Schneider, R., 2001. Environmental processes of the ice age: land,oceans, glaciers (EPILOG). Quaternary Science Reviews 20, 627e657.
Molloy, C., Shane, P.A., Augustinus, P.C., 2009. Eruption recurrence rates in a basalticvolcanic field based on tephra layers in maar sediments: implications forhazards in the Auckland volcanic field. Geological Society of America Bulletin121, 1666e1677.
Moore, P.D., Webb, J.A., Collinson, M.E., 1991. Pollen Analysis, second ed. BlackwellScientific, London. 216 p.
Mortlock, R.A., Froelich, P.N., 1989. A simple method for the rapid determination ofbiogenic opal in pelagicmarine sediments. Deep Sea Research 36, 1415e1426.
Nesbitt, H.W., Wilson, R.E., 1992. Recent chemical weathering of basalts. AmericanJournal of Science 292, 740e777.
Newnham, R.M., Lowe, D.J., 1991. Holocene vegetation and volcanic activity, Auck-land Isthmus, New Zealand. Journal of Quaternary Science 6, 177e193.
Newnham, R.M., Lowe, D.J., Giles, T., Alloway, B.V., 2007. Vegetation and climate ofAuckland, New Zealand, since ca. 32 000 cal. yr ago: support for an extendedLGM. Journal of Quaternary Science 22, 517e534.
Pepper, A.C., Shulmeister, J., Nobes, D.C., Augustinus, P., 2004. ENSO signals duringthe Last Glacial Maximum, the Antarctic Cold Reversal and the early Holocenefrom New Zealand. Geophysical Research Letters 31 (15), L15206. doi:10.1029/2004GL020236.
Pillans, B.J., 1990. New Zealand Quaternary stratigraphy: an overview. QuaternaryScience Reviews 10, 405e418.
Prokopenko, A.A., Karabanov, E.B., Williams, F.W., Kuzmin, M.I., Shackleton, N.J.,Crowhurst, S.J., Peck, J.A., Gvozdkov, A.N., King, J.W., 2001. Biogenic silica recordof the Lake Baikal response to climatic forcing during the Brunhes. QuaternaryResearch 55, 123e132.
Reid, M., 2005. Diatom-based models for reconstructing past water quality andproductivity in New Zealand lakes. Journal of Paleolimnology 33, 13e38.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., BronkRamsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M.,

P. Augustinus et al. / Quaternary International 253 (2012) 18e31 31
Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F.,Kromer, B., McCormac, F.G., Manning, S.W., Reimer, R.W., Richards, D.A.,Southon, J.R., Talamo, S., Turney, C., van der Plicht, J., Weyhenmeyer, C.E., 2009.IntCal09 and Marine09 radiocarbon age calibration curves, 0e50,000 years cal BP.Radiocarbon 51, 1111e1150.
Round, F.E., Crawford, R.M., Mann, D.G., 1990. The Diatoms: Biology andMorphology of the Genera. Cambridge University Press, Cambridge. 747 p.
Samson, C.R., Sikes, E.L., Howard, W.R., 2005. Deglacial paleoceanographic history ofthe Bay of Plenty, New Zealand. Paleoceanography 20 (PA4017). doi:10.1029/2004PA001088.
Sandiford, A., Newnham, R., Alloway, B.J., Ogden, J., 2003. A 28000e7600 cal yr BPpollen record of vegetation and climate change from Pukaki Crater, northernNew Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology 201, 235e247.
Schmidt, R., Kamenik, C., Lange-Bertalot, H., Klee, R., 2004. Fragilaria and Staurosira(Bacillariophyceae) from sediment surfaces of 40 lakes in the Austrian Alps inrelation to environmental variables, and their potential for palaeoclimatology.Journal of Limnology 63, 171e189.
Shane, P., 2000. Tephrochronology: a New Zealand case study. Earth ScienceReviews 49, 223e259.
Shane, P., Hoverd, J., 2002. Distal record of multi-sourced tephra in Onepoto basin,Auckland: implications for volcanic chronology, frequency and hazards. Bulletinof Volcanology 64, 441e454.
Shane, P., Sandiford, A., 2003. Paleovegetation of marine isotope stages 4 and 3 innorthern New Zealand and the age of the widespread Rotoehu tephra.Quaternary Research 59, 420e429.
Shulmeister, J., Goodwin, I., Renwick, J., Harle, K., Armand, L., McGlone, M.S.,Cook, E., Dodson, J., Hesse, P.P., Mayewski, P., Curran, M., 2004. The SouthernHemisphere westerlies in the Australasian sector during the last glaciationcycle: a synthesis. Quaternary International 118/119, 23e53.
Singer, C., Shulmeister, J., McLea, B., 1998. Evidence against a significant YoungerDryas cooling event in New Zealand. Science 281, 812e814.
Sonneman, J.A., Sincock, A., Fluin, J., Reid, M., Newall, P., Tibby, J., Gell, P., 2000. AnIllustrated Guide to Common Stream Diatom Species from Temperate Australia.Albury, Cooperative Research Centre for Freshwater Ecology. IdentificationGuide No. 33.
Street-Perrott, F.A., Ficken, K.J., Huang, Y., Eglinton, G., 2004. Late Quaternarychanges in carbon cycling on Mt Kenya, East Africa: an overview of the d13Crecord in lacustrine organic matter. Quaternary Science Reviews 23, 861e879.
Suggate, R.P., Almond, P., 2005. The Last Glacial Maximum (LGM) in western SouthIsland, New Zealand: implications for the global LGM and MIS2. QuaternaryScience Reviews 24, 1923e1940.
Tomlinson, A.I., Sansom, J., 1994a. Rainfall Normals for New Zealand for the Period1961 to 1990. National Institute of Water and Atmospheric Research,Wellington. 20 p.
Tomlinson, A.I., Sansom, J., 1994b. Temperature Normals for New Zealand for thePeriod 1961 to 1990. National Institute of Water and Atmospheric Research,Wellington. 18 p.
Turney, C.S.M., McGlone, M.S., Wilmshurst, J.M., 2003. Asynchronous climatechange between New Zealand and the North Atlantic during the last deglaci-ation. Geology 31, 223e226.
Van Dam, H., Mertens, A., Sinkeldam, J., 1994. A coded checklist and ecologicalindicator values of freshwater diatoms from the Netherlands. NetherlandsJournal of Aquatic Ecology 28, 117e133.
Vandergoes, M.J., Dieffenbacher-Krall, A.C., Newnham, R.M., Denton, G.H.,Blaauw, M., 2008. Cooling and changing seasonality in the Southern Alps, NewZealand during the Antarctic Cold Reversal. Quaternary Science Reviews 27,589e601.
Vos, P.C., de Wolf, H., 1988. Methodological aspects of paleo-ecological diatomresearch in coastal areas of the Netherlands. Geologie en Mijnbouw 67, 31e40.
Weckström, J., Korhola, A., 2001. Patterns in the distribution, composition anddiversity of diatom assemblages in relation to ecoclimatic factors in ArcticLapland. Journal of Biogeography 28, 31e45.
Whiteside, M.C., Williams, J.B., White, C.P., 1978. Seasonal abundance and pattern ofchydorid, Cladocera in mud and vegetative habitats. Ecology 59, 1177e1188.
Whiteside, M.C., Swindoll, M.R., 1988. Guidelines and limitations to cladoceranpalaeoecological interpretations. Palaeogeography, Palaeoclimatology, Palae-oecology 62, 405e412.
Williams, D.M., Round, F.E., 1987. Revision of the genus Fragilaria. Diatom Research2, 267e288.
Williams, P.W., King, D.N.T., Zhao, J.-X., Collerson, K.D., 2005. Late Pleistocene toHolocene composite speleothem 18O and 13C chronologies from South Island,New Zealand e did a global Younger Dryas really exist? Earth and PlanetaryScience Letters 230, 301e317.
Woodward, C.A., Shulmeister, J., 2005. A Holocene record of human induced andnatural environmental change from Lake Forsyth (Te Wairewa), New Zealand.Journal of Paleolimnology 34, 481e501.