internodal elongation and orientation of cellulose microfibrils and microtubules in deepwater rice
TRANSCRIPT

Planta (1993)190:354-362 P l a n t a
�9 Springer-Verlag 1993
Internodal elongation and orientation of cellulose microfibrils and microtubules in deepwater rice Margret Sauter i, Robert W. Seagull 2, Hans Kende 1
1 MSU-DOE Plant Research Laboratory, Michigan State University, East Lansing, MI 48824-1312, USA 2 Southern Regional Research Center, U.S.D.A./A.R.S. Southern Regional Laboratory, 1100 Robert E. Lee Blvd., New Orleans, LA 70124, USA
Received: 5 October 1992 / Accepted: 5 November 1992
Abstract. Excised stem sections of deepwater rice (Oryza sativa L.) containing the highest internode were used to study the induction of rapid internodal elongation by gibberellin (GA). It has been shown before that this growth response is based on enhanced cell division in the intercalary meristem and on increased cell elongation. In both GA-treated and control stem sections, the basal 5- mm region of the highest internode grows at the fastest rate. During 24 h of GA treatment, the internodal elon- gation zone expands from 15 to 35 mm. Gibberellin does not promote elongation of internodes from which the intercalary meristem has been excised. The orientation of cellulose microfibrils (CMFs) is a determining factor in cell growth. Elongation is favored when CMFs are ori- ented transversely to the direction of growth while elon- gation is limited when CMFs are oriented in the oblique or longitudinal direction. The orientation of CMFs in parenchymal cells of GA-treated and control internodes is transverse throughout the internode, indicating that CMFs do not restrict elongation of these cells. Changes in CMF orientation were observed in epidermal cells, however. In the basal 5-mm zone of the internode, which includes the intercalary meristem, CMFs of the epider- mal cell walls are transversely oriented in both GA-treat- ed and control stem sections. In slowly growing control internodes, CMF orientation changes to the oblique as cells are displaced from this basal 5-mm zone to the re- gion above it. In GA-treated rapidly growing internodes, the reorientation of CMFs from the transverse to the oblique is more gradual and extends over the 35-mm length of the elongation zone. The CMFs of older epider- mal cells are obliquely oriented in control and GA-treat- ed internodes. The orientation of the CMFs parallels that of the cortical microtubules. This is consistent with the hypothesis that cortical microtubules determine the di- rection of CMF deposition. We conclude that GA acts on cells that have transversely oriented CMFs but does not
Abbreviations: CMF(s)=cellulose microfibril(s); GA=gibberellin; GA3 =gibberellic acid; MT(s)=microtubule(s)
Correspondence to: H. Kende; FAX: 1 (517)3539168
promote growth of cells whose CMFs are already obliquely oriented at the start of GA treatment.
Key words: Cellulose microfibril - Gibberellin In- ternodal elongation Microtubule- Oryza (growth)
Introduction
Deepwater rice is induced to grow rapidly when it be- comes submerged. Enhanced internodal elongation en- ables the plant to keep part of its foliage above the rising water surface and to survive flooding. The low partial pressure of oxygen in submerged internodes promotes ethylene biosynthesis, and ethylene trapped under water increases the responsiveness of the internodal tissue to gibberellin (GA). Thus, the growth response is ultimately elicited by GA (for a review see Kende 1987). Gib- berellin-induced elongation in deepwater rice can be separated into three components (Sauter and Kende 1992). The initial growth response is based on enhanced cell elongation in the intercalary meristem followed by promotion of cell division activity. Cells that emerge from the intercalary meristem during treatment with GA reach an up to threefold greater length than do cor- responding cells in control internodes (see also Bleecker et al. 1986). These results indicate that cells in the interca- lary meristem are the primary targets of GA action.
The direction of cell expansion is determined primari- ly by the arrangement of the cellulose microfibrils (CMFs) in the cell wall (for reviews see Green 1980; Taiz 1984). In elongating cells, CMFs are deposited in a trans- verse orientation with respect to the direction of growth. It is assumed that this circumferential reinforcement by CMFs restricts growth in girth but permits growth in length. Conversely, deposition of CMFs in an oblique or longitudinal direction with respect to the length axis of an organ limits elongation. A close correlation between elongation and transverse orientation of CMFs has been demonstrated in developing cotton fibers (Seagull 1986),

M. Sauter et al. : Cellulose microfibril orientation in rice internodes 355
in maize coleoptiles (Bergfeld et al. 1988), in coleoptiles and mesocotyls of Arena ( Iwata and Hoge tsu 1989) and in Vigna angularis epicotyls (Takeda and Sh ibaoka 1981a, b).
There is a large body of c i rcumstan t ia l evidence that the o r ien ta t ion of newly deposi ted C M F s is de te rmined by the o r i en ta t ion of cortical mic ro tubu les (MTs) (for reviews see Hepler and Palevitz 1974; R o b i n s o n a nd Quader 1982; Lloyd 1984; Seagull 1989; Gidd ings and Staehelin 1991). T rea tmen t with G A results in t ransverse o r ien ta t ion of MTs in epicotyls of Viyna angularis and peas (Shibaoka 1974; Akashi and Sh ibaoka 1987), in mesocotyls of maize (Mita and K a t s u m i 1986; I sh ida a nd Ka t sumi 1991) and in hypocotyls of cucumber (Ishida and Ka t sumi 1992). T rea tmen t with G A also results in t ransverse deposi t ion of C M F s in epicotyls of Vigna angularis (Takeda and Sh ibaoka 1981b). Based on these observat ions , it has been suggested that G A induces e longat ion by causing r ea r rangemen t of cortical M T s into a pr imar i ly t ransverse di rect ion which, in turn , leads to transverse o r ien ta t ion of the newly formed cellulose microfibrils.
In the exper iments described below, we examined further the role of the in tercalary mer is tem in the growth response of deepwater rice, and we correlated the o r ien ta t ion of C M F s and MTs with the rate of growth in different regions of the in ternode. Our results show that the presence of the in tercalary mer is tem is necessary for the p r o m o t i o n of in te rnoda l growth by G A and that a t ransverse o r i en ta t ion of C M F s and MTs is m a i n t a i n e d over a longer distance in epidermal cells of GA- t rea t ed in ternodes t han in epidermal cells of cont ro l in ternodes . This s t ruc tura l feature of the cell wall may be the basis for G A - e n h a n c e d cell e longa t ion in deepwater rice.
Materials and methods
Plant material and growth conditions. Seeds of deepwater rice (Oryza sativa L., cv. Pin Gaew 56) were obtained from the International Rice Research Institute (Los Bafios, Philippines). Growth con- ditions were as described by Sauter and Kende (1992). Rice stem sections were excised from tillers according to Raskin and Kende (1984a). They had a length of 20 cm and included the youngest internode between the highest and the second highest node (see Fig. 2). Stem sections were placed in a 100-ml beaker containing 30 ml of distilled water or a solution of 50 laM gibberellic acid (GA3) and were incubated at 25 ~ C in a water-saturated atmosphere under continuous light (cool-white fluorescent tubes; General Electric, Cleveland., Ohio, USA; photosynthetic flux density 35 ~tmol. m-e . s-I).
Sites and rates of internodal growth. Stem sections were marked at 5-mm intervals starting above the second highest node by piercing a fine needle through the leaf sheath to give a small wound mark on the internode. Growth was determined by measuring the increase in the distance between the centers of the marks. Wounding reduced the overall growth response by 1~15% compared to unwounded control stem sections. To determine the distribution of growth rates correctly (Silk 1984), the displacement of the marks was kept small in relation to the distance between the marks. One batch of stem sections was treated with 50 laM GA3 for 24 h, marked, and growth rates were determined after 45 and 90 min. The growth rates were essentially the same over both time intervals. A second batch of stem sections was treated with 50 gM GA3 for 48 h, marked, and
growth rates were measured over a 45-min period. Control stem sections were preincubated in distilled water for 3 h to permit the burst of growth following excision to dissipate (Sauter and Kende 1992); they were then marked. Because the growth rate of control internodes after 24 h of incubation in water is approx. 25-fold lower than that of GA-treated internodes (see, e.g., Fig. 5A in Sauter and Kende 1992), it was impossible to determine the displacement of marks in control internodes over short time periods. To localize growth along the internode of control stem sections, measurements were taken 24 h after marking. The growth rates derived from these determinations represent, therefore, average values for the entire 24-h period.
Localization of the GA-responsive tissue. Stem sections were excised from rice tillers as described. After preincubation in distilled water for 3 h, they were shortened further according to the scheme shown in Fig. 2. The remaining part of each stem section, which included different regions of the internode, was incubated in distilled water or in 50 gM GA3 solution for 45 h under the conditions described above for whole stem sections. Internodes were also marked with a needle at 2.5-mm intervals as described above and incubated in 50 gM GA 3 solution or water immediately after marking.
Calcofluor stainh~g of CMFs. Excised stem sections were incubated in distilled water or in 50 laM GA 3 solution. After 48 h, four 1-cm segments were cut from the GA-treated internodes starting above the second highest node. A 1-cm segment from the oldest part of the internode just below the highest node was also excised. Control internodes were cut similarly except that the first 1-cm segment above the second highest node was further subdivided into two 5-ram segments. Two 5-mm segments were also isolated from the internodal region above the second highest node of air-grown plants at 0 h. These isolated segments were fixed in 4% paraformaldehyde in 0.1 M sodium-phosphate-buffer (pH 7.4), washed in the same buffer, embedded in mounting medium (Histoprep; Fisher Diag- nostic, Orangeburg, N.Y., USA) and frozen in liquid nitrogen. Longitudinal sections of 7 ~tm thickness were cut with a cryocut microtome (2800 Frigocut M, Reichert, Vienna, Austria) and attached to glass slides coated with gelatine. The slides were heated at 40 ~ C for 30 min, washed three times in phosphate-buffered saline [PBS= 15 mM phosphate buffer, 0.7% (w/v) NaC1, pH 7.5] for 3 min each wash, stained for 1 min with a 0.01% (w/v) solution of calcofluor (Fluorescence Brightener M2R, Sigma, St. Louis, Mo., USA) in PBS and washed again two times in PBS for 3 min each wash. Excess solution was removed by blotting, and the sections were embedded in medium containing 0.01% (w/v) n-propyl gallate and 50% glycerol in PBS. The sections were examined with a fluo- rescence microscope (Zeiss, Oberkochen, Germany). The orienta- tion of CMFs was determined from photomicrographs taken with Kodak TriX Pan film (400 ASA).
Electron microscopy. Stem sections were incubated for 48 h in distilled water or in 50 gM GA 3 solution to induce rapid growth. Cylindrical segments, 3 mm in length, were excised from control internodes 1-4 and 6-9 mm above the second highest node and also from the oldest part of the internode just below the highest node. In internodes treated with GA3, 3-mm segments were isolated 4-7 and 24-27 mm above the node and from the oldest part of the internode. The segments were cut lengthwise into four pieces and fixed in 4% glutaraldehyde in 0.1 M sodium cacodylate (pH 6.8), at room temperature for 1 d. They were washed three times in 0.1 M sodium cacodylate (pH 6.8), for 20 min each time and postfixed in 1% osmium tetroxide in sodium-cacodylate buffer. They were wash- ed again as before, dehydrated in ethanol and embedded in 25%, 50%, 75% and two times in 100% Spurr's resin (firm) (Spurr 1969) for 8-12 h at each gradation. The resin was baked at 65 ~ C for 20 h. Thin sections were cut on a Reichert Ultracut E microtome using a diamond knife and stained with saturated uranyl acetate in 100% methanol and 0.25% lead citrate (pH 12.5). Samples were examined using a Philips (Eindhoven, The Netherlands) 300 electron micros- cope and photographed on Kodak ISO 128 film.
356
0.04 A
M. Sauter et al. : Cellulose microfibril orientation in rice internodes
0.8
~ B
0.03
e-
E E
0.02
o
0.01
0.6
E o.s E Q)
0.4
~o 0.3
0 ~ 5 10 15 20 25 30 35 40 45 50
D i s t a n c e f r o m N o d e [ m m ]
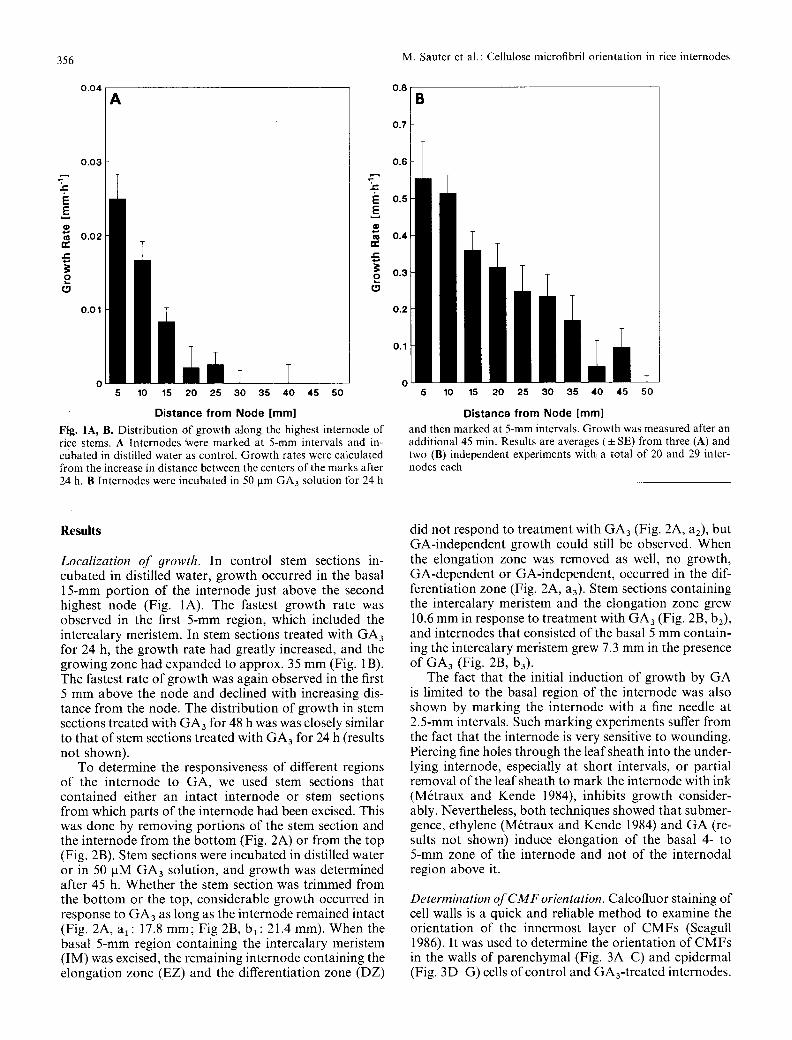
Fig. 1A, B. Distribution Of growth along the highest internode of rice stems. A Internodes Were marked at 5-mm intervals and in- cubated in distilled water as control. Growth rates were calculated from the increase in distance between the centers of the marks after 24 h. B Internodes were incubated in 50 gm GA3 solution for 24 h
0.2
0.1
5 10 15 20 25 30 35 40 45 50
Distance from Node [mm]
and then marked at 5-mm intervals. Growth was measured after an additional 45 min. Results are averages (• SE) from three (A) and two (B) independent experiments with a total of 20 and 29 inter- nodes each
Results
Localization of 9rowth. In control stem sections in- cubated in distilled water, growth occurred in the basal 15-mm port ion of the internode just above the second highest node (Fig. 1A). The fastest growth rate was observed in the first 5-mm region, which included the intercalary meristem. In stem sections treated with GA3 for 24 h, the growth rate had greatly increased, and the growing zone had expanded to approx. 35 mm (Fig. 1B). The fastest rate of growth was again observed in the first 5 mm above the node and declined with increasing dis- tance from the node. The distribution of growth in stem sections treated with GA3 for 48 h was was closely similar to that of stem sections treated with GA3 for 24 h (results not shown)�9
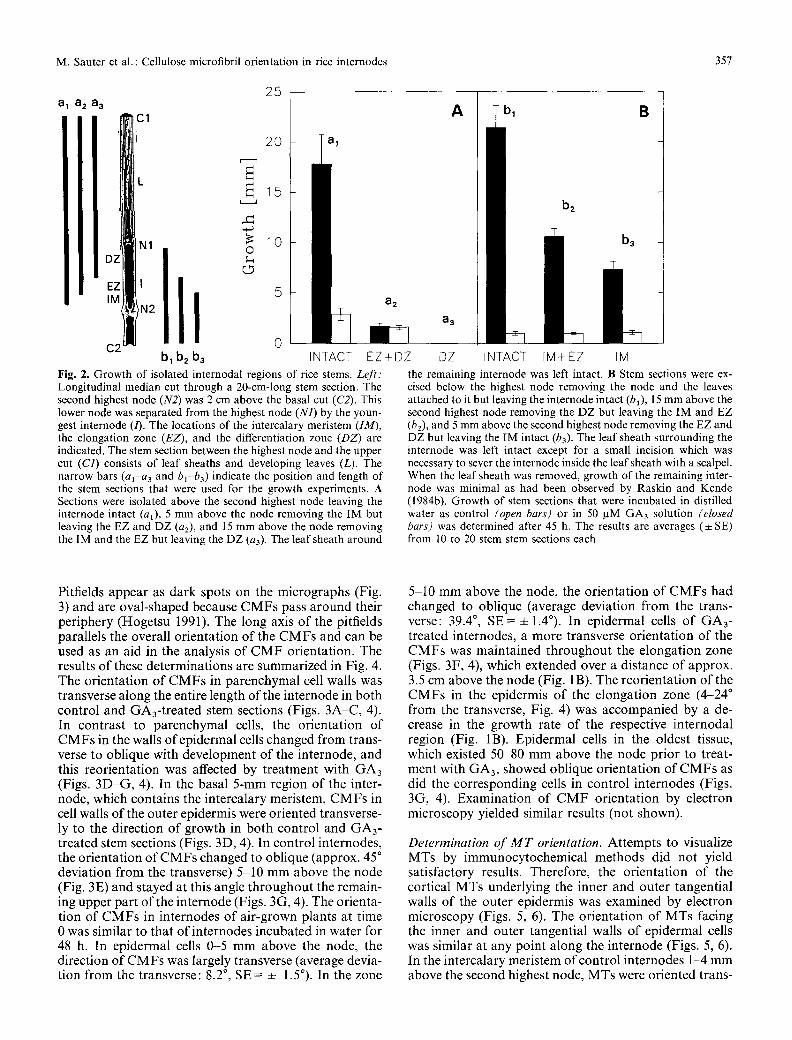
To determine the responsiveness of different regions of the internode to GA, we used stem sections that contained either an intact internode or stem sections from which parts of the internode had been excised. This was done by removing portions of the stem section and the internode from the bot tom (Fig. 2A) or from the top (Fig. 2B). Stem sections were incubated in distilled water or in 50 gM GA3 solution, and growth was determined after 45 h. Whether the stem section was trimmed from the bot tom or the top, considerable growth occurred in response to GA3 as long as the internode remained intact (Fig. 2A, al: 17.8 mm; Fig 2B, bl : 21.4 mm). When the basal 5-mm region containing the intercalary meristem (IM) was excised, the remaining internode containing the elongation zone (EZ) and the differentiation zone (DZ)
did not respond to treatment with GA3 (Fig. 2A, a2), but GA-independent growth could still be observed. When the elongation zone was removed as well, no growth, GA-dependent or GA-independent, occurred in the dif- ferentiation zone (Fig. 2A, a3). Stem sections containing the intercalary meristem and the elongation zone grew 10.6 mm in response to treatment with GA3 (Fig. 2B, b2) , and internodes that consisted of the basal 5 mm contain- ing the intercalary meristem grew 7.3 mm in the presence of GA3 (Fig. 2B, b3).
The fact that the initial induction of growth by GA is limited to the basal region of the internode was also shown by marking the internode with a fine needle at 2.5-mm intervals. Such marking experiments suffer from the fact that the internode is very sensitive to wounding. Piercing fine holes through the leaf sheath into the under- lying internode, especially at short intervals, or partial removal of the leaf sheath to mark the internode with ink (M6traux and Kende 1984), inhibits growth consider- ably. Nevertheless, both techniques showed that submer- gence, ethylene (M6traux and Kende 1984) and GA (re- sults not shown) induce elongation of the basal 4- to 5-mm zone of the internode and not of the internodal region above it.
Determination of CMF orientation. Calcofluor staining of cell walls is a quick and reliable method to examine the orientation of the innermost layer of CMFs (Seagull 1986). It was used to determine the orientation of CMFs in the walls of parenchymal (Fig. 3A-C) and epidermal (Fig. 3D-G) cells of control and GA3-treated internodes.
M. Sauter et al. : Cellulose microfibril orientation in rice internodes 357
a a 2 a 3 31
N1
:'" Itl IM .~N2
C2 bl b2 b3
25
20 i' 5
0 IN TACT
a2
EZ+DZ Fig. 2. Growth of isolated internodal regions of rice stems. Left: Longitudinal median cut through a 20-cm-long stem section. The second highest node (N2) was 2 cm above the basal cut (C2). This lower node was separated from the highest node (N1) by the youn- gest internode (/). The locations of the intercalary meristem (IM), the elongation zone (EZ), and the differentiation zone (DZ) are indicated. The stem section between the highest node and the upper cut (C1) consists of leaf sheaths and developing leaves (L). The narrow bars (al-a a and bl-ba) indicate the position and length of the stem sections that were used for the growth experiments. A Sections were isolated above the second highest node leaving the internode intact (al), 5 mm above the node removing the IM but leaving the EZ and DZ (a2), and 15 mm above the node removing the IM and the EZ but leaving the DZ (a3). The leaf sheath around
A bl B
bz
b3
a3
DZ INTACT IM+EZ IM the remaining internode was left intact. B Stem sections were ex- cised below the highest node removing the node and the leaves attached to it but leaving the internode intact (bl), 15 mm above the second highest node removing the DZ but leaving the IM and EZ (b2), and 5 mm above the second highest node removing the EZ and DZ but leaving the IM intact (b3). The leaf sheath surrounding the internode was left intact except for a small incision which was necessary to sever the internode inside the leaf sheath with a scalpel. When the leaf sheath was removed, growth of the remaining inter- node was minimal as had been observed by Raskin and Kende (1984b). Growth of stem sections that were incubated in distilled water as control (open bars) or in 50 /aM GA3 solution (closed bars) was determined after 45 h. The results are averages (• SE) from 10 to 20 stem stem sections each
Pitfields appear as dark spots on the micrographs (Fig. 3) and are oval-shaped because CMFs pass around their periphery (Hogetsu 1991). The long axis of the pitfields parallels the overall orientation of the CMFs and can be used as an aid in the analysis of C M F orientation. The results of these determinations are summarized in Fig. 4. The orientation of CMFs in parenchymal cell walls was transverse along the entire length of the internode in both control and GA3-treated stem sections (Figs. 3A-C, 4). In contrast to parenchymal cells, the orientation of CMFs in the walls o f epidermal cells changed f rom trans- verse to oblique with development of the internode, and this reorientation was affected by treatment with GA3 (Figs. 3D-G, 4). In the basal 5-mm region of the inter- node, which contains the intercalary meristem, CMFs in cell walls o f the outer epidermis were oriented transverse- ly to the direction of growth in both control and GA3- treated stem sections (Figs. 3D, 4). In control internodes, the orientation of CMFs changed to oblique (approx. 45 ~ deviation f rom the transverse) 5-10 m m above the node (Fig. 3E) and stayed at this angle throughout the remain- ing upper part of the internode (Figs. 3G, 4). The orienta- tion of CMFs in internodes of air-grown plants at time 0 was similar to that of internodes incubated in water for 48 h. In epidermal cells 0-5 m m above the node, the direction of CMFs was largely transverse (average devia- tion f rom the transverse: 8.2 ~ SE = =k 1.5~ In the zone
5-10 m m above the node, the orientation of CMFs had changed to oblique (average deviation f rom the trans- verse: 39.4 ~ SE = + 1.4~ In epidermal cells of GA3- treated internodes, a more transverse orientation of the CMFs was maintained throughout the elongation zone (Figs. 3F, 4), which extended over a distance of approx. 3.5 cm above the node (Fig. 1B). The reorientation of the CMFs in the epidermis of the elongation zone (4-24 ~ from the transverse, Fig. 4) was accompanied by a de- crease in the growth rate of the respective internodal region (Fig. 1B). Epidermal cells in the oldest tissue, which existed 50 80 m m above the node prior to treat- ment with GA3, showed oblique orientation of CMFs as did the corresponding cells in control internodes (Figs. 3G, 4). Examinat ion of C M F orientation by electron microscopy yielded similar results (not shown).
Determination of M T orientation. Attempts to visualize MTs by immunocytochemical methods did not yield satisfactory results. Therefore, the orientation of the cortical MTs underlying the inner and outer tangential walls of the outer epidermis was examined by electron microscopy (Figs. 5, 6). The orientation of MTs facing the inner and outer tangential walls of epidermal cells was similar at any point along the internode (Figs. 5, 6). In the intercalary meristem of control internodes 1-4 m m above the second highest node, MTs were oriented trans-
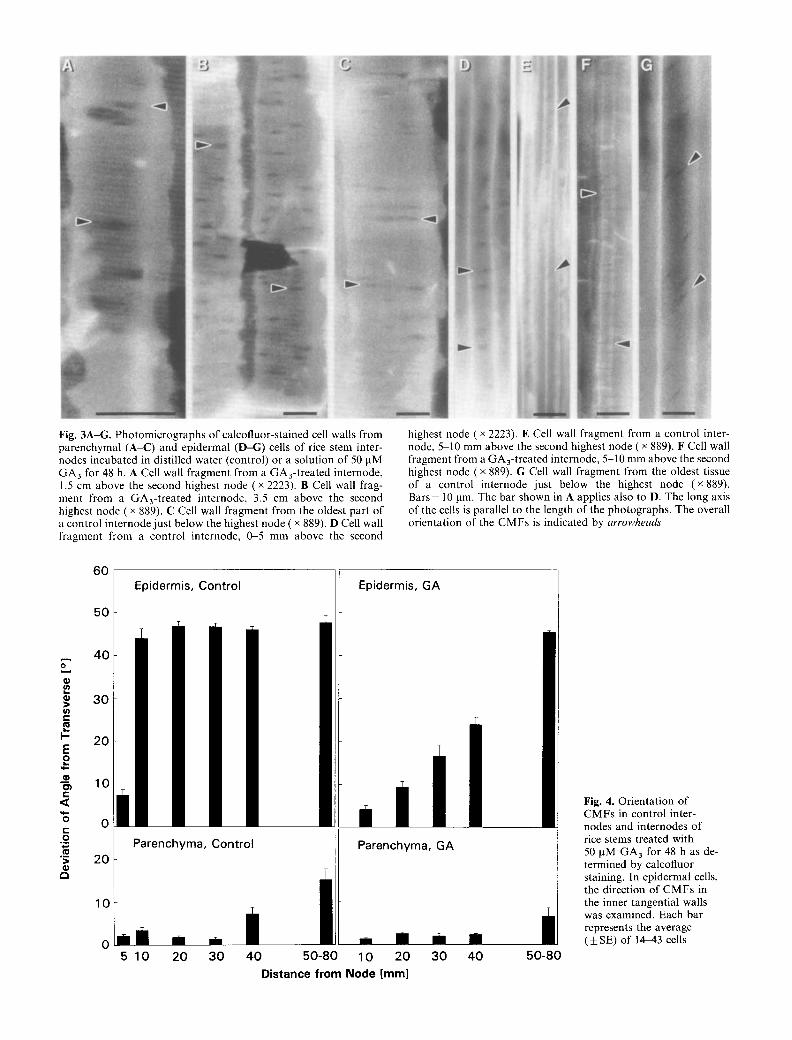
Fig. 3A-G. Photomicrographs of calcofluor-stained cell walls from parenchymal (A-C) and epidermal (D-G) cells of rice stem inter- nodes incubated in distilled water (control) or a solution of 50 pM GA3 for 48 h. A Cell wall fragment from a GA3-treated internode, 1.5 cm above the second highest node ( • 2223). B Cell wall frag- ment from a GA3-treated internode, 3.5 cm above the second highest node ( • 889). C Cell wall fragment from the oldest part of a control internode just below the highest node ( • 889). D Cell wall fragment from a control internode, 0-5 mm above the second
highest node ( x 2223). E Cell wall fragment from a control inter- node, 5 10 mm above the second highest node ( x 889). F Cell wall fragment from a GA3-treated internode, 5-10 mm above the second highest node ( x 889). G Cell wall fragment from the oldest tissue of a control internode just below the highest node (x 889). Bars = 10 lam. The bar shown in A applies also to D. The long axis of the cells is parallel to the length of the photographs. The overall orientation of the CMFs is indicated by arrowheads
60 ider , Control Epidermis, GA
5O
E 40
30
i! ; o t- o .=
:= r~
2 0
10
Parenchyma, Control
5 10 20 3 0 4 0 5 0 - 8 0 10 20 3 0 4 0 5 0 - 8 0
Distance from Node [mm]
Fig. 4. Orientation of CMFs in control inter- nodes and internodes of rice stems treated with 50 pM G A 3 for 48 h as de- termined by calcofluor staining. In epidermal cells, the direction of CMFs in the inner tangential walls was examined. Each bar represents the average (+_SE) of 1443 cells
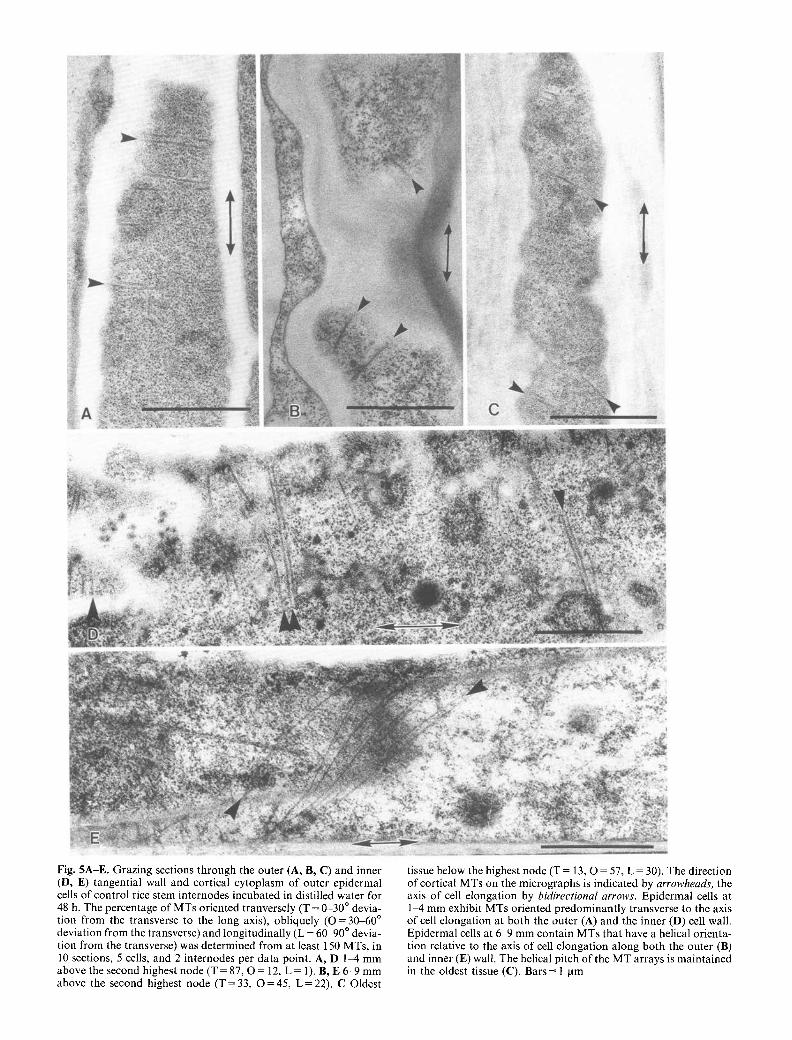
Fig. 5A-E. Grazing sections through the outer (A, B, C) and inner (D, E) tangential wall and cortical cytoplasm of outer epidermal cells of control rice stem internodes incubated in distilled water for 48 h. The percentage of MTs oriented tranversely (T = 0-30 ~ devia- tion from the transverse to the long axis), obliquely ( O = 30-60 ~ deviation from the transverse) and longitudinally (L = 6(~90 ~ devia- tion from the transverse) was determined from at least 150 MTs, in 10 sections, 5 cells, and 2 internodes per data point. A, D 1-4 mm above the second highest node (T = 87, O = 12, L = 1). B, E 6-9 mm above the second highest node (T = 33, O = 45, L = 22). C Oldest
tissue below the highest node (T = 13, O = 57, L = 30). The direction of cortical MTs on the micrographs is indicated by arrowheads, the axis of cell elongation by bidirectional arrows. Epidermal ceils at 1-4 mm exhibit MTs oriented predominantly transverse to the axis of cell elongation at both the outer (A) and the inner (D) cell wall. Epidermal cells at 6-9 mm contain MTs that have a helical orienta- tion relative to the axis of cell elongation along both the outer (B) and inner (E) wall. The helical pitch of the MT arrays is maintained in the oldest tissue (C). Bars = l lam
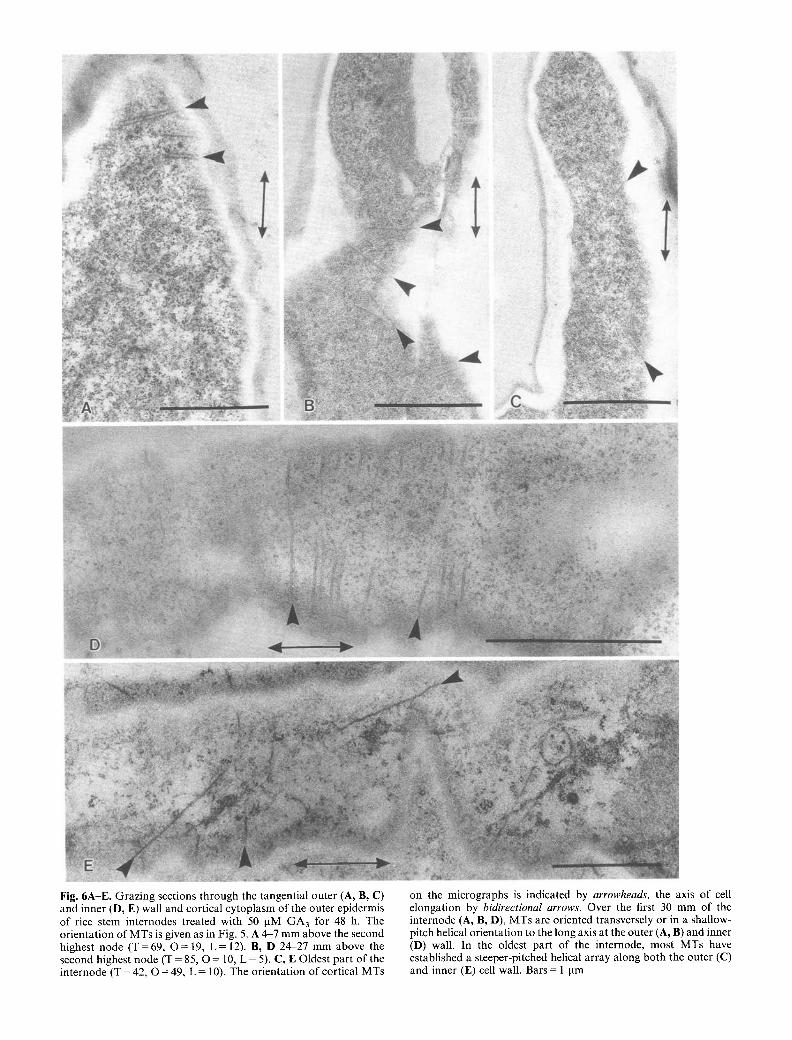
Fig. 6A-E. Grazing sections through the tangential outer (A, B, C) and inner (D, E) wall and cortical cytoplasm of the outer epidermis of rice stem internodes treated with 50 pM GA3 for 48 h. The orientation of MTs is given as in Fig. 5. A 4-7 mm above the second highest node (T=69 , O = 19, L = 12). B, D 24-27 mm above the second highest node (T = 85, O = 10, L = 5). C, E Oldest part of the internode (T = 42, O = 49, L = 10). The orientation of cortical MTs
on the micrographs is indicated by arrowheads, the axis of cell elongation by bidirectional arrows. Over the first 30 mm of the internode (A, B, D), MTs are oriented transversely or in a shallow- pitch helical orientation to the long axis at the outer (A, B) and inner (D) wall. In the oldest part of the internode, most MTs have established a steeper-pitched helical array along both the outer (C) and inner (E) cell wall. Bars = 1 pm

M. Sauter et al.: Cellulose microfibril orientation in rice internodes 361
versely to the direction of growth (Fig. 5A, D). At 6-9 mm above the node, MT orientation had changed to predominantly oblique (Fig. 5B, E). Mostly oblique orientation of MTs was also found in the oldest part of the internode just below the highest node (Fig. 5C). In contrast to the cortical MTs of control internodes, those of GA3-treated internodes remained in a more transverse orientation throughout the elongation zone (Fig. 6A, B, D). Microtubules in the oldest part of GA3-treated inter- nodes showed mostly oblique orientation (Fig. 6C, E).
Discussion
When rice internodes were induced to grow rapidly by treatment with GA3 for 24 or 48 h, the growing zone expanded from 1.5 to 3.5 cm (Fig. 1). The basal 5-mm zone of the internode, which included the intercalary meristem, grew at the fastest rate in both control and GA3-treated stem sections.
We have shown previously that the intercalary meri- stem is the primary site of GA action in deepwater-rice internodes (Sauter and Kende 1992). Upon treatment with GA3, cells in the intercalary meristem are induced to elongate and to divide. The intercalary meristem has been localized anatomically (Bleecker et al. 1986; Sauter and Kende 1992) and by autoradiography following la- beling with [3H]thymidine (M6traux and Kende 1984). Synthesis of DNA and mitotic activity in the intercalary meristem have been determined by measuring the incor- poration of [3H]thymidine into DNA and by flow cyto- metry, respectively (Sauter and Kende 1992). The inter- calary meristem occupies a zone of ca. 3 mm at the base of the youngest internode just above the second highest node. The excision experiments described above showed that the internodal zone containing the intercalary meri- stem was required and sufficient by itself for the induc- tion of growth by GA3 (Fig. 2A, B). Internodes from which this zone had been excised did not respond to GA3 (Fig. 2A), but growth of internodes which had been trimmed down to the basal 5-mm internodal zone con- taining the intercalary meristem and which lacked the elongation and differentiation zone was promoted by GA3 (Fig. 2B). The reduced growth response in GA3- treated stem sections lacking the elongation and/or the differentiation zone (Fig. 2B) might have been due, in part, to the loss of starch reserves which are normally mobilized from these older parts of the internode to sustain rapid elongation (Raskin and Kende 1984c). The results of our excision experiments are in agreement with marking experiments which showed that submergence and application of ethylene or GA 3 induced growth of only the basal 4-5 mm of the internode (M6traux and Kende 1992, and unpublished results).
Ceils that emerged from the intercalary meristem during GA treatment reached an up to threefold greater final length than did cells in control internodes (Sauter and Kende 1992). Gibberellin enhanced growth of cells that were in the intercalary meristem at the time of GA application but did not stimulate growth of cells that had entered the elongation zone prior to GA treatment
(Sauter and Kende 1992). This difference in response may be determined by the orientation of CMFs in the cell walls of the outer epidermis. In the internodal zone 0-5 mm above the node, which contains the intercalary meristem, epidermal cells had transversely oriented CMFs. In slowly growing, control internodes, the orientation of CMFs in the internodal zone 5-10 mm above the node had changed to the oblique (Fig. 4). This oblique orientation was maintained over the remainder of the internode. In rapidly growing, GA3-treated inter- nodes, the re-orientation of CMFs from the transverse to the oblique was gradual and extended over the entire elongation zone, i.e. over the basal 35-mm region of the internode (Figs. 1, 4). Hence, there was a close correla- tion between the rate of growth along the internode and the orientation of the CMFs in the walls of epidermal cells, as has been found in Arena coleoptiles and meso- cotyls, and pea epicotyls (Iwata and Hogetsu 1989), epi- cotyls of Vigna angularis (Takeda and Shibaoka 1981a, b) and corn coleoptiles (Bergfeld et al. 1988). The orienta- tion of CMFs in parenchyma cells was transverse along the entire internode of both control and GA3-treated stem sections (Fig. 4). Our results support the notion that an oblique to longitudinal orientation of CMFs restricts elongation (for reviews see Green 1980; Taiz 1984) and that the epidermal cell walls are growth-limiting struc- tures while the walls of parenchymal cells do not restrict growth (for a review see Kutschera 1989).
Cellulose microfibrils were oriented parallel to the cortical MTs at the tangential surfaces of the outer epi- dermal cells. This parallel arrangement was observed in both control and GA3-treated internodes and is consis- tent with the hypothesis that cortical MTs play a role in orienting CMFs (Hepler and Palevitz 1974; Robinson and Quader 1982; Lloyd 1984; Seagull 1989; Giddings and Staehelin, 1991). Since GA 3 was ineffective in pro- moting growth of cells that had been displaced from the zone of the intercalary meristem prior to start of hor- mone treatment and that had obliquely oriented MTs and CMFs, we propose that GA cannot cause reorienta- tion of MTs and CMFs; it can only enhance elongation of cells whose MTs and CMFs are still largely transverse in orientation. Herein may lie one of the basic differences between GA and auxin action. At least in rice internodes, induction of growth by GA is restricted to cells with transversely oriented cortical MTs and CMFs. In con- trast, auxin is known to cause reorientation of cortical MTs and CMFs from the oblique/longitudinal to the transverse and to promote elongation of cells that have stopped growing (Bergfeld et al. 1988).
It is not known whether GA has a direct effect on the orientation of MTs in deepwater-rice internodes. During GA-induced rapid growth, cells move through the elon- gation zone at a greatly accelerated rate (Sauter and Kende 1992). Therefore, the reorientation of MTs and CMFs in GA-treated rice internodes may be delayed as a function of position rather than time.
This work was supported by grant No. DCB 9103747 from the National Science Foundation and grant No. DE-FG02- 90ER20021 from the Department of Energy. During part of this

362 M. Sauter et al.: Cellulose microfibril orientation in rice internodes
work, M.S. was the recipient of a fellowship from the Deutsche Forschungsgemeinschaft. The authors thank Dr. Natasha Raikhel for use of her microscope facilities and Drs. Deborah Delmer and Paul Green for critically reading the manuscript of this paper.
References
Akashi, T., Shibaoka, H. (1987) Effects of gibberellin on the ar- rangement and the cold stability of cortical microtubules in epidermal cells of pea internodes. Plant Cell Physiol. 28, 339-348
Bergfeld, R., Speth, V., Schopfer, P. (1988) Reorientation of mi- crofibrils and microtubules at the outer epidermal wall of maize coleoptiles during auxin-mediated growth. Bot. Acta 101, 57-67
Bleecker, A.B., Schuette, J.L., Kende, H. (1986) Anatomical analy- sis of growth and developmental patterns of deepwater rice. Planta 169, 490-497
Giddings, T.H., Jr., Staehelin, L.A. (1991) Microtubule-mediated control of microfibril deposition: a re-examination of the hypothesis. In: The cytoskeletal basis of plant growth and form, pp. 85-99, Lloyd, C.W., ed. Academic Press, London
Green, P.B. (1980) Organogenesis- a biophysical view. Annu. Rev. Plant Physiol. 31, 51-82
Hepler, P.B., Palevitz, B.A. (1974) Microtubules and microfila- ments. Annu. Rev. Plant Physiol. 25, 309 362
Hogetsu, T. (1991) Mechanism for formation of the secondary wall thickening in tracheary elements: Microtubules and microfila- ments of tracheary elements of Pisum sativum L. and Comme- lina communis L. and the effects of aminoprophosmethyl. Planta 185, 19~200
Ishida, K., Katsumi, M. (1991) Immunofluorescence microscopical observation of cortical microtubule arrangement as affected by gibberellin in d 5 mutant of Zea mays L. Plant Cell Physiol. 32, 409~417
Ishida, K., Katsumi, M. (1992) Effects of gibberellin and abscisic acid on the cortical microtubule orientation in hypocotyl cells of light-grown cucumber seedlings. Int. J. Plant Sci. 153, 155-163
Iwata, K., Hogetsu, T. (1989) Orientation of wall microfibrils in Arena coleoptiles and mesocotyls and in Pisum epicotyls. Plant Cell Physiol. 30, 749-757
Kende, H. (1987) Studies on internodal growth using deepwater rice. In: Physiology of cell expansion during plant growth, pp. 227 238, Cosgrove, D.J., Knievel, D.P., eds. American Society of Plant Physiologists, Rockville, Md., USA
Kutschera, U. (1989) Tissue stress in growing plant organs. Physiol. Plant. 77, 157-163
Lloyd, C.W. (1984) Towards a dynamic helical model for the in- fluence of microtubules on wall patterns in plants. Int. Rev. Cytol. 86, 1-51
M6traux, J.-P., Kende, H. (1984) The cellular basis of the elonga- tion response in submerged deep-water rice. Planta 160, 73 77
Mira, T., Katsumi, M. (1986) Gibberellin control of microtubule arrangement in the mesocotyl epidermal cells of the d5 mutant of Zea mays L. Plant Cell Physiol. 27, 651-659
Raskin, I., Kende, H. (1984a) Regulation of growth in stem sections of deepwater rice. Planta 160, 66-72
Raskin, I., Kende, H. (1984b) Role of gibberellin in the growth response of deep water rice. Plant Physiol. 76, 947-950
Raskin, I., Kende, H. (1984c) Effect of submergence on transloca- tion, starch content and amylolytic activity in deep-water rice. Planta 162, 556-559
Robinson, D.G., Quader, H. (1982) The microtubule-microfibril syndrome. In : The cytoskeletal basis of plant growth and form, pp. 109-126, Lloyd, C.W., ed. Academic Press, London
Sauter, M., Kende, H. (1992) Gibberellin-induced growth and reg- ulation of the cell division cycle in deepwater rice. Planta 188, 362 368
Seagull, R.W. (1986) Changes in microtubule orientation and in wall microfibril orientation during in vitro cotton fiber develop- ment: an immunofluorescent study. Can. J. Bot. 64, 1373 1381
Seagull, R.W. (1989) The plant cytoskeleton. Crit. Rev. Plant Sci. 8, 131 167
Shibaoka, H. (1974) Involvement of wall microtubules in gib- berellin promotion and kinetin inhibition of stem elongation. Plant Cell Physiol. 15, 255-263
Silk, W.K. (1984) Quantitative descriptions of development. Annu. Rev. Plant Physiol. 35, 479-518
Spurr, A.R. (1969) A low-viscosity resin embedding medium for electron microscopy. J. Ultrastruct. Res. 26, 31-34
Taiz, L. (1984) Plant cell expansion: regulation of cell wall mechani- cal properties. Annu. Rev. Plant Physiol. 35, 585-657
Takeda, K., Shibaoka, H. (1981a) Changes in microfibril arrange- ment on the inner surface of the epidermal cell walls in the epicotyl of Vigna angularis Ohwi et Ohashi during cell growth. Planta 151, 385 392
Takeda, K, Shibaoka, H. (1981b) Effects of gibberellin and col- chicine on microfibril arrangement in epidermal cell walls of Vigna angularis Ohwi et Ohashi epicotyls. Planta 151, 393 398