isolation and characterization of class a4 heat shock transcription factor from alfalfa
TRANSCRIPT

Isolation and characterization of class A4 heat shock transcription
factor from alfalfa
Jeremy N. Friedberg a,1, Stephen R. Bowley a, Bryan D. McKersie a,2, William B. Gurley b,Eva Czarnecka-Verner b,*
a Department of Plant Agriculture, Crop Science Building, University of Guelph, Guelph, Ont. N1G 2W1, Canadab Microbiology and Cell Science Department, Program of Plant Molecular and Cellular Biology, University of Florida,
Bldg. 981, Gainesville, FL 32611-0700, USA
Received 4 October 2005; received in revised form 31 March 2006; accepted 10 April 2006
Available online 17 May 2006
Abstract
Plant heat shock transcription factors (HSFs) regulate transcription of heat shock (HS) genes. In Arabidopsis thaliana, 21 HSFs have been
classified into groups A–C. Members of class A act as typical transcriptional activators, whereas B HSFs function as coactivators or repressors
depending on promoter context. The function of class C HSFs is still unclear. Here, we present the isolation and characterization of the first HSF
from alfalfa (Medicago sativa L.) and designate it MsHSFA4 based on amino acid sequence analysis. The MsHSFA4 gene was determined to be
single copy and was detected at two separate genetic loci in the tetraploid Medicago sativa. Overexpression of MsHSFA4 in tobacco mesophyll
protoplasts resulted in weak transcriptional activity, similar to that exhibited by Arabidopsis AtHSFA4a. The MsHSFA4 proximal promoter contains
three putative HSE elements, and the gene itself is activated both by heat and cold stress.
# 2006 Elsevier Ireland Ltd. All rights reserved.
Keywords: Heat shock factor; C-terminal region; Promoter; Chromosomal localization
www.elsevier.com/locate/plantsci
Plant Science 171 (2006) 332–344
1. Introduction
Plants respond to the environmental stress by increased
production of molecular chaperones that protect the cell from
irreparable damage. These proteins produced in response to
elevated temperatures are known as heat shock proteins (HSPs).
The induction of HSPs is universal, very rapid, and controlled at
the transcriptional level by HSFs that bind to heat shock
response DNA elements (HSEs) present in heat shock gene
promoters (reviewed by [1]). The HSE is composed of a
palindromic penta-nucleotide repeat of the core sequence 50-nGAAn-30, or its complement 50-nTTCn-30 [2,3]. The optimal
core sequence for plants was shown to be 50-aGAAg-30, or 50-cTTCt-30 [4]. Plants differ from most other organisms by the
* Corresponding author. Tel.: +1 352 392 2400; fax: +1 352 392 5922.
E-mail address: [email protected] (E. Czarnecka-Verner).1 Present address: BASF Plant Science, 26 Davis Dr., Research Triangle Park,
NC 27709-3528, USA.2 Present address: Vive Technologies Inc., 32 St. Clair Gardens, Toronto,
Ont., Canada M6E 3V4.
0168-9452/$ – see front matter # 2006 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.plantsci.2006.04.007
presence of an extensive network of HSFs. Originally, these
were classified into two [5–8], and later three HSF classes [9]
based on the structural and functional characteristics of HSF
proteins. While all class A HSFs activate transcription [10–13],
they exhibit a large range in the activity, possibly reflecting a
diversity of function [14,15]. Some HSFs play a key role in the
early heat shock response and thermotolerance [16,17], while
others function in a secondary role to boost the activity of other
class A HSFs [18]. Transcriptional activation depends on the
AHA domains present in the C-terminal regions (CTR) of the
class A HSFs [8,10,15,19]. AHA motifs were not found in class
B HSFs. As a result, class B HSFs do not activate transcription
on their own, but rather seem to be specialized as active
repressors of transcription in certain circumstances [20],
perhaps as attenuators of the heat shock (HS) response
[13,21]. Studies in Arabidopsis suggest that the A and B class
HSFs have differential roles in the early versus late phases of
the HS response [16].
Impairment of class B HSFs coincided with the loss of ability
to transiently shutdown transcription of HSF target genes [16].
Interestingly, class B HSFs can also act as coactivators through

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 333
synergistic mechanisms that may involve interactions of their C-
terminal domains with general transcription factor TFIIB [20]
and coactivators such as HAC1 [22]. The function of class C
HSFs still remains largely unexplained. There are only two
known members of this class, and surprisingly, although
Arabidopsis AtHSFC1 lacks obvious AHA motifs, it seems to
weakly activate transcription in plant and yeast systems [15].
HSF proteins have a highly conserved structural motifs [7,9]
interspaced with regions that vary significantly in size and
sequence. The DNA binding domain (DBD) has the greatest
conservation and has become the defining feature of the HSFs.
It is located in the N-terminus and contains a helix-turn-helix
structure [23] related to the HNF3/forkhead winged helix
family of DNA binding motifs [24]. The DBD is connected to
the oligomerization domain (OD) by a flexible linker domain
[25]. In plant HSFs, the length of the flexible linker seems to be
longer for B HSFs than for the class A HSFs [9]. HSF binds to
DNA most commonly as a trimer [26]. The trimerization
domain forms a triple-stranded, a-helical coiled coil analogous
to the one present in influenza hemagglutinin [27], with two
subdomains of hydrophobic heptapeptide repeats, HR-A and
HR-B, separated by a break in the helicity of the OD [28].
Structural features of the ODs considered together with the
parsimony analysis of DBDs [6] support differentiation of plant
HSFs into various classes [5,9]. In human HSFs, the third
hydrophobic heptapeptide repeat HR-C masks the OD and a
nuclear localization sequence (NLS) resulting in an inactive,
monomeric form of HSF localized in the cytoplasm [29,30].
Plant class B HSFs do not seem to have a functional HR-C [5,9]
and are present in the nucleus under HS and non-HS conditions
[18]. The intracellular distribution of plant A HSFs depends on
the overall balance of nuclear import and export processes that
are directed by the strength and accessibility of the NLS and
nuclear export sequence (NES) [15,31]. Transcriptional
activation of class A HSFs is attributed to AHA motifs located
in the C-terminal region (CTR) of class A HSFs [10,13].
Detailed analysis of these motifs revealed the importance of
aromatic, leucine and proline residues, as well as the synergism
between adjacent AHA motifs [19,20]. These AHA motifs have
been shown to recruit members of the transcriptional
preinitiation complex (PIC) such as TBP and TFIIB [20].
The combination of AHA and NES constitutes a signature
domain that has facilitated the recognition of over 60 new class
A HSFs among the expressed sequence tag (EST) database of
plants [15].
The HS response has been well documented in a wide variety
of organisms subjected to elevated temperatures. Remarkably,
several groups have reported the induction of HSP gene
expression during low temperature stress [32–35], including
freezing [36]. There are indications that at least members of the
HSP90 [33], HSP70 [32,35] and some low molecular weight
(17–21 kDa) HSP families [34,36–38] are involved in
protection against chilling injury and acquisition of freezing
tolerance. HS and HSP accumulation can provide cross-
adaptation to cold stress. For example, the increase of HSP17.6
and HSP21 expression has been correlated with the protection
of tomato fruits from chilling injury [37,38]. Heat treatments
prior to chilling reduced the chilling-induced inhibition of rice
seedling radicle growth [39] and rates of ion leakage and
chloroplast aggregation in petioles of African violet [40].
Evidence for HSPs providing protection from both heat and
chilling injury is provided by experiments in E. coli when
CsHSP17.5 from chestnut conveyed increased bacterial cell
viability under heat stress and increased survivability at low
temperatures [41]. The same cytosolic CsHSP17.5 was shown
to play an important role in the acquisition of freezing tolerance
by protecting the cold-labile lactate dehydrogenase enzyme
from freezing-induced inactivation [42]. The mechanisms of
cryo-protection provided by HSPs still need to be elaborated,
but molecular chaperone activity as a protection against freeze-
induced protein denaturation has been suggested [43]. The
accumulation of HSPs during episodes of chilling clearly
suggests that HSPs may enhance a plants’ ability to cope with
low temperature stress. However, it is still not known if HSFs
are responsible for the induction of HSPs under the low
temperature stress. Although, HSFs are responsible for heat-
induced expression of HSPs, their role in chilling-induced
promoter activation has not been documented.
Here, we present the isolation and characterization of the
first HSF cloned from Medicago sativa L. (alfalfa). We confirm
its ability to function as a transcriptional activator and find the
gene encoding MsHSFA4 to be induced by heat and cold
stresses. The novel finding of cold activated transcription of a
HSF may be physiologically significant since alfalfa is
susceptible to low temperature stress and winter kill.
2. Materials and methods
2.1. cDNA library screening
The cDNA library was kindly provided by Drs. Suzanne
Cunningham and Jeffrey Volenec from Purdue University.
PolyA+ RNA was extracted from field-acclimated alfalfa (cv.
Norseman) bud tissue harvested in September, November and
December. For library construction, this RNA was pooled and
used as template for cDNA synthesis and cloning into the
lambda ZAP vector from the Lambda ZapTM library kit
(Stratagene, La Jolla, CA) according to the protocol supplied by
the manufacturer. Screening of the library was performed
according to methods detailed in Current Methods in Molecular
Biology [44] and the protocols supplied by Stratagene. The
open reading frame (ORF) of the Arabidopsis thaliana
AtHSFA4a cDNA (GenBank accession no. AAC31792) was
used as a heterologous probe for library screening. The
AtHSFA4a ORF was amplified from plasmid DNA [13] by
polymerase chain reaction (PCR) and subsequently radio-
actively labeled with 32P using the PrimIt II Labeling KitTM
(Stratagene). The AtHSFA4a probe had a specific activity of
2.3 � 109 cpm/mg. One million phage plaques were tested in
the first library screen. Putative positive candidates were then
subjected to three rounds of plaque purification resulting in a
pure plaque containing MsHSF. The MsHSF cDNA recovered
from the lambda ZAP vector was sequenced from both
directions using the T3 and T7 universal primers by Lab

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344334
Service, University of Guelph, and was designated MsHSFA4
(GenBank accession no. AF235958). The transcription start site
for MsHSFA4 was based on sequences derived from cDNA
clones and confirmed using a series of RT-PCR reactions with
mRNA template from control and heat stressed tissues and
primer pairs strategically located within the 50 region. The 50
end of MsHSFA4 mRNA was then independently determined
by primer extension assays (data not shown).
2.2. Genomic DNA libraries construction and screening
Alfalfa (cv. N442) genomic DNA was isolated from 1 g of
pooled mature and developing leaf tissue using the Trebuchet
genomic DNA extraction protocol (Bowley lab, University of
Guelph; protocol available upon request), dissolved in 100 ml
of distilled water and incubated overnight at 40 8C. Genomic
libraries were built using the BD GenomeWalkerTM Universal
Kit (ClonTech, Palo Alto, CA) according to the manufacturer’s
instructions. Genomic DNA was digested in four separate
reactions by restriction enzymes EcoR V, Dra I, Pvu II and Ssp I,
and adapter fragments containing unique primer sites were
ligated onto the ends of the digested genomic DNA from each
reaction. This process generated four genomic libraries that
were used as templates for two rounds of polymerase chain
reactions (PCR). This first amplification utilized a gene-specific
downstream primer and upstream adapter primer, followed by a
second amplification with nested primers. PCR fragments were
cloned using the TOPO TA cloning kit from Invitrogen. The
genomic DNA inserts were sequenced and 50 flanking
sequences were analyzed for promoter consensus elements
using the following programs:
PLACE (http://www.dna.affrc.go.jp/PLACE/fasta.html),
PROSCAN (http://bimas.dcrt.nih.gov/molbio/proscan/),
Promoter 2.0 (http://www.cbs.dtu.dk/services/Promoter/),
TESS (http://www.cbil.upenn.edu/tess/) and TRANSFAC
(http://www.gene-regulation.com/pub/databases.html).
2.3. Temperature stress treatments and RNA extractions
Approximately 500 mg of shoot tips (first three expanding
leaves) were harvested from alfalfa cultivar N442 and
incubated in 100 ml of temperature-acclimated SHK media
(derived from SH media [45] modified with 10 mM K2SO4),
with continuous rotation at 175 rpm in temperature-controlled
shakers. The treatments were continued for 0.5 and 4 h at the
following temperatures: heat shock at 41 8C, cold stress at 4 8Cand control at 28 8C. The samples were then immediately
frozen in liquid N2 and stored at�80 8C. An additional control
consisted of shoot meristems harvested from plants and
directly frozen in liquid N2. Frozen tissue samples were ground
under liquid N2 and total RNA was extracted using the
RNaqueous RNA Extraction Kit (Ambion, Austin, TX) in
conjunction with Plant RNA Isolation Aid (Ambion). Aliquots
of 20 mg of RNA were stored at �80 8C. Total RNA was
quantified using RiboGreen fluorescent RNA stain (Molecular
Probes, Eugene, OR) and detected with a Shimadzu RF-5000
spectrofluorophotometer with excitation at 480 nm and emi-
ssion at 580 nm.
2.4. Preparation of the digoxigenin-labeled DNA probe
and Northern blot hybridizations
The entire ORF of the alfalfa MsHSFA4 was used to generate
a digoxigenin (DIG)-labeled probe for Northern blot hybridi-
zations. The MsHSFA4 clone was digested using EcoR I and
Xho I restriction enzymes, and the 1.2 kb ORF insert was
isolated by purification on agarose gels. The MsHSFA4 insert
was used as a template in the DIG PCR reaction that was carried
out according to the procedure detailed in the DIG PCR Probe
Synthesis Kit (Roche Biochemicals, Laval, Quebec) with
primers specific to the full length ORF.
For Northern blots, 20 mg aliquots of total RNA were
electrophoresed on a 1.2% (w/v) agarose gel prepared with 1�MOPS buffer and 5.4% formaldehyde. The RNA was capillary-
transferred overnight in 10� SSC to a positively charged nylon
membrane (Roche Biochemicals) and then fixed to the
membrane via UV crosslinking. The membrane was prehy-
bridized for 2 h at 37 8C in EasyhybeTM (Roche Biochemicals)
prehybridization solution and then incubated overnight in a
rotating hybridization oven at 50 8C with DIG-labeled
MsHSFA4 probe. Subsequently, membranes were washed
twice at room temperature for 15 min each in 2� stringency
wash solution (2� SSC, 1% SDS (w/v)), followed by washes at
65 8C in 0.2� stringency wash solution (0.2� SSC, 1% SDS
(w/v)) and further processed according to protocols provided by
Roche Biochemicals. The membrane was placed on an acetate
sheet, overlaid with the chemiluminescent substrate solution
and heat-sealed. Hybridized blots were displayed by exposure
of the membrane to Kodak XGB X-ray film.
2.5. Protoplast transformations, GUS assays and
constructs
Protoplast transformation and evaluation of b-glucuroni-
dase (GUS) expression was performed as described pre-
viously [13]. Protoplast assays were repeated two to three
times and were conducted with triplicate replications. GUS
reporter activities were normalized against luciferase activity
as an internal standard. Two types of GUS reporter constructs
were used: 9� HSE/GUS for the full length HSF constructs
with the native HSF DBD, and 10� Gal4 DBS/GUS for
chimeric constructs containing CTR of HSFs fused to the
yeast Gal4 DBD. In either case, the multiple DNA recognition
sites were fused upstream to a minimal 35S CaMV promoter.
The effector constructs for testing with the 9� HSE/GUS
reporter consisted of (1) the full length MsHSFA4 ORF from
the first methionine to the last coding amino acid residues
cloned behind the yeast Gal4 leader sequence, and (2) the
complete cDNA of MsHSFA4, including its long 50
untranslated region (50UTR), cloned in the same Gal4 leader
vector. The full length Arabidopsis AtHSFA1a and AtHSFA4a
were used as positive examples of transcriptionally active
HSF genes, and vectors containing only a Gal4 leader
J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 335
sequence, or additionally encoding T7 tag peptide, served as
negative controls.
The second group of effector constructs tested with the 10�Gal4 DBS/GUS reporter had yeast Gal4 transcription factor
DBD (aa 1-147) present, either alone or fused to the alfalfa
MsHSFA4 CTR (aa 189-402, plus its native stop codon and
most of the 30 untranslated region). The CTR (aa 189-401) of
Arabidopsis AtHSFA4a was used as a positive control.
2.6. Fluorescent in situ hybridization
This protocol was performed following instructions of the
Cereal Genetics Laboratory In Situ Hybridization Manual
(USDA, 1999) using germinating alfalfa seed (cv. ABI 700). A
biotin-labeled probe was prepared from a plasmid containing
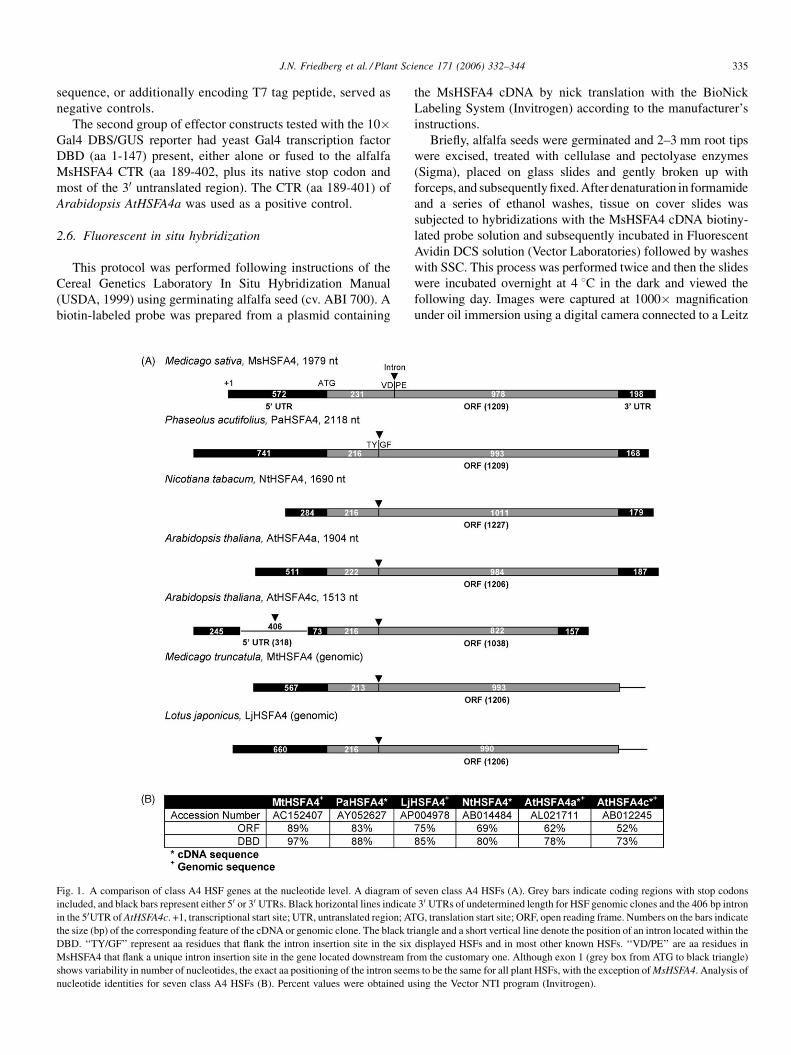
Fig. 1. A comparison of class A4 HSF genes at the nucleotide level. A diagram of
included, and black bars represent either 50 or 30 UTRs. Black horizontal lines indicat
in the 50UTR of AtHSFA4c. +1, transcriptional start site; UTR, untranslated region; AT
the size (bp) of the corresponding feature of the cDNA or genomic clone. The black tr
DBD. ‘‘TY/GF’’ represent aa residues that flank the intron insertion site in the six
MsHSFA4 that flank a unique intron insertion site in the gene located downstream fr
shows variability in number of nucleotides, the exact aa positioning of the intron seem
nucleotide identities for seven class A4 HSFs (B). Percent values were obtained u
the MsHSFA4 cDNA by nick translation with the BioNick
Labeling System (Invitrogen) according to the manufacturer’s
instructions.
Briefly, alfalfa seeds were germinated and 2–3 mm root tips
were excised, treated with cellulase and pectolyase enzymes
(Sigma), placed on glass slides and gently broken up with
forceps, and subsequently fixed. After denaturation in formamide
and a series of ethanol washes, tissue on cover slides was
subjected to hybridizations with the MsHSFA4 cDNA biotiny-
lated probe solution and subsequently incubated in Fluorescent
Avidin DCS solution (Vector Laboratories) followed by washes
with SSC. This process was performed twice and then the slides
were incubated overnight at 4 8C in the dark and viewed the
following day. Images were captured at 1000� magnification
under oil immersion using a digital camera connected to a Leitz
seven class A4 HSFs (A). Grey bars indicate coding regions with stop codons
e 30 UTRs of undetermined length for HSF genomic clones and the 406 bp intron
G, translation start site; ORF, open reading frame. Numbers on the bars indicate
iangle and a short vertical line denote the position of an intron located within the
displayed HSFs and in most other known HSFs. ‘‘VD/PE’’ are aa residues in
om the customary one. Although exon 1 (grey box from ATG to black triangle)
s to be the same for all plant HSFs, with the exception of MsHSFA4. Analysis of
sing the Vector NTI program (Invitrogen).

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344336
Fig. 2. Amino acid sequence comparison of plant class A4 HSFs. (A) The diagram of a typical class A4 HSF protein illustrates conserved regions. DBD, DNA
binding domain; OD, oligomerization domain; NLS, nuclear localization signal; AHA1 and AHA2, transcriptional activation motifs; NES, nuclear export
sequence; CTR, C-terminal regulatory region spans amino acid sequences from OD to the end of the HSF protein. (B) Amino acid sequence alignment of seven
classes of A4 plant HSFs: (1) Medicago sativa MsHSFA4 (GenBank accession no. AAF37579); (2) Phaseolus acutifolius PaHSFA4 (GenBank accession no.
AAL12248), (3) Nicotiana tabacum NtHSFA4, formerly known as NtHSF2 (GenBank accession no. BAA83711); (4) Arabidopsis thaliana AtHSFA4a, formerly

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 337
Fig. 3. HSF relatedness tree from CLUSTALW analysis of the HSF DNA binding domains. Only class A HSFs were used in the analysis. Although previous HSF
surveys [9] distinguished nine classes of A HSFs, this analysis puts AtHSFA5/4b (GenBank accession no. CAB10177) in class A4 alongside of ZmHSFA4 (GenBank
accession no. CAA58117), OsHSFA9 (GenBank accession no. AAQ23063) and OsHSFA10 (GenBank accession no. AAQ23064). Here, HaHSFA9 (GenBank
accession no. AAM43804) is placed in the HSFA2 subfamily.
phase contrast fluorescent microscope. Images of chromosome
spreads were captured using Northern Eclipse Image Analysis
Software (Empix Imaging, Mississauga, Ont., Canada). Indivi-
dual chromosomes [46,47] were karyotyped using MicroMea-
sure software (version 3.3, Aaron Reeves & Jim Tear, Colorado
State University).
3. Results
3.1. Alfalfa HSFA4 homologue
Sequencing of the positive cDNA clone isolated from the
alfalfa bud cDNA library revealed that it was approximately 2 kb
in length. BLASTx nucleotide analysis identified this clone to
exhibit a high degree of similarity to known genes encoding plant
HSFs, specifically, Medicago truncatula MtHSFA4 (GenBank
accession no. AC152407), Phaseolus acutifolius PaHSFA4
(GenBank accession no. AY052627), Lotus corniculatus var.
japonicus LjHSFA4 (GenBank accession no. AP004978),
Nicotiana tabacum NtHSFA4, formerly known as NtHSF2
(GenBank accession no. AB014484), Arabidopsis thaliana
AtHSFA4a, known previously as AtHSF21 (GenBank accession
nos. U68561 and AL021711) and Arabidopsis thaliana
AtHSFA4c (GenBank accession no. AB012245) (Fig. 1).
Comparison of the MsHSFA4 nucleotide sequence to PaHSFA4
known as AtHSF21 ([13], GenBank accession no. AAC31792); (5) Arabidopsis th
LjHSFA4 (GenBank accession no. AP004978) and Medicago truncatula MtHS
homology and correspond to textures in the diagram shown in A. Amino acid residu
represent hydrophobic heptapeptide repeats A and B shown in register. Two vertical
present in HSF C-terminal regions. The black triangle on the left indicates the most
its position within MsHSFA4 (see text). (C) Amino acid sequence identities of se
program.
and LjHSFA4 (all three Fabaceae) revealed a high degree of
similarity in both, the overall nucleotide sequence of the ORFs
(83% and 75% identical, respectively) and regions encoding the
DBD (respectively, 88 and 85% identical) (Fig. 1B). Tobacco
NtHSFA4 (Solanaceae) showed 80% similarity in the region
encoding the DBD and 69% over the entire ORF. In contrast, not
so closely related Arabidopsis AtHSFA4a and AtHSFA4c
(Brassicaceae) displayed only 62 and 52% similarity to
MsHSFA4 in the overall ORF sequence, with significantly
higher similarities (78 and 73%) in regions encoding DBDs. Not
surprisingly, MsHSFA4 and MtHSFA4 share the greatest
identity, especially in the nucleotide sequences of the ORF
region (89%) and the DBD region (97%). This high degree of
conservation indicates relatively recent divergence between both
Medicago species and an overall evolutionary need for a high
degree of amino acid conservation of the DBD.
A schematic representation of HSP genes related to
MsHSFA4 is shown in Fig. 1. Most have 50UTRs ranging
from approximately 250 bp to over 700 bp. The 50UTR for
MtHSFA4 was predicted based on the high degree of sequence
similarity with MsHSFA4 (not shown), and the 50UTR for
LjHSFA4 was based on the location of a putative promoter
sequence (TATAA and HS elements). Interestingly, the related
class A4 Spl7/OsHSF10 gene of rice also has an extensive
50UTR (approximately 600 bp), which contains an intron of
aliana AtHSFA4c (GenBank accession no. BAB09213); (6) Lotus japonicus
FA4 (GenBank accession no. ABE77517). Boxes indicate regions of high
es, written in bold type face are aligned in positions ‘‘d’’ and ‘‘a’’ of the OD and
boxes encompass highly conserved amino acid sequences of unknown function
common position of an intron within the DBD; the right triangle corresponds to
ven classes of A4 HSFs. Percent values were obtained using the Vector NTI

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344338
243 bp [48]. A similar case is observed for the AtHSFA4c gene
that has a 50UTR of 724 bp with an intron of 406 bp.
The MsHSFA4 cDNA encodes a predicted protein of 402
amino acids with a molecular weight of 46.2 kDa (GenBank
accession no. AAF37579). The full-length protein sequence
shows high amino acid similarity to LjHSFA4, PaHSFA4,
NtHSFA4, AtHSFA4a and AtHSFA4c of 67, 67, 50, 46 and
43%, respectively (Fig. 2C). When only the DBD amino acid
sequence of HSFs was considered, the similarity increased to
77–86% among these six HSFs. Overall, the MsHSFA4 protein
showed the highest degree of similarity to the barrel medic
MtHSFA4 ORF (90%) and DBD (95%), with a very significant
conservation of the C-terminal transcriptional regulatory region
(87% identity) (Fig. 2C). In contrast, amino acid similarity to
Arabidopsis AtHSFA4 CTRs was only 27–29%. CLUSTALW
[49] neighbor joining analysis of the MsHSFA4 cDNA
(GenBank accession no. AF235958) and available plant full-
length HSF ORF sequences (data not shown) and DBDs (Fig. 3)
confirmed the placement of alfalfa MsHSF in class A4 HSFs,
alongside of barrel medic, tepary bean, tobacco, tale cress, rice
and maize A4 HSFs.
Conserved domains of the MsHSFA4 protein are dia-
grammed in Fig. 2 and include: (1) the DBD located between aa
residues 11 and 103, (2) a typical class A HSF OD (aa 127-184)
consisting of HR-A and HR-B, (3) downstream-localized NLS
(aa 202-205), (4) two transcriptional activation AHA motifs (aa
257-262 and 340-345) and (5) NES located around QMGHV
sequence (aa 392-396). CTR is immediately adjacent to the OD
and encompasses NLS, AHA motifs and NES.
3.2. MsHSFA4 genomic clone and promoter sequence
Isolation and sequencing of the MsHSFA4 genomic clone
(GenBank accession no. AF494082) revealed this gene to be
2358 bp in length and have two exons of 231 and 978 bp that
flank a single intron of 104 bp (Fig. 4A). The intron is located
within the DBD; however, it is shifted 6 aa residues downstream
(D77/P78; Figs. 4A and 1A) from its position in other plant
HSFs (Y71/G72; Fig. 1A) [9]. The intron/exon boundries
contain the canonical 50/30 sequence (GT/AG) that commonly
denotes splice junctions.
The genomic clone contains 278 bp of DNA upstream of the
50 UTR. Within this region, a number of promoter elements
were identified; specifically, a putative TATA box, two TATA-
proximal heat shock elements (sites I and II), a third upstream
HSE III and several HSE core tri-nucleotide elements present in
both orientations (Fig. 4B). These isolated core sequences lack
flanking sequence identity required to meet the definition of
active head/tail modules described by Nover et al. [9], albeit
some show partial conservation with the optimal plant HSE
core consensus [4].
From the current sequence database, we selected and
analyzed genomic sequence data available for class A4 HSF
from Medicago truncatula. The promoter region of MtHSFA4
exhibits 89% homology with that of MsHSFA4. CLUSTALW
alignment of the MsHSFA4 and MtHSFA4 promoter regions
(Fig. 4C) indicated that, although most of the HSE cores
were conserved in sequence and location within promoters,
some point mutations and deletion/insertions of the promoter
sequences occurred. In general, the MtHSFA4 promoter is
less conserved than the MsHSFA4 promoter with respect to
the identity of HSE consensus sequence. The MtHSFA4
promoter, compared to MsHSFA4, displayed seven single or
double nucleotide differences within the HSE cores, as well
as the lack of five nucleotides within the putative HSE site II.
Although the differences occurring in the MtHSFA4 promoter
weakened the overall homology of cores to the consensus,
the missing nucleotides within HSE II seemed to preserve
the register of HSE arrays and brought six of them closer
together, still making it possible for two hypothetical
HSF trimers (designated by head-to-toe arrows) to bind
(Fig. 4D).
3.3. MsHSFA4 gene copy number and chromosomal
localization
Southern analysis using a DNA probe to the MsHSFA4 ORF
revealed a single strong band and a possible faint second band
(data not shown) suggesting that MsHSFA4 is a single copy
gene. Chromosome spreads exhibited the expected 32 visible
chromosomes (8 homologues), confirming the source plants as
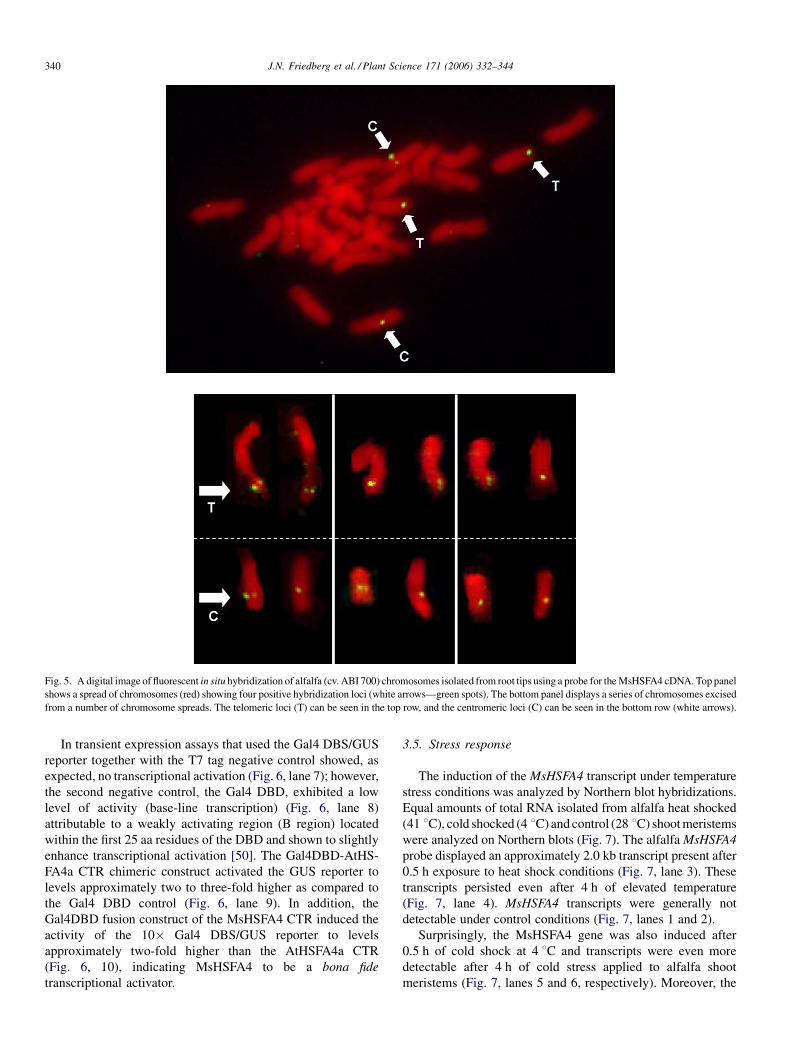
tetraploid alfalfa. Chromosomal localization of MsHSFA4
through fluorescent in situ hybridization revealed that this gene
exists in single copy number, with four copies in a tetraploid
genome located on chromosome 6, or 7. Among the four
homologous chromosomes, MsHSFA4 resides at one of two
genetic loci (Fig. 5). The first locus is found near the telomere,
and the second is found near the centromere.
3.4. Transcriptional activity of MsHSFA4 in tobacco
protoplasts
The goal of this series of experiments was two-fold. The first
was to determine if the alfalfa HSFA4 gene possessed
transcriptional activity, and the second was to determine the
ability of the isolated CTR of MsHSFA4 to act as a typical
transcriptional activator outside of the HS-dependent mechan-
ism. These experiments were conducted using transient assays
in tobacco mesophyll protoplasts. The reporter consisted of
GUS gene under control of an artificial HS promoter consisting
of the minimal CaMV 35S promoter (TATAA) with nine typical
HSE motifs located upstream. The second objective was met by
separating the requirements for trimerization and HSE binding
from transcriptional activation and, thereby, alleviating the
need for HS-mediated induction. The CTR of MsHSFA4 was
fused to the yeast Gal4 DBD by cloning and tested for the
ability to activate a GUS reporter driven by a minimal CaMV
35S promoter with 10 Gal4 DNA binding sites (Gal4 DBS)
located upstream.
Two types of vectors were used as negative controls: one
with transcript corresponding to the Gal4 untranslated 50-leader
and the other additionally encoding the T7 epitope. Both of
these controls showed no transcriptional activity, indicating that
there was little to no background GUS activity in tobacco

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 339
Fig. 4. Alfalfa MsHSFA4 gene promoter analysis. (A) Diagram of the MsHSFA4 genomic sequence shows the organization of the gene. VD/PE are the last two amino
acid residues in exon 1 and the first two amino acid residues in exon 2 of MsHSFA4. Numbers indicate the size (bp) of specific regions. (B) The diagram illustrates the
location of the putative TATA box (rectangular box), transcriptional start site (+1) and distribution of HSE cores (semi-square boxes) within the MsHSFA4 promoter.
Triangles mark conservation to the HSE pentanucleotide core consensus sequence [4]. Brackets indicate three potentially functional HSE elements. HSE; heat shock
element. (C) Alignment of promoter sequences for two closely related Medicago HSFA4 genes using CLUSTALW. Grey box indicates a putative TATA box. Clear
boxes encompass conserved HSE core sequences, and arrows illustrate their head-to-toe orientations.
protoplasts (Fig. 6, lanes 2 and 1, respectively). In contrast,
Arabidopsis class A1 HSF, AtHSFA1a ORF, displayed very
high levels of GUS activity (Fig. 6, lane 3) and served as the
positive control for transcriptional activation. The AtHSFA4a
ORF construct showed approximately 20% of transcriptional
activity as compared to the AtHSFA1a ORF (Fig. 5, lane 4), a
finding consistent with previous experiments designating
AtHSFA4a as a weak transcriptional activator [13,14,20].
The alfalfa MsHSFA4 ORF displayed comparable levels of
activity to its homologue Arabidopsis AtHSFA4a (Fig. 6,
lane5). However, the MsHSFA4 full length cDNA, that
included a long 50 UTR sequence, was unable to activate the
9� HSE/GUS reporter construct, indicating that the 50 UTR
may potentially act as a negative regulator (Fig. 6, lane 6).
J.N. Friedberg et al. / Plant Science 171 (2006) 332–344340
Fig. 5. A digital image of fluorescent in situ hybridization of alfalfa (cv. ABI 700) chromosomes isolated from root tips using a probe for the MsHSFA4 cDNA. Top panel
shows a spread of chromosomes (red) showing four positive hybridization loci (white arrows—green spots). The bottom panel displays a series of chromosomes excised
from a number of chromosome spreads. The telomeric loci (T) can be seen in the top row, and the centromeric loci (C) can be seen in the bottom row (white arrows).
In transient expression assays that used the Gal4 DBS/GUS
reporter together with the T7 tag negative control showed, as
expected, no transcriptional activation (Fig. 6, lane 7); however,
the second negative control, the Gal4 DBD, exhibited a low
level of activity (base-line transcription) (Fig. 6, lane 8)
attributable to a weakly activating region (B region) located
within the first 25 aa residues of the DBD and shown to slightly
enhance transcriptional activation [50]. The Gal4DBD-AtHS-
FA4a CTR chimeric construct activated the GUS reporter to
levels approximately two to three-fold higher as compared to
the Gal4 DBD control (Fig. 6, lane 9). In addition, the
Gal4DBD fusion construct of the MsHSFA4 CTR induced the
activity of the 10� Gal4 DBS/GUS reporter to levels
approximately two-fold higher than the AtHSFA4a CTR
(Fig. 6, 10), indicating MsHSFA4 to be a bona fide
transcriptional activator.
3.5. Stress response
The induction of the MsHSFA4 transcript under temperature
stress conditions was analyzed by Northern blot hybridizations.
Equal amounts of total RNA isolated from alfalfa heat shocked
(41 8C), cold shocked (4 8C) and control (28 8C) shoot meristems
were analyzed on Northern blots (Fig. 7). The alfalfa MsHSFA4
probe displayed an approximately 2.0 kb transcript present after
0.5 h exposure to heat shock conditions (Fig. 7, lane 3). These
transcripts persisted even after 4 h of elevated temperature
(Fig. 7, lane 4). MsHSFA4 transcripts were generally not
detectable under control conditions (Fig. 7, lanes 1 and 2).
Surprisingly, the MsHSFA4 gene was also induced after
0.5 h of cold shock at 4 8C and transcripts were even more
detectable after 4 h of cold stress applied to alfalfa shoot
meristems (Fig. 7, lanes 5 and 6, respectively). Moreover, the

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 341
Fig. 6. The full length MsHSFA4 and its isolated CTR activate transcription of reporter genes when expressed in tobacco mesophyll protoplasts. Transcriptional
activities of HSF effector constructs containing their native DBDs were tested using the 9� HSE/GUS reporter and are shown in lanes 1–6. Transcriptional activities
of isolated HSF CTRs in fusion constructs with yeast acidic activator Gal4 DBD for Arabidopsis and Medicago sativa class A4 HSFs are presented in lanes 7 through
10. Transcriptional activity of HSF CTRs was tested using the 10� Gal4 DBS/GUS reporter. Basal transcriptional activity of the tobacco protoplast system is
presented in lanes 1 and 2 for the 9� HSE/GUS reporter, and in lane 7 for 10� Gal4 DBS/GUS. Base-line activity using Gal4 DBD alone is reflected in lane 8. GUS
activity was normalized to the activity of the internal luciferase (Luc) standard that was driven by a full-length ubiquitin promoter. ORF (open reading frame), G4L
(Gal 4 leader sequence), 50 UTR (50 untranslated leader of MsHSFA4), T7 (T7 tag peptide). Graphed values represent means � S.D., where n = 3.
Fig. 7. Heat and cold stress induced the expression of MsHSFA4 gene. Northern
blots containing equal amounts of total RNA isolated from shoot meristems from
either control or stressed alfalfa were hybridized to the DIG-labeled probe of
MsHSFA4 ORF generated by PCR. A non-treated (nt) control grown at 28 8C and
harvested is shown in line 1. Shoot tips incubated in the incubation medium at
28 8C for 4 h are considered a treated control (liq) and are presented in lane 2. Heat
stressed samples are located in lanes 3 (41 8C for 30 min) and 4 (heat shock at
41 8C for 4 h), and cold stressed in lanes 5 (4 8C for 30 min), 6 (cold shock at 4 8Cfor 4 h) and 7 (bud tissue from field acclimated plants harvested in November).
The accuracy of RNA loading was confirmed by using RNA fluorescent dye and
through normalizing against an ethidium bromide stained ribosomal 18S and 28S
RNA (rRNA) bands. Molecular weight markers are denoted on the left side of the
top panel. This experiment was replicated three times.
same transcript was detected in the alfalfa shoot meristems
RNA sample isolated from field acclimated alfalfa (cv. N442)
crown buds collected in November 1999 (Fig. 7, lane 7), raising
the possibility of a role for this transcriptional regulator of the
HS response during periods of low temperature.
4. Discussion
We have isolated the first heat shock transcription factor
from Medicago sativa. MsHSFA4 is a single copy gene, and the
predicted protein exhibits structural and functional domains
highly conserved in other eukaryotes and specifically in plants.
At both the DNA and protein levels, the sequence conservation
for the DBD is higher than for the ORF as a whole. As expected,
at the DNA level, the sequence conservation directly correlates
with the overall degree of phylogenetic relatedness among the
species examined. For example, MsHSFA4 is more similar to
orthologs in Medicago truncatula, Phaseolus acutifolius and
Lotus japonicus, species belonging to the same family
Fabaceae; whereas Arabidopsis, which exhibited the least
similarity of the seven sequences surveyed, is found in the
Brassicaceae, and tobacco belongs to the Solanaceae. As its
designation indicates, MsHSFA4 is best categorized as a
member of the class A4 HSFs (Fig. 3). The phylogenetic tree
shown in Fig. 3 was based on CLUSTALW neighbor joining
analysis of the aa sequences and indicates that MsHSFA4 is
solidly grouped within the A4 clade. In addition, four other
more distantly related HSFs are located on the same branch
(OsHSF9, OsHSF10, ZmHSFA4 and AtHSFA5/A4b). Of these,
rice HSFs showed the greatest diversity within the CTRs;
blocks of sequences were inserted between AHA1 and AHA2
motifs for OsHSF10, and one such insertion seemed to
eliminate the OsHSF10 AHA1 domain. Interestingly, rice
spotted leaf gene Spl7 (identical to OsHSF10), whose DBD
mutant spl7 (W40/C40) is responsible for the lesion-mimic
plant phenotype, was identified previously to be class A4 HSF
and implicated in protection against necrosis caused by
environmental stress [48].
Alfalfa chromosomes are small and relatively similar in size,
which makes it extremely difficult to identify different
chromosomes by in situ hybridization procedures. As a result,
relative length and arm ratios were the only tools that could be
used to differentiate between chromosomes. Thus, through

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344342
repetitive analysis we concluded that MsHSFA4 is most likely
located on a single chromosome, either 6 or 7. These studies
have revealed two loci for this single copy gene. In an auto-
tetraploid, one expects to find a single copy gene at one locus
among the four homologous chromosomes. However, alfalfa
exhibits one locus near the telomere and the second locus near
the centromere. Cytologically, this type of locus orientation is
usually indicative of a large chromosomal inversion event.
However, since two homologues have the telomere locus and
the other two homologues have the centromere locus, a 2:2
locus ratio, as opposed to a 1:3 locus ratio, suggest that the
inversion event occurred before alfalfa’s fusion from diploid to
tetraploid. This is significant, since it suggests that alfalfa arose
from an ‘‘allo’’ doubling of chromosomes as opposed to the
current belief of an ‘‘auto’’ doubling of chromosomes. Thus, the
alfalfa genome may exist as a chimera, exhibiting character-
istics of diploid and tetraploid genetics, reminiscent of its
progenitor parents.
The MsHSFA4 protein contains all critical domains
important for it to function as a transcriptional activator, i.e.
DNA binding domain, oligomerization domain, nuclear
localization signal, nuclear export signal and two transcrip-
tional activation domains AHA1 and AHA2. MsHSFA4
contains a conserved amino acid sequence PVHSHS flanking
the C-terminus of the DBD that is also a feature of other class
A4 HSFs. Strikingly, the intron in the DNA encoding the
MsHSF DBD, that is located between amino acid residues TY-
GF in all other plant HSFs [9], is found in a different location, 6
aa residues downstream between VD–PE.
In general, the C-terminal halves of HSFs are the least
conserved portions of the protein; however, this region usually
contains recognizable transcriptional activation domains (AHA
motifs), a NES, and other blocks of conserved aa sequences. The
AHA2 transcription activation motif of MsHSFA4 shows strong
conservation to the consensus AHA, FWxxF/LF/I/L [15],
whereas AHA1 does not. Additionally, in the very extreme C-
terminus of MsHSFA4, there are two regions located upstream
from the NES that exhibit a high degree of sequence conservation
with other members of the A4 class. Perhaps these sequences
(EVQSERK and WWN) constitute additional hydrophilic and
hydrophobic patches for interactions with MsHSFA4 targets.
The transient expression study of MsHSFA4 in tobacco
mesophyll protoplasts demonstrated its capacity to activate HS
promoters as predicted by the presence of a typical HSF DBD,
an oligomerization domain and AHA motifs (Fig. 6). Its ability
to activate transcription was comparable to the Arabidopsis
thaliana A4 homologue, which is a much weaker transcrip-
tional activator than Arabidopsis thaliana A1a HSF, as
observed previously [14]. Transient expression assays using
Gal4 DBD fusion constructs showed that the weak activities of
Arabidopsis and alalfa HSFA4 were largely due to the presence
of weak AHA activation motifs located in the C-terminal half of
these proteins.
The expression studies in tobacco protoplasts raised the
possibility that the long native 50 UTR of MsHSFA4 may play a
regulatory role in expression, since the construct containing
these sequences had no transcriptional activity. Since the
protein product produced from the open reading frame was
functional, we can dismiss the possibility of post-translational
regulation. The inhibitory effect of the 50UTR may occur at the
level of transcription or translation, but there is insufficient data
to suggest its true mode of action. It is, however, possible that
the effect is caused by the context of the tobacco system as
opposed to alfalfa. Additional studies using the same
constructs, but in alfalfa protoplasts, are warranted.
Northern analysis revealed that the MsHSFA4 gene was not
expressed under non-stress conditions and induced by heat
shock, a pattern that is also seen in AtHSFA4a expression
(unpublished). However, MsHSFA4 appears to also be actively
transcribed under cold stress conditions in growth chambers
and under low temperature conditions in the field (Fig. 7).
Although the specific role of MsHSFA4 in activating heat shock
gene expression under cold stress conditions is unknown at the
present time, our results, in combination with similar reports in
the literature of HSP gene expression under low temperature
stress [32,33,38,43], suggest that this alfalfa HSF is an integral
component of the low temperature stress response. Further-
more, at least one report indicates that HSP expression may
enhance chilling tolerance in plants [51].
There are several transcription factors involved in cold
responsive mechanisms in plants. The CBF/DREB1 proteins, a
family of Arabidopsis transcription factors, have been
determined to control a regulon of cold-induced (COR) genes
that enhance plant freezing tolerance (reviewed by [52]). The
CBF transcription factors provide example that innate systems
exist in plants that respond specifically to cold stress and
regulate a set of genes that provide cellular protection during
low temperature stress. Our report begins to expand on cold
response mechanisms by involving aspects of the heat shock
response. Many HSP genes are up regulated in response to high
temperature stress and some to low temperature stress.
However, there is very little known about how HSFs are
induced by cold stress and how they may function to protect or
acclimate plants for coping with low temperature stress.
Interestingly, it has been shown in Drosophila that HSF can be
expressed in several alternatively spliced isoforms (dHSFb, c
and d). The ratio of dHSFb is increased upon heat shock, while
that of dHSFd is enhanced upon exposure to cold [53].
Our report provides the first evidence of plant HSF A4 gene
induction by a cold stress. It is necessary to further investigate
the activity of MsHSFA4 in the context of other HSP and HSF
expression since multiple HSFs tend to work together to
activate and attenuate the HS response. It is possible that
MsHSFA4 may regulate a subset of HSP and other genes that
have evolved to enhance survivability to chilling injury.
Acknowledgements
This project was supported in part by the National Science
and Engineering Research Council of Canada (NSERC) and
Ontario Forage Council. Additionally, this project was
supported in part by the Florida Institute of Food and
Agricultural Sciences (IFAS) and USDA Grant 9500959 to
W.B.G. and E.C.-V.

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344 343
Reference
[1] E. Czarnecka-Verner, M.D. Barros, W.B. Gurley, Regulation of heat shock
gene expression, in: A.S. Basra (Ed.), Stress-induced Gene Expression in
Plants, Harwood Academic Publishers, Switzerland, 1994, pp. 131–161.
[2] H.R.B. Pelham, A regulatory upstream promoter element in the Droso-
phila hsp70 heat shock gene, Cell 30 (1982) 517–528.
[3] L. Nover, Expression of heat shock genes in homologous and heterologous
systems, Enzyme Microb. Technol. 9 (1987) 130–144.
[4] M.D. Barros, E. Czarnecka, W.B. Gurley, Mutational analysis of plant heat
shock promoter, Plant Mol. Biol. 19 (1992) 665–675.
[5] E. Czarnecka-Verner, C.X. Yuan, P.C. Fox, W.B. Gurley, Isolation and
characterization of six heat shock transcription factor genes from soybean,
Plant Mol. Biol. 29 (1995) 37–51.
[6] L. Nover, K.-D. Scharf, D. Gagliardi, P. Vergne, E. Czarnecka-Verner,
W.B. Gurley, The Hsf world: classification and properties of plant heat
stress transcription factors, Cell Stress Chaperones 1 (1996) 215–223.
[7] E. Czarnecka-Verner, W.B. Gurley, Plant heat shock transcription factors:
divergence in structure and function, Biotechnologia 3 (1999) 125–142.
[8] E. Czarnecka-Verner, S. Pan, C.-X. Yuan, W.B. Gurley, Functional
specialization of plant class A and B HSFs, in: J.H. Cherry (Ed.), Plant
Tolerance to Abiotic Stresses in Agriculture: Role of Genetic Engineering,
Kluwer Academic Publishers, The Netherlands, 2000, pp. 3–28.
[9] L. Nover, K. Bharti, P. Doring, S.K. Mishra, A. Ganguli, K.-D. Scharf,
Arabidopsis and the heat stress transcription factor world: how many heat
stress transcription factors do we need? Cell Stress Chaperones 6 (2001)
177–189.
[10] E. Treuter, L. Nover, K. Ohme, K.D. Scharf, Promoter specificity and
deletion analysis of three heat stress transcription factors of tomato, Mol.
Gen. Genet. 240 (1993) 113–125.
[11] J.H. Lee, A. Hubel, F. Schoffl, Derepression of the activity of genetically
engineered heat shock factor causes constitutive synthesis of heat shock
proteins and increased thermotolerance in transgenic Arabidopsis, Plant J.
8 (1995) 603–612.
[12] A. Hubel, J.H. Lee, C. Wu, F. Schoffl, Arabidopsis heat shock factor is
constitutively active in Drosophila and human cells, Mol. Gen. Genet. 248
(1995) 136–141.
[13] E. Czarnecka-Verner, C.-X. Yuan, K.-D. Scharf, G. Englich, W.B. Gurley,
Plants contain a novel multi-member class of heat shock factors without
transcriptional activation potential, Plant Mol. Biol. 43 (2000) 459–471.
[14] E. Czarnecka-Verner, W.B. Gurley, Arabidopsis class A and B HSFs show
a spectrum of transcriptional activity, Biotechnologia 3 (2002) 15–27.
[15] S. Kotak, M. Port, A. Ganguli, F. Bicker, P. von Koskull-Doring, Char-
acterization of C-terminal domains of Arabidopsis heat stress transcription
factors (Hsfs) and identification of a new signature combination of plant
class A Hsfs with AHA and NES motifs essential for activator function
and intracellular localization, Plant J. 39 (2004) 98–112.
[16] C. Lohmann, G. Eggers-Schumacher, M. Wunderlich, F. Schoffl, Two
different heat shock transcription factors regulate immediate early expres-
sion of stress genes in Arabidopsis, Mol. Genet. Genomics 271 (2004)
11–21.
[17] S.K. Mishra, J. Tripp, S. Winkelhaus, B. Tschiersch, K. Theres, L. Nover,
K.-D. Scharf, In the complex family of heat stress transcription factors,
HsfA1 has a unique role as master regulator of thermotolerance in tomato,
Genes Dev. 16 (2002) 1555–1567.
[18] K.-D. Scharf, H. Heider, I. Hohfeld, R. Lyck, E. Schmidt, L. Nover, The
tomato Hsf system: HsfA2 needs interaction with HsfA1 for efficient
nuclear import and may be localized in cytoplasmic heat stress granules,
Mol. Cell. Biol. 18 (1998) 2240–2251.
[19] P. Doring, E. Treuter, C. Kistner, R. Lyck, A. Chen, L. Nover, The role of
AHA motifs in activator function of tomato heat stress transcription
factors HsfA1 and HsfA2, Plant Cell 12 (2000) 265–278.
[20] E. Czarnecka-Verner, S. Pan, T. Salem, W.B. Gurley, Plant class B HSFs
inhibit transcription and exhibit affinity for TFIIB and TBP, Plant Mol.
Biol. 56 (2004) 57–75.
[21] E. Czarnecka-Verner, C.-X. Yuan, L. Nover, K.-D. Scharf, G. Englich,
W.B. Gurley, Plant heat shock transcription factors: positive and negative
aspects of regulation, Acta Physiol. Plantarum 19 (1998) 529–537.
[22] K. Bharti, P. von Koskull-Doring, S. Bharti, P. Kumar, A. Tintschl-
Korbitzer, E. Treuter, L. Nover, Tomato heat stress transcription factor
HsfB1 represents a novel type of general transcriptional coactivator with a
histone-like motif interacting with the plant CREB binding protein
ortholog HAC1, Plant Cell 16 (2004) 1521–1535.
[23] F.F. Damberger, J.G. Pelton, C.J. Harrison, H.C. Nelson, D.E. Wemmer,
Solution structure of the DNA-binding domain of the heat shock tran-
scription factor determined by multidimentional heteronuclear magnetic
resonance spectroscopy, Protein Sci. 3 (1994) 1806–1821.
[24] K.L. Clark, E.D. Halay, E. Lai, S.K. Burley, Co-crystal structure of the
HNF-3/fork head DNA-recognition motif resembles histone H5, Nature
364 (1993) 412–420.
[25] K.E. Flick, L.J. Gonzales, C.J. Harrison, H.C.M. Nelson, Yeast heat shock
transcription factor contains a flexible linker between the DNA-binding
and trimerization domains, J. Biol. Chem. 269 (1994) 12475–12481.
[26] P.K. Sorger, H.C. Nelson, Trimerization of a yeast transcriptional activator
via a coiled-coil motif, Cell 59 (1989) 807–813.
[27] R. Peteranderl, H.C. Nelson, Trimerization of the heat shock transcription
factor by a triple-stranded alpha-helical coiled-coil, Biochemistry 31
(1992) 12272–12276.
[28] R. Peteranderl, M. Rabenstein, Y.-K. Shin, C.W. Liu, D.E. Wemmer, D.S.
King, H.C.M. Nelson, Biochemical and biophysical characterization of
the trimerization domain from the heat shock transcription factor, Bio-
chemistry 38 (1999) 3559–3569.
[29] L.A. Sheldon, R.E. Kingston, Hydrophobic coiled-coil domains regulate
the subcellular localization of human heat shock factor 2, Genes Dev. 7
(1993) 1549–1558.
[30] S.K. Rabindran, R.I. Haroun, J. Clos, J. Wisniewski, C. Wu, Regulation of
heat shock factor trimer formation: role of a conserved leucine zipper,
Science 259 (1993) 230–240.
[31] D. Heerklotz, P. Doring, F. Bonzelius, S. Winkelhaus, L. Nover, The
balance of nuclear import and export determines the intracellular dis-
tribution and function of tomato heat stress transcription factor HsfA2,
Mol. Cell. Biol. 21 (2001) 1759–1768.
[32] M. Cabane, P. Calvet, P. Vincens, A.M. Boudet, Characterization of
chilling-acclimation-related proteins in soybean and identification of
one as a member of the heat shock protein (HSP70) family, Planta 190
(1993) 346–353.
[33] P. Krishna, M. Sacco, J.F. Cherutti, S. Hill, Cold-Induced accumulation of
hsp90 transcripts in Brassica napus, Plant Physiol. 107 (1995) 915–923.
[34] A. Soto, I. Allona, C. Collada, M.A. Guevara, R. Casado, E. Rodriguez-
Cerezo, C. Aragoncillo, L. Gomez, Heterologous expression of plant small
heat-shock protein enhances Escherichia coli viability under heat and cold
stress, Plant Physiol. 120 (1999) 521–528.
[35] Q.B. Li, D.W. Haskell, C.L. Guy, Coordinate and non-coordinate expres-
sion of the stress 70 family and other molecular chaperones at high and
low temperature in spinach and tomato, Plant Mol. Biol. 39 (1999) 21–34.
[36] N. Ukaji, C. Kuwabara, D. Takezawa, K. Arakawa, S. Yoshida, S.
Fujikawa, Accumulation of small heat-shock protein homologs in the
endoplasmic reticulum of cortical parenchyma cells in mulberry in
association with seasonal cold acclimation, Plant Physiol. 120 (1999)
521–528.
[37] D.K. Kadyrzhanova, K.E. Vlachonasios, P. Ververidis, D.R. Dilley, Mole-
cular cloning of a novel heat induced/chilling tolerance related cDNA in
tomato fruit by use of mRNA differential display, Plant Mol. Biol. 36
(1998) 885–895.
[38] A. Sabehat, S. Lurie, D. Weiss, Expression of small heat-shock proteins at
low temperatures. A possible role in protecting against chilling injuries,
Plant Physiol. 117 (1998) 651–658.
[39] M.E. Saltveit, Heat shocks increase the chilling tolerance of rice (Oryza
sativa) seedling radicles, J. Agric. Food Chem. 50 (2002) 3232–3235.
[40] M.E. Saltveit, P.K. Hepler, Effect of heat shock on the chilling sensitivity
of trichomes and petioles of African violet (Saintpaulia ionantha),
Physiol. Plant 121 (2004) 35–43.
[41] A. Soto, I. Allona, C. Collada, M.A. Guevara, R. Casado, E. Rodriguez-
Cerezo, C. Aragoncillo, L. Gomez, Heterologous expression of a plant
small heat-shock protein enhances Escherichia coli viability under heat
and cold stress, Plant Physiol. 120 (1999) 521–528.

J.N. Friedberg et al. / Plant Science 171 (2006) 332–344344
[42] M.A. Lopez-Matas, P. Nunez, A. Soto, I. Allona, R. Casado, C. Collada,
M.A. Guevara, C. Aragoncillo, L. Gomez, Protein cryoprotective activity
of a cytosolic small heat shock protein that accumulates constitutively in
chestnut stems and is up-regulated by low and high temperatures, Plant
Physiol. 134 (2004) 1708–1717.
[43] C.L. Guy, D. Haskell, Q.B. Li, Association of proteins with the stress 70
molecular chaperones at low temperature: evidence for the existence of
cold labile proteins in spinach, Cryobiology 36 (1998) 301–314.
[44] F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A.
Smith, K. Struhl, Current Protocols in Molecular Biology, Greene
Publishing Associates and Wiley-Interscience, Toronto, Ont., Canada,
1989.
[45] R.U. Schenk, A.C. Hildebrandt, Medium and techniques for induction and
growth of monocotyledonous and dicotyledonous plant cell cultures, Can.
J. Bot. 50 (1972) 199–204.
[46] G.R. Bauchan, M.A. Hossain, Karyotypic analysis of C-banded chromo-
somes of diploid alfalfa: Medicago sativa ssp. caerulea and ssp. falcata
and their hybrid, J. Hered. 88 (1997) 533–537.
[47] G.R. Bauchan, M.A. Hossain, Karyotypic analysis of N-banded chromo-
somes of diploid alfalfa: Medicago sativa ssp. caerulea and ssp. falcata
and their hybrid, J. Hered. 89 (1998) 191–193.
[48] U. Yamanouchi, M. Yano, H. Lin, M. Ashikari, K. Yamada, A rice spotted
leaf gene, Spl7, encodes a heat stress transcription factor protein, Proc.
Natl. Acad. Sci. 99 (2002) 7530–7535.
[49] D. Higgins, J. Thompson, T. Gibson, J.D. Thompson, D.G. Higgins, T.J.
Gibson, CLUSTAL W: improving the sensitivity of progressive multiple
sequence alignment through sequence weighting, position-specific gap
penalties and weight matrix choice, Nucleic Acids Res. 22 (1994) 4673–
4680.
[50] J.C. Corton, E. Moreno, S.A. Johnston, Alterations in the GAL4 DNA-
binding domain can affect transcriptional activation independent of DNA
binding, J. Biol. Chem. 273 (1998) 13776–13780.
[51] L. Wang, C.M. Zhao, Y.J. Wang, J. Liu, Overexpression of chloroplast-
localized small molecular heat-shock protein enhances chilling tolerance
in tomato plant, J. Plant Physiol. Mol. Biol. 31 (2005) 167–174.
[52] M.F. Thomashow, Plant cold acclimation: freezing tolerance genes and
regulatory mechanisms, Annu. Rev. Plant Physiol. Plant Mol. Biol. 50
(1999) 571–599.
[53] N. Fujikake, Y. Nagai, H.A. Popiel, H. Kano, M. Yamaguchi, T. Toda,
Alternative splicing regulates the transcriptional activity of Drosophila
heat shock transcription factor in response to heat/cold stress, FEBS Lett.
579 (2005) 3842–3848.