large conductance ca2+-activated k+ (bk) channel: activation by ca2+ and voltage
TRANSCRIPT

385LATORRE & BRAUCHI Biol Res 39, 2006, 385-401Biol Res 39: 385-401, 2006 BR
Large conductance Ca2+-activated K+ (BK) channel:Activation by Ca2+ and voltage
RAMÓN LATORRE1 and SEBASTIAN BRAUCHI1, 2
1 Centro de Estudios Científicos, Valdivia, Chile.2 Universidad Austral de Chile, Valdivia, Chile.
ABSTRACT
Large conductance Ca2+-activated K+ (BK) channels belong to the S4 superfamily of K+ channels that includevoltage-dependent K+ (Kv) channels characterized by having six (S1-S6) transmembrane domains and apositively charged S4 domain. As Kv channels, BK channels contain a S4 domain, but they have an extra (S0)transmembrane domain that leads to an external NH2-terminus. The BK channel is activated by internal Ca2+,and using chimeric channels and mutagenesis, three distinct Ca2+-dependent regulatory mechanisms withdifferent divalent cation selectivity have been identified in its large COOH-terminus. Two of these putativeCa2+-binding domains activate the BK channel when cytoplasmic Ca2+ reaches micromolar concentrations,and a low Ca2+ affinity mechanism may be involved in the physiological regulation by Mg2+. The presence inthe BK channel of multiple Ca2+-binding sites explains the huge Ca2+ concentration range (0.1 μM-100 μM)in which the divalent cation influences channel gating. BK channels are also voltage-dependent, and all theexperimental evidence points toward the S4 domain as the domain in charge of sensing the voltage. Calciumcan open BK channels when all the voltage sensors are in their resting configuration, and voltage is able toactivate channels in the complete absence of Ca2+. Therefore, Ca2+ and voltage act independently to enhancechannel opening, and this behavior can be explained using a two-tiered allosteric gating mechanism.
Key terms: BK channel, Ca2+-binding sites, voltage dependence, allosteric models.
Corresponding author: Ramón Latorre, Centro de Estudios Científicos, Arturo Prat 514, Valdivia, Chile, E-mail: [email protected]
Received: July 22, 2005. Accepted: August 22, 2005
PROLOGUE: ON SQUIDS AND THE AMAZING
MONTEMAR LABORATORY
If you ask me (RL) why the laboratory inMontemar was so important in thedevelopment of biophysics in Chile, I wouldimmediately say: because the seniors weredoing good science, and they let the juniorsto do whatever they wanted. In this regard,our professors made it clear from the onsetof our research work that we were supposedto take one idea from the many floatingaround in the lab in those years and developthe experimental and theoretical frameworkin an absolutely independent manner.Freedom is a terrible thing when one isyoung and insecure, and learning to be onmy own was the first thing I was forced tomaster in Montemar. Our teachers were wiseenough and brave enough to give ussufficient intellectual ammo to make us
scientifically independent. In other words,all the scientists at Montemar followed thePeter Medawar’s dictum to the youngscientist apprentice: choose an importantproblem and become apprenticed to a seniorscientist. I fully agree with Perutz when hestates that “creativity in science, as in thearts, cannot be organized. It arisesspontaneously from individual talent”. Well-run laboratories, as was the case ofMontemar, can foster talent, but hierarchicalorganizations, inflexible bureaucratic rulesthat plague our universities, and mountainsof futile paperwork can kill it, as is usuallythe case in Chile.
The next thing I learned during my Ph.D.years at Montemar was that biology is aboveall an experimental field and that no theoryis as valuable as a well-done experiment.Actually, many experiments were needed,and the squid season was short, so you were
REVIEW

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401386
forced to work hard and efficiently andeven built your own equipment whennecessary (voltage clamps were notcommercially available at the time!). Thewhole lab environment forced you to realizethat “experimentation is a form of thinkingand that a wrong interpretation of anexperiment is forgivable, but anunrepeatable experimental result is not”(Medawar, 1979).
Paul Ehrlich, the father of immunology,used to say that scientists need the four Gs:Geschick, Geduld, Geld, und Glück (skill,patience, money, and luck). Science atMontemar was undertaken withconsiderable skill, patience and luck butvery little money; our advantage over therich laboratories of the United States wasthe squid (Dosidicus gigas). Biophysicsfirst became a Chilean specialty because“that’s where the squids were” as ChrisMiller stated in an interview that appearedin Science in 1995. Using the squid giantaxon, Mario Luxoro and his pupil EduardoRojas (“Guayo” to his colleagues andstudents) were the first to claim thatproteins were involved in the electricalexcitability of nerve (Rojas and Luxoro,1963). The axons of this monster, about onemillimeter in diameter, were, during the1960s, an attraction difficult to resist forscientists interested in nerve excitability.And this is another important reason whythe Montemar laboratory was so successful.Every summer, we were visited byscientists of the stature of Clay Armstrong,Ichigi Tasaki, Bob Taylor, and GerryEhrenstein. Clay, Gerry, and Guayo weremy first scientific idols, Clay because of hisamazing wit, Gerry because of the economyand precision of his thinking, and Guayobecause he was able to play Beethoven witha voltage-clamp set-up. Clay, Gerry andGuayo were the first to introduce me to theamazing field of ion channels. Thanks tothe squid, Montemar was a window to theworld, and Cecilia Hidalgo, PanchoBezanilla, and I had the opportunity (Glück)of doing postdocs in very good laboratoriesin the U.S. Thus, the “little ones” alsolearned at Montemar that if you have aunique biological preparation that enablesyou to understand a fundamental biological
process, you can use this knowledge to youradvantage in order to overcome poverty.
In Montemar, I realized that science isnot a quiet life. If you put together twoItalian descendants (Mario and Mitzy) inthe same space, you get an explosivemixture. Discussions about science, labspace, or politics were frequently heatedand always won by Fernando Vargasbecause during the course of a debate henever lost his temper, and his logic wasunbeatable. So, there was another lesson forus: scientists are generally passionatepeople who defend their territories and petideas as ferociously as bears. Above all,however, our teachers taught us that inMontemar the authority principle did notexist. If you were wrong, somebody wasthere to show you that you were mistaken,whether you were a graduate student,Luxoro, or the Pope himself. One of hischaracters in Fred Hoyle’s novel the BlackCloud remarks that scientists are alwayswrong, yet they always go on. What makesthem to continue? I think it is somethingthat was always latent in Montemar: thepassion for solving problems aesthetically.
Eduardo Rojas deserves a special spacein these remembrances. Guayo wasfundamental in my life as a scientist. Due tothe political situation in the country duringin the late 1960s, most of our professors leftMontemar to take different posts at theUniversidad de Chile, and Cecilia and Iwere left orphaned of advisers. During thesummer of 1969, Guayo adopted us, andthanks to his support (emotional andscientific), we were able to finish our Ph.D.theses. His generosity and kindness madepossible for me to be telling you my part ofthe Montemar adventure.
INTRODUCTION
When Mario Luxoro asked me (RL) towrite a chapter in this issue of BiologicalResearch dedicated to my teacher,colleague, and friend, Eduardo Rojas, Iimmediately thought that the best presentfor him would be to say something aboutthe ion channel that is closest to my heart.From the moment we discovered its

387LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
existence in a membrane preparation frommuscle T-tubule (Latorre et al., 1982), wewere confronted with a molecularPandora’s box: once opened, its electricallanguage left all of us bewitched. Others(e.g., Marty, 1981; Pallota et al., 1981)were as fascinated as we were with this“monster” of a single-channel conductanceclosed to the ceiling imposed by simplediffusion combined with an exquisite K+
selectivity. BK channels essentially areimpermeant to Na+ and conduct K+ 10- and200-fold more effectively than Rb+ and Cs+,respectively (large conductance channelswere not supposed to be so selective!)(Blatz and Magleby, 1984; Eisenman et al.,1986; Stefani et al., 1997). At the sametime, the channel was activated by voltageand cytoplasmic Ca2+. This latter propertyled Meech (1978) to hypothesize that thisconductance system was perfect l inkbetween cell metabolism and electricalactivity, and he was right on the mark.Because of its large conductance, thisvoltage and calcium-activated K+ channelwas christened “maxi K” (Latorre andMiller, 1983) or “BK” (for big K+; Blatzand Magleby, 1987).
Muscle contraction, neurosecretion,chromaffin cell electrical activity, and haircell tuning are some of the keyphysiological processes that require anincrease in cytoplasmic Ca2+ to develop.Most often, this Ca2+ increase is mediatedby the Ca2+ entry into the cells throughvoltage-dependent Ca2+ channels (VDCCs).The increase in internal Ca2+, however, alsoputs into action a negative feedback thatwill serve to stop or to dampen excitatoryphenomena induced by the opening ofVDCCs. This negative feedback appears asa consequence of the activation of BK(about 250 pS in 100 mM symmetrical KCl;Marty, 1981; Pallota et al., 1981; Latorre etal., 1982; for reviews see Latorre et al.,1989; McManus, 1991). Thus, BK channelshave the largest single-channel conductanceof all K+ selective channels. To ensuremaximum efficiency of the negativefeedback, BK channels functionally co-localize with VDCCs (Marrion and Tavalin,1998; Prakriya and Lingle, 1999). Despitebeing coded by a single gene (Slowpoke),
the diversity of BK channels is great.Regulatory β-subunits, splicing, andmetabolic regulation create this diversityfundamental to the adequate function ofmany tissues (Vergara et al., 1998; Latorreet al., 2000; Orio et al., 2002) In a bird’s-eye view of this fantastic molecularmachine, we will discuss its molecularproperties and how these propertiesdetermine the BK channel opening andclosing.
GROSS BK CHANNEL STRUCTURE AND THE CA2+-
SENSING ELEMENTS
The cloning of the BK channel fromDrosophila (Atkinson et al., 1991; Adelmanet al., 1992) showed that the BK channelwas a member of the S4 superfamilyencompassing voltage-dependent K+ (Kv),Na+ and Ca2+ channels. The gene coding forBK was called Slowpoke or Slo and, afterthe cloning and expression of Slo2 (Yuan etal., 2000; Yuan et al., 2003) and Slo3(Schreiber et al., 1998), was renamed Slo1.In the case of Kv channels, the channel-forming protein possesses sixtransmembrane domains (S1-S6) containingthe pore-forming domain S5-P-S6 and anS4 voltage-sensing element. Like Kvchannels, the BK channel is a tetramer(Shen et al., 1994); unlike Kv channels,however, the BK channel protein consistsof seven transmembrane domains (S0-S6)that lead to an exoplasmic N-terminus (Fig.1; Meera et al., 1997; Wallner et al., 1996;Toro et al., 1998). The large C-terminus(containing about twice as many aminoacids as S0-S6) has four hydrophobicdomains (S7-S10), and Salkoff’s groupidentified two molecular domains (S0-S8,“core” and S9-S10, “tail”) that, whenexpressed together, were able to producefunctional channels (Wei et al., 1994).
SEARCHING FOR THE HIGH AFFINITY CA2+-
BINDING SITES
Taking advantage of the fact that theDrosophila Slo (dSlo1) channel has adifferent Ca2+ sensitivity than the mouse
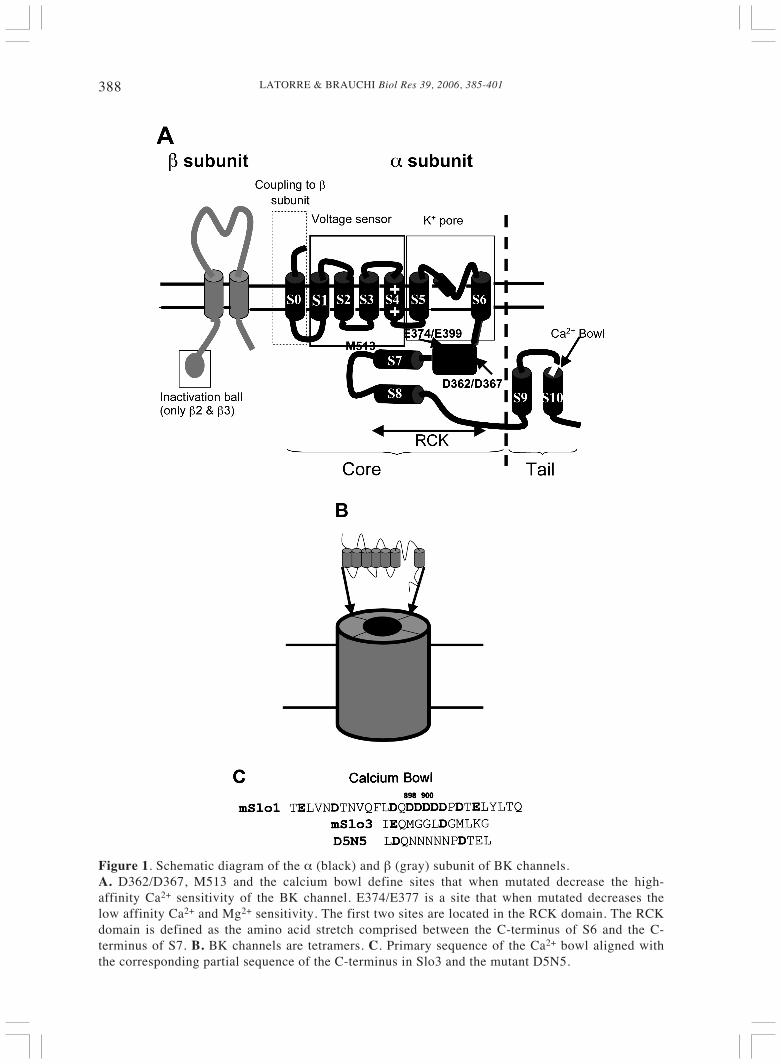
LATORRE & BRAUCHI Biol Res 39, 2006, 385-401388
Figure 1. Schematic diagram of the α (black) and β (gray) subunit of BK channels.A. D362/D367, M513 and the calcium bowl define sites that when mutated decrease the high-affinity Ca2+ sensitivity of the BK channel. E374/E377 is a site that when mutated decreases thelow affinity Ca2+ and Mg2+ sensitivity. The first two sites are located in the RCK domain. The RCKdomain is defined as the amino acid stretch comprised between the C-terminus of S6 and the C-terminus of S7. B. BK channels are tetramers. C. Primary sequence of the Ca2+ bowl aligned withthe corresponding partial sequence of the C-terminus in Slo3 and the mutant D5N5.

389LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
Slo (mSlo1) channel, Wei et al. (1994)showed that coexpression of mSlo coretogether with dSlo tail produced channelswith a Ca2+ sensitivity similar to that of thedSlo channel. These experiments providedthe first indication that the tail was playingthe role of the calcium-sensing element.Closer inspection of the tail showed thepresence of a domain consisting of 28amino acids containing nine acidic residuesincluding a string of five conservedaspartates residues, the “calcium bowl”(Fig. 1A, C). Partial deletion or pointmutations of the aspartates contained in thecalcium bowl produced BK channel thatwere less sensitive to Ca2+ - at the sameCa2+ concentration, the calcium bowlmutant conductance-voltage (G-V) curveswere right-shifted compared to the wild-type BK channel G-V curve (Schreiber andSalkoff, 1997; Bian et al., 2001; Braun andSy, 2001). If the Ca2+ bowl is disrupted bydeleting crucial aspartates, this high Ca2+
affinity regulatory site is lost, but themutant and the wild-type BK showed thesame sensitivity to Cd2+ ions. This findingprovided evidence of the presence of twodifferent Ca2+-binding sites; a site sensitiveto Ca2+ but not to Cd2+ and another site ableto bind with similar affinity Ca2+ and Cd2+
(Schreiber and Salkoff, 1997). Clearly, itwould be difficult to explain theobservation that BK activation is sensitiveto a range of Ca2+ concentrations that spansover four orders of magnitude on the basisof a single Ca2+-binding site (see Fig. 6).The Slo3 channel lacks the calcium bowl(Fig. 1C) and is not sensitive to Ca2+. Co-expression of mSlo1 core and mSlo3 tailgave origin to Ca2+-insensitive channels inthe range comprised between 0-10 μMinternal Ca2+, but the data presentedindicated that the chimeric Slo channelswere activated by Ca2+ at concentrations≥300 μM (Schreiber et al. , 1999).Interestingly, mSlo1 core-mSlo3 tailchannels opened at much lower voltagesthan mSlo1 at zero internal Ca2+
(conductance-voltage curves for thechimeric channel were shifted by about 60mV to the left along the voltage axis).These results suggest the presence of aCa2+-binding site different from the Ca2+
bowl and that, in the absence of Ca2+, theSlo1 tail inhibits voltage-dependent BKchannel gating, inhibition that is relieved byCa2+. This inhibition may help to keep thechannel closed in the resting cell (lowinternal Ca2+ and negative membranepotential). A detailed single-channelanalysis confirmed the results of Schreiberet al. (1999) and suggested that in additionto playing a role as a Ca2+ sensing domain,the tail domain also modulates the gatingand conductance properties of BK channels(Moss and Magleby, 2001).
In all the experiments described above,Ca2+-binding sites were inferred from theeffects of Ca2+ on channel activation (i.e.,changes in the BK open probability). Bianet al. (2001) measured direct binding ofradioactive calcium to a COOH-terminusfragment of the Drosophila BK containingthe calcium bowl and showed that mutatingthe aspartates to asparagines (D5N5 mutant;Fig. 1C) was able to reduce the Ca2+-binding affinity by about 60%. This resultdemonstrates a direct correlation betweenCa2+ binding and Ca2+ sensitivity forchannel activation (see also Braun and Sy,2001). The D5N5 mutant expressed in cellsexhibits a lower Ca2+ sensitivity foractivation and reduces the dSlo channel’sHill coefficient 2-fold, as if one binding siteper monomer is lost in the D5N5 mutant.The most economic way to account forthese results is to assume the existence oftwo distinct Ca2+-binding sites per channelmonomer (Bian et al., 2001; cf. Schreiberand Salkoff, 1997). Bao et al. (2004)explored the relative importance of thedifferent acidic amino acids contained inthe calcium bowl using alanine scanningmutagenesis of 20 residues contained in thisdomain. These experiments were done inthe background mutant, M513I; a mutantthat eliminates one of the highCa2+sensitivity regulatory sites (see below)allowing determining the isolated Ca2+-binding properties of the calcium bowl.They found that mutation to alanines of twocritical aspartates (D898 and D900; Fig.1C) has the greatest effect on the change inthe G(V) curve half voltage (ΔV0.5) uponaddition of Ca2+. The point mutations alsowere able to decrease binding of 45Ca by

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401390
about 50% to a peptide composed of afusion protein consisting of glutathione-S-transferase and a 207-amino acid part of themSlo1 tail that includes the calcium bowl.These results indicate that there are “hot”residues in the calcium bowl and gavefurther support to the hypothesis that thisdomain is the one of the high affinity sitesthat couples Ca2+ binding to channelopening. (1)
The molecular origin of the remainingBK Ca2+ sensitivity after neutralization ofmost of the acidic residues present in theCa2+ bowl was elucidated by mutatingnegatively charged residues outside theCa2+ bowl and, in particular, in theregulator of conductance for K+ (RCK;Jiang et al., 2001; Fig. 1-3). The RCKdomain in the BK channel was unveiled byMacKinnon’s group (Jiang et al., 2001) bymultiple sequence alignment of the BKchannel with prokaryotic K+ channels andother proteins known to possess the RCKdomain structure. The structure of the RCKdomain of a six transmembrane domain K+
channel from E. coli (Fig. 2A) solved at 2.4Å resolution has a Rossmann-fold topology,a very common structural motif found inenzymes and ligand-binding proteins.Rossmann-fold secondary structures areorganized into two linked β−α−β−α−βunits (see Figs. 2B and 3A) and were firstidentified in a number of NAD+-dependentdehydrogenases (Darby and Creighton,1993).
The hypothesis that BK channels containa RCK domain was put to test by disturbinga well-conserved salt bridge, not commonlypresent among Rossmann-fold proteins(K448-D481 in hSlo1; Fig. 3B). If eitherposition is mutated (e.g., K448D orD481K), the channel becomes less Ca2+sensitive. However, the double mutantK448D/D481K recovers the Ca-sensitivityshown by the wild-type BK channel.Therefore, these results strongly suggest thepresence of a salt bridge as predicted fromthe E.coli K+ channel RCK structure andsupport the hypothesis that the BK channelcontains an RCK domain on its C-terminus.
The double mSlo1 RCK mutant D362A/D367A (Fig. 3C) produced a markedreduction in the Ca2+ sensibility of the BK
channel, and the mutant D362A/D367A-5D5N is still gated by voltage, with avoltage dependence similar to that of thewild type, but Ca2+ is unable to activate thechannel at [Ca2+] < 10-3 M (Xia et al.,2002). Another mutation, M513I, a residueimmediately following the S7 domain (Figs.2B and 3B), together with a deletion mutantthat eliminates most of the aspartatescontained in the calcium bowl (Δ896-903),similar to the D362A/D367A-5D5N mutant,was shown to completely remove highaffinity responsiveness to Ca2+ (Bao et al.,2002). The behavior of the M513I andΔ896-903 mutants was analyzed in terms ofallosteric models (Cox and Aldrich, 2000;Rothberg and Magleby, 2000; Horrigan andAldrich, 2002; see below) that allow for thequantitative determination of thecontribution of the different mutations tothe BK channel activation. The mainfinding of this study was that in the absenceof an applied voltage, each regulatory sitecontributes almost equally to the freeenergy difference between open and closedstates. The estimated dissociation constantswere in the micromolar range. Thus,although no direct evidence supports theidea that D362/D367 or M513 form part ofa structure able to coordinate Ca2+ ions, thedata of Xia et al. (2002) and Bao et al.(2002) suggest that the BK channelcontains at least two high affinity Ca2+-binding sites. The results of Schreiber andSalkoff (1997) pointed towards theexistence of a second site able to bind Cd2+
and gave a clear indication of the existenceof a second Ca2+-binding site with differentdivalent cation selectivity. Oberhauser et al.(1988), on the other hand, found that theBK channel can be activated by a series ofdivalent cations including Sr2+, Cd2+, Mn2+,Fe2+, and Co2+. Taking advantage of theseobservations, Zeng et al. (2005) performeda series of elegant divalent cationselectivity experiments using the D362A/D367A mutant to investigate the selectivityof the Ca2+ bowl or the D5N5 mutant todetermine the divalent cations able toactivate the channel by interacting with theD362/D367 regulatory mechanism. Theresults indicated that the calcium bowl wasable to selectively bind Ca2+ and Sr2+, while

391LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
Figure 2. mSlo1 RCK secondary structure prediction.A. Sequence alignment used for the homology modeling. mSlo1 C-terminus primary sequence usedwas directly taken from the protein data bank (sequence number NP_034740). AvailableKCH_ECOLI-RCK domain coordinates (P31069, 1ID1) obtained previously by X-raycrystallography (Jiang et al., 2001) were used as template. The alignment used for the homologymodeling was performed using LIALIGN. There is ~20% identity between these proteins, but theyare similar regarding their secondary structure. The homology modeling was performed usingModeler 7v7 (Marti-Renom et al., 2000). LOOPS routine was used for the unaligned sequences.The models that show the lowest energy profiles were selected. WATH IF web interface was usedto check the obtained structures and remove bumps. B. The secondary structure prediction of thehSlo1 RCK domain was performed using JPred (Professor Barton group, Dundee University,Scotland, UK) and PredictProtein Server (Columbia University Bioinformatics Center) (Karplus,2003; Rost, 2003). Secondary structure prediction was used to evaluate the structural similaritybetween RCK-Domain from E.coli 6TM channel (KCH_ECOLI) and mSlo1 RCK. Previousalignment between these two sequences was carried out by Jiang et al. (2001). Secondary structureprediction is marked as E (Strand) and H (Helix). The order of Rossmann structures β−α−β ispredicted for mSlo1 from βA through αD, after this point, the a helices from Jiang et al. (2001)original structure are conserved but there are no more predicted beta formations. Moreover, originalβE in Jiang et al. (2001) structure was replaced by a α helix in our prediction, named αX(highlighted). Buried (B) amino acids also are predicted and indicate that the hydrophobic segmentS7 is buried almost completely. Important charges involved in calcium sensitivity are highlightedwith red asterisks. Residues forming salt bridges are marked with dotted lines. Notice that E374 ispredicted to form a salt bridge with H350. Solid line over the conserved sequence IMRVI, shows asequence involved in the interaction between domains.
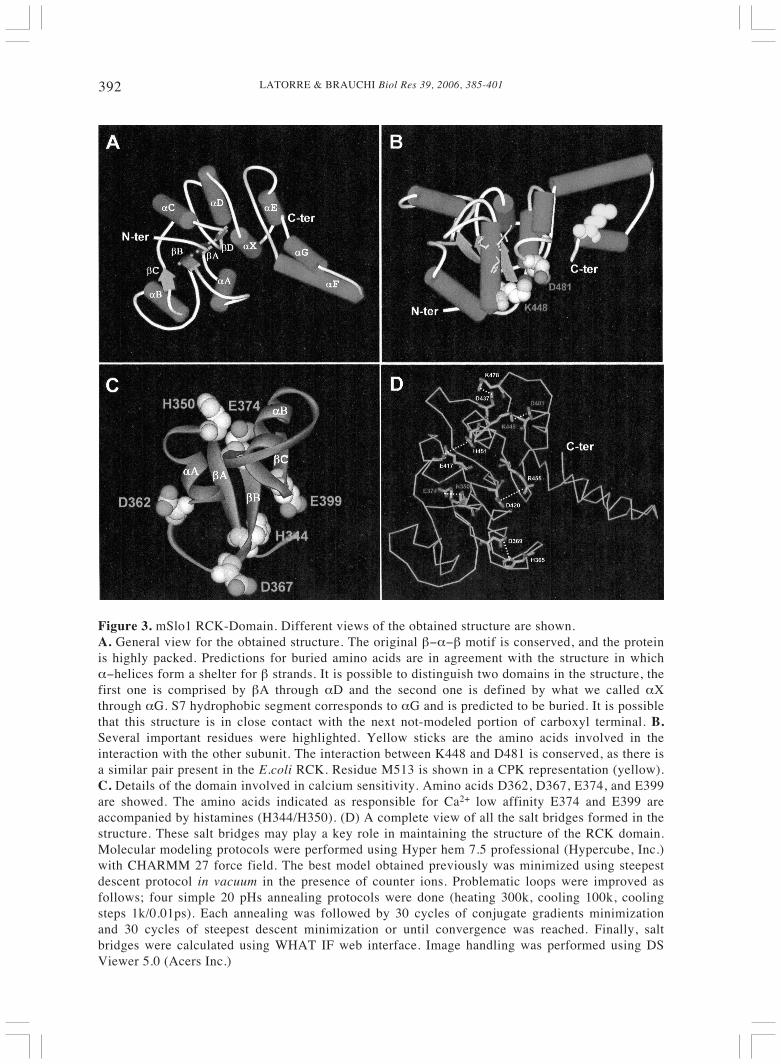
LATORRE & BRAUCHI Biol Res 39, 2006, 385-401392
Figure 3. mSlo1 RCK-Domain. Different views of the obtained structure are shown.A. General view for the obtained structure. The original β−α−β motif is conserved, and the proteinis highly packed. Predictions for buried amino acids are in agreement with the structure in whichα−helices form a shelter for β strands. It is possible to distinguish two domains in the structure, thefirst one is comprised by βA through αD and the second one is defined by what we called αXthrough αG. S7 hydrophobic segment corresponds to αG and is predicted to be buried. It is possiblethat this structure is in close contact with the next not-modeled portion of carboxyl terminal. B.Several important residues were highlighted. Yellow sticks are the amino acids involved in theinteraction with the other subunit. The interaction between K448 and D481 is conserved, as there isa similar pair present in the E.coli RCK. Residue M513 is shown in a CPK representation (yellow).C. Details of the domain involved in calcium sensitivity. Amino acids D362, D367, E374, and E399are showed. The amino acids indicated as responsible for Ca2+ low affinity E374 and E399 areaccompanied by histamines (H344/H350). (D) A complete view of all the salt bridges formed in thestructure. These salt bridges may play a key role in maintaining the structure of the RCK domain.Molecular modeling protocols were performed using Hyper hem 7.5 professional (Hypercube, Inc.)with CHARMM 27 force field. The best model obtained previously was minimized using steepestdescent protocol in vacuum in the presence of counter ions. Problematic loops were improved asfollows; four simple 20 pHs annealing protocols were done (heating 300k, cooling 100k, coolingsteps 1k/0.01ps). Each annealing was followed by 30 cycles of conjugate gradients minimizationand 30 cycles of steepest descent minimization or until convergence was reached. Finally, saltbridges were calculated using WHAT IF web interface. Image handling was performed using DSViewer 5.0 (Acers Inc.)

393LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
regulation defined by D362/D367 can beactivated by Ca2+, Sr2+, and Cd2+. A third,low affinity mechanism (discussed below),can be put into action by Ca2+, Sr2+, Cd2+,Mn2+, Ni2+, and Co2+. In the absence ofbinding assays like those performed withBK carboxyl terminus fusion proteins (Bianet al., 2001; Braun and Sy, 2001), thereexists the possibility, however, that all theresults are due to coincidental allostericeffects on Ca2+ binding to a region that liesoutside the RCK region. Independent ofthese considerations, the unique divalentcation selectivity of the different BKregulatory mechanisms supports theexistence of at least three distinct divalentcation binding sites and that the sites actindependently of each other.
Although rooted in solid experimentaldata, this picture of a BK channelcontaining all his Ca2+-sensing machinerylocated in the carboxyl terminal, needsfurther confirmation both from binding andstructural studies, in particular, in view ofthe report by Piskorowski and Aldrich(2002), who claimed that a channel lackingthe calcium bowl and RCK domain can stillbe activated by Ca2+. However, byconstructing chimeric channels in which thecarboxyl termini between the Slo1 channel(Ca2+ activated) and the Slo3 (pHdependent) were switched, Lingle’s groupshowed that the C-terminus of Slo1 is arequisite for channel Ca2+ sensitivity (Xiaet al., 2004). The chimeric channel Slo3-C-teminus Slo1 was activated by Ca2+, and thechimeric channel Slo1-C-terminus Slo3induced pH-sensitive currents.
LOW AFFINITY CA2+-BINDING SITES AND THE
MECHANISM OF MG2+ ACTIVATION OF BK
Golowash et al. (1986) found thatmillimolar amounts of Mg2+ were able topotentiate BK channel activation by Ca2+
and to increase the Hill coefficientdescribing the BK Ca2+ activation curves.Other divalent cations, such as Cd2+, Mn2+ ,Fe2+, Co2+, and Ni2+, also were able toenhance activation and increase the Hillcoefficient of channels already activated byCa2+ (Oberhauser et al., 1988). Recently, it
was discovered that most of these cationsare able to activate the BK channel in theabsence of Ca2+ by binding to a low affinitysite (Shi and Cui, 2001; Zhang et al., 2001;Shi et al., 2002; Zeng et al., 2005).
Notice in Fig. 4A, that Ca2+ shift theprobability of opening (Po) vs voltagecurves towards the left along the voltageaxis and that the data is well fitted by usinga Boltzmann function of the type:
(1)
where z describes the channel voltagedependence, F is Faraday’s constant, V isthe applied voltage, and V0.5 , the voltage atwhich Po = 0.5. A convenient way ofdetermining the ability of Ca2+ to activatethe BK channel is by plotting the V0.5 as afunction of Ca2+ concentration (Fig. 4B)because is directly related with the freeenergy, ΔG to open the channel since
(2)
where the values of z obtained from the fit ofthe data shown in Figure 4A to a Boltzmannfunction are given in Fig. 4C. This type ofplot was extensively used in thecharacterization of BK channels and gavethe first hint of the existence of a lowaffinity Ca2+ site (Wei et al., 1994; Cui et al.,1997). Fig. 4B shows that V0.5 shifts to morenegative voltages as the Ca2+ concentrationis increased, but above 10 μM, there is atendency towards saturation, suggesting thata new binding site has began to play a role inBK channel activation. This low affinity siteis unselective (see above) and Shi and Cui(2001) and Zhang et al. (2001) showed thatMg2+ was able to produce similar ΔV0.5 inthe presence or in the absence of Ca2+. Inother words, Mg2+ activates BK channelsindependently of Ca2+. Mg2+ does not affectthe activation but slows down thedeactivation kinetics, suggesting that it bindspreferentially to the open state. The possiblelocation of the Mg2+ regulatory domain wasrevealed by co-expressing Slo1 core andSlo3 tail. These chimeric channels, which donot contain the Ca2+ bowl, are still sensitive
RTVVzFo eP /)( 5.01
1−−+
=
ΔG zFV= 0 5.
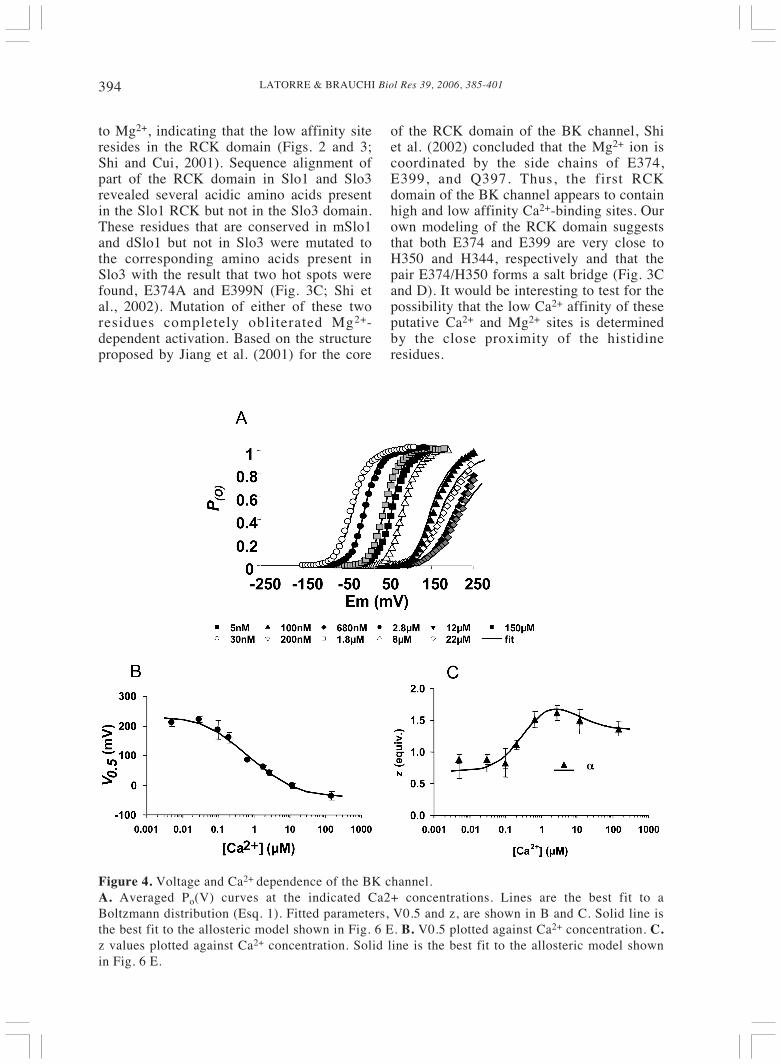
LATORRE & BRAUCHI Biol Res 39, 2006, 385-401394
to Mg2+, indicating that the low affinity siteresides in the RCK domain (Figs. 2 and 3;Shi and Cui, 2001). Sequence alignment ofpart of the RCK domain in Slo1 and Slo3revealed several acidic amino acids presentin the Slo1 RCK but not in the Slo3 domain.These residues that are conserved in mSlo1and dSlo1 but not in Slo3 were mutated tothe corresponding amino acids present inSlo3 with the result that two hot spots werefound, E374A and E399N (Fig. 3C; Shi etal., 2002). Mutation of either of these tworesidues completely obliterated Mg2+-dependent activation. Based on the structureproposed by Jiang et al. (2001) for the core
of the RCK domain of the BK channel, Shiet al. (2002) concluded that the Mg2+ ion iscoordinated by the side chains of E374,E399, and Q397. Thus, the first RCKdomain of the BK channel appears to containhigh and low affinity Ca2+-binding sites. Ourown modeling of the RCK domain suggeststhat both E374 and E399 are very close toH350 and H344, respectively and that thepair E374/H350 forms a salt bridge (Fig. 3Cand D). It would be interesting to test for thepossibility that the low Ca2+ affinity of theseputative Ca2+ and Mg2+ sites is determinedby the close proximity of the histidineresidues.
Figure 4. Voltage and Ca2+ dependence of the BK channel.A. Averaged Po(V) curves at the indicated Ca2+ concentrations. Lines are the best fit to aBoltzmann distribution (Esq. 1). Fitted parameters, V0.5 and z, are shown in B and C. Solid line isthe best fit to the allosteric model shown in Fig. 6 E. B. V0.5 plotted against Ca2+ concentration. C.z values plotted against Ca2+ concentration. Solid line is the best fit to the allosteric model shownin Fig. 6 E.

395LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
ACTIVATION BY VOLTAGE AND THE VOLTAGE
SENSOR
Despite the fact that it contains an S4domain, the molecular nature of the BKchannel voltage-dependence remainedunclear until it was unequivocallydemonstrated that depolarizing voltages areable to activate the BK channel in theabsence of Ca2+ (Pallota, 1985; Meera et al.,1996; Cui et al., 1997; Cox et al., 1997;Stefani et al., 1997; Horrigan and Aldrich,1999; Rothberg and Magleby, 2000;Nimigean and Magleby, 2000; Taludker andAldrich, 2000). One of the first observationsin support of the presence of an integralvoltage sensor in the BK channel-formingprotein is depicted in Figure 4B. Figure 4Bclearly shows a region comprised between 5and about 100 nM Ca2+, where the V0.5 isindependent of the Ca2+ concentration(Meera et al., 1996). Moreover, in this Ca2+
concentration interval, the channel can bemaximally activated by voltage, indicatingthat in the absence of Ca2+, voltages highenough can increase the Po to its maximumvalue (Cui et al., 1997).
In voltage-dependent channels,membrane depolarization promotes the
displacement of the charged residuescontained in the S4-inducing gatingcurrents (Armstrong and Bezanilla, 1973;Schneider and Chandler, 1973; Keynes andRojas, 1974; Yang and Horn, 1995;Aggarwal and MacKinnon, 1996; Seoh etal., 1996; Larsson et al., 1996; Yang et al.,1996; Yusaf et al., 1996; Ahern and Horn,2004). Gating currents in BK channels weredetected by Stefani et al., (1997) in theabsence of Ca2+ and showed that like theG(V) curves, the gating charge-voltage(Q(V)) curves were left-shifted in thepresence of Ca2+. The maximum number ofgating charges per channel, obtained bydividing the limiting charge (Qmax) by thenumber of channels in the patch, was about4, much less than those obtained in the caseof the voltage-dependent Shaker K+ channel(12-14 electronic charges; Schoppa et al.,1992; Seoh et al., 1996; Aggarwal andMacKinnon, 1996; Noceti et al., 1996). Thereason for this large difference in chargeper channel between BK and Shaker is stillunclear, since a primary alignment of hSloand Shaker charged domains S2, S3 and S4shows that most of the negative charges inS2 and S3 and positive charges in S4 areconserved (Fig. 5).
Figure 5. Alignment of the S2, S3 and S4 transmembrane domains of the human (hSlo) and ShakerK+ channel. Horizontal bars denote the transmembrane segments in both channels. Numbersindicate the position of the far right residues in the primary sequence of their respective proteins.Amino acids in bold are the charged residues conserved in Kv channels and hSlo channels.

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401396
In Shaker, four arginines (R362, R365,R368, and R371) contribute to the gatingcharge (Seoh et al., 1996; Aggarwal andMacKinnon, 1996) and Bezanilla’s grouppresented elegant evidence that these fourcharges move the entire electric field(Bezanilla, 2000; Starace and Bezanilla,2001; Starace and Bezanilla, 2004). In hSlo,three of these arginines are conserved(R207, R210, and R213) but only two ofthem (R210 and R213) were found to beable to alter the maximal slope of the Po(V)relationship (Diaz et al., 1998; cf., Cui andAldrich, 2000). More recently, Ma andHorrigan (2005) reported that charge D153in S2 and R213 in S4 reduce the maximumvoltage dependence of Po. (2). If R213 isthe only gating charge, it should move allthe way across the electric field (one chargeper channel subunit). Considering that inKv channels the voltage sensors structure(S1-S4) are essentially self-contained,independent domains inside the membrane(Long et al. , 2005a, b) and that BKchannels contain an extra transmembranedomain (S0), i t is probable that thestructure of the voltage sensor of the BKchannel will prove to be slightly differentfrom that of Kv channels. The clue of thelarge difference in gating changes betweenBK and Kv channels may lie in structuraldifferences in the voltage sensor.
EXPLAINING BK CHANNEL ACTIVITY USING
ALLOSTERIC MODELS
In this section, I will emphasize some of thehighlights of the two-tiered allosteric modelthat is commonly used to explain the BKchannel activity. For details, the readershould consult the excellent review byMagleby (2003). In the absence of Ca2+,Nimigean and Magleby (2000) andTaludker and Aldrich (2000) found 4-5exponential components in the dwell timedistribution for the closed states and 2-3components in the dwell time distributionfor the open states for the BK channel.Linear models in which only one pathwayled to the open states were excluded sinceMcManus et al. (1985) found an inverserelationship between the duration of
adjacent open and closed intervals: shorterclosed intervals are preferentially adjacentto longer open intervals and longer closedintervals. Given the large number of closedand open states, the correlation betweenadjacent intervals and the fact that thechannel is a tetramer, in the absence ofCa2+, BK channel gating is consistent withthe 10-state model indicated in Fig. 6A.One of the predictions of this model is thateven in the absence of voltage sensoractivation, described by the equilibriumconstant J, the channel can open throughthe reaction described by the equilibriumconstant L . (3) This prediction wasconfirmed by Horrigan et al. (1999), whofound that even in the absence of Ca2+ andat very negative voltages, the channel canopen with a very low, but measurable, Po(about 10-6) and a low voltage dependence,zL, not related to the voltage sensors. Onthe other hand, a detailed analysis of thegating currents in the absence of Ca2+
suggests a two-state model, resting-activated (R-A) suffices to explain thevoltage sensor movement (Horrigan andAldrich, 1999). The simple behavior of thegating currents, Q(V) curves which are welldescribed by a Boltzmann function, themonoexponential kinetics of the fastcomponent of the gating current, and thelack of a gating current rising phase alsoare consistent with the kinetic modelproposed in Fig. 6A in which the voltagesensors act independently.
In the allosteric model described inFigure 6A, for each voltage sensoractivated, the equilibrium constant forchannel opening, L, is multiply by anallosteric factor D, so the opening processis facilitated as more voltage sensors areactivated. The observation that even whenall voltage sensor are resting, Po can beincreased by augmenting intracellular Ca2+
(Horrigan and Aldrich, 2002) is the basisfor postulating the allosteric kinetic modeldepicted in Figure 6B under theassumption that there is only one Ca2+-binding site per channel subunit. In thiscase, for each Ca2+-binding site occupiedthe equilibrium constant L is multiply byan allosteric factor C. Figures 5A and Bdefine the key feature of BK channels:
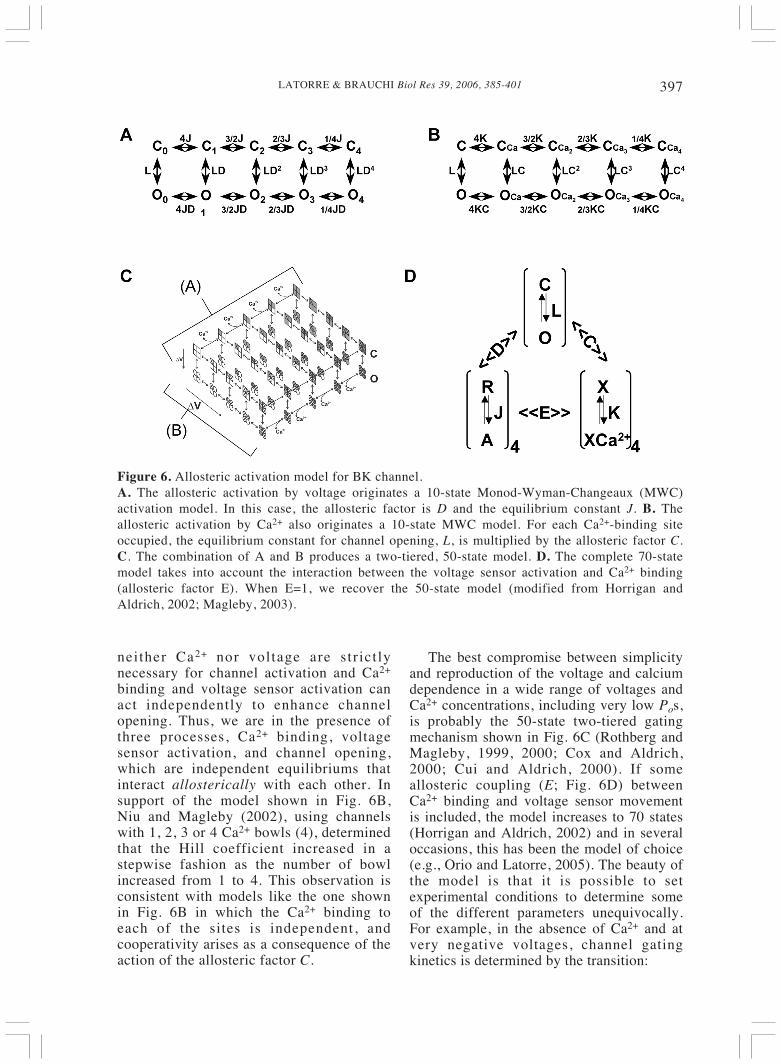
397LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
neither Ca2+ nor voltage are str ict lynecessary for channel activation and Ca2+
binding and voltage sensor activation canact independently to enhance channelopening. Thus, we are in the presence ofthree processes, Ca2+ binding, voltagesensor activation, and channel opening,which are independent equilibriums thatinteract allosterically with each other. Insupport of the model shown in Fig. 6B,Niu and Magleby (2002), using channelswith 1, 2, 3 or 4 Ca2+ bowls (4), determinedthat the Hill coefficient increased in astepwise fashion as the number of bowlincreased from 1 to 4. This observation isconsistent with models like the one shownin Fig. 6B in which the Ca2+ binding toeach of the sites is independent, andcooperativity arises as a consequence of theaction of the allosteric factor C.
Figure 6. Allosteric activation model for BK channel.A. The allosteric activation by voltage originates a 10-state Monod-Wyman-Changeaux (MWC)activation model. In this case, the allosteric factor is D and the equilibrium constant J. B. Theallosteric activation by Ca2+ also originates a 10-state MWC model. For each Ca2+-binding siteoccupied, the equilibrium constant for channel opening, L, is multiplied by the allosteric factor C.C. The combination of A and B produces a two-tiered, 50-state model. D. The complete 70-statemodel takes into account the interaction between the voltage sensor activation and Ca2+ binding(allosteric factor E). When E=1, we recover the 50-state model (modified from Horrigan andAldrich, 2002; Magleby, 2003).
The best compromise between simplicityand reproduction of the voltage and calciumdependence in a wide range of voltages andCa2+ concentrations, including very low Pos,is probably the 50-state two-tiered gatingmechanism shown in Fig. 6C (Rothberg andMagleby, 1999, 2000; Cox and Aldrich,2000; Cui and Aldrich, 2000). If someallosteric coupling (E; Fig. 6D) betweenCa2+ binding and voltage sensor movementis included, the model increases to 70 states(Horrigan and Aldrich, 2002) and in severaloccasions, this has been the model of choice(e.g., Orio and Latorre, 2005). The beauty ofthe model is that it is possible to setexperimental conditions to determine someof the different parameters unequivocally.For example, in the absence of Ca2+ and atvery negative voltages, channel gatingkinetics is determined by the transition:

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401398
LC↔O
and
(3)
as L << 1, Po ~ L = Loexp(zLFV/RT). Thus,under these experimental conditions, we areable to determine two parameters: Lo andzL. This exercise allows us to arrive atanother important conclusion: for the BKchannel, the limiting slope is actuallydetermined by the lesser voltage-dependenttransition and does not reflect the voltagesensor charge effectively coupled tochannel activation (see Almers, 1978; Siggand Bezanilla, 1997). The existence of twoor maybe three Ca2+-binding sites withdifferent affinities (see above) makes thepicture more complicated, however, andraises almost exponentially the number ofstates in a model.
SOME INFERENCES ABOUT THE MOLECULAR
NATURE OF CA2+ ACTIVATION
Jiang et al. (2002) determined the crystalstructure of the Ca2+-bound openMethanobacterium thermoautotrophicumK+ (MthK) channel. The structure showsthat each of the four channel subunitscontributes with two RCK domains forminga gating ring where two Ca2+ ions bind persubunit. The site behaves as the low Ca2+
affinity binding site in BK, since millimolaramounts of Ca2+ are necessary to activatethe MthK channel. Jiang et al. (2002) putforward a model in which Ca2+ bindingpromotes displacement mediated by aflexible interface of the two RCK domainsexpanding the diameter of the gating ring;this expansion is used to mechanically openthe pore to an open channel. Whether thismechanism applies to the BK channel isunclear, but the presence of RCK domainsin BK channels is undeniable (Jiang et al.,2001; Figs. 2 and 3). Actually two RCKdomains have been proposed to be presentin BK channels, which can act as a gatingring similar to that of MthK channels (e.g.,
Zeng et al., 2005), but evidence for thesecond RCK domain is lacking.
Niu et al. (2004) studied the effect ofchanging the length of the linker joining theS6 with the RCK domain. The authors use asimple and elegant model in which thecoupling could be explained by using aspring-like model. They found thatshortening the linker increase channelactivity and increasing linker lengthdecreases channel activity. Theseobservations suggest that, in the absence ofCa2+, the linker-RCK (gating ring?)complex behaves as a passive springattached to the channel gates. This linker-gating complex in the absence of Ca2+
applies force to the gates, an observationconsistent with the finding that compared tothe Slo3 tail, the Slo1 tail inhibits channelopening. Ca2+ binding, as in the case of theMthK channel, transform the passive springinto a force generating that uses the freeenergy of Ca2+ binding to pull open the S6helices to permit ion conduction.
ACKNOWLEDGEMENTS
We thank all the members of the LatorreLaboratory for their enthusiasm and muchdiscussion over the years about theworkings of ion channels. This work wassupported by grants from the FondoNacional de Investigación Científica yTecnológica (FONDECYT 103-0830 to R.Latorre and FONDECYT 104-0254 to F.González), the Human Frontiers in ScienceProgram. S. Brauchi was recipient of aPh.D. fellowship from the ComisiónNacional de Ciencia y Tecnología. TheCentro de Estudios Científicos is aMillennium Institute and is funded in partby a grant from Fundación Andes.
NOTES
1. The correlation between the DV0.5induced by the mutations and the effect on45Ca binding was lost when the mutationD899A was tested. This mutation has noeffect on DV0.5 but a large effect on the45Ca-binding signal. Bao et al. (2004) have
111
−+=
+=
LCOOPo

399LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
argued that the structure of the calciumbowl in the gel-overlay assay may take aconformation that is different to the oneadopted in its native conformation.
2. In Shaker, the equivalent negativecharge E283 does not contribute to the totalgating charge but E293 does (Sheo et al.,1996); in the equivalent position of E293,hSlo contains a tyrosine (Y163).
3. Voltage sensor activation is describedby the equilibrium constant J =J(0)exp(zjF(V - Vh(J)/RT), where J(0) isthe equilibrium constant at zero voltage; zj,the sensor charge; and Vh(J), the halfactivation voltage for each sensor. L =L(0)exp(zLFV/RT), where zL is the voltagedependence of L and L(0) is the equilibriumconstant for channel opening with the thevoltage sensor in resting state and no Ca2+
bound at zero voltage.4. The Ca2+ bowl of mSlo1 was
disrupted by deleting aspartates 965 and966, and the number of bowls per channeldetermined by mixing TEA-insensitivesubunits with bowl-disrupted subunits.Stoichiometry is obtained from the single-channel current amplitudes (see also Shenet al., 1994).
REFERENCES
ADELMAN JP, SHEN KZ, KAVANAUGH MP, WARRENRA, WU YN, LAGRUTTA BOND ACT, NORTH RA(1992) Calcium-activated potassium channelsexpressed from cloned complementary DNAs. Neuron9: 209-216
AGGARWAL SK, MACKINNON R (1996) Contributionof the S4 segment to gating charge in the Shaker K+
channel. Neuron 16: 1169-1177AHERN A, HORN R (2004) Specificity of charge-carrying
residues in the voltage sensor of potassium channels. JGen Physiol 123: 205-216
ALMERS W (1978) Gating currents and charge movementsin excitable membranes. Rev Physiol BiochemPharmacol 82: 96-190
ARMSTRONG CM, BEZANILLA F (1973) Currentsrelated to movement of gating particles of sodiumchannels. Nature 242: 459-461
ATKINSON NS, ROBERTSON GA, GANETZKY B(1991) A component of calcium-activated potassiumchannels encoded by the Drosophila slo locus. Science253: 551-555
BAO L, KALDANY C, HOLMSTRAND EC, COX DH(2004) Mapping the BKCa channel’s “Ca2+ bowl”: Sidechains essential for Ca2+ sensing. J Gen Physiol 123:475-489
BAO L, RAOIN AM, HOLMSTRAND EC, COX DH(2002) Elimination of the BKCa channel’s high-affinityCa2+ sensitivity. J Gen Physiol 120: 173-189
BEZANILLA F (2000) The voltage sensor in voltage-dependent ion channels. Physiol Rev 80: 555-592
BIAN S, FAVRE I, MOCZYDLOWSKI E (2001) Ca2+-binding activity of a COOH-terminal fragment of theDrosophila BK channel involved in Ca2+-dependentactivation. Proc Natl Acad Sci USA 98: 4776-4781
BLATZ AL, MAGLEBY, KL (1984) Ion conductance andselectivity of single calcium-activated potassiumchannels in cultured rat muscle. J Gen Physiol 84: 1-23
BLATZ AL, MAGLEBY KL (1987) Calcium-activatedpotassium channels. Trends Neurosci 10: 463-467
BRAUN AF, SY L (2001) Contribution of potential EFhand motifs to the calcium-dependent gating of amouse brain large conductance calcium-sensitive K+
channel. J Physiol 533: 681-695COX DH, CUI J, ALDRICH RW (1997) Allosteric gating
of a large conductance Ca-activated K+ channel. J GenPhysiol 110: 257-281
COX DH, ALDRICH RW (2000) Role of the beta1 subunitin large-conductance Ca(2+)-activated K(+) channelgating energetics. Mechanisms of enhanced Ca(2+)
sensitivity. J Gen Physiol 116: 411-432CUI J, ALDRICH RW (2000) Allosteric linkage between
voltage and Ca(2+)-dependent activation of BK-typemslo1 K(+) channels. Biochemistry 39: 15612-15619
CUI J, COX DH, ALDRICH RW (1997) Intrinsic voltagedependence and Ca2+ regulation of mSlo largeconductance Ca-activated K+ channels. J Gen Physiol109: 647-673
DARBY NJ, CREIGHTON TE (1993) Protein Structure.IRL Press at Oxford University Press, Oxford
DÍAZ L, MEERA P, AMIGO J, STEFANI E, ÁLVAREZO, TORO L, LATORRE R (1998) Role of the S4segment in a voltage-dependent calcium-sensitivepotassium (hSlo) channel. J Biol Chem 273: 32430-32436
EISENMAN G, LATORRE R, MILLER C (1986) Multi-ion conduction and selectivity in the high-conductanceCa++-activated K+ channel from skeletal muscle.Biophys J 50: 1025-1034
GOLOWASH J, KIRWOOD A, MILLER C (1986)Allosteric effects of Mg2+ on the gating of Ca2+-activated K+ channels from mammalian skeletalmuscle. J exp Biol 124: 5-13
HORRIGAN FT, ALDRICH RW (1999) Allosteric voltagegating of potassium channels II. Mslo channel gatingcharge movement in the absence of Ca(2+). J GenPhysiol 114: 305-336
HORRIGAN FT, ALDRICH RW (2002) Coupling betweenvoltage sensor activation, Ca2+ binding and channelopening in large conductance (BK) potassium channels.J Gen Physiol 120: 267-305
HORRIGAN FT, CUI J, ALDRICH RW (1999) Allostericvoltage gating of potassium channels I. Mslo ioniccurrents in the absence of Ca(2+). J Gen Physiol 114:277-304
JIANG Y, LEE A, CHEN J, CADENE M, CHAIT BT,MACKINNON R (2002) Crystal structure andmechanism of a calcium-gated potassium channel.Nature 417: 515-522
JIANG Y, PICO A, CADENE M, CHAIT BT,MACKINNON R (2001) Structure of the RCK domainfrom the E. coli K+ channel and demonstration of itspresence in the human BK channel. Neuron 29: 593-601
KARPLUS K, KARCHIN R, DRAPER J, CASPER J,MANDEL-GUTFREUND Y, DIEKHANS M,HUGHEY R (2003). Combining local-structure, fold-recognition, and new-fold methods for protein structureprediction. Proteins: Structure Function and Genetics53(S6): 491-496

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401400
KEYNES RD, ROJAS E (1974) Kinetics and steady-stateproperties of the charged system in controlling sodiumconductance in the squid giant axon. J Physiol (Lond)239: 393-434
LARSSON HP, BAKER OS, DHILLON DS, ISACOFF EY(1996) Transmembrane movement of the Shaker K+
channel S4 domain. Neuron 16: 387-397LATORRE R, MILLER C (1983) Conduction and
selectivity in potassium channels. J Membr Biol 71:11-30
LATORRE R, OBERHAUSER A, LABARCA P,ÁLVAREZ O (1989) Varieties of calcium-activatedpotassium channels. Ann Rev Physiol 51: 385-399
LATORRE R, VERGARA C, HIDALGO C (1982)Reconstitution in planar lipid bilayers of a Ca2+-dependent K+ channel from transverse tubulemembranes isolated from rabbit skeletal muscle. ProcNatl Acad Sci USA 79: 805-809
LATORRE R, VERGARA C, ÁLVAREZ O, STEFANI E,TORO L (2000) Voltage-gated calcium-modulatedpotassium channels of large unitary conductance:structure, diversi ty and pharmacology. In:Pharmacology of Ion Channel Functions. HandbookExp. Pharmacol. 147: 197-233
LONG SB, CAMPBELL EB, MACKINNON R (2005a)Crystal structure of a mammalian voltage-dependentShaker family K+ channel. Science 309: 897-903
LONG SB, CAMPBELL EB, MACKINNON R (2005b)Voltage sensor of Kv1.2: Structural basis ofelectromechanical coupling. Science 309: 903-908
MA Z, HORRIGAN FT (2005) Voltage-sensing residues inthe S2 and S4 segments of the BK channel. Biophys J88: 100a (Abstract)
MAGLEBY KL 2003 Gating Mechanism of BK (Slo1)Channels: So Near, Yet So Far. J Gen Physiol 121: 81-96
MARTI-RENOM MA, STUART A, FISER A, SÁNCHEZR, MELO F, SALI A (2000) Comparative proteinstructure modeling of genes and genomes. Annu RevBiophys Biomol Struct 29, 291-325
MARRION NV, TAVALIN SJ (1998) Selective activationof Ca2+-activated K+ channels by co-localized Ca2+
channels in hippocampal neurons. Nature 395: 900-904MARTY A (1981) Ca-dependent K channels with large
unitary conductance in chromaffin cell membranes.Nature 291: 497-500
MCMANUS OB (1991) Calcium-activated potassiumchannels: Regulation by calcium. J BioenergBiomembr 23: 537-560
MCMANUS OB, BLATZ AL, MAGLEBY KL (1985)Inverse relationship of the durations of adjacent openand shut intervals for Cl and K channels. Nature 317:625-627
MEDAWAR PB (1979) Advice to a Young Scientist. NewYork: Harper and Row
MEECH RW (1978) Calcium-dependent potassiumactivation in nervous tissues. Annu Rev BiophysBioeng 7: 1-18
MEERA P, WALLNER M, JIANG Z, TORO L (1996) Acalcium switch for the functional coupling betweenalpha (hslo) and beta subunits (Kv, cabeta) of maxi Kchannels. FEBS Lett 385: 127-128
MEERA P, WALLNER M, SONG M, TORO L (1997)Large conductance voltage- and calcium-dependent K+
channel, a distinct member of voltage-dependent ionchannels with seven N-terminal transmembranesegments (S0-S6), an extracellular N terminus, and anintracellular (S9-S10) C terminus. Proc Natl Acad SciUSA 94: 14066-14071
MOSS BL, MAGLEBY KL (2001) Gating and conductance
properties of BK channels are modulated by the S9-S10tail domain of the alpha subunit. A study of mSlo1 andmSlo3 wild-type and chimeric channels. J Gen Physiol118: 711-734
NIMIGEAN CM, MAGLEBY KL (2000) Functionalcoupling of the beta(1) subunit to the largeconductance Ca(2+)-activated K(+) channel in theabsence of Ca(2+). Increased Ca(2+) sensitivity from aCa(2+)-independent mechanism. J Gen Physiol 115:719-736
NIU X, QIAN X, MAGLEBY KL (2004) Linker-gatingring complex as passive spring and Ca(2+)-dependentmachine for a voltage- and Ca(2+)-activated potassiumchannel. Neuron 42: 745-756
NIU X, MAGLEBY KL (2002) Stepwise contribution ofeach subunit to the cooperative activation of BKchannels by Ca2+. Proc Natl Acad Sci USA. 99: 11441-11446
NOCETI F, BALDELLI P, WEI X, QIN N, TORO L,BIRNBAUMER L, SEFANI E (1996) Effective gatingcharges per channel in voltage dependent K+ and Ca2+
channels. J Gen Physiol 108: 143-155OBERHAUSER A, ÁLVAREZ O, LATORRE R (1988)
Activation by divalent cations of a Ca2+-activated K+
channel from skeletal muscle membrane. J Gen Physiol92: 67-86
ORIO P, LATORRE R (2005) Differential Effects of beta1and beta2 Subunits on BK Channel Activity. J GenPhysiol 125: 395-411
ORIO P, ROJAS P, FERREIRA G, LATORRE R (2002)New Disguises for an Old Channel: MaxiK Channel b-Subunits. News Physiol Sci 17: 156-161
PALLOTTA BS, MAGLEBY KL, BARRETT JN (1981)Single channel recordings of Ca2+-activated K+
currents in rat muscle cell culture. Nature 293: 471-474PALLOTTA BS (1985) N-Bromoacetamide removes a
calcium-dependent component of channel opening fromcalcium-activated potassium channels in rat skeletalmuscle. J Gen Physiol 86: 601-611
PISKOROWSKI R, ALDRICH RW (2002) Calciumactivation of BKCa potassium channels lackingcalcium bowl and RCK domains. Nature 420: 499-502
PRAKRIYA M, LINGLE CJ (1999) BK channel activationby brief depolarizations requires Ca2+ influx through L-and Q-type of Ca2+ channels in rat chromaffin cells. JNeurophysiol 81: 2267-2278
ROJAS E, LUXORO M (1963) Micro-injection of trypsininto axons of squid. Nature 199: 78
ROTHBERG BS, MAGLEBY KL (1999). Gating kineticsof single large-conductance Ca2+-activated K+channels in high Ca2+ suggest a two-tiered allostericgating mechanism. J Gen Physiol 114: 93-124
ROTHBERG BS, MAGLEBY KL (2000) Voltage and Ca2+
activation of single large-conductance Ca2+-activatedK+ channels described by a two-tiered allosteric gatingmechanism. J Gen Physiol 116: 75-99
ROST B, YACHDAV G, LIU J (2003) The PredictProteinServer. Nucleic Acids Research 32(online issue):W321-W326
SCHNEIDER MF, CHANDLER WK (1973) Voltagedependent charge movement in skeletal muscle: Apossible step in excitation-contraction coupling. Nature242: 244-246
SCHOPPA NE, MCCORMACK K, TANOUYE MA,SIGWORTH FJ (1992) The size of gating charge inwild-type and mutant Shaker potassium channels.Science 255: 1712-1715
SCHREIBER M, SALKOFF L (1997) A novel calcium-sensing domain in the BK channel. Biophys J 73: 1355-1363

401LATORRE & BRAUCHI Biol Res 39, 2006, 385-401
SCHREIBER M, WEI A, YUAN A, GAUT J, SAITO M,SALKOFF L (1998) Slo3, a novel pH-dependentsensitive K+ channel from mammalian spermatocytes. JBiol Chem 273: 3509-3516
SCHREIBER M, YUAN A, SALKOFF L (1999)Transplantable sites confer calcium sensitivity to BKchannels. Nat Neurosci 2: 416-421
SEOH SA, SIGG D, PAPAZIAN DM, BEZANILLA F (1996)Voltage-sensing residues in the S2 and S4 segments of theShaker K channel. Neuron 16: 1159-1167
SHEN KZ, LAGRUTTA A, DAVIES NW, STANDENNB, ADELMAN JP, NORTH RA (1994)Tetraethylammonium block of Slowpoke calcium-activated potassium channels expressed in Xenopusoocytes: Evidence for tetrameric channel formation.Pflugers Arch 426: 440-445
SHI J, CUI J, (2001) Intracellular Mg2+ Enhances theFunction of BK-type Ca2+-activated K+ Channels. JGen Physiol 118: 589-606
SHI J, KRISHNAMOORTHYET G, YANG Y, HU L,CHATURVEDI N, HARITAL D, QIN J, CUI J (2002)Mechanism of magnesium activation of calcium-activated potassium channels. Nature 418: 876-880
SIGG D, BEZANILLA F (1997) Total charge movementper channel. The relation between gating chargedisplacement and the voltage sensitivity of activation. JGen Physiol 109: 27-39
STARACE D, BEZANILLA F (2001) Histidine scanningmutagenesis of basic residues of the S4 segment of theShaker K+ channel. J Gen Physiol 117: 469-490
STARACE D, BEZANILLA F (2004) A proton pore in apotassium channel voltage sensor reveals a focusedelectric field. Nature 427: 548-553
STEFANI E, OTTOLIA M, NOCETI F, OLCESE R,WALLNER M, LATORRE R, TORO L (1997)Voltage-controlled gating in a large conductance Ca2+-sensitive K+ channel (hslo). Proc Natl Acad Sci USA94: 5427-5431
TALUKDER G, ALDRICH RW (2000) Complex voltage-dependent behavior of single unliganded calcium-sensitive potassium channels. Biophys J 78: 761-772
TORO L, WALLNER M, MEERA P, TANAKA Y (1998)Maxi-K(Ca), a unique member of the voltage-gated Kchannel superfamily. News Physiol Sci 13: 112-117
VERGARA C, LATORRE R, MARRION NV, ADELMANJP (1998) Calcium-activated potassium channels. CurrOpinion Neurobiol 8: 321-329
WALLNER M, MEERA P, TORO L (1996) Determinantfor beta-subunit regulation in high-conductancevoltage-activated and Ca(2+)-sensitive K+ channels: Anadditional transmembrane region at the N terminus.Proc Natl Acad Sci USA 93: 14922-14927
WEI C, SOLARO R, LINGLE CJ, SALKOFF L (1994)Calcium sensi t iv i ty of BK-type KCa channelsdetermined by a separable domain. Neuron 13: 671-681
XIA XM, ZENG X, LINGLE CJ (2002) Multipleregulatory sites in large-conductance calcium-activatedpotassium channels. Nature 418: 880-884
XIA XM, ZHANG X, LINGLE CJ (2004) Ligand-dependent activation of Slo family channels is definedby interchangeable cytosolic domains. J Neurosci 24:5585-5591
YANG N, HORN R (1995) Evidence for voltage-dependentS4 movement in sodium channels. Neuron 15: 213-218
YANG N, GEORGE AL Jr, HORN R (1996) Molecularbasis of charge movement in voltage-gated sodiumchannels. Neuron 16 (1): 113-122
YUAN A, DOURADO M, BUTLER A, WALTON N, WEIA, SALKOFF L (2000) Slo-2, a K+ channel with anunusual Cl-dependence. Nature Neurosci 3: 771-779.
YUAN A, SANTI CM, WEI A, WANG ZW, POLLAK K,NONET M, KACZMAREK L, CROWDWER CM,SALKOFF L (2003) The sodium-activated potassiumchannel is encoded by a member of the Slo genefamily. Neuron 37: 765-773
YUSAF SP, WRAY D, SIVAPRASADARO A (1997)Measurement of the movement of the S4 segmentduring the activation of a voltage-gated potassiumchannel. Pflugers Arch 433: 91-97
ZENG XH, XIA, XM, LINGLE CJ (2005) Divalent cationsensitivity of BK channel activation supports theexistence of three distinct binding sites. J Gen Physiol125: 273-286
ZHANG X, SOLARO CR, LINGLE CJ (2001) Allostericregulation of BK channel gating by Ca(2+) and Mg(2+)
through a nonselective, low affinity divalent cationsite. J Gen Physiol 118: 607-636

LATORRE & BRAUCHI Biol Res 39, 2006, 385-401402