iron deprivation in cyanobacteria
TRANSCRIPT

Journal of Applied Phycology 6: 199-210, 1994. 199( 1994 Kluwer Academic Publishers. Printed in Belgium.
Iron deprivation in cyanobacteria
Fernando Ferreira & Neil A. Straus*Department of Botany, University of Toronto, Toronto, Ontario, Canada M5S 3B2(*Author for correspondence; phone (416) 978-3532; fax (416) 978-5878; E-mail: [email protected])
Received 30 August 1993; revised 10 January 1994; accepted 12 January 1994
Key words: cyanobacteria, iron deficiency, photosynthesis, siderophores, flavodoxin
Abstract
Iron is an essential component of electron transport in almost all living organisms. It is particularly important tophototrophs like cyanobacteria because 22-23 irons are required for a complete functional photosynthetic apparatus.Since the low solubility of Fe ++ above neutral pH in oxic ecosystems severely limits the biological availability ofiron to aquatic microorganisms, cyanobacteria and other microbes have developed a number of responses to copewith iron deficiency. Cyanobacterial responses to iron stress include the synthesis of an efficient, siderophore-basedsystem to scavenge iron and the substitution of ferredoxin with flavodoxin. An additional response in cyanobacteriainvolves the alteration of the light-harvesting apparatus that includes the appearance of a new, iron-stress-induced,photosystem II, chlorophyll-binding protein. Although cytochrome c-553 has a potential non-iron-containingreplacement in plastocyanin, a copper-containing protein, iron stress appears to favor the utilization of cytochromec-553 because siderophores also bind copper and form a complex that is excluded from the cell.
This paper is intended primarily as a review of molecular and physiological responses of actively growingcyanobacterial cultures to conditions of iron stress, where iron is present but essentially insoluble, and to differentiatethese responses from iron starvation, where the amount of iron in the system is not sufficient for cell growth.
Introduction
Since iron is the fourth most abundant element byweight in the earth's crust, one would not expectiron stress to be a recurring problem to living organ-ism. However, in aqueous oxic environments Fe++is quickly oxidized to Fe+ + +, and, at physiologicalpH's, Fe+ + + forms highly insoluble hydroxides. Forexample, the free Fe+++ concentration at pH 7 is onlyabout 103 ions per ml (Braun et al., 1990). Since a sin-gle bacterial cell (Escherichia coli) is estimated to con-tain about 106 iron ions (Archibald, 1983), an activelygrowing bacterial culture of 109 cells ml-' suffers anincredible deficit of iron when compared to the solubleiron ions in the environment.
Cyanobacteria and other bacteria have developedtwo basic responses to maintain growth under iron-stress conditions. In one response, they greatly im-prove their ability to scavenge iron by synthesizingsiderophores that solubilize iron as siderophore-iron
complexes. The siderophore-iron complexes are boundby specific receptor proteins in the outer membrane andthen passed into the cytoplasm by transport proteins inthe cytoplasmic membrane (reviewed by Braun & Han-tke, 1991). In the other response, the cellular demandfor iron is reduced by altering the population of proteinsso as to reduce the number of proteins that contain ironor whose synthesis requires iron. In cyanobacteria andother bacteria, the mobile electron carrier flavodoxinreplaces iron-sulfur-containing ferredoxin under con-ditions of iron deficiency (Smillie, 1965a, b; Hutber etal., 1977). In unicellular cyanobacteria, the gene forflavodoxin was shown to be the second open-readingframe of a dicistronic operon whose expression wastightly regulated by iron (Laudenbach et al., 1988;Leonhardt & Straus, 1992). The first gene, isiA, codesfor a protein that is very similar to CP43, a chlorophyll-binding core-protein of photosystem II (PS II) (Lau-denbach & Straus, 1988; Leonhardt & Straus, 1992)and forms an iron-stress-induced, chlorophyll-binding

200
protein complex (Burnap et al., 1993). Under ex-treme iron stress, cyanobacteria show reduced levels ofchlorophyll, phycobilisomes, iron-sulfur centers, andother components of the photosynthetic electron trans-port chain (Sandmann & Malkin, 1983, Sandmann,1985). This paper presents an overview of iron stressin cyanobacteria and discusses implications of the ironstress response to the regulation of gene expression incyanobacteria.
Discussion
Iron in photosynthetic electron transport
Cyanobacteria are obligate phototrophs. Iron playsa major role in the photosynthetic electron transportchain in cyanobacteria and is an essential componentof every membrane-bound protein complex of the lightreaction.
The organization and function of the componentsof PS II and its associated light-harvesting complexeshave been recently reviewed in detail for cyanobac-teria and eukaryotes (Vermaas & Ikeuchi, 1991). Themajor protein components of PS II include: the re-action center proteins D1 and D2, the chlorophyll-binding proteins CP47 and CP43, and cytochromeb-559. Primary charge separation has been observedin D1-D2-cytochrome b-559 complexes isolated fromchloroplasts and cyanobacteria, indicating that thechlorophyll-binding proteins CP47 and CP43 are prob-ably antenna proteins responsible for the transfer oflight energy to P680. However, both CP43 and CP47appear to be required for stable PS II assembly in thecell, since mutants of Synechocystis PCC 6803 thatlack either CP47 or CP43 are not able to grow pho-toautotrophically (Vermaas et al., 1988; Yu & Ver-maas, 1990; Rtgner et al., 1991). Cytochrome b-559is a heterodimer of two polypeptides that bind a singleheme. Cytochrome b-559 appears to play an essen-tial role in photosynthesis since Synechocystis PCC6803 mutants with deletions in the genes encodingcytochrome b-559 lack PS II activity (Pakrasi et al.,1988, 1989). Although there has been some debate asto whether one or two cytochromes b-559 are in thecore complex, the prevailing opinion favors two cy-tochromes per P680 (Vermaas & Ikeuchi, 1991). PSII also contains a non-heme iron between the primaryelectron acceptor QA, which is bound to D2, and QB,the quinone that binds to D1 but dissociates upon com-plete reduction. The function of this iron is not known
and no redox turnover has been observed under physi-ological conditions. However, its presence is essentialsince extraction or replacement with other ions resultsin a loss of PS II activity (Vermaas & Ikeuchi, 1991).Therefore, three iron atoms appear to be necessary fora functional PS II complex.
The electrons of reduced QB are passed to thecytochrome b6lf complex. The complex functions asan electrogenic proton translocator; it passes elec-trons from plastoquinone to either plastocyanin or cy-tochrome c-553 and translocates protons across thethylakoid membrane in the process, thus contributingto the protonmotive force that drives ATP synthesis(Hurt et al., 1982). It consists of four major proteins:cytochrome b6 which contains two hemes, cytochromefwhich is a c type cytochrome with one heme, a ReiskeFeS protein that has a 2Fe-2S center, and a 17-kDpolypeptide frequently referred to as subunit IV. Fi-nally, photosystem I (PS I) receives electrons fromreduced plastocyanin or cytochrome c-553 to catalyzethe photo-reduction of ferredoxin. The structure andfunction of this complex has been reviewed by Chitnisand Nelson (1991). PS I contains 7-8 protein subunitsand three 4Fe-4S centers (Fx, FA and FB) that partic-ipate in electron transfer within the complex.
Thus, under iron replete conditions, 22-23 ironatoms are needed for a functional photosynthetic ap-paratus in cyanobacteria under normal nutrient condi-tions. The positions of these irons are shown in Fig.1. PS II requires 3 irons, the cytochrome b6/lf com-plex has 5 irons, PS I contains 12 irons, and ferre-doxin has 2 irons in its single 2Fe-2S center. Thepresence of the 23rd iron depends on whether plas-tocyanin or cytochrome c-553 is being used. Sincecytochrome c-553 and ferredoxin are mobile electroncarriers that act as sinks for electrons passed throughthe membrane-bound photosynthetic complexes, onewould not expect a stoichiometric relationship to existbetween these carriers and one complete photosyn-thetic complex. In fact, the mobile carriers exist aspools in the cells, so that 23 irons per functioningphotosynthetic unit is probably a gross underestimate.Certainly ferredoxin would be expected to form a sub-stantial pool because it is used as the reducing agent ina multitude of cellular reactions.
Iron stress and photosynthesis
Iron deficiency has been shown to induce a number ofrecurring responses in the photosynthetic physiologyof cyanobacteria. In Synechococcus PCC 7002, Syne-

201
Ferredoxin (2 irons)(2Fe-2S1
Plastoquinone
Cyt b6/f complexey 6 (2 irons)
Veiie 2Fe2S (2 irons)(total = 5 irons)
PlastocyaninCyt 53 ( iron)rCy c553 (1 iron)
PS IFx.FA, FB3 X (4Fe-4S)(12 irons)
PS IID1 & D2 + non-hemne iron2 X y b559 (2 irons)CP4CP43(total=3 irons)
Total iron = 22-23
cytochrome c553 and ferredoxin are found in pools
Fig. 1. A diagrammatic representation of photosynthesis,indicating the locations of iron in the process.
chococcus PCC 7942 and Aphanocapsa PCC 6714,there are decreases in the amount of phycocyanin andchlorophyll (Hardie et al., 1983a; Guikema & Sher-man, 1983; Sandmann, 1985). Ultrastructurally, theselosses in S. PCC 7002 and S. PCC 7942 are paralleledby decreases in the amounts of phycobilisomes andmembranes (Hardie et al., 1983b; Sherman & Sher-man, 1983). In addition, all the iron-containing com-ponents of photosynthetic electron transport that havebeen examined in A. PCC 6714 - including cytochromeb-559 of PS II, cytochromes b andf of the cytochromeb6/fcomplex, and all the Fe-S centers (especially thoseof PS I) - show iron-stress-induced decreases (Sand-mann & Malkin, 1983; Sandmann, 1985).
Iron stress also results in qualitative changes tothe light-harvesting structures of Synechococcus PCC7942 and PCC 6301. These include large increases inoriginal and maximal fluorescence at room tempera-ture and dramatic alterations in the low temperaturefluorescence profile (Oquist, 1974a; Guikema & Sher-man, 1983; Guikema, 1985). Absorption studies onwhole cells and phycocyanin-free lamellae display ablue shift of 5-6 nanometers in the main red chloro-phyll absorption band (Oquist, 1971, 1974b; Guikema& Sherman, 1983). These spectral shifts are paral-leled by the appearance of an iron-stress-induced PSII chlorophyll-protein complex, CPVI-4, with absorp-
tion and fluorescence characteristics similar to those re-ported for whole cells under iron stress; this complexcontains a new 34 kDa, chlorophyll-binding protein(Pakrasi et al., 1985; Riethman & Sherman, 1988).
Iron-stress conditions were also shown to inducethe synthesis of flavodoxin in Anacystis nidulans(Smillie, 1965a, 1965b). Subsequently, it was observedthat, in Nostoc MAC and in Aphanocapsa 6714, theappearance of flavodoxin is accompanied by a paralleldecrease in the amount of ferredoxin (Hutber et al.,1977; Sandmann & Malkin, 1983). Flavodoxin hasbeen shown to be a functional alternative to ferredoxinin the photoreduction of NADP (Entsch & Smillie,1972) and in the donation of electrons to nitrogenase(Fillat et al., 1988; Sandmann et al., 1990), and isthought to be a general substitute for ferredoxin in ahost of reactions in iron-stressed cells (Rogers, 1987).
Iron-regulated genes involved in photosynthesis
The genesforfiavodoxin andferredoxin:The genes for flavodoxin and ferredoxin I have beencloned fiom a number of cyanobacteria. In the uni-cellular cyanobacteria, Synechococcus PCC 7942 andSynechococcus PCC 7002, the gene for flavodoxin,petl (also called isiB), is the second-open reading framein an operon whose transcription is tightly regulatedby iron (Laudenbach et al., 1988; Leonhardt & Straus,1992). The first gene, isiA, codes for a protein that isvery similar to CP43 (Laudenbach & Straus, 1988).Two transcripts are produced in cells grown in iron-deficient media: an abundant, monocistronic messagethat only contains isiA and a much less abundant di-cistronic message. Neither of these messages is de-tectable in cells grown in iron-replete medium. If ironis added to actively growing cultures, both of thesemessages, are barely detectable within a period of onehour. We have also found that these genes are presentand are arrTanged in the same way in another unicellularcyanobacterium, Synechocystis PCC 6803 (Ferreira &Straus, unpublished data).
In Anabaena PCC 7120, petl and isiA are on sepa-rate operons (Leonhardt & Straus, 1994). In this organ-ism, petI is the first gene of a dicistronic operon; thesecond open-reading frame, ORF2, contains a gene ofunknown function (Leonhardt & Straus; 1989; Leon-hardt & Straus, EMBL accession # X14577). Thisoperon produces messages of three size categories, anabundant monocistronic message containing only petl,0.9 kb, and much less abundant messages of 1.5 kb
I

202
and 1.8 kb containing both genes. The 0.9 kb andthe 1.8 kb messages are under the control of an iron-regulated promoter. The third message is constitutivelyproduced from a separate promoter, P2 (Leonhardt &Straus, in press). The constitutive expression of thisgene is in agreement with a report by Sandmann et al.(1990) that indicated constitutive production of flavo-doxin in heterocysts of all nitrogen-fixing, heterocys-tous cyanobacteria at high iron concentrations. Figure2 compares the organization of the operons containingisiA and petI from unicellular cyanobacteria and fromAnabaena PCC 7120. In Anabaena PCC 7119, onlya monocistronic message has been reported when ironwas limiting and no detectable message was seen underiron-replete conditions (Fillat et al., 1991). Since flavo-doxin has been detected in nitrogen-fixing cyanobac-teria in iron-replete conditions, at least some of thedifference in the transcript levels of flavodoxin be-tween PCC 7119 and PCC 7120 could be accountedfor if the method of RNA isolation used by Fillat etal. (1991) did not rupture heterocysts. However, thedifferences in the number of transcripts are difficult toexplain away because there is no difference betweenthe reported amino acid sequences of flavodoxin, indi-cating that the strains are very closely related. In anycase, these differences will have to be examined inmore detail to see if the strains do have different genearrangements outside of petI and different modes ofgene regulation.
In E. coli and other bacteria the expression of iron-regulated genes is controlled by a repressor protein,Fur, that binds to specific sequences in the promoterregion of the operon and stops transcription in iron-replete conditions (for review, see Braun & Hantke,1991). An examination of the sequences upstream fromthe transcriptional start of those flavodoxin genes thathave been sequenced reveals a series of sequencesthat share similarities with the Fur-binding consen-sus sequences of iron-regulated genes in E. coli andother bacteria (Laudenbach et al., 1988; Leonhardt &Straus, 1992; Leonhardt & Straus, EMBL accession# X14577). It is generally assumed that cyanobacteriaalso contain afur repressor gene that is responsible forthe regulation of iron stress genes.
Experiments in our laboratory indicate that tran-script levels of ferredoxin I are similar in iron-deficientand iron-replete media for all cyanobacteria examined,including the unicellular species Synechococcus PCC7942 and PCC 7002, and the filamentous species An-abaena PCC 7120 (Laudenbach et al., 1988; Leon-hardt & Straus, 1992; Leonhardt & Straus, in press).
A
IsA peti
t~~~~~~~~~~~-ic~
B
pet I ORF2
21
isiA1I.l
Fig. 2. The arrangement and transcription of IsiA and petl invarious cyanobacteria: (A) Synechococcus PCC 7942, Syne-chococcus PCC 7002 and Synechocystis PCC 6803; (B) An-abaena PCC 7120.
Contradictory results have been published by the groupworking at the University of Utrecht. In agreement withour observations, an earlier paper showed little differ-ence in the level of ferredoxin I transcript from Syne-chococcus PCC 7942 in iron-stress and iron-repletemedia (Van der Plas et al., 1988). However, morerecently, they published results that indicate strikingreductions in the levels of the ferredoxin transcript inSynechococcus PCC 7942, and no detectable transcriptin Anabaena PCC 7937 from iron-deficient medium. Inthe more recent report, the amounts of ferredoxin tran-script returned to iron-replete levels within one hourof the addition of iron to the medium; the more recentobservations were attributed to differential mRNA sta-bility (Bovy et al., 1993). In any case, all of the ev-idence to date indicates that the appearance of ferre-doxin is regulated post-transcriptionally. However, thisissue should be reexamined to see if differences in theabundance of ferredoxin mRNA could be the result ofdifferent growing conditions.
In addition to vegetative cell ferredoxins, thenitrogen-fixing cyanobacteria Anabaena PCC 7937and PCC 7120 contain an additional heterocyst-specific ferredoxin that only appears to be expressed
1
f.r,
1
.n
1
r_

203
under nitrogen-fixing conditions (Schrautemeier &Bihme, 1985; Bohme & Schrautemeier, 1987; Bbhme& Haselkorn, 1988).
IsiA:The first open reading frames (isiA) of the flavodoxinoperons from Synechococcus PCC 7942 and Syne-chococcus sp. PCC 7002 code for polypeptides with agreat deal of similarity to CP43 (Laudenbach & Straus,1988; Leonhardt & Straus, 1992). Recently, the prod-uct of isiA was identified as the 34 kD protein of theiron-stress-induced, PS II, chlorophyll-protein com-plex, CPVI-4 (Burnap et al., 1993).
Many features of the deduced isiA protein sequenceindicate that it may have functions similar to thoseof CP43. All the hydrophobic regions of CP43 havebeen conserved in IsiA, indicating that both proba-bly have similar membrane-spanning structures. In ad-dition, the conservation of histidine residues in themembrane-spanning regions suggests similar chloro-phyll ligand capabilities (Laudenbach & Straus, 1988;Bricker, 1990; Vermaas & Ikeuchi, 1991). Figure 3compares the amino acid sequences for CP43 and IsiAfrom a number of cyanobacteria. Regions of high se-quence identity occur in all membrane-spanning re-gions and in lumenar loops A and C. The most strikingdifference between the putative isiA products and CP43consists of a large deletion in the fifth hydrophilic lu-menar loop, the E loop (Bricker, 1990).
The function of IsiA remains obscure. There is in-direct evidence that IsiA may have a functional role inPS II, because in Anabaena PCC 7120 isiA is located ina monocistronic operon whose transcription is tightlyregulated by iron (Leonhardt & Straus, 1994). If IsiAhad a PS II function, then the separation of petl andisiA would make sense in heterocystous nitrogen fixingcyanobacteria, where flavodoxin is needed as an elec-tron donor for nitrogenase, but proteins involved in theevolution of oxygen are detrimental because oxygeninactivates nitrogenase.
Because of the similarities between CP43 and IsiA,it is tempting to speculate that under iron stress IsiAmay compensate for some as yet unknown iron-relatedfunction of CP43. Since a number of mutants have beencreated for psbC in Synechocystis PCC 6803 (Vermaasetal., 1988; Yu & Vermaas, 1990; Rtgneretal., 1991),one should be able to use these mutants to see if IsiAcan functionally replace CP43 under iron stress. Wehave attempted to do this, by comparing oxygen evo-lution of iron-stressed and iron-replete wild-type and
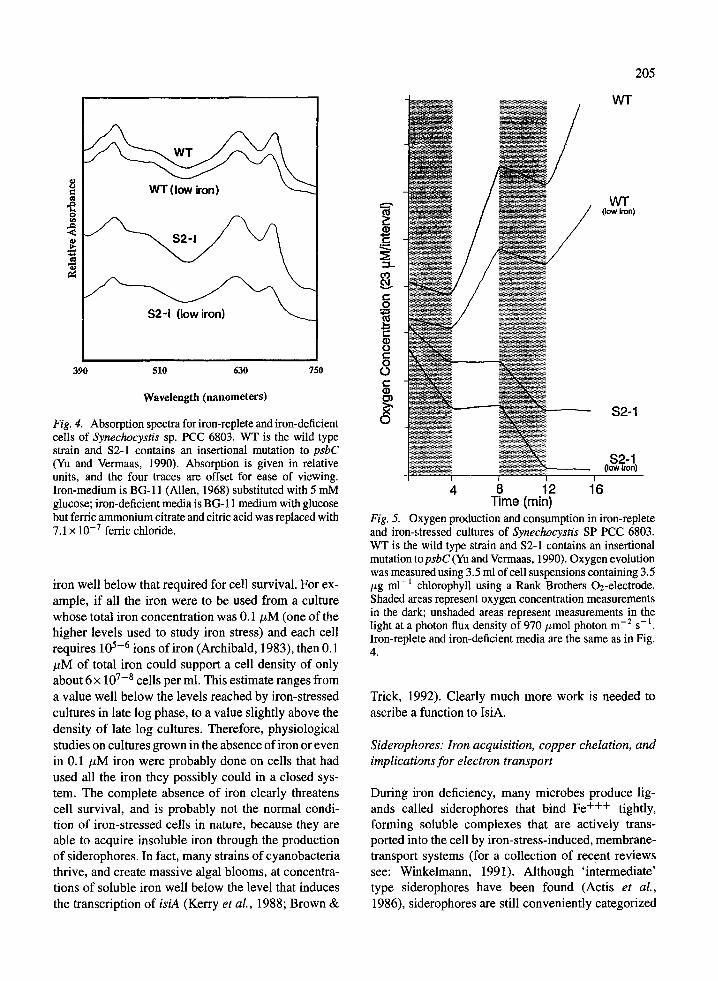
mutant cultures, including deletion and insertion mu-tants of psbC (Vermaas et al., 1988; Yu & Vermaas,1990) and a psbC start codon mutant in a phycocyanindeficient strain (Rogner et al., 1991). All experimentshad similar results (authors, unpublished). Figure 4shows the whole cell absorption spectra of wild-typecultures and cultures of the insertional mutant S2-1(Yu & Vermaas, 1990) for both iron-stressed and iron-replete conditions. Both wild-type and mutant cellsshow the blue shift of the main red chlorophyll absorp-tion peak that occurs during iron stress and probablycharacterizes the appearance of the CPVI-4 complex.Figure 5 shows the light and dark evolution, and con-sumption, of oxygen by these cells in iron-deficient andiron-replete conditions. Wild type cells evolved oxy-gen in the light under both iron-stress and iron-repleteconditions and consumed oxygen via respiration in thedark. Mutants of psbC were not able to evolve oxygenin the light although they did not appear to consumeit either; however, in the dark they respired quite ac-tively. The oxygen concentration curves for these PS IImutant cells indicate that these cells had not lost PS Iactivity; in the light oxygen was not consumed, indicat-ing that the electrons were diverted from cytochromeoxidase to PS I. Since a similar response was observedin a series of different CP43 mutants, it appears thatIsiA cannot functionally replace CP43 in iron-stressconditions.
Riethman and Sherman (1988) proposed two otherfunctions for the CPVI-4 complex. First, they proposedthat it might function as an auxiliary light-harvestingcomponent that compensates for phycobilisome lossduring iron stress; second, they proposed that it mayserve as a reservoir for chlorophyll that is used bythe cell during recovery from iron stress. Reductionand/or loss of phycobilisomes and phycobiliproteinshave been reported as a symptom of iron stress incyanobacteria (Hardie et al., 1983a; Sherman & Sher-man, 1983; Guikema & Sherman, 1983; Sandmann,1985). However, these losses are paralleled by lossesin chlorophyll and other components of photosyntheticelectron transport (Hardie et al., 1983a; Sandmann,1985) so that the overall ratio of phycocyanin to photo-synthetic centers may not be greatly reduced. Certainlythe absorption spectra of Fig. 4 for both wild type andmutant cells (where a direct comparison can be madebetween the phycocyanin peak, at approximately 625nm, and the last chlorophyll peak) shows that there isnot a disproportionate loss of phycocyanin during ironstress. Similar whole cell spectra have been reportedby others (Guikema & Sherman, 1983). Therefore the

204
N-terminus. STROMA C MEMBRANE I A-Loop LUMEN A-Loop [ MEMBRANE* ~~~***** *** *** ** * * *~ * * *** * * **** ** ****
7942 CP43 MVTLSSPSVIAGGRDIDSTGYAWW SGNARLNLGKLLGAHVAHAGL IVFWAGAMTLFEVAHFVPEKPMYEGILLSHLATLGWGVGPGGEVVTFPYFGVLHLISSAVLGLG
7002 CP43 .....N AP.S.....E...F...A....................................
6803 CP43 .....N TSSS....LP...F.....................R.............................L...P. ......... A...T....F.................
7120 CP43 .....N TL-G ....E.S.FAW .......................................I ....V....L...P.V .......V....I....................F.
7942 IsiA .Q.YNN.E.T D...A....FA....LFIA .... Q.A..M .... F..Y.ISWLTAOOSG. .. L. .P......L...D..Q.T .Y.L....AV ,.. ..... A.7002 IsiA .Q.YDN.D.K .E..A....FAO...OFI.......A........F....ISY.D.TL..G..NL...P......L.IEGN.AIN. E....I.AI ........ AA.6803 IsiA .Q.YGNDT.Q .E..A....FADQ..LFIA....Q.A.TA ..... F....ISR.D.TQA.GD..L ... P......W...D..Q]...Y....I.RI...A.....A.7120 IsiA .Q.YDN.N.K .D..A....FA....LFI. .... A.TTL....F.W..ISRYK....G...L...P.. F...VS.Q..N.Y ..... A .......... A.
] B-Loop STROM4A B-Loop [ MEMBRANE I C-Loop LUMEN C-Loop [MEMBRANE* * * * * *** ** **t* **l ** ** * ** *** * * ** * ** ***
7942 CP43 GIYHALRGPESLEEYSTF FSQOWKDKNQMTNI IGYM LILLGLGAFLLVFKAMFFGGVYDTWAPGGGDVR IISNPTLNPAVIFGYLLKSPFGGDGWI VSVDNLEDVIGGH IWIG
7002 CP43 ..... V....T..... . .GY .................... .L ............. ........................... F.A .... E.MIG. M.. ..... YV6803 CP43 .......... V.....S. .................. .... L ........................ V.T . ............. A....E.. ....... .7120 CP43 ..... V....T...... .GY ......K. .....F.I..C.L...L.................... V.T .................... E......M .......... A7942 IsiA ALF.TF.A.SD.AAA.GAAKR.HF.N.PK.LGL.L.H. .LF..V..L.. A..TTW..L.. A. ST..LVTE........Y.. QTH. ASI.....LV...VYV.7002 IsiA GLF.VL... OD.KTATGPARR.HF..E.PK.LGL.L.N.L .......... A...Y. .. L.. T TQT..LVTE........Y.. QTH. AT......IV .... YV.6803 IsiA ALF.TL.A.AD.STLKGQGKK.HFT.ENPQ.LGL.H.I .A.L. .AG...YW..L.. . TTQT..LV.Q...D.L..Y., QTH. ASISS...LV....FV.7120 IsiA ALF.TFK .RN.KNTTGSARK.HFE.N.PK.LGL.L.H. .LF..MA.L...G ... W..L.. TTQV..WVNH .... F..Y.. TH. AS.N....LV .... YV.
O-Loop STROMA MEMBRANE I E-Loop LUMEN E-Loop LUMEN****** * * *** ***** * * * * ** ***
7942 CP43 LICISGGIWHILTKPFWGVGRAF NGGEALSYSLAFTFLVRDALGAGAQPTGLGKYLRPTGE IFGETMR
7002 CP43 ....F.............AR..LV.S ...............A...TCF..F...A. ...N ........ LI..A...SM...V..................... . L.......
6803 CP43 .................. AR..L.S .................... VF..F...A.....Y...GM ............... R.....A................S...........
7120 CP43 ........... F......AR.....S....................C............ P... ..A .......R....V.................S...........
7942 IsiA VML.A.......VP..Q.TKKVL.YS...I......GIA.A..V.AYFCAV.TLA..V..Y APL.IKLGVTPY.ADTV.LPFGAH
7002 IsiA VLLVA .....VP.LO.AKKVLLFS... I.......IA.A..V.AYFCAV.T.A..V..Y. PVLDVKL.IVPY.ADTIELPMNEH
6803 IsiA FLLIG .......VP.L..AKKVLLFS... I......GIA.A..V.AYFCAV.TLA..P..Y. PPLAIKLGIFPY.AOTVELPMHAH
7120 IsiA ..L.G......VKE.LP.AKKLL.FS... I......GIA.A..V.AYFCAV.TLA..V..Y. APL.LKFGVTPY.ADTVKLADGGY
E-Loop LUMEN E-Loop C MEMBRANE STROK4A C-terminus** .* * ****dr** **
7942 CP43 FW FRGPCVEPLRGPNGLDLDKLTND IPWQARRAAEYMTHAPLGSLNSVGGATE INSVNFVSPRAWLATSPFVLAF FLVGLWHAGRARAAAAGFEKGIDRATEPVLAMRDLD
7002 CP43 ....... WL ............. IR....... L.... . ..... NA. ..... I..P. ... NL.Q...G.H...S.................E.........E.....N.NE..6803 CP43 ....... WL .............. R....... V........ ............. I.V..F.Y .......... ... G...........................E...T.F.P...7120 CP43 .. .. ........ . ...... .G..... ................. EN..AM..G...7942 IsiA T.....SNAH.F .. .C. ...... L..M .. DFRRVEKALSSVEA7002 IsiA TS.....NAH.FF.....Q......L..M .. DFRRVEKVLSDPLOA6803 IsiA TS.....NAH.F......Q......L..L .. DFKRVNKRLIPCKPSH7120 IsiA A.....NAH.F......I......L..I .VDFRQ.EKSLNAI'SSAE
Fig. 3. Amino acid sequence comparison of CP43 and IsiA from various cyanobacteria. Amino acids are given the single letterdesignation. A dot means that the amino acid is identical to that of the first listed sequence. Spaces represent a deletion. Theregions of the proteins that are located in the stroma, lumen and membrane are marked above the sequence. The designationsof the different hydrophilic loops are indicated above the sequence. Loci that have the same amino acid in all four proteinsequences are indicated with a *. The following are the source references for sequences of CP43: from Synechococcus sp.PCC 7942, Golden and Steams, 1988; from Synechococcus sp. PCC 7002, Gingrich et al., 1990; from Synechocystis sp. PCC6803, Chisholm and Williams, 1988; from Anabaena sp. PCC 7120, Leonhardt and Straus, 1994. The following are the sourcereferences for sequences of IsiA: from Synechococcus sp. PCC 7942, Laudenbach and Straus, 1988; from Synechococcus sp.PCC 7002, Leonhardt and Straus, 1992; from Synechocystis sp. PCC 6803, Ferreira and Straus, 1993 Genbank AccessionNumber L26530; from Anabaena sp. PCC 7120, Leonhardt and Straus, 1993.
proposal that IsiA is needed to compensate for losses for chlorophyll that is used in times of recovery fromin phycocyanin seems to be based on the misconcep- iron stress, seems to imply that iron-stressed cells aretion that phycocyanin is disproportionately decreased not healthy. This concept may have been influencedduring iron stress, by earlier work on iron stress that studied cells grow-
The second proposal, that IsiA serves as a reservoir ing in the absence of iron or at concentrations of total
205
0U
0*0
Ci
Ci
390 510 630 750
Wavelength (nanometers)
Fig. 4. Absorption spectra for iron-replete and iron-deficientcells of Synechocystis sp. PCC 6803. WT is the wild typestrain and S2-1 contains an insertional mutation to psbC(Yu and Vermaas, 1990). Absorption is given in relativeunits, and the four traces are offset for ease of viewing.Iron-medium is BG-11 (Allen, 1968) substituted with 5 mMglucose; iron-deficient media is BG- 11 medium with glucosebut ferric ammonium citrate and citric acid was replaced with7.1 x10 - 7 ferric chloride.
iron well below that required for cell survival. For ex-ample, if all the iron were to be used from a culturewhose total iron concentration was 0.1 atM (one of thehigher levels used to study iron stress) and each cellrequires 105-6 ions of iron (Archibald, 1983), then 0.1/tM of total iron could support a cell density of onlyabout 6 x 107-8 cells per ml. This estimate ranges froma value well below the levels reached by iron-stressedcultures in late log phase, to a value slightly above thedensity of late log cultures. Therefore, physiologicalstudies on cultures grown in the absence of iron or evenin 0.1 atM iron were probably done on cells that hadused all the iron they possibly could in a closed sys-tem. The complete absence of iron clearly threatenscell survival, and is probably not the normal condi-tion of iron-stressed cells in nature, because they areable to acquire insoluble iron through the productionof siderophores. In fact, many strains of cyanobacteriathrive, and create massive algal blooms, at concentra-tions of soluble iron well below the level that inducesthe transcription of isiA (Kerry et al., 1988; Brown &
WT
WT(low Iron)
S2-1
0)4-
c
a,0
ca,00C
0
S2-1(low Iron)
I4 8 12 16
Time (min)Fig. 5. Oxygen production and consumption in iron-repleteand iron-stressed cultures of Synechocystis SP PCC 6803.WT is the wild type strain and S2-1 contains an insertionalmutation topsbC (Yu and Vermaas, 1990). Oxygen evolutionwas measured using 3.5 ml of cell suspensions containing 3.5/tg ml-' chlorophyll using a Rank Brothers 02-electrode.Shaded areas represent oxygen concentration measurementsin the dark; unshaded areas represent measurements in thelight at a photon flux density of 970 /imol photon m-2 s-.Iron-replete and iron-deficient media are the same as in Fig.4.
Trick, 1992). Clearly much more work is needed toascribe a function to IsiA.
Siderophores: Iron acquisition, copper chelation, andimplicationsfor electron transport
During iron deficiency, many microbes produce lig-ands called siderophores that bind Fe+++ tightly,forming soluble complexes that are actively trans-ported into the cell by iron-stress-induced, membrane-transport systems (for a collection of recent reviewssee: Winkelmann, 1991). Although 'intermediate'type siderophores have been found (Actis et al.,1986), siderophores are still conveniently categorized

206
into two classes, hydroxamate-type and catechol-typesiderophores. In general catechol-type siderophoreshave a higher affinity for iron than the hydroxamate-type siderophores do.
Cyanobacterial research on siderophores has notapproached the complexity of that for other bacteria(Boyer et al., 1987). Although many siderophoreshave been characterized for other bacteria, onlythe hydroxymate-type siderophore, schizokinen, hasbeen structurally characterized from a cyanobac-terium (Simpson & Neilands, 1976). However, manycyanobacteria have been shown to be capable of pro-ducing and transporting the ferric siderophore com-plex, and other cyanobacteria that apparently cannotsynthesize siderophores are able to utilize siderophoresproduced by other bacteria. For example, of ten speciesexamined by Lange (1974), six could grow axenicallyunder iron stress-conditions. Although the remainingfour species could not grow in normal iron-deficientmedium, they were able to grow in iron-deficient fil-trates from the siderophore-producing strains. In fact,it has been suggested that the ability of cyanobacteriato sequester iron through the production and utiliza-tion of siderophores allows them to dominate eukary-otic algae during blue-green algal blooms (Estep etal., 1975; Murphy et al., 1976). Table 1 contains listsof cyanobacteria that are presumed to be capable ofsiderophore production by one or more tests. Chemicaltests include the Csaky test for hydroxamates (Csaky,1948; Gillam et al., 1981) and the catechol tests devel-oped by Arnow (1937) and Rioux etal. (1983). Biolog-ical tests include the ability to grow axenically underconditions of iron deprivation, the ability to promotegrowth of bacterial siderophore auxotrophs (Burnham& Neilands, 1961) and the ability to survive toxic levelsof copper in axenic iron-stress conditions (McKnight &Morel, 1980). Table 1 attempts to interpret the resultsof these disparate techniques in terms of siderophoreproduction; however, a table of species listed accord-ing to technique used is found in a review article byBoyer et al. (1987).
Siderophores are capable of binding to cupric ionsas well as ferric ions; however, cyanobacteria donot transport the copper-siderophore complex. There-fore, siderophore secretion has the effect of detoxi-fying medium containing toxic levels of copper(McKnight & Morel, 1980), and the ability of iron-stressed cyanobacteria to grow in the presence of toxiclevels of copper has been used as an indication ofsiderophore production (Boyer et al., 1987). Clarke etal. (1987) showed that the growth of Anabaena PCC
7120 was inhibited by the addition of 2.1 to 6.5 /uMcopper and abolished by concentrations above 10 /M.However, the addition of schizokinen to the mediawas able to alleviate the toxic effects of copper; in theabsence of schizokinen, copper was taken up by thecells, but when schizokinen was added to the mediumschizokinen-bound copper remained in the medium.
The exclusion of copper from iron-stressed cellshas other implications for photosynthetic electrontransport, since plastocyanin is a copper-containingmobile electron carrier between the cytochrome b6lfcomplex and PS I. McKnight and Morel (1980)have shown that during cyanobacterial blooms thesiderophore-copper complex is the dominant form ofcopper in the environment. Since the complexing ofcopper to siderophores during iron stress excludes itfrom the cell, high levels of siderophore accumulationduring iron stress could induce a condition of copperstress. This condition in turn would induce the syn-thesis of cytochrome c-553, an iron-containing mobileelectron carrier (Ho & Krogmann, 1984). The induc-tion of an iron-containing mobile electron carrier underconditions of iron stress does not appear to make muchsense on the surface; however, it may be a small priceto pay for the utilization of the siderophore system.Since cytochrome c-553 has a relatively limited func-tion, its pool may be small relative to that of ferredoxin,which is required for many essential cellular reactions.Therefore, the transition from ferredoxin to flavodoxinprobably represents a far greater economy in the uti-lization of iron than does the demand for iron createdby the induction of cytochrome c-553. In this contextit is significant that many species of cyanobacteria ap-pear to have lost the gene for plastocyanin and utilizecytochrome c-553 only in photosynthesis (Ho & Krog-mann, 1984; Sandmann, 1986). In fact, the creation ofan oxygen atmosphere by cyanobacteria, and the re-sultant competition for iron, may have selected forcyanobacterial strains that utilize only cytochrome c-553. In addition, although cytochrome c-553 is used bycyanobacteria to pass electrons from the cytochromeb6 /f complex to PS I, it does not appear to be anabsolute requirement for this function. In Synechococ-cus PCC 7942, which lacks the gene for plastocyanin,deletion mutants lacking cytochrome c-553 grow aswell as wild-type cells under normal light conditions(Laudenbach et al., 1990).

207
Table 1. Iron acquisition in Cyanobacteriaa" b
Strains Capable of High Affinity or Catechol Siderophore ProductionAnabaena sp. PCC 7937 (1), Oscillatoria tenuis UTEX 428 (2), Spirulina maxima(1), Spirulina platensis (1)
Strains Producing Siderophores (Probably Hydroxymates), but ApparentlyLacking High Affinity SiderophoresAgmenellum quadruplicatum UTEX 2268 (1); Anabaena sp. PCC 7120 (3),PCC7122 (3,4), UTEX 1611(3); Anacystis sp. UCB No. 69 (4); Chroococcus sp.PCC 6501(4); LPP group ATCC 29126 (1); Nostoc sp. PCC 6411 (4); Nostocmuscorum UTEX 1038 (1); Phormidium autumnale UTEX 1580 (1);Synechococcus sp. PCC 6307 (4), PCC 7002 (5), PCC 7003 (5), PCC 7942 (1)
Strains Producing Siderophores (Probably hydroxamates), Not Known IfHigh Affinity Siderophores PresentAnabaena cylindrica IU 629 (6), UTEX 1611 (7,8);Gloeocapsa alpicola No 6308 (7); Lyngbya sp. IU 487 (6);Microcystis aeruginosa W1036 (6), UTEX 2061 (7); A'ostoc muscorum W1013 (6);Phormidiumfoveolarum IU 427 (6); Synechococcus sp. PCC 6301 (8)
Strains Apparently Lacking Iron Scavenging SystemAnabaena circinalis W1038 (6); Anabaena sp. PCC 7938 (1); Aphanizomenonflos-aquae W1062 (6); Fischerella sp. PCC 73103 (1); Gloeotrichia echinulataW1052 (6); Nostoc sp. PCC 7524 (1); Oscillatoria rubescens EAWAG 18 clone51(6); Plectonema sp. PCC 6306 (1), Synechococcus sp. PCC 6301(1)
References: (1) Kerry et al. 1988; (2) Brown and Trick, 1992; (3) Goldman et al.,1983; (4) Simpson and Neilands, 1976; (5) Armstrong and van Baalen, 1979; (6)Lange, 1974; (7) McKnight and Morel, 1979; (8) McKnight and Morel, 1980.
a Species names have been converted to the Pasteur Culture Collection numberwhere possible.b The presence or absence of siderophores, and type of siderophore have beeninterpreted from disparate data including growth in media lacking a chelator, growthin iron minus medium, ability to compete with EDDA for iron, resistance to coppertoxicity while under iron stress, and chemical tests for siderophores.
Concluding remarks
A substantial amount of descriptive work has beendone on the effect of iron stress on cyanobacteria. Un-fortunately, most of this work has studied a terminalform of iron deprivation where iron is either totallyabsent from the media or is present in amounts toosmall to support cell growth. However, in the environ-ment, there is a naturally occurring condition of ironstress in which iron is actually present but is not avail-able because it is not in a soluble form that the cellcan readily use. Under these conditions, cyanobacteria(and other bacteria) respond to iron stress by producing
siderophores to acquire iron and by using flavodoxininstead of ferredoxin to reduce the cellular require-ments for iron. In unicellular cyanobacteria, the tran-scriptional regulation of the flavodoxin gene is tiedto the expression of a gene for a chlorophyll-bindingprotein, IsiA, with many similarities to CP43. Thefunction of this protein is not known at present; how-ever, it does not appear to functionally replace CP43under iron-stress conditions. More work is needed tounderstand the function of IsiA, keeping in mind thatthe pathways for chlorophyll and heme synthesis sharecommon intermediates up to protoporphyrin IX, andthat perturbations in the synthesis of heme may be felt

208
by connected pathways.On a more basic level, iron stress experiments have
to be redesigned to separate symptoms of iron starva-tion leading to cell death from iron stress leading tothe production of siderophores and a more economicuse of iron in the cell. One solution is the use of aniron chelator that is not toxic to the cell but forces
the cell to produce siderophores to scavenge iron fromthe chelator. With this strategy, Kerry et al. (1988)were able to divide cyanobacteria into three categories
by using ethylenediamine-di(o-hydroxyphenyl-aceticacid) (EDDA); these categories included cells with a
high affinity iron-scavenging system, cells with a rel-atively weak iron-scavenging system, and cells withno siderophore system. Two practical disadvantagesto this approach are the high cost of EDDA and the
difficulty in finding suppliers. A simpler and cheapersolution may be found in using iron-containing mediathat lack citrate and other chelators (Lange 1974).
Acknowledgements
We would like to acknowledge the excellent techni-
cal assistance of Bess Wong and Junyoung Park, andfinancial support from the Natural Sciences and Engi-neering Research Council of Canada.
References
Actis LA, Fish W, Crosa JH, Kellerman K, Ellenberger SR,Hauser FM, Sanders-Loher J (1986) Characterization ofanguibactin, a novel siderophore from Vibrio anguillarum775 (pJMl). J. Bact. 167: 57-65.
Allen M (1968) Simple conditions for growth of unicellularblue-green algae on plates. J. Phycol. 4: 1-3.
Archibald F (1983) Lactobacillus plantarum, an organismnot requiring iron. FEMS Microbiol. Lett. 19: 29-32.
Armstrong JE, van Baalen C (1979) Iron transport in microal-gae: the isolation and biological activity of a hydroxya-mate siderophore from the blue-green algae Agmenellumquadruplicatum. J. gen. Microbiol. 111: 253-262.
Arnow LE (1937) Colorimetric determination of the compo-nents of 3,4-dihydroxyphenylalanine-tyrosine mixtures.J. biol. Chem. 118: 531-537.
BOhme H, Haselkorn R (1988) Molecular cloning and nu-cleotide sequence analysis of the gene coding for hete-rocyst ferredoxin from the cyanobacterium Anabaena sp.strain PCC 7120. Mol. gen. Genet. 214: 278-285.
Bohme H, Schrautemeier B (1987) Electron donation to ni-trogenase in a cell-free system from heterocysts of An-abaena variabilis. Biochim. Biophys. Acta 891: 115-120.
Bovy A, de Vrieze G, Lugones L, van Horssen P, van denBerg C, Borrias WE, Weisbeek P (1993) Iron-dependent
stability of the ferredoxin I transcripts from cyanobacterialstrains Synechococcus species PCC 7942 and Anabaenaspecies PCC 7937. Mol. Microbiol. 7: 429-439.
Boyer GL, Gillam AH, Trick C (1987) Iron chelation anduptake. In: Fay P, Van Baalen C (eds), The Cyanobacteria.Elsevier, Amsterdam, New York & Oxford: 415-436.
Braun V, Hantke K (1991) Genetics of bacterial iron trans-port. In: Winkelmann G (ed.), CRC Handbook of micro-bial iron chelates. CRC Press, Boca Raton: 107-138.
Braun V, Schaffer S, Hantke K, Troger W (1990) Regulationof gene expression by iron. In: Hauska G, Thauer R (eds),The molecular basis of bacterial metabolism. Springer-Verlag, Berlin: 164-179.
Bricker TM (1990) The structure and function of CPa-1 andCPa-2 in photosystem II. Photosyn. Res. 24: 1-13.
Brown CM, Trick CG (1992) Response of the cyanobac-terium, Oscillatoria tenuis, to low iron environments: theeffect on growth rate and evidence for siderophore pro-duction. Arch. Microbiol. 157: 349-354.
Burnap RL, Troyan T, Sherman LA (1993) The highly abun-dant chlorophyll-protein complex of iron-deficient Syne-chococcus sp. PCC7942(CP43') is encoded by the isiAgene. Plant Physiol. 103: 893-902.
Burnham BF, Neilands JB (1961) Studies on the metabolicfunction of the ferrichrome compounds. J. biol. Chem.236: 554-559.
Chisholm D, Williams JGK (1988) Nucleotide sequence ofpsbC, the gene encoding the CP-43 chlorophyll a-bindingprotein of Photosystem II, in the cyanobacterium Syne-chocystis 6803. Plant mol. Biol. 10: 293-301.
Chitnis PR, Nelson N (1991) Photosystem I. In: Bogorad L,Vasil IK (eds), Cell Culture and Somatic Cell Genetics ofPlants, Vol 7B. Academic Press, San Diego: 177-224.
Clarke SE, Stuart J, Sanders-Loehr J (1987) Induction ofsiderophore activity in Anabaena spp. and its moderationof copper toxicity. Appl. environ. Microbiol. 53: 917-922.
Csaky TZ (1948) On the estimation of bound hydroxylaminein biological materials. Acta Chem. Scand. 2: 450-454.
Entsch B, Smillie RM (1972) Oxidation-reduction proper-ties of phytoflavin, a flavoprotein from blue-green algae.Arch. Biochem. Biophy. 151: 378-386.
Estep M, Armstrong JE, van Baalen C (1975) Evidence forthe occurrence of specific iron (III)-binding compoundsin near-shore marine ecosystems. Appl. Microbiol. 30:186-188.
Fillat MF, Borrias WE, Weisbeek PJ (1991) Isolation andoverexpression in Escherichia coli of the flavodoxin genefrom Anabaena PCC 7119. Biochem. J. 280: 187-191.
Fillat MF, Sandmann G, Gomez-Moreno C (1988) Flavo-doxin from the nitrogen-fixing cyanobacterium AnabaenaPCC 7119. Arch. Microbiol. 150: 160-164.
Gillam AH, Lewis AG, Andersen RJ (1981) Quantitativedetermination of hydroxamic acids. Anal. Chem. 53: 841-844.
Gingrich JC, Gasparich GE, Sauer K, Bryant DA (1990)Nucleotide sequence and expression of the two genes en-coding D2 protein and the single gene encoding the CP43protein of photosystem II in the cyanobacterium Syne-chococcus sp. PCC 7002. Photosyn. Res. 24: 137-150.
Golden SS, Steams GW (1988) Nucleotide sequence andtranscript analysis of three photosystem II genes from the

209
cyanobacterium Synechococcus sp. PCC 7942. Gene 67:85-96.
Goldman SJ, Lammers PJ, Berman MS, Sanders-Loehr J(1983) Siderophore-mediated iron uptake in differentstrains of Anabaena sp. J. Bact. 156: 1144-1150.
Guikema JA (1985) Fluorescence induction characteristics ofAnacystis nidulans during recovery from iron deficiency.J. Plant Nutr. 8: 891-908.
Guikema JA, Sherman LA (1983) Organization and functionof chlorophyll in membranes of cyanobacteria during ironstarvation. Plant Physiol. 73: 250-256.
Hardie LP, Balkwill DL, Stevens SE Jr (1983a) Effects ofIron starvation on the physiology of the cyanobacteriumAgmenellum quadruplicatum. Appl. environ. Microbiol.45: 999-1006.
Hardie LP, Balkwill DL, Stevens SE Jr (1983b) Effects ofiron starvation on the ultrastructure of the cyanobacteriumAgmenellum quadruplicatum. Appl. environ. Microbiol.45: 1007-1017.
Ho KK, Krogmann DW (1984) Electron donors to P700 incyanobacteria and algae, an instance of unusual geneticvariability. Biochim. Biophys. Acta 766: 310-316.
Hurt EC, Hauska G, Shahak Y (1982) Electron protontranslocation by the chloroplast cytochrome b6lfcomplexreconstituted into phospholipid vesicles. FEBS Lett. 149:211-216.
Hutber GN, Hutson KG, Rogers LJ (1977) Effect of irondeficiency on levels of two ferredoxins and flavodoxinsin a cyanobacterium. FEMS Microbiol. Lett. 1: 193-196.
Kerry A, Laudenbach DE, Trick CG (1988) Influence of ironlimitation and nitrogen source on growth and siderophoreproduction by cyanobacteria. J. Phycol. 24: 566-571.
Lange W (1974) Chelating agents and blue-green algae. Can.J. Microbiol. 20: 1311-1321.
Laudenbach DE, Straus NA (1988) Characterization of acyanobacterial iron stress-induced gene similar to psbC.J. Bact. 170: 5018-5026.
Laudenbach DE, Reith ME, Straus NA (1988) Isolation, se-quence analysis, and transcriptional studies of the flavo-doxin gene from Anacystis nidulans R2. J. Bact. 170:258-264.
Laudenbach DE, Herbert SK, McDowell C, Fork DC, Gross-man AR, Straus NA (1990) Cytochrome c-553 is not re-quired for photosynthetic activity in the cyanobacteriumSynechococcus. Plant Cell 2: 913-924.
Leonhardt KG, Straus NA (1992) An iron stress operon in-volved in photosynthetic electron transport in the marinecyanobacterium Synechococcus sp. PCC 7002. J. gen. Mi-crobiol. 138: 1613-1621.
Leonhardt KG, Straus NA (1994) Photosystem II genes isiA,psbDI and psbC in Anabaena sp. PCC 7120: Cloning, se-quencing and the transcriptional regulation in iron stressedand iron replete cells. Plant mol. Biol. (in press).
McKnight DM, Morel FMM (1979) Release of weakand strong copper-complexing agents by algae. Limnol.Oceanogr. 24: 823-837.
McKnight DM, Morel FMM (1980) Copper complexation bysiderophores from filamentous blue-green algae. Limnol.Oceanogr. 25: 62-71.
Murphy TP, Lean DRS, Nalewajko C (1976) Blue-green al-gae: Their excretion of iron-selective chelators enables
them to dominate other algae. Science 192: 900-902.Oquist G (1971) Changes in pigment composition and pho-
tosynthesis induced by iron-deficiency in the blue-greenalga Anacystis nidulans. Physiol. Plant. 25: 188-191.
6quist G (1974a) Iron deficiency in the blue-green algaAnacystis nidulans: fluorescence and absorption spectrarecorded at 770 K. Physiol. Plant. 31: 55-58.
Oquist G 1974b) Iron deficiency in the blue-green alga Ana-cystis nidulans: Changes in pigmentation and photosyn-thesis. Physiol. Plant. 30: 30-37.
Pakrasi HB, Riethman HC, Sherman LA (1985) Organizationof pigment proteins in the photosystem II complex of thecyanobacterium Anacystis nidulans R2. Proc. natl. Acad.Sci. USA 82: 6903-6907.
Pakrasi HB, Williams JGK, Arntzen CJ (1988) Targeted mu-tagenesis of the psbE and psbF genes blocks photosyn-thetic electron transport: Evidence for a functional role ofcytochrome b-559 in photosystem II. EMBO J. 7: 325-332.
Pakrasi HB, Diner BA, Williams JGK, Arntzen CJ (1989)Deletion mutagenesis of the cytochrome b-559 proteininactivates the reaction center of photosystem II. PlantCell 1: 591-597.
Riethman HC, Sherman LA (1988) Purification andcharaclerization of an iron stress-induced chlorophyll-protein from the cyanobacterium Anacystis nidulans R2.Biochim. Biophys. Acta 935: 141-151.
Rioux C, Jordan DC, Rattray JBM (1983) Colorimetric deter-mination of catechol siderophores in microbial cultures.Anal. Biochem. 133: 163-169.
Rogers LJ (1987) Ferredoxins, flavodoxins and related pro-teins: structure, function and evolution. In: Fay P, VanBaalen C (eds), The Cyanobacteria. Elsevier, Amsterdam,New York & Oxford: 35-67.
Rogner M, Chisholm DA, Diner BA (1991) Site-directedmutagenesis of the psbC gene of photosystem II: Isolationand functional characterization of CP43-less photosystemII core complexes. Biochemistry 30: 5387-5395.
Sandmann G (1985) Consequences of iron deficiency on pho-tosynthetic electron transport in blue green algae. Photo-syn. Res. 6: 261-271.
Sandmann G (1986) Formation of plastocyanin and cy-tochrome c-553 in different species of blue-green algae.Arch. Microbiol. 145: 76-79.
Sandmann G, Malkin R (1983) Iron-sulfur centers and activi-ties of the photosynthetic electron transport chain in iron-deficient cultures of the blue-green alga Aphanocapsa.Plant Physiol. 73: 724-728.
Sandmann G, Peleato ML, Fillat MF, Lazaro MC, Gomez-Moreno C (1990) Consequences of iron-dependent forma-tion of ferredoxin and flavodoxin on photosynthesis andnitrogen fixation on Anabaena strains. Photosyn. Res. 26:119-125.
Schrautenmeier B, Bhme H (1985) A distinct ferredoxinfor nitrogen fixation isolated from heterocysts of thecyanobacterium Anabaena variabilis. FEBS Lett. 184:304-308.
Sherman DM, Sherman LA (1983) Effect of iron deficiencyand iron restoration on ultrastructure of Anacystis nidu-lans. J. Bacteriol. 156: 393-401.
Simpson FB, Neilands JB (1976) Siderochromes in

210
Cyanophyceae: Isolation and characterization of schizoki-nen from Anabaena sp. J. Phycol. 12: 44-48.
Smillie RM (1965a) Isolation of phytoflavin, a flavoproteinwith chloroplast ferredoxin activity. Plant Physiol. 40:1124-1128.
Smillie RM (1965b) Isolation of two proteins with chloro-plast ferredoxin activity from a blue-green alga. Biochem.biophys. Res. Comm. 20: 621-629.
Van der Plas J, de Groot R, Woortman FC, Borrias M, VanArkel G, Weisbeek P (1988) Genes encoding ferredoxinsfrom Anabaena sp. PCC 7937 and Synechococcus sp.PCC 7942: structure and regulation. Photosyn. Res. 18:179-204.
Vermaas WFJ, Ikeuchi M (1991) Photosystem II. In: Bo-gorad L, Vasil IK (eds), The Photosynthetic Apparatus:Molecular Biology and Operation. Academic Press, SanDiego: 25-111.
Vermaas WFJ, Ikeuchi M, Inoue Y (1988) Protein compo-sition of the photosystem II core complex in geneticallyengineered mutants of the cyanobacterium Synechocystissp. PCC 6803. Photosyn. Res. 17: 97-113.
Winkelmann G (1991) CRC Handbook of Microbial IronChelates. CRC Press, Bacon Raton.
Yu J, Vermaas WFJ (1990) Transcript levels and synthesisof photosystem II components in cyanobacterial mutantswith inactivated photosystem II genes. Plant Cell 2: 315-322.