characterization of phycotoxins produced by cyanobacteria
TRANSCRIPT

O c e a n o l o g i c a l a n d H y d r o b i o l o g i c a l S t u d i e s I n t e r n a t i o n a l J o u r n a l o f O c e a n o g r a p h y a n d H y d r o b i o l o g y
Vol. XXXV, No.1
Institute of Oceanography (85-109) 2006
University of Gdańsk
Research Article
Characterization of phycotoxins produced by cyanobacteria
HANNA MAZUR-MARZEC
Department of Marine Biology and Ecology, University of Gdańsk Marszałka Piłsudskiego 46, 81-378 Gdynia, Poland
[email protected] Key words: Harmful Algal Blooms, cyanobacteria, toxins
Abstract
Cyanobacteria (blue-green algae), photosynthetic prokaryotes, are essential elements of aquatic ecosystems. They produce a great variety of secondary metabolites; some of which have potentially useful pharmaceutical properties as anti-tumour, antibacterial and antiviral agents. Some species of cyanobacteria form blooms and become dominant over other forms of aquatic life. Blooms can deteriorate water quality in a variety of ways. Blooms formed by toxic species are the most harmful, as they can cause poisoning and death of organisms that come into contact with them.
In this review, the structure and activity of cyanobacterial toxins are described. The toxins are classified into: hepatotoxins, neurotoxins, dermatotoxins and endotoxic lipopolysaccharides. Hepatotoxic cyclic pentapeptides, microcystins and nodularins, are the most common cyanobacterial toxins that have been identified in strains of Microcystis, Anabaena, Nodularia, Planktothrix (Oscillatoria), Nostoc, Hapalosiphon and Anabaenopsis. Cylindrospermopsin, another cyanobacterial hepatotoxin, is produced mainly by Cylindrospermopsis raciborskii. There are several neurotoxins produced by the cyanobacterial genera Anabaena, Aphanizomenon, Planktothrix and Cylindrospermopsis. Based on their activity, these neurotoxins were classified into anatoxin-a, anatoxin-a(S) and saxitoxins. BMAA is a novel cyanobacterial neurotoxin implicated in the ALS/PDC syndrome in the Chamorro people of Guam. The occurrence and harmful effects of cyanobacterial dermatotoxins and endotoxic lipopolysaccharides are less well recognised.

H. Mazur-Marzec 86
HARMFUL ALGAL BLOOMS
Phytoplanktonic organisms, as primary producers, constitute an important element of marine and freshwater food sources. However, excessive growth of these organisms, called blooms, has a negative impact on the sustainable functioning of the ecosystem as well as on environmental health. It has been estimated that approximately 300 marine phytoplankton species can, at times, proliferate to such an extent that they form blooms that change the colour of the surface water to red (red tides) or green (Hallegraeff 1993). Out of these, over 100 species produce toxins. If toxic blooms occur in areas where fish and shellfish are farmed, they can cause serious economic losses as well as threats to humans’ health. The occurrence of algal blooms is a natural phenomenon; however, in recent years, an increase in their frequency and intensity has been observed.
One of the primary reasons for excessive algal proliferation is a disturbance in the ecological equilibrium, caused by anthropogenic impact. High nutrient loading from domestic, industrial and agricultural wastes has increased the level of eutrophication, and created favourable conditions for algal growth. In some instances the natural balance between macronutrients has been changed. A decline in the Si:P and N:P ratios has contributed to the proliferation of nuisance organisms and reduced the biodiversity of aquatic ecosystems. The growth of phytoplanktonic organisms is also stimulated by global climate change and the warming of surface waters. There is also strong evidence that toxic blooms can spread to distant places due to the transfer of the settled spores in ship water ballast tanks.
The term “Harmful Algal Bloom” (HAB) refers to some (at least three) negative aspects of the phenomenon. First, the bloom may be harmful when the number of algal cells exceeds one million per 1 dm3. At such a high abundance of microorganisms, oxygen consumption by the processes of respiration and organic matter mineralization leads to anoxic conditions and the formation of toxic sulphide. This deteriorates water quality and may cause the death of fish and other aquatic organisms.
Second, the blooms may be formed by species producing ichthyotoxins, primarily haemolysins (Cambella 2003). In spectacular incidents of fish death, the raphidophytes Heterosigma akashiwo and Chattonella marina, prymnesiophytes Chrysochromulina polylepsis and Prymnesium parvum, as well as the dinoflagellates Karenia mikimotoi and Pfisteria piscicida have been involved.
From the human perspective, the most harmful blooms are formed by phytoplankton species producing substances toxic to mammals. The blooms have been divided into several classes according to the clinical symptoms

Phycotoxins – Review, part I 87
associated with toxication and the mode of action of the toxins. Uptake of the toxins may result in serious diseases, defined as: paralytic shellfish poisoning (PSP), diarrhetic shellfish poisoning (DSP), amnesic shellfish poisoning (ASP), neurotoxic shellfish poisoning (NSP), ciguatera fish poisoning (CFT) and cyanobacterial toxin poisoning (CTP). The toxins are synthesized mainly by marine dinoflagellates (PSP, DSP, NSP, CFT), diatoms of the Pseudo-nitzschia genus (ASP), and by freshwater, brackish and marine cyanobacteria (CTP, PSP). The biosynthetic pathways of the toxins are unique, so they constitute a structurally and functionally diverse group of compounds. Generally, they either interact with the cell membrane or affect signal transduction at receptor sites of the target cell. Dinoflagellates of the genera Alexandrium, Dinophysis, Prorocentrum and the benthic species Gambierdiscus toxicus are responsible for most cases of human intoxication after the consumption of contaminated seafood products.
The role of phycotoxins in the life of their producer has been widely discussed. Generally, they are not essential for metabolic processes nor for the growth of the organism. There is some evidence that the toxins function as allelochemical agents used to survive in an ecological niche, or contribute to chemical defence against predators from higher trophic levels. Phycotoxins may take part in chemical signalling or may simply be an evolutionary relic. It has been demonstrated that the production of toxins is genetically determined, and within one species both toxin-producing and non-toxic strains can be found. The toxin content in different strains belonging to the same species can vary significantly. To a lesser degree, toxin production depends on the growth phase of the organism as well as on environmental conditions such as temperature, light intensity and nutrient concentration (Granéli and Johansson 2003).
CYANOBACTERIAL BLOOMS
Cyanobacteria (blue-green algae) are photosynthetic prokaryotic microorganisms possessing chlorophyll a. They can grow as single cells, colonies or filamentous forms. Cyanobacteria have developed some adaptive strategies that enable them to survive in unfavourable environmental conditions. Cyanobacterial accessory pigments protect them against excessive solar radiation and enable them to effectively absorb the available light. Some filamentous forms of Nostocales and Stigonematales produce heterocysts – specialised cells that fix atmospheric nitrogen. These cyanobacteria are independent of other nitrogen sources. Through the production or collapse of gas vacuoles, certain species can adjust their position in the water column to obtain optimal light and nutrient conditions (Pearl 2000). In stable, stratified water this may lead to surface accumulations of cells and bloom formation (Fig.

H. Mazur-Marzec 88
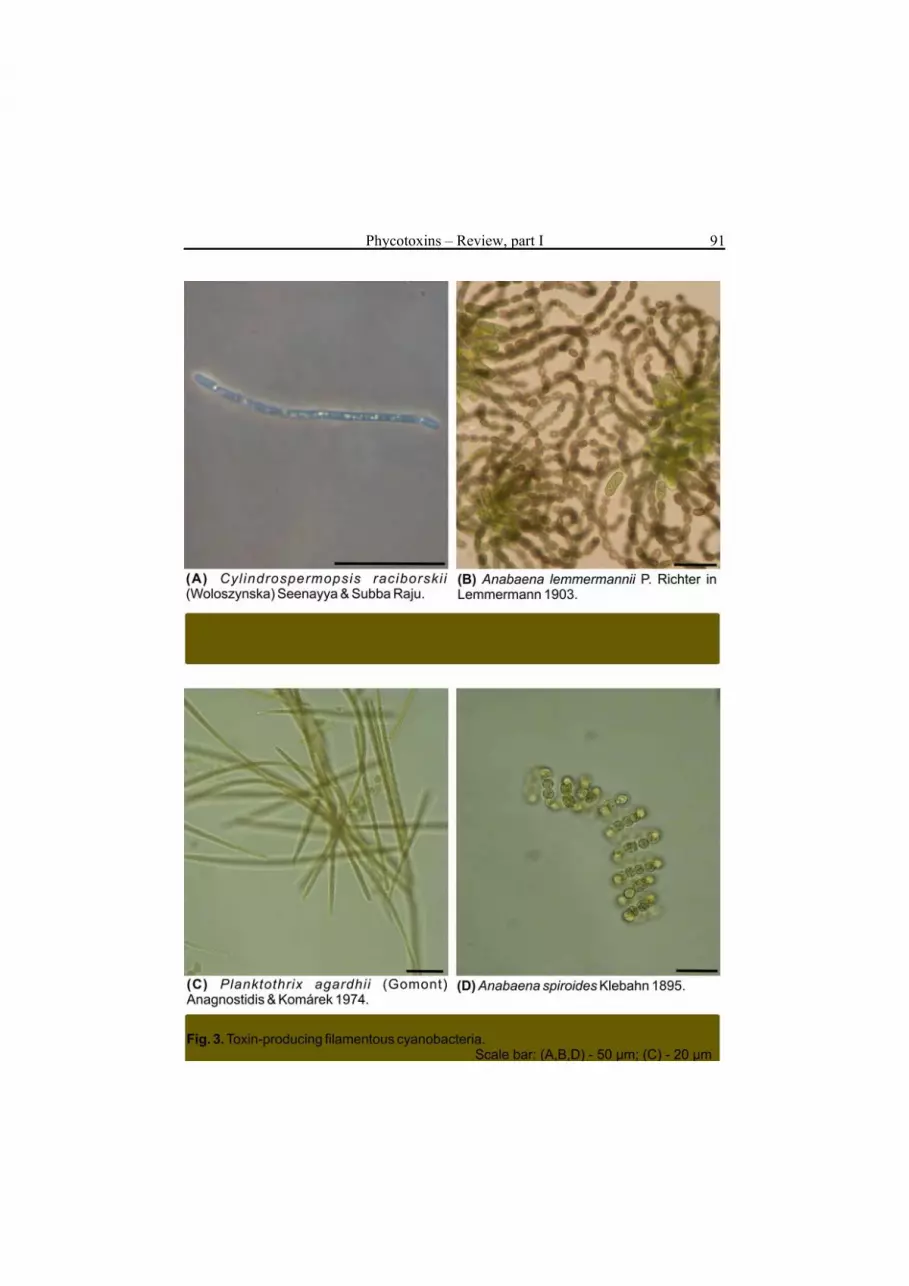
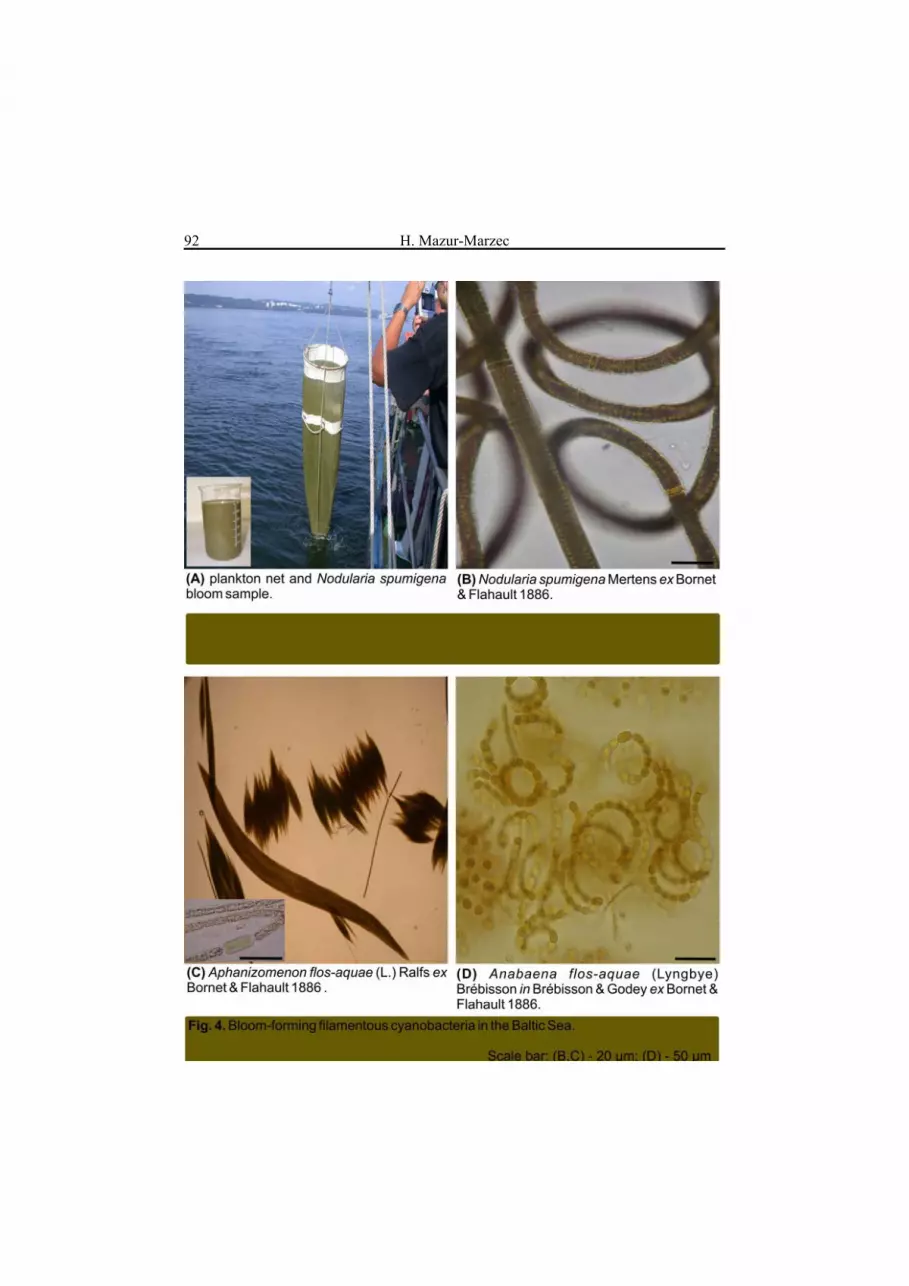
1). The conditions that induce and favour bloom development include: high nutrient concentration and low N/P ratio, light intensity, seawater temperature of 15-30°C, pH> 6 and thermal stratification. It has been estimated that between 25% and 75% of cyanobacterial blooms are formed by toxin producing species, primarily of freshwater origin (Sivonen 1996). They are especially abundant in shallow, warm, eutrophic waters. In lakes, species of the genera Microcystis (Fig. 2), Anabaena and Planktothrix (Fig. 3) usually dominate. Even if the blooms last only a few days, they reduce water quality and can pose a serious threat to drinking water supplies. In the open and coastal ocean, the N2-fixing non-heterocystous cyanobacterium Trichodesmium spp. forms the largest phytoplankton aggregations ever recorded (Sellner 1997). The cyanobacterium was implicated in incidents of fish, oyster, and crab mortality. In brackish waters of the Baltic Sea, summer blooms of the heterocystous cyanobacteria Aphanizomenon flos-aquae, Nodularia spumigena and Anabaena sp. (Fig. 4) can cover an area of over 100.000 km2 (Fig. 1) (Helcom 2004). N-fixing organisms are efficient suppliers of nitrogen to ecosystems; however, their growth requires sufficient concentrations of phosphorus and iron. The latter element is important for physiological processes such as photosynthesis, nitrogen assimilation, respiration and chlorophyll a synthesis. Large-scale blooms of N2-fixing organisms are frequent phenomena in freshwater and marine environments.
CYANOBACTERIAL TOXINS
Over 40 species of cyanobacteria, belonging mainly to the Chroococcales, Oscillatoriales or Nostocales orders, produce such a great variety of toxins that they can be regarded as the primary source of compounds in aquatic environments. A substantial amount of data has been collected on the harmful effects of cyanobacterial toxins for human health; currently, more attention is paid to the problem. In many countries people are chronically exposed to low concentrations of cyanobacterial toxins in tap water. This can take place when cyanobacterial blooms occur in the drinking water supplies, and the processes of toxin removal are not totally effective. Another route of exposure includes skin contact and/or inhalation during recreational use of waters affected by toxic blooms. Numerous incidents of human and animal toxication have been described by Kupier-Goodman et al. (1999) and Falconer (2005). The first record in the scientific literature comes from 1878, when farm animal poisoning occurred around the coastal Lake Alexandrina, close to the Murray River in South Australia (Francis 1878). Consumption of scum containing Nodularia spumigena caused the death of sheep, horses, dogs and pigs. The most serious and well-documented human poisonings occurred in Brazil. First in 1988, when

Phycotoxins – Review, part I 89

H. Mazur-Marzec 90
Phycotoxins – Review, part I 91
H. Mazur-Marzec 92

Phycotoxins – Review, part I 93
exposure to cyanobacterial toxins in drinking water resulted in over 2000 cases of gastroenteritis and 88 deaths in a 42-day period. Then, in 1996, when 116 out of 131 dialyzed patients were affected by gastric disturbances in a dialysis clinic in Caruaru, Pernambuco State. Out of these, 100 developed liver failure and 76 patients died (Pouria et al. 1998). Later studies proved the presence of cyanobacterial hepatotoxins, microcystin and cylindrospermopsin, in water used for dialysis (Carmichael et al. 2001). Microcystin was also detected in the patients’ liver samples (50-471 ng g-1) and serum (2.2 ng dm-3).
Based on the mode of action, cyanobacterial toxins can be categorized into the following groups: hepatotoxins (most commonly encountered in cyanobacterial blooms), neurotoxins, cytotoxins, dermatotoxins and lipopolysacharides (LPS). One strain of cyanobacteria may produce more than one toxin, and even some toxins of different classes.
As far as the role of cyanobacterial toxins, it cannot be excluded that they take part in allelochemical interactions with other phytoplanktonic organisms. There are some reports of their effect on the growth of green algae and other cyanobacteria by the inhibition of electron transfer in photosystem II or inhibition of protein and chlorophyll a synthesis and DNA replication (Doan et al. 2001, Singh et al. 2001, Hu et al. 2004).
Hepatotoxins
Microcystins and nodularins This group includes the cyclic heptapeptide microcystins (Mcst) and the
structurally and functionally similar cyclic pentapeptide nodularins. The general structure of microcystins (Fig. 5) is cyclo-(D-Ala1-X2-D-MeAsp3-Z4-Adda5-D-Glu6-Mdha7), where X and Z are variable L-amino acid (leucine (L), arginine
O O
OO
O
OH
H3C
H3CH
CH3
CH3 HH
H
H
H
H
H
H3CCH2
H
HNN
NH
HN
NH
NH
OX
CH3
CH3
COOH
O
COOH
Z
1
2
34
5
6 7
Fig. 5. General structure of microcystins (MCYST). X, Z – L-amino acids, in MCYST-LR, X=L-Leucine (L), Z=L-Arginine (R).

H. Mazur-Marzec 94
(R), alanine (A), tyrosine (Y), methionine (M), tryptophane (W), phenylalanine (F)), MeAsp3 is D-erythro-β-methylaspartic acid, and Mdha7 is N-methyldehydroalanine. The Adda5 is a C20 β-amino acid residue unique to cyanobacterial hepatotoxins: (2S,3S,8S,9S)-3-amino-9-methoxy-2,6,8,-trimethyl-10-phenyldeca-4(E),6(E)-dienoic acid.
Microcystins are named according to their variable amino acid – for example Mcst-LR contains leucine and arginine residues. There are over 76 microcystin analogues, which differ primarily with respect to the two L-amino acid components and the presence or absence of methyl groups on MeAsp, Mdhb and Adda (Sivonen and Jones 1999, Spoof 2003). There are also variants with an acetoxy group in the Adda residue, instead of a methoxy group. Nodularin (Fig. 6) is cyclo(-D-MeAsp1-L-Arg2-Adda3-D-Glu4-Mdhb5), where Mdhb stands for 2-(methylamino)-2(Z)-dehydrobutiric acid. Some minor nodularin analogues were characterized in bloom ([DMAdda3]nodularin, [6(Z)-Adda3]nodularin) and culture ([d-Asp1]nodularin) N. spumigena samples from Lake Ellesmere in New Zealand (Namikoshi et al. 1994). A Nodularin analogue without a methyl group on Asp [D-Asp1] nodularin was also detected in N. spumigena and blue-mussels from the Baltic Sea (Karlsson et al. 2003). There are two other nodularins: [L-Val2]nodularin, probably produced by microbial symbionts of a marine sponge (de Silva et al. 1992), and [L-Har2]nodularin detected in Nodularia PCC7804 (Pasteur Culture Collection) isolated from a thermal spring in France (Beattie et al. 2000).
NH
O
O
O
O
N
N NH
O
COOH
H
NH
NH
CH2N
NH
CH3
CH3 CH3
H3C
O
Adda
D-Glu
M dhb
D-MeAsp
CH3
CH3
CH3
Arg
COOH3
4
5
12
Fig. 6. Structure of nodularin.

Phycotoxins – Review, part I 95
Changes in the hepatotoxin structure have a significant effect on their biological activity. The Adda -Glu part of the molecule and its configuration are essential for the toxicity of the compounds. Esterification of the free carboxyl group in Glu results in an inactive compound. The isolated Adda was inactive in a mouse test and protein phosphatase inhibition assay (PPIA) (Harada et al. 2004). Studies on the structure-activity relationship demonstrated that the formation of a [6(Z)Adda3] stereoisomer, saturation of the diene in Adda or linearization render the microcystins and nodularins non-toxic or lead to a significant decrease in toxicity (Rinehart et al. 1988, An and Carmichael 1994). A lethal dose (LD50) of most cyanobacterial peptide hepatotoxins in mouse tests range from 50 μg kg-1 body weight (b.w.) (for nodularin and Mcst-LR) to 600 μg kg-1 b.w.(for Msct-RR) as administered through intraperitoneal injection (i.p.) (Sivonen and Jones 1999).
It has been shown that microcystins bind covalently to protein phosphatase (PP) and inhibit activity of the enzymes in eukaryotic organisms (Yoshizawa et al. 1990). Nodularin does not form covalent bounds, but strong interactions of the enzymes with the Adda-Glu part of the molecule still occur. Protein phosphatases play a role in maintaining cellular homeostasis and take part in many important processes in the cell (carbohydrate and lipid metabolism, signal transduction, cell division). Inhibition of the enzymes leads to hyperphosphorylation of cytoskeletal proteins, disruption of cytoskeletal structure and massive hepatic haemorrhage – the direct cause of death. Experiments on animals exposed to sub-lethal doses of cyanobacterial hepatotoxins revealed their activity as liver cancer promoters (microcystins) and direct carcinogens (nodularin) (Ohta et al. 1994, Falconer and Humpage 1996). Recently, it has been shown that in cultured hepatocytes the toxins induced oxidative stress, which was expressed by an enhanced production of reactive oxygen species (ROS), lipid peroxidation and depletion of intracellular glutathione (Lankoff et al. 2002, Ding et al. 2003, Bouaїcha and Maatouk 2004). Ding and Ong (2003) hypothesised that microcystin-induced oxidative stress, alterations in the mitochondrial membrane potential, and a transition in mitochondrial permeability may lead to programmed cell death, called apoptosis. After microcystin exposure, in addition to liver damage, intestinal and kidney injuries have been observed.
Kondo et al. (1992) showed that in vitro microcystins form conjugates with glutathione and cysteine (Cys). These structures were found in the livers of animals treated with the toxins. Environmental and laboratory studies revealed hepatotoxin accumulation in zooplankton, mussels, liver and muscles of Atlantic salmon and other fish. Detoxication of microcystins, as in many other xenobiotics, proceeds through the formation of conjugates with reduced

H. Mazur-Marzec 96
glutathione (GSH) in a reaction catalyzed by the enzyme glutathione S-transferases (GST) (Pflugmacher et al. 1998). The Mcst-GSH and Mcst-Cys conjugates are less toxic and more polar, and they can be excreted through the kidney. Typical symptoms of acute poisoning in mammals exposed to hepatotoxic microcystins include diarrhoea, vomiting, weakness, anorexia, pallor and, in extreme situations, death due to liver haemorrhage (Codd et al. 1999). Chronic exposure to toxins in drinking water is of particular concern. According to the World Health Organization (WHO), the concentration of microcystin-LR in drinking water should not exceed 1 μg dm-3 (WHO 1999). However, cyanobacterial hepatotoxins are liver tumour promoters and even much lower concentrations may prove harmful to man. One positive fact is that the ability of the toxins, as polar compounds, to enter cells is limited. They can cross lipid membranes only by the carrier mediated bile-acid transport system, which operates in liver cells (Eriksson et al. 1990). This explains why the liver is a target organ for microcystins and nodularins. Some more hydrophobic and less common microcystins can cross the cell membrane by passive diffusion.
Microcystins are primarily produced by freshwater cyanobacteria of the genera Microcystis (Fig. 2), Anabaena, Planktothrix (Oscillatoria) (Fig. 3), Nostoc, Hapalosiphon and Anabaenopsis. However, the occurrence of the toxins in the Baltic Sea, in association with the presence of Anabaena, has also been reported (Mazur et al. 2003, Karlsson et al. 2005). Cell-bound microcystin concentration rarely exceeds 100 μg dm-3; in lyophilized phytoplankton samples it may reach up to 7,300 μg g-1 d.w. (Sivonen and Jones 1999). Within the same bloom both toxic and non-toxic strains can be found. In some strains the occurrence of specific microcystin variants is more probable. Cyanobacteria of the Planktothrix genus (Fig. 3 c) are reported to produce mainly demethylated microcystin variants, e.g. [D-Asp]Mcst-RR or [Dha]Msct-RR. Dimethylated Mcst-YR is typical of the Scandinavian Microcystis spp. In most studies of microcystin presence in freshwater blooms, the co-occurrence of Mcst-LR, RR and YR has been reported (e.g. Park and Watanabe 1996, Sivonen and Jones 1999, Jurczak et al. 2004). This is partially due to the fact that standards of these three toxins are commercially available; hence the toxins can be more easily identified. Microcystin-LR has also been found in picoplanktonic cyanobacteria, e.g. Aphanocapsa cumulus (Domingos et al. 1999). Nodularin is produced only by N. spumigena, which forms blooms primarily in the Baltic Sea (Fig. 4 b) and some estuaries and coastal embayments in Australia and New Zealand. In the Baltic Sea, the average concentration of cell-bound nodularin during a N. spumigena bloom is 100-2,400 μg g-1, but values as high as 18,100 μg g-1 have also been recorded (Sivonen et al. 1989, Kononen et al. 1993, Mazur and Pliński 2003).

Phycotoxins – Review, part I 97
The toxins are endogenous substances, and in exponential growth phase they occur predominantly inside the cell. They are localized in the thylakoid area, nucleoid and, to a lesser extent, in the cell wall and sheath area (Shi et al. 1995). They are non-ribosomally synthesised in the presence of large multifunctional enzyme complexes (Tillet et al. 2000). The release of toxins to the surrounding water occurs during the senescence and decomposition of cells. As a result, after the cyanobacterial bloom, the concentrations of dissolved toxins increase. There are some natural processes that lead to the reduction of toxin levels in water; they include dilution by uncontaminated water masses, adsorption on suspended particles and sediments, thermal decomposition, photodegradation and microbial degradation. Very few bacteria have been identified that are responsible for degradation of the toxins; most of them belong to the genus Sphingomonas. Jones and Orr (1994) suggested biphasic degradation of microcistins by sequential induction of two different bacterial populations. One of them utilized the toxin as a source of carbon and energy, and the second metabolized the remaining microcystin-LR. Bourne et al. (1996) characterized the intermediates of microcystin enzymatic degradation as the linearized form of the toxin and tetrapeptide NH2-Adda-Glu (iso)-methyldehydroalanine-Ala-OH. Much less is known about the biodegradation pathway of nodularins. Both kinds of cyanobacterial toxins are degraded by natural sunlight into non-toxic geometrical isomers (Tsuji et al. 1994, Twist and Codd 1997). Takenaka and Watanabe (1997) demonstrated that the bacterium Pseudomonas aeruginosa isolated from a Japanese lake can metabolize microcystin-LR into (2S,3S,8S)-3-amino-2,6,8,-trimethyl-10-phenyldeca-4(E),6(E)-dienoic acid (DMAdda). The same degradation product was found in the liver of mouse treated with microcistin-LR (Takenaka 1998). Cyanobacterial peptide hepatotoxins are chemically very stable; they cannot easily be destroyed by oxidation at neutral pH or heating. In a solution of pH 1 and temperature of 40°C, their half-life was about three weeks (Harada 1996).
Studies on the effect of hepatotoxic cyanobacteria on zooplankton have shown different response reactions depending on the zooplankton species DeMotte et al. (1991) suggested that the different susceptibility to toxic cyanobacteria may be a result of natural selection favouring the most resistant species. It is unclear whether the effects can be attributed to poor nutritional value of the cyanobacteria and inhibition of zooplankton grazing activity or simply the activity of toxins or other biologically active compounds. In fact, some zooplanktonic organisms, including Daphnia, avoid grazing on toxic cyanobacterial species, but still feed on non-toxic ones (Rohrlack et al. 1999). In this way, selective enrichment of toxic forms can occur. Additionally, long cyanobacterial filaments or large colonies can be difficult for zooplankton to

H. Mazur-Marzec 98
ingest. Therefore, other possible routes of toxin administration have been considered (Reinikainen et al. 2002). In experiments with radiolabeled nodularin, Karjalainen et al. (2003) demonstrated that uptake and accumulation of dissolved toxin occurred in Baltic Sea zooplankton. The transfer of toxins through faecal pellets produced by herbivorous zooplankton feeding on cyanobacteria to other aquatic organisms, including bivalves, has also been found (Lehtiniemi et al. 2002, Svensen et al. 2005).
Mussels may also accumulate high quantities of toxins through ingestion of contaminated food or direct uptake from water. In Mytilus galloprovincialis fed on freshwater cyanobacteria, up to 10.7 μg of microcystin per 1 g mussel d.w. was measured (Amorim and Vasconcelos 1999). During an N. spumigena bloom, Mytilus edulis from the Baltic Sea accumulated up to 2.1 μg g-1 d.w. of nodularin (Sipiä et al. 2002). In laboratory experiments, tissue of a mussel fed for 48 hrs with N. spumigena cells contained 80±21.2 μg g-1 of nodularin. In the digestive gland, 245±35 μg g-1 was detected (Svensen et al. 2005). Lower nodularin concentrations were found (399±5 ng g-1) in flounders that consume mussels (Sipiä et al. 2001). Detection of nodularin in eiders feeding on blue mussels (3-180 ng g-1) confirmed the transfer of cyanobacterial toxins to higher trophic levels (Sipia et al. 2003). The published results show that the concentration of the toxin decreases in the trophic chain from mussel to birds. This indicates that biodilution rather than bioaccumulation of cyanobacterial toxins takes place. This might be due to higher toxin content in mussel food and/or more effective detoxification processes in fish and birds. Even at low concentrations, however, microcystins and nodularin can cause similar symptoms in fish and laboratory mammals. Fladmark et al. (1998) found that cyanobacterial toxins added to suspension cultures of rat and salmon hepatocytes evoked stronger apoptotic effects in the latter. Significant histo-pathological changes in the liver of fish exposed to the toxins were observed (Bury et al. 1997, Kankaanpää et al. 2002). In reared Atlantic salmon, a Net-pen Liver Disease attributed to microcystin toxication caused significant economic losses (Andersen et al. 1993). The toxic effects of cyanobacteria on fish early life stages have been studied by Wiegand et al. (1999) and Tymińska et al. (2005), among others.
Since the cyanobacterial hepatotoxins accumulate in aquatic biota, all seafood products from a bloom affected area should be checked for the presence of microcystins and nodularin. The tolerable daily intake (TDI) of microcystin-LR has been established as 40 ng kg-1 b.w. (body weight) (Kupier-Goodman et al. 1999).
Cylindrospermpsin Cylindrospermopsin (CYN) (LD50=2.1 mg kg-1) is a cyclic guanidine

Phycotoxins – Review, part I 99
OH
H
O3SO
H3C
O
O
NHNH
NH
HNN
+
alkaloid with cyto- and hepatotoxic activity (Fig. 7). The toxin is produced mainly by Cylindrospermopsis raciborskii (Fig. 3a) in tropical and subtropical waters of Australia. It also has been identified in Aphanizomenon ovalisporum, Umezakia natans, Anabaena bergii and Raphidopsis curvata.
These species vary significantly in their level of toxicity. At present, cylindrospermopsin-producing cyanobacteria can be found in many European, Central Asian and American water bodies (Falconer 2005). Several analogues of the toxin have been identified, including 7-epi-cylindrosprmopsin and deoxycylindrospermopsin. It has been demonstrated that the pyrimidine ring, the presence of a hydroxyl group on the uracil bridge and keto-enol status of uracil is essential for biological activity of the compound. Cylindrospermopsin is water soluble, but as a small compound is likely to be taken up by cells through diffusion; a small fraction of the toxin was found to enter hepatocytes by bile acid carriers. Cylindrospermopsin strongly and irreversibly inhibits protein biosynthesis and induces apoptotic changes in liver cells. The toxin binds covalently to DNA and significantly affects its structure; therefore, mutagenic activity of the toxin can be expected. Runnegar et al. (1995) showed that cylindrospermopsin inhibits glutathione synthesis, but GSH is not employed in the biotransformation of the toxin. One significant finding from studies on the mechanism of CYN toxicity is that the toxin is metabolized to more potent compounds by cytochrome P450 oxidation. In fact, the products of CYN oxidation by cytochrome P450 caused the death of cultured hepatocytes within 8-24 hrs, while the inhibition of protein synthesis by CYN did not. It was also revealed that pure toxin had a deleterious effect on liver cells, while cylindrospermopsin-containing cyanobacterial extract can induce pathological changes in the kidney, thymus, and heart.
Fig. 7. Structure of cylindrospermopsin.
In 1979, cylindrospermopsin was implicated in the human poisoning on Palm Island, Queensland, Australia (Griffith and Saker 2003). After chronic exposure to toxin in drinking water, 138 children and 10 adults were affected. Of these, over 100 children had to be admitted to the hospital with symptoms of gastroenteritis: swollen and painful livers, abdominal pain, constipation and vomiting. Additionally, kidney damage was recorded, probably because it is the primary route of toxin excretion. Epidemiological studies confirmed that a bloom of C. raciborskii was the most likely cause of the poisoning. During the

H. Mazur-Marzec 100
bloom of the cyanobacterium in Australian waters, the toxin concentration ranged from 0.1 to 800 μg dm-3 (Shaw et al. 1999, Falconer 2005). A poor correlation between cylindrospermopsin concentration and the abundance of C. raciborskii cells has been observed (Griffth and Saker 2003). Unlike microcystin and nodularin, a high portion of cylindrospermopsin is extracellular, and during a bloom over 60% of the toxin is in the water. There is some evidence that cylindrospermopsin accumulates in aquatic organisms, such as red claw crayfish and freshwater mussels (Saker et al. 2004). In toxicity tests, it had a negative impact on growth and protein synthesis in brine shrimp Artemia salina (Metcalf et al. 2002).
Neurotoxins
Anatoxin-a The alkaloide anatoxin-a (Fig. 8) (LD50 = 200 μg kg-1 b.w.) was isolated for
the first time from Anabaena flos-aquae. Anatoxin-a is also produced by other species of filamentous cyanobacteria of the genera Anabaena, Planktothrix, Aphanizomenon and Cylindrospermum (Sivonen and Jones 1999). Recently, production of the toxin by the benthic cyanobacterium Phormidium favosum was reported by Gugger et al. (2005). Park et al. (1993) detected anatoxin-a in Microcystis aeruginosa. The structurally similar homoanatoxin-a (LD50 = 288-578 μg kg-1 b.w.) has been found in Planktothrix formosa by Skulberg et al. (1992) Anatoxin-a is a structural analogue of cocaine and the neurotransmitter acetylcholine. The toxin acts as a post-synaptic neuromuscular blocking agent. Its irreversible binding to the nicotinic acetylcholine receptor causes sodium channel opening and the constant inflow of sodium ions to cells. As a result of membrane depolarization, overstimulation of the muscle cells occurs. When the
respiratory muscles are affected, lack of oxygen in the brain may lead to convulsions and finally to death by suffocation.
NH CH3
O
There were incidents of dog and sheep mortality attributed to the blooms of anatoxin-a producing cyanobacterial species. The toxin was also implicated in the mass mortality of flamingos in Kenya (Krienitz et al. 2003). Typical signs of anatoxin-a poisoning in animals include staggering, muscle
fasciculation, gasping, convulsions and in birds, opisthotonos.
Fig. 8. Structure of anatoxin-a.

Phycotoxins – Review, part I 101
Anatoxin-a(S) Anatoxin-a(S), a guanidine methyl phosphate ester, was isolated from
Anabaena flos-aquae and A. lammermannii. Although anatoxin-a(S) (LD50 = 20 μg kg-1 b.w.) produces many symptoms similar to anatoxin-a, the two toxins differ with respect to their structures and function. The activity of anatoxin-a(S) is quite similar to the organophosphate insecticides parathion and malathion. The toxin inhibits the activity of acetylcholinesterase so that the acetylcholine-receptor binding cannot be degraded and the muscles become constantly stimulated (Matsunaga et al. 1989). Anatoxin-a(S) was reported to be a source of livestock death in North America. The letter S in the name of the toxin stands for salivation, as this is one of the most apparent syndromes of anatoxin-a(S) toxication in vertebrates. The low stability of the toxin diminishes the risk of poisoning.
Saxitoxins
NH2
H2N N
N
OH
O
RO
H2N
NH
H
OH
NH+
+
Saxitoxins (LD50 = 10 μg kg-1 b.w.), responsible for the PSP syndrome, are primarily produced by marine dinoflagellates (Anderson 1989). These tricyclic guanidinium alkaloids (Fig. 9) have also been identified in Anabaena circinalis (Australia), Aphanizomenon flos-aquae, Lyngbya wollei (USA), and Cylindrospermopsis raciborskii (Brazil) (Humpage et al. 1994).
In nerve axon membranes, saxitoxins block sodium ion channels and stop ion inflow to the cell. This leads to a disturbance in the propagation of action potential to muscle cells. The presence of a carbamoyl group and hydroxyl groups in the molecule are essential for toxicity. Depending on the dose, saxitoxin poisoning may give such symptoms as tingling and numbness around the lips or, in extreme situations, paralysis and death caused by respiratory failure. Saxitoxins accumulate in aquatic organisms; they are harmful to mammals, fish and zooplankton. In the summer of 1990, mass development of the saxitoxin producing A. circinalis, covering over 1000 km of the Darling River in Australia, was recorded (Humpage et al. 1994, Falconer 2005). The bloom caused the death of approximately 2000 sheep and cattle along the river. Saxitoxins were also found in the drinking water supply.
Fig. 9. Structure of saxitoxin, R=H.

H. Mazur-Marzec 102
Β-N-methylamino-L-alanine (BMAA) BMAA, a non-protein amino acid
(Fig. 10), was originally identified in cycad seeds, where it probably plays a role as a chemical deterrent to herbivorous organisms. The toxin was also found in cyanobacterial (Nostoc) symbionts on cycad tree roots. Cox et al. (2003) reported significant biomagnification of BMAA within the Guam ecosystem. In the trophic chain: cyanobacteria - cycad tree roots - flying foxes - the Chamorro people of Guam, a 100.000-fold increase in the toxin concentration was recorded. BMAA accumulates in the brain tissue of Chamorro people living in Guam, where the flying fox is a traditional delicacy. Since the toxin was found only in people who died of degenerative disease, BMAA was suggested to be a possible cause of the amotrophic lateral sclerosis/parkinsonism-dementia complex (ALS/PDC). The rate of ALS-PDC among Chamorros is 50-100 times higher than elsewhere. The toxin is stored by binding with proteins, and is slowly released in the organism when proteins are metabolised (“slow toxin”). It has been postulated that BMAA is produced by many different groups of cyanobacteria (Cox et al. 2005). The toxin was detected in 95% of all genera of cyanobacteria tested. It should not be ruled out that BMAA is responsible for the neurotoxic activity of the marine cyanobacteria Trichodesmium thiebautii and Trichodesmium erythraeum. According to Cox et al. (2005), the common occurrence of BMAA in different cyanobacterial orders indicates that the production of toxin is a conserved evolutionary feature characteristic for the group of organisms as a whole.
H
The ecological function of BMAA in cyanobacteria is unclear.
Dermatotoxins
Some bethic cyanobacteria of the genera Lyngbya, Planktothrix and Schizothrix produce compounds causing irritant dermatitis and conjunctivitis after skin contact. These are the alkaloids: lyngbiatoxins (LA-A, B, C), aplysiatoxins (AT) and debromoaplysiatoxins (DAT). These toxins are strong tumour promoters and protein kinase C activators. The benthic filamentous marine cyanobacterium Lyngbya majuscula has been responsible for numerous incidents of dermatitis and gastrointestinal inflammation among swimmers in tropical and subtropical waters (Osborne et al. 2001). Skin irritation was more intense when cyanobacterial cells were caught under the swimsuit. In 1958, on
NH2
NH
OH
O3C
Fig. 10. Structure of BMAA (β-methylamino-L-alanine).

Phycotoxins – Review, part I 103
the windward beaches of Hawaii, 125 people were reported to suffer acute dermatitis. A few hours after contact with Lyngbya majuscula, they had symptoms of erythema and burning sensations, followed by blister formation and deep desquamation, lasting up to several days. Similar symptoms were developed in nearly 90% of people bathing on beaches in Okinawa in 1968. The parts of the body most severely affected were the genitals, eyes and lips (Burke and Tester 2002). The toxins produced by Lyngbya majuscula accumulated in fish and caused mass mortality of the reared blue-shrimp Penaeus stylirostris.
Lipopolysacharides (LPS)
As in Gram-negative bacteria, lipopolysaccharides are components of the outer cell envelope in many cyanobacterial strains. So far they have been identified in the chroococcalean genera Synechococcus, Synechocystis and Microcystis, the filamentous heterocystous Anabaena, and filamentous non-nitrogen fixing Phormidium, Oscillatoria and Schizothrix (Rapala et al. 2002). LPS are complex polymers composed of lipid A, a core oligosaccharide, and an O-specific polysaccharide chain consisting of repeating oligosaccharide units. Unlike in bacteria, they do not contain any phosphate in the lipid A core. LPS have endotoxic activity and are recognised as contact irritants. They can be involved in septic shock syndrome. It has been reported that LPS endotoxins may reinforce the effects of microcystins through inhibiting the activity of GST – the key enzymes involved in the detoxication of microcystins (Wiegand and Pflugmacher 2005). In animals and humans, they elicit pyrogenic, irritant and allergic responses. LPS produced by cyanobacteria are 10-times less toxic in mouse tests than those produced by pathogenic bacteria, such as Salmonella spp. (Keleti and Sykora 1982). However, high concentrations of the toxins were measured in cyanobacterial bloom samples. It is presumed that most of the toxins were produced by the associated Gram-negative bacteria (Rapala et al. 2002). Outbreaks of gastroenteritis, bath water fever and allergic disorders associated with high concentrations of endotoxins were reported in Pennsylvania, USA and Finland (Rapala et al. 2002). Due to a lack of axenic strains and the need for more advanced analytical techniques, our knowledge on LPS in cyanobacteria is limited.
CONCLUSIONS
To a large degree, harmful blooms of cyanobacteria have increased in frequency and intensity as a consequence of human activity. This phenomenon illustrates the complex web of interaction between biotic and abiotic environmental factors. There have been some spectacular events of human and animal acute poisoning caused by cyanobacterial blooms. However, health

H. Mazur-Marzec 104
hazards due to chronic exposure to the toxins is of greatest concern, as it may affect a wider population of people. Moreover, the harmful effects of such exposure have not been fully recognised. There are also some gaps in our knowledge about the occurrence and health impact of cyanobacterial toxins other than microcystins. The wider introduction of modern analytical techniques, such as LC-MS/MS, in laboratories monitoring water quality will probably result in new findings on other cyanobacterial toxins.
Due to climatic changes and high levels of eutrophication, the problem of harmful algal blooms is unlikely to improve in the near future. Therefore, some initiatives to establish guidelines for safe water usage have been undertaken. In many countries, the maximum acceptable levels for microcystins in drinking and recreational water have been determined based on biological, toxicological and epidemiological data. Still, there is a need for improved management of water resources and a limited input of nutrients to aquatic ecosytems.
The question is open of why cyanobacteria expend energy on the production of toxins that are most harmful to organisms not sharing the same habitat.
ACKNOWLEDGEMENTS
I wish to thank the State Committee for Scientific Research in Poland for financial assistance (project 0366/PO4/2003/25). Adam Krężel is warmly thanked for satellite images, Justyna Kobos and Agata Błaszczyk for their help with other figures.
REFERENCES Amorim A., Vasconcelos V., 1999. Dynamics of microcystins in the mussels Mytilus
galloprovincialis. Toxicon, 37, 1041-1052. An J., Carmichael W.W., 1994. Use of a colorimetric protein phosphatase inhibition assay and
enzyme linked immuno sorbent assay for the study of microcystin and nodularin. Toxicon 32, 1495-1507.
Anderson D.M., Kulis D.M., Doucette G.J., Gallagher J.C., Balech E., 1989. Biogeography of toxic dinoflagellates in the genus Alexandrium from the northern United States and Canada. Mar. Biol. 120, 467-478.
Beattie K.A., Kaya K., Codd G.A., 2000. The cyanobacterium Nodularia PCC 7804, of freshwater origin, produces [L-Har2] nodularin. Phytochemistry 54, 57-61.
Bouaïcha N., Maatouk I., 2004. Microcystin-LR and nodularin induce intracellular glutatione alteration, reactive oxygen species production and lipid peroxidation in primary cultured rat hepatocytes. Toxicology Letters 148, 53-63.
Bourne D.G., Jones G.J., Blakeley R.L., Jones A., Negri A.P., Riddles P., 1996. Enzymathic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin-LR. Appl. Environ. Microbiol. 62 (11), 4086-4094.
Burke W.A., Tester P.A., 2002. Skin problems related to non-infectious coastal micro organisms. Dermatologic Therapy 15, 10-17.

Phycotoxins – Review, part I 105
Bury N.R., McGeer J.C., Eddy F.B., Codd G.A., 1997. Liver damage in brown trout, Salmo trutta L., and rainbow trout, Onchorhynchus mykiss (Walbaum), following the administration of the cyanobacterial hepatotoxin microcystin-LR via the dorsal aorta. J. Fish Diseases 20, 209-215.
Cambella A.D., 2003. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 42, 420-447.
Carmichael W.W., Azevedo S.M.F.O., 2001. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 109, 663-668.
Codd G.A., Bell S.G., Kaya K., Ward C.J., Beattie K.A., Metcalf J.S., 1999. Cyanobacterial toxins, exposure routes and human health. Eur. J. Phycol. 34, 405-415.
Cox P.A., Banack S.A., Murch S.J., 2003. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among Chamorro people of Guam. Proc. Natl. Acad. Sci. USA, 100, 13380-13383.
Cox P.A., Banack S.A., Murch S.J., Rasmussen U., Tien G., Bidigare R.R., Metcalf J.S., Morrison L.F., Codd G.A., Bergman B., 2005. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA, 102, 5074-5078.
DeMott W.R., Hang Q., Carmichael W.W., 1991. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Oceanogr. 36, 1346-1357.
DeSilva E.D., Williams D.E., Andersen R.J., Klix H., Holmes C.F.B., Allen T.M., 1992. Motuporin, a potent protein phosphatases inhibitor isolated from the Papua New Guinea sponge Theonella swinhoei Gray. Tetrahedron Letters 33, 1561-1564.
Ding W.-X., Ong Ch.N., 2003. Role of oxidative stress and mitochondrial changes in cyanobacteria-induced apoptosis and hepatotoxicity. FEMS Microbiol. Lett. 220, 1-7.
Doan N.T., Rickards R.W., Rothschild J.M., Smith G.D., 2000. Allelopathic actions of the alkaloid 12-epi-hapalindole E isonitrile and calothrixin A from cyanobacteria of the genera Fischerella and Calothrix. J. Appl. Phycol. 12, 409-416.
Domingos P., Rubim T.K., Molica R.J.R., Azevedo S.M.F.O., Carmichael W.W., 1999. First report of microcystin production by picoplanktonic cyanobacteria isolated from a Northeast Brazilian drinking water supply. Environ. Toxicol. 14, 31-35.
Eriksson J.E., Grönberg L., Nygård S., Slotte J.P., Meriluoto J.A.O., 1990. Hepatocellular uptake of 3H-dihydromicrocystin-LR, a cyclic peptide toxin. Biochem. Biophys. Acta 1025, 60-66.
Falconer I.R., Humpage A.R., 1996. Tumour promotion by cyanobacterial toxins. Phycologia, 35, 74-79.
Falconer I.R., 2005. Cyanobacterial poisoning of livestock and people. [in:] Cyanobacterial toxins of drinking water supplies. Cylindrospermopsins and microcystins. I.R. Falconer (ed.), CRC Press, Boca Raton, Florida.
Fladmark K.E., Serres M.H., Lersen N.L., Yasumoto T., Aune T., Døskeland S.O., 1998. Sensitive detection of apoptogenic toxins in suspention cultures of rat and salmon hepatocytes. Toxicon 36, 1101-1114.
Francis G., 1878. Poisonous Australian lake. Nature (London) 18, 11-12. Granéli E. and Johansson N., 2003. Increase in the production of allelopathic substances by
Prymnesium parvum cells grown under N- or P- deficient conditions. Harmful Algae, 2, 135-145.
Griffiths D.J., Saker M.L., 2003. The Palm Island mystery disease 20 years on: a review of the research on cyanotoxin cylindrosprmopsin. Environ. Toxicol. 18, 78-93.
GuggerM., Lenoir S., Berger C., Ledreux A., Druart J.C., Humbert J.F., Guette C., Bernard C., 2005. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 45, 919-928.

H. Mazur-Marzec 106
Hallegraeff G.M., 1993. A review of harmful algal blooms and their apparent global increase. Phycologia, 33, 79-99.
Harada K.-I., 1996. Chemistry and detection of microcystins. [in:] Toxic Microcystis, M.F. Watanabe, K.-I. Harada, W.W. Carmichael, H. Fujiki (eds), CRC Press, Boca Raton, London, 103-148.
Harada K-I., Imanishi S., Kato H., Mizuno M., Ito E., Tsuji K., 2004. Isolation of Adda from microcystin-LR by microbial degradation. Toxicon 44, 107-109.
Helcom 2004, http://www.helcom.fi/environment2/ifs/ifs2004/en_GB/cyanobacteria/Hu Z., Liu Y., Li D., 2004. Physiological and biochemical analyses of microcystin-RR toxicity to
the cyanobacterium Synechococcus elongatus. Environ. Toxicol. 19, 571-577. Humpage K.R., Rositano J., Bretag A.H., Brown R., Baker P.D., Nicholson B.C., Steffensen
D.A., 1994. Paralytic shellfish poisons from Australian cyanobacterial blooms. Aust. J. Mar. Freshwater Res. 45 (5), 761-771.
Jones G.J., Orr P.T., 1994. Release and degradation of microcystin following algicide treatment of a Microcystis aeruginosa bloom in recreational lake, as determined by HPLC and protein phosphatase inhibition assay. Wat. Res. 28, 871-876.
Jurczak T., Tarczyńska M., Karlsson K., Meriluoto J., 2004. Characterization and diversity of cyanobacterial hepatotoxins (microcystins) in blooms from Polish freshwaters identified by liquid chromatography-electrospray ionisation mass spectrometry. Chromatogr. 59, 571-578.
Kankaanpää H., Vuorinen P.J., Sipiä V., Keinänen M., 2002. Acute effects and bioaccumulation of nodularin in sea trout (Salmo trutta m. trutta L.) exposed orally to Nodularia spumigena under laboratory conditions. Aquat Toxicol 61, 155-168.
Karjalainen M., Reinikainen M., Lindvall F., Spoof L., Meriluoto J.A.O., 2003. Uptake and accumulation of dissolved, radiolabeled nodularin in the Baltic Sea zooplankton. Environ Toxicol 18, 52-60.
Karlsson K.M., Kankaanpää H., Huttunen M., Meriluoto J., 2005. First observation of microcystin-LR in pelagic cyanobacterial blooms in the northern Baltic Sea. Harmful Algae 4, 163-166.
Keleti F., Sykora J.L., 1982. Production and properties of cyanobacterial endotoxins. Appl. Environ. Microbiol., 38, 471-477.
Kondo .F, Ikai Y., Oka H., Okumara M., Ishikawa N., Harada K.-I., Matsuura K., Murata H., Suzuki M., 1992. Formation, characterisation and toxicity of the glutathione and cysteine conjugate of toxic heptapeptide microcystins Chem. Res. Toxicol 5, 591-596.
Kononen K., Sivonen K., Lehtimäki J., 1993. Toxicity of phytoplankton blooms in the Gulf of Finland and Gulf of Bothnia, Baltic Sea. [in:] Toxic phytoplankton blooms in the sea, T.J. Smayda & Y. Shimizu (eds), Elsevier Sciences Publishers B.V., 269-273.
Krienitz L., Ballot A., Kotut K., Wiegand C., Pütz S., Metcalf J.S., Codd G.A., Pflugmacher S., 2003. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingoes at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 43, 141-148.
Kupier-Goodman T., Falconer I., Fitzgerald J., 1999. Human health aspect. [in:] Toxic cyanobacteria in water: a guide to their public health consequences, monitoring and management, I. Chorus & J. Bartram (eds.), WHO Publ., E. & F.N. Spon, London-New York, 41-111.
Lankoff A., Banasik A., Nowak M., 2002. Protective effect of melatonin against nodularin-induced oxidative stress. Arch. Toxicol. 76, 158-165.
Lehtiniemi M., Engström-Öst J., Karjalainen M., Kozlowski-Suzuki B., Viitasalo M., 2002. Fate of cyanobacterial toxins in the pelagic food web: transfer to copepods or to faecal pellets?, Mar. Ecol. Prog. Ser. 241, 13-21.

Phycotoxins – Review, part I 107
Matsunaga S., Moore R.E., Niemczura W.P., Carmichael W.W., 1989. Anatoxin-a(s) a potent anticholinesterase from Anabaena flos-aquae. J. Am. Chem.Soc. 111, 8021-8023.
Mazur H., Pliński M. 2003. Nodularia spumigena blooms and the occurrence of hepatotoxin in the Gulf of Gdańsk. Oceanologia 45, 305-316.
Mazur H., Lewandowska J., Błaszczyk A., Kot A., Pliński M., 2003. Cyanobacterial toxins in fresh and brackish waters of Pomorskie Province (Northern Poland). Oceanol. Hydrobiol. Stud. 32 (1), 15-26.
Metcalf J.S., Lindsay J., Beattie K.A., Birmingham S., Saker M.L., Trokne A.K., Codd G.A., 2002. Toxicity of cylindrospermopsin to the brine shrimp Artemia salina: comparison with protein synthesis inhibitors and microcystins. Toxicon 40, 1115-1120.
Namikoshi M., Choi B.W., Sakai R., Sun F., Rinehart K.L., Carmichael W.W.. Evans W.R., Cruz P., Munro M.H.G., Blunt J.W. 1994. New nodularin: A general method for structure assignment. J. Org. Chem. 59, 2349-2357.
Ohta T., Sueoka E., Iida N., Komori A., Suganuma M., Nishiwaki R., Tatematsu M., Kim S.J., Carmichael W.W., Fujiki H., 1994. Nodularin, a potent Inhibitor of protein phosphatases 1 and 2A, is a new environmental carcinogen in male F344 rat liver. Cancer Res. 54, 6402-6406.
Osborne N.J.T., Webb P.M., Shaw G.R., 2001. The toxins of Lyngbya majuscula and their human and ecological health effects. Environ. Int. 27, 381-392.
Park H.D., Watanabe M.F., Harada K.I., Nagai H., Suzuki M., Watanabe M., Hayashi H., 1993. Hepatotoxin (microcystin) and neurotoxin (anatoxin-a) contained in natural blooms and strains of cyanobacteria from Japanese waters. Nat. Toxins 1, 353-360.
Park H.-D., Watanabe M.F., 1996. Toxic Microcystis in eutrophic lakes. [in:] M.F. Watanabe, K.-I. Harada, W.W. Carmichael, Fujiki H. (edrs) Toxic Microcystis, CRC Press, Boca Raton, London, 57-77.
Pearl H.W., 2000. Marine Plankton. [in:] B.A. Whitton and M. Potts (eds), The Ecology of Cyanobacteria, Kluwer Academic Publishers, Netherlands. 121-148.
Pflugmacher S., Wiegand C., Oberman A., Beattie K.A., Krause E., Codd G.A., Steinberg C.E.W., 1998. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-LR: the first step of detoxication. Biochem. Biophys. Act 1425, 527-533.
Pouria S., de Andrade A., 1998. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 352, 21-26.
Rapala J., Lahti K., Räsänen L.A., Esala A.L., Niemelä, Sivonen K., 2002. Endotoxins associated with cyanobacteria and their removal during drinking water treatment. Water Res., 36, 2627-2635.
Reinikainen M., Lindvall F., Meriluoto J.A.O., Repka S., Sivonen K., Spoof L., Wahlsten M., 2002. Effects of dissolved cyanobacterial toxins on the survival and egg hatching of estuarine calanoid copepods. Mar. Biol. 140, 577-583.
Rinehart K.L., Harada K., Namikoshi M., Chen C., Harris C.E., Munro M.H.G., Blunt J.W., Mulligan P.E., Beasley V.R., Dahlem A.M., Carmichael W.W., 1988. Nodularin, microcystin and configuration of Adda. J. Am. Chem. Soc., 110, 8557-8558.
Rohrlack T., Dittmann E., Henning M., Börner T., Kohl J.-G., 1999. Role of microcystins in poisoning and food ingestion inhibition of Daphnia galeata caused by the cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 65, 737-739.
Runnegar M.T., Kong S.M., Zhong Y.Z., Lu S.C., 1995. Inhibition of reduced glutathione synthesis by cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Biopharmacol. 49, 219-225.

H. Mazur-Marzec 108
Saker M.L., Metcalf J.S., Codd G.A., Vasconcelos V.M., 2004. Accumulation and depuration of the cyanobacterial toxin cylindrospermopsin in the freshwater mussel Anodonta cygnea. Toxicon 43, 185-194.
Sellner K.G., 1997. Physiology, ecology, and toxic properties of marine cyanobacteria blooms. Limnol. Oceanogr., 42, 1089-1104.
Shawn G.R., Sukenik A., Livne A., Chiswell R.K., Smith M.J., Seawright A.A., Norris R.L., Eaglesham G.K., Moor M.R., 1999. Blooms of the cylindrospermopsin containing cyanobacterium Aphanizomenon ovalisporum (Forti), in newly constructed lakes, Queensland, Australia. Environ. Toxicol. 14, 167-177.
Shi L., Carmichael W.W., Miller I., 1995. Immuno-gold localisation of hepatotoxins in cyanobacterial cells. Arch. Microbiol. 163 (1), 7-15.
Singh D.P., Tyagi M.B., Kumar A., Thakur J.K., Kumar A., 2001. Antialgal activity of a hepatotoxin-producing cyanobacterium Microcystis aeruginosa. World J. Microbiol. Biotechnol. 17, 15-22.
Sipiä V.O., Kankaanpää H.T., Pflugmacher S., Flinkman J., Furey A., James K.J., 2002. Bioaccumulation and detoxication of nodularin in tissues of flounder (Platichthys flesus), mussels (Mytilus edulis, Dreissena polymorpha), and clams (Macoma balthica) from the northern Baltic Sea. Ecotoxicol. Environmental Safety 53, 305-311.
Sipiä V.O., Kankaanpää H., Flinkman J., Lahti K., Meriluoto J.A.O., 2001. Time-dependent accumulation of cyanobacterial hepatotoxins in flounders (Platichthys flesus) and mussels (Mytilus edulis) from the northern Baltic Sea. Environ. Toxicol. 16 (4), 330-336.
Sipiä V.O., Karlsson KM, Meriluoto JO, Kankaanpää HT., 2003. Eiders (Somateria mollissima) obtain nodularin, a cyanobacterial hepatotoxin, in Baltic Sea food web. Environ Toxicol 23, 1256-1260.
Sivonen K., Kononen K., Carmichael W.W., Dahlem A.M., Rinehart K.L., Kiviranta J., Niemelä I., 1989. Occurrence of the hepatotoxic cyanobacterium Nodularia spumigena in the Baltic Sea and structure of the toxin. Appl. Environ. Microbiol. 55 (8), 1990-1995.
Sivonen K., 1996. Cyanobacterial toxins and toxin production. Phycologia 35, 12-24. Sivonen K., Jones G. 1999. Cyanobacterial toxins. [in:] Toxic cyanobacteria in water: a guide to
their public health consequences, monitoring and management, I. Chorus & Bartram (red.), WHO Publ., E. & F.N. Spon, London-New York, 41-111.
Spoof L., Vesterkvist P., Lindholm T., Meriluoto J., 2003. Screening for cyanobacterial hepatotoxins, microcystins and nodularins in environmental water samples by reversed-phase liquid chromatography-electrospray ionization mass spectrometry. J. Chromatogr. 1020, 105-119.
Skulberg O.M., Carmichael W.W., Anderson R.A., Matsunaga S., Moore R.E., Skulberg R. 1992. Investigation of a neurotoxic Oscillatorialean strain (cyanophyceae) and its toxin. Isolation and characterisation of homoanatoxin-a. Environ. Toxicol. Chem. 11, 321-329.
Svensen C., Strogyloudi E., Wexels Riser C., Dahlmann J., Legrand C., Wassmann P., Granéli E., Pagou K., 2005. Reduction of cyanobacterial toxins through coprophagy in Mytilus edulis. Harmful Algae 4, 329-336.
Takenaka S., Watanabe M.F., 1997. Microcystin-LR degradation by Pseudomonas aeruginosa alkaline protease. Chemosphere 34:749-757.
Takenaka S. 1998. Formation of 3-amino-2,6,8-trimethyl-10-phenyldeca-4E, 6E-dienoic acid from microcystin-LR by the treatment with various proteases and its detection in mouse liver. Chemosphere 36: 2277-2282.
Tillett D., Dittmann E., Erhard M., von Döhren H., Börner T., Neilan B.A., 2000. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC 7806: an integrated peptide-polyketide synthetase system. Chem. Biol., 7, 753-764.

Phycotoxins – Review, part I 109
Tsuji K., Nalto S., Kondo F., Ishikawa N., Watanabe M.F., Suzuki M., Harada K.-I. 1994. Stability of microcystins from cyanobactreia: effect of light on decomposition and isomerization. Environ. Sci. Technol. 28, 173-177.
Twist H., Codd G.A. 1997. Degradation of the cyanobacterial hepatotoxin, nodularin, under light and dark conditions. FEMS Microbiol. Lett. 151, 83-88.
Tymińska A., Mazur-Marzec H., Graczyk M., Pliński M., 2005. Effect of two different Nodularia spumigena extracts on Danio rerio eggs mortality and hatching time. Oceanological and Hydrobiol. Stud. 34 Suppl. 3, 149-159.
Yoshizawa S., Matsushima R., Watanabe M.F., Harada K.-I., Ichihara A., Carmichael W.W. and Fujiki H. 1990. Inhibition of protein phosphatases by microcystins and nodularin associated with hepatotoxicity. J.Cancer Res. Clin. Oncol. 116, 609-614.
Wiegand C., Pflugmacher S., Oberemm A., Meems N., Beattie K., Steinberg C.E.W., Codd G.A., 1999. Uptake and effects of microcystin LR on detoxication enzymes of early life stages of the zebrafish (Danio rerio). Environ. Toxicol. 14, 89-95.
Wiegand C., Pflugmacher S., 2005. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. Appl. Pharm. 203, 201-218.
World Health Organization 1998. Guidelines for drinking water quality, 2 ed. Addendum to Vol. 1, pp. 13-14. Recommendations. Geneva, World Health Organization.