international conference on - biodiversity for sustainable
TRANSCRIPT

BIODIVERSITYInternational Conference on
PROCEEDINGS
May 22(Wed) - 24(Fri), 2019 Centara Grand & Bangkok Convention Centre at CentralWorldBangkok, Thailand
www.ibd2019exhibition.org
Sirindhorn thailandiensisAdamski & Malikul

Proceedings of International Conference on Biodiversity: IBD2019
on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
Organized by:
Ministry of Higher Education, Science, Research and Innovation
National Science and Technology Development Agency
Ministry of Natural Resources and Environment
Biodiversity-Based Economy Development Office
Supported by:
Fund for Sustainable Education (FUSE) – SAVITA FOUNDATION
Thailand Convention & Exhibition Bureau
In Cooperation with:
Office of Natural Resources and Environmental Policy and Planning
National Science Museum
National Research Council of Thailand
Department of National Park, Wildlife and Plant Conservation
Department of Marine and Coastal Resources
Royal Forest Department
The Zoological Park Organization under the Royal Patronage of His Majesty The King
King Mongkut’s University of Technology Thonburi
Center of Excellence on Biodiversity, Office of the Higher Education Commission
Sponsored by:
Electricity Generating Authority of Thailand
Toyota Motor Thailand Company Limited
Syngenta Crop Protection Limited

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand ii
Proceedings of
International Conference on Biodiversity: IBD2019
Biodiversity for Sustainable Bioeconomy on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
First Edition, September 2019
Copyright © 2019 by:
National Science and Technology Development Agency
Ministry of Higher Education, Science, Research and Innovation
Published by National Science and Technology Development Agency
Ministry of Higher Education, Science, Research and Innovation
111 Thailand Science Park, Phahonyothin Road,
Khlong Nueng, Khlong Luang, Pathum Thani 12120
TEL: +66 2644 8150 Ext. 81917
FAX: +66 2644 8106
Website: https://www.ibd2019exhibition.org
Designed by Program: Biodiversity Management and Sustainable use
Mrs. Rungsima Tantalakha
Mr. Chutdanaiphakorn Buraso
Ms. Sunthari Sueakham
Mrs. Sangdao Klangklai Mr. Pornpong Narasri
Academic committee Asst. Prof. Dr. Apiradee Saelim Prince of Songkla University, Pattani campus
Prof. Dr. Benchamas Silayoi Kasetsart University
Assoc. Prof. Dr. Boonsatien Boonsoong Kasetsart University
Asst. Prof. Dr.Chitchol Phalaraksh Chiang Mai University
Assoc. Prof. Dr. Decha Wiwatwittaya Kasetsart University
Dr. Weerachai Nanakorn Queen Sirikit Park
Assoc. Prof. Dr. Mullica Jaroensutasinee Walailak University
Assoc. Prof. Dr. Narumon Sangpradub Khon Kaen University
Asst. Prof. Dr. Noppadon Kitana Chulalongkorn University
Assoc. Prof. Philip D. Round Mahidol University
Asst. Prof. Dr. Pratueng Jintasakul Khorat Fossil Museum
Dr. Putarak Chomnuti Mae Fah Luang University
Prof. Dr. Saisamorn Lumyong Chiang Mai University
Mr. Sakanan Plathong Prince of Songkla University
Asst. Prof. Dr. Santi Watthana Suranaree University of Technology
Prof. Dr. Savitree Limtong Kasetsart University
Dr.Somying Thunhikorn Department of National Parks, Wildlife and Plant Conservation
Dr. Tanit Changthavorn Biodiversity-Based Economy Development Office
Prof. Dr. Wanchai De-Eknamkul Chulalongkorn University
Asst. Prof. Dr. Wangworn Sankamethawee Khon Kaen University
Prof. Dr. Warren Brockelman National Center for Genetic Engineering and Biotechnology
Asst. Prof. Dr. Witsanu Attavanich Kasetsart University
All right reserved. No part of this book may be reproduced, stored in retrieval system or transmitted in any form or by any mean: electronic, electrostatic, magnetic, tape, mechanical photocopying, recording or otherwise without permission from the publishers.

Proceedings of International Conference on Biodiversity: IBD2019 On 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand iii
CONTENT
Discover of giant chili in Nan Province, Thailand 1 - 4
Vichai Puripunyavanich, Narisra Suwan, Taweepong Na Nan
and Penjan Sutthanukul
Participatory approaches for Volkameria inermis conservation through herbal bioproducts at
the Sirinart Rajini Mangrove Ecosystem Learning Center, Prachuap Khiri Khan Province
5 - 8
Nudchanard Rukklin, Jedsada Kongkasurichay, Panisa Rodpai,
Nidanuch Sungpia and Komson Hongpadharakiree
Endophytic fungi from wild plants and their antifungal and plant-growth promoting properties 9 - 15
Sakuntala Siri-Udom, Nattamon Somjai and Wanwipa wongchaiya
Diversity of entomopathogenic fungi in protected forest in the Eastern of Thailand 16 - 20 Winanda Himaman, Panrada Jangsantear, Baramee Sakolruk,
Kittima Duengkae, Suchada Mongkolsamrit, Wasana Noisripoom,
Janet Jennifer Luangsa-Ard and Isarapong Vorapab
Diversity of mushrooms at Mu Ko Chang National Park, Trat Province 21 - 32
Baramee Sakolrak, Panrada Jangsantear, Winanda Himaman,
Tiplada Tongtapao, Chanjira Ayawong and Kittima Duengkae
Health benefit screening on 45 Thai human probiotics (TISTR strains): their safety and
immunomodulatory activity in macrophage function
33 - 44
Bhusita Wannissorn, Weerasak Taengphan, Prapaipat Klungsupya,
Supatjaree Ruengsomwong and Thanchanok Muangman
Records of potential antimicrobial activity of soil actinomycetes isolated from
community forest, Ban Khoklam Sang Aram, Udon Thani Province
41 - 47
Metinee Wasoontharawat and Panthong Kulsantiwong
Screening on anti-inflammatory property of 45 Thai human probiotics from
Biodiversity Research Centre of TISTR
48 - 54
Prapaipat Klungsupya, Weerasak Taengphan, Bhusita Wannissorn
Supatjaree Ruengsomwong and Thanchanok Muangman
Effect of water temperature on growth, survival and health status
of East Asian Bullfrog (Hoplobatrachus rugulosus) Larvae
55 - 61
Sornsawan Liamtong, Kanokporn Saenphet , Monruedee Chaiyapo and Supap Saenphet
Antioxidant activity and hair growth promoting activity of flavonoid extracts
from Phyllodium pulchellum and Uvaria rufa Blume on cultured mouse vibrissa hair follicles
62 - 66
Patcharida Penpakkula, Kanokporn Saenphetb and Supap Saenphet
Chemical composition and antibacterial activities of essential oils of Lavandula × hybrid
‘boysemberry ruffles’, L. pedunculata ‘princess’ and L. × hybrid ‘high five purple’
67 - 72
Suchawadee Insawang and Patcharee Pripdeevech
Discovery of plant antimicrobial peptides and laboratory scale production 73 - 76
Parichart Burns, Jutatape Watcharachaiyakup, Patchima Sithisarn,
Pimpilai Saengmanee, Vinitchan Ruanjaichon and Sonthichai Chanpreme
Diversity of indigenous vegetables in Puparn Royal Development Study Centre,
Sakon Nakhon Province, Thailand
77 - 85
Hathairat Chokthaweepanich, Kwankamol Tawaitakam and Noppadol Kaewkumsai
Phytochemical screening and antimicrobial activity of Indian almond leaf extracts 86 - 88
Ratthayaporn Thansuwan, Jirapron Jitturongarpron, Jittima Buatib,
Thanyaporn Tangjaroenchai and Noppadon Muangsue

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand iv
Study of chemical composition and antioxidant properties of Sangyod and
Tubtimchumpae rice bran oil
89 - 93
Pavinee Yampeng , Hathairat Rimkeeree and Supanida Winitchai
Checklist of bee genus Megachile Latreille, 1802 in Thailand (Hymenoptera:Megachilidae) 94 - 101
Nontawat Chatthanabun and Natapot Warrit
Diversity of caterpillars (Order Lepidoptera) in KhaoYai National Park,
Nakhon Ratchasima Province
102 - 115
Paradorn Dokchan, Nanthasak Pinkaew, Sunisa Sanguansub
and Sravut Klorvuttimontara
Proposal to reduce anthocyanin-deficient banana Musa siamensis to a M. rubra variety 116 - 121
Wandee Inta, Panida Kongsawadworakul, Unchera Viboonjun,
Paweena Chuenwarin, Paweena Traiperm and Sasivimon Chomchalow Swangpol
Species richness and distribution of marine spider crabs (Majoidea) in Thailand 122 - 128
Kamonchanok Wongissarakul, Pattanee Jantrarotai and Puntip Wisespongpand
The effectiveness of basic bait traps for collecting adult flies 129 - 135
Amornrat Ninon and Taeng On Prommi
An assessment of habitat connectivity for the endangered Malayan tapir in Thailand 136 - 142
Damisa Kaminsin and Naparat Suttidate
Satun UNESCO Global Geopark, the readiness for sustainable tourism 143 - 147
Suwimon Hengpatana, Phannipha Anuruksakornkul, Apinya Wanaset,
Taweesin Tangseng and Danuvas Suwanwong
Analysis of factors related to bamboo structure for preventing erosion and restoration of coastal
area on the Upper Gulf of Thailand
148 - 155
Patcharaporn Yaowasooth
Exploring the suitability map of wild banana (Musa serpentina Swangpol & Somana)
in Thailand using species distribution models with the limited occurrence data
156 - 162
Thanayut Changruenngam and Jantrararuk Tovaranonte
Stimulating the access to biodiversity and technologies to combat climate change 163 - 173
Fabrice Mattei

International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 1
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 1 - 4
Discover of giant chili in Nan Province, Thailand
Vichai Puripunyavanich1*, Narisra Suwan2, Taweepong Na Nan2 and Penjan Sutthanukul3
1Thailand Institute of Nuclear Technology, Ongkharak District, Nakhon Nayok, Thailand 2Nan Agricultural Research and Development Center, Muang District, Nan, Thailand
3Sukhothai Horticultural Research Center, Si Satchanalai District, Sukhothai, Thailand
*Corresponding author e-mail: [email protected]
Abstract: From the research project of anthracnose resistance breeding in chili, the native chili in northern and
north eastern region of Thailand were explored for its diversities of native wildtype. The result showed that most
of the chili cultivars that Thai farmer planting after rice harvesting season were commercial F1 hybrid. However,
some farmers still planted native wild types chilies. The objective of Thai chili farmer was to trade the product to
the middle merchants or directly sale to seasoning factory. Diversities of Thai native chili was rather narrow genetic
base because Thai farmers select only dark red colour and large size fruit that the factory is needed and discarded
the other characters. By incident, giant chili cultivar was noticed in remote valley of Tha Wangpha District, Nan
Province, Thailand. Some giant chili fruits are up to 25-30 cm in length and 3-5 cm in width with dark red colour.
This giant chili was an open pollinated cultivar which only few farmers in that area planted and collected it
continuously for more than ten years. The chili specialist at the Department of Agriculture also has never found
the giant chili at this size before. However, after the researcher group requested some seeds from a farmer and
brought to planted at Sukhothai Horticultural Research Center, most of the giant chili were died or became dwarf
plants. It is indicated that the giant chili cultivar may have highly specific to a certain area, thus, further
investigation and research study on the giant chili are needed.
Keywords: Nan, chili, fruit size, Capsicum annuum
Introduction
Chili pepper or Prik Chifah in Thai is a Thai local vegetable that become widely known among consumers
as it has hot taste and color right to the recipes. Chili is considered as one of the most important commercial spice
crops and is widely used as universal spice, named as wonder spice. Different varieties are cultivated for various
uses like vegetable, pickles, spice and condiments. Chili pepper has botanically known as Capsicum annuum L.
and belongs to the genus Capsicum, under the Solanaceae family which closed relatives used as vegetables: potato,
tomato, and eggplant. Chili originated in Latin America and was introduced to Asia by Spain and Portugal around
three hundred years ago. Most cultivated chili belonging to one of two major species groups.
1. Capsicum annuum includes chili pepper, sweet peppers and the pimiento (pimento), or Spanish pepper.
Sometime they have grown principally for use in commercial chili sauces.
2. Capsicum frutescens includes the "hot" chili and cayenne peppers; these have extreme pungency.
C. frutescens species may contain as much as 1 percent capsaicin and is one of the most important ingredients in
many different cuisines throughout the world as it adds pungency taste, flavor and color to the dishes (Ministry of
agriculture of India, 2009). The world production was reported by FAO in 2010 approximately 26.8 and 2.8 ton
for fresh and dried fruit. Usually, chili pepper is an annual plant but it can grow as a woody biennial in warm
climates. Thai farmers often started planting it in the nursery as seedling and then transplanted to the production
field. Chili pepper is a dicotyledonous plant. Flowers are insect pollinated and harvest green or red fruit. Fruit
color of chili peppers changes from mature green to full red color or yellow depending on its varieties. Their fruit
varies substantially in shape, pericarp thickness, color and pungency. The pungency of chili peppers is caused by
compound capsaicin. Chili peppers are range from 2,000 to 25,000 unit capsaicin whereas hot chilies are range
between 60,000 to 80,000 unit capsaicin. In Thailand, fresh edible chili peppers are usually harvested when red
while fresh chili peppers in commercial chili sauces are only harvested in red or dark red.
In Thailand, farmer usually grow chili pepper in November to April after harvesting rice. The area for
chili production was about 2-4 rais / family. The chili cultivar was determinate by the middle merchants or dealer
which was deal with the factory. Company F1 hybrid of Mae-ping 8000 was the introduced cultivar, following by
Kweawmanee 26, Sunkampang, Xian and Yoksiam. In some case, if there were contacted with collector, the farmer
may harvest only ripe chilies. Chili peppers fruit was usually sent to factory within 24 hours after harvesting. Thick
flesh and dark red color chili pepper were highly needed for the commercial chili sauces factory. The important
problem for the production is the pest infected by both insects and diseases. Major diseases of chili are anthracnose,
bacterial spot, and Phytophthora blight while major insects are broad mite, red spider mite, whitefly, etc. Most of
the Thai farmer buy chili seeds from a company. Usually F1 hybrid seeds are served for the farmers from the
company through a dealer. Very few Thai farmers developed open pollinated seed from natural germplasm by

International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 2
themselves. On the beginning of this project, researchers tried to discover the anthracnose resistance in chilli which
may be hidden in natural resource conditions in north and north eastern region of Thailand. However, sometimes
the result did not correspond to the project target that had been set as some hybrid chili-pepper with vigour
characters hidden in the deep remote area of the country could be find.
Materials and Methods
1. Exploration of chili peppers samples were done in north and north eastern region of Thailand. Samples
of leaves, flowers, seeds and fruits were collected, observed, studied and took photographs in some areas at
Yasothon and Mukdahan, Chiang Mai, Sakhon Nakhon, Nan, Phrae, Roi Ed, Khon Kaen, Ubon Ratchathani,
Phitsanulok, Sukhothai, etc.
2. Around 40 sample of farmers were interviewed
- Farmer bought the seed from whom?
- Do farmers use native chili varieties?
- Are there contracts between farmers and company?
- Pest control process
- Period of planting
3. Morphology and taxonomy studies at Sukhothai Horticultural Research Center
4. Field plot test conducts at Sukhothai Horticultural Research Center for some interested cultivars.
Result & Discussion
From field exploration around the north and north eastern Thailand, it is founded that most of chili
cultivars which Thai farmer planting after rice field season were commercial F1 hybrid. Maeping 8000, Keaw-
manee 26 and Yok-siam which generally introduced by the company through the middle man in both north and
north eastern regions. Some farmers had contracted with the company whereas some farmers were no contracted.
Those contracted farmers are received price guarantee from the company. However, some farmers did not accept
the guarantee price and sale their products by themselves through independent middle merchants or directly to
chili sauces factory somewhere such as at Sukhothai, Sisaket, Ubon Ratchathani Province. There was setting up
the community enterprise or agricultural co-operative units but most of them were not strong. The price guarantee
from company was attracted the farmers to make decision to plant chili after their rice field season. Although the
objective of Thai chili farmer was mostly trade to the middle merchants or directly sent to seasoning factory. Only
thick flesh and dark red color chili pepper were needed for the commercial chili sauces factory and some products
which the factory did not accept, the farmer would sale it in the local market nearby their village. At that time, the
exploration of anthracnose disease indicated that it was widely spread in both chili pepper (Capsicum annuum)
and hot chili (Capsicum frutescens). But the farmers use only chemical disease control such as benomyl, mancozeb,
carbendazim or prochloraz to control this disease, so there was no anthracnose resistance in chili cultivars or native
cultivars in Thailand.
Although most of Thai farmers planted F1 hybrid chili which bought from the company, however, few of
them still planted native wild types. Diversities of Thai native chili was rather narrow genetic base because Thai
farmers select only dark red colour and large size fruit which factory needed and discard the other characters. The
exploration in Nan Province were conducted deep to the remote valley area of Tumbol Jomphra, Tha Wangpha
District. Some farmers planted the commercial F1 hybrid chili peppers cultivar under company contract. Some
farmers planted the commercial F1 hybrid chili peppers paralleled with their native inbred lines. All of their chili
pepper cultivars are open pollinated cultivars which different from F1 hybrid. The quality and quantity of the
cultivars look similar to the company F1 hybrid. It did not name after their native cultivars were breed, therefore,
all those cultivars were called by coding with Nan 1001, 1002, 1003, 1004…etc. Usually, the farmers sold only F1
hybrid chili pepper products to the company under contract through a dealer. But their native chili pepper products
were sold to freelance middle man or sold in local market. There was no farmer sold their chili pepper fruits
directly to chili sauce factory because there was no any factory nearby. In addition, the native ‘Sornthong Nan’
cultivar was investigated. Mr. Sawai Bhrammanee was the cultivar breeder which was not under contract from any
company. He explained ‘Sornthong Nan’ cultivar has very thick flesh and sweet. There was freelance dealer sold
his products to Nam Prik Num (Northern Thai Green Chilli Dip) factories because ‘Sornthong Nan’ has very thick
flesh and sweet fruit. By incident, giant chili cultivar was noticed. The land owner and the giant chili pepper
breeder is Mr. Kamsarn Sutthisan. He told the research group that the breeding of giant chili pepper was done for
more than ten years from native varieties crossed over with commercial varieties and did selection year by year
until receiving the right chili pepper line. However, this genetical line is still not so stable, there possesses a range
from giant chili pepper size to the normal chili pepper size in the same field. The giant chili pepper was sold in
the local market. There was no middle man from any company bought this giant chili pepper because it was not
under contract with any company. Some giant chili fruits are up to 25-30 cm in length and 3-5 cm in width with
dark red colour and thick flesh fruits. This giant chili was an open pollinated cultivar. After receive giant chili

Proceedings of International Conference on Biodiversity: IBD2019 On 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 3
pepper samples from the owner, it was brought to Sukhothai Horticulture Research Centre. The chili specialist of
the Department of Agriculture said that the chili with giant size has never been found before. Although the giant
chili has dark red colour and thick flesh fruit but it produces rather few seeds of only 40-50 seeds/fruit. The giant
chili pepper seed were planted in Sukhothai Horticultural Research Centre whereas nearby many chili seasoning
factories. Most of them were died or became dwarf plants and got a few normal size fruits. It is indicated that this
giant chili cultivar was highly specific to area base Nan Province or may be highly specific to area only in Tumbol
Jomphra, Tha Wangpha District.
To avoid patent conflict and to breed a new cultivar base on this giant chili pepper, mutation breeding
program by gamma irradiation was setting up. The research hope to discover a giant chili pepper that able to adapt
and survive at Sukhothai Horticulture Research Centre where there are chili seasoning factories in Sukhothai
Province, therefore, it is expected that the breeding of a mutant line will be accomplished to support the Sukhothai
area. Large size, dark red colour and thick flesh fruit characters will be selected in the research program for the
near future research.
Conclusion
Although the first target of anthracnose resistance in native chili exploration was not discovered in north
and north eastern region of Thailand. Most of chili cultivars were commercial F1 hybrid from a company. From
the exploration in Nan Province, some of farmers planted native wild types or inbred lines that they did breeding
it themselves. Diversities of Thai native chili was rather narrow genetic base. There were some open pollinated
chili pepper cultivars developed by several farmers. Quality and Quantity of their chili peppers fruit were similar
to F1 hybrid of any company. ‘Sornthong Nan’ and Giant chili pepper cultivar were discovered in Nan Province.
The native ‘Sornthong Nan’ is suitable to make Nam Prik Num (Northern Thai Green Chilli Dip). Some giant chili
pepper fruits were very big up to 25-30 cm in length, dark red colour and thick flesh fruits are suited for chili sauce
factory requirement. The giant chili pepper and native ‘Sornthong Nan’ were also open pollinated cultivars.
However, most of the giant chili pepper were died or became dwarf plants after they were planted in Sukhothai
Province.
Acknowledgement
We would like to thank AVRDC (Asian Vegetable Research and Development Centre), Kasetsart
University, Kamphaengsaen Campus, Nakhon Pathom) for their valuable suggestions and comments. We also
thank all authorities and their staff in every agricultural station and especially researchers from Department of
Agriculture for their technical supports.
References
Department of agriculture and co-operation, Ministry of agriculture of India. 2009. Postharvest profile of chilli. p.80
Kraikruan W., Supkweaw U., Sachati S. and Suthanigool P. 2014. Supply chain of processing chili production in
Sukhothai Province. The 53th Kasetsart University Annual Conference. Feb. 3-6, 2015. p. 279-286 (in Thai script).
Mahasuk P., Chinthaisong J. and Mongkolporn O. 2013. Differential resistances to anthracnose in Capsicum baccatum as responding to two Colletotrichum pathotypes and inoculation methods. Breed Science. 2013 Sep; 63(3): 333 – 338.
Mongkolporn O., Montri P., Supakaew T. and Taylor P.W.J. 2010. Differential reactions on mature green and
ripe chili fruit infected by three Colletotrichum spp. Plant Disease Journal. 94: 306–310.
Puripunyavanich V., Phadvibulya V. and Boonsirichai K. 2007. Effect of Gamma Irradiation on Chili Mutation.
The 10th Conference on Nuclear Science and Technology. Bangkok International Trade & Exhibition
Centre. June 16-17, 2007.
Ratanacherdchai K., Wang H. K., Lin F.C. and Soytong K. 2007. RAPD analysis of Colletotrichum species causing
chilli anthracnose disease in Thailand. Journal of Agricultural Technology. 2007; 3:211–219.
Than P.P., Prihastuti, H., Phoulivong S., Taylor P.W.J. and Hyde K.D. 2008. Chilli anthracnose disease caused
by Colletotrichum species. Journal of Zhejiang University Science B. 2008 Oct; 9(10): 764–778.

International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 4
Figure 1. Nan 1008, Nan 1009 and Nan 1015 chili pepper lines.
Figure 2. Nan 1014, Nan 1016 and Nan 1017 chili pepper lines.
Figure 3. Sorn Thing Nan, Xian F1 hybrid (Jia Tai) and Sunkampang F1 hybrid (Sorn Dang) chili pepper lines.
Figure 4. Giant chili pepper line.
Figure 5. Giant chili pepper line breeder and ‘Sorn Thong Nan’ breeder.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 5
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 5 - 8
Participatory approaches for Volkameria inermis conservation through herbal
bioproducts at the Sirinart Rajini Mangrove Ecosystem Learning Center,
Prachuap Khiri Khan Province
Nudchanard Rukklin*, Jedsada Kongkasurichay, Panisa Rodpai, Nidanuch Sungpia,
and Komson Hongpadharakiree
Sirinart Rajini Mangrove Ecosystem Learning Center, Pranburi District, Prachuab Khiri Khan, Thailand
*Corresponding author e-mail: [email protected]
Abstract: The study aimed to examine participatory approach using commercial values of herbal bioproducts and
local attitudes to expand the plant habitats inside the Sirinart Rajini Mangrove Ecosystem Learning Center at Pak
Nam Pran Sub-district. Volkameria inermis was chosen among other mangrove plants for its values. Forty
respondents, twenty of them whose incomes were promoted from Volkameria inermis herbal products, while
another twenty respondents lived adjacent the center. Their attitudes were collected by questionnaire surveys and
analyzed by descriptive statistical analysis including percentage, mean, and One-way ANOVA with a significance
level (α) of 0.05. Incomes increased by added values of mangrove herbal products made from the plant were
analyzed. The study showed that respondents whose incomes were promoted through the herbal products had high
level of awareness at 61.77% on the plant value and the mangrove forest conservation while another group of
respondents showed their moderate awareness with 44.28%. The results showed significant influence (p-value =
0.024) of knowledge on the plant benefits and the higher incomes earned from the herbal products on the attitude
towards a plant and mangrove natural resource conservation. A total income obtained from selling the plant
products of center during January 2016 to September 2018 was totally 226,990 baht. Among the products, the
incomes from selling the soap and hot balm made from the plant were 222,535 baht and 4,455 baht respectively,
made by total fresh leaves of plant 109.34 kilograms or 2,076 baht per kilogram wet weights. It can be concluded
that a community income promotion using local mangrove forest resources was an approach to the community’s
values and awareness on the mangrove forest conservation, leading to community participation in tree planting,
caring, and restoring of targeted plant species as well as using the natural resources in sustainable ways.
Keywords: Participatory approach, conservation, Volkameria inermis, herbal bioproducts
Introduction
Mangrove forest conservation with participatory policy under the Sirinart Rajini Mangrove Ecosystem
Learning Center, at the Klong Kao – Klong Koy National Reserved Forest, Pak Nam Pran Sub-district, Pran Buri
District in Prachuap Khiri Khan Province has been ecological restored since 1981 with the area of 786 rai (126
hectares) (Paphavasit et al., 2014). The policy on community participation on the mangrove forest was applied to
the mangrove management following His Majesty King Rama IX’s speeches "The locals benefit from the
mangrove forest, so they must help to conserve it" on 16 November 2002 by His Majesty King Rama IX delivered
during the Public Company Limited’s 1 Million-Rai Reforestation Project in Commemoration of His Majesty the
King Rama IX’s 50th Anniversary of His Accession to the Throne. Following the Speech, becoming a policy and
actions, the restored mangrove forest tracts of Forest plantation target 29 and forest plantation target 29/3 provided
the forest ecosystem benefits with the building the participation of the community in the conservation of mangrove
planting to sustainability among communities at Pak Nam Pran Sub-district of Pran Buri District. To achieve the
target on community participation on the mangrove forest management, there were four strategic issues were
included to the forest operations which were knowledge, community and society, environment, and ecotourism.
Social entrepreneurship is an effective tool for promoting participatory resource-based management. Network of
villagers who earned benefits from herbal plants of mangrove forest was established under the Center as a group
of mangrove forest herbal bioproducts. The social entrepreneurship is known as a process of creating value by
combining resources in new concepts of social networking to seek for opportunities regarding social values and
needs. Management of herbal bioproduct values to promote more socio-incomes among the members and
strengthen the mangrove forest was anchored as foundation of capacity building conservation expanding around
the Center. The process of creating value of mangrove plant bioproducts regarding local needs such as hot balm
oils for the development of product values of mangrove plants and encourage the sustainable mangrove forest
management under the Center. Promotion of mangrove forest services for local communities in particular, various
mangrove plant species growing in the center’s area is for occupation promotion and socio-incomes of group
members. There are more than 20 plant species that could be utilized as herbal bioproducts with simple production
process and meet to the market needs of local consumption. Their medicinal characteristics were reported and
utilized by local wisdoms and scientific research studies. For example, boiled water from bark of Rhizophora

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 6
apiculata can heal diarrhea, nausea, vomiting, fever, or chronic wounds clean (Faculty of Science Chulalongkorn
University as cited in Paphavasit et al., 2014) as well as the boil water or blended bark of Xylocarpus granatum
can cure wounds and bruise. The boiled water of green leaves of Samma Nga or Volkameria inermis is used to
clean wounds, treat skin diseases, and bathing to reduce rash (Kung Krabaen Bay Royal Development Study
Center. 2006). Among those plant speices such as Rhizophora apiculata, Rhizophora mucronata, Xylocarpus
moluccensis, Xylocarpus granatum, Thespesia populnea, Avicenniaalba, Avicennia marina, Bruguiera cylindrical,
Acanthus ilicifolius, Volkameria inermis, and etc, the local plant named Samma Nga (Volkameria inermis) was
selected for participatory management model using commercial values to expand the area of plantation in the
Center area.
The plant contains flavone derivatives including hispidulin, acacetin, and diosmetin. The hispidulin is
found in Volkameria inermis reported as a potential medicinal plant species by Faculty of Pharmacy, Mahidol
University. Potential therapeutic role of hispidulin extracted from the plant leaves is volatile substance that
decongestive bronchial congestion. More substances of acacetin and diosmetin were found in the plant leaves and
exhibits several pharmacological and medicinal properties especially their roles in anti-inflammation of the skin,
rash, bruise and sprain.
From highly potential therapeutic roles of chemical substances found in Volkameria inermis, but in very
small quantity of leaves available at the Center, the plant will be promoted through commercial values and local
attitudes to expand the plant habitats inside the Center. Social mechanisms especially social entrepreneurship using
the potential of herbal bioproducts to generate the diversified socio-incomes and community participation in the
programs of Volkameria inermis planting is assessed. Assessment of effectiveness of conservation practices that
related to Sirinart Rajini Center programs including knowledge transfer and occupation promotion using benefits
from herbal bioproducts of Volkameria inermis. The results of assessment would be the management model for
sustainable forest management using participatory approach together with commercial values development of
herbal bioproducts (Figure. 1).
Materials and Methods
In order to find the participatory approaches using the social entrepreneurship operating the herbal
bioproduct values from Volkameria inermis available at the Sirinart Rajini Mangrove Ecosystem Learning Center,
Pak Nam Pran Sub-district, field survey was conducted using semi-structure questionnaires. The contents of
questionnaires are divided into three data sets.
A. Demographic information the 40 respondents.
B. Benefits of Volkameria inermis to promote the mangrove forest conservation.
C. Participatory approaches for Volkameria inermis planting in the Center area to promote the
mangrove forest conservation.
Attitudes survey, totally forty respondents was conducted by selecting the twenty respondents from
communities in the center and another twenty respondents from adjacent communities. The collected data sets
were analyzed using the Statistical Package for Social Sciences program for averages percentage and mean values.
One-way ANOVA at a significance level (α) of 0.05 was chosen to reveal the significant differences of two groups
of respondents on the studied data sets. Moreover, using the literature reviews and experts opinions for qualitative
data analysis, the records of socio-incomes obtained from the benefits of herbal bioproducts made from Volkameria
inermis in 2016 to 2018 was analyzed.
Result & Discussion
The forty respondents were in the middle ages older than 51 years old. The twenty respondents from the
observed group who earned benefits form the Volkameria inemis herbal bioproducts were mainly female at 65%
at 51-60 years old. Their education information showed that 30% of this group graduated in an elementary school
or lower and 50% of the respondents were graduated in high school. For the source of main incomes, there were
30% of respondents earning their incomes from fishery, and 30% of those had their total incomes around 5,000-
8,000 baht per month.
The other twenty respondents who lived adjacent mangrove forest of Sirinart Rajini Mangrove Ecosystem
Learning Center were found females 60% and age of 60 years old of respondents were found around 30%. There
were about 30% of respondents graduated in high school, and among the respondents they were found being a
merchant. There were 40% of them earned incomes 5,000-8,000 baht per month and another 60% of them earned
incomes more than 15,000 baht.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 7
The results of interviews of all respondents showed that their attitudes on the benefits of herbal plant and
importance of mangrove forest, the respondents whose incomes were earned from the herbal bioproducts were
found at 61.77% while another group of respondents showed their moderate awareness with 44.28%.
The results showed significant attitudes (p-value = 0.024) on the plant benefits and the higher incomes
earned from the herbal bioproducts on the attitude towards a plant and mangrove natural resource conservation.
The demographic factors including gender, age, education, occupation and incomes per month did not significantly
affect (p-value = 0.534, 0.824, 0.636, 0.752, and 0.983, respectively) to the attitudes a plant and mangrove natural
resource conservation.
In addition, the results showed the respondents whose their incomes were from herbal bioproducts needed
training programs for the knowledge related to the benefits of Volkameria inermis and should developed other
herbal bioproducts to provide alternative choices for consumers. The Volkameria inermis may be reduced if
harvested continuously, so more areas inside the Center should be for Volkameria inermis plantation as well as the
determination of timing of restoring the Volkameria inermis back to the area.
Socio-incomes obtained from the benefits of herbal bioproducts made from Volkameria inermis January
2016 to September 2018 revealed that the plant provided the respondents more incomes. The center with the group
of mangrove forest herbal bioproducts produced the herbal bioproducts earned more incomes from selling the
herbal bioproducts totally 226,990 baht. Among the different types of bioproducts, the incomes from selling the
soap made from the plant were 31,975, 87,600 and 107,145 baht in 2016, 2017 and 2018 respectively. The soap
made from total fresh leaves of Volkameria inermis 109.34 kilograms or 2,076 baht per kilogram wet weights.
Conclusion
The results showed the significant attitudes on the benefits of herbal plant, Volkameria inermis
conservation through incomes promotion of selected stakeholders from the community at Pak Nam Pran Sub-
district, Pran Buri District in Prachuap Khiri Khan Province and from the Center adjacent communities. The
respondents whose incomes were promoted through the herbal bioproducts had high level of awareness at 61.77%
on the plant value and the mangrove forest conservation while another group of respondents showed their moderate
awareness with 44.28%. The results showed significant attitudes (p-value = 0.024) on the plant benefits and the
higher incomes earned from the herbal bioproducts on the attitude towards a plant and mangrove natural resource
conservation. Moreover, they needed knowledge related to the benefits of Volkameria inermis and other herbal
bioproducts to provide alternative choices for consumers. To increase the numbers of this herbal plant, more areas
inside the Center should be provided for plantation as well as the determination of timing of restoring the
Volkameria inermis back to the area.
The occupation promotion to community was an approach to the community’s values and awareness on
the mangrove forest conservation, leading to community participation in planting, caring, and restoring of targeted
plant species as well as generating the incomes and using the natural resources in sustainable ways.
Acknowledgement
The authors gratefully thank the respondents and officers of the Sirinart Rajini Mangrove Ecosystem
Learning Center for their information. Also, thank you very much to the PTT Public Company Limited for
supporting the research budget. Finally, we would like to thank Dr. Kallaya Suntornvongsagul, Environmental
Research Institute of Chulalongkorn University for useful comments and suggestions on the English language and
structure of our manuscript.
References
Faculty of Pharmacy Mahidol University. 2017. Herbal Plants: Volkameria inermis. Retrieved from
https://pharmacy.mahidol.ac.th/siri/index.php?page=search_detail&medicinal_id=38.
Forestry Kung Krabaen Bay Royal Development Study Center. 2006. Medicinal plants of beach forests and
mangrove forests in the Kung Krabaen Bay Royal Development Study Center area. Chanthaburi.
Paphavasit, N., Siriboon, S., Jaiperm, J. and Mookui, P. 2014. Sirinath Rajini Mangrove Ecosystem Learning
Center. From mangrove plantation to mangrove forest enhancing human development (1ed.).
Bangkok: PTT co., Ltd and Department of Science, Chulalongkorn University. Bangkok.
Paphavasit, N., Chawasiri, W., and Theerathanathor, V. 2012. Herbal plants in the mangrove forest Thung Tase
in Trang Province. Yves Rocher (France) and Yves Rocher (Thailand) Ltd. And Department of
Science, Chulalongkorn University. Pra Suk Chai Printing Part., Ltd. Bangkok.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 8
Figure 1. Research framework diagram.
Sustainable Forest Management using Participatory Forest Conservation
Community Based Participatory Approach
Attitudes on Benefits of Forest
Conservation
Benefits for Socio-economics:
-Herbal medicines
-Food
Sustainable Management
Tools

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 9
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 9 - 15
Endophytic fungi from wild plants and their antifungal and
plant-growth promoting properties
Sakuntala Siri-Udom*, Nattamon Somjai and Wanwipa wongchaiya Department of Biology, Faculty of Science, Udon Thani Rajabhat University, Muang District,
Udon Thani, Thailand
*Corresponding author e-mail: [email protected]
Abstract: A total of 118 endophytic fungi were isolated from 21 wild plants in northeastern of Thailand. Most
endophyte were isolated from Maesa ramentacea (Roxb.) A. DC. (12.7%), followed by Baeckea frutescens
(11.9%) and Anneslea fragrans Wall. (6.8%), respectively. Endophytic fungi were more prevalent in the leaves
(59.3%) than the branches/stems and the flowers. Isolate Gu02 and Gu03 from flowers of Gluta usitata (Wall.)
Ding Hou exhibited board range of antifungal activity above 50% growth inhibition by dual culture technique
toward to Colletotrichum gloeosporioides, Phellinus noxius and Rigidoporus microporus. Isolate Gu03 inhibited
mycelial growth of the pathogenic fungi, P. noxius and R. microporus causing root rot disease of rubber tree with
74.1% and 88.1% inhibition, respectively. Isolate Gu03 produced plant growth hormone, indole acetic acid (IAA)
at the concentration of 331.5 µg/ml. Volatile metabolite-producing endophytic fungus, isolate Cs05 was isolated
from leaf tissues of Cycas siamensis Miq. The volatile organic compounds (VOCs) of isolate Cs05 were active
against P. noxius and R. microporus with 100% growth inhibition.
Keywords: Antifungal activity, endophytic fungi, indole acetic acid, wild plant
Introduction
Endophytic fungi live in symbiosis with plant tissues and play an important role in plant growth (Young-
Hyun et al., 2012). They have been identified in nearly 300,000 plant species (Strobel and Daisy, 2003) and dwell
in root, stem, leaf, flower, fruit and seed (Yu et al., 2018). Most endophytic fungi are members of the Ascomycota,
with only a few reports of basidiomycetous endophytes, these often being orchid mycorrhizas (Rungjindamai et
al., 2008). Natural select the evolution of beneficial endophyte strains and several endophytes are found to produce
bioactive compounds that protect plant from insect pests and pathogens (Saikkonen et al., 2004). They play an
important role in plant defense including the function as growth promoter and enable the host survival under
extreme conditions (Rosa et al., 2012). Although, chemical pesticides were used to protect plant disease, but
pesticide use may lead to environmental pollution and might threaten human health (Yu et al., 2018). Recently,
the development and spread of drug-resistant pathogens are still a global problem and there is a need to search for
new active agents with antimicrobial activity (Espinel et al., 2001). Thus, endophytic fungi are source of novel or
bioactive metabolites for pharmacological and agricultural applications (Idris et al., 2013; Deshmukh et al., 2018).
Natural products from fungal endophyte showed antagonistic activity to inhibit several pathogenetic
organism such as bacteria, fungi, viruses and protozoans. Furthermore, they could be plant growth regulators such
as indole acetic acid (IAA), one of the most physiologically active auxins. Fungi produce a wide variety of plant
hormone such as gibberellins (GAs), abscisic acid (ABA), and auxin (Young-Hyun et al., 2012; Khan et al., 2017).
IAA is a product of L-tryptophan metabolism by various microorganism including Plant growth promoting
rhizobacteria (PGPR) (Ahmad et al., 2005). The volatile metabolite-producing endophytic fungi have not been
commonly reported. But some endophytic fungi that belong to the families Diaporthaceae, Hypocreaceae,
Stachybotryaceae and Xylariaceae of the phylum Ascomycota are notable for their capacity to form volatile
metabolites with antimicrobial activity (Stinson et al., 2003; Banerjee et al., 2010; Suwannarach et al., 2013; Siri-
Udom et al., 2015). The mixture of fungal volatile compounds that have been identified were aldehydes, alcohols,
benzene derivatives, cyclohexanes, ketones, hydrocabons, heterocycles, phenol, thioalcohols, thioesters and their
derivatives (Mercier et al., 2007; Morath et al., 2012). In agriculture, fungal VOCs with antimicrobial activity may
be used as a mycofumigant to control plant disease.
In this study, we aimed to evaluate the diversity of endophytic fungi that live in association with wild
plants. Furthermore, to assessed the potential of the effective fungi that capable of producing bioactive agents,
plant growth promoting metabolites and volatile metabolites against fungal pathogens.
Materials and Methods
Preparation and surface sterilization of plant materials
Twenty-one healthy wild plant species were used as source for isolation of endophytic fungi. Eight plants
were collected from The Royal-initiated Lam Huai Bong Forest Area Development Project (16°54′23″N, 102°
24′47″E), located in, Nong Bua Lam Phu Province. Six plants were collected in Dong Kheng community forestry,

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 10
Nong Bua Lam Phu Province (16°56′49″N, 102°34′31″E). Seven plants were collected from Ban Thai Seri
community forestry, Bueng Kan Province (18°00 ′54″N, 103°57′49″E). Plant materials were cut randomly into
small segments (leaf and flower, 5 mm x 5 mm; stem/branch 5 mm long). All segments were sterilized by soaking
in 75% ethanol for 30 s, 2% sodium hypochlorite for 3 min, and 95% ethanol for 30s under a laminar flow hood
(Suwannarach et al., 2010).
Isolation of endophytic fungi
Each plant materials were placed on potato dextrose agar (PDA) containing Rose bengal (0.033 g/l) and
chloramphenicol (50 mg/l). Petri dishes were sealed with Parafilm® M (Bemis company, Inc., USA) and incubated
at room temperature (25±2 C) for 2 weeks. Emerging fungi were transferred to fresh PDA plates. The medium
was also be used for subculture and stock culture. Identification of endophytic fungi was carried out on the basis
of morphological characteristics (Huang et al., 2008). The fungal isolates were grown on PDA and incubated at
room temperature to observe morphological characteristic such as color, shape and size of spores. Spore production
of endophytic fungi was studied on different media, such as malt agar (MA), PDA and water agar (WA).
Antagonism against fungal pathogens by dual culture method
The antagonistic analysis was observed from the interaction between fungal endophytes and the pathogens
using dual culture technique. An agar plug (6 mm diameter) 4-day-old fungal endophytes growing on PDA was
inoculated on PDA part of the petri dish, then an agar plug of fungal pathogens was inoculated opposite side of
the Petri dish. The control plates were inoculated with either pathogens. All Petri dishes were wrapped with
Parafilm® M and incubated at room temperature (25±2 °C) for 7 days. The percentage of inhibition of fungal
growth after the dual culture test was calculated with the following equation: [(R1- R2)×100] ÷ R1 , where R 1
was the average colony radius of each tested fungi measured in the control plates (without the tested fungi), and R
2 was the average colony radius that calculated from the tested plates.
Antimicrobial assay of VOCs
The parallel-growth isolation technique was adapted for the antagonism test of fungal VOCs. An agar
plug of volatile-producing endophytic fungi was inoculated on the PDA part of a two-compartment Petri dish, and
allowed to grow at room temperature (25±2 °C) for 2-4 days. Then, an agar plug (6-mm diameter) 4-day-old fungal
pathogens growing on PDA was inoculated on PDA on the opposite side of the Petri dish. All Petri dishes were
wrapped with parafilm® M and incubated at room temperature (25±2°C) for 3-7 days. The percent inhibition of
fungal growth after the dual culture test was calculated. The viability of the test fungal pathogens was observed
by transferring them from the test plates and re-growing in fresh PDA (Strobel et al. 2001).
Indole acetic acid (IAA) production
All isolates of endophytic fungi were inoculated in 5 ml potato dextrose broth (PDB) with L-tryptophan
(2 mg/ml) and incubated in the dark at room temperature (25±2 °C) with shaking at 150 rpm on a shaker for 5
days. The broth cultures were filtrated by two layers of gauze cloth to separate the broth cultures and mycelia. A
modified method described by Ahmad et al., 2005 was used for screening IAA production. All filtrates (1 ml) were
mixed with 2 ml of Salkowski’s reagent (1 ml of 0.5 M FeCl3; 50 ml of 35% Perchloric acid (HClO4)) and
incubated in the dark for 30 min. The development of pink color indicated IAA production and the absorbance at
530 nm was measured. The level of IAA production was estimated by standard IAA graph.
Result & Discussion
Isolation of endophytic fungi
A total of 118 endophytic fungi were isolated from 21 plant species, the most of them were isolated from
Maesa ramentacea (Roxb.) A. DC. (12.7%), followed by Baeckea frutescens (11.9%) and Anneslea fragrans Wall.
(6.8%), respectively (Table 1). Most of them colonized in leaf (59.3%) and only 3 isolates colonized in flower of
plant sample (Figure 1. and Table 1). Furthermore, 6 volatile producing endophytic fungi were isolated from 5
wild plants, including M. ramentacea. Isolate Cs05 did not produce spore and belonged to mycelia sterilia group.
Endophytic fungi are presented in most of plant section, especially healthy leaf tissue (Kharwar et al., 2011;
Karunai and Balagengatharathilagam, 2014; Yu et al., 2018) according to this study, most endophytic fungi were
more prevalent in the leaves than the branches/stems and the flower. Although, Zheng et al. (2016) proposed that
the diversity of endophytic fungi is generally significantly higher in the stems than in the leaves.
Endophytic population varies from plants to plants and species to species (Nair and Padmavathy, 2014).
However, the endophytes population was affected by the environmental conditions under which the host is growing
and the endophyte profile may be is more diversified in tropical areas (Karunai and Balagengatharathilagam,
2014). The isolate numbers of endophytic fungi are closely correlated with the sampling range of the plant age,
such as increasing the collecting plant specimens of different ages, to enable the isolation of additional fungal
endophytes (Yu et al., 2018).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 11
Wild plant in family Myrsinaceae can potentially be explored for estrogenic activity and for sources of
phytoestrogens (Jamal et al., 2012), genus Maesa was reported the fungal endophyte in genus Diaporthe lived in
fruit (Gomes et al., 2013). Coniochaeta ligniaria, an endophytic fungus with antifungal activity was isolated from
leaves of Baeckea frutescens (family Myrtaceae) obtained from the Phu Luang wildlife sanctuary in Thailand
(Kokaew et al., 2011). Fungal endophyte can live in various wild plant tissue, genus Fusarium was the most
common fungal isolate from root of wild banana (Musa acuminate, family Musaceae), which is native to Southeast
Asia and is an ancestor of the edible banana (Zakaria et al., 2016). Several endophytic fungi, such as Colletotrichum
gloeosporioide, Phomopsis sp., Phyllosticta capitalensis and Corynespora were consistently isolated from the
different tree in tropical forest of southern India. In dry season, endophytic diversity was greater in the dry thorn
forest than in the dry deciduous forest (Murali et al., 2007).
Antagonistic activity of endophytic fungi
Based on the results, 22 isolates of endophytic fungi could inhibit the mycelial growth of fungal pathogens
and only 16 isolates showed the percentage of inhibition above 50%. Isolate Gu03 showed the greatest percentage
of inhibition of mycelial growth with R. microporus (88.1%) and P. noxius (74.1%), respectively (Figure 2. and
Table 2). Isolate Cc12 showed the greatest percentage of inhibition of mycelial growth with C. gloeosporioides
(64.9%) by overgrowth of the tested pathogen (Table 2).
Root rot disease is the most serious problem of rubber tree plantation and present in many countries,
including Thailand. In this study, the antagonistic test showed that the causing pathogen, R. microporus and
P. noxius was against by isolate Gu02 and Gu03 with the highest percentage of inhibition. The antagonistic activity
of fungi to inhibit mycelial growth of the root rot pathogens was considered to control root rot disease in the field
according to the previous studies such as the use of Trichoderma hazianum (Jayasuriya and Thennakoon, 2007)
and Chaetomium cupreum to control R. microporus in vivo (Kaewchai and Soytong, 2010).
The bioactive compounds were produced by the plant but in a mutualistic association with the host plant,
endophytes may enhance bioactive metabolites that process bioactivity such as antibacterial and antifungal activity
in host plant (Radu and Kqueen, 2002; Tejesvi et al., 2007).
Antimicrobial assay of VOCs
The volatile metabolites produce by endophytic fungi were tested for their antimicrobial activity. Isolate
Cs05 showed the greatest antimicrobial activity with all test organisms (Table 3). It was isolated from leaf tissues
of Cycas siamensis (family Cycadaceae), which did not produce spores on several media, including MA, PDA and
WA, and had rope-like mycelium with coiled structures. Many endophytic fungi are known to produce the
bioactive compounds in the form of volatile metabolites with anti-microbial, anti-oxidant and anti-proliferative
activities, cytotoxicity, and fuel production (Naik, 2018). In the family Xylariaceae, Muscodor albus is the first
known fungal endophyte isolated from Cinnamomum zeylanicum, which produces bioactive volatile metabolites
(Strobel et al., 2001; Ezra et al., 2004). An endophytic fungus, Nodulisporium sp. produced the mixture of volatile
compounds were active against plant pathogens. The most abundant identified compound was 1, 8 cineole, 1-
butanol, 2-methyl, and phenyl ethanol alcohol and most importantly cyclohexane, propyl, which is a common
ingredient of diesel fuel (Hassan et al., 2013).
Indole acetic acid (IAA) production
Eighteen isolates of endophytic fungi showed their ability to produce IAA in preliminary test. The range
of IAA production was 11.3 µg/ml to 331.5 µg/ml (Table 2). Endophytic fungus, isolate Gu03 produced maximum
concentration of IAA, following by isolate Gu01 and Cc12, respectively (Table 2). This result was supported by
the previous studies that showed several endophytic fungi, which produced IAA and increased seed germination
and plant growth (Khan et al., 2012, 2015, 2017; Zhou et al., 2013; Kedar et al., 2014). Endophytic fungi enhance
plant growth by produce various secondary metabolites, including IAA, flavonoids and flavnols. These compounds
were determined in the culture filtrate of Aspergillus fumigatus, an endophyte in leaves of wild plant in family
Solanaceae, Withania somnifera (Mehmood et al., 2018). IAA is essential for crop growth and development
because it enhances root, flower development, and other processes (Reinhardt et al., 2000). However, there are
reports that IAA is important tool to against fungal pathogens, such as Colletotrichum spp. (Yue et al., 2000).
Conclusion
Endophytic fungi colonized in plant tissue, especially healthy leaf tissue. They had antimicrobial and
plant-growth promoting properties. Biological control by endophytic fungi to prevent disease and promote plant
growth offer an attractive alternative method for disease management without the negative impact of the chemical
control. Furthermore, volatile metabolite-producing endophytes may be an alternative biological approach as
biofumigation in control of plant diseases.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 12
Acknowledgement
This study was supported by grants from Research and Development Institute Udon Thani Rajabhat
University.
References
Ahmad, F., Ahmad, I. and Khan, M.S. 2005. Indole acetic acid production by the indigenous isolates of
Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turkish Journal of
Biology 29: 29-34.
Banerjee, D., Strobel, G.A., Booth, E., Geary, B., Sears, J., Spakoxicz, D. and Busse, S. 2010. An endophytic
Myrothecium inundatum producing volatile organic compounds. Mycosphere 1: 229-240.
Deshmukh, S.K., Gupta, M.K., Prakash, V. and Saxena, S. 2018. Endophytic fungi: A source of potential
antifungal compounds. Journal of Fungi 4: 1-42.
Espinel, M.A., Laszlo, A., Simonsen, L., Boulahbal, F., Kim, S.J., Reniero, A., Hoffner, S., Rieder, H.L., Binkin,
N., Dye, C., Williams, R. and Raviglione, M.C. 2001. Global trends in resistance to antituberculosis
drugs. The New England Journal of Medicine 344:1294-1303.
Ezra, D., Hess, W.M. and Strobel, G.A. 2004. New endophytic isolates of Muscodor albus, a volatile-antibiotic-
producing fungus. Microbiology 150: 4023-4031.
Gomes, R.R., Glienke, C., Videira, S.I.R., Lombard, L., Groenewald, J.Z. and Crous, P.W. 2013. Diaporthe: a
genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 31: 1-41.
Hassan, S.R., Strobel, G.A., Geary, B. and Sears, J. 2013. An endophytic Nodulisporium sp. from Central America
producing volatile organic compounds with both biological and fuel potential. Journal of Microbiology
and Biotechnology 23: 29-35.
Huang, Z., Cai, X., Shao, C., She, Z., Xia, X., Chen, Y., Yang, J., Zhou, S. and Lin, Y. 2008. Chemistry and weak
antimicrobial activities of phomopsins produced by mangrove endophytic fungus Phomopsis sp. ZSU-
H76. Phytochemistry 69: 1604-1608.
Idris, A., Al-tahir, I. and Idris, E. 2013. Antibacterial activity of endophytic fungi extracts from the medicinal plant
Kigelia africana. Egyptian Academic Journal of Biological Science 5: 1-9.
Jamal, J.A., Ramli, N., Stanslas, J. and Husain, K. 2012. Estrogenic activity of selected Myrsinaceae species in
MCF-7 human breast cancer cells. International Journal of Pharmacy and Pharmaceutical Sciences 4:
547-553.
Jayasuriya, K.E. and Thennakoon, B.I. 2007. Biological control of Rigidoporus microporus, the cause of white
root disease in rubber. Ceylon Journal of Science (Biological Sciences) 36: 9-16.
Kaewchai, S. and Soytong, K. 2010. Application of biofungicides against Rigidoporus microporus causing white
root disease of rubber trees. Journal of Agricultural Technology 2: 349-363.
Karunai, S.B. and Balagengatharathilagam, P. 2014. Isolation and screening of endophytic fungi from medicinal
plants of Virudhunagar district for antimicrobial activity. International Journal of Science and Nature 5:
147-155.
Kedar, A., Rathod, D., Yadav, A., Agarkar, G. and Rai, M. 2014. Endophytic Phoma sp. isolated from medicinal
plants promote the growth of Zea mays. Nusantara Bioscience 6:132-139.
Khan, A.L., Gilani, S.A., Waqas, M., Al-Hosni, K., Al-Khiziri, S., Kim, Y., Ali, L., Kang, S., Asaf, S., Shahzad,
R., Hussain, J., Lee, I. and Al-Harrasi, A. 2017. Endophytes from medicinal plants and their potential for
producing indole-acetic acid, improving seed germination and mitigating oxidative stress. Journal of
Zhejiang University Science B. 18: 125-137.
Khan, S.A., Hamayun, M., Khan, A.L., Lee, I., Shinwari, Z.K. and Kim, J. 2012. Isolation of plant growth
promoting endophytic fungi from dicots inhabiting coastal sand dunes of Korea. Pakistan Journal of
Botany 44: 1453-1460.
Khan, A.R., Ullah, I., Waqas, M., Shahzad, R., Hong, S.J., Park, G.S., Jung, B.K., Lee, I.J. and Shin, J.H. 2015.
Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World
Journal of Microbiology and Biotechnology 31: 1461-1466.
Kharwar, R.N., Verma, S.K., Mishra, A., Gond, S.K., Sharma, V.K., Afreen, T. and Kumar, A. 2011. Assessment
of diversity, distribution and antibacterial activity of endophytic fungi isolated from a medicinal plant
Adenocalymma alliaceum Miers. Symbiosis 55:39-46.
Kokeaw, J., Manoch, L., Worapong, J., Chamswarng, C., Singburaudom, N., Visarathanonth, N., Piasai, O. and
Strobel, G. 2011. Coniochaeta. ligniaria an endophytic fungus from Baeckea frutescens and its
antagonistic effects against plant pathogenic fungi. Thai Journalof Agricultural Science 44: 123-131.
Mehmood, A., Hussain, A., Irshad, M., Khan, N., Hamayun, M., Ismail, Afridi, S.G. and Lee, I. 2018. IAA and
flavonoids modulates the association between maize roots and phytostimulant endophytic Aspergillus
fumigatus greenish. Journal of Plant Interactions 13: 532-542.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 13
Mercier, J., Jiménez-Santamaría, J.I. and Tamez-Guerra, P. 2007. Development of the volatile-producing fungus
Muscodor albus Worapong, Strobel, and Hess as a novel antimicrobial biofumigant. Revista Mexicana
de Fitopatología 25: 173-179.
Morath, S.U., Hung, R. and Bennett, J.W. 2012. Fungal volatile organic compounds: a review with emphasis on
their biotechnological potential. Fungal Biology Reviews 26: 73-83.
Murali, T.S., Suryanarayanan, T.S. and Venkatesan, G. 2007. Fungal endophyte communities in two tropical
forests of southern India: diversity and host affiliation. Mycological Progress 6: 191–199.
Naik, B.S. 2018. Volatile hydrocarbons from endophytic fungi and their efficacy in fuel production and disease
control. Egyptian Journal of Biological Pest Control 28: 1-9.
Nair, D.N. and Padmavathy, S. 2014. Impact of endophytic microorganisms on plants, environment and humans.
The Scientific World Journal 2014: 1-11.
Radu, S. and Kqueen, C.Y. 2002. Preliminary screening of endophytic fungi from medicinal plants in Malaysia
for antimicrobial and antitumor activity. Malaysian Journal of Medical Sciences 9: 23-33.
Reinhardt, D., Mandel, T. and Kuhlemeier, C. 2000. Auxin regulates the initiation and radial position of plant
lateral organs. Plant Cell 12: 507-518.
Rosa, L.H., Tanabca, N., Techen, N., Pan, Z. and Wedge, D.E. 2012. Antifungal activity of extracts from
endophytic fungi associated with Smallanthus maintained in vitro as autotrophic cultures and as pot plants
in the greenhouse. Canadian Journal of Microbiology 58: 1202-1211.
Rungjindamai, N., Pinruan, U., Choeyklin, R., Hattori, T. and Jones, E.B.G. 2008. Molecular characterization of
basidiomycetous endophytes isolated from leaves, rachis and petioles of the oil palm, Elaeis guineensis,
in Thailand. Fungal Diversity 33: 139-161.
Saikkonen, K., Wäli, P., Helander, M. and Faeth, S.H. 2004. Evolution of endophyte-plant symbioses. Trends in
Plant Science 9: 275-280.
Siri-Udom, S., Suwannarach, N. and Lumyong, S. 2 0 1 5 Existence of Muscodor vitigenus, M. equiseti and M.
heveae sp. nov. in leaves of the rubber tree (Hevea brasiliensis Müll.Arg.), and their biocontrol potential.
Annals of Microbiology. 66 :437-448. .
Stinson, M., Ezra, D., Hess, W.M., Sears, J. and Strobel, G. 2003. An endophytic Gliocladium sp. of Eucryphia
cordifolia producing selective volatile antimicrobial compounds. Plant Science 165: 913-922.
Strobel, G. 2006. Harnessing endophytes for industrial microbiology. Current Opinion in Microbiology 9: 240-
244.
Strobel, G. and Daisy, B. 2003. Bioprospecting for microbial endophytes and their natural products. Microbiology
and Molecular Biology Reviews 67: 491–502.
Strobel, G.A., Dirkse, E., Sears, J. and Markworth, C. 2001. Volatile antimicrobials from Muscodor albus, a novel
endophytic fungus. Microbiology 147: 2943-2950.
Suwannarach, N., Bussaban, B., Hyde, K.D. and Lumyong, S. 2010. Muscodor cinnamomi, a new endophytic
species from Cinnamomum bejolghota. Mycotaxon 114: 15-23.
Suwannarach, N., Kumla, J., Bussaban, B., Nuangmek, W., Matsui, K. and Lumyong, S. 2013. Biofumigation with
the endophytic fungus Nodulisporium spp. CMU-UPE34 to control postharvest decay of citrus fruit. Crop
Protection 45: 63 -70.
Tejesvi, M.V., Nalini, M.S., Mahesh, B., Prakash, H.S., Kini, K.R., Shetty, H.S. and Subbiah, V. 2007. New hopes
from endophytic fungal secondary metabolites. Boletín de la Sociedad Química de México 1: 19-26.
Young-Hyun, Y., Yoon, H., Kang, S., Shin, J., Choo, Y., Lee, I., Lee, J. and Kim, J. 2012. Fungal diversity and
plant growth promotion of endophytic fungi from six halophytes in Suncheon Bay. Journal of
Microbiology and Biotechnology 22: 1549-1556.
Yu, J., Wu, Y., He, Z., Li, M., Zhu, K. and Gao, B. 2018. Diversity and antifungal activity of endophytic fungi
associated with Camellia oleifera. Mycobiology 46: 85-91.
Yue, Q., Miller, C.J., White, J.F. and Richardson, M.D. 2000. Isolation and characterization of fungal inhibitors
from Epichloë festucae. Journal of Agricultural and Food Chemistry 48: 4687-4692.
Zakaria, L., Jamil, M.I.M. and Anuar, I.S.M. 2016. Molecular characterisation of endophytic fungi from roots of
wild banana (Musa acuminata). Tropical Life Sciences Research 27: 153-162.
Zheng, Y., Qiao, X., Miao, C., Liu, K., Chen, Y., Xu, L. and Zhao, L. 2016. Diversity, distribution and
biotechnological potential of endophytic fungi. Annals of Microbiology 66: 529-542.
Zhou, Z., Zhang, C., Zhou, W., Li, W., Chu, L., Yan, J. and Li, H. 2013. Diversity and plant growth-promoting
abilty of endophytic fungi from the five flower plant species collected from Yunnan, Southwest Chaina.
Journal of Plant Interactions 9: 585-591.
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 14
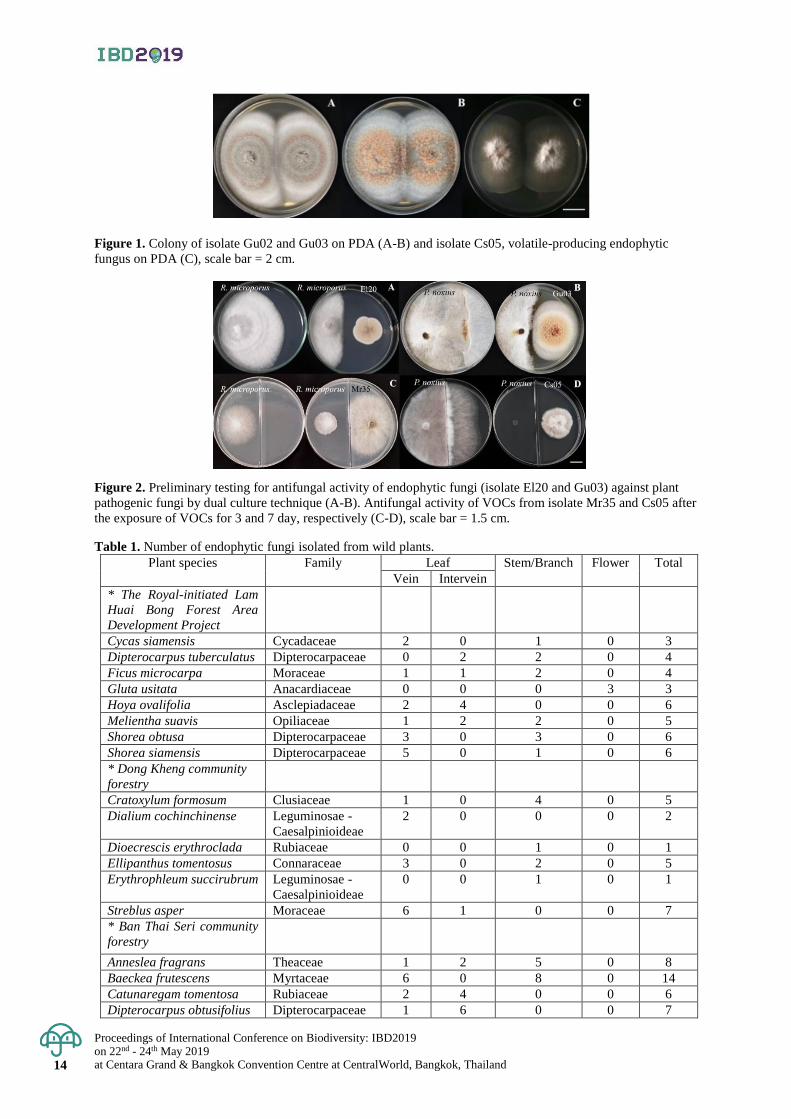
Figure 1. Colony of isolate Gu02 and Gu03 on PDA (A-B) and isolate Cs05, volatile-producing endophytic
fungus on PDA (C), scale bar = 2 cm.
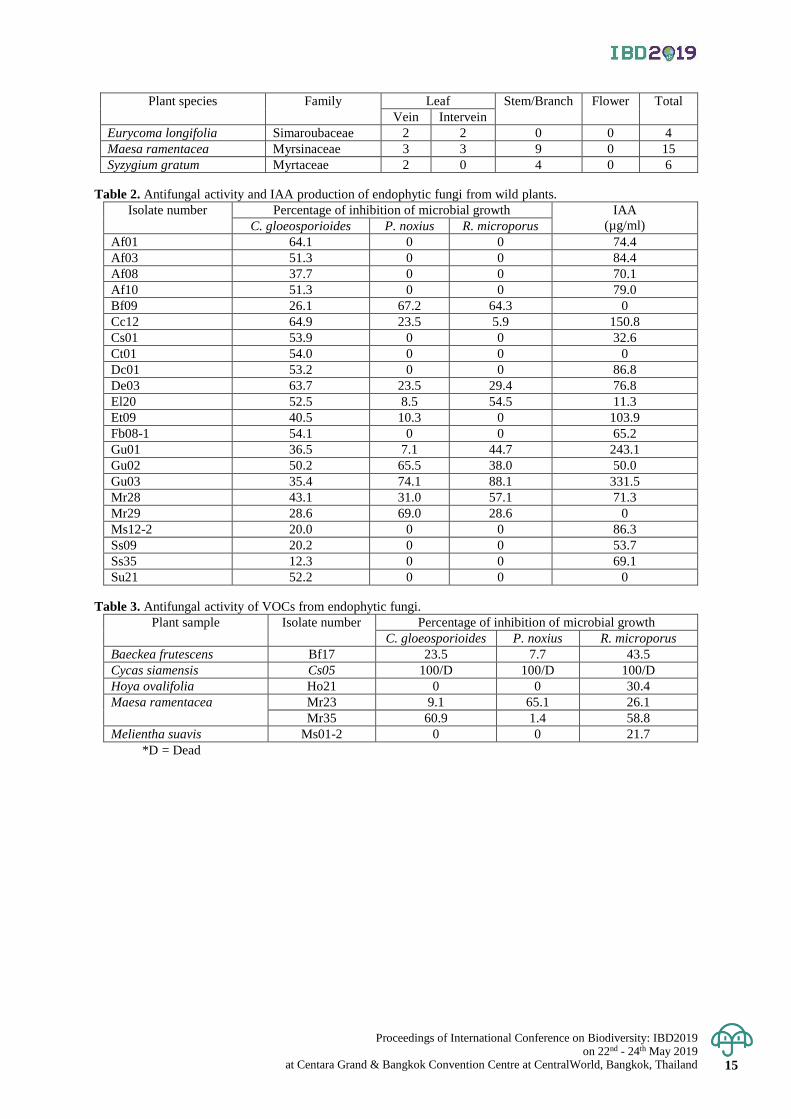
Figure 2. Preliminary testing for antifungal activity of endophytic fungi (isolate El20 and Gu03) against plant
pathogenic fungi by dual culture technique (A-B). Antifungal activity of VOCs from isolate Mr35 and Cs05 after
the exposure of VOCs for 3 and 7 day, respectively (C-D), scale bar = 1.5 cm.
Table 1. Number of endophytic fungi isolated from wild plants.
Plant species Family Leaf Stem/Branch Flower Total
Vein Intervein
* The Royal-initiated Lam
Huai Bong Forest Area
Development Project
Cycas siamensis Cycadaceae 2 0 1 0 3
Dipterocarpus tuberculatus Dipterocarpaceae 0 2 2 0 4
Ficus microcarpa Moraceae 1 1 2 0 4
Gluta usitata Anacardiaceae 0 0 0 3 3
Hoya ovalifolia Asclepiadaceae 2 4 0 0 6
Melientha suavis Opiliaceae 1 2 2 0 5
Shorea obtusa Dipterocarpaceae 3 0 3 0 6
Shorea siamensis Dipterocarpaceae 5 0 1 0 6
* Dong Kheng community
forestry
Cratoxylum formosum Clusiaceae 1 0 4 0 5
Dialium cochinchinense Leguminosae -
Caesalpinioideae
2 0 0 0 2
Dioecrescis erythroclada Rubiaceae 0 0 1 0 1
Ellipanthus tomentosus Connaraceae 3 0 2 0 5
Erythrophleum succirubrum Leguminosae -
Caesalpinioideae
0 0 1 0 1
Streblus asper Moraceae 6 1 0 0 7
* Ban Thai Seri community
forestry
Anneslea fragrans Theaceae 1 2 5 0 8
Baeckea frutescens Myrtaceae 6 0 8 0 14
Catunaregam tomentosa Rubiaceae 2 4 0 0 6
Dipterocarpus obtusifolius Dipterocarpaceae 1 6 0 0 7
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 15
Plant species Family Leaf Stem/Branch Flower Total
Vein Intervein
Eurycoma longifolia Simaroubaceae 2 2 0 0 4
Maesa ramentacea Myrsinaceae 3 3 9 0 15
Syzygium gratum Myrtaceae 2 0 4 0 6
Table 2. Antifungal activity and IAA production of endophytic fungi from wild plants.
Isolate number Percentage of inhibition of microbial growth IAA
(µg/ml) C. gloeosporioides P. noxius R. microporus
Af01 64.1 0 0 74.4
Af03 51.3 0 0 84.4
Af08 37.7 0 0 70.1
Af10 51.3 0 0 79.0
Bf09 26.1 67.2 64.3 0
Cc12 64.9 23.5 5.9 150.8
Cs01 53.9 0 0 32.6
Ct01 54.0 0 0 0
Dc01 53.2 0 0 86.8
De03 63.7 23.5 29.4 76.8
El20 52.5 8.5 54.5 11.3
Et09 40.5 10.3 0 103.9
Fb08-1 54.1 0 0 65.2
Gu01 36.5 7.1 44.7 243.1
Gu02 50.2 65.5 38.0 50.0
Gu03 35.4 74.1 88.1 331.5
Mr28 43.1 31.0 57.1 71.3
Mr29 28.6 69.0 28.6 0
Ms12-2 20.0 0 0 86.3
Ss09 20.2 0 0 53.7
Ss35 12.3 0 0 69.1
Su21 52.2 0 0 0
Table 3. Antifungal activity of VOCs from endophytic fungi.
Plant sample Isolate number Percentage of inhibition of microbial growth
C. gloeosporioides P. noxius R. microporus
Baeckea frutescens Bf17 23.5 7.7 43.5
Cycas siamensis Cs05 100/D 100/D 100/D
Hoya ovalifolia Ho21 0 0 30.4
Maesa ramentacea Mr23 9.1 65.1 26.1
Mr35 60.9 1.4 58.8
Melientha suavis Ms01-2 0 0 21.7
*D = Dead

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 16
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 16 - 20
Diversity of entomopathogenic fungi in protected forest
in the Eastern of Thailand
Winanda Himaman1*, Panrada Jangsantear1, Baramee Sakolruk1,
Kittima Duengkae1, Suchada Mongkolsamrit2, Wasana Noisripoom2,
Janet Jennifer Luangsa-Ard2 and Isarapong Vorapab1
1Department of National Parks, Wildlife and Plant Conservation (DNP), Chatuchak District, Bangkok, Thailand 2National Center of Genetic Engineering and Biotechnology (BIOTEC), National Science and Technology
Development Agency (NSTDA), Khlong Luang District, Pathum Thani, Thailand
*Corresponding author e-mail: [email protected]
Abstract: The entomopathogenic fungi or insect fungi are widespread in nature. They are a well-known as a rich
source of bioactive compounds which prove to useful in medicinal and agricultural applications. There are about
400 species recorded from natural forests in Thailand. However, our knowledge of entomopathogenic fungi
species diversity in the eastern of Thailand is limited. During June 2017-December 2018, we studied species
diversity of entomopathogenic fungi in Khao Soi Dao Wildlife Sanctuary, Chanthaburi Province and Mu Ko Chang
National Park, Trat Province. A total of 537 samples were classified to 3 families; Clavicipitaceae, Cordycipitaceae and
Ophiocordycipitaceae in the order Hypocreales. These fungi were revealed into 43 species based on morphological
character study. The infected insects were in 8 major orders including Coleoptera, Hemiptera, Hymenoptera,
Isoptera, Lepidoptera, Neuroptera, Orthoptera and spider in Araneae order of class Arachnida. The most infected
insects were in Hymenoptera order (44.87 percentage). The most abundant species was Ophiocordyceps unilateralis.
In addition, Ophiocordyceps species found on coleoptera larvae and orthoptera in Mu Ko Chang National Park will
be tentatively described as new species based on morphology and phylogenetic analysis.
Keywords: Entomopathogenic Fungi, Diversity, Taxonomy, Protected Forest, Eastern
Introduction
The term entomopathogenic refers to those microorganisms that are capable of attacking insects using
them as hosts to develop part of their life cycle (Delgado and Murcia, 2011). Entomopathogenic fungi are fungi
that parasitizes the wide range of insects and spiders. The entomopathogenic species are found in almost all
ecosystems. The largest numbers of fungal species that are pathogenic to insects belong to the order (Molnar et
al., 2010). For more than one thousand years, medicinal Ophiocordyceps sinensis has been known as a unique
Tibet's prized parasitic fungus in the Qinghai-Tibetan Plateau for its mysterious life history (Lo et al., 2013).
The genus Beauveria and Metarhizium (Cordycipitaceae) play an important role in controlling insect populations
and have been increasingly utilized as biological control agents of insect pests throughout the world. Nowadays,
the entomopathogenic fungi are becoming increasingly popular from medical and pharmacological researchers
because these fungi are abundant source of useful natural products with various biological activities (Das et al.,
2010). There are about 700 species of entomopathogenic fungi worldwide (Roy et al., 2006).
Thailand is rich in biodiversity located in the tropical areas (Luangsa-ard et al. 2010). At this present,
more than 400 species of entomopathogenic species were reported from Thailand (Luangsa-ard et al. 2010). The
fungus Cordyceps gentilis on a hornet from northern Thailand was the first species recorded by Petch (1932)
which was later considered as Cordyceps sphecocephala by Hywel-Jones (1995).
The diversity of entomopathogenic fungi of the eastern part is poorly known and few collections have
been made. The objectives of the present study were to explore diversity of entomopathogenic fungi in eastern
Thailand. Specifically, Department of National Parks, Wildlife and Plant Conservation (DNP) and BIOTEC have
contributed to the studying of diversity of entomopathogenic fungi in Khao Soi Dao Wildlife Sanctuary (KSD),
Chanthaburi Province and Mu Ko Chang National Park (MKC), Trat Province, Thailand.
Materials and Methods
Collecting area
Entomopathogenic fungi specimens were collected in Khao Soi Dao Wildlife Sanctuary and Mu Ko
Chang National Park during June 2017-December 2018.
Specimen collection and isolation
The undersides and upper sides of leaves, stems and leaf litter in the forest were examined for fungi
growing on dead insects. Collected specimens were placed in plastic boxes and returned to the laboratory for
further examination. Fresh materials were examined and isolated in pure culture from both the anamorph and

International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 17
teleomorph using the technique described by Luangsa-ard et al. (2012). Strains were isolated from the infected
specimens on potato dextrose agar (PDA). Cultures were incubated at 25°C for up to 10 weeks with high moisture
provided by a moist cotton sheets. Slow growing isolates were daily checked. After incubation, fungal growth
was transferred with a sterile needle to new PDA plates. Fungal culture were deposited in the Forest and Plant
Conservation Research Office and the BIOTEC Culture Collection (BCC).
Morphological study Fruiting bodies were examined using free hand sections under a stereo-microscope (Olympus SZX16).
Water-mounted slides were prepared for a microscope study and photographed under a compound microscope
(Olympus CX41). Morphological characteristics of the fungus were examined using classical mycological
techniques based on growth rate and macroscopic and microscopic characteristics. All specimens were dried at
50oC using food dehydrator. The IF numbers refer to herbarium material and to cultures deposited at the Forest
herbarium (mushroom section), DNP and the BCC, Thailand. In this study, clearly distinguishable morphological
samples will be studied molecular taxonomy according to Luangsa-ard et al. (2018). However, the data of
molecular taxonomy will not be showed and discussed in this paper.
Result & Discussion
During this study, we have collected infected insects 397 and 140 samples from KSD and MKC,
respectively. A total of 537 entomopathogenic fungi were collected and then identification on the basis of morphological
characters at Forest Microbiology Laboratory, DNP. The specimens were mostly founded during rainy season
(May to July). All samples were identified 15 genera belonging to 3 families; Clavicipitaceae, Cordycipitaceae
and Ophiocordycipitaceae in the order Hypocreales. These fungi were revealed into 43 species as shown in table
1. Currently, the strains of entomopathogenic fungi are listed in the following orders: Hypocreales (various genera),
Onygenales (Ascosphaera genus), Entomophthorales, and Neozygitales (Entomophthoromycota) (Mora et al., 2017).
Fungal species were varied on collections made over different site. Among these species, some were reported as
biocontrol agents, some with medicinal properties. Ophiocordyceps unilateralis was the most abundant entomopathogenic
fungi species of affixed to the underside of a leaves. Kobmoo et al. (2015) mentioned that O. unilateralis is an
ubiquitous pathogen of ants and in addition the species had hidden phylogenetic diversity associated with host
specificity. The occurrence of B. bassiana and M. anisopliae were zero number in KSD comparing with both were
common in MKC.
The host insects were in 8 major orders including Coleoptera, Hemiptera, Hymenoptera, Isoptera, Lepidoptera,
Neuroptera, Orthoptera and spider in Araneae of class Arachnida. In fact, the hosts are spread among 20 of the 31
orders of insects, in all developmental stages: eggs, larvae, pupae, nymphs, and adults (Araújo and Hughes, 2016).
In this study, the highest frequency of infection insects were in Hymenoptera order (44.87 percentage). Some
fungal species were highly host specific, whereas others had broad host. The genera Aschersonia, Hypocrella and
Moelleriella were specific to scale insects. The Gibellula specimens were regularly collected on spider hosts that
indicate its specific presence. Ants were found to be infected by Ophiocordyceps irangiensis/myrmecophila on
the ground or underneath the leaf litter. A thin single yellow synnema of the O. irangiensis/ myrmecophila arising
from the host was produced. In this case, the hosts were found on the ground, or underneath the leaf litter. In this
study, we found that there were 19 species with hosts in the order Hemiptera. This has clearly shown that Hemiptera
is wide host range. Faria and Wraight (2007) reported approximately 60% of insect diseases are caused by pathogenic fungi.
The isolation of these fungal species in culture was successful in some fungal species. However, the
isolation of O. unilateralis was needed a complicated method and culture medium according by Wongsa et al. (2005).
Furthermore, two Ophiocordyceps species will be described as new species based on morphology and
phylogenetic analysis. The Ophiocordyceps sp. 1 resemble Ophiocordyceps bruneipunctata, buried in the forest
soil, were found parasitizing coleoptera larvae collected from dry evergreen forest at Mu Ko Chang National Park.
The fungus produced pale pink stroma on morphological character. The Ophiocordyceps sp. 2 was collected from
rotting log. Hosts were immature orthopteran. Their morphological character and phylogenetic study clearly
distinct from genera Ophiocordyceps. The surveys of entomopathogenic fungi in Thailand have a very long history
with BIOTEC staff. Their several field studies have reported the new species of entomopathogenic fungi from
natural forests (Luangsa-ard et al., 2017; Luangsa-ard et al., 2018; Khonsanit et al., 2019; Kuephadungphan et al.,
2019). These findings encourage further research on the fungus in protected forests including national parks and
wildlife sanctuaries in Thailand.
Conclusion
Entomopathogenic fungi were mainly found in order Hypocreales (Clavicipitaceae, Cordycipitaceae and
Ophiocordycipitaceae). Fungus diversity was 43 species from both protected forests in the eastern part of Thailand.
Fungal common to both study sites was Ophiocordyceps irangiensis/ myrmecophila. Two species of Ophiocordyceps
proved to be undescribed and will be proposed as new species after confirmed with molecular results. At this

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 18
present, studying biodiversity need to explore the potential economic benefit from entomopathogenic fungi
concurrently. Thus, there is an important to contribute the collaboration with many research groups including
biotechnologists, pharmacologists, analytical chemists, medical scientists etc. (Shrestha et al., 2016).
Acknowledgement
We would like to thank the staff of Khao Soi Dao Wildlife Sanctuary and Head of Mu Ko Chang National
Parks for splendid hospitality whilst we were on field trips. We also would like to thank Forest and Plant
Conservation Research Office, Department of National Parks, Wildlife and Plant Conservation for the support of
the project "Diversity of invertebrate-pathogenic fungi and wild mushrooms in protected forest in eastern and
northeastern of Thailand".
References
Araújo, J.P., and Hughes, D.P. 2016. Diversity of entomopathogenic fungi: which groups conquered the insect
body?. In Advances in genetics. Vol. 94: 1–39. Academic Press.
Das, S.K., Masuda, M., Sakurai, A., and Sakakibara, M. 2010. Medicinal uses of the mushroom Cordyceps
militaris: current state and prospects. Fitoterapia 81(8): 961–968.
Delgado, P.A.M. and Murcia, O.P. 2011. Hongos entomopatógenos: uma alternativa para la obtención de Biopesticidas.Ambi-
Agua6: 77–90.
Faria, M.R., and Wraight, S.P. 2007. Mycoinsecticides and myco-acaricides: a comprehensive list with worldwide
coverageand international classification of formulation types. BiolControl 43: 237–256.
Hywel-Jones, N., 1995. Cordyceps sphecocephala and a Hymenostilbe sp. infecting wasps and bees in Thailand.
Mycological Research 2(99): 154–158.
Khonsanit, A., Luangsa-ard, J.J., Thanakitpipattana, D., Kobmoo, N., and Piasai, O. 2019. Cryptic species within Ophiocordyceps
myrmecophila complex on formicine ants from Thailand. Mycological Progress 18(1–2): 147–161.
Kobmoo, N., Mongkolsamrit, S., Wutikhun, T., Tasanathai, K., Khonsanit, A., Thanakitpipattana, D., and
Luangsa-ard, J.J. 2015. New species of Ophiocordyceps unilateralis, an ubiquitous pathogen of ants from
Thailand. Fungal Biology 119(1): 44–52.
Kuephadungphan, W., Macabeo, A.P.G., Luangsa-ard, J.J., Tasanathai, K., Thanakitpipattana, D., Phongpaichit,
S., Yuyama, K., and Stadler, M. 2019. Studies on the biologically active secondary metabolites of the
new spider parasitic fungus Gibellula gamsii. Mycological Progress 18(1–2): 135–146.
Lo, H.C., Hsieh, C., Lin, F.Y., and Hsu, T.H. 2013. A Systematic review of the mysterious caterpillar fungus
Ophiocordyceps sinensis in Dong-ChongXiaCao (Dōng Chóng Xià Cǎo) and related bioactive
ingredients. Journal of Traditional and Complementary Medicine 3(1): 16–32.
Luangsa-ard, J.J., Tasanathai, K., Mongkolsamrit, S., and Hywel-Jones, N.L. 2010. Atlas of Invertebrate-
Pathogenic Fungi of Thailand Volume 3. National Center for Genetic Engineering and Biotechnology.
National Science and Technology Development Agency. Bangkok.
Luangsa-ard, J.J., Tasanathai, K., Mongkolsamrit, S., and Hywel-Jones, N.L. 2012. Atlas of Invertebrate-
Pathogenic Fungi of Thailand. Volume 4. National Center for Genetic Engineering and Biotechnology.
National Science and Technology Development Agency. Bangkok.
Luangsa-ard, J.J., Mongkolsamrit, S., Noisripoom, W., Thanakitpipattana, D., Khonsanit, A., and Wutikhun, T.,
2017. Helicocollum, a new clavicipitalean genus pathogenic to scale insects (Hemiptera) in Thailand.
Mycological progress 16(4): 419–431.
Luangsa-ard, J.J., Tasanathai, K., Thanakitpipattana, D., Khonsanit, A., and Stadler, M. 2018. Novel and interesting
Ophiocordyceps spp. (Ophiocordycipitaceae, Hypocreales) with superficial perithecia from Thailand. Studies in
Mycology 89: 125–142.
Molnar, I., Gibson, D.M., and Krasnoff, S.B. 2010. Secondary metabolites from entomopathogenic Hypocrealean
fungi. Natural Product Reports 27(9): 1241–1275.
Mora, M.A.E., Castilho, A.M.C., and Fraga, M.E. 2017. Classification and infection mechanism of
entomopathogenic fungi. Arquivos do Instituto Biológico 84: 1–10.
Petch, T., 1932. Notes on entomogenous fungi. Transactions of the British Mycological Society, 16(4): 209–245.
Roy, H.E., Steinkraus, D.C., Eilenberg, J., Hajek, A.E., and Pell, J.K. 2006. Bizarre interactions and endgames:
entomopathogenic fungi and their arthropod hosts. Annu Rev Entomol 51: 331–357.
Shrestha, B., Tanaka, E., Hyun, M.W., Han, J.G., Kim, C.S., Jo, J.W., Han, S.K., Oh, J., and Sung, G.H. 2016.
Coleopteran and lepidopteran hosts of the entomopathogenic genus Cordyceps sensu lato. Journal of
Mycology. 1–15.
Wongsa, P., Tasanatai, K., Watts, P., and Hywel-Jones, N. 2005. Isolation and in vitro cultivation of the insect
pathogenic fungus Cordyceps unilateralis. Mycological Research 109(8): 936–940.
doi:10.1017/S0953756205003321.
International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 19
Table 1. Diversity of entomopathogenic fungi in KSD, Khao Soi Dao Wildlife Sanctuary and MKC, Mu Ko
Chang National Park in eastern, Thailand (June 2017 to December 2018).
Genus/species Host/Insect Order KSD MKC
Family Clavicipitaceae
Aschersonia badia
Aschersonia confluens
Aschersonia luteola
Aschersonia marginata
Aschersonia placenta
Aschersonia sp.
Aschersonia oxystoma
Conoideocrella luteorostrata
Conoideocrella tenuis
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
0
2
0
1
6
2
0
2
3
1
8
3
1
1
1
3
7
0
Hypocrella badia
Hypocrella calendulina
Hypocrella discoidea
Hypocrella sp.
Hypocrella luteola
Metarhizium anisopliae
Moelleriella mollii
Moelleriella raciborskii
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Scale insect/Hemiptera
Coleopteran/Coleoptera
Scale insect/Hemiptera
Scale insect/Hemiptera
1
5
2
1
0
0
3
8
0
0
6
0
1
1
7
4
Family Cordycipitaceae
Akanthomyces pistillariiformis
Akanthomyces sp.
Beauveria bassiana
Beauveria sp.
Cordyceps javanica
Cordyceps sp.
Cordyceps tenuipes
Cordyceps tuberculata
Engyodontium sp.
Gibellula pulcha
Gibellula sp.1
Gibellula sp.2
Isaria sp.
Torrubiella hemipterigenum
Moth/Lepidoptera
Moth/Lepidoptera
Coleopteran/Coleoptera
Coleopteran/Coleoptera
Spider/Araneae
Spider/Araneae
Lepidopteran pupae/Lepidoptera
Moth/Lepidoptera
Scale insect/Hemiptera
Spider/Araneae
Spider/Araneae
Spider/Araneae
Spider/Araneae
Spider/Araneae
Leafhopper/ Hemiptera
0
0
0
0
1
1
1
0
0
0
1
1
8
0
1
1
1
21
5
0
0
2
3
1
1
2
3
0
1
3

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 20
Table 1. (continued)
Genus/species Host/Insect Order KSD MKC
Family Ophiocordycipitaceae
Ophiocordyceps irangiensis/myrmecophila
Ophiocordyceps longissima
Ophiocordyceps sporangifera
Ophiocordyceps sp. 1
Ophiocordyceps sp. 2
Ant/Hymenoptera
Cicada/Hemiptera
Elateridae larvae/Coleoptera
Coleopteran larvae/Coleoptera
Orthopteran/Orthoptera
52
3
0
0
0
29
7
1
28
7
Ophiocordyceps sphecocephala
Ophiocordyceps unilateralis
Ophiocordyceps cf. communis
Ophiocordyceps nutan
Ophiocordyceps houaynhangensis
Polycephalomyces nipponicus
Purpureocillium takamizusanensis
Wasp/Hymenoptera
Ant/Hymenoptera
Termite/Isoptera
Stink bug/Hemiptera
Lepidopteran/Lepidoptera
Neuroptera
Stink bug/Hemiptera
0
141
51
2
49
10
1
3
15
0
0
0
0
0
Total 397 140
Figure 1. a) Ophiocoryceps sp. 1 in habitat showing pale pink stroma emerging from coleopteran larva
underneath the forest ground b) Ophiocoryceps sp. 2 with mushroom-like head on orthopteran.
a b

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 21
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 21 - 32
Diversity of mushrooms at Mu Ko Chang National Park, Trat Province
Baramee Sakolrak*, Panrada Jangsantear, Winanda Himaman, Tiplada Tongtapao,
Chanjira Ayawong and Kittima Duengkae
Forest and Plant Conservation Research Office, Department of National Parks,
Wildlife and Plant Conservation, Chatuchak District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Diversity of mushrooms at Mu Ko Chang National Park was carried out by surveying the mushrooms
along natural trails inside the national park. During December 2017 to August 2018, a total of 246 samples were
classified to 2 phyla Fungi; Ascomycota and Basidiomycota. These mushrooms were revealed into 203 species
based on their morphological characteristic. They were classified into species level (78 species) , generic level
(103 species) and unidentified (22 species). All of them were divided into 4 groups according to their ecological
roles in the forest ecosystem, namely, saprophytic mushrooms 138 species (67.98%), ectomycorrhizal mushrooms
51 species (25.12%) , plant parasitic mushrooms 6 species (2.96%) termite mushroom 1 species (0.49%). Six
species (2.96%) were unknown ecological roles and 1 species as Boletellus emodensis (Berk.) Singer are both of
the ectomycorrhizal and plant parasitic mushroom. The edibility of these mushrooms were edible (29 species) ,
inedible (8 species) and unknown edibility (166 species). Eleven medicinal mushroom species were recorded in
this study. The most interesting result is Spongiforma thailandica Desjardin, et al. has been found, the first report
found after the first discovery in 2009 at Khao Yai National Park by E. Horak, et al.
Keywords: Species list, ecological roles, edibility, protected area, Spongiforma thailandica
Introduction
Mushroom is a group of fungi which has the reproductive part known as the fruit body or fruiting body
and develops to form and distribute the spores. The mushrooms are a very large class of organisms which in their
structure have some similarities to plants, by they lack chlorophyll and are thus unable to build up the carbon
compounds essential to life. Instead, they draw their sustenance ready-made from living or dead plants or even
animals, as animal do (Phillips, 2006). They can be epigeous or hypogeous and are divided into two distinct groups
according to sexual reproductive forms as Basidiomycetous and Ascomycetous mushrooms. The Basidiomycetous
mushrooms are a diverse group of fungi composed of agarics, boletes, tooth fungi, chanterelles, coral fungi,
polypores, puffballs, earth stars, stinkhorns, bird’s nest fungi and jelly fungi. The Ascomycetous mushrooms are
included the cup fungi, morels, truffles, earth tongues, and saddle fungi. There are approximately 1.5 million species
of fungi found on earth (Hawksworth, 2001; 2004) and approximately 41,000 species of mushrooms (Deshmukh,
2004) . An estimated 1,069 mushroom species have been reported as being used for food purposes (Boa, 2004) .
According to Sangwanit et al.(2013) the numbers of mushrooms recorded in Thailand were over 2,500 species.
Mu Ko Chang National Park is a big archipelago of over 40 islands in Trat Province, east of Thailand. It
became established in 1982 as 45th national park of Thailand. The total area is 650 square kilometers of which
two-thirds is marine area. Its landscape is mostly high mountains and stone cliffs. The highest mountain is Khao
Salak Phet (743 meters above mean sea level) . The forests are very abundant as tropical rain forest, mangrove
forest and beach forest. There are many waterfalls on the island such as Than Mayom Waterfall, Khlong Phlu
Waterfall, etc. There are many plants, including Dipterocarpus alatus, Anisoptera costala, Hopea odorata,
Terminalia catappa, Calophyllum inophyllum and Acanthus ebracteatus. The park’s wildlife includes 29 mammal
species, 74 types of birds and 42 kinds of reptiles such as Ko Chang Frog (Limnonectes kohchangae Smith) , the
endemic animal of Ko Chang (Department of National Parks, Wildlife and Plant Conservation, 2019) . In 1902,
Rostrup, the first recorder species of mushroom in Ko Chang, reported 94 species of fungi including 1 rust fungus,
2 Myxomycetes and 91 mushrooms species (Rostrup, 1902) . This research is the first mushroom survey and
reported after 117 years ago.
Materials and Methods
Study areas
The mushroom specimens were collected along the nature trails inside Ko Chang main islands showed
in Figure 1. such as Than Mayom Waterfall, Khlong Phlu Waterfall, Khlong Nonsi Waterfall, Salak Phet trail and
Ban Bang Bao trail in rainy and winter seasons of the years from 2017 to 2018 (December 2017 to August 2018).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
22
22
Mushroom identification All collected mushrooms were studied on macroscopic and microscopic characteristics. Macroscopic
characters required for identification such as size and color of the pileus and stipe, habit, habitat, substratum, odor,
presence or absence of veil and volva were noted from the fresh fruit bodies in the field and photographed in their
natural habitat (Largent, 1973). Microscopic characters required such as shape, size, color and ornamented spores,
basidia, cystidia and chemical reaction on Melzer’s solution (Largent et al. , 1977). All characteristics were used
to identify all mushrooms into species if possible by various identification keys and books (Corner and Bas, 1962;
Dring, 1964; Corner, 1966; 1972; 1981; Lowy, 1951; 1952; Moser, 1973; Pegler, 1986; Largent and Baroni, 1988;
Ruksawong and Flegel, 2001; Imazeki et al. , 2005; Phillips, 2006; Miller and Miller, 2006; Chandrasrikul et al. ,
2008 Sanoamuang, 2010; Sangwanit et al. , 2013; Bandara et al. , 2017) . Besides these keys and books, some
authentic websites were accessed such as www.mushroomexpert.com. After proper identification, the current name
of the identified mushrooms and their taxonomic details were accessed from the website: www.catalogueoflife.org and
www.indexfungorum.org (accessed on March 6, 2019). All dried specimens are deposited at the Forest Herbarium
(Mushrooms Section), under the supervision of Department of National Parks, Wildlife and Plant Conservation.
Result & Discussion
The 246 mushroom specimens were collected from various nature trails in Mu Ko Chang National Park
in main island. They were classified to 2 phyla; Ascomycota and Basidiomycota. These mushrooms were revealed
into 203 species based on their morphological characteristic only. They were classified into species level (78
species) , generic level ( 103 species) and unidentified ( 22 species) ( Table 1.) . Since this study used only
morphological characteristics for classification. If they were studied more by molecular techniques. The number
and species of collected mushrooms may be changed.
All of collected mushrooms were divided into 4 groups according to their ecological roles in the forest
ecosystem, namely, saprophytic mushrooms 138 species ( 67. 98% ) , ectomycorrhizal mushrooms 51 species
(25.12%), plant parasitic mushrooms 6 species (2.96%) termite mushroom 1 species (0.49%). Six species (2.96%)
were unknown ecological roles and 1 species as Boletellus emodensis (Berk. ) Singer are both of the ectomycorrhizal
and plant parasitic mushroom (Table 1. and Figure 2.).
The edibility of these mushrooms were edible (29 species) , inedible (8 species) and unknown edibility
(166 species). The eleven medicinal mushroom species were recorded (Table 1. and Figure 3.). Most of edible mushrooms
were in the genus Russula which were found at least five species such as R. alboareolata Hongo, R. cyanoxantha
(Schaeff.) Fr., R. delica Fr., etc. This genus was well known as one of the finest foods derived from the forest especially
in the north and northeast of Thailand and had a high price around 270 baht/kg (Sakolrak et al., 2018). According
to this study, none of Ko Chang villagers collected wild mushrooms for eating or selling. It is possible that the
economic system in this area tied to tourism.
In 1902, Rostrup, the first mushroom species recorder in Ko Chang, reported 94 species of fungi
including 1 rust fungus, 2 Myxomycetes and 91 mushrooms species. There were only four mushroom species that
were found in this study as Microporus xanthopus ( Fr. ) Kuntze, Pycnoporus sanguineus ( L. ) Murrill,
Schizophyllum commune Fr. and Stereum ostrea (Blume & T. Nees) Fr. So the summary numbers of mushroom
species are reported from Mu Ko Chang National Park are 290 species.
The most interesting result is the first report found of Spongiforma thailandica Desjardin, et al. outside
the Khao Yai National Park, the first discovery in 2009 by E. Horak, et al. (Desjardin et al., 2009). The specimens
were collected in December 2017. The research team has tried to track more specimens but still cannot collect
more specimens. The macroscopic and microscopic characters of S. thailandica, which found at Mu Ko Chang
National Park (Figure 4.), has quite similar to previous publication (Desjardin et al., 2009) except some chemical
reaction tests for example 10% potassium hydroxide (10% KOH) that it was not tested in this study. The fresh
fruit bodies that found at Kho Yai National Park were immediately deep purple to purplish black in 10% KOH
(Desjardin et al. , 2009) . In this occasion, the research team tried to extract DNA according to Desjardin et al.
(2009) method. The PCR reaction was performed for internal transcribed spacer ( ITS) regions using the primer
combinations ITS1-ITS4 and sequencing was done commercially (Marcrogen, Korea). The ITS gene sequencing
results were compared with the GenBank database (http: / /www.ncbi.nlm.nih.gov/ ) using BLAST search. The
resultant ITS gene sequences were aligned using CLUSTAL W software (Larkin et al. , 2007) in the program
BioEdit (Hall, 2011). The results showed 98.32% similar to Spongiforma thailandica BBH DED 7873. However,
98.32% similar is still not strong enough. If we study in more other genes regions. The difference may be found
and leading to being a new species in the future.
Conclusion
This research was the second time to survey diversity of mushrooms at Mu Ko Chang National Park after
117 years ago. Two hundred and three mushrooms species were collected inside Ko Chang island, the main island
of Mu Ko Chang National Park, from various nature trails. The consisted of saprophytic mushrooms, ectomycorrhizal

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 23
mushrooms, plant parasitic mushrooms, termite mushroom, unknown ecological roles and both of the ectomycorrhizal
and plant parasitic mushroom are 138, 51, 6, 6, 1 and 1 species respectively. The twenty nine species of edible
mushrooms were recorded in this survey. The most interesting result is that the first report found that Spongiforma
thailandica Desjardin, et al. were spread outside the Khao Yai National Park after the first discovery in 2009.
Acknowledgement
This research was funded by Department of National Parks, Wildlife and Plant Conservation (DNP) ,
Ministry of Natural Resources and Environment. We would like to thank Mr. Itsarapong Voraphab and Mr. Boonsong
Sriyotsombat for their kind helps on fieldwork and Mr. Kosit Nilrat, Head of Mu Ko Chang National Parks, and his
staffs for various facilities.
References
Bandara, A. R. , Karunarathna, S. C. , Phillips, A. J. L. , Mortimer, P.E. , Xu, J. , Kakumyan, P. , and Hyce, K.D.
2017. Diversity of Auricularia (Auriculariaceae, Auriculariales) in Thailand. Phytotaxa 292: 19–34.
Boa, E. 2004. Wild edible fungi: a global overview of their use and importance to people. NonWood Forest Products,
No. 17, FAO, Forestry Department, Rome.
Catalogue of Life. 2019. Catalogue of Life: 2018 Annual Checklist. Available Source:
http://www.catalogueoflife.org/annual-checklist/2018, March 6, 2019.
Chandrasrikul, A., Suwanarit, P., Sangwanit, U., Morinaga, T., Nishizawa, Y., and Murakami, Y. 2008. Diversity of Mushrooms
and Macrofungi in Thailand. Kasetsart University Press, Bangkok.
Corner, E. L. H., and Bas, C. 1962. The genus Amanita in Singapore and Malaya. Persoonia 2: 241–304.
Corner, E. L. H. 1966. A monograph of Cantharelloid fungi, Ann. Bot. Mem. 2. Oxford University Press, London.
Corner, E. L. H. 1972. Boletus in Malaysia. The Government Printing Office, Singapore.
Corner, E. L. H. 1981. The agaric genera Lentinus, Panus and Pleurotus with particular reference to Maraysian
species, Beih. Nova Hedwigia 69. J. Cramer, Hirschberg.
Department of National Parks, Wildlife and Plant Conservation (DNP) . 2019. Mu Ko Chang National Park.
Available Source: http://portal.dnp.go.th/DNP/FileSystem/download?uuid=8d3f6ee9-6e15-468b-ad21-
1701b528ae07.pdf, March 5, 2019.
Deshmukh, S. K. 2004. Biodiversity of tropical basidiomycetes as sources of novel secondary metabolites, pp.
121–140. In P. C. Jain ( ed. ) . Microbiology and Biotechnology for Sustainable Development. CBS
Publisher and Distributors, New Delhi.
Desjardin, D. E. , Binder, M. , Roekring, S. , and Flegel, T. 2009. Spongiforma, a new genus of gasteroid boletes
from Thailand. Fungal Diversity 37: 1–8.
Dring, D. M. 1964. Gasteromycetes of West Tropical Africa, Mycological Paper No. 98. Commonwealth
Mycological Institute, Surrey.
Hall, T. 2011. BioEdit: an important software for molecular biology. GERF Bull Biosci. 2: 60–61.
Hawksworth, D. L. 2001. The magnitude of fungal diversity: the 1. 5 million species estimate revisited.
Mycological Research 105: 1422–1432.
Imazeki, R., Otani, Y., and Hongo, T. 2005. Fungi of Japan, 2nd edition. YAMA-KEI Publisher Co., Ltd., Tokyo. (in Japanese)
Index Fungorum Partnership. 2019. Index Fungorum – Search Page. Available Source:
http://www.indexfungorum.org/Names/Names.asp, March 6, 2019.
Kuo, M. 2019. Mushroomexpert.com homepage. Available Source: http://www.mushroomexpert.com, December
20, 2017 – January 31, 2019.
Largent, L. D. 1973. How to Identify Mushrooms to Genus I: Macroscopic Features. Mad River Press, Inc., California.
Largent, L. D. , Johnson, D. , and Watling, R. 1977. How to Identify Mushrooms to Genus III: Microscopic
Features. Mad River Press, Inc., California.
Largent, L. D., and Baroni, T. J. 1988. How to Identify Mushrooms to Genus VI: Modern Genera. Mad River Press, Inc.,
California.
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam, H., Valenti, F., Wallace, I. M.,
Wilm, A., Lopez, R., Thompson, J. D., Gibson, T. J., and Higgins, D. G. 2007. Glustal W and Clustal X
version 2.0. Bioinformatics 23: 2947–2948.
Lowy, B. 1951. A morphological basis for classifying the species of Auricularia. Mycologia 43: 351–358.
Lowy, B. 1952. The genus Auricularia. Mycologia 44: 656–692.
Miller, O. K. Jr., and Miller, H.H. 2006. North American Mushrooms: a Field Guide to Edible and Inedible Fungi.
Globe Pequot Press, Connecticut.
Moser, M. 1973. Keys to Agaricus and Boleti (Polyporales, Boletales, Agaricales, Russulales). Whitefriars Press Ltd.,
Tongridge.
Pegler, D. N. 1986. Agaric Flora of Sri Lanka, Kew Bulletin Additional Series XII. Royal Botanic Gardens, Kew.
Phillips, R. 2006. Mushrooms. Pan Macmillan Ltd. Oxford.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
24
24
Ruksawong, P., and Flegel, T. W. 2001. Thai Mushrooms and Other Fungi. National Center for Genetic
Engineering and Biotechnology and National Science and Technology Development Agency, BIOTEC,
Bangkok. (in Thai)
Rostrup, E. 1902. Flora of Koh Chang; Contribution to the knowledge of the vegetation in the Gulf of Siam, Part
6. Botanisk Tidsskrift 24: 355–363.
Sakolrak, B., Srisuwun, S., Himaman, W., Duengkae, K., Jangsantear, P. and Pongpanich, K. 2018. Utilization
of mushrooms around Nam Phong National Park and Phu Wiang National Park, Khon Kaen Province.
Proceeding of the 5th National Meeting on Biodiversity Management in Thailand, July 10th–14th, 2018,
Surat Thani, Thailand p. 1–12. (in Thai)
Sangwanit, U., Suwanarit, P., Payappanon, A., Luangsa-ard, J.J., Chandrasrikul, A., and Sakolrak, B. 2013.
Checklists of Biological Properties: Mushrooms. Biodiversity-Based Economy Development Office
(Public Organization), Bangkok. (in Thai)
Sanoamuang, N. 2010. Wild Mushrooms of Thailand: Biodiversity and Utilization. Universal Graphic and
Trading Co., Ltd., Bangkok. (in Thai)
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 25
Figure 1. Map of Mu Ko Chang National Park (DNP, 2019); 1 = Khlong Nonsi Waterfall, 2 = Khlong Phlu
Waterfall, 3 = Than Mayom Waterfall, 4 = Salak Phet trail, and 5 = Ban Bang Bao trail.
Figure 2. The ecological roles of mushrooms collected from Mu Ko Chang National Park.
Figure 3. The edibility information of mushrooms collected from Mu Ko Chang National Park.
1
2 3
4
5

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
26
26
Figure 4. Spongiforma thailandica were found at Mu Ko Chang National Park; a. fruit bodies, b. basidiospores.
Table 1. Species list of mushrooms collected from Mu Ko Chang National Park, Trat Province, their edibility
information (EI) and ecological roles (ER).
Scientific Name EI ER
Phylum Ascomycota
Class Geoglossomycetes
Order Geoglossales
Family Geoglossaceae
Trichoglossum hirsutum (Pers.) Boud. Une Unk Class Leotiomycetes
Order Helotiales
Family Helotiaceae
Bisporella citrina (Batsch) Korf & S. E. Carp. Une Sap
Class Pezizomycetes
Order Pezizales
Family Pyronemataceae
Aleuria luteonitens (Berk. & Broome) Gillet Une Sap Family Sarcoscyphaceae
Cookeina sulcipes (Berk.) Kuntze Une Sap Cookeina tricholoma (Mont.) Kuntze Une Sap
Class Sordariomycetes
Order Xylariales
Family Xylariaceae
Xylaria allantoidea (Berk.) Fr. Une/Med Sap Xylaria polymorpha (Pers.) Grev. Une/Med Sap Xylaria sp.1 Une Sap Xylaria sp.2 Une Sap Xylaria sp.3 Une Sap
Phylum Basidiomycota
Class Agaricomycetes
Order Agaricales
Family Agaricaceae
Agaricus trisulphuratus Berk. Une Sap Agaricus sp.1 Une Sap Agaricus sp.2 Une Sap Calvatia gardneri (Berk.) Lloyd Edi Sap Coprinus sp.1 Une Sap Cyathus striatus (Huds.) Willd. Ine Sap
a. b.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 27
Scientific Name EI ER Cyathus sp.1 Une Sap Lepiota sp.1 Une Sap Lepiota sp.2 Une Sap Leucocoprinus fragilissimus (Ravenel ex Berk. & M.A. Curtis) Pat. Une Sap Lycoperdon sp.1 Une Sap Lycoperdon sp.2 Une Sap Unidentified 1 Une Sap
Family Amanitaceae
Amanita javanica (Corner & Bas) T. Oda, C. Tanaka & Tsuda Edi Ect Amanita mira Corner & Bas Une Ect Amanita virgineoides Bas Edi Ect Amanita sp.1 Une Ect
Family Clavariaceae
Clavaria fumosa Pers. Une Sap Clavaria sp.1 Une Sap Clavaria sp.2 Une Sap Clavaria sp.3 Une Sap Clavulinopsis helvola (Pers.) Corner Une Sap
Family Entolomataceae
Entocybe nitida (Quél.) T.J. Baroni, Largent & V. Hofst. Une Sap Entoloma sp.1 Une Sap Entoloma sp.2 Une Sap Entoloma sp.3 Une Sap
Family Hygrophoraceae
Hygrocybe cantharellus (Fr.) Murrill Edi Sap Hygrocybe sp.1 Une Sap Hygrocybe sp.2 Une Sap Hygrocybe sp.3 Une Sap
Family Hymenogastraceae
Gymnopilus sp.1 Une Sap Naematoloma sp.1 Une Sap Psilocybe sp.1 Une Sap
Family Inocybaceae
Crepidotus sp.1 Une Sap Inocybe rimosa (Bull.) P. Kumm. Une Ect
Family Lyophyllaceae
Termitomyces sp.1 Edi Ter Family Marasmiaceae
Campanella junghuhnii (Mont.) Singer Une Sap Campanella sp.1 Une Sap Campanella sp.2 Une Sap Marasmius siccus (Schwein.) Fr. Une Sap Marasmius sp.1 Une Sap Tetrapyrgos nigripes (Fr.) E. Horak Une Sap Trogia infundibuliformis Berk. & Broome Une Sap
Family Mycenaceae
Mycena sp.1 Une Sap Mycena sp.2 Une Sap

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
28
28
Scientific Name EI ER Mycena sp.3 Une Sap Mycena sp.4 Une Sap Mycena sp.5 Une Sap Panellus pusillus (Pers. ex Lév.) Burds. & O.K. Mill. Une Sap Xeromphalina tenuipes (Schwein.) A.H. Sm. Une Sap
Family Omphalotaceae
Gymnopus sp.1 Une Sap Family Physalacriaceae
Oudemansiella canarii (Jungh.) Höhn. Edi Sap
Family Pleurotaceae
Pleurotus sp.1 Une Sap Family Psathyrellaceae
Psathyrella candolleana (Fr.) Maire Edi Sap
Family Pterulaceae
Deflexula fascicularis (Bres. & Pat.) Corner Une Sap Family Schizophyllaceae
Schizophyllum commune Fr. Edi/Med Sap
Family Tricholomataceae
Clitocybe sp.1 Une Sap Collybia sp.1 Une Sap Collybia sp.2 Une Sap Collybia sp.3 Une Sap Collybia sp.4 Une Sap Collybia sp.5 Une Sap Resupinatus applicatus (Batsch) Gray Une Sap
Unidentified 2 Une Sap Unidentified 3 Une Unk Unidentified 4 Une Unk Unidentified 5 Une Sap
Order Auriculariales
Family Auriculariaceae
Auricularia nigricans (Sw.) Birkebak, Looney & Sánchez-García Edi/Med Sap Auricularia thailandica Bandara & K.D. Hyde Edi Sap Auricularia sp.1 Une Sap
Order Boletales
Family Boletaceae
Boletellus emodensis (Berk.) Singer Edi Ect/Sap Boletus sp.1 Une Ect Boletus sp.2 Une Ect Boletus sp.3 Une Ect Boletus sp.4 Une Ect Boletus sp.5 Une Ect Boletus sp.6 Une Ect Boletus sp.7 Une Ect Boletus sp.8 Une Ect Chalciporus piperatus (Bull.) Bataille Edi Ect Phylloporus bellus (Massee) Corner Edi Ect Phylloporus sp.1 Edi Ect

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 29
Scientific Name EI ER Spongiforma thailandica Desjardin, Manfr. Binder, Roekring & Flegel
Une Ect Strobilomyces strobilaceus (Scop.) Berk. Edi Ect
Family Calostomataceae
Calostoma japonicum Henn. Une Ect Family Sclerodermataceae
Scleroderma sinnamariense Mont. Une Ect Scleroderma sp.1 Une Ect Scleroderma sp.2 Une Ect Scleroderma sp.3 Une Ect
Order Cantharellales
Family Cantharellaceae
Cantharellus sp.1 Une Ect Craterellus aureus Berk. & M.A. Curtis Une Ect Craterellus odoratus (Schwein.) Fr. Une Ect
Family Hydnaceae
Hydnum rufescens Pers. Une Ect Order Geastrales
Family Geastraceae
Geastrum mirabile Mont. Une Sap Geastrum rufescens Pers. Une Sap Geastrum sp.1 Une Sap
Family Gomphaceae
Phaeoclavulina cyanocephala (Berk. & M.A. Curtis) Giachini Edi Ect Ramaria sp.1 Une Sap Ramaria sp.2 Une Sap Ramaria sp.3 Une Ect
Order Hymenochaetales
Family Hymenochaetaceae
Coltricia cinnamomea (Jacq.) Murrill Ine Sap Hymenochaete rubiginosa (Dicks.) Lév. Une Sap Hymenochaete sp.1 Une Sap Hymenochaete sp.2 Une Par Hymenochaete sp.3 Une Par Hymenochaete sp.4 Une Sap Hymenochaete sp.5 Une Sap Unidentified 6 Une Sap Unidentified 7 Une Par Unidentified 8 Une Par
Order Phallales
Family Phallaceae
Phallus indusiatus Vent. Edi Sap Unidentified 9 Une Sap
Order Polyporales
Family Fomitopsidaceae
Daedalea sp.1 Une Sap Daedalea sp.2 Une Sap

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
30
30
Scientific Name EI ER Family Ganodermataceae
Amauroderma rude (Berk.) Torrend Edi/Med Sap Amauroderma rugosum (Blume & T. Nees) Torrend Edi Sap Ganoderma applanatum (Pers.) Pat. Edi Sap Ganoderma sp.1 Une Sap Ganoderma sp.2 Une Sap Ganoderma sp.3 Une Sap Ganoderma sp.4 Une Sap
Family Meruliaceae
Cymatoderma elegans Jungh. Ine Sap Podoscypha sp.1 Une Sap
Family Phanerochaetaceae
Byssomerulius corium (Pers.) Parmasto Une Sap Family Polyporaceae
Cerioporus mollis (Sommerf.) Zmitr. & Kovalenko Une Sap Hexagonia tenuis (Fr.) Fr. Une Sap Lentinus arcularius (Batsch) Zmitr. Une Sap Lentinus fasciatus Berk. Une Sap Lentinus squarrosulus Mont. Edi/Med Sap Microporus xanthopus (Fr.) Kuntze Ine Sap Nigrofomes sp.1 Ine Sap Picipes badius (Pers.) Zmitr. & Kovalenko Une Sap Polyporus grammocephalus Berk. Une Sap Polyporus retirugus (Bres.) Ryvarden Une Sap Polyporus sp.1 Une Sap Polyporus sp.2 Une Sap Polyporus sp.3 Une Sap Polyporus sp.4 Une Sap Polyporus sp.5 Une Sap Polyporus sp.6 Une Sap Pycnoporus sanguineus (L.) Murrill Ine Sap Trametes cingulata Berk. Ine Sap Trametes versicolor (L.) Lloyd Edi/Med Sap Trametes sp.1 Une Sap Trametes sp.2 Une Sap Trametes sp.3 Une Sap Trametes sp.4 Une Sap Trametes sp.5 Une Sap Unidentified 10 Une Sap
Unidentified 11 Une Sap Unidentified 12 Une Sap
Order Russulales
Family Peniophoraceae
Peniophora sp.1 Une Sap Peniophora sp.2 Une Sap
Family Russulaceae
Lactarius piperatus (L.) Pers. Edi/Med Ect Lactarius sp.1 Une Ect

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 31
Scientific Name EI ER Lactarius sp.2 Une Ect Lactarius sp.3 Une Ect Lactarius sp.4 Une Ect Russula alboareolata Hongo Edi/Med Ect Russula cyanoxantha (Schaeff.) Fr. Edi Ect Russula delica Fr. Edi/Med Ect Russula densifolia Secr. ex Gillet Edi Ect Russula nigricans Fr. Edi Ect Russula rosea Pers. Une Ect Russula violeipes Quél. Edi Ect Russula sp.1 Une Ect Russula sp.2 Une Ect Russula sp.3 Une Ect Russula sp.4 Une Ect Russula sp.5 Une Ect Russula sp.6 Une Ect Russula sp.7 Une Ect Russula sp.8 Une Ect Russula sp.9 Une Ect
Family Stereaceae
Stereum ostrea (Blume & T. Nees) Fr. Ine Sap Stereum sp.1 Une Sap
Order Stereopsidales
Family Stereopsidaceae
Stereopsis sp.1 Une Sap Order Thelephorales
Family Thelephoraceae
Thelephora vialis Schwein. Une Ect
Unidentified 13 Une Unk Unidentified 14 Une Sap Unidentified 15 Une Sap Unidentified 16 Une Sap Unidentified 17 Une Unk Unidentified 18 Une Sap Unidentified 19 Une Unk Unidentified 20 Une Par Unidentified 21 Une Par Unidentified 22 Une Sap
Class Dacrymycetes
Order Dacrymycetales
Family Dacrymycetaceae
Calocera viscosa (Pers.) Fr. Une Sap Calocera sp.1 Une Sap Dacrymyces minor Peck Une Sap
Class Tremellomycetes
Order Tremellales
Family Tremellaceae

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand
32
32
Scientific Name EI ER Tremella mesenterica Retz. Edi/Med Sap Tremella sp.1 Une Sap
Remark: Abbreviations of edibility information (EI): Edi = edible mushroom; Ine = inedible mushroom;
Med = medicinal mushroom and Une = unknown edibility and abbreviations of ecological roles (ER):
Ect = ectomycorrhizal mushroom; Sap = saprophytic mushroom; Ter = termite mushroom and
Unk = unknown ecological role

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 33
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 33 - 40
Health benefit screening on 45 Thai human probiotics (TISTR strains): their safety and
immunomodulatory activity in macrophage function
Bhusita Wannissorn1*, Weerasak Taengphan2, Prapaipat Klungsupya2, Supatjaree Ruengsomwong1
and Thanchanok Muangman2
1Biodiversity Research Centre (BRC), Thailand Institute of Scientific and Technological Research (TISTR),
Techno Polis, Khlong Luang District, Pathum Thani, Thailand 2Expert Centre of Innovative Herbal Products (InnoHerb), Thailand Institute of Scientific and Technological
Research (TISTR), Techno Polis, Khlong Luang District, Pathum Thani, Thailand
*Corresponding author e-mail: [email protected]
Abstract: A total of 45 human probiotic strains (namely Med 1 to Med 45) from the probiotic bank of Thailand
Institute of Scientific and Technological Research (TISTR) were analyzed for their cytotoxic and phagocytic
activities on macrophage RAW 264.7 cells. Their heat killed supernatants were prepared from 48 hr-culture
anaerobically grown in de Man Rogosa and Sharpe (MRS) broth supplemented with 0.05% L-cystiene and were
tested without dilutions (at 100% concentration). Results obtained from the water-soluble tetrazolium salt (WST-1)
assay suggested that 27 out of 45 strains were non-cytotoxic to macrophage RAW 264.7 cells. Among them, 10
strains clearly demonstrated a cell-proliferation promoting activity with cell survival rate greater than 100%
compared with the untreated cells. The phagocytic activity of all 45 TISTR probiotics were determined by their
capability on up-taking phenol red particles which acted as foreign bodies. The results revealed that 16 out of 45
TISTR probiotic strains possessed positive phagocytosis in macrophage RAW 264.7 cells at various degrees.
A significant increase in percentage of phagocytosis (% phagocytic activity) in RAW 264.7 cells induced by these
16 TISTR probiotic strains was found in a range of 2.58 to 39.89% in comparison to control (0% phagocytic
activity, untreated cells) and β-glucan (64.43% phagocytic activity, positive control). Among 45 TISTR probiotic
strains, Med 1, Med 31, Med 39 and Med 42 are considered to be the most active strains regarding to their safety
and phagocytic activities on macrophage RAW 264.7 cells when tested under conditions described in this study.
Keywords: Cytotoxicity, immunomodulation, macrophages RAW 264.7, phagocytosis, probiotics
Introduction
Due to their beneficial effects on health, probiotics, live microorganisms conferring health benefit to the
host when administered in sufficient amount (FAO and WHO, 2011), have been widely applied as ingredients in
functional foods and dietary supplements. The Markets and Markets reported that probiotics market is estimated
to grow at USD 49.4 billion in 2018 and is projected to grow at a compound annual growth rate (CAGR) of 7.0%
from 2018, to reach a value of USD 69.3 billion by 2023 due to highly rising health awareness among consumers
worldwide. Apart from the abilities in enhancement intestinal barrier, preventing of pathogens via acidification,
production of antimicrobial substances and competitive exclusion, stimulation of immune system is considered as
one of the probiotics’ mechanisms which in turn generating health benefits to the host (Kumar Bajaj et al., 2015).
However, these mechanisms are strain-specific (Collado et al., 2012; Lee et al., 2016), not all probiotics can show
equally effectiveness. Hence, screening of a large number of candidate probiotics in order to obtain the effective
strains are still required.
Regarding the Innovative Center for Production of Industrially Used Microorganisms (ICPIM), Thailand
Institute of Scientific and Technological Research (TISTR), we have set up the probiotic bank with the aim to
collect and conserve local probiotics for commercial uses. Extensive isolation of effective probiotics has been
done from various sample sources in Thailand. Recently, 45 strains have been isolated from flowers and fruits
locally grown in the northern part of Thailand. All isolates were selected for probiotic properties based on
notification of Ministry of Public Health (2011) such as non-hemolytic activity, sensitivity to antibiotic, tolerance
to gastric- and intestinal juices, adhesion to human epithelial cell lines and antimicrobial activity against certain
intestinal pathogens. In this study, we further characterized immunomodulatory effects of these 45 isolates on
cytokine profiles using macrophage RAW 264.7 cell line.
Materials and Methods
Probiotics and growth conditions
Forty five strains of TISTR human probiotics (namely Med 1 to Med 45) from the probiotic bank of
Culture Collection of Microorganism Centre, TISTR were anaerobically grown in de Man Rogosa and Sharpe
(MRS) broth supplemented with 0.05% L-cysteine for 48 hr. Each 48 hr grown culture was adjusted with normal
saline to obtain the final concentration of 108 CFU/ml. Then, cell pellets of each culture were collected by centrifugation

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 34
for 5 min at 8,500 rpm at 25°C. After 3 times washing with normal saline, the washed pellets were suspended in
Dulbecco's Modified Eagle Medium (DMEM) high glucose (Biowest, USA) without antibiotics before being
heated at 110 °C for 10 min. The 45 heat killed supernatants were tested without dilutions (at 100% concentration).
Macrophages culture
The mouse monocyte macrophage cell line, RAW 264.7 (ATCC TIB-71TM) was purchased from American
Type Culture Collection (ATCC) (Manassas, VA, USA). Cells were grown as adherent in Dulbecco’s Modified
Eagle Medium (DMEM) (Biowest, USA) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS)
(GIBCO®, USA) and 1% (v/v) penicillin-streptomycin (GIBCO®, USA) in humidified atmosphere incubator with
5% CO2 at 37°C. The cells were collected by centrifugation (Kobota, Japan) at 1500 rpm, 4°C for 5 min, aspirated
and fresh culture medium at similar volume was added before dispensing into new culture flasks. All steps of
culturing and maintaining of RAW 264.7 cells were conducted in aseptic condition using the Biological Safety
Cabinet Class II (SANYO, Japan). For experimentation, cells were seeded onto 96-well plates at a density of
2x104 cells/well and incubated for 24 hrs before treatments.
Cytotoxicity analysis by WST-1 assay
The cytotoxic property of 45 TISTR human probiotics (heat killed supernatants) on macrophage RAW
264.7 cells were determined using 4-[3-(4-Iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio]-1, 3-benzene disulfonate
(WST-1) assay. Its principle is based upon the conversion of tetrazolium salt to soluble formazan crystals by
succinate dehydrogenase in the mitochondria of metabolically active cells. Therefore, in the dead cells this reaction
will not be occurred. After incubation, the probiotic supernatants were removed and washed once with 10 µl of
PBS and then 100 µl of WST-1 solution (Sigma, USA) was added to each well. The plates were incubated in
darkness with 5% CO2 in a humidified atmosphere incubator at 37°C for 30 min. The cell viability was determined
by the optical density (OD) at the wavelength of 450 nm with microplate reader (Tecan Infinite 200 Pro, Austria).
Mitomycin C (MMC) solution (100 µl) at 20 µg/ml prepared in DMEM was used as positive control. The percentage
viability of cells related to control wells containing cell culture medium (DMEM) without samples was calculated
as following equation:
% Cell viability = (Absorbance of treated cells / Absorbance of treated cells) x 100
Phagocytic activity by neutral red uptake assay
The phagocytosis ability of macrophages was measured by colorimetric neutral red uptake assay. The assay
was performed to determine phagocytosis ability of macrophage RAW 264.7 cells when treated with heat killed
supernatants of 45 TISTR human probiotics. The principle of this assay is based upon membrane permeability
and lysosomal activity of viable cells in response to pharmaceuticals, chemicals and environmental compounds
as well as nutrients. In this study, RAW 264.7 cells were treated (at 200 µl total volume) with 100% concentration
of heat killed supernatants of 45 TISTR human probiotics or or β-glucan (Sigma, USA) at 100 µg/ml for 24 hr.
By the end of incubation time, the treatments were removed and cells were washed 3 times with PBS followed by
adding of 100 µl of 0.075% neutral red solution (Sigma, USA) to each well. The reactions were kept at 37C for
60 min. The supernatant was removed and 100 µg/ml cell lysis solution (ethanol and 0.01% acetic acid at the ratio
of 1:1) was added to lysed cells at 37C for 120 min. The quality of dye incorporated into RAW 264.7 cells in
measured by spectrophotometry at 540 nm wavelength. The percentage of phagocytosis activity was calculated
as following equation:
% Phagocytic activity = Absorbance of sample – Absorbance of control x 100
Absorbance of control
Statistical analysis
All experimental data were expressed as mean ± standard error (Mean ± SE) of triplicated determination.
The significance of difference was used to compare mean (p<0.05). All analyses were performed using One-way
ANOVA and Tukey’s Honestly Significant Difference test.
Result & Discussion
Effects of probiotics on macrophage cell viability
MMC is a chemotherapeutic drug that has been used in cancers treatment. It works by intercalating the DNA
together. Hence, DNA cannot come apart during cell division resulting in no cells division and apoptosis. Damage
to nuclear DNA is thought to be its primary mechanism of cell death in MMC action (Snodgrass et al., 2013).
In this study, MMC (20 µg/ml concentration) was used as a positive control for cell proliferation test using the
WST-1 assay. After RAW 264.7 cells were treated with heat killed supernatants probiotics at 100 % concentration
and assessed by WST-1 assay, all probiotics exhibited the % cell viability as illustrated in Table1. Morphologies
of cells following treatments were demonstrated in Figure 1. The results were expressed as % cell inhibition

Proceedings of International Conference on Biodiversity: IBD2019
on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 35
compared with the control cells. It was found that most TISTR probiotic strains (27 out of 45 strains) showed no
significant cytotoxic activity against RAW 264.7 cells. Among them, 9 strains including Med1, Med2, Med3,
Med4, Med5, Med7, Med8, Med10 and Med11 clearly demonstrated a cell-proliferation promoting activity with
cell survival rate greater than 100% compared with the untreated cells. However, the highest cytotoxic effect
against RAW 264.7 cells was observed in 6 strains (Med23, Med27, Med34, Med35, Med44, Med45) with cell
growth inhibition greater than 90% (cell survival rate less than 10%). RAW 264.7 cells treated with MMC
exhibited a marked decrease in cell survival to 17.04 ± 0.43 % (Table 1.)
Phagocytic activity of probiotics in macrophage function
It has been well-known that probiotics promote various health benefits (Sanders, 2008). They can inhibit
overgrowth of pathogenic bacteria by antibacterial substance production and stimulating the immune system. They
have been demonstrated that the effects on the variety of immune cells such as phagocytes (macrophages and
neutrophils), lymphocytes and natural killer (NK) cells. The neutral red uptake assay is a method to detect cell
viability or lysosome activity assay based on the ability of viable cells to incorporate and bind neutral red within
lysosomes (Guan et al., 2011). It is generally performed on adherent cells. The quantity of dye incorporated into
cells is measured by spectrometry at 540 nm, and is directly proportional to the number of cells with an intact
membrane. In this study, stimulation of phagocytic function by 45 TISTR probiotics in macrophage RAW 264.7
cells was investigated by phenol red assay meaning the amount of neutral red internalized in phagosomes (Figure1).
β-glucans are naturally occurring polysaccharides that are produced by bacteria, yeast, fungi and many plants
especially mushrooms (Han et al., 2008). β-glucan is recognized by the dectin-1 and C3 receptors on macrophages
that triggers phagocytosis (Goodridge et al., 2009). We selected β-glucan (at 100 µg/ml concentration) and used
as a positive control in this study. As demonstrated in Table 2, the results from phenol red uptake assay revealed
that 16 out of 45 TISTR probiotic strains possessed positive phagocytosis in macrophage RAW 264.7 cells. An
increase in percentage of phagocytosis (% phagocytic activity) in RAW 264.7 cells induced by these 16 TISTR
probiotic strains (Med1, Med2, Med3, Med4, Med6, Med10, Med11, Med14, Med16, Med30, Med31, Med36,
Med39, Med40, Med41, Med42) was found in a range of 2.58 to 39.89% in comparison to control (0% phagocytic
activity, untreated cells) and β-glucan (64.43% phagocytic activity, positive control) (Table 2). Though we found
out that these TISTR probiotics were effective on activation of macrophages function (phagocytosis), their
mechanisms are still unknown. They may or may not function like β -glucans by binding to dectin-1 receptor on
macrophages and lead to activation of macrophages to engulf pathogens through phagocytosis and digestion them
with lysosomal enzymes. Further studies on this matter are required.
Conclusion
Among 45 heat-killed TISTR probiotic strains, Med 1, Med 31, Med 39 and Med 42 are considered to
be the most active strains regarding to their safety and phagocytic activities on macrophage RAW 264.7 cells.
These results suggest that administration of these four TISTR probiotic strains may result in an initiation of
immune reaction against foreign materials. Therefore, they can be employed as a natural resource for development
to healthy dietary supplement such as functional food products.
Acknowledgement
This work was performed by Thailand Institute of Scientific and Technological Research (TISTR) under
financial support by a grant from the Ministry of Science and Technological (MOST) during 2018-2020.
References
Collado, C., Bauerl, C., and Perez-Martinez, G. 2012. Defining the microbiota for developing new probiotics.
Microb Ecol Health Dis 23(1):35-39.
Food and Agriculture Organization of the United Nations (FAO) and World Health Organization (WHO).2004.
Health and nutritional properties of probiotics in food including powder milk with live lactic acid
bacteria. FAO and WHO Joint and Expert Committee Report.
Goodridge, S., Underhill, D., and N. Touret. 2012. Mechanisms of fc receptor and dectin-1 activation for phagocytosis.
Traffic. 13 (8):1062-1071.
Guan, D., Zhang, Z., Yingnan, Y., Gouqing, X., and Jinqiang, L. 2011. Immunomodulatory activity of polysaccharide
form the roots of Actinidia kolomikta on macrophage. Int J Biol. 3 (2): 3-10.
Han, M., Han, Y., Hyun, S., and Shin, H. 2008.Solubilization of water-insoluble β-glucan isolated from
Ganoderma lucidum. J Env Biol. 29 (2): 234-242.
Kumar Bajaj, B., Claes, I., and Lebeer, S. 2015. Functional mechanisms of probiotics. J Microbiol Biotech Food
Sci. 4 (4): 321-327.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 36
Lee, C., Van, G., Taverne, N., Meijerink, M., Bron, A., Spano, G., and Kleerebezem, M. 2016. Strain-specific
features of extracellular.
polysaccharides and their impact on Lactobacillus plantarum-host interactions. Appl Environ Microbiol 82
(13):3959–3970.
Sanders, M. 2008. Probiotics: Definition, Sources, Selection, and Uses. Clin Infec Dis. 46 (2): 58-61.
Snodgrass, G., Collier, A., Coon, A., and Pritsos, A. 2010. Mitomycin C inhibits ribosomal RNA: A novel
cytotoxic mechanism for bioreductive drug. Cold Spring Harb. Perspect. Biol.5:1-16.

Proceedings of International Conference on Biodiversity: IBD2019
on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 37
Table 1. Cytotoxicity determination of 45 TISTR human probiotics against RAW 264.7 cells by WST-1 assay.
% Cell viability
range Probiotic stains
Cytotoxicity
(% cell viability)
80-100
Med1 102.60 ± 3.18a
Med2 109.92 ± 6.08a
Med3 110.85 ± 6.70a
Med4 108.98 ± 6.89a
Med5 103.98 ± 8.30a
Med6 99.00± 4.67a
Med7 104.59 ± 14.13a
Med8 111.58 ± 1.65a
Med10 117.74 ± 4.05a
Med11 108.34 ± 5.55a
Med14 87.40 ± 0.24a
Med16 91.48 ± 10.04a
Med30 84.90 ± 15.91a
Med31 89.94 ± 2.90a
Med39 86.07± 1.65a
Med41 82.62 ± 4.52a
Med42 85.73 ± 6.91a
60-79
Med9 79.11 ± 5.01a
Med17 68.13 ± 2.09c
Med19 65.15 ± 5.19c
Med21 62.13 ± 8.60c
Med36 77.21 ± 3.17a
Med38 69.50 ± 3.30c
Med40 73.26 ± 8.64c
40-59
Med18 49.30 ± 9.78d
Med24 51.66 ± 7.90d
Med26 48.39 ± 1.60d
Med28 58.66 ± 17.06c
20-39
Med13 28.62 ± 4.54c
Med22 39.73 ± 1.66d
Med25 30.40 ± 8.48d
Med32 30.06± 5.39d
0-19
Med12 10.41 ± 1.16b
Med15 10.40 ± 3.64b
Med20 14.06 ± 10.28b
Med23 8.18 ± 0.35b
Med27 9.95 ± 1.53b
Med29 16.09 ± 13.21b
Med33 11.90 ± 4.75b
Med34 9.14 ± 1.81b
Med35 9.00 ± 0.75b
Med37 15.59 ± 6.00b
Med43 11.90 ± 6.33b
Med44 9.33 ± 1.71b
Med45 9.82 ± 1.97b
Positive control MMC (20 µg/ml) 17.04 ± 0.43b
* Each value is mean ± SD (n=3). One way ANOVA and Tukey’s Honestly Significant Difference (P<0.05).
Difference later in the same row are significantly different.
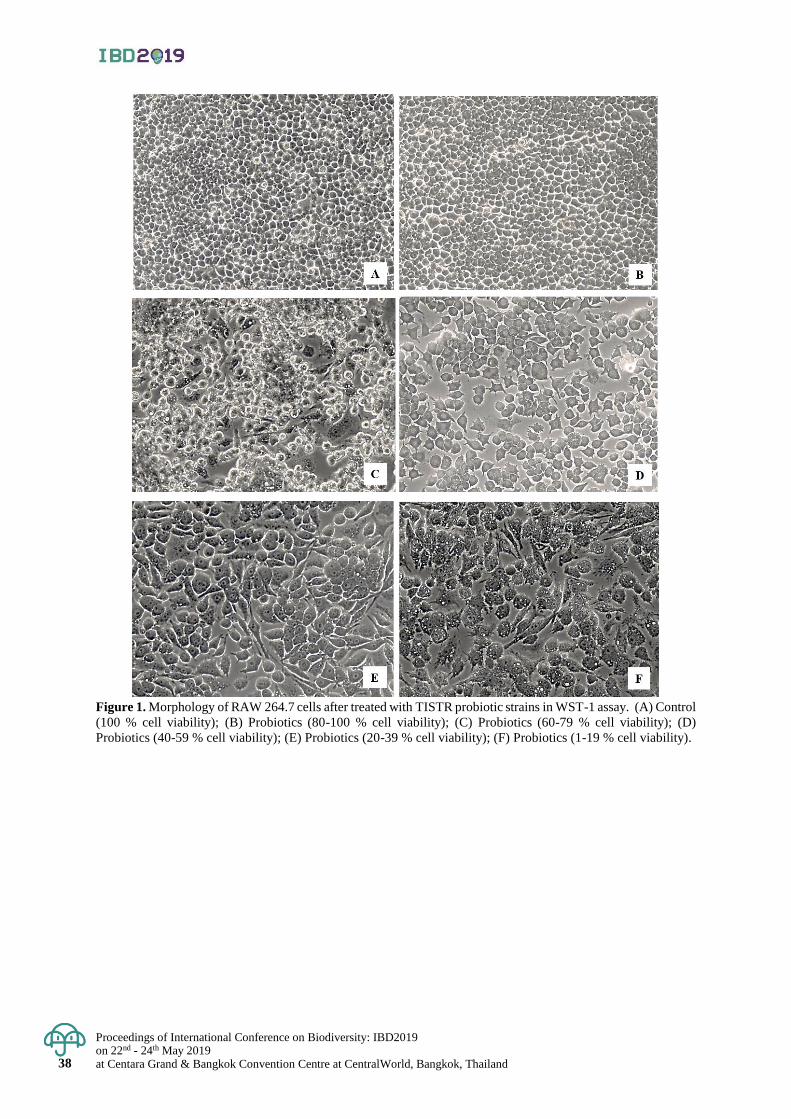
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 38
Figure 1. Morphology of RAW 264.7 cells after treated with TISTR probiotic strains in WST-1 assay. (A) Control
(100 % cell viability); (B) Probiotics (80-100 % cell viability); (C) Probiotics (60-79 % cell viability); (D)
Probiotics (40-59 % cell viability); (E) Probiotics (20-39 % cell viability); (F) Probiotics (1-19 % cell viability).
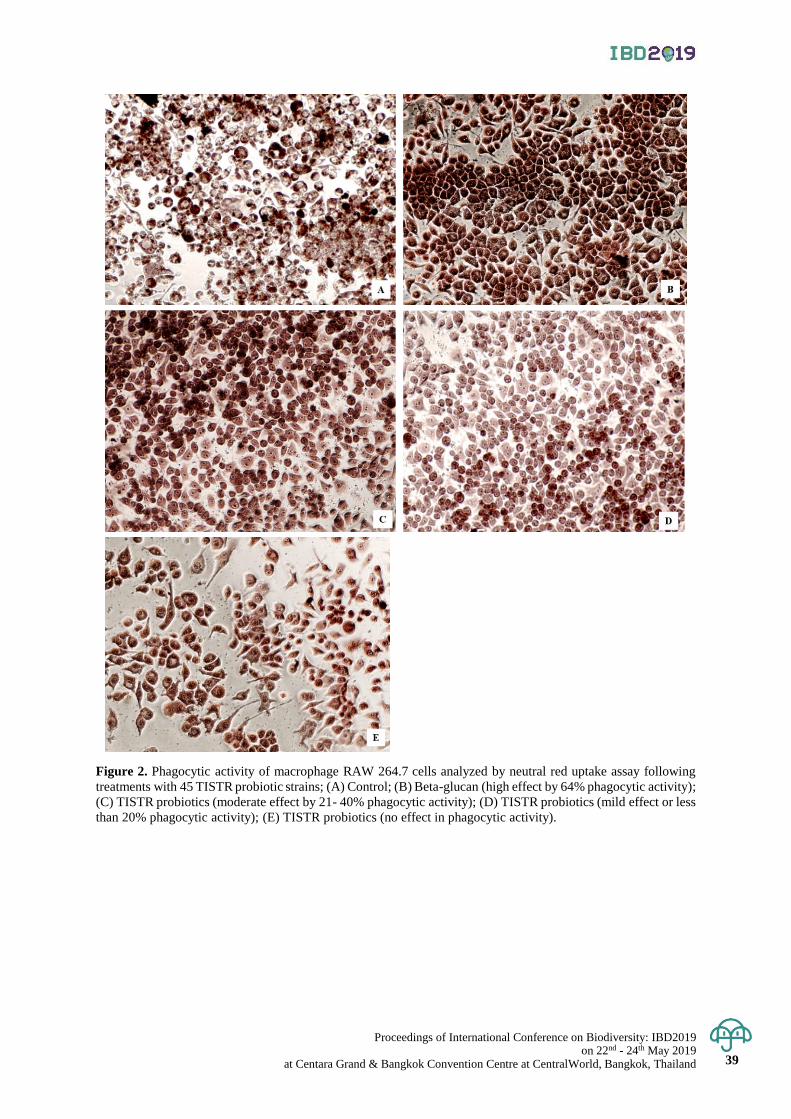
Proceedings of International Conference on Biodiversity: IBD2019
on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 39
Figure 2. Phagocytic activity of macrophage RAW 264.7 cells analyzed by neutral red uptake assay following
treatments with 45 TISTR probiotic strains; (A) Control; (B) Beta-glucan (high effect by 64% phagocytic activity);
(C) TISTR probiotics (moderate effect by 21- 40% phagocytic activity); (D) TISTR probiotics (mild effect or less
than 20% phagocytic activity); (E) TISTR probiotics (no effect in phagocytic activity).
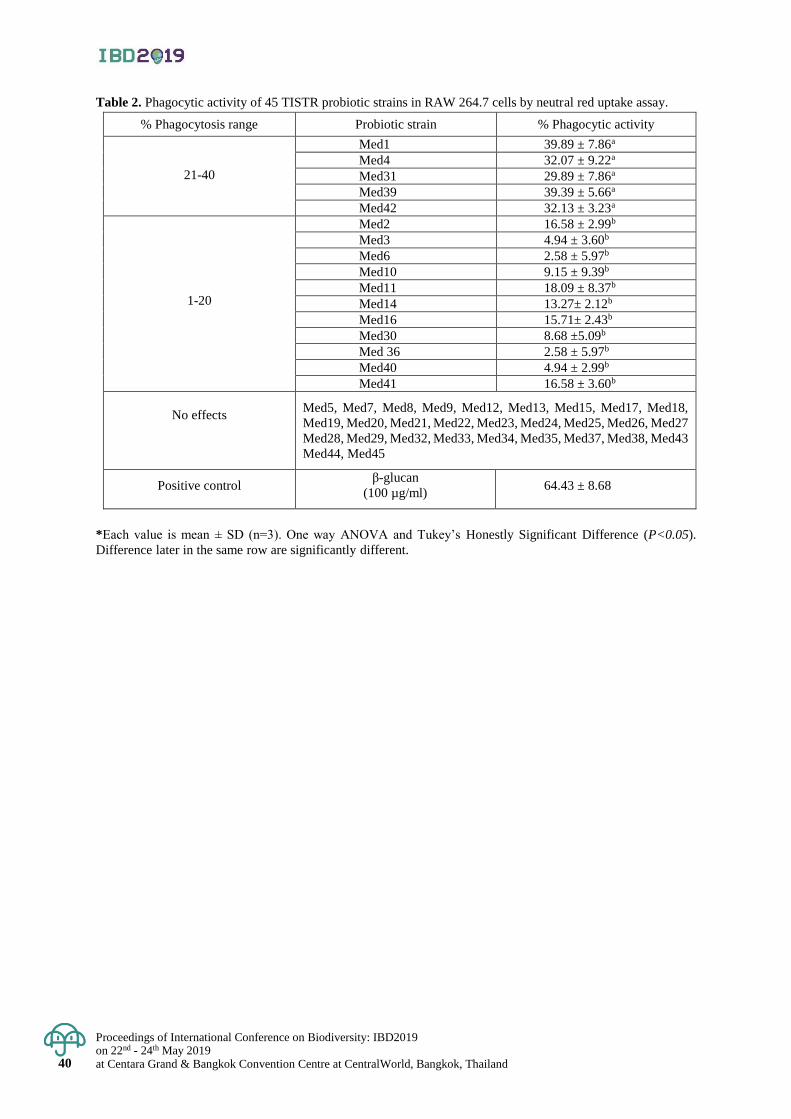
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 40
Table 2. Phagocytic activity of 45 TISTR probiotic strains in RAW 264.7 cells by neutral red uptake assay.
% Phagocytosis range Probiotic strain % Phagocytic activity
21-40
Med1 39.89 ± 7.86a
Med4 32.07 ± 9.22a
Med31 29.89 ± 7.86a
Med39 39.39 ± 5.66a
Med42 32.13 ± 3.23a
1-20
Med2 16.58 ± 2.99b
Med3 4.94 ± 3.60b
Med6 2.58 ± 5.97b
Med10 9.15 ± 9.39b
Med11 18.09 ± 8.37b
Med14 13.27± 2.12b
Med16 15.71± 2.43b
Med30 8.68 ±5.09b
Med 36 2.58 ± 5.97b
Med40 4.94 ± 2.99b
Med41 16.58 ± 3.60b
No effects Med5, Med7, Med8, Med9, Med12, Med13, Med15, Med17, Med18,
Med19, Med20, Med21, Med22, Med23, Med24, Med25, Med26, Med27
Med28, Med29, Med32, Med33, Med34, Med35, Med37, Med38, Med43
Med44, Med45
Positive control β-glucan
(100 µg/ml) 64.43 ± 8.68
*Each value is mean ± SD (n=3). One way ANOVA and Tukey’s Honestly Significant Difference (P<0.05).
Difference later in the same row are significantly different.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 41
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 41 - 47
Records of potential antimicrobial activity of soil actinomycetes isolated from
a community forest in Ban Khoklam Sang Aram, Udon Thani Province
Metinee Wasoontharawat 1* and Panthong Kulsantiwong 2 1Department of Biology, Faculty of Science, Udon Thani Rajabhat University, Muang District, Udon Thani, Thailand 2Department of Biology, Faculty of Science, Udon Thani Rajabhat University, Muang District, Udon Thani, Thailand
*Corresponding author e-mail: [email protected]
Abstract: The WHO (2015) released a report of an increasing number of infections due to the global antimicrobial
drug resistance crisis. Unexplored areas may be possible for sources of actinomycetes with potent antimicrobial
compounds. This study was directed towards isolation of actinomycetes that produce antimicrobial substances
from unexplored soil in a community forest in Ban Khoklam Sang Aram, Udon Thani Province. The survey
showed diversity of actinomycetes and rare actinomycetes in this area. Ten of 232 isolated actinomycetes showed
potent antimicrobial activity by growth inhibition of Bacillus subtillis, Candida albicans, Escherichia coli,
Pseudomonas aeroginosa and Staphylococcus spp. after testing using agar disc diffusion and agar well diffusion
tests. These isolates were identified as species in the genera Streptomyces (9 isolates) and Microbispora (1 isolate).
Keywords: Microbial diversity, actinomycetes, Antimicrobial activity, Soil microorganism, Pathogenic microorganism
Introduction
Actinomycetes are Gram-positive bacteria. They are unique in their formation of branching filaments with
fungi like hyphae and asexual spores. Their bacterial DNA has a high G+C content, 55-75% (Lechevalier and
Lechevalier, 1967; Embley and Stackebrandt, 1994). Actinomycetes is a major microbial population that is widely
distributed and inhabits the soil (Jaralla et al., 2014). They play role in soil biodegradation and humus formation
(McCathy and Williams, 1992; Agadagba, 2014). Moreover, many actinomycetes are pharmaceutically important
for production of secondary metabolites as antibiotics, vitamins and enzymes (Shahidi et al., 2004; Xu et al., 2005;
Arifuzzaman et al., 2010). Soil actinomycetes produce many of currently used antibiotics including Erythromycin,
Gentamycin, Rifamycin and Streptomycin (Jeffrey, 2008). Therefore actinomycetes are potentially important
strains for producing new bioactive compounds, especially antibiotics that have diverse clinical effects and have
activity against many pathogens (Atta and Ahamd, 2009; Usha et al., 2011).
This work was aimed to investigate diversity of an actinomycetes population in the soil of a community
forest in Ban Khoklam Sang Aram, Udon Thani Province and their antimicrobial activities, for the first time.
This forest has not yet been studied. Additionally, this area yields Streptomyces species and rare actinomycetes
strains with high antimicrobial activity.
Materials and Methods
Collection of soil samples
Seven soil samples were randomly collected from various sites in a community forest in Ban Khoklam
Sang Aram, Udon Thani Province. The seventh sites where the collected soil sample were takes were diverse.
They include 1) loam with a leaf covering, 2) loam with a decayed wood covering, 3) fine sand with no vegetation
and a leaf covering that served as drainage for rainfall and 4) sandy loam surrounding plant roots. The soil was
taken at a depth of 1-10 cm from the soil surface. Approximately 500 grams of each soil sample was collected
and was placed in sterile polyethylene bags and kept at room temperature. These were promptly transferred to a
laboratory for analysis of their actinomycetes population.
Preparation of soil samples
The soil samples were air-dried at room temperature until a constant weight was attained and were stored
at 4C prior to processing for actinomycetes isolation. They were pretreated to stimulate actinomycetes growth
and eliminate unwanted Gram-negative bacteria. Each soil sample divided into two parts for separate pretreatment
methods. One part was pretreated with wet heat and the other was pretreated with air dry plus calcium carbonate
(CaCO3) following the methodology of Fang et al. (2017). Then the samples were further examined for diversity
of actinomycetes colonies and actinomycetes isolates.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 42
Enumeration isolation and purification of actinomycetes
Actinomycetes in each soil were isolated using a 10-fold serial dilution of soil and spread plate technique.
Diluted soil solution was spread on a humic acid (HA) medium (Hayakawa and Nonomura, 1987) and starch-casein
agar (SCA) (Okazaki et al., 1983) supplement with 1 mg/mL of nalidixic acid and 1 mg/mL of cycloheximide. All
spread plates were incubated at 37C for a period of 14 days.
A number of diverse morphological colonies appeared in each dilution on both the HA and SCA media on
day 14. The various colony morphologies were carefully counted and recorded. Then, the density of the actinomycetes
population was determined in units of CFU/g dried soil as the number colonies on a plate times the dilution factor.
Various colonies on HA and SCA media were then picked with sterile needles and transferred to an
International Streptomyces Project 2 (ISP2) medium. Each isolate was purified using a streak plate technique and
cultured at 37C for a period of 3-5 days until pure single colonies appeared.
Preliminary Screening for antimicrobial activity
Preliminary screening of the antimicrobial activity of purified isolates was done using a streak plate technique
following Lemons et al. (1985). One isolate was long single streak at the corner of ISP2 solid medium plate. Four
pathogenic strains were used to investigate the antagonistic activity of actinomycetes isolates. They were Bacillus subtilis,
Staphylococcus aureus, Pseudomonas aeruginosa and Candida albicans. These pathogenic microorganisms were
suspended (0.5 McFarland) and streaked perpendicular to the actinomycetes strain using a sterilized loop. The plate was
then incubated at 37C for a period of 14 days. Clear zones, indicating microbial inhibition, and their widths were determined
in millimeters.
Secondary screening for antimicrobial activity
Each purified isolate expressing growth inhibition in the preliminary screening was cultured in 100 ml ISP2 broth
on a 250 ml rotary shaker (180 revolutions per minute, rpm) at 28+2°C for 4 days. They were then transferred to
a new YEME broth supplemented with 0.1% CaCO3, as a production medium. The cultures were incubated a rotary
shaker (180 rpm) at room temperature for 14 days. The culture broth was filtered through Whatman No.1 filter paper
to separate the mycelium. A clear supernatants were obtained that were extracted three times with ethyl acetate at a 1:1
(v/v) ratio with vigorous shaking for 1 hour according the method of Rao et al. (2017). The bioactive compounds were in
the ethyl acetate phase that separated from the aqueous phase. The ethyl acetate phase was evaporated under a reduced
vacuum at 80C. The obtained bioactive crude extract was used to test the antimicrobial activity of isolates using agar disc
diffusion and agar well diffusion tests.
Secondary screening for antimicrobial activity
The antimicrobial activity of purified isolates expressing growth inhibition in the preliminary screening were
confirmed by agar disc diffusion and agar well diffusion methods as described by Mohseni et al. (2013) and Omar et al.
(2000), respectively. Nystatin and nalidixic acid agents were used as controls for anti-Gram positive and anti-Gram
negative bacterial activity, respectively.
Morphological identification of actinomycetes
Preliminary identification of actinomycetes isolates grouped the bacterial as Streptomyces and non- Streptomyces,
(rare actinomycetes). It was performed by observing the growth rates of bacteria on an International Streptomyces
Project (ISP) medium (Shirling and Gottli, 1966). Streptomyces grew faster than non- Streptomyces, as was also
reported by Abdullah et al. (2016).
Isolates were identified at the genus level based on microscopic and macroscopic studies of actinomycetes
following Shirling and Gottileb (1966), Muthu et al. (2013) and Barka et al. (2015). Macroscopic examination
considered colony features such as size, shape, color, absence or presence of aerial mycelium and spore formation.
Microscopic characteristics observed were aerial and substrate mycelium, conidia, spore type, and especially spore chains.
Result & Discussion
Enumeration and isolation of actinomycetes
Actinomycetes in soil samples were isolated using HA medium and SCA medium supplement with
nalidixic acid (as antifungal agent) and cycloheximide (as antibacterial agent) at 37C for 7-14 days. The diversity
of the actinomycetes population was viewed from various types of colony morphology, size, color, elevation, surface
and pigmentation, as they grew on a solid medium.
The results indicated a high diversity of actinomycetes from soil samples collected from a community
forest in Ban Khoklam Sang Aram, Udon Thani Province, on both media. This data is shown in Figure 1.
The various soil samples had actinomycetes populatons in the range of 1.1 x 102 - 5.3 x 102 CFU/g dried soil on
HA plates and 2.1 x 102 - 9.4 x 102 CFU/g dried soil on SCA plates. The various colonies had obviously different
colony characteristic on the media. The highest numbers of colonies with different morphology were found on the
SCA medium as it supported significantly more growth than the HA medium. Most actinomycetes isolates grew

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 43
better when hydrolyzing starch and casein than the humid acid in the HA medium (Ravel et al., 2000). The types
of colonies grown on SCA medium are shown in Figure 2.
The most populated sample had 9.4 x 102 CFU/g dried soil on SCA medium, from the fourth site in the
forest. Its soil was a fine sand without vegetation and the soil was saturated with water. This may have been due
to a recent rainfall. Furthermore, the soil might contain soil nutrients, such as organic carbon, that originated from
other sites. According to Stackebrandt et al. (1991) and Baby et al. (2002), fertile soils or soil containing organic
carbon, nitrogen, and phosphorus is an important soil feature that can enhance the grow the rate and diversity of
an actinomycetes population. Alternatively, the highest number of diverse colonies on HA medium was 5.3 x 102
CFU/g dried soil from soil at the seventh site. This soil was a loam with a decayed wood covering. The colony
morphology from soil taken at this site was different than from other soil sources. So, it was of interest to
investigate these bacteria. Actinomycetes isolates have been found on decomposing wood since some actinomycetes
can produce hydrolytic enzymes to break down the lignocellulose of wood (Bontemps et al., 2013; Ding et al.,
2004). The lowest number of colonies on both media was 2.1 x 102 CFU/g dried soil on SCA medium from soil
at third site, as fine sand without vegetation. It was in an area where there was water flow due to rainwater runoff.
A total of 232 actinomycetes were isolated from seven soil samples collected at various locations within
this community forest.
Primary screening for antimicrobial activity All actinomycetes isolates underwent a preliminary screening for antimicrobial activity using an agar cross
streak method. It found that ten isolates possess inhibitory activity against pathogenic fungi and bacteria. These
were the 8.2.4HV, 8.2.5HV, 6.1HV-1, 20.2.6, 15.1, RH13-26, 11.1.2, 10. 4.1, 10.5, and RH12-1 isolates.
Secondary screening for antimicrobial activity
The ten isolates were further tested to confirm their antimicrobial activity using a crude ethyl acetate extract
with disc diffusion and well diffusion methods. Their antimicrobial activity was measured in terms of a clear zone
that formed. As shown in Table 1, many isolates were active against more than one of the tested pathogenic
microorganisms. Bacillus subtillis growth was inhibited by eight isolates. These were 6.1HV-1, 20.2.6, 15.1, 11.1.2,
10.5, 8.2.4HV, 10.4.1 and RH12-1, ranked from highest to lowest antimicrobial activity. Maximum and minimum
inhibition zones of B. subtillis of these isolates were 10 and 1 mm, respectively. Staphylococcus aureus growth
was inhibited by seven isolates viz., 15.1, 11.1.2, 10.5, 8.2.4HV, 8.2.5HV, 10.4.1, and RH12-1. The isolates
displayed maximum and minimum inhibition zones for S. aureus of 8 and 4 mm, respectively. Pseudomonas
aeruginosa growth was inhibited by eight isolates viz., 8.2.4HV, RH13-26, 15.1, 10.5, 8.2.5HV, 11.1.2, 10.4.1, and
20.2.6. The isolates displayed maximum and minimum inhibition zones for P. aeruginosa inhibition of 20 and
4 mm, respectively. Candida albicans growth was inhibited by three isolates viz., 8.2.4HV, 10.5, and 8.2.5HV.
The isolates displayed maximum and minimum inhibition zones for C. albicans of 10 and 6 mm, respectively. No
isolate showed growth inhibition of E. coli. Isolates 8.2.4HV and 10.5 showed inhibition against four pathogenic
microorganisms, but not against E. coli. Some isolates (10.4.1 and RH12-1) showed no inhibition of pathogenic
microorganisms. Actinomycetes isolates all produced secondary metabolites, such as bioactive antibiotics, to
inhibit growth of some pathogens. These results are similar to those of Thangapandian et al. (2007) and
Gebreyohannes et al. (2013) who studied antimicrobial actinomycetes isolated from rhizosphere soil, water, and
sediment.
The isolate 10.5 displayed antimicrobial activity against four pathogens including Bacillus subtilis, S.
aureus, P. aeruginosa, and C. albicans, comparing with other isolates. The results showed in Figure 3.
Identification of actinomycetes
A total of 232 microorganisms were preliminary identified as Streptomyces and non- Streptomyces (rare
actinomycetes), by observing their growth rate on ISP medium. It was found that most of the isolates (206) showed
the typical morphology of Streptomyces and few isolates (26) were found to be rare actinomycetes.
Only ten isolates expressed antimicrobial activity. They were identified at the genus level by examining
their morphological characteristics, i.e., presence of aerial mycelium, conidia, pigment per Barka et al. (2015) and
Shirling and Gottileb (1966). The results indicate that these isolates can be divided into two genera, Streptomyces
and Microbispore. Isolates 8.2.4HV, 8.2.5HV, 6.1HV-1, 20.2.6, 15.1, 11.1.2, 10.5, RH13-26, and RH12-1 are
Streptomyces. They have diverse types of spores including retinaculiapert, oligosporous, spira and spiral spores.
Only isolate, 10.4.1, was a Microbispore spp. and it showed disporus spores as depicted in Figure 4.
Conclusion
This work clearly showed that soil samples collected from a community forest in Ban Khoklam Sang
Aram, Udon Thani Province contain actinomycetes that possess antibacterial and antifungal activities. The data
demonstrates the diversity of actinomycetes population in the form of colony morphology. Additionally, these
isolates were effective in producing antimicrobial metabolites to against B. subtilis, S. aureus, P. aeruginosa and
C. albicans. It is revealed that this location is a good source of antimicrobial actinomycetes, both Streptomyces

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 44
and rare-actinomycetes. Further work will be performed to determine their potential application to control human
and plant pathogens.
Acknowledgement
The authors acknowledge Udon Thani Rajabhat University for support for research funding. We are
especially thankful to Prof. Dr. Somboon Tanasupawat, Department of Biochemistry and Microbiology, Faculty
of Pharmaceutical Sciences, Chulalongkorn University for advice and support with some agents and pathogens in
this work. We thank Mr. Prechawut Pimphakdee, Department of Biology, Faculty of Science, Udon Thani
Rajabhat University, Udon Thani for his technical assistance.
References
Abdullah, Q. Y. M., Al-Maqtari, M.A., Al-Awadhi, O.A.A., Al-Mahdi, A.Y. 2016. New records of Streptomyces
and non Streptomyces actinomycetes isolated from soils surrounding Sana’a high mountain. International
Journal of research in pharmacy and biosciences 3(3): 19-31.
Agadagba, S.K. 2014. Isolation of actinomycetes from soil. Journal of microbiology research 4(3): 136-140.
Arifuzzaman, M., Khatun, M.R., and Rahman, H. (2010). Isolation and screening of actinomycetes from
sundarbans soil for antibacterial activity. African journal of biotechnology 9: 4615-4619.
Atta, H.M., and Ahamd, M.X. 2009. Antimycin-A antibiotic biosynthesis produced by Streptomyces sp., AZ-AR-
262: Taxonomy, fermentation, purification and biological activities. Australian journal of basic and applied
sciences 3: 126-135.
Baby, U.I., Tensingh Baliah, N., Ponmurugan, P., and Premkumar, R. 2002. Population level of certain of certain
beneficial microorganisms in tea soils. UPASI Tea Res. Found Newslett 12: 3.
Barka, E.A., Vatsa, P., Sanchez, L., Gaveau-Vaillant, N., Jacquard, C., Klenk, H.P., Clément, C., Ouhdouch, Y.
and van Wezel, G.P. 2 0 1 5 . Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol
Mol Biol Rev 80(1): 1-43.
Bontemps, C., Toussaint, M., Revol, P.-V., Hotel, L., and Jeanbille, M. 2013. Taxonomic and functional diversity of
Streptomyces in forest soil. FEMS Microbiology Letters 342(2): 157-167.
Ding, C.H., Jiang, Z.Q., Li, X.T., Li, L.T., and Kusakabe, I. 2004. High activity xylanase production by Streptomyces
olivaceoviridis E-86. World journal of microbiology and biotechnology 20: 7-10.
Embley, T., and Stackebrandt, E. 1994. The molecular phylogeny and systematics of the actinomycetes. Annual
Review of microbiology 48: 257-289.
Fang, B.-Z., Salam, N., Han, M.-X., Jiao, J.-Y., Cheng, J., Wei, D.-Q., Xiao, M., and Li, W.-J. 2017. Frontiers in
microbiology 8: 1-9.
Gebreyohannes, G., Moges, F., Sahile, S., and Raja, N. 2013. Isolation and characterization of potential antibiotic
producing actinomycetes from water and sediments of Lake Tana Ethiopia. Asian pacific journal of tropical
biomedicine 3(6); 426-435.
Jaralla, E.M., Al-Dabbagh, N.N., Hameed, N.M., and Abdul-Hussain, N. 2014. Screening for enzymatic
production ability and antimicrobial activity of actinomycetes isolated from soil in Hillah/Iraq. IOSR
Journal of pharmacy and biological sciences 9(5): 42-47.
Jeffrey, L.S. 2008. Isolation characterization and identification of actinomycetes from agriculture soils at
Semongok, Sarawak. African journal of biotechnology 7: 3697-3702.
Lechevalier, H., and Lechevalier, M. 1967. Biology of actinomycetes. Annual review of microbiology 21: 71-99.
Lemons, M.L., Toranzo, A.E., Barja, J.L. 1985. Antibiotic activity of epiphytic bacteria isolated from intertidal seaweeds.
Microbial ecology 11: 149-163.
Hayakawa, M. and Nonomura, H. (1987). Humic Acid-Vitamin Agar, a New Medium for the Selective Isolation of Soil
Actinomycetes. Journal of Fermentation Technology 65(5): 501-509.
McCathy, A.J., and Williams, S.T. 1992. Actinomycetes as agents of biodegradation in the environment-a review.
Gene 115(1-2): 189-192.
Mohseni, M, Norouzi, H., Hamedi, J., and Roohi, A. 2013. Screening of antibacterial producing actinomycetes from
sediments of the Caspian Sea. International journal of molecular and cellular medicine 2(2): 64-71.
Muthu, M.R., Subbaiya, R., Balasubramanian, M., Ponmurugan, P., and Selvam, M. 2013. Isolation and
identification of actinomycetes Isoptericola variabilis from Cauvery river soil sample. International
Journal of current microbiology and applied sciences 2(6): 236-245.
Okazaki, H., Serizawa, N., Enokita, R., Torikata, A. and Terahera, A. 1983. Taxonomy of actinomycetes capable of
hydroxylation of ML236B compactin. Journal of Antibiotics 36: 1176–1183.
Omar, S., Lemonnier, B., Jones, N., Ficker, C., Smith, M.L., Neema, C., Towers, G.H.N., and Arnason, J.T. 2000.
Antimicrobial activity of extracts of eastern North American hardwood trees and relation to traditional medicines.
Journal of Ethnophrmacology 73: 161-170.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 45
Rao, K.V.R., Mani, P., Satyanarayana, B., Rao, T.R. 2017. Purification and structural elucidation of three bioactive
compounds isolated from Streptomyces coelicoflavus BC 01 and their biological activity. Biotechnology 7: 24.
Ravel, J., Wellington, W.H., and Hill, R.T. 2000. Interspecific transfer of Streptomyces liner plasmids in sterile
amended soil microorganism. Applied and environmental microbiology 66: 529-534.
Shahidi, B.G.H., Fooladi, M.H., Mahadavi, M.J., Shahghasi, A. 2004. Broad spectrum a novel antibacterial from
Streptomyces sp. Biotechnology 3: 126-130.
Shirling, E. B., and Gottlieb, D. 1966. Methods for characterization of Streptomyces species. International Journal
Systematic Bacteriology 16: 313-340.
Stackebrandt, E., Wilt, D., Kemmerling, C., Kroppenstedt, M., and Liesack, W. 1991. Designation of
Streptomyces 16S and 23S rRNA based target regions of oligonucleotide probes. Applied and
environmental microbiology 57: 1468-1477.
Thangapandian, V., Ponmurugan, P., and Ponmurugan, K. 2007. Actinomycetes diversity in the rhizosphere soils of
different medicinal plants in Kolly Hills-Tamilnadu, India, for secondary metabolite production. Asian journal of
plant sciences 6(1): 66-70.
Usha, R.J., Shenpagam, N.H., Devi, D.K. 2011. Antagonistic activity of actinomycetes isolates against human
pathogen. Journal of microbiology and biotechnology research 1: 74-79.
World Health Organization antimicrobial resistance: global report on surveillance 2014. Geneva, Switzerland:
WHO; 2014.
Xu, L.H., Jiang, Y., Li, W.J., Wen, M.L., Li, M.G., Jiang, C.L. 2005. Streptomyces roseoalbus sp. Nov., and
actinomycetes isolated from soil in Yunnan, China. Antoine van Leeuwenhoek 87: 189-194.
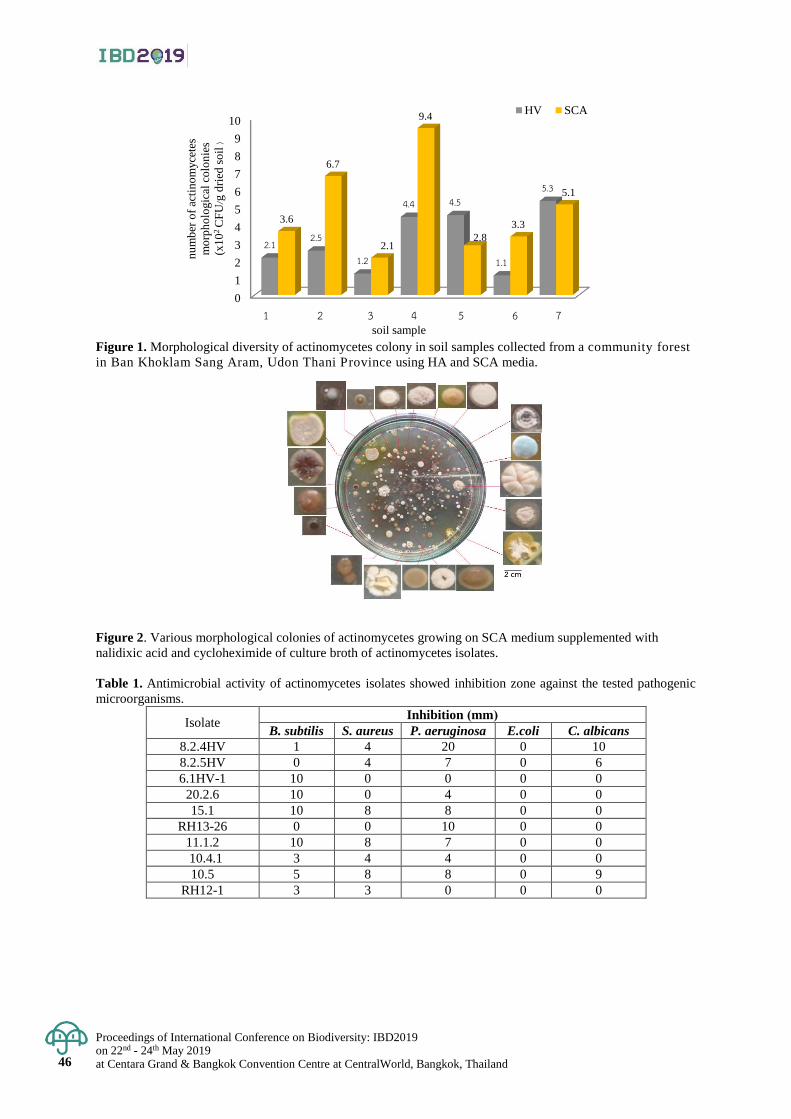
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 46
0
1
2
3
4
5
6
7
8
9
10
6 8 10 13 11 15 20
2.1 2.5
1.2
4.4 4.5
1.1
5.3
3.6
6.7
2.1
9.4
2.8
3.3
5.1
HV SCA
soil sample
nu
mb
er o
f ac
tin
om
yce
tes
mo
rph
olo
gic
al c
olo
nie
s
(x1
02 C
FU
/gd
ried
so
il)
Figure 1. Morphological diversity of actinomycetes colony in soil samples collected from a community forest
in Ban Khoklam Sang Aram, Udon Thani Province using HA and SCA media.
Figure 2. Various morphological colonies of actinomycetes growing on SCA medium supplemented with
nalidixic acid and cycloheximide of culture broth of actinomycetes isolates.
Table 1. Antimicrobial activity of actinomycetes isolates showed inhibition zone against the tested pathogenic
microorganisms.
Isolate Inhibition (mm)
B. subtilis S. aureus P. aeruginosa E.coli C. albicans
8.2.4HV 1 4 20 0 10
8.2.5HV 0 4 7 0 6
6.1HV-1 10 0 0 0 0
20.2.6 10 0 4 0 0
15.1 10 8 8 0 0
RH13-26 0 0 10 0 0
11.1.2 10 8 7 0 0
10.4.1 3 4 4 0 0
10.5 5 8 8 0 9
RH12-1 3 3 0 0 0
1 2 3 4 5 6 7
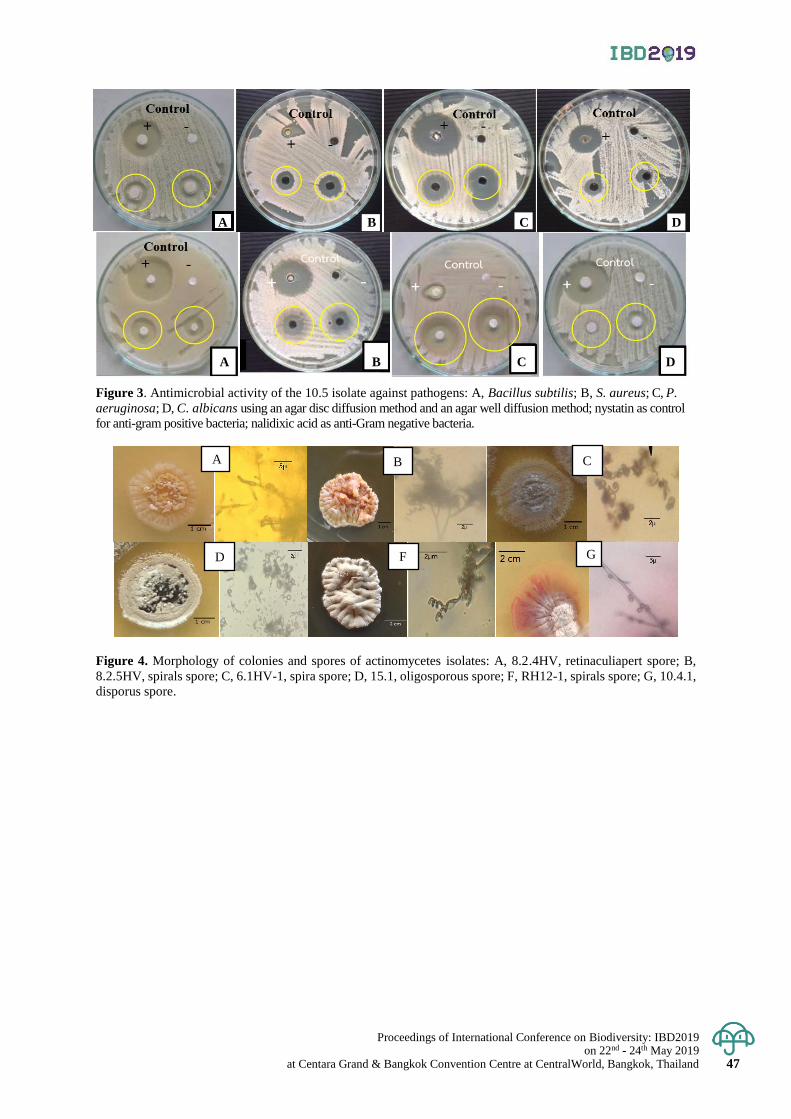
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 47
Figure 3. Antimicrobial activity of the 10.5 isolate against pathogens: A, Bacillus subtilis; B, S. aureus; C, P.
aeruginosa; D, C. albicans using an agar disc diffusion method and an agar well diffusion method; nystatin as control
for anti-gram positive bacteria; nalidixic acid as anti-Gram negative bacteria.
Figure 4. Morphology of colonies and spores of actinomycetes isolates: A, 8.2.4HV, retinaculiapert spore; B,
8.2.5HV, spirals spore; C, 6.1HV-1, spira spore; D, 15.1, oligosporous spore; F, RH12-1, spirals spore; G, 10.4.1,
disporus spore.
A C
D F G
A B C D
A B C D
B

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 48
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 48 - 54
Screening on anti-inflammatory property of 45 Thai human probiotics from
Biodiversity Research Centre of TISTR
Prapaipat Klungsupya1*, Weerasak Taengphan1, Bhusita Wannissorn2, Supatjaree Ruengsomwong2
and Thanchanok Muangman1
1Expert Centre of Innovative Herbal Products (InnoHerb), Thailand Institute of Scientific and Technological
Research (TISTR), Techno Polis, Khlong Luang Distric, Pathum Thani, Thailand 2Biodiversity Research Centre (BRC), Thailand Institute of Scientific and Technological Research (TISTR),
Techno Polis, Khlong Luang District, Pathum Thani, Thailand.
*Corresponding author e-mail: [email protected]
Abstract: Probiotics are defined as living microorganisms which can provide various health benefits. They can
inhibit overgrowth of pathogenic bacteria by antibacterial substance production and stimulating the health
function in gut system. Moreover, probiotics can promote the affect on innate immunity such as phagocytosis,
and anti-inflammatory activity. The anti-inflammatory effect of probiotics is attributed to the released cytokines
from immune cells such as lymphocytes, mast cells and macrophages. The aim of this study was to determine
the anti-inflammatory activity of 45 TISTR human probiotic strains on lipopolysaccharide-induced macrophage
RAW 264.7 cell line. Heat-killed supernatants of these 45 probiotic strains (Med1 to Med45) prepared and tested
at 100% concentration. Lipopolysaccharides (LPS, 10 µg/ml) of 100 µl was used to induce inflammation. Results
were collected based on the measurements of nitric oxide (NO), tumor necrosis factor-α (TNF-α) and interleukin-6
(IL-6) productions using ELISA Kit assays. The results revealed that 20 out of 45 TISTR probiotics yielded various
degrees of their anti-inflammatory capacity with %NO inhibition ranging from 19.91±1.50 to 82.04±3.71% compared
with β-glucan (83.93±0.07%, positive control). Stimulation of RAW 264.7 macrophages with LPS induced a high
production of NO by 378.61 pg/ml. The reduction in TNF-α levels (% TNF-α inhibition) was seen in 16 TISTR
probiotics with the highest activity at 52.46% (Med 8 strain) and the lowest at 5.69% (Med 9 strain). Considering
on % IL-6 inhibition, among 45 TISTR probiotic strains, 18 strains expressed positive results by 5.79 to 68.79%.
The results suggest that many TISTR probiotic strains are potential of anti-inflammatory activity on macrophage
RAW 264.7 cells. These findings can support the utilization of TISTR probiotics in preventing and down-regulating
inflammation.
Keywords: Anti-inflammation, interleukin, macrophages, nitric oxide, probiotics, tumor necrosis factor-α
Introduction
Thailand Institute of Scientific and Technological Research (TISTR) by Biodiversity Research Centre
(BRC) is responsible for culture collection of bacteria, yeasts, molds and seaweeds which are beneficial to
industrial and environmental usage. At the moment, BRC has more than 500 human probiotics collected and
isolated from Thai population kept in the bank. Probiotics have been promoted the affecting on innate immunity
such as phagocytosis, bactericidal activity and anti-inflammatory activity. The anti-inflammatory effect of
probiotics is attributed to the released cytokines from immune cells such as lymphocytes, mast cells and
macrophages (Thomas and Versalovic, 2010). Macrophages express a variety of surface receptors that including
cytokine receptors (CRs), the interleukin-1 receptor (IL-1R) and complement receptor for pathogen associated
molecular pathways (PAMPs) such as lipopolysaccharide (LPS). The activated macrophages also produce and
secrete cytokines such as TNF-α, IL-1, IL-6, IL-8, IL-12 and IFN)-γ, that are important for inflammatory and
acquired immune response (Guan et al., 2011; Ariel et al., 2012). Rodes et al., (2013) reported that the pro-inflammatory
cytokine concentration of TNF-α, IL-1beta was reduced by probiotics Lactobasilus and Bifidobacterium but anti-
inflammatory cytokine was increased on lipopolysaccharide induced-RAW 264.7 macrophages. Therefore, macrophages
behave as critical play in the initiation, maintenance, and resolution of inflammation through the release of variety
of factors in response to an activating stimulus (Moro et al., 2012). The aim of the present study was to determine
the anti-inflammatory mediator effect of 45 TISTR human probiotic strains on lipopolysaccharide-induced macrophage
RAW 264.7 cell line.
To assort local probiotic strains for Thailand Institute of Scientific and Technological Research (TISTR)
probiotic bank, we randomly collected various sources of flowers and fruits grown in the northern part of Thailand
to isolate bacterial strains. All isolated bacterial strains were screened for probiotic properties according to a
notification of Thai Food and Drug Administration (FDA), Ministry of Public Health (MOPH) and World Health
Organization (WHO) starting from gram straining, catalase-oxidase activity, hemolytic activity, antimicrobial
activity against potentially pathogens, antibiotic susceptibility, gastric acidity resistance (pH 2 and 3), bile salt tolerance,
adherence to human epithelial cell line, and immunomodulatory activity in macrophage function. They were gram

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 49
positive, rod shape, catalase-negative, and negative hemolytic activity bacteria. All of them have antimicrobial
activity against pathogens such as Escherichia coli, Staphylococcus aureus. The antibiotic susceptibility, gastric acidity
and bile salt resistance and adherence to human epithelial cell lines of them were already proved. After probiotic
properties screening, we got 45 probiotic strains in total. Therefore, this study was carried out to evaluate anti-inflammation
activity of these 45 TISTR probiotics.
Materials and Methods
Probiotic strains and growth condition
A total of 45 TISTR human probiotic strains (namely Med 1 to Med 45) from the TISTR culture collection
bank of Biodiversity Research Centre (BRC) were cultivated in de Man Rogosa and Sharpe (MRS) broth (Merck,
Germany) with 0.05% L-cysteine (Merck, Germany) under anaerobic condition at 37C for 48 hr. T hey were
adjusted their turbidity to the concentration to match that of McFarland No. 0.5 (approximate 108 CFU/ml). Cell pellets
were obtained by centrifugation at 8,500 rpm for 5 min at 25°C and washed 3 times with 0.85% NaCl. After
washing, they were resuspended with 10 ml of Dulbecco's Modified Eagle Medium (DMEM) high glucose
(Biowest, USA) without antibiotics before heating at 110°C for 10 min. The supernatants of each probiotic strain
was kept at -20°C until use.
Macrophages culture
RAW 264.7 macrophage cell line (ATCC TIB-71TM) was purchased from American Type Culture Collection
(ATCC) (Manassas, VA, USA). The cells were cultured as adherent in Dulbecco’s Modified Eagle Medium
(DMEM) (Biowest, USA) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS) (GIBCO®,
USA) and 1% (v/v) penicillin-streptomycin (GIBCO®, USA) in humidified atmosphere incubator with 5% CO2
at 37°C. The cells were collected by centrifugation (Kobota, Japan) at 1500 rpm, 4°C for 5 min, aspirated and
fresh culture medium was added before dispensing into new culture flasks. All steps of culturing and maintaining
of RAW 264.7 macrophage cells were conducted in aseptic condition using the Biological Safety Cabinet Class II
(SANYO, Japan).
Determination of the anti-inflammatory activity in RAW 264.7 macrophage cells.
Nitric oxide measurement by Griess reagent assay Griess reagent system is based on the chemical reaction which uses sulfanilamide and N-1-napthylethylenediamine
dihydrochloride (NEDD) under phosphoric acid conditions. When sulfanilamide is added, the nitrites (NO2–) form
a sulphonamide. Then, the NEDD was added, the sulphonamide was formed a pink colour of diazonium. To determine
the anti-inflammatory activity, 200 µl of RAW 264.7 macrophage cells in DMEM were seed onto 96-well plates
at a density of 2x104 cells/well and incubated for 24 hr at 37°C in humidified atmosphere incubator with 5% CO2.
The overnight culture media were removed and RAW 264.7 macrophage cells (2x106 cells/well) were separately
treated with 100 µl of probiotics (Med1-Med45) or β-glucan at 100 µg/ml (positive control) and incubated for 24
hr. Culture media were replaced by 100 µl (10 µg/ml concentration) of lipopolysaccharide (LPS) from Escherichia
coli (Sigma, USA) and incubated at 37°C in humidified atmosphere incubator with 5% CO2 for 24 hr. After
incubation, the culture media was analyzed for nitric oxide (NO) production using the Griess reaction. Briefly,
the culture media was mixed with 100 μl of Griess reagent (1% sulfanilamide, 5% phosphoric acid and 1%
Naphthyletylenediamine dihydrochloride) and incubated at room temperature for 10 min. Evaluation of the concentration
of NO in the supernatants of the RAW 264.7 cell culture was carried out with enzyme-linked immunosorbent
assay (ELISA) kit (Thermo, USA) according to the manufacturer’s instruction. Then, the absorbance at 540 nm
was measured using microplate reader (Tecan Infinite 200 Pro, Austria). The amount of nitrite presented in the
samples was measured with the sodium nitrite serial dilution standard curve.
Cytokines measurement
RAW 264.7 macrophage cells were prepared in 96-well plate as previously described in NO measurement
assay. Following removal of overnight culture medium, cells (2x106 cells/well) were treated with 100 µl of each TISTR
probiotics (Med1-Med45) or β-glucan (Sigma, USA) at 100 µg/ml which was served as a positive control. Cells were
incubated at 37°Cin humidified atmosphere incubator with 5% CO2 for 24 hr. Then, culture media were replaced
with 100 µl of LPS (Sigma, USA) at 10 µg/ml and incubated for 24 hr. The supernatants of treated-RAW 264.7
macrophage cells were collected by centrifugation at 1500 rpm, 4°C for 5 min and analyzed for two cytokine
measurements including interleukin-6 (IL-6) and tumor necrosis- alpha (TNF-α) by Human IL-6 ELISA kit
(Thermo, USA) and Human TNF-α ELISA kit (Thermo, USA), respectively. The secreted cytokine concentrations
were quantified on the basis of a linear dose-response standard curve.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 50
Statistical analysis
All data were expressed as mean ± standard error (Mean ± SE) of triplicated determination. The
significance of difference was used to compare mean (p<0.05). All analyses were performed using One-way
ANOVA and Tukey’s Honestly Significant Difference test.
Result & Discussion
In this study, the anti-inflammatory capacity of 45 TISTR probiotic strains was determined on % inhibition
of NO, TNF-α and IL-6 productions. Following treatments, the morphology of RAW 264.7 macrophage cells was
examined under the inverted microscopy as demonstrated in Figure 1. It was observed that 25-26 TISTR probiotic
strains exhibited cytotoxic property (cell viability less than 50%) against RAW 264.7 macrophage cells when tested
at 100% concentration (undiluted heat-killed supernatants). This observation was confirmed by cell viability and
proliferation test using WST-1 assay whose principle is based upon a conversion of tetrazolium salts to colored
formazan compounds by mitochondrial succinate-tetrazolium reductase which exists in viable cells. The results
of WST-1 assay were analyzed by spectrophotometric quantification. However, results of WST-1 assay suggested
that at least 27 out of 45 TISTR probiotic strains were non-cytotoxic to RAW 264.7 macrophage cells. Among them,
10 strains demonstrated a cell-proliferation promoting activity with cell survival rate greater than 100% compared
with the untreated cells.
Macrophages are a type of phagocytic cells of the innate immune system. They play a crucial role and function
as one of the first line of host defense mechanism against invading pathogen, foreign materials and inflammation
(Flannagan et al., 2007; Ariel et al., 2012). Macrophages are activated in the inflammatory process including cytokines,
bacterial lipopolysaccharide (LPS), extracelluar matrix proteins, and other chemical mediators. Activated
macrophages release several different chemical mediators, including ROS, NO, IL-6, TNF-α, and PGE2, that
perpetuate the pro-inflammatory response (Shaikh 2011; Soromou et al., 2012). To determine the effect of TISTR
probiotics on NO and pro-inflammatory mediators (TNF-α, IL-6) productions, RAW 264.7 cells were treated with
45 heat-killed TISTR probiotic strains (Med1 - Med45) at 100 % concentration for 24 hr prior to stimulate with
LPS at a concentration of 10 µg/ml for 24 hr. Then, concentrations of NO, TNF-α and IL-6 mediators in medium
were determined in cell supernatants. Under the stimulus with LPS which was a pro-inflammatory molecule,
RAW 264.7 macrophage cells exhibited high concentration levels of NO (up to 126.17±5.33 µM), TNF-α (up to
378.61±0.52 pg/ml) and IL-6 (up to 149.73±1.52 pg/ml). Overall of 45 heat-killed TISTR probiotic strains, 19
strains were able to reduce the production of NO demonstrating as “% NO inhibition” (Figure 2). Overall, Med2,
Med6 and Med10 showed the best anti-inflammatory activity on RAW264.7 macrophage cells. Their anti-
inflammatory capacity was close to β-glucan (83.93±0.07%, positive control). The decrease of NO production
was not related to viability of cells, which was not affected by LPS stimulation (data not shown).
Anti-inflammatory activity of 45 heat-killed TISTR probiotic strains was confirmed by inhibition of
productions of pro-inflammatory cytokines focusing on TNF-α and IL-6. RAW264.7 cells were first pre-treated
with TISTR probiotic samples (Med1 - Med45) and then stimulated with LPS for verifying their protective anti-
inflammatory activity. The known anti-inflammatory compound, β-glucan at concentration of 100 µg/ml was used
as a positive control. β-glucans are naturally occurring polysaccharides that are produced by bacteria, yeast, fungi
and many plants (Han et al., 2008). Results on % inhibition of TNF-α and IL-6 productions were shown in Figures
3 and 4, respectively. It was found that 19 out of 45 TISTR probiotic strains yielded various degrees of their anti-
inflammatory capacity against LPS-induced TNF-α and IL-6 mediators in RAW264.7 macrophage cells. The
reduction in TNF-α levels (% TNF-α inhibition) was found in 18 TISTR probiotics with the highest activity by
52.46±2.98% (Med 8) and the lowest at 5.56±32.66% (Med 9) (Figure 3). Regarding % IL-6 inhibition, among
45 TISTR probiotic strains, 18 strains expressed positive results by 5.79±2.80% (Med40) to 68.53±1.63% (Med1)
- 68.79±2.79% (Med41) (Figure 4). However, the anti-inflammatory capacity was not found in 26 TISTR probiotic
strains tested including Med12, Med13, Med15, Med17, Med18, Med19, Med20, Med21, Med22, Med23, Med24,
Med25, Med26, Med27, Med28, Med29, Med32, Med33, Med34, Med35, Med36, Med37, Med38, Med43 and
Med45 strains. This might be due to their cytotoxic property when tested at 100% concentration as described in
WST-1 assay results.
Conclusion
Besides increasing inflammation and stimulating the immune system, macrophages play an important
anti-inflammatory role and can decrease immune reactions through the release of cytokines. The results generated
by this study suggest that many TISTR probiotic strains (positive results found in 18-20 strains among 45 strains
tested) potentially exhibit anti-inflammatory activity on RAW 264.7 macrophage cells. These findings can support
the utilization of TISTR probiotics in preventing and down-regulating inflammation. They can be utilized as
ingredients for functional foods and dietary supplement products.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 51
Acknowledgement
This work was performed by Thailand Institute of Scientific and Technological Research (TISTR) under
financial support by a grant from the Ministry of Science and Technological (MOST) during 2018-2020.
Referenes
Ariel, A., Maridonneau-Parini, I., Rovere-Querini, P., Levine, J., and Mühl, H. 2012. Macrophages in inflammation
and its resolution. Front Immunol 324 (3): 1-2.
Flannagan, S., Harrison, E., Yip, M., Jaqaman, K., and Grinstein, J. 2007. Macrophages are crucial frontline cells
in the body's defense against infection. J Leukoc Biol 82(2):417-428.
Guan, D., Zhang, Z., Yingnan, Y., Gouqing, X., and Jinqiang, L. 2011. Immunomodulatory activity of polysaccharide
form the roots of Actinidia kolomikta on macrophage. Inter J Biol 3 (2): 3-10.
Han, M., Han, Y., Hyun, S., and Shin, H. 2008.Solubilization of water-insoluble β-glucan isolated from
Ganoderma lucidum. J Env Biol 29 (2): 234-242.
Moro, C., Palacios, I., Lozano, M., D’Arrigo, M., Guillamón, E., Villares, A., Martínez, J., and García- Lafuentea,
A.2012. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated
RAW 264.7 macrophages. Food Chem 130(2): 350-355.
Shaikh, Z. 2011.Cytokine & their physiologic and pharmacologic functions in inflammation: a review. Inter J
Pharma Life Sci 2 (11):1247-1263.
Soromou, W., Zhang, R,. Li, R., Chen, N., Guo, W., Huo, M., Guan, S., Lu, J., and Deng, X. 2012. Regulation of
inflammatory cytokine in lipopolysachharide-stimulated RAW. 264.7 murine macrophage by 7-O-
methyl-naringenin. Molecules 17 (3): 3574-3585.
Thomas, M., and Versalovic, J. 2010. Probiotics-host communication modulation of signaling pathways in the
intestine. Gut Microbes.1 (3): 148-163.
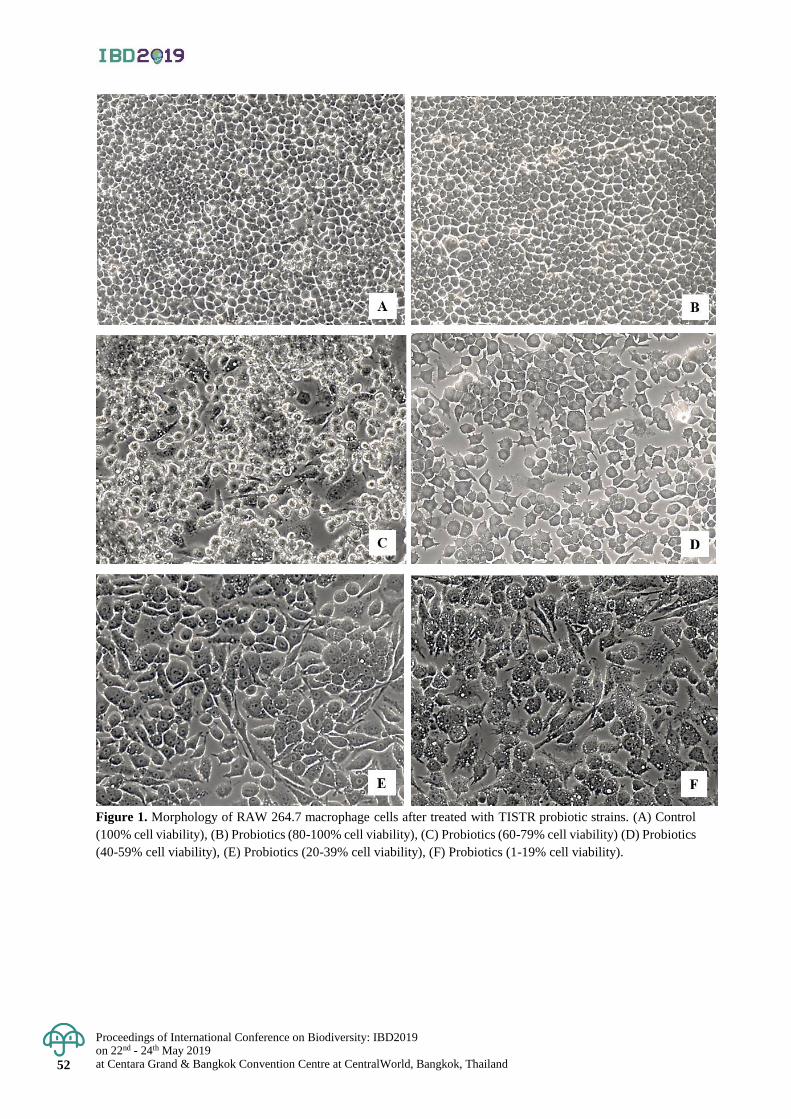
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 52
Figure 1. Morphology of RAW 264.7 macrophage cells after treated with TISTR probiotic strains. (A) Control
(100% cell viability), (B) Probiotics (80-100% cell viability), (C) Probiotics (60-79% cell viability) (D) Probiotics
(40-59% cell viability), (E) Probiotics (20-39% cell viability), (F) Probiotics (1-19% cell viability).
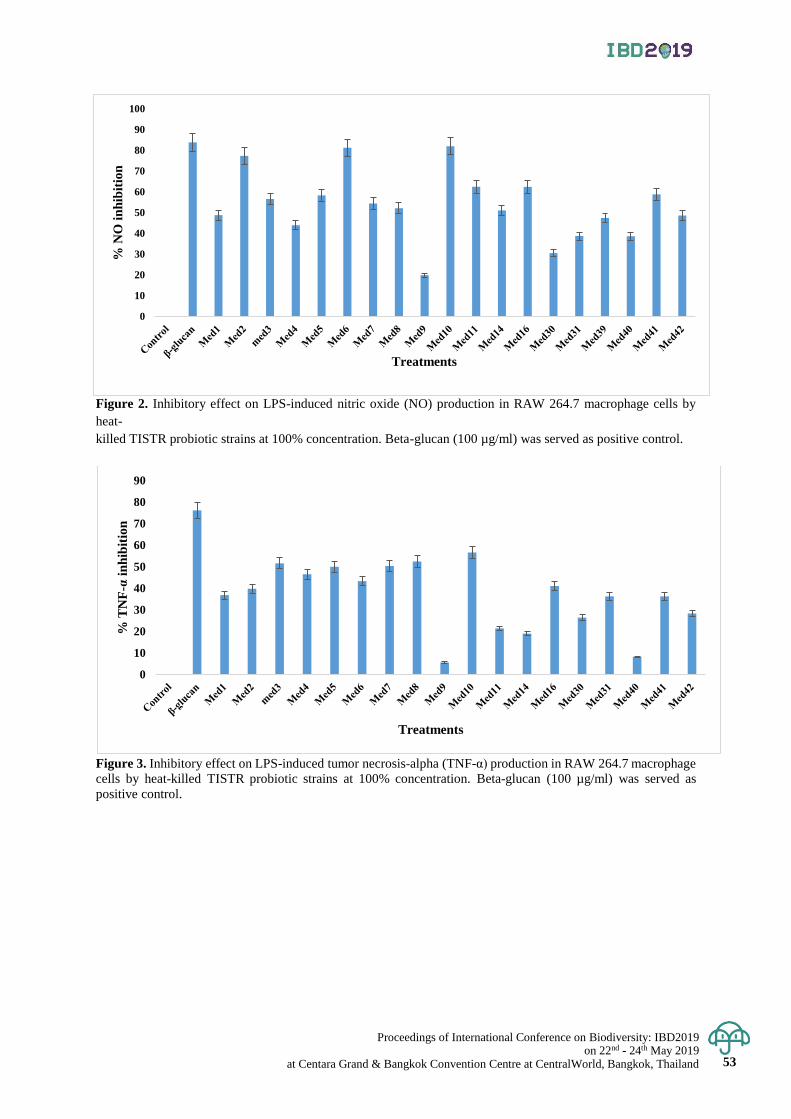
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 53
Figure 2. Inhibitory effect on LPS-induced nitric oxide (NO) production in RAW 264.7 macrophage cells by
heat-
killed TISTR probiotic strains at 100% concentration. Beta-glucan (100 µg/ml) was served as positive control.
Figure 3. Inhibitory effect on LPS-induced tumor necrosis-alpha (TNF-α) production in RAW 264.7 macrophage
cells by heat-killed TISTR probiotic strains at 100% concentration. Beta-glucan (100 µg/ml) was served as
positive control.
0
10
20
30
40
50
60
70
80
90
% T
NF
-αin
hib
itio
n
Treatments
0
10
20
30
40
50
60
70
80
90
100%
NO
in
hib
itio
n
Treatments
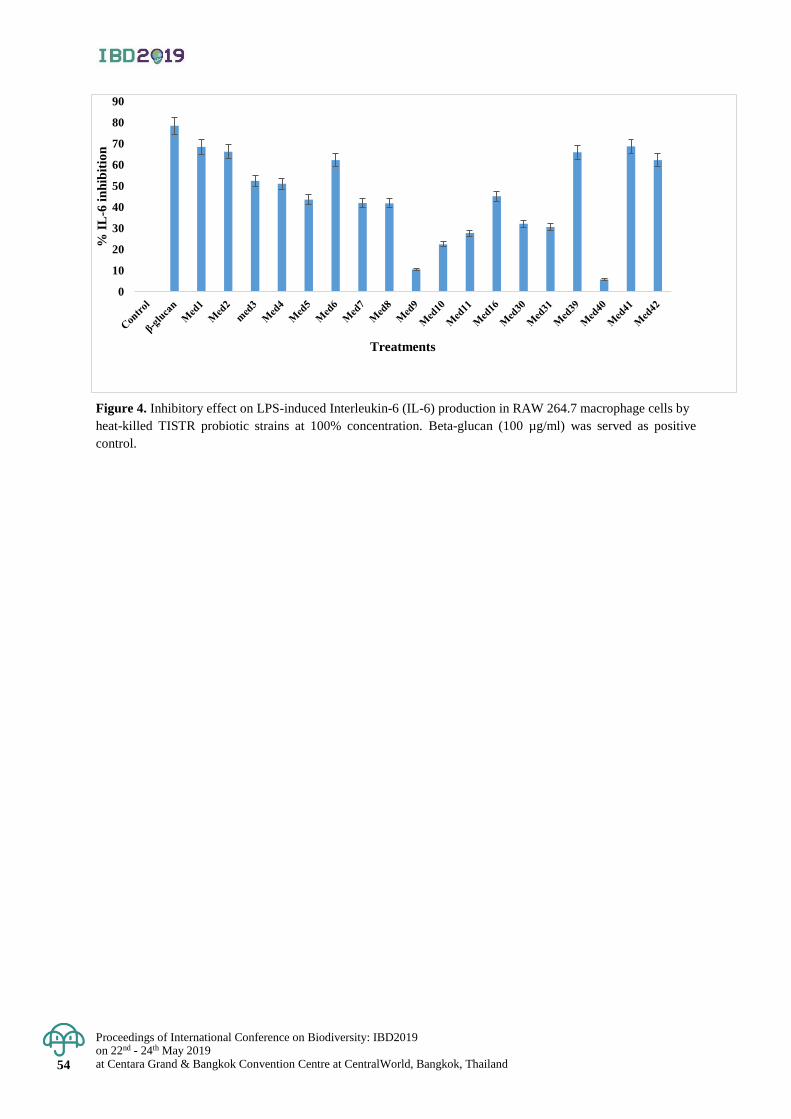
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 54
Figure 4. Inhibitory effect on LPS-induced Interleukin-6 (IL-6) production in RAW 264.7 macrophage cells by
heat-killed TISTR probiotic strains at 100% concentration. Beta-glucan (100 µg/ml) was served as positive
control.
0
10
20
30
40
50
60
70
80
90%
IL
-6 i
nh
ibit
ion
Treatments

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 55
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 55 - 61
Effect of water temperature on growth, survival and health status
of East Asian Bullfrog (Hoplobatrachus rugulosus) Larvae
Sornsawan Liamtong1, Kanokporn Saenphet2, Monruedee Chaiyapo2 and Supap Saenphet2*
1Master's Degree Program in Biology, Faculty of Science, Chiang Mai University, Muang District, Chiang Mai, Thailand 2Department of Biology, Faculty of Science, Chiang Mai University, Muang District, Chiang Mai, Thailand
*Corresponding author e-mail: [email protected]
Abstract: East Asian Bullfrog (Hoplobatrachus rugulosus) is an economically and nutritionally important animal
in Thailand. However, many factors including UV radiation, infection and especially the fluctuation of temperature
adversely affect the East Asian Bullfrog culture. Thus, the aims of this study was to investigate the effects of water
temperature changes on the survival and growth as well as oxidative stress of the H. rugulosus tadpoles. The result
showed that cold shock and heat shock significantly decreased the survival rate of the tadpoles when compared to
those of control groups. Cold shock at 23°C showed the highest survival rates among the cold shocked groups and
the heat shock at 27°C showed the highest survival rates among the heat shocked groups. However, there was no
significant difference (p>0.05) of their body length at day 0 and day 7 among the experimental groups. Histological
examination showed blood congestion and a large number of melanomacrophage and Kupffer cell in the liver
tissue of tadpoles exposed to cold and heat shock when compared to those of the controls. Moreover, blood congestion
was found in tadpoles exposed to the cold shock (17°C). In addition, cold and heat shock groups revealed
significantly increased malondialdehyde (MDA) level in whole body of the tadpoles compared with the control
group. Moreover, the level of MDA in the cold shock group was significantly greater than that in the heat shock
group. The data obtained from this study will help to raise awareness of temperature shock in farm-raised frogs
and others wild amphibian species. Keywords: Growth, Hoplobatrachus rugulosus, Survival, Tadpole, Temperature.
Introduction
East Asian Bullfrog (Hoplobatrachus rugulosus, Wiegmann, 1835) is an anuran amphibian with a widespread
distribution from central China, Myanmar, Peninsular, Malaysia including Thailand. They are commonly found
in wetlands and paddy fields (Diesmos et al., 2004; Gratwicke et al., 2009). This species is economically important
in Thailand because it is used as food by the local people and they are a very good source of protein. Moreover,
frozen frog legs of farm-raised frogs are a part of the exports to many countries (Oza, 1990). However, they are
susceptible to changes in their natural environment brought on, for example, by pollution, pesticides, diseases,
especially the climate change.
Frogs are ectothermic animals, water temperature is one of the most important environmental factors affecting
the physiology of organisms because they do not have a thermoregulatory mechanism (Goldstein et al., 2017).
Earlier studies indicated that temperature fluctuation could influence their survival, growth and physiological functions
in the wild and farmed frogs (Wang et al., 2012; Bellakhal et al., 2014). Moreover, Sahoo and Kara (2014) demonstrated
that cold shock on Tiger frogs (H. rugulosus) caused acute mortality in the larvae. Thus, these phenomena were the main
causes of the mortality and the decline in frog populations. In addition, Kern et al. (2015) found that water temperature
fluctuation on Australian frogs (Limnodynastes peronii) inhibited growths and developments by significant
decrease in total length (TL), body mass (Mb) and body length (BL) when compared with those reared in constant
water temperature. Furthermore, temperature fluctuation can lead to oxidative stress in organisms by increased
production of reactive oxygen species (ROS), which cause lipid peroxidation (Sahin, 2004). Sahoo and Kara
(2014) reported that the exposure of short-term cold stress to common Indian toads (Bufo melanostictus)
significantly increased ROS that was indicated by the rising of lipid peroxidation in the liver, kidney and brain
tissues. Additionally, thermal stresses can also effectively induce histological alterations in the liver of other
ectotherms, such as fat droplets accumulation (Liu et al., 2018), mitochondrial swelling (Xu et al., 2018), hepatocyte
degeneration and necrosis (Ates et al., 2006). It has been reported that histological examination in the liver tissue
of giant spiny frog (Quasipaa spinosa) exposed to cold and heat shock, erythrocyte extravasation as hemorrhage
and congestion was observed. Inflammatory cell infiltration was conspicuous and enlargement of sinusoids was
also presented (Liu et al, 2018). Therefore, the present study aimed to investigate the effects of water temperature
changes on the survival and growth as well as oxidative stress of the H. rugulosus tadpoles.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 56
Materials and Methods
Animals and experimental design
Experiments were performed with H. rugulosus larvae. Tadpoles, Gosner stage 26, were obtained from
Thunpisit frog farm, Chiang Mai province. They were kept in dechlorinated water with laboratory conditions at
least 24 hours for acclimation. The tadpoles were fed twice daily with commercial tadpole diet. Experimental designs
were as followed:
Experiment 1 – The frog tadpoles were divided into 7 groups comprising of 30 tadpoles each. Group 1-3
were served as cold shock groups and groups 4-6 were served as heat shock groups. For cold shock groups, the initial
temperature was set at 25°C and then decreased to 23°C, 20°C and 17°C within 4 hours. For heat shock groups,
the initial temperature was set at 25°C and then increased to 27°C, 30°C and 33°C within 4 hours. The control
group was maintained at 25°C throughout the experimental period. Each treatment was conducted in triplicate.
Dead larvae, if present, were counted and removed daily. After 7 days of exposure, the survived tadpoles were
measured for their body length to determine the growth rate. The liver tissues of frog tadpoles were collected and
processed for histological investigation.
Experiment 2 – The frog tadpoles were divided into 3 groups comprising of 30 tadpoles each. Group 1 was
served as control group and the temperature was set at 25°C. Group 2 was set as cold shock group and the temperature
was started at 25°C and then decreased to 23°C (the temperature which was produced the highest viability of cold
shock-exposed larvae from the experiment I). Group 3 was heat shock group and the temperature was started at
25°C and then increased to 27°C (the temperature which was produced the highest viability of heat shock-exposed
larvae from the experiment I). Each treatment was conducted in triplicate. The tadpoles were exposed to heat and
cold temperatures until they reached the Gosner stage 42. They were then collected for determining oxidative
stress by evaluating the level of malondialdehyde (MDA).
Histopathological examinations The tadpoles at day 7 were collected and whole body were fixed in Bouin's solution. After fixation for
24-30 h, tissues were dehydrated through a graded series of ethanol, cleared in xylene, and then embedded in
paraffin. Six μm sections were made and stained with hematoxylin and eosin (H&E). The sample was analyzed
by assessing the histopathological alterations under a light microscope.
Estimation of lipid peroxidation in tadpoles tissue. Twenty milligrams of homogenated tadpoles tissue were mixed in 1 ml of 0.1 M Phosphate Buffer
Solution (PBS), pH 7.4 at -4 °C. The mixed sample was centrifuged at 1,155 x g for 10 minutes and the supernatant
was stored at -40°C for examination of MDA within 24 hours. The degree of lipid peroxidation was determined
colorimetrically using the method of Buege and Aust (1976). Briefly, 100 µl of sample was mixed with 450 µl of normal
saline, 200 µl of thiobarbituric acid (TBA) and 1,000 µl of trichloroacetic Acid (TCA). The mixture was heated
at 100°C for 30 minutes, cooled with running tap water and then added with 2,000 µl of distilled water, mixed
with vortex mixer and centrifuged at 1,155 x g for 10 minutes. The absorbance at 532 nm was then measured and
calculated for MDA concentration against the MDA standard curve.
Estimation of protein in tadpoles tissue.
The level of protein content from each tissue was estimated following the method described by Bradford
(1976) and the bovine serum albumin (BSA) was used as a standard protein. Five milligrams of Coomassie Blue G
were dissolved in 1 ml of 25% (v/v) methanol containing 42.5% (v/v) H3PO4. One volume of the prepared dye
was diluted with four volumes of distilled water, and 0.04 mL of each sample was mixed with 2 mL of diluted
dye. The absorbance was read at 590 nm and calculated for protein concentration against the protein standard curve.
Statistical Analyses All statistical analyses were managed with Statistical Package for Social Science (SPSS) software version
20.0 for Mac OSX (SPSS Inc., IBM). Values were verified for normality and homogeneity of variance by Shapiro-
Wilk test and Levene's tests, respectively. Mean comparisons were carried out by one way analysis of variance
(ANOVA) followed by Tukey's HSD test.
Result & Discussion
Survival and growth of tadpoles
As shown in Figure 1, cold shock and heat shock groups revealed significantly decreased survival rate of
the tadpoles when compared to those of the control groups (p<0.05). Moreover, the survival rate of tadpoles in
cold shock groups was not significantly different from that of heat shock groups. The highest survival rates in cold
and heat shock were found at 23°C and 27°C, respectively. However, there was no significant difference (p>0.05)
of their body length at day 7 among the experimental groups. (Figure 2) The highest survival rates obtained from
our study were slightly associated with the studies of Wang et al. (2012) which reported that the mortality rate of

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 57
Tiger frog (H. rugulosus) exposed to cold shock were 28.1% at 12°C, 87. 5% at 10°C and 100% at 8°C.
Additionally, Bellakhal et al. (2014) reported that African green frog (Pelophylax saharicus) survival was reduced
at temperatures above 28 °C. These inferred that the temperature fluctuation on the survival rate among species
may be determined by differences in their thermal optimum for survival. The larval tadpole H. rugulosus used by
Wang et al. and P. saharicus used by Bellakhal might have greater thermal optimum range than the older larvae
used in our study. In addition, the body length of tadpoles obtained from our study are contradiction with the
studies of Kern et al. (2015) which, reported that body length of L. tasmaniensis tadpoles raised in temperature
shock was significantly lower than those of tadpoles in the constant temperature. As a result, tadpoles obtained
from cold and heat shock had reduced survival when compared with tadpoles reared in the constant temperature
since the temperature has been well known as the most pervasive abiotic factor to influence physiological function
because of thermodynamic effects on biochemical reactions which underline survival and growth (Somero, 2002).
Environmental temperature determines body temperature in most ectotherms (Guderley, 2004; Seebacher and
Murray, 2007). Consequently, the temperature fluctuation could affect the physiology, survival and growth of
tadpole as seen in our study.
Histopathological changes in tadpole liver
Histopathological changes of liver were observed in all treated group (Figure 3). A larger number of
melanomacrophage and Kupffer cells were observed in the liver tissue of tadpoles exposed to cold and heat shock
Moreover, blood congestion was also found in the liver of the tadpoles exposed to the cold shock (17°C) (Figure 3D).
Our results were consistent with the studies of Corsaro et al. (1990) which reported that melanomacrophages
in R. esculenta increased during the winter (about 5–10°C) and decreased during the summer (about 20°C –25°C).
The liver has long been well known to be the main organ for detoxification that could suffer serious morphological
alterations in tadpoles exposed to temperature shock. Therefore, its histological changes could be used as biomarkers
for environmental stress (Sichel et al., 1987). The hematopoietic organs of ectothermic animals commonly display
pigmented cells with phagocytic activity called melanomacrophage (Barni et al., 1999). They are focal accumulations
of pigmented macrophages in the liver. These may contain four types of brown to black pigments, namely melanin,
lipofuscin, ceroid, and hemosiderin/ferritin, which absorbs and neutralizes free radicals, cations, and other potentially
toxic agents derived from the degradation of phagocytized cellular material. Thus, it appears that these cells are
sensitive to stimuli as a way to adapt to stress conditions (Sayed, et al., 2015). In addition, Kupffer cells are
specialized resident macrophages in the liver and the activated kupffer cells appears to modulate acute hepatocyte
injury and chronic liver responses. They constitute the macrophage population of the body to encounter the bacteria
which derived from the gastrointestinal tract and transported to the liver via the portal vein. Thus, the increase in
the number of Kupffer cells in the liver tissue might be the response of the defense system when animals are in
contact with environmental stress. Moreover, it has been reported that blood congestion was present in the liver
tissues of giant spiny frog (Q. spinosa) under cold and heat stresses (Liu et al., 2018) which was consistent with
our results. It could be attributed to the destruction of hepatic erythrocytes and newly produced erythrocytes
accumulated in the liver (Maekawa et al., 2012).
Lipid peroxidation in tadpole tissue
Biochemical analysis of tadpole tissue in the present experiments indicated that cold shock (23°C)
(0.079±0.009 mM/mg protein) and heat shock groups (27°C) (0.050±0.002 mM/mg protein) induced a significant
increase in MDA level compared with the control group. (0.021±0.006 mM/mg protein). Moreover, the level of MDA
in the cold shock group (0.079±0.009 mM/mg protein) was significantly greater than those in the heat shock group
(0.050±0.002 mM/mg protein) (Figure 4).
This is in agreement with the findings of previous studies of Sahoo and Kara (2014) which report that
MDA level in the liver, kidney and brain tissues of the common Indian toad (B. melanostictus) exposed in cold
and heat shock were higher than those of the tadpoles exposed in constant temperature. In addition, the level of MDA
of the liver of goldfish (Carassius auratus) obtained in heat shock was higher than that was obtained in the constant
temperature. As a result, temperature change induces a significant increase in the MDA level in tissue of ectotherm
animals. (Abele et al., 2002). It is commonly known that the MDA is one of the important biochemical compounds
used as an indicative of reactive oxygen species (ROS) being generated from lipid peroxidation. In general, ROS,
which can be neutralized by a variety of antioxidants, are generated by cellular metabolism (Lushchak and
Bagnyukova, 2006). However, excessive ROS would damage various chemical and biological membranes and it
has been suggested as a cause of toxicity in several organs. Furthermore, oxidative stress has also been shown to
affect organ dysfunction and tissue damage. The cold and heat stress, which induces elevated thermogenesis by
enhanced substrate combustion along with increased oxygen consumption, has been reported to produce ROS,
which cause lipid peroxidation and play an important role in tissue injury (De Quiroga et al., 1991).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 58
Conclusion
In summary, we concluded that the temperature change caused decrease survival and growth of East
Asian Bullfrog (H. rugulosus). In addition, temperature shock caused oxidative stress that was demonstrated by
the increased MDA level and triggered the histopathological alterations in the livers of H. rugulosus tadpoles.
It could be suggested that the data obtained from this study would help to raise awareness of temperature shock in
farm-raised frogs and others wild amphibian species.
Acknowledgement
This work was supported by the Research and Researchers for Industries (RRi)(MSD6110063) and the
Graduate School, Chiang Mai University.
References
Abele, D., Heise, K., Pörtner, H.O., Puntarulo, S. 2002. Temperature-dependence of mitochondrial function and production
of reactive oxygen species in the intertidal mud clam Mya arenaria. Journal of Experimental Biology.205:
1831-1841.
Ates, B., Dogru, M.L., Gul, M., Erdogan, A., and Dogru, A.K. 2006. Protective role of caffeic acid phenethyl
esterin the liver of rats exposed to cold stress. Fundamental & Clinical Pharmacology 20: 283–289.
Barni, S., Bertone, V., Croce, A.C., and Bottiroli, G.C. 1999. Increase in liver pigmentation during natural
hibernation in some amphibians. The Journal of Anatomy. 195: 19-25.
Bellakhal, M., Neveu A., Missaoui, H., and Aleya, L. 2014. Effects of temperature, density and food quality
on larval growth and metamorphosis in the north African green frog Pelophylax saharicus. Journal of Thermal
Biology. 45: 81-86.
Bradford, M.A. 1976. Rapid and sensitive method for the quantification of microgram quantities of protein utilizing
the principle of proteindye binding. Annual Review of Biochemistry. 72: 248–254.
Buege, j.A., and Aust, S.D. 1976. Lactoperoxidase-catalyzed lipid peroxidation of microsomal and artificial
membranes. Biochimica et Biophysica Acta. 444: 192-201.
Corsaro, C., Scalia, M., Sinatra, F., and Sichel, G. 1990. Circannual Rhythm of the Melanin Content in Frog Liver
(Rana esculenta L.). Pigment Cell Research. 3: 120-122.
De Quiroga, G.B., López-Torres, M., Pérez-Campo, R., Abelenda, M., Paz Nava, M., and Puerta, M.L. 1991.Effect
of cold acclimation on GSH, antioxidant enzymes and lipid peroxidation in brown adipose tissue.
Biochemical Journal. 277: 289-292.
Diesmos, A.C., Brown, R.M., and Genevieve, V.A. 2004. Preliminary report on the amphibians and reptiles of
Balbalasang-Balbalan National Park, Luzon Island, Philippines. The Technical Journal of Philippine
Ecosystems and Natural Resources 13: 63-80.
Goldstein, J.A., Seckendorr Ho, K.V., and Hillyard S.D. 2017. The effect of temperature on development and
behaviour of relict leopard frog tadpoles. Conservation Physiology. 5: 1-8.
Gratwicke, B., Evans, M.J., Jenkins, P.T., Kusrini, M.D., Moore, R.D., Sevin, J., and Wildt, D. 2009. Is the
international frog legs trade a potential vector for deadly amphibian pathogens? Frontiers in Ecology
and the Environment. 8: 438-442.
Guderley, H. 2004. Locomotor performance and muscle metabolic capacities: impact of temperature and energetic
status. Comparative Biochemistry and Physiology. 139: 371-382.
Kern, P., Cramp, R.L., and Franklin, C.E. 2015, Physiological responses of ectotherms to daily temperature
variation. Journal of Experimental Biology. 218: 3068-3076.
Liu, Z.P., Gu, W.B., and Tu, D.D. 2018. Effects of both cold and heat stresses on the liver of giant spiny frog
Quasipaa spinosa: stress response and histological changes. Journal of Experimental Biology. 221:
1-11.
Lushchak, V.I., and Bagnyukova, T.V. 2006. Temperature increase results in oxidative stress in goldfish tissues.
Indices of oxidative stress. Comparative Biochemistry and Physiology. 143: 30–35.
Maekawa, S., Lemura, H., Kuramochi, Y., Nishikawa, H., Okui, T., Aizawa, T., and Kato, T. 2012. Hepatic
confinement of newly produced erythrocytes caused by low-temperature exposure in Xenopus
laevis. Journal of Experimental Biology. 215: 3087-3095.
Oza, G.M. 1990. Ecological effects of the frog’s legs trade. The Environmentalist 10 (1): 39-42.
Sayed, A.E.H., Elballouz, A.I., and Wassif, E.T. 2015. Histological and histochemical studies on the early
developmental stages of the egyptian toad Bufo regularis reuss. Journal of Animal Sciences. 5: 142- 156.
Sahin, E. 2004. Cold-stress-induced modulation of antioxidant defence: role of stressed conditions in tissue
injury followed by protein oxidation and lipid peroxidation. International Journal of Biometeorology.
48: 165-171

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 59
Sahoo, D.D., and Kara, T.C. 2014. Cold stress-induced lipid peroxidation and non-enzymatic antioxidant defense
in tissues of the common indian toad, Bufo melanostictus. Archives of Biological Sciences. 66:1303-1310.
Somero, G.N. 2002. Thermal physiology and vertical zonation of intertidal animals: optima, limits, and costs of living.
Integrative and Comparative Biology. 42: 780–789.
Seebacher, F., and Murray S.A. 2007. Transient receptor potential ion channels control thermoregulatory behaviour
in reptiles. Thermal Reception in Reptiles. 2: e281.
Sichel, G., C. Corsaro, M. Scalia, S. Sciuto., and E. Geremia. 1987. Relationship between melanin content and
superoxide dismutase (SOD) activity in the liver of various species of animals. Journal of Cellular
Biochemistry. 5: 123-128.
Wang, N., Shao, C., Xie, Z., and Ling, Y. 2012. Viability and changes of physiological functions in the tiger frog
(Hoplobatrachus rugulosus) exposed to cold stress. Journal of Guangzhou Physical Education Institute.11: 670-680.
Xu, C., Li, E., Suo, Y., Lu, M., and Qin, J.G. 2018. Histological and transcriptomic responses of two immune
organs, the spleen and head kidney, in Nile tilapia (Oreochromis niloticus) to long-term hypersaline stress.
Fish and Shell Fish Immunology. 76: 48-58.
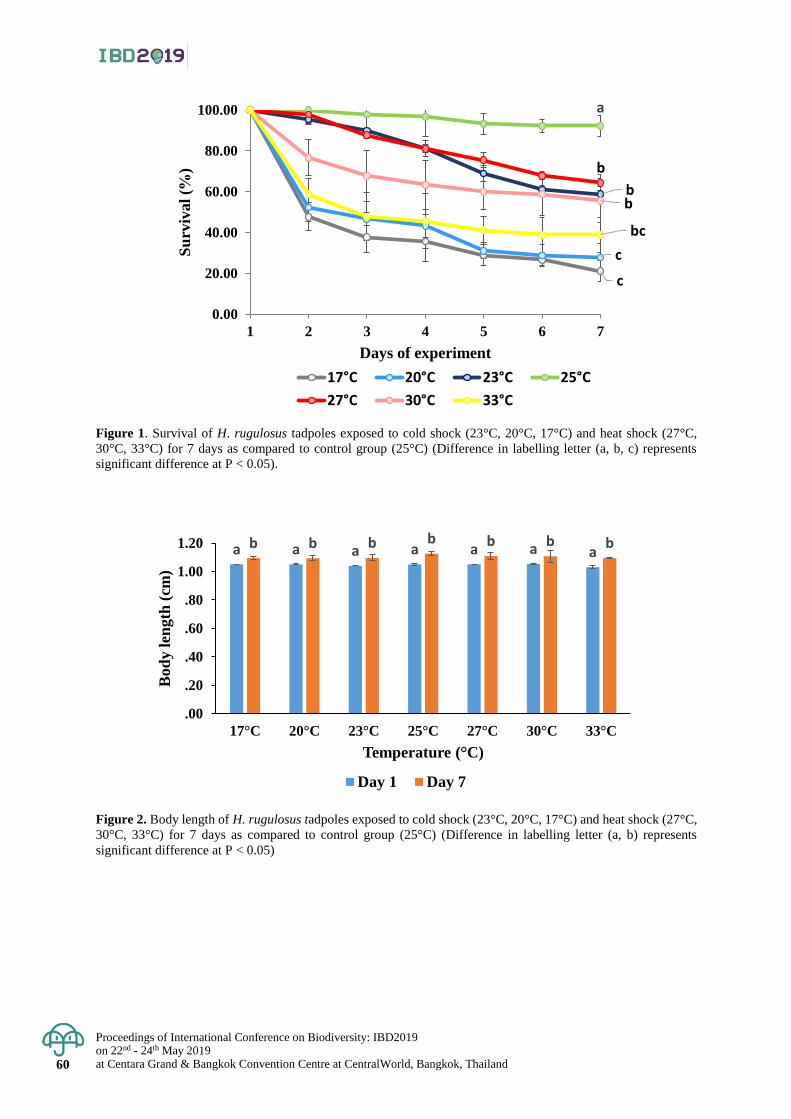
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 60
Figure 1. Survival of H. rugulosus tadpoles exposed to cold shock (23°C, 20°C, 17°C) and heat shock (27°C,
30°C, 33°C) for 7 days as compared to control group (25°C) (Difference in labelling letter (a, b, c) represents
significant difference at P < 0.05).
Figure 2. Body length of H. rugulosus tadpoles exposed to cold shock (23°C, 20°C, 17°C) and heat shock (27°C,
30°C, 33°C) for 7 days as compared to control group (25°C) (Difference in labelling letter (a, b) represents
significant difference at P < 0.05)
c
c
b
a
b
b
bc
0.00
20.00
40.00
60.00
80.00
100.00
1 2 3 4 5 6 7
Su
rviv
al
(%)
Days of experiment
17°C 20°C 23°C 25°C
27°C 30°C 33°C
a a a a a a ab b b b b b b
.00
.20
.40
.60
.80
1.00
1.20
17°C 20°C 23°C 25°C 27°C 30°C 33°C
Bod
y l
ength
(cm
)
Temperature (°C)
Day 1 Day 7
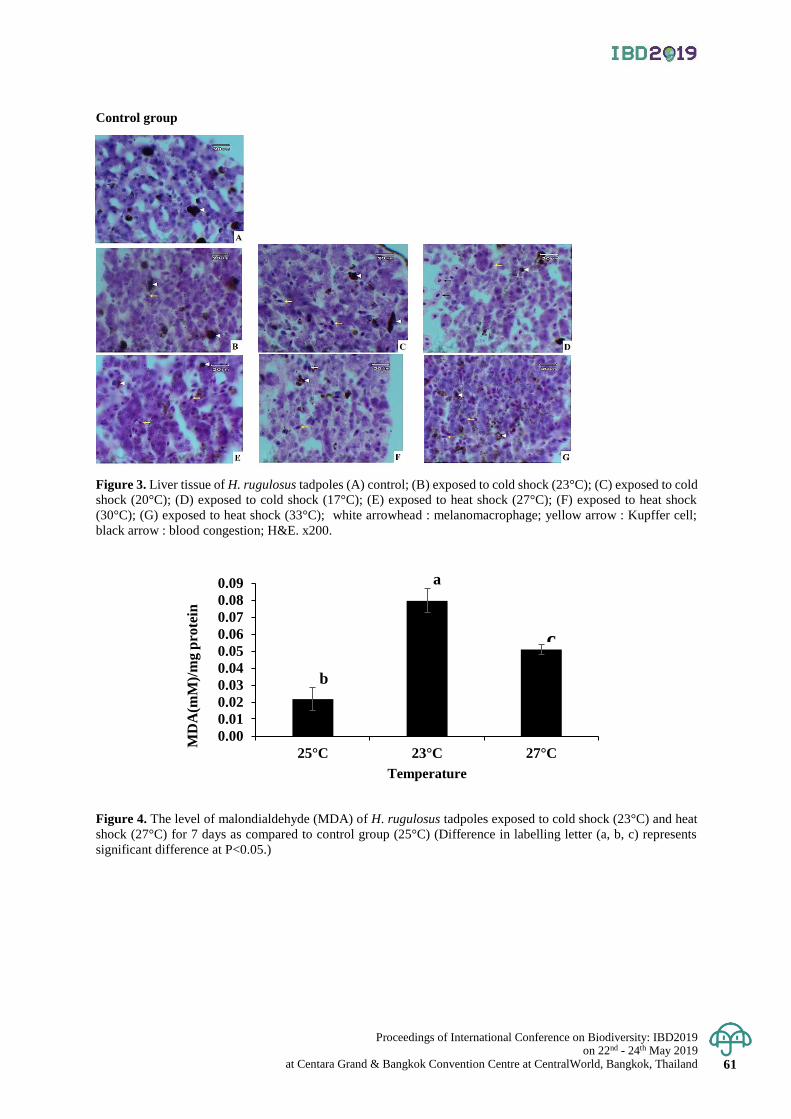
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 61
Control group
Cold shock group
Heat shock group
Figure 3. Liver tissue of H. rugulosus tadpoles (A) control; (B) exposed to cold shock (23°C); (C) exposed to cold
shock (20°C); (D) exposed to cold shock (17°C); (E) exposed to heat shock (27°C); (F) exposed to heat shock
(30°C); (G) exposed to heat shock (33°C); white arrowhead : melanomacrophage; yellow arrow : Kupffer cell;
black arrow : blood congestion; H&E. x200.
Figure 4. The level of malondialdehyde (MDA) of H. rugulosus tadpoles exposed to cold shock (23°C) and heat
shock (27°C) for 7 days as compared to control group (25°C) (Difference in labelling letter (a, b, c) represents
significant difference at P<0.05.)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
25°C 23°C 27°C
MD
A(m
M)/
mg p
rote
in
Temperature
a
b
c

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 62
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 62 - 66
Antioxidant activity and hair growth promoting activity of flavonoid extracts
from Phyllodium pulchellum and Uvaria rufa Blume on cultured mouse vibrissa hair follicles
Patcharida Penpakkul1, Kanokporn Saenphet2* and Supap Saenphet3 1Master’s Degree Program in Biology, Faculty of Science, Chiang Mai University, Muang District, Chiang Mai, Thailand
2Department of Biology, Faculty of Science, Chiang Mai University, Muang District, Chiang Mai, Thailand
*Corresponding author e-mail: [email protected]
Abstract: This study aims to determine antioxidant activity and hair growth promoting activity of flavonoid extracts
from the aerial parts of Phyllodium pulchellum and the stems of Uvaria rufa Blume on cultured mouse vibrissa hair
follicles. Flavonoid extracts of both plants were tested for their antioxidant activities using total antioxidant assay,
2,2’-azino-bis (3-ethylbenzthiazoline-6-sulphonic acid) (ABTS), 2,2-diphenyl-1-picrylhydrazyl (DPPH) and nitric
oxide radical scavenging assays. To investigate hair growth promotion, vibrissa follicles of mouse were isolated and
cultured in 95 % CO2, 5 % O2 and 37⁰C. Cultured vibrissa follicles were treated with flavonoid extracts of P. pulchellum
(PPE) and U. rufa (URE) at concentrations of 0.01-20 µg/ml for three days. The length of hair follicles was
measured every 24 hours for three days. Total flavonoids contents of PPE and URE were 10.14 ± 0.20, 10.28 ±
0.60 mg quercetin per extract, respectively. We found that PPE and URE had total antioxidant activities at 10.53 ± 0.26
and 11.29 ± 0.04 mg ascorbic acid equivalent per gram extract, respectively. Moreover, PPE and URE markedly
exhibited antioxidant activities with the IC50 values at 1.67 mg/ml and 2.44 mg/ml for ABTS assay, and 1.34 mg/mL
and 2.45 mg/ml for DPPH assay, and 0.44 mg/ml and 0.34 mg/ml for nitric oxide radical scavenging assay. PPE
at 1 µg/ml significantly increased the length of hair follicles at 24 and 48 hours while URE at 0.01-1 µg/ ml
significantly elongated hair follicles at 48 and 72 hours when compared to those of controls. The results suggest that
the flavonoid extracts from P. pulchellum and U. rufa are the new sources of antioxidants and the potential promoting
hair growth agents.
Keywords: Antioxidant, hair growth, Phyllodium pulchellum, Uvaria rufa Blume
Introduction
Hair loss is a concern problem which can occur in both men and women. This problem brings loss of
self-esteem and mental health problems (Williamson et al., 2001). Although there are effective drugs for treating hair loss
which are finasteride and minoxidil, they contribute several adverse side effects. Finasteride caused male infertility
and gynecomastia while minoxidil can lead to skin irritation, itching and dryness and hypotension (Chilba et al., 2011;
Ramot et al., 2009; Rossi et al., 2012; Mehta et al., 1975). Due to the limited usage of synthetic drugs, natural product
has become attractive. Hair growth promoting activities of various plants; Centella asiatica Linn., Panax ginseng,
Avicennia marina and Thujae occidentalis were previously reported (Saansoomchai et al. , 2018; Matsuda et al. ,
2003; Jain et al., 2015; Park et al., 2003). The major phytochemical constituent in these plants is flavonoids, which
is well-known for its antioxidant properties. Since oxidative stress is related to hair loss, consumption of antioxidants
can prevent the miniaturization of hair. Over-production of free radicals in scalp skin can trigger dermal papilla cell
apoptosis, eventually leads to hair shedding (Trüeb, 2015). Therefore, natural antioxidant treatment for maintaining
healthy scalp is necessary.
Phyllodium pulchellum and Uvaria rufa Blume are distributed in Thailand and used in traditional
medicine as remedy for hair loss. P. pulchellum was reported to have hepatoprotective effect while U. rufa was
reported as effective treatment against benign prostatic hyperplasia in experimental animals (Fan et al. , 2018;
Buncharoen et al. , 2016) . However, there is no evidence for hair growth promoting activity of these two plants.
Thus, this study aims to investigate hair growth promoting activity as well as antioxidant of extracts from P.
Pulchellum and U. rufa to confirm their traditional usage.
Materials and Methods
Reagents
2,2’-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid), 2,4-dinitrophenylhydrazine (DNPH), basal medium,
bovine calf serum, bovine pituitary extract, fibroblast growth factor, gallic acid and insulin were purchased from
Sigma Aldrich (MO, USA). Ammonium molybdate was obtained from Unilab (Sydney, Australia). Ascorbic acid
was purchased from Prolabo (Leuven, Belgium). Penicillin/streptomycin was obtained from Merck (Darmstadt, Germany).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 63
Plant materials
The aerial parts of P. pulchellum were collected in Chiang Mai Province and the stems of U. rufa were
obtained from Buriram Province, Thailand.
Extraction
All plant materials were chopped, cleaned and dried. The dried sample was defatted with petroleum ether
using a Soxhlet extractor at 40-60⁰C. Then, the residue was extracted with ethyl acetate. The ethyl acetate extracts
were evaporated by a rotary evaporator at 50⁰C (Peleg et al., 1991) to obtain the flavonoid extracts of P. Pulchellum
(PPE) and U. rufa (URE). Total flavonoid contents were determined by using aluminium chloride solution at 420 nm
(Ordonez et al. , 2006) . The flavonoids in PPE and URE were expressed as mg quercetin equivalent per gram
extract (mgQE/g extract).
Measurement of antioxidant capacity
ABTS radical scavenging activity
The scavenging of ABTS radical was tested according to the method described by Re et al (1998). Briefly,
the working ABTS solution was prepared by mixing 7 mM ABTS solution with 2. 45 mM potassium persulfate.
The mixture was allowed to react in the dark at room temperature for 12 h. The ABTS•+ solution was then diluted
with deionized water to obtain an absorbance of 0.7 ± 0.02 at 734 nm. Different concentrations of each sample
were mixed with 1 ml of the ABTS+. The absorbance was measured at 734 nm at 0 (initial absorbance) and 1 (final
absorbance) minute of incubation. Gallic acid was used as the standard control. The inhibition of ABTS•+ was calculated
by using the following equation:
% inhibition of ABTS•+ = [Abs (initial)- Abs(final) / Abs (initial)] x 100
The results were expressed as median inhibitory concentration (IC50) value.
DPPH radical scavenging activity
The DPPH assay was done according to the method of Susanti et al. (2007). 100 µl of each sample was added
to 2 ml of 0.1 mM DPPH·solution. The mixture was vigorously shaken and the absorbance (initial absorbance) of the mixture
was recorded immediately at 517 nm. Then, it was incubated in the dark for 30 min at room temperature and the final
absorbance at 517 nm of the mixture was measured. Gallic acid was used as the standard control. The DPPH·scavenging
ability was calculated as:
% inhibition of DPPH· = [Abs (initial)- Abs(final) / Abs (initial)] x 100
The IC50 value of each sample as well as antioxidant standard were calculated. Each sample was performed in triplicate.
Nitric oxide scavenging activity
The nitric oxide scavenging activity assay was conducted according to the procedure described by
Marcocci et al. (1994). Sodium nitroprusside (10 mM) in phosphate buffer saline was prepared to produce nitric oxide
radicals, which was measured by using Griess reagent. 500 µl of sodium nitroprusside solution was then mixed
with 1 ml of various concentrations of each extract. The mixture was incubated at room temperature for 180 min.
After incubation, 500 µl of Griess reagent was added. Gallic acid was used as the standard positive control. The absorbance
of the mixture was taken at 546 nm.
Total antioxidant capacity
The phosphomolybdenum method was used to investigate the total antioxidant capacity in PPE and URE
extracts (Oyaizu, 1986. Phosphomolybdenum reagent was prepared by mixing 0.6 M sulfuric acid, 28 mM sodium
phosphate and 1% of ammonium molybdate. The reagent was then mixed with 500 µl of each sample. The mixtures
were incubated at 95⁰C for 10 min. The absorbance of the mixture was subsequently measured at 695 mm. The results
were expressed as mg ascorbic acid equivalent per gram extract (mgAAE/g extract)
Isolation and culture of mouse vibrissa follicles
For isolation experiment, five-week-old ICR mice were isolated as previously described ( Jindo et al. ,
1994). Briefly, the mystacial pads were removed from upper lip region. Intact vibrissa follicles were then carefully
disaggregated under a stereomicroscope using a scalpel and tweezers. Isolated vibrissa follicles were placed in
24-well plates containing 1 ml of basal medium supplemented with 0.002 ml/ml bovine pituitary extract, 0.5µg/ml
bovine insulin, 0.05 ml/ml calf serum, 0.5 ng/ml fibroblast growth factor, 50 U/ml penicillin and 50 µg/ ml
streptomycin ( Junlatat and Sripanidkulchai, 2014) . The follicles were cultured in an incubator with 5% CO2 at
37⁰C. For each assay, PPE and URE were diluted with DMSO and added to the medium to yield various
concentrations (0.001 - 10 µg/ml). DMSO (0.1%) was used as a control. The medium was discarded and replaced
every 24 h. During the period of culture (72 h), the cells were photographed at 24, 48 and 72 h using a stereo microscope.
The difference of the length of vibrissa hair follicles were calculated using ImageJ software and expressed as
mean ± S.D.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 64
Statistical analysis
All values were expressed as mean ± S.D. of at least three samples. The significant differences among
experimental groups were determined by one-way analysis of variance (ANOVA), followed by Duncan’s test using
SPSS version 16. P-values less than 0.05 were regarded as significant.
Result & Discussion
Antioxidant activity of PPE and URE
Oxidative stress is the disturbance which leads to damaging of cellular structures and may be a factor of
hair loss (Trüeb, 2015) . The association between oxidative stress and hair loss has been reported by Bahta et al.
(2008) that the dermal papilla cells were susceptible to oxidative stress and resulting in miniaturization of hair
follicles. In addition, Beoy et al. (2010) have suggested that consuming the supplement containing antioxidants
can decrease oxidative stress in the hair scalp as well as increase the numbers of hairs.
The low IC50 values of all parameters obtained from our results indicated that PPE and URE extracts
exhibited the ability to scavenge free radicals. PPE and URE markedly performed antioxidant activities with the
IC50 values at 1.67 mg/ml and 2.44 mg/ml for ABTS assay, and 1.34 mg/mL and 2.45 mg/ml for DPPH assay,
respectively. Moreover, URE also exhibited excellent nitric oxide scavenging activity, especially URE which IC50
value was not much higher than that of gallic acid (0.34 mg/ml for URE and 0.23 mg/ml for gallic acid) . PPE,
however, exhibited lower capacity to scavenge nitric oxide than URE by showing IC50 value at 0.44 mg/ml (Table
1). Determination of total antioxidant capacity of PPE and URE expressed values with 10.53 ± 0.26 and 11.29 ±
0.04 mg AAE/g extract, respectively. Total flavonoid contents of PPE and URE were 10.14 ± 0.20, 10.28 ± 0.60
mgQE/g extract. The potent antioxidant activities of PPE and URE found in our study might contribute to their
flavonoid compounds.
As previous study reported that flavonoids are powerful antioxidants which can reduce oxidative stress
in the hair scalp (Pietta, 2000; Beoy et al., 2010). Thus, flavonoids in P. pulchellum and U. rufa may be the agents
that reduce the free radicals in the hair scalp. To ensure hair growth promoting activity of PPE and URE via
decreasing oxidative stress, determination of hair growth promoting activity in vivo needs to be further evaluated.
Elongation of vibrissa hair follicles
Flavonoids could enhance hair quality not only by decreasing free radicals in the hair scalp, but also promoting
elongation of hair follicles. The previous study reported that green tea extract containing flavonoid promoted hair
growth by inducing elongation of hair follicles and inhibiting apoptosis of dermal papilla cells (Kwon et al., 2007).
In addition, flavonoids contained in Panax ginseng performed significant stimulation on hair growth of vibrissa
follicles after 48 h of culture (Matsuda et al., 2003).
In this study, the vibrissa follicles treated with PPE at 1 µg/ml significantly increased the elongation of
hair follicles after 24 and 48 h of culture (128.02 ± 21.97 and 116.81 ± 28.05 µm, respectively) when compared
with the control (Figures 1, 3) . In the URE group, the follicles were significantly longer than those treated with
vehicle control after 48 and 72 h of culture (Figure 3). The follicles treated with URE at 0.01, 0.1 and 1 µg/ml for
24 h expressed elongation with values 123.76 ± 69.58, 136.47 ± 12.46 and 115.04 ± 26.20 µm, respectively. For
72 h of culture, the URE treatment at 0.01, 0.1 and 1 µg/ml enhanced the hair growth by 111.38 ± 17.82, 113.47
± 2.83 and 149.83 ± 25.59 µm, respectively (Figure 2) . Hence, these results suggested that PPE and URE have
potential hair growth promoting activity in vitro resulting from the presence of flavonoids as the major constituents.
Conclusion
In conclusion, the flavonoid extracts from P. pulchellum and U. rufa are potent antioxidants and able to
promote hair growth by inducing hair elongation. The antioxidant capacity of these two extracts is recommended
as the mechanism underlying their hair growth promoting activity.
Acknowledgement
This research was supported by the Research and Researchers for Industries, Thailand Research Fund
(MSD61I0046). We also gratefully acknowledge Graduate School, Chiang Mai University.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 65
References
Bahta, A. W., Farjo, N., Farjo, B., and Philpott, M. P. 2008. Premature senescence of balding dermal papilla cells
in vitro is associated with p16INK4a expression. Journal of Investigative Dermatology 128(5): 1088-1094.
Beoy, L. A., Woei, W. J., and Hay, Y. K. 2010. Effects of tocotrienol supplementation on hair growth in human
volunteers. Tropical life sciences research, 21(2), 91-99.
Buncharoen, W., Saenphet, K., Saenphet, S., and Thitaram, C. 2016. Uvaria rufa Blume attenuates benign prostatic
hyperplasia via inhibiting 5α- reductase and enhancing antioxidant status. Journal of ethnopharmacology
194: 483-494.
Chiba, K. , Yamaguchi, K. , Li, F. , Ando, M. , and Fujisawa, M. 2011. Finasteride-associated male infertility.
Fertility and sterility 95(5): 1786e9-1786e11.
Fan, Y. C., Yue, S. J., Guo, Z. L., Xin, L. T., Wang, C. Y., Zhao, D. L., Gaun, H.S. and Wang, C. Y. 2018. Phytochemical
Composition, Hepatoprotective, and Antioxidant Activities of Phyllodium pulchellum (L.) Desv. Molecules
23(6): 1361.
Jain, R., Monthakantirat, O., Tengamnuay, P., and De-Eknamkul, W. 2015. Identification of a new plant extract for
androgenic alopecia treatment using a non-radioactive human hair dermal papilla cell-based assay. BMC
complementary and alternative medicine 16(1):18.
Jindo, T., Tsuboi, R., Imai, R., Takamori, K., Rubin, J. S., and Ogawa, H. 1994. Hepatocyte growth factor/scatter
factor stimulates hair growth of mouse vibrissae in organ culture. Journal of investigative dermatology
103(3): 306-309.
Junlatat, J. , and Sripanidkulchai, B. 2014. Hair Growth‐Promoting Effect of Carthamus tinctorius Floret Extract.
Phytotherapy research 28(7): 1030-1036.
Kwon, O. S., Han, J. H., Yoo, H. G., Chung, J. H., Cho, K. H., Eun, H. C., and Kim, K. H. 2007. Human hair growth
enhancement in vitro by green tea epigallocatechin-3-gallate (EGCG). Phytomedicine 14(7-8): 551-555.
Marcocci, L., Packer, L., Droy-Lefaix, M. T., Sekaki, A., and Gardès-Albert, M. 1994. Antioxidant action of Ginkgo
biloba extract EGb 761. In Methods in enzymology 234: 462-475.
Matsuda, H. , Yamazaki, M. , Asanuma, Y. , and Kubo, M. 2003. Promotion of hair growth by ginseng radix on
cultured mouse vibrissal hair follicles. Phytotherapy Research 17(7): 797-800.
Matsuda, H., Yamazaki, M., Asanuma, Y., and Kubo, M. 2003. Promotion of hair growth by Ginseng radix on cultured
mouse vibrissal hair follicles. Phytotherapy Research 17(7): 797-800.
Mehta, P. K., Mamdani, B., Shansky, R. M., Mahurkar, S. D., and Dunea, G. 1975. Severe hypertension: Treatment
with minoxidil. Jama 233(3): 249-252.
Ordonez, A. A. L., Gomez, J. D., and Vattuone, M. A. 2006. Antioxidant activities of Sechium edule (Jacq.) Swartz
extracts. Food chemistry 97(3): 452-458.
Oyaizu, M. 1986. Studies on products of browning reaction: antioxidative activity of products of browning reaction
prepared from glucosamine. The Japanese Journal of Nutrition and Dietetics 44:307-315.
Park, W. S., Lee, C. H., Lee, B. G., and Chang, I. S. 2003. The extract of Thujae occidentalis semen inhibited 5α-
reductase and androchronogenetic alopecia of B6CBAF1/ j hybrid mouse. Journal of dermatological
science 31(2); 91-98.
Peleg, H., Naim, M., Rouseff, R. L., and Zehavi, U. 1991. Distribution of bound and free phenolic acids in oranges
(Citrus sinensis) and grapefruits (Citrus paradisi). Journal of the Science of Food and Agriculture 57(3): 417-426.
Pietta, P. G. 2000. Flavonoids as antioxidants. Journal of natural products 63(7): 1035-1042.
Ramot, Y., Czarnowicki, T., and Zlotogorski, A. 2009. Finasteride induced Gynecomastia: Case report and Review
of the Literature. International journal of trichology.
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., and Rice-Evans, C. 1999. Antioxidant activity applying
an improved ABTS radical cation decolorization assay. Free radical biology and medicine 26(9-10): 1231-1237.
Rossi, A., Cantisani, C., Melis, L., Iorio, A., Scali, E., and Calvieri, S. 2012. Minoxidil use in dermatology, side effects
and recent patents. Recent patents on inflammation & allergy drug discovery 6(2): 130-136.
Saansoomchai, P., Limmongkon, A., Surangkul, D., Chewonarin, T., and Srikummool, M. 2018. Enhanced VEGF
Expression in Hair Follicle Dermal Papilla Cells by Centella asiatica Linn. Chiang Mai University
Journal of Natural Sciences. 17(1): 25-37.
Susanti, D., Sirat, H. M., Ahmad, F., Ali, R. M., Aimi, N., and Kitajima, M. 2007. Antioxidant and cytotoxic flavonoids
from the flowers of Melastoma malabathricum L. Food Chemistry 103(3): 710-716.
Trüeb, R. M. 2015. The impact of oxidative stress on hair. International journal of cosmetic science 37: 25- 30.
Willianson, D. , Gonzalez, M. and Finlay, AY. 2001. The effect of hair loss on quality of life. Journal of the
European Academy of Dermatology and Venereology15(2): 137-139.
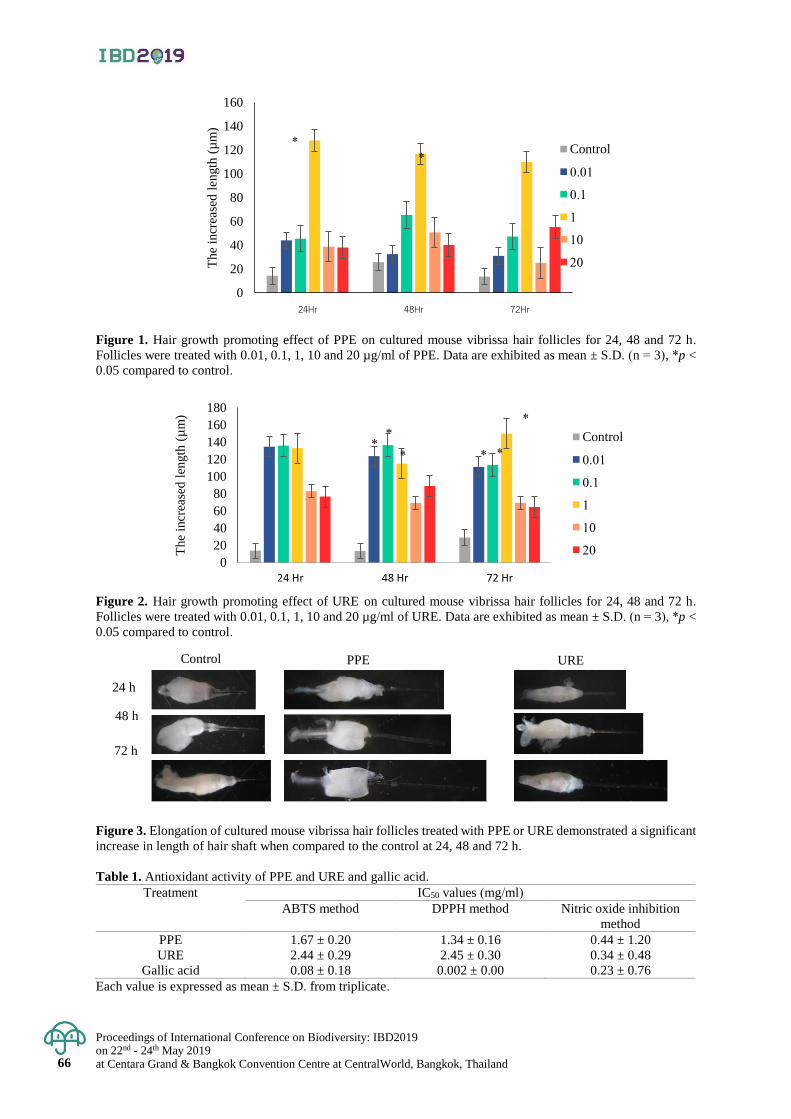
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 66
Figure 1. Hair growth promoting effect of PPE on cultured mouse vibrissa hair follicles for 24, 48 and 72 h.
Follicles were treated with 0.01, 0.1, 1, 10 and 20 µg/ml of PPE. Data are exhibited as mean ± S.D. (n = 3), *p <
0.05 compared to control.
Figure 2. Hair growth promoting effect of URE on cultured mouse vibrissa hair follicles for 24, 48 and 72 h.
Follicles were treated with 0.01, 0.1, 1, 10 and 20 µg/ml of URE. Data are exhibited as mean ± S.D. (n = 3), *p <
0.05 compared to control.
Figure 3. Elongation of cultured mouse vibrissa hair follicles treated with PPE or URE demonstrated a significant
increase in length of hair shaft when compared to the control at 24, 48 and 72 h.
Table 1. Antioxidant activity of PPE and URE and gallic acid.
Treatment IC50 values (mg/ml)
ABTS method DPPH method Nitric oxide inhibition
method
PPE 1.67 ± 0.20 1.34 ± 0.16 0.44 ± 1.20
URE 2.44 ± 0.29 2.45 ± 0.30 0.34 ± 0.48
Gallic acid 0.08 ± 0.18 0.002 ± 0.00 0.23 ± 0.76
Each value is expressed as mean ± S.D. from triplicate.
0
20
40
60
80
100
120
140
160
24Hr 48Hr 72Hr
The
incr
ease
d l
ength
(µ
m)
Control
0.01
0.1
1
10
20
0
20
40
60
80
100
120
140
160
180
24 Hr 48 Hr 72 Hr
The
incr
ease
d l
ength
(µ
m)
Control
0.01
0.1
1
10
20
**
* *
*
*
*
*
Control PPE URE
24 h
48 h
72 h

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 67
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 67 - 72
Chemical composition and antibacterial activities of essential oils of Lavandula × hybrid
‘boysemberry ruffles’, L. pedunculata ‘princess’ and L. × hybrid ‘high five purple’
Suchawadee Insawang1 and Patcharee Pripdeevech1,2* 1School of Science, Mae Fah Luang University, Muang District, Chiang Rai, Thailand
2Center of Chemical Innovation for Sustainability (CIS), Mae Fah Luang University, Muang District, Chiang Rai, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Essential oils of Lavandula plants are used in perfumery and therapeutic applications. They are mainly
planted in Mediterranean area and several regions in Asia. According to its beneficial property, chemical composition
and bioactivity of various species of Lavandula plants have been studied. Thus, this study aimed to extract the essential
oils of three species from Lavandula plants grown in Thailand including L. × hybrid ‘boysemberry ruffles’, L. pedunculata
‘princess’ and L. × hybrid ‘high five purple’. The yield of essential oils obtained among these samples were 0.11-0.24 %w/w.
The chemical composition of all essential oils were investigated using gas chromatography- mass spectrometry.
A total of 89 volatile compounds were identified. Four main compounds including 1,8-cineole, fenchone, camphor
and α-pinene were detected in all samples with different contents. For the antibacterial activity assay, the experiment
was performed by agar disc diffusion method against four Gram-negative bacteria (Klebsiella pneumonia, Salmonella
typhimurium, Escherichia coli and Pseudomonas aeruginosa) and five Gram-positive bacteria (Staphylococcus
epidermidis, S. aureus, Bacillus subtilis, Enterococcus faecium and Streptococcus pyogens). The result showed that
the great antibacterial activity was obtained from the essential oil of L. pedunculata ‘princess’ compared to those
found in essential oils of L. × hybrid ‘boysemberry ruffles’ and L. × hybrid ‘high five purple’, respectively. Different
antibacterial activities among these samples could be correlated to major components such as 1,8-cineole, fenchone
and camphor. It was indicated that essential oil of L. pedunculata ‘princess’ may be used as natural antibacterial agent in
therapeutic applications.
Keywords: Antibacterial, chemical composition, essential oil, gas chromatography-mass spectrometry, Lavandula.
Introduction
Essential oils from plant species ‘Lavandula’ have been receiving many interests due to their applications
in many industries such as food, pharmaceutical, and cosmetic. In cosmetic industry, the oils can be used in the a production
of perfumes, colognes, body wash and other cosmetics (Cavanagh and Wilkinson, 2005; Lesage-Meessen et al., 2015).
The use of lavender essential oil might depend on their cultivar source due to the variety of composition. The volatiles
constituents in the extracted oil are feasibly depend on many factors such as cultivar and extraction method. It has
been found that linalool, linalyl acetate, 1,8-cineole, β-ocimene (both cis- and trans-), terpinen-4-ol and camphor are
the main constituents of lavender oil. (Cavanagh and Wilkinson, 2002; Costa et al., 2012). Lavandula essential oil
has the antimicrobial activity against various fungi and bacteria causing skin infection (Kunicka‐Styczyńska et al., 2009).
It was found that there is a relation between volatiles in the essential oil and antimicrobial strength. Linalool, a volatile
components in lavender essential oil, has a strong antimicrobial strength (Guo et al., 2018). In other studies, it is
reported that lavender oil is used for relieving symptoms of psoriasis, dermatitis, and eczema (Cavanagh and
Wilkinson, 2002; Matos et al., 2009). Another factor that affect quality and yield of the oil is an extraction method.
Hydrodistillation is chosen to extract essential oils in this study due to it is a popular method using less heat and
retaining most of the volatiles in the sample (Costa et al., 2012). Therefore, the aims of this study are to extract
essential oils from selected species, analyze volatile compounds using gas chromatography-mass spectrometry
(GC-MS) and screen their antibacterial activities.
Materials and Methods
Plant materials
The sample Lavandula plants including L. pedunculatas ‘princess’, L. x hybrid ‘high five purple’ and L.
x hybrid ‘boysemberry ruffles’ were collected from Angkhang Royal Agricultural Station, Chiang Mai, Thailand
on April 2018.
Extraction of essential oils
Fresh arial part of each sample (250 g) was subjected to hydrodistillation. The essential oils were extracted
for three h. The obtained essential oils were dried over anhydrous Na2SO4, and diluted with dichloromethane
(1:100 v/v) prior subjecting to GC-MS injection port.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 68
Analysis of volatile compounds
The volatile components were also identified by GC-MS using a Hewlett–Packard 5973 MSD apparatus
(Agilent 5973 network mass selective detector) from Agilent Technologies, USA with fused-silica capillary columns.
The temperature program was started at 60°C and raised up to 220°C with a rate of 3°C/min. Helium was used as
carrier gas with a flow rate of 1 mL/min. Injector and detector temperatures were set at 250 °C and 280°C, respectively.
Electron impact ionization was employed with electron energy of 70 eV. The acquisition was performed in scanning
mode (mass range m/z 35-300 u). Identification of all volatile compounds was performed by comparing their mass
spectra to those obtained from NIST 05 and Wiley 7N libraries and literature (Angioni et al., 2006; Adams, 2017).
Moreover, linear retention indices correlating with C9-C18 n-alkanes were used to confirm identification.
Antibacterial activity assay
Antibacterial activity was performed using the paper disc diffusion method of Zaidan (2005). The bacteria
strains including Staphylococcus aureus ATCC 25923, S. epidermidis ATCC 12228, S. pyrogens DMST 17020,
Bacillus subtilis TISTR 008, Enterococcus faecium ATCC 29212, Escherichia coli ATCC 25922, Klebsiella
pneumoniae ATCC 700603, Pseudomonas aeruginosa ATCC 27853 and Salmonella typhimurium ATCC 13311 were
obtained from the Department of Medical Science, Ministry of Health, Bangkok, Thailand. All strains were subcultured
in Müller-Hinton Broth (YM, Difco, USA) at 37°C for 24 h. All pathogenic bacterial was further spread in Müller-
Hinton agar. Different concentrations of essential oils were prepared by two-fold dilution method using hexane
as solvent until the final concentration was obtained as follows: 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78 and 0.39 mg/mL.
Thirty microliters of each essential oil sample were dropped on a sterilized 6 mm diameter paper disc (WhatmanTM, USA),
and the Petri dish plates were placed in culture plates prior incubating at 37°C for 24 h. Hexane and chloramphenicol
antibiotic were used as negative and positive control, respectively. The result was reported as minimum inhibition
concentration (MIC) and zone inhibition diameter. Each experiment was carried out in triplicate.
Result & Discussion
Essential oils from three Lavandula species, L. pedunculatas ‘princess’ , L. x hybrid ‘high five purple’
and L. x hybrid ‘boysemberry ruffles’ were extracted by hydrodistillation method. All essential oils were clear
with pale yellow and strong piquant smell. All essential oils yielded ranging between 0.11-0.24 %w/w. The results
of volatile constituents in the essential oils from Lavandula species analyzed by GC-MS are listed in Table 1.
A total of eighty-nine volatile constituents were detected in the essential oils extracted from the selected Lavandula species.
Forty-five compounds were observed in the essential oil of L. x hybrid ‘boysemberry ruffles’ while the essential
oils from L. pedunculatas ‘princess’ and L. x hybrid ‘high five purple’ contained 42 and 38 volatile compounds,
respectively. It was also found that 1,8-cineole, fenchone, camphor and α-pinene were the main compounds in all
Lavandula species with different contents. This might be the result of their genetic diversity. Camphor found to be
the most abundant volatile in L. pedunculata ‘princess’ was similar to those found previously in the study of
Gonçalves and Romano (2013). Camphor is known as antimicrobial (Zuccarini, 2009), thus the essential oil with
the highest amount of camphor could be expected to provide the greatest antibacterial activity in this study. As show
in Table 2. the essential oil of L. pedunculatas ‘princess’ showed the best result in the microbial inhibitory
activities, considering from its ability to inhibit microbial growth by MIC and zone inhibition diameter. It was
able to inhibit most of the bacterial pathogens used in this study. S. aureus was evaluated as the most sensitive
bacteria with MIC and zone inhibition diameter of L. pedunculatas ‘princess’ essential oil being 0.39 mg/mL and
7.37 mm, respectively. On the other hand, K. pneumoniae and B. subtilis were considered to be less sensitive
when being tested with all essential oils. The greatest antibacterial activity of L. pedunculatas ‘princess’ might be
due to the presence of major volatile compounds, such as 1,8-cineole, fenchone, camphor (Vakilian et al., 2011).
Conclusion
This study reports the essential oil composition and antibacterial activities of three different Lavandula
species grown in Thailand. Each Lavandula species appeared to have unique volatile profiles and antibacterial
activities. The essential oil of L. pedunculata ‘princess’ showed a broad spectrum of antibacterial activities,
suggesting thatit may be used as antimicrobial agent in the food, cosmetic and therapeutic applications.
Acknowledgement
The authors would like to thank Angkhang Royal Agricultural Station for providing access to the lavender
samples.The author thanks Mae Fah Luang University for supporting of financial and GC-MS.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 69
References
Adams, R.P. 2017. Identification of essential oil components by gas chromatography/mass spectroscopy, ed. 4.1.
Allured Publishing Corporation, Illinois, USA.
Angioni, A., Barra, A., Coroneo, V., Dessi, S., and Cabras, P. 2006. Chemical composition, seasonal variability,
and antifungal activity of Lavandula stoechas L. ssp. stoechas essential oils from stem/leaves and
flowers. Journal of Agricultural and Food Chemistry 54(12):4364-4370.
Cavanagh, H., and Wilkinson, J. 2002. Biological activities of lavender essential oil. Phytotherapy Research
16(4):301-308.
Cavanagh, H. M., and Wilkinson, J. M. 2005. Lavender essential oil: a review. Australian Infection Control 10(1):35-37.
Costa, P., Grosso, C., Gonçalves, S., Andrade, P. B., Valentão, P., Bernardo-Gil, M. G., and Romano, A. 2012.
Supercritical fluid extraction and hydrodistillation for the recovery of bioactive compounds from Lavandula viridis
L’Hér. Food Chemistry 135(1):112-121.
Gonçalves, S., and Romano, A. 2013. In vitro culture of lavenders (Lavandula spp.) and the production of secondary
metabolites. Biotechnology Advances 31(2):166-174.
Guo, J.-j., Gao, Z.-p., Xia, J.-l., Ritenour, M. A., Li, G.-y., and Shan, Y. 2018. Comparative analysis of chemical
composition, antimicrobial and antioxidant activity of citrus essential oils from the main cultivated
varieties in China. LWT 97:825-839.
Kunicka‐Styczyńska, A., Sikora, M., and Kalemba, D. 2009. Antimicrobial activity of lavender, tea tree and lemon
oils in cosmetic preservative systems. Journal of Applied Microbiology 107(6):1903-1911.
Lesage-Meessen, L., Bou, M., Sigoillot, J.-C., Faulds, C. B., and Lomascolo, A. 2015. Essential oils and distilled
straws of lavender and lavandin: a review of current use and potential application in white biotechnology.
Applied Microbiology and Biotechnology 99(8):3375-3385.
Matos, F., Miguel, M. G., Duarte, J., Venâncio, F., Moiteiro, C., Correia, A. I., and Pedro, L. G. 2009. Antioxidant
capacity of the essential oils from Lavandula luisieri, L. stoechas subsp. lusitanica, L. stoechas subsp.
lusitanica x L. luisieri and L. viridis grown in Algarve (Portugal). Journal of Essential Oil Research 21(4):327-336.
Vakilian, K., Atarha, M., Bekhradi, R., and Chaman, R. 2011. Healing advantages of lavender essential oil during
episiotomy recovery: a clinical trial. Complementary therapies in clinical practice, 17(1), 50-53.
Zuccarini, P. 2009. Camphor: risks and benefits of a widely used natural product. Journal of Applied Sciences
and Environmental Management, 13(2).
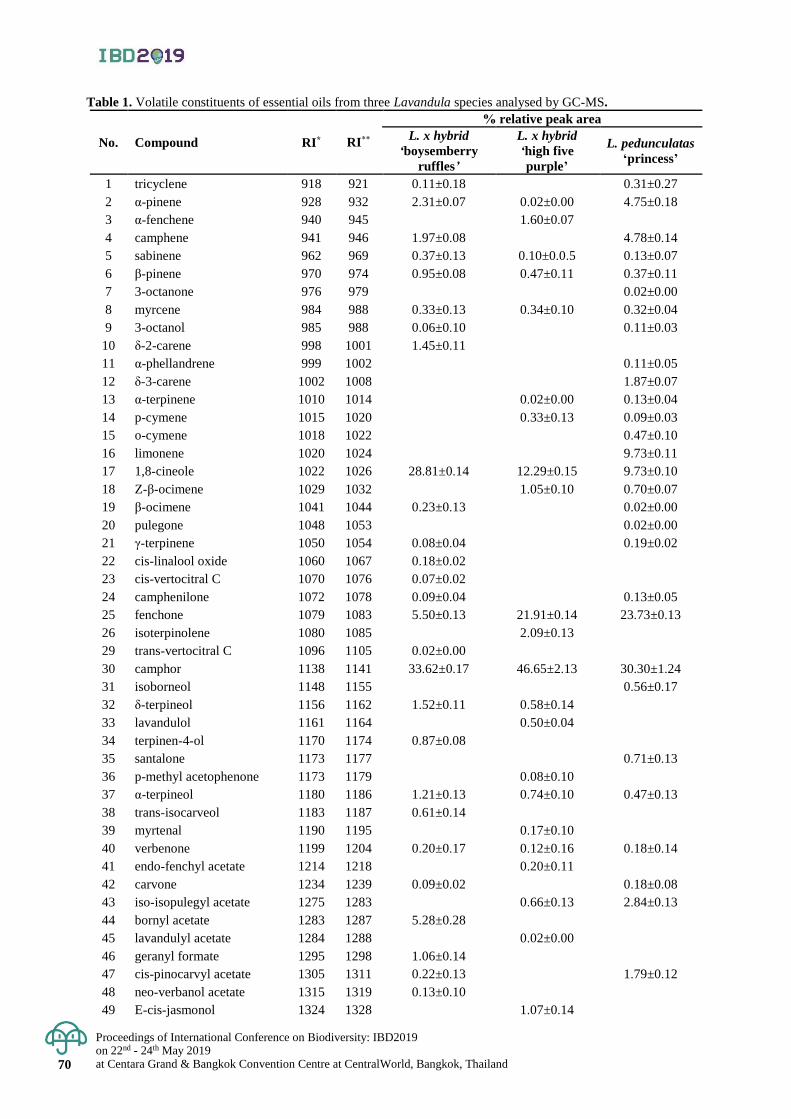
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 70
Table 1. Volatile constituents of essential oils from three Lavandula species analysed by GC-MS.
No. Compound RI* RI**
% relative peak area
L. x hybrid
‘boysemberry
ruffles’
L. x hybrid
‘high five
purple’
L. pedunculatas
‘princess’
1 tricyclene 918 921 0.11±0.18 0.31±0.27
2 α-pinene 928 932 2.31±0.07 0.02±0.00 4.75±0.18
3 α-fenchene 940 945 1.60±0.07
4 camphene 941 946 1.97±0.08 4.78±0.14
5 sabinene 962 969 0.37±0.13 0.10±0.0.5 0.13±0.07
6 β-pinene 970 974 0.95±0.08 0.47±0.11 0.37±0.11
7 3-octanone 976 979 0.02±0.00
8 myrcene 984 988 0.33±0.13 0.34±0.10 0.32±0.04
9 3-octanol 985 988 0.06±0.10 0.11±0.03
10 δ-2-carene 998 1001 1.45±0.11
11 α-phellandrene 999 1002 0.11±0.05
12 δ-3-carene 1002 1008 1.87±0.07
13 α-terpinene 1010 1014 0.02±0.00 0.13±0.04
14 p-cymene 1015 1020 0.33±0.13 0.09±0.03
15 o-cymene 1018 1022 0.47±0.10
16 limonene 1020 1024 9.73±0.11
17 1,8-cineole 1022 1026 28.81±0.14 12.29±0.15 9.73±0.10
18 Z-β-ocimene 1029 1032 1.05±0.10 0.70±0.07
19 β-ocimene 1041 1044 0.23±0.13 0.02±0.00
20 pulegone 1048 1053 0.02±0.00
21 γ-terpinene 1050 1054 0.08±0.04 0.19±0.02
22 cis-linalool oxide 1060 1067 0.18±0.02
23 cis-vertocitral C 1070 1076 0.07±0.02
24 camphenilone 1072 1078 0.09±0.04 0.13±0.05
25 fenchone 1079 1083 5.50±0.13 21.91±0.14 23.73±0.13
26 isoterpinolene 1080 1085 2.09±0.13
29 trans-vertocitral C 1096 1105 0.02±0.00
30 camphor 1138 1141 33.62±0.17 46.65±2.13 30.30±1.24
31 isoborneol 1148 1155 0.56±0.17
32 δ-terpineol 1156 1162 1.52±0.11 0.58±0.14
33 lavandulol 1161 1164 0.50±0.04
34 terpinen-4-ol 1170 1174 0.87±0.08
35 santalone 1173 1177 0.71±0.13
36 p-methyl acetophenone 1173 1179 0.08±0.10
37 α-terpineol 1180 1186 1.21±0.13 0.74±0.10 0.47±0.13
38 trans-isocarveol 1183 1187 0.61±0.14
39 myrtenal 1190 1195 0.17±0.10
40 verbenone 1199 1204 0.20±0.17 0.12±0.16 0.18±0.14
41 endo-fenchyl acetate 1214 1218 0.20±0.11
42 carvone 1234 1239 0.09±0.02 0.18±0.08
43 iso-isopulegyl acetate 1275 1283 0.66±0.13 2.84±0.13
44 bornyl acetate 1283 1287 5.28±0.28
45 lavandulyl acetate 1284 1288 0.02±0.00
46 geranyl formate 1295 1298 1.06±0.14
47 cis-pinocarvyl acetate 1305 1311 0.22±0.13 1.79±0.12
48 neo-verbanol acetate 1315 1319 0.13±0.10
49 E-cis-jasmonol 1324 1328 1.07±0.14

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 71
No. Compound RI* RI**
% relative peak area
L. x hybrid
‘boysemberry
ruffles’
L. x hybrid
‘high five
purple’
L. pedunculatas
‘princess’
50 trans-carvyl acetate 1335 1339 3.16±0.28
51 α-ylangene 1370 1373 0.19±0.14
52 α-duprezianene 1383 1387 0.85±0.13
53 β-cubebene 1383 1387 0.02±0.00
54 iso-longifolene 1384 1389 0.13±0.07 0.05±0.02
55 sesquithujene 1400 1405 0.05±0.02
56 acora-3,7(14)-diene 1401 1407 0.20±0.13
57 α-barbatene 1403 1407 0.02±0.00
58 α-gurjunene 1406 1409 0.22±0.11 0.02±0.00
59 α-cedrene 1407 1410 0.15±0.05
60 aromadendrene 1433 1439 0.08±0.02
61 Z-β-farnesene 1435 1440 0.02±0.00
62 γ-muurolene 1472 1478 0.15±0.04 0.02±0.00
63 amorpha-4,7(11)-diene 1474 1479 0.05±0.02 0.10±0.07
64 β-vetispirene 1491 1493 0.18±0.04
65 valencene 1494 1496 0.06±0.02
66 epizonarene 1496 1501 0.05±0.02
67 α-chamigrene 1498 1503 0.07±0.02
68 premnaspirodiene 1499 1505 1.36±0.13
69 Z-α-bisabolene 1499 1506 0.11±0.04
70 δ-amorphene 1507 1511 0.09±0.02
71 γ-cadinene 1508 1513 0.32±0.04
72 sesquicineole 1509 1518 0.02±0.00
73 7-epi-α-selinene 1517 1521 0.05±0.02
74 10-epi-cubebol 1525 1533 0.11±0.04
75 E-β-ionol acetate 1531 1537 0.07±0.02
76 α-calacorene 1538 1542 0.19±0.13
77 trans-dauca-4(11),7-diene 1547 1556 0.22±0.11
78 E-nerolidol 1552 1562 0.10±0.03
79 longipinanol 1560 1567 0.40±0.07
80 zierone 1565 1574 0.37±0.13
81 santalenone 1567 1576 0.02±0.00 0.05±0.02
82 β-copaen-4α-ol 1581 1590 0.93±0.07 0.51±0.07 0.02±0.00
83 khusimone 1591 1604 0.18±0.14
84 β-humulene epoxide 1595 1614 0.06±0.02
85 epi-cedrol 1605 1618 0.37±0.15
86 β-acorenol 1619 1637 0.12±0.14
87 α-muurolol 1629 1644 0.65±0.08
88 himachalol 1648 1652 0.17±0.08
89 α-bisabolol oxide B 1649 1658 0.02±0.00
%yield (w/w) 0.24±0.13 0.11±0.32 0.22±0.18
Total of volatile components 45 38 42
RI*; calculated retention indices, RI**; retention indices from literature Adams (2017).
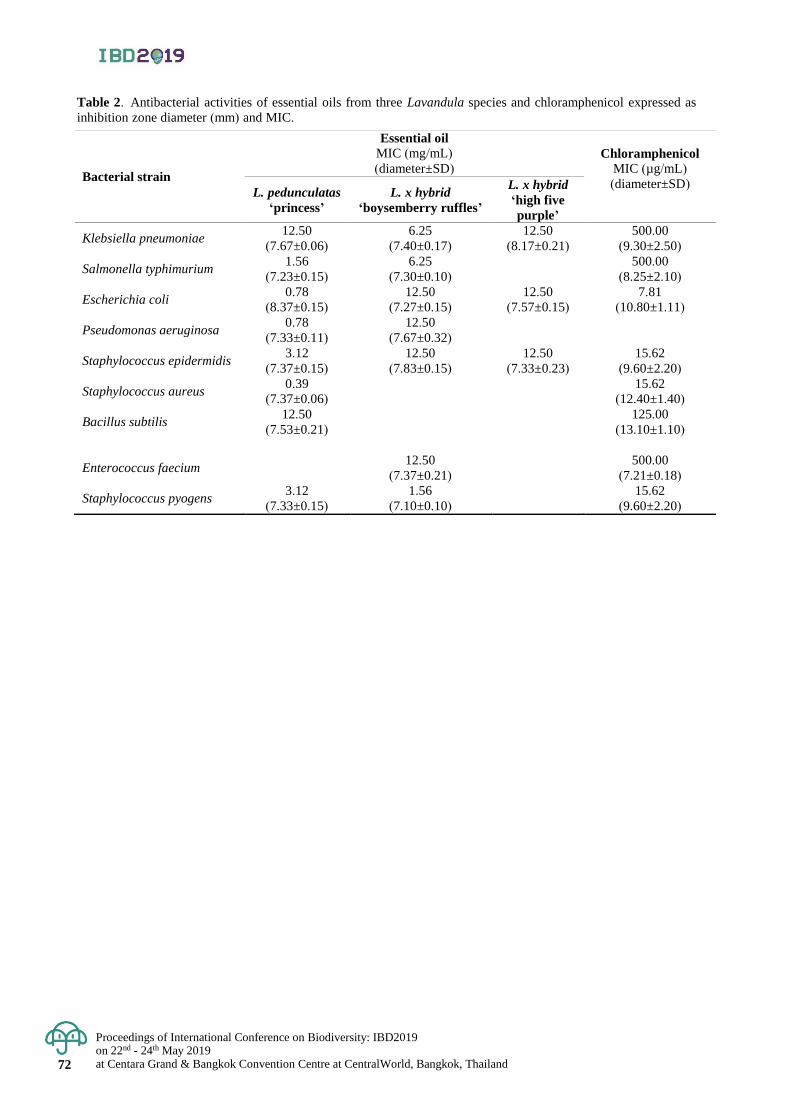
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 72
Table 2. Antibacterial activities of essential oils from three Lavandula species and chloramphenicol expressed as
inhibition zone diameter (mm) and MIC.
Bacterial strain
Essential oil MIC (mg/mL)
(diameter±SD)
Chloramphenicol MIC (µg/mL)
(diameter±SD) L. pedunculatas
‘princess’
L. x hybrid
‘boysemberry ruffles’
L. x hybrid
‘high five
purple’
Klebsiella pneumoniae 12.50
(7.67±0.06)
6.25
(7.40±0.17)
12.50
(8.17±0.21)
500.00
(9.30±2.50)
Salmonella typhimurium 1.56
(7.23±0.15)
6.25
(7.30±0.10)
500.00
(8.25±2.10)
Escherichia coli 0.78
(8.37±0.15)
12.50
(7.27±0.15)
12.50
(7.57±0.15)
7.81
(10.80±1.11)
Pseudomonas aeruginosa 0.78
(7.33±0.11)
12.50
(7.67±0.32)
Staphylococcus epidermidis 3.12
(7.37±0.15)
12.50
(7.83±0.15)
12.50
(7.33±0.23)
15.62
(9.60±2.20)
Staphylococcus aureus 0.39
(7.37±0.06)
15.62
(12.40±1.40)
Bacillus subtilis 12.50
(7.53±0.21)
125.00
(13.10±1.10)
Enterococcus faecium
12.50
(7.37±0.21)
500.00
(7.21±0.18)
Staphylococcus pyogens 3.12
(7.33±0.15)
1.56
(7.10±0.10)
15.62
(9.60±2.20)

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 73
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 73 - 76
Discovery of plant antimicrobial peptides and laboratory scale production
Parichart Burns1,2,3*, Jutatape Watcharachaiyakup2,3, Patchima Sithisarn4, Pimpilai Saengmanee2,
Vinitchan Ruanjaichon1 and Sonthichai Chanpreme5
1National Center for Genetic Engineering and Biotechnology (BIOTEC), National Science and Technology
Development Agency, Khlong Luang District, Pathum Thani, Thailand
2Center for Agricultural Biotechnology, Kasetsart University, Kamphaeng Saen District, Nakhorn Pathom, Thailand 3Center of Excellence on Agricultural Biotechnology, Chatuchak Distance, Bangkok, Thailand
4Department of Veterinary Public health, Faculty of Veterinary Medicine, Kasetsart University Kamphaeng Saen District,,
Nakhorn Pathom, Thailand 5Department of Agronomy, Faculty of Agriculture at Kamphaeng Saen, Kasetsart University, Kamphaeng Saen District,
Nakhorn Pathom, Thailand *Corresponding author e-mail: [email protected]
Abstract: Plant antimicrobial peptides (PAMPs) are natural peptide molecules derived from plants with antimicrobial
properties. Their targets range from viruses, bacteria to fungi with the activities of replication or growth inhibition,
slow and abnormality growth induction, bactericides and fungicides. With the availability of genome, transcriptome
and metabolome databases, bioinformatics has been successfully identified these beneficial molecules and in silico
studied of their properties. Four peptides including P5, P9, P25 and P36 were selected from retrieved genomic
and transcriptome data of monocots including maize, sorghum, rice and sugarcane. Their properties were analysed
online tools such as the predicted protein server, Phyre2 and AMPA. The molecules had potential to be antimicrobial
peptides. In order to determine their activities, E. coli expression system was used to produce these peptides in
fusion form. The fusion proteins were selected, digested, purified and concentrated using affinity absorption and
size selection methods. Spectrophotometry indicated that the purified peptides had high concentration and can be
used for further study.
Keywords: defensin, bioinformatics, E. coli expression system, laboratory scale production, protein stability
Introduction
Antimicrobial peptides (AMPs) are found in invertebrates, vertebrates, plant and bacteria. Most AMPs
are cationic or amphiphilic. Hence their interactions are to anionic charge molecules. Modes of AMP functions
include 1) binding to negatively charged surfaces of gram negative bacterial outer membrane, 2) interact with
bacterial cell wall and 3) interact with bacterial cell membrane. These actions cause the restriction cell division,
causing cell wall remodeling and subsequently cell lysis. In some cases, AMPs interact with specific targets within
the bacterial cells causing the disruption of cellular functions such as protein synthesis (Bechinger and Gorr, 2018).
They are, therefore, valuable not only for plant pathogen protection but also medical and veterinary treatment.
Some plant AMPs reportedly had antifungal and antibacterial activities against both plant and human pathogens
(Sathoff et al., 2019). Although share modes of functions, plant microbial peptides have a distinct character of high
cysteine residues which form multiple disulfide bonds. Most of them are low and moderate in size (molecular weight).
These features provides advantages of chemical, thermal and proteolytic activities. There are seven major PAMP
families; thionins, defensin, hevein, knottin, hairpinin, lipid transfer proteins (LTPs) and snakins (Tam et al., 2015).
Recently, many plant genome and transcriptome databases are available in public domain. Therefore,
bioinformatics approach becomes a power tool in identifying and retrieving PAMPs data. Schmitt et al., (2016)
reported the link between variations of AMP amino acid sequences and AMP binding capacity to bacterial membranes.
Plant AMPs are generally accumulated from specific organs such as leaves, flower and seeds. Several purification
steps are required for purified AMPs (Tang et al., 2018). Alternatively, in silico approach combining with de novo
synthesis via chemical reaction has become an alternative approach to conventional method for production and
an improvement of AMP (Mikut et al., 2016). The aim of this study is to 1) identify target AMPs and retrieve
the genome/transcriptome data from monocots databases using in silico study 2) to amplify AMP genes from monocots
and 3) produce purified PAMPs in laboratory scale.
Materials and Methods
Plant databases
There are many plant genome and transcriptome databases available on line. However, only those with
monocots data (sugarcane, rice, maize and sorghum) were included (Table 1). Criteria such as keywords and
sequence similarity were used in BLAST and FASTA search. The outcome from various sources were compared
and analyzed. The PAMP genomic DNA and cDNA sequences were retrieved. They were aligned using MacVector 12.6.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 74
In silico study on protein properties and antimicrobial activity
Protein secondary and tertiary structures and other important characters including solubility, transmembrane
probability and stability were investigated. The antimicrobial activities were determined using on line software (Table 1).
Primer design and PCR amplification
Primers were designed based on retrieved sequences using Primer3 (Untergasser et al., 2012). The target
genes were amplified using Reverse transcription PCR. The DNA were cloned and automate sequenced. The authenticity
were determined using BLASTX software (Altschul et al., 1997).
E. coli Expression system
DNA fragments were cloned into pMAL-c2 (New Englan Biolab, USA) and recombinant plasmids were
transformed into E. coli. The optimal temperature were investigated in small scale conditions. The fusion protein
was produced in laboratory scale under optimized conditions. It was purified and digested with factor Xa. PAMPs
was further purified and concentrated.
Result & Discussion
Four plant microbial peptides (P5, P9, P25 and P36) were successfully retrieved from plant databases.
They were identified as defensin and snakin. They displayed antimicrobial activities and relatively stable in
cytoplasm. The coding regions were cloned into pMALc2 E. coli expression vector. The work processes were
summarized in Figure. 1 E. coli expression system is widely used for heterologous protein expression because of
the cost effective and routine/standard protocol for E. coli production (Chen, 2012).The temperature, aeration and
duration for induction were tested to fine optimized condition for protein production (Papaneophytou and Kontopidis,
2014; Singha et al., 2017). The fusion proteins were produced in laboratory scale and maintained within bacterial
cells. They were released using sonication and purified from other bacterial proteins via specific binding. PAMPs
were digested out of fusion proteins, purified and concentrated using size exclusion chromatography. The concentration
and protein content were summarized in Table 2. The results indicated variation of protein recovery and yield.
Interestingly, the deviation results from the last steps of factor Xa digestion and protein recovery. This could be
specific protein tertiary structure that reduce availability of factor Xa site or the current process caused misfolding
of specific protein (Overton, 2013).
Conclusion
Plant antimicrobial peptides (PAMPs) were effectively produced in laboratory scale using bioinformatics
approaches combining with E. coli expression system. This method can be applied to other AMPs production.
Acknowledgement
This research is financially supported by Thailand Research Fund (TRF) through National Research
Council of Thailand (NRCT). National Center for Genetic Engineering and Biotechnology (BIOTE) and Center
for Agricultural Biotechnology (CAB) provided laboratory facilities and equipment.
References
Bechinger, B., and Gorr, S. U. 2017. Antimicrobial peptides: mechanisms of action and resistance. Journal of Dental
research 96(3): 254260.
Chen, R. 2012. Bacterial expression system for recombinant protein production: E. coli and beyond. Biotechnology
Advances 30: 11021107.
Mikut, R., Ruden, S., Reischl, M., Breitling, F., Volkmer, R., and Hilper, K. 2016. Improving short antimicrobial peptides
despite elusive rules for activity. Biochimica et Biophysica Acta (BBA)-Biomembranes 1858:10241033.
Overton, T. W. 2014. Recombinant protein production in bacterial hosts. Drug Discovery Today 19(5): 590601.
Papaneophytou, C.P. and Kontopidis, G. 2014. Statistical approaches to maximize recombinant protein expression
in Escherichia coli: A general view. Protein Expression and Purification 94: 2232.
Sathoff, A. E., Velivelli, S., Shah, D. M. and Samac, D. A. 2019. Plant defensing peptides have antifungal and
antibacterial activity against human and plant pathogens. Phytopathology 109: 402408.
Schmitt, P., Rosa, R. D. and Destoumieux-Garzón, D. 2016. An intimate link between antimicrobial peptide
sequence diversity and binding to essential components of bacterial membranes. Biochimica et Biophysica
Acta (BBA)-Biomembranes 1858: 958–970.
Singha, T. K., Gulati, P., Mohanty, A., Khasa, Y., Kapoor, R. K. and Kumar, S. 2017. Efficient genetic approaches
for improvement of plasmid based expression of recombinant protein in Escherichia coli: A review.
Process Biochemistry 55: 1731.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 75
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., and Lipman D. J. 1997. Gapped BLAST
and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research. 25(17)
: 33893402.
Tam, J. P., Wang, S., Wong, K. H. and Tan, W. L. 2015. Antimicrobial peptides from plants. Pharmaceuticals 8(4): 711757.
Tang, S. S., Prodhan, Z H., Biswas, S. K., Le, C. F. and Sekaran, S. D. 2018. Antimicrobial peptides from different
plant sources: isolation, characterization and purification. Phytochemistry 154: 94105.
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B.C., Remm, M. and Rozen, S.G. 2012. Primer3-
new capabilities and interfaces. Nucleic Acids Research 40(15): e115.
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 76
Figure 1. The overview of plant AMP protein production at laboratory scale.
Table 1. Websites used in this study for A) Plant genome and transcriptome databases and B) Protein analysis
and antimicrobial activity.
Name Location Tools
Ensembl Plants
Home http://plants.ensembl.org/species.html BLAST/BLAT
NCBI https://www.ncbi.nlm.nih.gov/ BLAST, keyword search
Phytozome12 https://phytozome.jgi.doe.gov/pz/portal.html BLAST, keyword search
PlantGDB http://www.plantgdb.org/prj/GenomeBrowser/ BLAST, pattern search
Sugarcane genome
hub http://sugarcane-genome.cirad.fr/
BLAST, keyword search, primer
design
PredyFlexy http://www.dsimb.inserm.fr/dsimb_tools/predyfl
exy/index.html
Flexibility and Local Structure
prediction from sequence
Pro Pi https://www.protpi.ch Protein tools, peptide tools
AntiBP2 http://crdd.osdd.net/raghava/antibp2/index.html Server for antibacterial peptide
prediction
Antimicrobial
Sequence Scanning
System (AMPA)
http://tcoffee.crg.cat/apps/ampa/do
A theoretical approach to spot
active regions in antimicrobial
proteins.
Table 2. Protein concentration and total protein content of four plant antimicrobial peptides. Bradford assay was
used for determination of protein concentration with bovine serum albumin (BSA) as a protein standard.
Step Protein concentration
(mg/mL)
Total protein (milligram)
Cell suspension 0.455 - 1.083 182-433.2
Cell disruption 7.65 -8.27 153.0-169.5
Fusion protein 4.15-7.19 4.15-7.19
Purified protein 0.014-0.765 0.0032-137.7

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 77
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 77 - 85
Diversity of indigenous vegetables in Puparn Royal Development
Study Centre, Sakon Nakhon Province, Thailand
Hathairat Chokthaweepanich1*, Kwankamol Tawaitakam1and Noppadol Kaewkumsai2 1Department of Agriculture and Resources, Faculty of Natural Resources and Agro-Industry,
Kasetsart University Chalermphrakiat Sakonnakhon province Campus, Muang District, Sakon Nakhon, Thailand 2The Puparn Royal Development Study Centre, Muang District, Sakon Nakhon, Thailand
*Corresponding author e-mail: [email protected]
Abstract: A study on diversity of indigenous vegetables in Puparn Royal Development Study Centre, Sakon
Nakhon province during June 2015 to May 2016 was purposed to collect and classify the indigenous vegetables
with revealing their potential as food resources, edible parts and utilization method. Sixty-three species, sixty
genera and forty-three families were identified. The four majorities of plant families were Zingiberaceae,
Fabaceae, Araceae, and Asteraceae, respectively. Additionally, identified indigenous vegetables could be
classified into six groups based on consumed part including underground stem (4 species), leaves and young shoots
(35 species), stem (2 species), flower (4 species), fruit (6 species) and whole plant (21 species). Most of utilization
methods were used as salad, steamed or boiled vegetables eaten with chili paste, minced meat, or seasoning in
food ingredients. Keywords: indigenous vegetables, diversity, plant utilization, Sakon Nakhon province
Introduction
The Puparn Royal Development Study Centre is established in Ban Na Nok Khao, Huai Yang sub district,
Muang district, Sakon Nakhon province on November 25, 1982. This centre is one of six development study
centres that is settled in northeastern region of Thailand. A total land area is approximately 2,128 hectares (13,300
rais) comprising about 368 hectares (2,300 rais) of an agricultural development zone and 1,760 hectares (11,000
rais) of peripheral areas earmarked for forestry development that is located within Pa Phu Lom Khao – Phu Peck
National Reserved Forests and Mai Kraya Loei Namphung Forest Project. Most of ecological systems in this
location are natural forest consisted of Dipterocarp forest (60%), dry evergreen forest (13%), mixed deciduous
forest (12%) and agricultural area (12%). Additionally, people who live in this area and nearby can get benefits
from plants in this forest [Office of the Royal Development Projects Board (ORDPB), 2018].
There are many species of indigenous vegetables in Thailand especially northeastern region. Deewiset
and Khumklang (1998) collected 139 species of indigenous vegetables in northeastern region. Khumklang et al.
(2000) reported that there were more than 220 species of indigenous vegetables in Thailand and majority of them
were found in the northeastern region. Indigenous vegetables can be consumed either wholly or in parts of plant.
The edible parts of vegetables may be root, stem, leave, flower, fruit, rhizome or bulb. Moreover, some mushroom
can also be consumed as vegetable (Shirai and Rambo, 2008; Nnamani et al., 2010; Chokthaweepanich et al.,
2016). These local vegetables are not only an excellent source of vitamins and minerals, but they can also be used
for several purposes such as food, medicinal plants, ornamental plants and household incomes (Omara-Achong et
al., 2012; Abukutsa-Onyango, 2014; Srisawat et al., 2016; Sseremba et al., 2017; Phumthum et al., 2018).
Indigenous vegetables can be prepared from locally available resources for food and nutritional security.
However, the local vegetables are known and take benefit in different patterns depend on traditional knowledge,
tasting culture, location, seasonal availability and social group (Lee et al., 2008; Omara-Achong et al., 2012;
Abukutsa-Onyango, 2014; Khuankaew et al., 2014). Although economic vegetables are widely eaten in Thailand
at the present, indigenous vegetables are still important to people who live in the rural area (Agoreyo et al., 2012;
Ayaz et al., 2015; Sseremba et al., 2017) because indigenous vegetables are cheap and substantial source of amino
acid, protein, vitamins and minerals (Okafor, 1983). Moreover, many researchers reported beneficial effects from
local vegetable consumption such as high antioxidant (Hong et al., 2004; Tangkanakul et al., 2006; Panpipat et
al., 2010; Chao et al., 2014; Baang et al., 2015; Kongkachuichai et al., 2015; Tinrat, 2016), flavonoid contents
(Baang et al., 2015; Kankanamge and Amarathunga, 2017), phenolic compound (Panpipat et al., 2010; Tinrat,
2016; Kankanamge and Amarathunga, 2017; Yakoh et al., 2018), preventing hypertension (Chuchote et al., 2015),
etc.
Unfortunately, traditional thai vegetables are risk to loss since the younger generations who prefer the
introduced species (e.g. broccoli, cabbage, cauliflower, lettuce, tomato, etc.), don’t know cooking techniques or
recipe using indigenous vegetables (Somnasang and Moreno-Black, 2000; Kolsuwat, 2002; Setalaphruk and Price,
2007). Homemade meals are substituting with fast food in Thailand (Kosulwat, 2002). Economic or introduced

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 78
vegetables are key ingredients in many dishes. Although many indigenous vegetables are sale in the local market,
some people may know their plant name, kinds, cooking process (Shirai and Rambo, 2008). Therefore, it will be
a worthwhile attempt to study on diversity and classification of indigenous vegetables covering their used parts
and utilization method in the Puparn Royal Development Study Centre.
Materials and Methods
Study Area
This study was conducted in the Puparn Royal Development Study Centre located in Sakon Nakhon
province, Thailand during June 2015 to May 2016. It is located between latitudes 170 04’ to 170 07’ north and
longitudes 1040 00’ to 1040 04’ East (Figure 1). The range of height is between 180-420 meters above moderate
sea level. The average temperature in this area is approximately 26.40C. The lowest temperature is around 12.10C
and highest is around 37.90C. The average precipitation is 362.1 mm per year and average annual relative humidity
is 72.9%. Soil condition in this study area is sandstone of conglomerate type and shale with lower layers of soil
comprised shale, sandstone and limestone (ORDPB, 2018).
Data Collection
Indigenous vegetables were collected and photographed from three forest areas including Dipterocarp
forest, dry evergreen forest and mixed deciduous forest. All samples were recorded plant ecology and
morphological characteristics especially disappeared traits in herbarium specimens or preserved plants in alcohol-
glycerine such as habit, latex, color, taste and flavor. Moreover, the local name, used part, and utilization methods
also were gathered from local people. The collected plants were identified in the fields and laboratory by
taxonomists or comparing with voucher specimens at Bangkok Herbarium Museum, Department of Agriculture,
Bangkok. Finally, all specimens were reconfirmed using literature on the flora of Thailand, Flora of China and
Thai plant names (Revised edition 2014) book (Smitinand, 2014).
Result & Discussion
Sixty-three indigenous vegetable species belong to 60 genera and 43 families (Table 1) were collected in
the Puparn Royal Development Study Centre. They could be classified into 4 main groups including
monocotyledonous plants 15 species, dicotyledonous plants 41 species, fern 2 species and fungi 5 species. The
most discovered plant families were 5 species (7.94%) in Zingiberaceae, 4 species (6.35%) in Fabaceae, 3 species
(4.76%) each in Araceae and Asteraceae. Moreover, They could be classified into 9 groups (Figure 2) depend on
plant habit (Smitinand, 2014) consisted of tree 9 species (14.29%), shrub 9 species (14.29%), herb 25 species
(39.68%), climbing 9 species (14.29%), scandent 1 species (1.59%), palm 1 species (1.59%), bamboo 2 species
(3.17%), fern 2 species (3.17%) and fungi 5 species (7.94%). All fungi were 5 species of mushrooms that grew in
the rainy season. Herbs were the dominant indigenous vegetables in this study because they contained less woody
tissue in all vegetative parts than tree and shrub, whereas tree and shrub took longer time for new branch, shoot
and fruit (Thongpukdee et al., 2014). Besides, the main features of surveyed areas were deciduous forest, open
canopy and abundance of herbs in the ground layer (Rundel and Boonpragob, 1995).
The majority species of indigenous plants were found in Dipterocarp forest (34 species), mixed deciduous
forest (30 species), wet areas (19 species) and dry evergreen forest (17 species), respectively. The Dipterocarp
forest contained more indigenous species than others because this forest type covered 60% of all areas (ORDPB,
2018), while the mixed deciduous forest was indicated higher plant diversity when species-area ratio was
examined (Figure 3). Dipterocarp forest was considered to be less diversity than evergreen forests or wet forests
due to the conditions of low moisture, high temperature and low annual rainfall averages (Murphy and Lugo, 1986;
Powers et al., 2018). Besides, most of trees in Dipterocarp forest are shedding their leaves in the dry season
(Janzsn, 1998; Himmapan and Kaitpraneet, 2008; Elliiott et al., 2013).
The plant parts used from local people both in single or multiple preparations (Figure 4 and Table 1) were
underground stem 4 species (6.35%), leave and young shoot 35 species (55.56%), stem 2 species (3.17%), flower
4 species (6.35%), fruit 6 species (9.52%) and whole plant 21 species (33.33%). The results revealed that the
greater vegetable consumption was leave and young shoot because this part was young and appreciated for
cooking. Additionally, leafy vegetables were quick to prepare and an admired source of food and medicine
(Turreira-García et al., 2017). The investigation of this study reported that most utilization methods were used in
cooked or fresh vegetables as salad, steamed or boiled vegetables eaten with chili paste, spicy minced meat, or
seasoning in food ingredient. Furthermore, they could be cooked in a variety of dishes; for example, soup, stir-
fried, curry or as a sweet dessert. Some lifestyle diet was similar to other regions in Thailand (Ngamsiri and
Thananoppakun, 2014; Turreira-García et al., 2017) and other countries such as in Kenya (Gido et al., 2017),
Nepal (Uperty et al., 2012), Nigeria (Nnamani et al., 2010; Arowosegbe et al., 2018).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 79
Various indigenous species have been presented to be used several ways as well as vegetables, for
instance, fruit (e.g. Aegle marmelos, Antidesma ghaesembilla, Passiflora foetida, Garcinia cowa, Schleichera
oleosa); medicinal plants (e.g. Acmella oleracea, Alpinia sp., Limnophila geoffrayi, Phlogacanthus pulcherrimus,
Glinus oppositifolius); ornamental plants (e.g. Ardisia crenata, Curcuma sessilis, Curcuma singularis, Cratoxylum
formosum, Caryota mitis, Nymphaea pubescens); and sweet dessert (e.g. Cissampelos pareira, Dioscorea alata,
Dioscorea sp.). Some local vegetables should not be eaten raw because of highly toxic prussic acid and must be
cooked or fermented before consuming to neutralize the hydrocyanic acid (CSIR, 1950; Coursey, 1967) such as
wild yam (Dioscorea sp.), giant taro (Colocasia gigantea) and unicorn plant (Lasia spinosa). Some indigenous
vegetables in this study were reported as a good source of minerals and vitamins (Office of the Primary Health
Commission. 1997; Nutrition Division, 2001; Sirival, 2008; Kingkachuichai et al., 2015): phosphorus (e.g.
Dracaena angustifolia, Lobelia chinensis, Persicaria odorata); potassium (e.g. Blumea napifolia, Dracaena
angustifolia, Lobelia chinensis,); calcium (e.g. Blumea napifolia, Cissampelos pareira, Lobelia chinensis); iron
(e.g. Diplazium esculentum, Hydrocotyle umbellata); vitamin A (e.g. Careya sphaerica, Centella asiatica,
Cissampelos pareira, Hydrocotyle umbellata, Lasia spinosa); and vitamin C (e.g. Aegle marmelos, Careya
sphaerica, Cissus hastate, Melientha suavis, Passiflora foetida). Consequently, the local vegetables can help the
rural people to prevent malnutrition (Omara-Achong et al., 2012; Kongkachuichai et al., 2015; Phumthum et al.,
2018) and to get rid of common illness (Chotchoungchatchai et al., 2012; Khuankaew et al., 2014; Phumthum et
al., 2018).
Conclusion
The Puparn Royal Development Study Centre can be represented as a major source of plant diversity.
Forty-three families with sixty-three species of indigenous vegetables were encountered in different four
ecosystems. A number of vegetables were found in the Dipterocarp forest. Leave and young shoot of diverse
species were preferred to food preparation. There were alternative methods to make use of indigenous plants
except vegetables such as medicine, ornamental plant or dessert. These vegetables are a good option for contributes
to food and nutrition security.
Acknowledgement
We are grateful to Office of the Royal Development Projects Board (ORDPB) for providing research
funding. We thank the Puparn Royal Development Study Centre for transportation and assistance during
fieldwork. Finally, we are also thankful to Dr. Yingyong Paisooksantivattana and Dr. Sranya Vajrodaya (Kasetsart
University) for some plant identification
References
Abukitsu-Onyango, M. 2014. Strategic repositioning African indigenous vegetables and fruits with nutrition,
economic and climate change resilience potential. In A. Gurib-Fakim (Eds), Novel Plant Bioresources
(pp. 361-369). John Wiley and Sons, London.
Agoreyo, B. O., Obansa, E. S., Obanor, E. O. 2012. Comparative nutritional and phytochemical analyses of
two varieties of Solanum melongena. Sci. World J. 7(1): 5-8.
Arowosegbe, S., Olanipekun, M. K., and Adeloye, I. A. 2018. Ethnobotanical survey of indigenous leafy
vegetables consumed in Ekiti Sate, Nigeria. European Journal of Biology and Medicinal Science
Research 6(1): 7-14.
Ayaz, F. A., Colak, N., Topuz, M., Tarkowski, P., Jaworek, P., Seiler, G., and Inceer, H. 2015. Comparison of
nutrient content in fruit of commercial cultivars of eggplant (Solanum melongena L.). Pol. J. Food
Nutr. Sci. 65(4): 251-260.
Baang, R. P., Rosario, R. M. D., and Palmes, N. D. 2015. Phytochemical profiles and antioxidant activity of
selected indigenous vegetables in Northern Mindanao, Philippines. International Journal of
Bioengineering and Life Sciences 9(8): 870-875.
Chao, P.-Y., Lin, S.-Y., Lin, K.-H., Liu, Y.-F., Hsu, J.-I., Yang, C.-M., and Lai, J.-Y. 2014. Antioxidant activity
in extracts of 27 indigenous Taiwanese vegetables. Nutrients 6: 2115-2130.
Chokthaweepanich, H., Phuburom, K., and Wongsridakulkaew, Y. 2016. Diversity of indigenous vegetables in
Ta-Rae market Sakon Nakhon province. Songklanakarin Journal of Plant Science 3(Suppl. III): M10/7-
12.
Chotchoungchatchai, S., Saralamp, P., Jenjittikul, T., Pornsiriponge, S., and Prathanturarug, S. 2012. Medicinal
plants used with Thai traditional medicine in modern healthcare services: A case study in Kabchoeng
Hospital, Surin Province, Thailand. Journal of Ethnopharmacology 141: 193-205.
Chuchote, C., Pathompak., P., and Charoonratana, T. 2015. Screening of angiotensin I-converting enzyme
inhibition of Thai indigenous vegetables. Bulletin of Health, Science and Technology 13(1): 38-42.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 80
Coursey, D. G. 1967. Yams, Longman, London. 230pp. CSIR. 1950. Wealth of India. Vol. II, CSIR, New Delhi.
Deewiset, K. and Khumklang, A. 1998. Local vegetables in the northeast. Veterans Organization Publishing
House, Bangkok.
Elliott, S., Blakesley, D., and Hardwick, K. 2013. Restoring tropical forests: A practical guide. Kew: Royal
Botanical Gardens.
Gido, E. O., Ayuya, O. I., Owuor, G., and Bokelmann, W. 2017. Consumption intensity of leafy African
indigenous vegetables: towards enhancing nutritional security in rural and urban dwellers in Kenya.
Agricultural and Food Economics 5:14.
Himmapan, W., and Kaitpraneet. S. 2008. Effect of fire on vegetation in the dry deciduous dipterocarp forest
at Huai Kha Khaeng wildlife sanctuary, Uthai Thani province. Thai Journal of Forestry 27(1): 43-55.
Hong, J. H., Manochai, B., Trakoontivakorn, G., and Na Thalang, V. 2004. Fibrinolytic activity of Thai indigenous
vegetables. Kasetsart J. (Nat. Sci.) 38: 241-246.
Janzen, D. H. 1998. Tropical dry forests: The most endangered major tropical ecosystem. In E. O. Wilson
(Ed.), Biodiversity (pp. 130-137). National Academy of Sciences, U.S.A.
Kankanamge, S. U. and Amarathunga, A. A. M. D. A. N. 2017. Phytochemical and ethno-pharmacological
properties of Lasia spinosa (Kohila): A review. World Journal of Pharmaceutical Research 6(13):1-9.
Khuankaew, S., Srithi, K., Tiansawat, P., Jampeetong, A., Inta, A. and Wangpakapattanawong, P. 2014.
Ethnobotanical study of medicinal plants used by Tai Yai in Northern Thailand. Journal of
Ethnopharnacology 151: 829-838.
Khumklang, A., Sakulku, S., and Phranakorn, R. 2000. “Eat line” Isan Food. Sakon Nakhon Agricultural
Research and Training Institute, Rajamangala Institute of Technology Ministry of Education.
Kongkachuichai, R., Charoensiri, R., Yakon, K., Kringkasemsee, A., and Insung, P. 2015. Nutrients value and
antioxidant content of indigenous vegetables from Southern Thailand. Food Chemistry 173: 838- 846.
Kosulwat, V. 2002. The nutrition and health transition in Thailand. Public Health Nutrition 5(1A): 183-189.
Lee, S., Xiao, C., Pei, S. 2008. Ethnobotanical survey of medicinal plants at periodic markets of
Honghe Prefecture in Yunnan Province, SW China. Journal of Ethnopharmacology 117: 362-377.
Murphy, P. G. and Lugo, A. E. 1986. Ecology of tropical dry forest. Annual Review of Ecology and Systematics
17: 67-88.
Nnamani, C. V., Oselebe, H. O., and Okporie, E. O. 2010. Aspect of ethnobotany of traditional leafy vegetables
utilized as human food in rural tropical communities. Animal Research International 7(1): 1110-1115.
Ngamsiri, Y. and Thananoppakun, K. 2014. Species diversity and utilization of local vegetable in Khirimat
District, Sukhothai province, Proceeding of the 12th Agricultural Conference of Naresuan University,
October 27th- November 2nd, 2014, Sukhothai, Thailand. p. 158-163.
Nutrition Division, Department of health, Ministry of Public Health. 2001. Nutritive values of Thai foods.
Veterans Organization Publishing House, Bangkok. Office of the Primary Health Commission. 1997.
Local vegetables: the meaning and wisdom of the Thai people. Institute of Thai Traditional
Medicine, Department of Medical Services, Ministry of Public Health, Bangkok, Thailand.
Office of the Royal Development Projects Board (ORDPB). 2018. History of the Royal Development
Study Centres. Amarin Printing and Publishing Plc., Bangkok, Thailand.
Okafor, J. C. 1983. Horticulturally promising indigenous wild plant species of the Nigerian forest zone. Acta
Horticulturae 123: 165-176.
Omara-Achong, T. E., Edwin-Wosu, N. L., Edu, E. A., and Nkang, A.E. 2012. Survey of indigenous vegetables
species in parts of Ogaja and Cala Bar, Cross River State, Nigeria. European Journal of Experimental
Biology 2(4): 1289-1301.
Pantipat, W., Suttirak, W., and Chaijan, M. 2010. Free radical scavenging activity and reducing capacity of five
Southern Thai indigenous vegetables extracts. Walailak J. Sci. & Tech. 7(1): 51-60.
Phumthum, M., Srithi, K., Inta, A., Junsongduang, A., Tangjitman, K., Pongamornkul, W., Trisonthi, C., and
Balslev, H. 2018. Ethnomedicinal plant diversity in Thailand. Journal of Ethnopharmacology 214: 90-98.
Powers, J. S., Feng, X., Sanchez-Azofeifa, A., and Medvigy, D. 2018. Focus on tropical dry forest ecosystems
and ecosystem services in the face of global change. Environmental Research Letters 13: 090201.
Rundel, P. W., and Boonpragob, K. 1995. Dry forest ecosystem of Thailand. In S. H. Bullock, H. A. Mooney,
and E. Medina (Eds.), Seasonally dry tropical forests (pp. 93-123). Cambridge University Press,
New York.
Setalaphruk, C. and Price, L. L. 2007. Children’s traditional ecological knowledge of wild food resources: a
case study in a rural village in Northeast Thailand. Journal of Ethnobiology and Ethnomedicine 3:33.
Shirai, Y., and Rambo, A. T. 2008. The economic value of edible wild and semi-domesticated species sold in an
urban market in Khon Kaen municipality in Northeast Thailand. Khon Kaen Agr. J. 36: 69-78.
Sirival, N. 2008. The determination of elements in local vegetables, Sakonnakhon province. Proceeding of the
46th Kasetsart University Annual Conference, Bangkok, January 29th – Febuary 1st, 2008, Bangkok,
Thailand. p. 624-630.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 81
Somnasang, P. and Moreno-Black. 2000. Knowing, Gathering and Eating: Knowledge and attitude about wild
food in an Isan village in Northeastern Thailand. Journal of Ethnobiology 20(2): 197-216.
Smitinand, T. 2014. Thai plant names. The Forest Herbarium, Royal Forest Department, Bangkok, Thailand.
Srisawat, T., Suvarnasingh, A., and Maneenoon, K. 2016. Traditional medicinal plants notably used
to treat skin disorders nearby Khao Luang mountain hills region, Nakhon Si Thammarat, Southern
Thailand. Journal of Herbs, Spices and Medicinal Plants 22: 35-56.
Sseremba, G., Kabod, N. P., Kasharu, A. K., Jaggwe, J. N., Masanza, M., and Kizito, E. B. 2017. Diversity and
distribution of African indigenous vegetable species in Uganda. International Journal of Biodiversity
and Conservation 9(11): 334-341.
Tangkanakul, P., Trakoontivakorn, G., Auttaviboonkul, P., Niyomvit, B., and Wongkrajang, K. 2006. Antioxidant
activity of Northern and Northeastern Thai foods containing indigenous vegetables. Kasetsart J.
(Nat. Sci.) 40 (Suppl.): 47-58.
Tinrat, S. 2016. Antioxidant activities and total phenolic content of multi-colored fruits and vegetables in
Thailand. KKU Res. J. 21(1): 1-11.
Thongpukdee, A., Theppisit, C., and Thammaso, C. 2014. Ethnobotany survey of vetgetable plants
traditionally used in Kalasin Thailand. International Scholarly and Scientific Research and Innovation
8(7): 692-695.
Turreira-García, N., Vilkamaa, A. M., Byg, A., and Theilade, I. 2017. Diversity, knowledge and use of leafy
vegetables in Northern Thailand – maintenance and transmission of ethnobotanical knowledge
during urbanisation. Nat. Hist. Bull. Siam Soc. 62(1): 85-105.
Uperty, Y., Poudel, R. C., Shrestha, K. K., Rajbhandary, S., Tiwari, N. N., Shrestha, U. B., and Asselin, H. 2012.
Diversity of use and local knowledge of wild edible plant resources in Nepal. Journal of Ethnobiology
and Ethnomedicine 8: 16.
Yakoh, K., Weschasat, T., Suwannachote, P. 2018. Phenolic compound of 10 indigenous vegetables from
Surat Thani province. Journal of Thai Traditional and Alternative Medicine 16: 185-194.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 82
Figure 1. Map of the Puparn Royal Development Study Centre, Sakon Nakhon, Thailand
Source: https://www.iucn.org/content/combining-science-local-knowledge-ecosystem-based-adaptation-kok-
klang-village-sakon-nakhon
Figure 2. Classification of indigenous vegetables in the Puparn Royal Development Study Centre based on plant
habit
Figure 3. A comparison of species number within each forest type in the Puparn Royal Development Study
Centre
3%
14%
3%
8%
40%
2%
2%
14%
14%bamboo
climbing
fern
fungi
herb
palm
scandent
shrub
tree
0% 20% 40% 60% 80% 100%
Dipterocarp forest
Mixed Deciduous forest
Dry Evergreen forest
Wet areas
Percentage of area size
Number of species
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 83
Figure 4. Edible part of indigenous vegetables in the Puparn Royal Development Study Centre
Table 1. Habit, ecosystem and edible parts of indigenous vegetables in Puparn Royal Development Study Centre
Family Scientific name Vernacular
name
Habit Ecosystem
*
Edible parts**
D
F
M
D
F
D
E
F
W
A
U
st
S
t
S
h
F
l
F
r
W
h
Acanthaceae Phlogacanthus
pulcherrimus T.
Anderson.
Di pla chon Shrub / /
Alismataceae Limnocharis flava (L.)
Buchenau
Phak phai Herb / / /
Amanitaceae Amanita sp. Het ra ngok Fungi /
Amaranthacea
e
Alternanthera sessilis (L.)
R. Br. ex DC.
Phak pet thai Herb / / /
Ancistroclada
ceae
Ancistrocladus tectorius
(Lour.) Merr. Hu kwang
Climb
ing
/
Apiaceae Centella asiatica (L.)
Urb.
Phak nok Herb / / /
Hydrocotyle umbellata L. Phak waen
kaeo Herb / / /
Apocynaceae Aganonerion
polymorphum Pierre ex
Spire
Som lom Climb
ing
/ / /
Amphineurion
marginatum (Roxb.) D.J.
Middleton
Khruea sai tan Climb
ing
/ / /
Araceae Amorphophallus sp. Buk Herb / / / / /
Colocasia gigantea
(Blume) Hook f. Bon Herb / / /
Lasia spinosa (L.)
Thwaites
Phak nam Herb / / /
Arecaceae Calamus sp. Wai Scand
ent
/ / /
Caryota mitis Lour. Tao rang Palm / /
Asparagaceae Dracaena angustifolia
(Medik.) Roxb. Khon Khan Shrub
/ /
Asteraceae Acmella oleracea (L.)
R.K. Jansen
Phak khrat hua
waen Herb / / /
Blumea napifolia DC. Phak hon han Herb / /
Crassocephalum
crepidioides (Benth.) S.
Moors.
Phak kho on Herb / / /
Athyriaceae Diplazium esculentum
(Retz.) Sw. Phak kut Fern / / /
Auriculariacea
e
Auricularia auricula-
judae (Bull.) J.Schröt.
Het hu nu Fungi
/
Campanulacea
e Lobelia chinensis Lour. Phak khi som Herb / / /
Clusiaceae Garcinia cowa Roxb. ex
Choisy Cha muang Shrub / /
0
5
10
15
20
25
30
35
underground
stem
leave & shoot stem flower fruit whole plant
Nu
mb
er o
f p
lan
t sp
ecie
s
Edible parts of indigenous vegetables
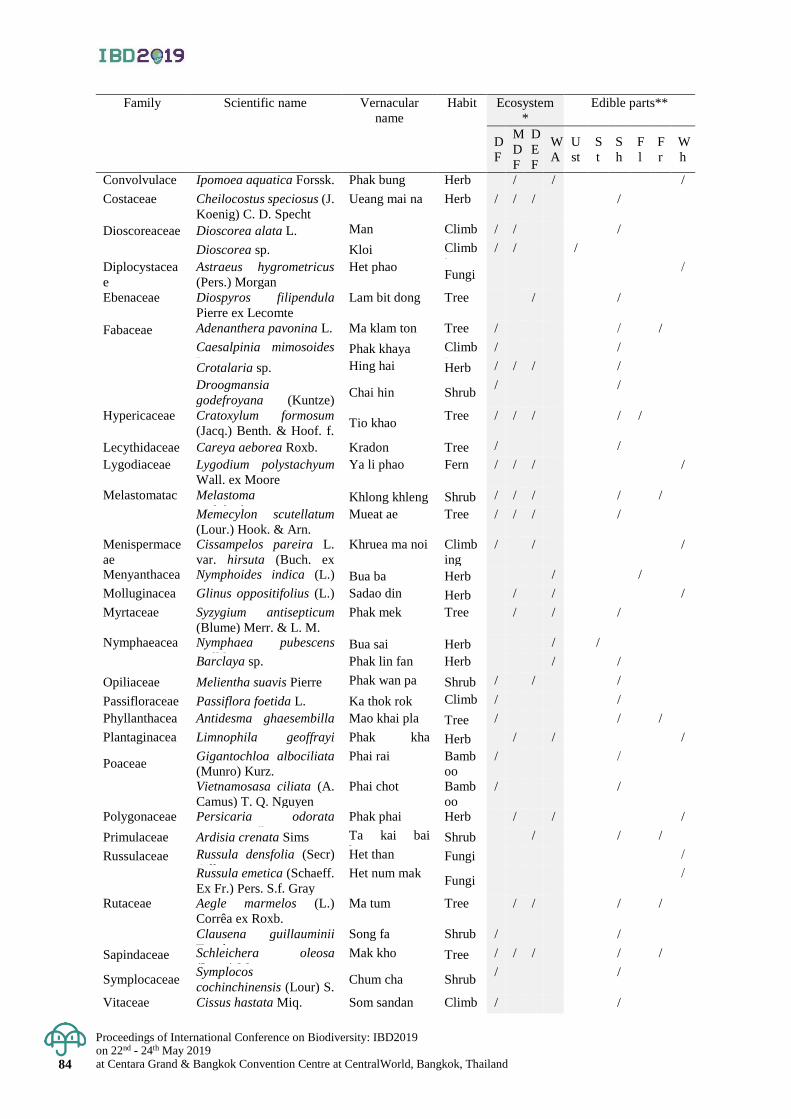
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 84
Family Scientific name Vernacular
name
Habit Ecosystem
*
Edible parts**
D
F
M
D
F
D
E
F
W
A
U
st
S
t
S
h
F
l
F
r
W
h
Convolvulace
ae
Ipomoea aquatica Forssk. Phak bung Herb / / /
Costaceae Cheilocostus speciosus (J.
Koenig) C. D. Specht
Ueang mai na Herb / / / /
Dioscoreaceae Dioscorea alata L. Man Climb
ing
/ / /
Dioscorea sp. Kloi Climb
ing
/ / /
Diplocystacea
e
Astraeus hygrometricus
(Pers.) Morgan
Het phao Fungi
/
Ebenaceae Diospyros filipendula
Pierre ex Lecomte
Lam bit dong Tree / /
Fabaceae Adenanthera pavonina L. Ma klam ton Tree / / /
Caesalpinia mimosoides
Lam. Phak khaya Climb
ing
/ /
Crotalaria sp. Hing hai Herb / / / /
Droogmansia
godefroyana (Kuntze)
Schindl.
Chai hin Shrub / /
Hypericaceae Cratoxylum formosum
(Jacq.) Benth. & Hoof. f.
ex Dyer subsp. formosum
Tio khao Tree / / / / /
Lecythidaceae Careya aeborea Roxb. Kradon Tree / /
Lygodiaceae Lygodium polystachyum
Wall. ex Moore
Ya li phao Fern / / / /
Melastomatac
eae
Melastoma
malabathricum L. Khlong khleng Shrub / / / / /
Memecylon scutellatum
(Lour.) Hook. & Arn.
Mueat ae Tree / / / /
Menispermace
ae
Cissampelos pareira L.
var. hirsuta (Buch. ex
DC.) Forman
Khruea ma noi Climb
ing
/ / /
Menyanthacea
e
Nymphoides indica (L.)
Kuntze Bua ba Herb / /
Molluginacea
e
Glinus oppositifolius (L.)
A. DC.
Sadao din Herb / / /
Myrtaceae Syzygium antisepticum
(Blume) Merr. & L. M.
Phak mek Tree / / /
Nymphaeacea
e
Nymphaea pubescens
Willd Bua sai Herb / /
Barclaya sp. Phak lin fan Herb / /
Opiliaceae Melientha suavis Pierre Phak wan pa Shrub / / /
Passifloraceae Passiflora foetida L. Ka thok rok Climb
ing
/ /
Phyllanthacea
e
Antidesma ghaesembilla
Gaertn.
Mao khai pla Tree / / /
Plantaginacea
e
Limnophila geoffrayi
Bonati
Phak kha
yaeng Herb / / /
Poaceae Gigantochloa albociliata
(Munro) Kurz.
Phai rai Bamb
oo
/ /
Vietnamosasa ciliata (A.
Camus) T. Q. Nguyen
Phai chot Bamb
oo
/ /
Polygonaceae Persicaria odorata
(Lour.) Soják
Phak phai Herb / / /
Primulaceae Ardisia crenata Sims Ta kai bai
kwang Shrub / / /
Russulaceae Russula densfolia (Secr)
Gill.
Het than Fungi /
Russula emetica (Schaeff.
Ex Fr.) Pers. S.f. Gray
Het num mak Fungi
/
Rutaceae Aegle marmelos (L.)
Corrêa ex Roxb.
Ma tum Tree / / / /
Clausena guillauminii
Tanaka
Song fa Shrub / /
Sapindaceae Schleichera oleosa
(Lour.) Merr.
Mak kho Tree / / / / /
Symplocaceae Symplocos
cochinchinensis (Lour) S.
Moore
Chum cha Shrub / /
Vitaceae Cissus hastata Miq. Som sandan Climb
ing
/ /
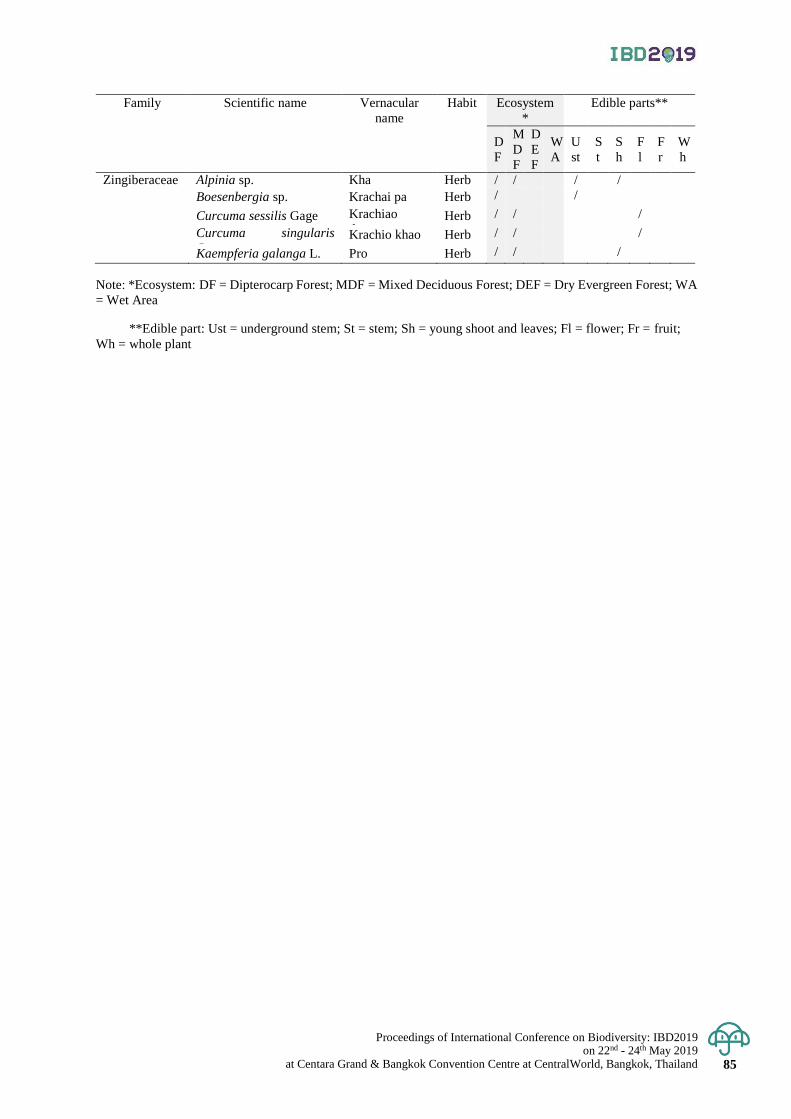
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 85
Family Scientific name Vernacular
name
Habit Ecosystem
*
Edible parts**
D
F
M
D
F
D
E
F
W
A
U
st
S
t
S
h
F
l
F
r
W
h
Zingiberaceae Alpinia sp. Kha Herb / / / /
Boesenbergia sp. Krachai pa Herb / /
Curcuma sessilis Gage Krachiao
daeng Herb / / /
Curcuma singularis
Gagnep. Krachio khao Herb / / /
Kaempferia galanga L. Pro Herb / / /
Note: *Ecosystem: DF = Dipterocarp Forest; MDF = Mixed Deciduous Forest; DEF = Dry Evergreen Forest; WA
= Wet Area
**Edible part: Ust = underground stem; St = stem; Sh = young shoot and leaves; Fl = flower; Fr = fruit;
Wh = whole plant

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 86
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 86 - 88
Phytochemical screening and antimicrobial activity of Indian almond leaf extracts
Ratthayaporn Thansuwan1*, Jirapron Jitturongarpron1, Jittima Buatib1,
Thanyaporn Tangjaroenchai1,2and Noppadon Muangsue3 1Demonstration School University of Phayao, Muang District, Phayao, Thailand
2School of Science, University of Phayao, Muang District, Phayao, Thailand 3School of Allied Health Sciences, University of Phayao, Muang District, Phayao, Thailand
*Corresponding author e-mail: [email protected]
Abstract: The antimicrobial activity of Indian almond leaf extracts, prepared by extraction using water, %70 ethyl
alcohol and 9 5% ethyl alcohol, were tested for their efficacy by paper disc diffusion method on Muller Hinton
Agar (MHA). The results showed that all three extracts could inhibit mainly the growth of Staphylococcus aureus
with the inhibition zones of 4.33 mm, 3.33 mm and 6.67 mm. respectively. No growth inhibition was observed with
other microbial cultures of Escherichia coli, Klebsiella sp., Citrobacter sp. and Enterobacter sp. Chemical analysis
by Gas Chromatography–Mass Spectrometry (GC-MS) revealed that there were 31 compounds that were matched with
the data in the library,of which, benzenethiol 1,2,3- benzenetriol or “Pyrogallol” was found to be the most abundance.
Keywords: Antimicrobial activity, Indian almond leaf extract, Phytochemical analysis, GC-MS
Introduction
Indian almond tree (Terminalia catappa) is a Combretaceous plant belonging to the tropical almond family.
It is widely distributed throughout all regions of Thailand. In aquaculture, the Indian almond leaves have been claimed
to have active compound(s) that promote wound healing. It has been used in Thailand for curing injured siamese
fighting fishs, presumably to increase thickness of keratin layer of the fish. In the literature, the leaf part of the plant
has been shown to have a potential to use as an alternative treatment for chemical substances and antibiotics [1].
The objectives of this study were to evaluate the antibacterial activity of the Indian almond leaf extracts in order to get
the most effective extracts that could inhibit the growth of microbial pathogens, and to evaluate the potential antimicrobial
compounds in the Indian almond leaf extracts by Gas Chromatography–Mass Spectrometry (GC-MS) analysis.
Materials and Methods
Extraction of Indian almond leaves
The Indian almond leaves, collected from School of Agriculture and Natural Resources, University of Phayao,
were washed with deionized water and immediately dried in an oven at 50 °C for 24 hours. The fried leaves were
then ground into powder by using motar and pestle. The leaf powder was then extracted separately with three
types of solvent, namely water, 70% ethanol and 95% ethanol for 3 days (100 g per 1000 ml). After filtration, the
extract was evaporated by using a rotary evaporator to dryness. The extracts were kept at 37°C until being used.
Antibacterial activity of Indian almond leaf extracts
Antibacterial tests were performed using disc agar diffusion method. The bacterial suspension of
Staphylococcus aureus, Escherichia coli, Klebsiella sp., Citrobacter sp. and Enterobacter sp. was each diluted to
1.5×108 CFU/ml and swabbed on the surface of MHA agar. Plates were incubated for 24 hours at room temperature.
Antimicrobial activity was detected by measuring the inhibition zone around the disc (in mm.). Tetracycline (TE 30 μg/disc)
was used as positive control.
Phytochemical analysis of Indian almond leaf extracts by gas chromatography – mass spectrometry (GC-MS)
The Indian almond leaf extracts were subjected to Gas Chromatography-Mass Spectrometry (GC-MS)
analysis and compounds were matched with data in the library.
Result & Discussion
The efficacy test for antimicrobial activity of the leaf extracts was conducted by disc diffusion method
on Muller Hinton agar (MHA). The lowest concentrations of the three Indian almond leaf extracts prepared by using
water, 70% ethanol and 95% ethanol were 31.25, 31.25 and 15.63 mg/ml, respectively. The results obtained from
the disc diffusion method showed that Staphylococcus aureus the inhibition zone could only be observed with
Staphylococcus aureus with the diameters of 4.33 mm for the water extract, 3.33 mm for the 70% ethanol and
6.67 mm for the 95% ethanol (Table 1). No significant inhibition zones were observed with other microbial cultures,
including Escherichia coli, Klebsiella sp., Citrobacter sp. and Enterobacter sp. Therefore, it was suggested that
the antimicrobial activities of the extracts are relatively specific.
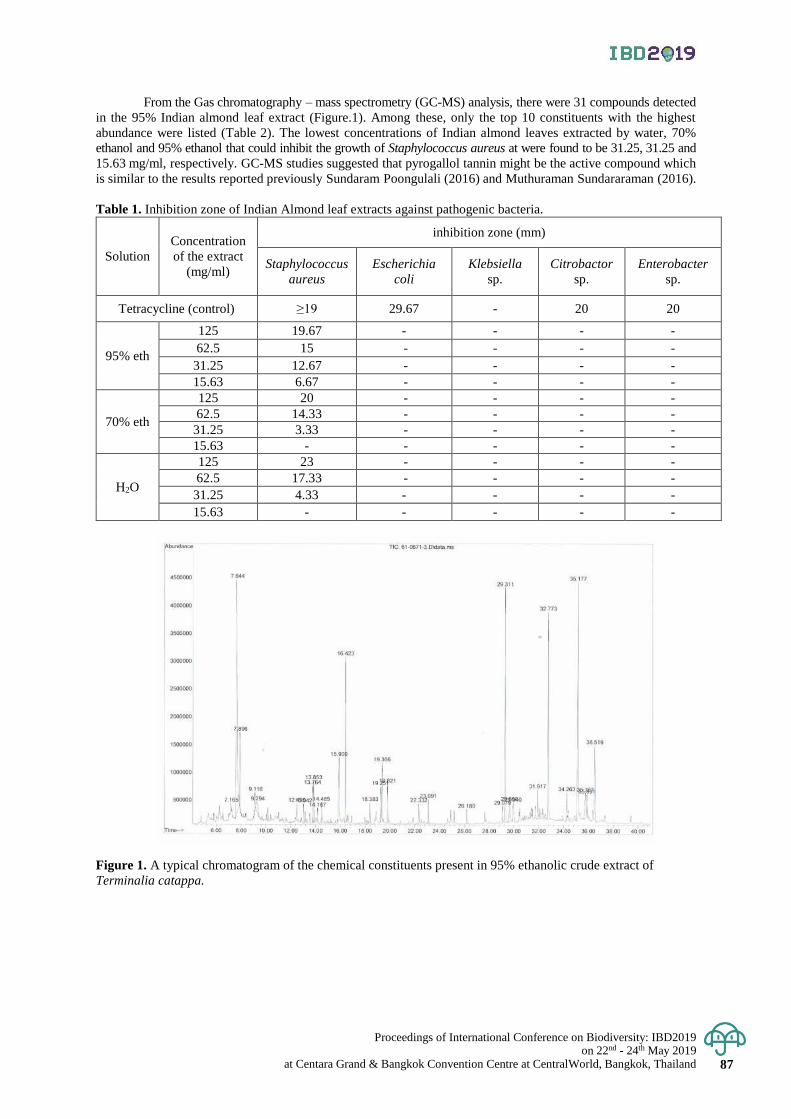
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 87
From the Gas chromatography – mass spectrometry (GC-MS) analysis, there were 31 compounds detected
in the 95% Indian almond leaf extract (Figure.1). Among these, only the top 10 constituents with the highest
abundance were listed (Table 2). The lowest concentrations of Indian almond leaves extracted by water, 70%
ethanol and 95% ethanol that could inhibit the growth of Staphylococcus aureus at were found to be 31.25, 31.25 and
15.63 mg/ml, respectively. GC-MS studies suggested that pyrogallol tannin might be the active compound which
is similar to the results reported previously Sundaram Poongulali (2016) and Muthuraman Sundararaman (2016).
Table 1. Inhibition zone of Indian Almond leaf extracts against pathogenic bacteria.
Solution
Concentration
of the extract
(mg/ml)
inhibition zone (mm)
Staphylococcus
aureus
Escherichia
coli
Klebsiella
sp.
Citrobactor
sp.
Enterobacter
sp.
Tetracycline (control) ≥19 29.67 - 20 20
95% eth
125 19.67 - - - -
62.5 15 - - - -
31.25 12.67 - - - -
15.63 6.67 - - - -
70% eth
125 20 - - - -
62.5 14.33 - - - -
31.25 3.33 - - - -
15.63 - - - - -
H2O
125 23 - - - -
62.5 17.33 - - - -
31.25 4.33 - - - -
15.63 - - - - -
Figure 1. A typical chromatogram of the chemical constituents present in 95% ethanolic crude extract of
Terminalia catappa.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 88
Table 2. Major constituents of Indian almond leaves extracted by 95% ethanol based on by Gas chromatography – mass
spectrometry (GC-MS).
Conclusion
The antimicrobial efficacy of Indian Almond leaf extracts prepared by using three kinds of solvent water,70%
ethanol and 95% ethanol were conducted by testing the ability of the extracts to inhibit the growth of bacteria.
The results showed that the 95% ethanol extract are the best in inhibiting the growth of Staphylococcus aureus.
Water is lower ability to inhibit Staphylococcus aureus and 70% ethanol is the lowest. But it has not affect the inhibition
of Escherichia coli.
Acknowledgement
This project was supported by the Science Classroom in University Affiliated School (SCiUS) under the
Ministry of Science and Technology, jointed by Demonstration School, School of Science University of Phayao
and School of Allied Health Sciences, University of Phayao.
References
Francisco José Mininel, Carlos Sérgio Leonardo Junior, Lívia Greghi Espanha, Flávia Aparecida Resende, Eliana
Aparecida Varanda, Clarice Queico Fujimura Leite. et al. 2014. Characterization and Quantification of
Compounds in the Hydroalcoholic Extract of the Leaves from Terminalia catappa Linn. (Combretaceae)
and Their Mutagenic Activity. Evidence-Based Complementary and Alternative Medicine : Hindawi
Publishing Corporation.
K.-T. CHUNG, Z. LU, M. W. CHOU. 2006. Mechanism of Inhibition of Tannic Acid and Related Compounds
on the Growth of Intestinal Bacteria. Food and Chemical Toxicology. 36(12), 1053-60.
Nantarika Chansue and Nongnut Assawawongkasem. 2008. The in vitro Antibacterial Activity and Ornamental
Fish Toxicity of the Water Extract of Indian almond leaves (Terminalia catappa Linn.). KKU veterinary
journal. 18(1), 36-45.
Sundaram Poongulali and Muthuraman Sundararaman. 2016. Antimycobacterial, Anticandidal and Antioxidant
Properties of Terminalia catappa and Analysis of Their Bioactive Chemicals. International Journal of
Pharmacy and Biological Sciences. 6(2), 69-83.
W. Purivirojkul and N. Areechon. 2006. Antibacterial Activity and Toxicity of Indian Almond
(Terminalia catappa) extract in Siamese Fighting Fish (Betta splendens Regan). Proceedings of 44th
Kasetsart University Annual Conference: Fisheries. 109-116.
No. R.T. min Compounds Molecular formula Molecular weight % peak area
1 7.644 Benzenetriol Benzene-1,2,3-
triol Pyrogallol C6H6O3 126.111 g/mol 100.00%
2 35.177 Stigmast-5-en-3-ol C29H50O 414.718 g/mol 74.31%
3 29.311 Tetracosahexaene C24H38 326.568 g/mol. 48.60%
4 32.773 Vitamin E C29H50O2 430.706 g/mol 42.40%
5 16.423 Hexadecanoic acid ethyl ester C18H36O2 284.48 g/mol 27.45%
6 7.896 1,3a-epoxy-3ah-indene - - 22.24%
7 36.519 Aristol-a-en-8-one C15H22O 218.34 g/mol 22.47%
8 15.9 Hexadecanoic acid
Hexadecanoate C16H32O2 256.4 g/mol 19.71%
9 19.366 Ethyl-9-octadecenoate C20H38O2 310.522 g/mol 10.23%
10 35.767 Olean-12-en-3-ol C32H52O2 468.766 g/mol 8.72%

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 89
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 89 - 93
Study of chemical composition and antioxidant properties of Sangyod and
Tubtimchumpae rice bran oil
Pavinee Yampeng1*, Hathairat Rimkeeree1 and Supanida Winitchai2
1Faculty of Agro-Industry, Kasetsart University, Chatuchak District, Bangkok, Thailand 2 Kasetsart Agricultural and Agro-Industrial Product Improvement Institute (KAPI), Kasetsart University,
Chatuchak District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Sangyod rice bran oil is an oil extracted from a local rice strain grown in Phattalung Province,Thailand.
Tubtimchumpae rice bran oil is another kind oil obtained from Tubtimchumpae rice grown in Kamphaengphet Province,
Thailand. Both types of red rice containing high anthocyanin were prepared for the oils by cold-press extraction.
Since they do not pass through heat processing, they are rich in nutritional values. The aim of this research was to study
the chemical compositions and antioxidant properties of Sangyod and Tubtimchumpae rice bran oils. The results
showed that the chemical compositions of in terms of free fatty acids, acid value, peroxide value, unsaponifiable
matter and saponification of the two rice bran oils were significantly different (p≤0.05). Both rice bran oils consisted
of high unsaturated fatty acid contents (80.46 and 80.35%, respectively), with oleic acid and linoleic acid as main
components. However, Tubtimchumpae rice bran oil contained higher tocopherol than Sangyod rice bran oil
(11.88 and 9.13 mg/100g, respectively). Total phenolic contents were found to be 1.48±0.05 and 1.54±0.03 mg
gallic acid equipvalent/100 g oil, respectively. DPPH (IC50) values were 13.90 and 13.97 µ g/ml, ABTS (IC50)
were 15.99 and 16.09 µ g/ml, respectively. In addition, the two types of rice bran oil were found to possess
important nutritional values, such as unsaturated fatty acids, antioxidants and vitamin E (α-tocopherol). Therefore,
it was suggested that these oils can be used to increase health benefits in food products.
Keywords: Sangyod rice bran oil, Tubtimchumpae rice bran oil, chemical composition, antioxidant properties.
Introduction
Rice bran oil (RBO) is the oil extracted from the germ and inner husk of rice. RBO contains significant
unsaturated fatty acids, saturated fatty acids and micronutrients, such as tocopherol, tocotrienol and oryzanol.
Several studies have shown that RBO has an effect on metabolic activities, causing the decrease of bad cholesterol LDL
but without significant affecting the blood level of HDL cholesterol (Yoshino et al., 1989, Qureshi et al., 1991,
Hegsted et al., 1990, Kahlon et al., 1992, Hegsted and Windhauser 1993). RBO containing high antioxidant activities
is known to have health benefits and for improving stability of foods during the storage (Shahid et al., 2005). RBO
also contains high concentration of tocopherols compared with other seed oils (Kao and Luh, 2010). Previous
studies have also shown that the rice grains with dark red or purple color contain high amount of anthocynanins,
a group of natural pigments found in deep purple or reddish fruits and vegetables (Yawadio et al., 2013). On the other
hand, rice grains with red color consist high amount of polyphenols, phenolics and flavonoids (Gunaratne et al., 2013;
Sompong et al., 2011). Sangyod rice is a traditional rice variety grown in Phatthalung province for more than a hundred years.
Sangyod Muang Phatthalung rice is soft with approximately 14-15% of the amylose content. The analysis of its nutrient
volume has been shown that Sangyod rice has high nutritional value, iron, vitamin B and niacin (Department of Health, 2004).
For Tubtimchumphae rice, it is a new Thai rice varieties which is a hybrid between Jasmin rice and Sangyod rice.
It contains high amount of phenolic, flavonoid, Vitamin E (α-tocopherol and γ-oryzanol) (Rice Department, 2016).
The objective of this research work was to study the chemical composition and antioxidant properties of RBOs obtained
from the two rice varieties, in order to provide information for their uses in food and supplementary products.
Materials and Methods
Materials
Sangyod rice bran oil (SRBO) was obtained from Phattalung Province’s Community Enterprise and
Tubtimchumpae rice bran oil (TRBO) from Kamphaengphet Province’s Community Enterprise.
Physical parameters
The color of RBOs was measured by the CIE L*, a*, b* with standard illuminant D65 and standard observer 10°
by Spectrophotometer (CM-3500d, Minolta, Japan)

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 90
Chemical parameters
Acid value (AV), free fatty acid (FFA), Peroxide value (PV), iodine value (IV), saponification value (SV) and
unsaponifiable matter were determined according to the methods described in AOAC (2000).
Determination of fatty acid composition
The fatty acid compositions were determined by GC-FID according to the methods described by Kasetsart
Agricultural and Agro-Industrial Product Improvement Institute, Thailand. Capillary GC (Agilent 6890 Serie) equipped
with a capillary column (BPX 70 SGE; 70% Cyanopropyl Polysilphenylene-siloxane; 30 m×0.25 mm–0.25 μm
film) was used. The GC conditions operated at the initial temperature of 160 °c for 0.5 min, then increased to 160-200 °c
at 10°c /min and the final temperature of 200 °c for 4.6 min with a total run time of 9.1 min. The flow rate of gas (Nitrogen)
was 1 ml/min. injection volume of 1 μl were used. The fatty acid composition was obtained by comparison of the peak
retention times with the respective fatty acids standards.
Determination of total phenolic content
The total phenolic content (TPC) of the crude RBO extracts was determined using Folin–Ciocalteu’sreagent
method as described by Singleton et al., (1999) Oil extracts (300 µL) were mixed with 1.5 ml of Folin-Ciocalteu’s
reagent, left for 6 min and subsequently 1.25 ml of 7% sodium carbonate solution was added. The mixture was diluted
to 3 ml with deionized water. The absorbance of the resulting mixture was read at 725 nm using a UV visible
spectrophotometer (UV 1601, Shimadzu, Japan) after leaving for 30 min at the room temperature (25 °C). The measurement
was compared to a standard curve of gallic acid concentrations, which were expressed mg gallic acid equivalents/ g sample.
Determination of α-tocopherol
The α-tocopherol content in RBO was determined by Institute of Food Research and Product Development,
Thailand, using an in-house method based on BS EN 12823-1:2000
Determination of DPPH radical scavenging capacity
DPPH radical-scavenging effects were determined by using the method modified from that reported previously
by Brand-Williams et al., (1995). A solution of DPPH was prepared in methanol, and the solution of RBO was dissolved
in DMSO to obtain the concentration between 5 and 30 µg/mL. Then, RBO and 1 ml of 0.1 mM DPPH solution
were mixed in test tube. The mixture was shaken and incubated for 30 min in the dark at room temperature. The absorbance
was determined at 517 nm by UV-VIS spectrophotometer (Shimadzu 160A, Japan). The radical scavenging activity
was expressed as IC50 value that denotes the concentration of the sample required to scavenge 50% of DPPH radicals.
Determination of ABTS radical scavenging capacity
ABTS assay was determined according to the method described by Re et al., (1999) ABTS radical was generated
by reacting 7 mM ABTS and 2.45 mM potassium persulfate at room temperature in dark for 16 h. The ABTS solution
was diluted with 80 % ethanol to an absorbance around 0.700 at 734 nm. 2 ml of ABTS solution was added to 0.1 ml
of the extracts and mixed thoroughly. The reaction mixture was kept at room temperature for 6 min, absorbance
was measured at 734 nm by UV-VIS spectrophotometer (Shimadzu 160A, Japan).
Result & Discussion
Physical parameters: The CIE Lab color measurement method was determined by its color coordinates: L* represent
the difference between light (L*= 100) and dark (L* = 0), a* represent the difference between green (–a*) and red
(+a*) and b* represent the difference between blue (-b*) and yellow (+b*). It was found that the color of SRBO
(L* = 56.48, a* = 15.60 and b* = 93.43) was darker, more green and yellow than the color of TRBO (L* = 43.64,
a* = 13.97 and b* = 74.11). Therefore, the color of two varieties rice bran oil were significantly different (p≤0.05).
based on the mentioned physical parameters.
Chemical parameters: The AV and FFA values in the samples of oil or fat were determined after the hydrolysis
of triglycerides by the enzyme lipase in which its activity was increased rapidly after the milling process. In terms
of lipid quality, the value of PV content is used as a parameter, and the values of IV is used to indicate the amount
of unsaturation in RBO. The results showed that the values of AV, FFA, PV and IV of SRBO were 13.36 mg KOH/g,
6.71%, 1.78 mg eq/kg oil and 96.86 g Iodine/100 g oil, respectively. While the values of TRBO were 7.12 mg
KOH/g oil, 3.58 %, 1.91 mg eq/kg oil and 100.74 g Iodine/100 g oil, respectively. CODEX standard (1999) has
suggested that the maximum levels of AV for Crude rice bran oil is 4 mg KOH/g, the maximum level of PV is 10
mg eq/kg oil, and the recommended IV value is between 90–105 g Iodine/100 g oil. According to Tao et al.(1993),
the level of FFA value for RBO should be not over 5%. Our results showed that the RBO samples had the AV
and FFA contents higher than the recommendations. This could be affected by the raw material quality, as the
bran did not stabilize before being used. The bran quality deterioration may be due to the presence lipase activity.
For the PV and IV contents, the values of both rice bran oils were lower than those of the CODEX standard.
Saponification value is the reaction that free hydroxide breaks the ester bonds between the fatty acids and glycerol
of a triglyceride, resulting in free fatty acids and glycerol. Unsaponification value is the measure of non-lipids

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 91
constituents (sterols, pigments, and hydrocarbons) in the oil (Bodger et al. 1982). In this study, the saponification
value and unsaponifiable matter of SRBO were determined to be 196.48 mg KOH/g oil and 2.29%, and of TRBO
were 189.49 mg KOH/g oil and 3.49%, respectively. Statistical analysis showed that there were significant
differences (p≤0.05) in the saponification values and unsaponifiable matter of two varieties oils.
Total phenolic content, DPPH and ABTS radical scavenging capacity: The result of total phenolic content
(TPC) was expressed as gallic acid equivalent, showing that the phenolic compounds in RBO contributes to their
antioxidant properties as summarized in Table 2. The total phenolic contents of SRBO (1.48 mg GAE/100g) and
TRBO (1.54 mg GAE/100g) were not significantly different (p>0.05). For DPPH and ABTS radical scavenging
activities of the RBO extracts, their values were expressed as IC50 which is inversely proportional to the antioxidant
activity. Our results showed that the DPPH and ABTS radical scavenging activities of the two varieties RBO extracts
were not significantly different (p>0.05). On the other hand, the α-tocopherol contents were found to be higher in TRBO
(9.13 mg/100g oil) than in SRBO (11.88 mg/100g oil), which were significantly different in both varieties (p ≤ 0.05).
It is well known that tocopherols possesses high antioxidant properties and the greatest vitamin E potency (Chen
and Bergman, 2015). Therefore, DPPH, ABTS radical scavenging activity and Total phenolic content of TRBO
were found to be higher than those values in SRBO.
Fatty Acid Composition: The fatty acid compositions of SRBO and TRBO are showed in Table 3.
There were eight fatty acids identified including oleic acid (C18:1), linoleic acid (C18:2), palmitic acid (C16:0),
stearic acid (C18:0), linolenic acid (C18:3), arachidonic acid (C20:0), myristic acid (C14:0) and palmitoleic
(C16:1). The saturated fatty acids were found in both rice bran oils such as myristic, palmitic, stearic and arachidonic
acids with the concentrations of 0.34–0.45%, 15.80–16.35%, 2.23–2.40% and 0.97-0.99%, respectively. The unsaturated
fatty acids were found to be palmitoleic, oleic, linoleic and linolenic acids with the concentration of 0–0.14%,
41.74–42.04%, 36.22–36.89% and 0.97-0.99%, respectively. The main components of the fatty acids were oleic
acid, linoleic acid and palmitic acid. However, the fatty acid contents of SRBO and TRBO were not significantly
different (p ≥ 0.05). According to Mingyail. et al., (2017) the major fatty acids in the RBO samples are oleic acid,
linoleic acid and palmitic acid. The content of the unsaturated fatty acids was apparently higher than the saturated
fatty acids.
Conclusion
The results of this study showed that the fatty acid profiles of SRBO and TRBO contain a high percentage
of the unsaturated fatty acids (approximately 80%), with the main components of oleic acid and linoleic acid. The results
also suggested that, the two rice bran oils can be used as a raw material in the production of functional foods. They
provided different physicochemical and antioxidant properties, which are potentially beneficial for the nutrition and
health of human.
Acknowledgement
This wor was supported by Faculty of Agro-Industry, Kasetsart University, Kasetsart Agricultural and Agro
Industrial Product Improvement Institute (KAPI) and Kasetsart University Research and Development Institute (KURDI).
Referenes
AOAC 2000, Oils & fats. In H. William (Ed.). Official methods of analysis of AOAC International (pp. 1-69),
USA: AOAC International.
Bodger. D., Davis, J.B., Farmery. D., Hammonds. T.W., Harper, A.J., Harris, R.V., Hebb, L., MacFarlane, N.,
Shanks, P., and Southwell, K. 1982. An investigation of the extraction, refining and composition of oil
from Winged bean (Psophocarpus tetragonolobus D.C.). J Am Oil Chem Soc. 59:523–530.
Brand-Williams, W., Cuvelier, M.E., and Berset, C. 1995. Use of a Free Radical Method to Evaluate Antioxidant
Activity. Lebensmittel Wissenschaft und Technologie, 28: 25–30.
Chen, M.H., and Bergman, C.J. 2005. A rapid procedure for analysing rice bran tocopherol, tocotrienol and
c-orizanol contents. J Food Anal 18:139–151.
CODEX STANDARD. 1999. CODEX standard for named vegetable oils. Department of Health. 2004. Nutrition
analysis of Sanyod Phatthalung rice. Department of Health, Ministry of Public Health, Thailand.
Gunaratne, A., Wu, K., Li, D., Bentota, A., Corke, H., and Cai, Y.Z. 2013. Antioxidant activity and nutritional
quality of traditional red-grained rice varieties containing proanthocyanidins. Food Chem 138: 1153-1161.
Hegsted, M., Windhauser, M., Lester, S., and Morris, S. 1990. Stabilized rice bran and germ product on hypercholersterolemia.
Food Chem. News 32(37).
Hegsted, M., and Windhauser, M. M. 1993. Reducing human heart disease risk with rice bran. La. Agric. 36(3):22-24.
Institute of food research and product development: Kasetsart University. Thailand: IFRPD [Online] Available
from:http://ifrpd.ku.ac.th/en/about/index.php.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 92
Kahlon, T., Saunders, R., Sayre, R., Chow, F., Chin, M., and Betschart, A. 1992. Cholesterol-lowering effects
of rice bran and rice bran oil fraction in hypercholesterolemic hamsters. Cereal Chem. 69:485-489.
Kao, C., and Luh, B. S. 1991. in Rice Utilization. Vol. 2. B. S. Luh, ed. Avi: New York. Pages 295-311.
Qureshi, A. A., Qureshi, N., and Wright, J. J. K. 1991. Lowering of serum cholesterol in hypercholesterolemic
humans by tocotrienols (palmvitee). Am. Clin. Nutr. 53:s1021-s1026.
Mingyai1, S., Kettawan, A., Srikaeo, K., and Singanusong, Riantong. 2017. Physicochemical and Antioxidant
Properties of Rice Bran Oils Produced from Colored Rice Using Different Extraction Methods. J Oleo
Sci. 66, (6) 565-572.
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., and Yang, M. 1999. Rice-Evans, C. (1999) Antioxidant
Activity Applying An Improved ABTS Radical Cation Decolorization Assay. Free Radical Biology and Medicine
26: 1231–1237.
Rice Department. 2016. Tubtimchumpae rice (RD69). Rice Department, Ministry of Agriculture and Cooperatives,
Thailand. Shahid, I., Bhanger, M.I., and Farooq, A. 2005. Antioxidant properties and components of
some commercially available varieties of rice bran in Pakistan. Food Chem 93: 265-272.
Singleton, V.L., Orthofer, R., and Lamuela-Ravento, R.M. 1999. Analysis of total phenols and other oxidation
substrates and antioxidants by means of Folin-Ciocalteu Reagent. Methods in Enzymology 299: 152-178.
Sompong, R., Siebenhandl, E.S., Linsberger, M.G., and Berghofer, E. 2011. Physicochemical and antioxidative
properties of red and black rice varieties from Thailand, China and Sri Lanka. Food Chem 124: 132-140.
Tao, J., Rao, R., and Liuzzo, J. 1993. Microwave heating for rice bran stabilization. J. Microw. Power
Electromagn. Energy 28,156-164.
Yawadio, R., Kikuzaki, H., and Konishi, Y. 2007. Antioxidant activity of various extracts and fractions of Chenopodium
quinoa and Amaranthus spp. seeds. Food Chem 106: 760-766.
Yoshino, G., Kazumi, T., and Amano, M. 1989. Effects of y-oryzanol and probucol on hyperlipidemia. Current
Ther. Res. 45:975-982.
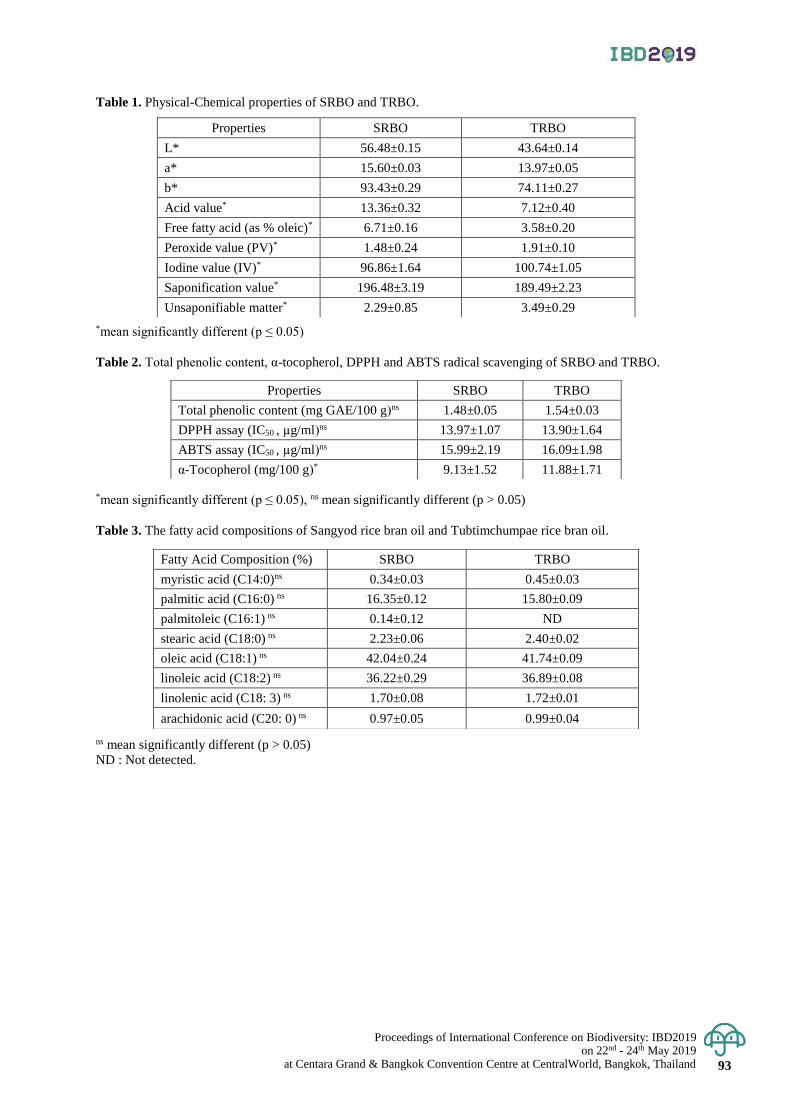
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 93
Table 1. Physical-Chemical properties of SRBO and TRBO.
*mean significantly different (p ≤ 0.05)
Table 2. Total phenolic content, α-tocopherol, DPPH and ABTS radical scavenging of SRBO and TRBO.
*mean significantly different (p ≤ 0.05), ns mean significantly different (p > 0.05)
Table 3. The fatty acid compositions of Sangyod rice bran oil and Tubtimchumpae rice bran oil.
ns mean significantly different (p > 0.05)
ND : Not detected.
Properties SRBO TRBO
L* 56.48±0.15 43.64±0.14
a* 15.60±0.03 13.97±0.05
b* 93.43±0.29 74.11±0.27
Acid value* 13.36±0.32 7.12±0.40
Free fatty acid (as % oleic)* 6.71±0.16 3.58±0.20
Peroxide value (PV)* 1.48±0.24 1.91±0.10
Iodine value (IV)* 96.86±1.64 100.74±1.05
Saponification value* 196.48±3.19 189.49±2.23
Unsaponifiable matter* 2.29±0.85 3.49±0.29
Properties SRBO TRBO
Total phenolic content (mg GAE/100 g)ns 1.48±0.05 1.54±0.03
DPPH assay (IC50 , µg/ml)ns 13.97±1.07 13.90±1.64
ABTS assay (IC50 , µg/ml)ns 15.99±2.19 16.09±1.98
α-Tocopherol (mg/100 g)* 9.13±1.52 11.88±1.71
Fatty Acid Composition (%) SRBO TRBO
myristic acid (C14:0)ns 0.34±0.03 0.45±0.03
palmitic acid (C16:0) ns 16.35±0.12 15.80±0.09
palmitoleic (C16:1) ns 0.14±0.12 ND
stearic acid (C18:0) ns 2.23±0.06 2.40±0.02
oleic acid (C18:1) ns 42.04±0.24 41.74±0.09
linoleic acid (C18:2) ns 36.22±0.29 36.89±0.08
linolenic acid (C18: 3) ns 1.70±0.08 1.72±0.01
arachidonic acid (C20: 0) ns 0.97±0.05 0.99±0.04

Proceeding of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 94
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 94 - 101
Checklist of bee genus Megachile Latreille, 1802 in Thailand (Hymenoptera: Megachilidae)
Nontawat Chatthanabun and Natapot Warrit*
Department of Biology & Center of Excellence in Entomology, Faculty of Science, Chulalongkorn University,
Pathum Wan District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: A checklist of Thai megachilid bees is reported for the first time. Species names listed were recorded
from specimens collected throughout Thailand and deposited at the Chulalongkorn University Natural History
Museum, Bangkok and through literature review. A total of 30 Thai Megachile species are listed with locality
information and notes on floral records, diagnostic characters, and nesting habitats (when appropriate). Due to
their abundance, we suggest that Megachile (Amegachile) bicolor, M. (Callomegachile) disjuncta, M. (Creightonella) fraterna,
and M. (Pseudomegachile) velutina are good candidates for being future crop pollinators.
Keywords: Diversity, Leafcutter bee, Pollination, Pollinator, Resin bee.
Introduction
Megachile Latreille, 1802 (Hymenoptera; Megachilidae) is one of the most diverse bee genera that include
leafcutter, resin, and mason bee. Megachile has a wide distribution and diverse habitats in most regions of the world
(Michener, 2007; Gonzalez et al., 2012). Presently, over 1,500 species are already named and described (Michener, 2007;
Ascher & Pickering, 2019). These bees include many important pollinators for natural ecosystems and agricultural
areas, e.g. Megachile rotundata, a well-known alfalfa crop pollinator (Delaplane & Mayer, 2000; Michener, 2007).
Historically, Southeast Asian megachilid bees are poorly known, underrepresented in collections, and inadequately
explored (Engel & Gonzalez, 2011; Gonzalez & Engel, 2012). Therefore, megachilid bees in Thailand are not well
recognized and protected from various anthropogenic threats such as habitat destruction and insecticide usages in
agriculture areas. This work attempts to contribute to the basic knowledge of Thai megachilid bee fauna. The checklist
comprises number of species and valid names, their distributions in Thailand, and notes on some species.
Materials and Methods
588 specimens (368♀, 220♂) collected throughout Thailand and deposited at the Chulalongkorn University
Natural History Museum (CUNHM) since 1962 were examined (see in Table 1). In addition, literature on Thai Megachile
taxonomic treatments were reviewed following Tadauchi and Tasen (2009), Engel and Gonzalez (2011), Gonzalez
& Engel, 2012, Ascher et al., (2016) and the Discoverlife Bee Species Guide and World Checklist (Ascher and
Pickering, 2019).
Result & Discussion
This checklist recorded 30 species of Megachile in Thailand through specimens examination and literature
review. For generic and subgeneric classification we follow Michener (2007) and Trunz et al. (2016), whereas for species
level identification, the specimens were verified with their original descriptions. In addition, notes on floral records,
diagnostic character, and nesting habitats are provided below. Species and collecting localities names are alphabetically
ordered.
Family: Megachilidae
Genus: Megachile Latreille, 1802
Subgenus: Aethomegachile Engel and Baker, 2006
Megachile (Aethomegachile) conjuncta Smith, 1853
Distribution: Chiang Mai.
Note: There is a sexual dimorphism between male and female bees.
Megachile (Aethomegachile) laticeps Smith, 1853
Distribution: Bangkok, Surat Thani.
Note: Superficially resembling to M. (Callomegachile) umbripennis but it can be easily
distinguished by mandible with an incomplete cutting edge in the second interspace and
hidden cutting edge in the third interspace.
Megachile (Aethomegachile) siamensis Cockerell, 1927
Distribution: Chiang Mai, Mae Hong Son.
Note: Only male was described.
Megachile (Aethomegachile) trichorhytisma Engel, 2006

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 95
Distribution: Chiang Rai, Loei
Note: Male can be easily recognized by the modification of its last flagellomere.
Subgenus: Amegachile Friese, 1909
Megachile (Amegachile) bicolor (Fabricius, 1781)
Distribution: Chiang Mai, Kanchanaburi, Mukdahan, Nakhon Nayok, Nakhon Pathom,
Nong Khai, Phayao, Ratchaburi, Saraburi, Tak.
Note: A widespread species in Oriental region, particularly India and southern China. This
species is commonly found in sunhemp field (Crotalaria juncea L.) and its nesting biology
was reported back from 1955 by Von Rudolf Altevogt.
Subgenus: Callomegachile Michener, 1962
Megachile (Callomegachile) atratiformis (Meade-Waldo, 1914)
Distribution: Uthai Thani.
Note: Engel and Gonzalez (2011) reported the species was collected on Dipterocarpus
obtusifolius Teijsman & Miquel.
Megachile (Callomegachile) disjuncta (Fabricius, 1781)
Distribution: Bangkok, Chiang Mai, Phayao, Surat Thani, Yala.
Note: This species forages on numerous types of plants, especially in Fabaceae. In addition,
Soh (2014) was first to report its nest construction using resins.
Megachile (Callomegachile) faceta (Bingham, 1897)
Distribution: No specific location provided.
Megachile (Callomegachile) facetula (Cockerell, 1918)
Distribution: No specific location provided.
Note: This species is superficially similar to M. (Callomegachile) faceta though can be
distinguished by the presence of dense fulvous hairs on the vertex of the former species.
Megachile (Callomegachile) fulvipennis (Smith, 1879)
Distribution: Surat Thani.
Note: Soh (2014) reported the species using resin for nest construction and visited
Peltophorum pterocarpum (Fabaceae) and Memecylon sp. (Melastomataceae).
Megachile (Callomegachile) impressa (Friese, 1903)
Distribution: No specific location provided.
Note: Only male was described.
Megachile (Callomegachile) monticola (Smith, 1858)
Distribution: No specific location provided
Note: This species has a wide distribution in south China.
Megachile (Callomegachile) odontophora Engel, 2011
Distribution: Nakhon Ratchasima
Note: Engel and Gonzalez (2011) reported the species was collected on Sindora siamensis
Teijsman & Miquel (Fabaceae).
Megachile (Callomegachile) ornata (Smith, 1853)
Distribution: Chiang Mai
Note: Tadauchi and Tasen (2009) identified only one specimen from a teak plantation.
Megachile (Callomegachile) tuberculata (Smith, 1858)
Distribution: No specific location provided
Note: Ascher et al. (2016) reported the species was collected on Grammatophyllum
speciosum Blume (Orchidaceae).
Megachile (Callomegachile) umbripennis (Smith, 1853)
Distribution: Bangkok, Phayao.
Note: This species has a wide distribution range in Southeast Asia and was reported to be
adventive in the US.
Subgenus: Carinula Michener, McGinley, and Danforth, 1944
Megachile (Carinula) fruhstorferi Friese, 1903
Distribution: Saraburi.
Note: This species is superficially similar to M. (Callomegachile) stulta though can be easily
distinguished by the presence of yellow felted hairs on the metasomal terga.
Subgenus: Chelostomoda Michener, 1962
Megachile (Chelostomoda) aureocincta Cockerell, 1927
Distribution: Nakhon Si Thammarat.
Megachile (Chelostomoda) nematocera Cockerell, 1929
Distribution: Chiang Mai.

Proceeding of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 96
Subgenus: Creightonella Cockerell, 1908
Megachile (Creightonella) fraterna Smith, 1853
Distribution: Ayutthaya, Chiang Mai, Kanchanaburi, Petchabun, Phayao, Ratchaburi,
Saraburi, Thonburi.
Note: This species usually forages on sunhemp flower (Crotalaria juncea L.) and constructs
its nest underground in the sun hemp field.
Megachile (Creightonella) atrata Smith, 1853
Distribution: Chumphon, Surat Thani, Yala.
Note: This species only occur in south Thailand. The bee forages on sunhemp flower
(Crotalaria juncea L.) and constructs its nests underground in the sun hemp field.
Subgenus: Eutricharaea Thomson, 1872
Megachile (Eutricharaea) gathela Cameron, 1908
Distribution: No specific location provided.
Megachile (Eutricharaea) griseopicta Radoszkowski, 1882
Distribution: Chiang Mai.
Note: This species is superficially similar to M. (Callomegachile) faceta though can be
easily distinguished by the presence of the hidden cutting in the third interspace of the
mandible.
Megachile (Eutricharaea) hera Bingham, 1897
Distribution: Kamphaeng Phet, Mae Hong Son, Nakhon Pathom, Petchabun, Phetchaburi,
Phang Nga, Phayao, Sakon Nakhon, Samut Sakorn, Ubon Ratchathani, Udon Thani.
Note: This species has a wide distribution and commonly found in Thailand.
Subgenus: Lophanthedon Gonzalez and Engel, 2011
Megachile (Lophanthedon) amputata Smith, 1857
Distribution: No specific location provided.
Note: This species is superficially similar to M. (Aethomegachile) laticeps though can be
easily distinguished by the presence of the outer ridge on the mandible.
Megachile (Lophanthedon) dimidiata Smith, 1853
Distribution: Lumpang.
Note: This species is superficially similar to M. (Pseudomegachile) velutina though can be
easily distinguished by the presence of the outer ridge on the mandible and absence of
dense long hairs on the inner adductor interspace of the mandible.
Megachile (Lophanthedon) erythropoda Cameron, 1901
Distribution: Songkhla.
Megachile (Lophanthedon) ferruginea Friese, 1903
Distribution: Nong Bua Lam Phu.
Note: This species is superficially similar to M. (Lophanthedon) dimidiata though can be
easily distinguished by the presence of fulvous hairs on the apical margin of the metasomal
terga.
Subgenus: Pseudomegachile Friese, 1898
Megachile (Pseudomegachile) lanata (Fabricius, 1775)
Distribution: No specific location provided.
Note: A common species found in India.
Megachile (Pseudomegachile) velutina Smith, 1853
Distribution: Lumpang, Nakhon Si Thammarat, Pattani.
Note: Commonly found in sunhemp field (Crotalaria juncea L.).
This report listed 30 Megachile species recorded in Thailand, which consist of 18 species of mason and
resin bees from 4 subgenera: Callomegachile, Carinula, Lophanthedon, and Pseudomegachile. Leafcutter bees
are represented for 12 species from 5 subgenera: Aethomegachile, Amegachile, Chelostomoda, Creightonella, and
Eutricharaea. Callomegachile has the highest number of species for Thai megachilid bees, whereas others are
infrequently reported. Despite Megachile wide distribution in Thailand, historical records are sporadic and rare.
Here, we documented 27 provinces in Thailand for the occurrence of Megachile. Of these 8 provinces are new
records for Thailand: Ayutthaya, Chumphon, Kanchanaburi, Mukdahan, Nakhon Pathom, Nong Khai, Petchabun,
and Thonburi.
Remarkably, three of the most abundance in agricultural areas, the leafcutter bees Megachile (Amegachile)
bicolor (Figure 1a), Megachile (Creightonella) fraterna (Figure 2a) and mason bee Megachile (Pseudomegachile)
velutina (Figure 2e), live and nest in sunhemp field. According to above, we suggest that these bees are potential
candidates for future pollination study. Another candidate is Megachile (Callomegachile) disjuncta (Figure 1e),
usually foraging on fabaceae and easy adoption human artifacts.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 97
Two close species of Thai Megachile, which have fulvous hairs on prosoma and mesosoma with the yellow
wings: the female of Megachile (Lophanthedon) dimidiata and Megachile (Pseudomegachile) velutina. These bees
have similar appearance, but differ in outer ridge on mandible, which are reaching to mandible base and reaching
only three-fourth mandible respectively. Female of Megachile (Aethomegachile) conjuncta and Megachile
(Callomegachile) disjuncta, which are black with white hairs on propodeum and first metasomal tergite, closely
resemble and they are sympatric species, but differ in scopal color, orange-red and black respectively. The male
of Megachile (Aethomegachile) conjuncta and Megachile (Aethomegachile) laticeps, black covered with fulvous
hairs, are very similar but differ in apical of gonostylus. Anonther interesting case is the Müllerian mimicry that
occurs to Megachile (Callomegachile) fulvipennis and Megachile (Callomegachile) tuberculata (Ascher et al., 2016).
Of the 5 species of Thai Megachile, include, Megachile (Aethomegachile) siamensis, Megachile (Aethomegachile)
trichorhytisma, Megachile (Callomegachile) monticola, Megachile (Callomegachile) tuberculata and Megachile
(Lophanthedon) dimidiate, are collected since past decades and there is no any recent appearance. For this reason,
we suggest that these bees may be extinct from Thailand. Nevertheless, these list may be not completely verifiable
because unclear distribution pattern, poor records and observations are difficult to determine and remain questionable.
In addition, anthropogenic activities such as deforestation, expansion of urban areas, and irresponsible tourism
are cause of habitat destruction and reduction of pollinators (Kjøhl et al., 2011).
Conclusion
30 species of Thai Megachile are recorded in 27 provinces. Here, we note some informations of Thai Megachile
such as diagnostic characters, food plants or nesting biology. Due to their high abundance and commonly found
in crop fields in Thailand, we suggest that M. (Amegachile) bicolor, M. (Callomegachile) disjuncta, M. (Creightonella)
fraterna, and M. (Pseudomegachile) velutina may be potential candidates for future pollination studies in Thailand
for food crops. In Addition, threats to the decline of Megachile species in Thailand should be of great concern since
they can be affected through anthropogenic activities.
Acknowledgement
The authors would like to thank the following institutions and persons as follow: Chawakorn Kunsete,
Pakorn Nalinrachatakan, Patsavee Utaipanon, Puttita Pasukdee, Sirat Lertjintanakit and Varat Sivayyapram, CU
Bee and Spider Research Lab, Chulalongkorn University, Thailand. We would like to express their gratitude towards
to The Scholarship from the Graduate School, Chulalongkorn University to commemorate the 72nd anniversary of
his Majesty King Bhumibol Adulyadej is gratefully acknowledged and The 90th Anniversary Chulalongkorn University
Fund (Ratchadaphiseksomphot Endowment Fund) (GCUGR1125604070M).
Referenes
Altevogt, R. 1955. Zur Technik der Blattschneidebienen Megachile bicolor F. und M. maritime Schck. Beitrage
zur Entomologie 5: 152–165.
Ascher, J. S. and Pickering, J. 2019. Discover Life bee species guide and world checklist (Hymenoptera:
Apoidea: Anthophila). [Online]. Available from:
http://www.discoverlife.org/mp/20q?guide=Apoidea_species [20 February 2019].
Ascher, J. S., Risch, S., Soh, Z. W. W., Lee J. X. Q., and Soh, E. J. Y. 2016. Megachile leaf-cutter and resin bees
of Singapore (Hymenoptera: Apoidea: Megachilidae). Raffles bulletin of zoology Supplement 32: 33-55.
Bingham, C. T. 1897. The fauna of British India, including Ceylon and Burma. Hymenoptera. Vol. I. wasps and
bees, Taylor & Francis, London, xxix+579 pp.
Cameron, P. 1908. A contribution to the Aculeate Hymenoptera of the Bombay presidency. The journal of the
Bombay Natural History Society 18: 649–659.
Cockerell, T. D. A. 1918. Descriptions and Records of Bees-LXXX. The annals and magazine of natural history,
including zoology, botany, and geology 2: 380–394.
Cockerell, T. D. A. 1927. Some bees, principally from Formosa and China. American Museum Novitates 274: 1–16.
Delaplane, K. S., and Mayer, D. F. 2000. Crop Pollination by Bees. CABI Publishing, New York, 322 pp.
Engel, M. S. and Gonzalez, V. H. 2011. Alocanthedon, a new subgenus of Chalicodoma from Southeast Asia
(Hymenoptera, Megachilidae). ZooKeys 101: 51–80.
Fabricius, J. C. 1775. Systema Entomologiae, Sistens Insectorum Classes, Ordines, Genera, Species, Adiectis
Synonymis, Locis, Descriptionibus, Observationibus, Flensburgi et Lipsiae, Korte, xxviii + 832 pp.
Fabricius, J. C. 1781. Species Insectorvm Exhibentes eorvn differentias specificas, synonyma avctorvm, loca
natalia, metamorphosin adiectis observationibvs, descriptionibvs, Hamburgi et Kilonii:Impensis Carol.
Ernest, Bohnii, viii+552 pp.

Proceeding of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 98
Friese, J. H. 1903. Neue Megachile-Arten des Sunda-Archipels. Zeitschrift für systematische Hymenopterologie
and Dipterologie 3: 349–359.
Gonzalez, V. H., and Engel, M. S., 2012. African and Southeast Asian Chalicodoma (Hymenoptera: Megachilidae):
new subgenus, new species, and notes on the composition of Pseudomegachile and Largella. Annales
Zoologici 62: 599–617.
Gonzalez, V. H., Griswold, T., Praz C. J., and Danforth, B. 2012. Phylogeny of the bee family Megachilidae
(Hymenoptera: Apoidea) based on adult morphology. Systematic Entomology 37: 261–286.
Kjøhl, M., Nielsen, A. and Stenseth, N. C. 2011. Potential effects of climate change on crop pollination. FAO,
Rome, 38 pp.
Meade-Waldo, M. A. G. 1 9 1 4 . Note on the Hymenoptera in the Collection of the British Museum, with
Descriptions of new Species. The annals and magazine of natural history, including zoology, botany,
and geology 14: 450–464.
Michener, C. D. 2007. The Bees of the World [2nd Edition]. Johns Hopkins University Press, Baltimore, MD,
xvi+[i]+953 pp.
Radoszkowski, O. 1882. Opisanie nowych gatunków błonkoskrzydłych (Hymenoptera). Wiadomości z Nauk
Przyrodzonych (Warszawa) 2: 72–81.
Smith, F. 1853. Catalogue of Hymenopterous insects in the collection of the British Museum. Taylor and
Francis, London, [i]+197 pp.
Smith, F. 1858. Catalogue of the Hymenopterous Insects collected at Sarawak, Borneo; Mount Ophir,
Malacca; and at Singapore, by A. R. Wallace. Journal of the proceeding of the Linnean Society 2: 42–52.
Smith, F. 1879. Descriptions of new species of Hymenoptera in the collection of the British Museum. Taylor
and Francis, London, xxi+240 pp.
Soh, E. J. Y. 2014. Nesting of two species of resin bees (genus Megachile) in a wooden bench. Singapore Biodiversity
Records 2014: 280–282.
Tadauchi, O., and Tasen, W. 2009. Bees of natural forests, teak plantations and agricultural fields in Thailand.
Esakia 49: 7–13.
Trunz, V., Packer, L., Vieu, J., Arrigo, N., and Praz, C. J. 2016. Comprehensive phylogeny, biogeography and
new classification of the diverse bee tribe Megachilini: Can we use DNA barcodes in phylogenies of
large genera?. Molecular Phylogenetics and Evolution 103(2016): 245–259.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 99
Figure 1. Pictures of candidates for future pollination studies (a–d) Megachile (Amegachile) bicolor (a) dorsal
view (b) lateral view (c) frontal view of mandible (d) lateral view of scopa (e–h) Megachile (Callomegachile)
disjuncta (e) dorsal view (f) lateral view (g) frontal view of mandible (h) lateral view of scopa.

Proceeding of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 100
Figure 2. Pictures of candidates for future pollination studies (a–d) Megachile (Creightonella) fraterna (a) dorsal
view (b) lateral view (c) frontal view of mandible (d) lateral view of scopa (e–h) Megachile (Pseudomegachile)
velutina (e) dorsal view (f) lateral view (g) frontal view of mandible (h) lateral view of scopa.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 101
Table 1. Examined specimens.
Subgenus sex
locality female male
Aethomegachile 55 43 Bangkok, Chiang Mai, Chiang Rai, Loei, Mae Hong Son,
Surat Thani
Amegachile 72 20 Chiang Mai, Kanchanaburi, Mukdahan, Nakhon Nayok,
Nakhon Pathom, Nong Khai, Phayao, Ratchaburi,
Saraburi, Tak
Callomegachile 120 62 Bangkok, Chiang Mai, Nakhon Ratchasima, Phayao,
Surat Thani, Uthai Thani, Yala
Creightonella 56 18 Ayutthaya, Chiang Mai, Chumphon Kanchanaburi,
Petchabun, Phayao, Ratchaburi, Saraburi, Thonburi.
Eutricharaea 24 70 Chiang Mai, Kamphaeng Phet, Mae Hong Son, Nakhon
Pathom, Petchabun, Phetchaburi, Phang Nga, Phayao,
Sakon Nakhon, Samut Sakorn, Ubon Ratchathani, Udon
Thani.
Lophanthedon 1 - Lumpang
Pseudomegachile 40 7 Lumpang, Nakhon Si Thammarat, Pattani

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 102
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 102 - 115
Diversity of Caterpillars (Order Lepidoptera) in KhaoYai National Park,
Nakhon Ratchasima Province
Paradorn Dokchan1,2*, Nanthasak Pinkaew1, Sunisa Sanguansub1 and Sravut Klorvuttimontara3 1Department of Entomology, Faculty of Agriculture at Kamphaeng Saen, Kasetsart University
KamphaengSaen Campus, Kamphaeng Saen Dictrict, Nakhon Pathom, Thailand 2Environmental Entomology Research and Development Centre, Faculty of Agriculture at KamphaengSaen,
Kasetsart University KamphaengSaen Campus, KamphaengSaen District, Nakhon Pathom, Thailand 3Faculty of Liberal Arts and Science, Kasetsart University Kamphaeng Saen Campus, Kamphaeng Saen
District, Nakhon Pathom, Thailand *Corresponding author e-mail:[email protected]
Abstract: The study of caterpillars diversity was started by sampled caterpillars from 500 meters line transect every
100 meters above mean sea level from 100 meters above mean sea level thru 1,200 meters above sea level in
KhaoYai National Park. Caterpillars were sampled every month from January 2017 – June 2017. A total of 3,434 specimens
were identified to 86 species, 55 genera, and 19 families and 37 morphospecies. The most abundant species was
Euremablanda (n=1,280). The highest diversity was found in 500 meters above mean sea level (H'= 2.66) and the similarity
of caterpillar that occurred in different elevation was low. Keywords: caterpillars, elevation, diversity, KhaoYai National Park.
Introduction
Khao Yai National Park is a Thailand's first national park, it is the third largest national park in Thailand.
Situated mainly in Nakhon Ratchasima Province. Khao Yai is part of Dong Phayayen-Khao Yai Forest Complex,
a world heritage site declared by UNESCO. In at least five different forest type, Khao Yai National Park has complex
ecosystem with richness of plant and animal such as mammal bird reptile and insects. Butterflies and Moths are
familiar insects with people. There are recorded of 214 butterflies species in Dong Phayayen-Khao Yai Forest Complex
include Khao Yai National Park (Lapkratok and Suwanwaree, 2010). Considering their abundance, moths and
butterflies caterpillars are not well known. It’s just a few records of caterpillars and foodplant in Khao Yai National Park.
The main objectives of the research were to determine the species of caterpillars and food plants in Khao Yai
National Park and to investigate the caterpillar diversity in different seasons and effect by biotic factor and abiotic factor.
Materials and Methods
Caterpillars collecting and rearing
The line transects walk modified from Pollard (1977). In different elevation, total 12 line transect are setting
in every 100 meters above mean sea level (=MMSL) from 100 – 1,200 MMSL. Every line transects are 500 meters length
and 5 meters width each both side along the transect. The transect walked is starting monthly from January – June 2017.
Caterpillars are sampled from every line transect by visual searching from ground surface up to 2 meters
on the host plant. The individual caterpillars are put in the plastic box. The caterpillar food plants were cut and put in
the plastic bag. The food plants were used for feed caterpillar and another purpose was for plant identified.
After field surveys, caterpillars will be moved to a bigger container. Leaves are changed and clean the container
every day along the development from larva to pupa. After pupation, the pupa are transferred into other plastic boxes
separately for rearing until adult emerge.
Specimens preservation and identification
Specimens are pinned and preserved on the setting board. After that, the setting boards were moved to a
hot air oven with 50 o C for two weeks. The specimens are moved to the insect boxes with code number and
insect’s information label. Then all of the specimens were identified by All specimens are identified by Cerny and
Pinratana (2009), Ek-Amnuay (2012), Holloway (1983; 1985; 1986; 1987; 1988a; 1988b; 1989; 1993; 1996; 1997;
1999; 2003; 2005), Inoue et al. (1997), Khanal and Smith (1997), Kononenko and Pinratana (2005), Pinratana
(1981; 1983; 1985),Pinratana and Eliot (1992; 1996), Robinson et al. (1994),Schintlmeister and Pinratana (2007),Smith
(1997), Solovyev (2008) Tan and Khoon (2012) and Zolotuhin and Pinratana (2005)

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 103
Caterpillar diversity calculation
Caterpillar diversities are calculated by diversity indexes such as Shannon index, Simpson index, evenness
index and Similarity index as follows:
where H' = Shannon Index
pi = The proportion abundance of the i species
i = number of i species individual
ln = Natural logarithm
Simpson’s index
Simpson’s index = 1- pi2
where pi = The proportion abundance of the i species
i = number of i species individual
Shannon species evenness index
J' = H'/Hmax
where Hmax = ln S
J' = H'/ ln S
H' = Shannon Index
ln = Natural logarithm
S = Number of species at each site
Result & Discussion
Total 3,434 individuals were collected and divide by 19 Family, 55 genera, 86 species and 37 morphospecies of
caterpillars. The most diverse species were Noctuidae and Pyralidae (15 species). In each elevation, The highest
number of individual was found in 1,000 MMSL (n=1,280) (figure 1). The most abundance was found in 500 MMSL
(figure 1.) and the highest diversity was found in 500 meters above mean sea level (H'= 2.66) (table 1.) consist
with McCoy (1990) studied on The distribution of insects include butterflies and moths along elevational gradients.
He reported that the highest species richness does frequently occur among insect in mid-elevation. In 800, 900
and 1,000 MMSL, we found that elevations were very high number of individual but their Shannon index values
were quite low (H' = 0.842, 1.135 and 0.460 respectively) and considered with Shannon species evenness index
values were low too (J' = 0.311, 0.367 and 0.237 respectively) (table 1). The Shannon index and Shannon species
evenness index values can reveal that some dominat species can be found. The highest individual of caterpillar
species was Eurema blanda (figure). A total 1,280 individuals were found and 788 individual were found in 1000 MMSL.
The similarity of caterpillar that occurred in different elevation represented by similarity index value.
The similarity measures between each elevation and another elevation were quite low (similarity index > 0.5),
some were absolutely different (similarity index = 0). It was because of the hostplant that occurred in each
elevation were different (table 2.)
Conclusion
Khao Yai National Park has a richness and high diversity of caterpillar. A total of 3,434 individuals were
identified into 19 Family, 55 genera, 86 species and 37 morphospecies. The most diverse species were Noctuidae
and Pyralidae. The dominant species was Eurema blanda. The highest species diversity was found in 500 MMSL.
The similarity measures between each elevation and another elevation is quite low, some were absolutely different.
However, the study period was quite short for higher analysis. A longer term sampling and monitoring are needed
to continue and observe changes in diversity. The physical and biological factor that be influencing to diversity
of caterpillars in different area are the futher study.
Acknowledgement
We thank all our friends who involve with this research, especially Miss Sopita Muadsub for caterpillar
image edited. We also would like to thank all of Khao Yai National Park rangers and officer for kindly supported.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 104
Reference
Cerny, K. and A. Pinratana. 2009. Moths of Thailand Vol. Six Arctiidae. Brothers of Saint Gabriel inThailand, Bangkok.
Ek-Amnuay, P. 2012. Butterflies of Thailand. 2nd ed. Amarin Book, Bangkok.
Holloway, J.D. 1983. The Moths of Borneo, Part 4, Family Notodontidae. Malay. Nat. J. 37: 1-107.
1985. The Moths of Borneo, Part 14: Noctuidae: Euteliinae, Stictopterinae, Plusiinae, Pantheinae.
Malay. Nat. J. 38:157-317.
1986. The Moths of Borneo, Part 1: Key to Families: Families Cossidae, Metarbelidae, Ratardidae,
Dudgeoneidae, EpipyropidaeandLimacodidae. Malay. Nat. J. 40: 1-166.
1988. The Moths of Borneo, Part 8: Castniidae, Callidulidae, Drepanidae, Uraniidae. Malay. Nat. J.
52: 1-155.
1989. The Moths of Borneo, Part 12: Family Noctuidae, trifine Subfamilies: Noctuinae, Heliothinae,
Hadeninae, Acronictinae, Amphipyrinae, Agaristinae. Malay. Nat. J. 42: 57-226.
1993. The Moths of Borneo, Part 11: Family Geometridae, Subfamily Ennominae. Malay. Nat. J. 47:1–309.
1996. The Moths of Borneo, Part 9: Geometridae (Incl. Orthostixini) Oenochrominae,
Desmobathrinae, Geometrinae, Ennominae addenda. Malay. Nat. J. 49: 147-326.
1997. The Moths of Borneo, Part 10: Family Geometridae, Subfamilies Sterrhinae and Larentiinae.
Malay. Nat. J. 51: 1–242.
1999. The Moths of Borneo, Part 5: Lymantriidae. Malay. Nat. J. 53: 1–188.
Inoue, H.R., D. Kennett and I.J. Kitching. 1997. Moths of Thailand Vol. Two Sphingidae. Brothers of Saint
Gabriel in Thailand, Bangkok.
Khanal, B. and C. Smith. 1997. Butterflies of Kathmandu Valley, Nepal. Tecpress Books, Bangkok.
Kononenko, V S. and A. Pinratana. 2005. Moths of Thailand Vol. Three Noctuidae. Brothers of Saint Gabriel
in Thailand, Bangkok.
Lapkratok, S. and Suwanwaree, P. 2010. Butterflies species diversity comparision among some waterfalls in
Dong Phayayen-Khao Yai Forest Complex World Heritage, Thailand.in Proceedings of 2010
International Conference on Environmental Science and Development (CESD 2010) : 26-28 February
2010.Singapore.
McCoy, E. 1990. The Distribution of Insects along elevational gradients. Oikos, 58: 313-322.
Pinratana, A. 1981. Butterflies in Thailand Vol. Four Lycaenidae. Brothers of Saint Gabriel in Thailand, Bangkok.
1983. Butterflies in Thailand Vol. Two Pieridae and Amathusiidae. Brothers of Saint Gabriel in
Thailand, Bangkok.
1985. Butterflies in Thailand Vol. Five Hesperiidae. Brothers of Saint Gabriel in Thailand, Bangkok.
Pinratana, A. and J. N. Eliot. 1992. Butterflies in Thailand Vol. One Papilionidae and Danaidae. 3rd edition.
Brothers of Saint Gabriel in Thailand, Bangkok.
1996. Butterflies in Thailand Vol. Three Nymphalidae. 2nd eds. Brothers of Saint Gabriel in Thailand, Bangkok.
Robinson, G.S., K.R. Tuck and M. Shaffer. 1994. A Field Guide to the Smaller Moths of South-East Asia.
The Natural History Museum, London.
Schintlmeister, A. and A. Pinratana. 2007. Moths of Thailand Vol. Five Notodontidae. Brothers of Saint
Gabriel in Thailand, Bangkok.
Smith, C. 1997. Butterflies of Royal Chitwan National Park, Nepal. Tecpress Books, Bangkok.
Tan, H. and K.S. Khoon, 2012. Caterpillars of Singapore’s Butterflies. National Park Borad, Singapore.
Zolotuhin, V.V. and A. Pinratana. 2005. Moths of Thailand Vol. Four Lasiocampidae. Brothers of Saint
Gabriel in Thailand, Bangkok.
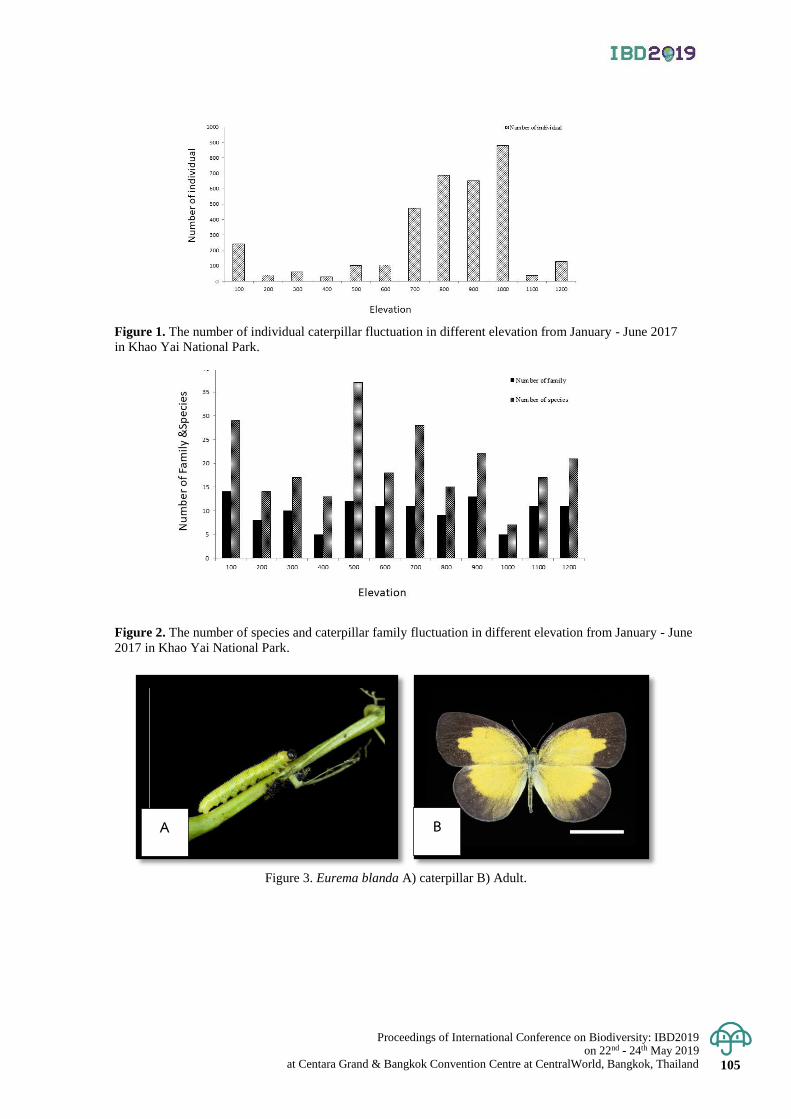
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 105
Figure 1. The number of individual caterpillar fluctuation in different elevation from January - June 2017
in Khao Yai National Park.
Figure 2. The number of species and caterpillar family fluctuation in different elevation from January - June
2017 in Khao Yai National Park.
Figure 3. Eurema blanda A) caterpillar B) Adult.
A B
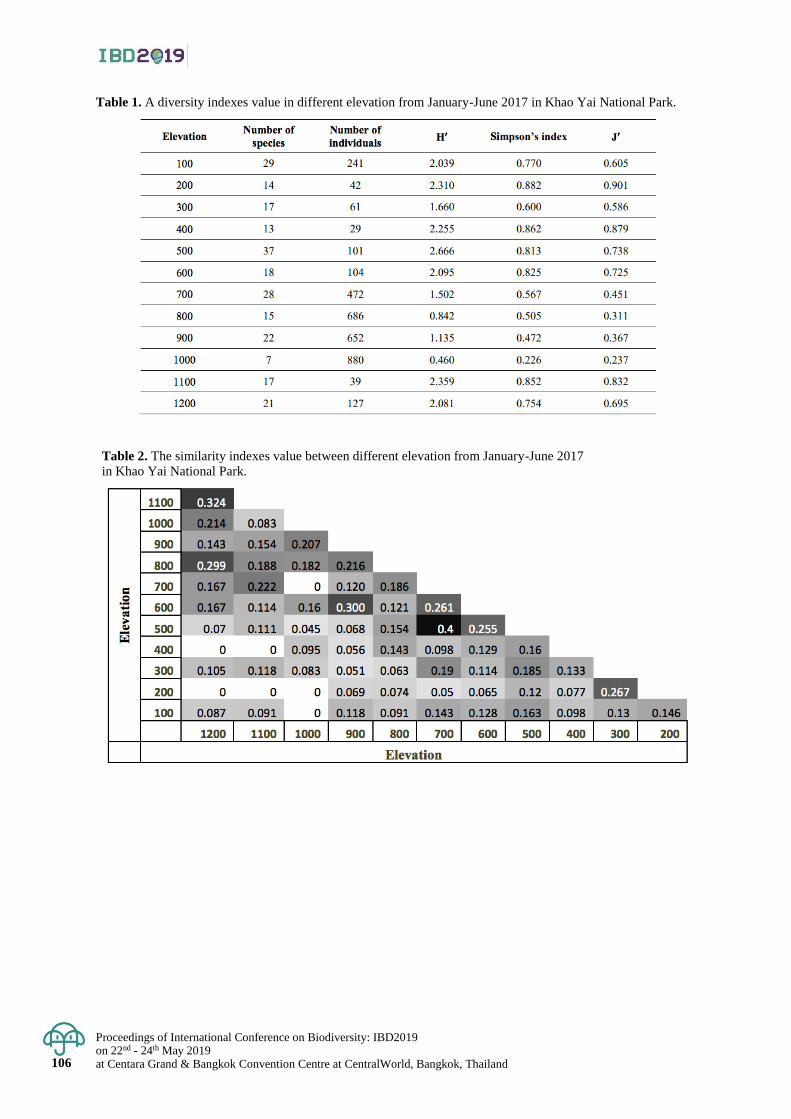
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 106
Table 1. A diversity indexes value in different elevation from January-June 2017 in Khao Yai National Park.
Table 2. The similarity indexes value between different elevation from January-June 2017
in Khao Yai National Park.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 107
APPENDIX
Appendix figure 1. The Buterfly caterpilars: F. Danaidae A (Tirumala septentrionis, F. Hesperiidae)
B (Bibasis sena) C (Burara gomata) D (Capitha purreea) E (Choaspes furcate) F (Hasora schoenherr)
G ( Notocrypta paralysos) H (Tagiades vajuna)
A
D
B
G
E
C
H
F

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 108
Appendix figure 2. The Buterfly caterpilars: F. Lycaenidae A (Arhopala abseus) B (Cheritra freja)
C (Flos sp.) D (Heliophorus epicles) E (Horaga sp.) F (Nacaduba sp.) G (Spindasis lohita)
H (Surendar quercetorum)
A
D
B
G
E
C
H
F
A B

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 109
Appendix figure 3. Butterfly caterpilars: F. Nymphalidae A (Athyma nefte) B (Charaxes bernardus)
C (Coelites nothis) D (Cyrestis thyodamas) E (Discophora sondaica) F (Faunis canens) G (Herona marathus)
H (Lebadea Martha)
D
G
E
C
H
F
A
D
B
C

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 110
Appendix figure 4. Butterfly caterpilars: F. Nymphalidae (cont.) A (Moduza procris) B (Neptis hylas)
C (Parasarpa dudu) D (Prothoe franck) E (Rohana parisatis) F (Tanaecia julii)
G (Tanaecia lepidea) H (Zeuxidia amethystus)
G
E
H
F
A
D
B
E
C
F

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 111
Appendix figure 5. Butterfly caterpilars: F. Papilionidae A (Graphium Agamemnon) B (Graphium antiphates)
C (Graphium doson) D (Graphium megarus) E (Lamproptera megas)
F (Losaria coon) G (Papilio Memnon) H (Papilio paris)
Appendix figure 6. Butterfly caterpilars: F. Pieridae A (Eurema hecabe) B (Eurema simulatrix)
G H
A B
A
D
B
C

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 112
Appendix figure 7. Moth caterpilars: F. Bombycidae A (Prismosticta tiretta F. Drepanidae) B (Drepana sp. F.
Gemetridae) C (Dyaphania militaris) D (Genusa simplex) E (Hyposidra sp.) F (Pingasa sp. F. Lasiocampidae)
G (Gastropacha sp.) H (Kunugia divaricate)
G
E
H
F
A
D
B
E
C
F

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 113
Appendix figure 8. Moth caterpilars: F. Lasiocampidae A (Kunugia sp.) B (Trabala vishnu) C (Trabala sp. F. Limacodidae)
D (Darna sp.) E (Parasa sinica) F (Setora sp. F. Lymantriidae) G (Calliteara horsfieldii) H (Orgyia postica)
Appendix figure 9. Moth caterpilars: F. Notodontodae A (Phalera grotei) B (Stauropus sp. F. Pyralidae)
C (Parotis marginata) D (Pygospila tyres F. Saturniidae) E (Antheraea assamensis) F (Attacus atlas)
G H
A
D
B
C
E F

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 114
Appendix figure 10. Moth caterpilars: F. Saturniidae A (Actias maenas) B (Cricula trifenestrata F. Sphingidae)
C (Acherontia Lachesis) D (Acosmeryx anceus) E (Cephonodes hylas) F (Daphnis hypothous) G (Meganoton
nyctiphanes) H (Neogurelca himachala)
A
D
B
G
E
C
H
F
A B

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 115
Appendix figure 11. Moth caterpilars: F. Sphingidae (cont.) A (Pergesa acteus) B (Theretra clotho)
C (Theretra oldenlandiae F. Thyrididae) D (Calindoea atripunctalis) E (Glanycus tricolor F. Zygaenidae)
F (Cyclosia panthona) G (Cyclosia papilionaria) H (Prosopandrophila distincta)
D
G
E
C
H
F

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 116
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 116 - 121
Proposal to reduce anthocyanin-deficient banana Musa siamensis to a M. rubra variety
Wandee Inta, Panida Kongsawadworakul, Unchera Viboonjun, Paweena Chuenwarin,
Paweena Traiperm and Sasivimon Chomchalow Swangpol*
Department of Plant Science, Faculty of Science, Mahidol University, Ratchathewi District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Musa rubra Wall. ex Kurz and M. siamensis Häkkinen & Rich. H. Wallace are small wild banana species
with erect inflorescences of orange and yellow bracts, respectively. While M. rubra is commonly found in North-East
India, Myanmar to Western Thailand, M. siamensis, described from a specimen collected in Eastern Thailand
bordering Cambodia, is known only in cultivation. Striking characters shared by both M. rubra and M. siamensis
include extended rhizome, shape and texture of inflorescence bracts, morphological characters of flowers and
smooth-surface seeds. Bract coloration caused by changes in cellular anthocyanin contents and revealed by expression
levels of genes involved e.g. dihydroflavonol 4-reductase (DFR) is the lone recognized difference between the two
taxa. It is, therefore, proposed here to reduce this yellow-bract taxon to M. rubra var. siamensis. These and other
bananas with a wide range of colorful bracts are useful as proper models for genetics, biochemistry, cytology and
biotechnology in flavonoid biosynthesis pathway. Keywords: Chek Meas banana, Leucoanthocyanidin, Musa laterita Cheesman, Rhodochlamys, Thai Gold banana
Introduction
Ornamental banana species possess attractive bract colors such as orange, pink, bright red, purple, and also,
though less frequently found, yellow and green. Among these colorful taxa, we have suspected that two species
in the Rhodoclamys sections, Musa rubra Wall. ex Kurz with orange bracts and M. siamensis Häkkinen & Rich.
H. Wallace with yellow ones, are conspecific.
The Musa genus has been classified into sections by Cheesman in 1947. Two out of his four sections,
Musa and Rhodoclamys, both having basic chromosome number x=11, are proved later by the genetic evidences
to be very closely allied (Cheesman 1947, Wong et al. 2002). They possess less glaucous, rarely or never polished,
non- or less overlapping inflorescence bracts, often dorsiventrally compressed, sometimes subglobose to irregular
angulate seeds, though the two sections are distinguished by different rachis directions; Musa is with hanging to
horizontal inflorescences and Rhodoclamys with erect ones.
Musa rubra was first discovered by Wallich in Burma (Myanmar) and grown in Calcutta Botanical Gardens
at least since as earlier as 1845 (Voigt, 1845; Hooker, 1895). It was later described and published in 1867 by Kurz
from specimens he collected by himself. Afterward, Cheesman (1949) separately described a banana grown from
seeds also collected from that country as M. laterita, however this name is recently declared as a synonym of M. rubra
by Joe et al (2016).
In 2007, Häkkinen and Wallace described M. siamensis from a specimen collected in 2002 in Eastern
Thailand. It has been called “Chek Meas”and used as ornamental plants by the Cambodians for a long time and
later commercialized by Thai nursery industry under the name “Thai Gold.” This species is closely related to M. rubra
by having similar itinerant rhizomes, smooth-surface seeds, and molecular and cytogenetic evidences (Häkkinen
and Wallace 2007; Čížková et al. 2015).
Color variation of banana male inflorescence bracts is of aesthetic value for ornamental purposes and
undoubtedly intended to attract pollinators. Among other factors, anthocyanin components and combinations are
stated as the cause of this diversity (Kitdamrongsont et al. 2008, Kongsawadworakul et al. 2016). Several anthocyanin
pigments, genes and biosynthetic pathways have been known to pay roles. Though it is rare for a single species
to contain genes encoding the entire spectrum of flower colors (Ahmed et al. 2014), bract colors of related Musaceae
species widely range from orange to pink and deep purple and could be useful in the study of anthocyanin biosynthesis.
Identification of anthocyanins by high performance liquid chromatography (HPLC) , mass spectrometry (MS) ,
and tandem mass spectrometry (MS/MS) revealed six major derivatives in Musaceae, i.e. delphinidin-3-rutinoside,
cyanidin-3-rutinoside, petunidin-3-rutinoside, pelargonidin-3-rutinoside, peonidin-3-rutinoside and malvidin-3-
rutinoside. It was found that bracts of M. rubra contain only non-methylated anthocyanin, i.e., delphinidin-3-rutinoside,
and cyanidin-3-rutinoside. On the other hand, in M. acuminata with yellow bracts and Ensete glaucum (Musaceae)
with green bracts, no anthocyanin could be detected. It was suspected that anthocyanin biosynthetic pathway in these
anthocyanin-deficient bananas may have been obstructed at leucoanthocyanidin- to-anthocyanidin step or prior
( Kitdamrongsan et al. 2008, Figure. 1) . One of the enzymes involving in this pathway is dihydroflavonol 4-

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 117
reductase (DFR) which catalyzes the production of flavan-3,4-diols ( leucoanthocyanidins) via the reduction of
colorless dihydroflavonols (Tian et al. 2017).
Apart from collected morphological data and investigation of floral morphological characters, we studied
expression levels of DFR from specimens of the two taxa with two different bract colors, one of which is suspected
to be a mutant of the other. The results of this study would clarify taxonomic statuses of these bananas and will
be beneficial in genetic resource selection for breedings and future studies on morphology, molecular biology and
phylogenetics. These bananas with colorful bracts would be proper models in research involving flavonoid
biosynthesis pathway and evolution of bract color variation in plants.
Materials and Methods
Two accessions of rare M. siamensis Häkkinen & Rich. H. Wallace from cultivations were investigated
along with three accessions of M. Rubra Wall. ex Kurz collected from two natural populations and a market.Three
accessions of M. Acuminate Colla subsp. siamea N.W. Simmonds and two accessions of M. ornata Roxb. were
also examined for comparison (Table 1). Morphological features were investigated and photographed in the fields.
Bracts were collected and stored in ice box and refrigerator before transferring to deep freezer and kept at -8 0 ⁰C
until used. Mature flowers from male inflorescences were preserved in 70% ethanol for morphological study.
Anthocyanin extraction and analysis
Frozen bract samples were ground to a fine powder with a mortar and pestle. The powdered bracts (0.2 g)
were extracted with 5 ml of methanol:HCL (99:1, v/v) at 4°C in darkness overnight. The extracts were centrifuged
at 10,000 g for 15 min at 4°C to precipitate the debris. The clear supernatant was transferred to a new tube, and the absorption
of the extracts at 530 and 657 nm was determined. Total anthocyanin content was calculated according to the
method of Mehrtens et al. (2005) using the following formula modification: QAnthocyanins = A530-0.25xA657,
where QAnthocyanins is the amount of anthocyanins and A530 and A657 are the absorption values at the indicated
wavelengths. The obtained data were the mean of two independent replicates.
Expression analysis by quantitative real-time PCR (qRT-PCR)
The qRT-PCR analysis was performed with the ABI-7500 real-time PCR machine (Applied Biosystem).
Twenty µl of PCR reactions containing 1 µl of 5-fold cDNA dilution, 0.4 µM of each primer, 0.2 mM dNTP mix,
2 mM MgCl2, 0.8 U of Platinum Taq DNA polymerase (Invitrogen) and 1000-fold dilution of SYBR green I (Sigma)
in 1X PCR buffer. The Ct (cycle threshold) data were determined using default threshold settings. The relative
expression was calculated as 2-∆Ct where ∆Ct = (Ct of candidate gene – Ct of internal control actin). The analyses
were performed in triplicate.
Result & Discussion
Musa acuminata is quite different from other species in this study. It has pendent rachis and fruits in two
rows and was previously grouped in the Musa section (Cheesman 1947) apart from the Rhodoclamys with erect type
of inflorescence and fruits in one row, which M. ornata, M. rubra and M. siamensis belong to. Besides inflorescence
bract colors, the latter three species are distinct in several aspects. Similar to most banana species in the Musa genus,
M. ornata has suckers and angular rough seeds. On the other hand, M. rubra and M. siamensis possess short
pseudostem of 1-1.5 m with extended rhizomes and subglobose seeds with smooth seed surface. These characters
are unique and rarely found among the members of the Musa genus. Though M. itinerans Cheesman also has long
travelling rhizome, its pseudostems are vigorous at 4-6 m tall. Moreover, with close investigation of floral morphology,
it was found that the flowers of M. rubra and M. siamensis having median inner tepal with acuminate apex and wing
apex as long as apex or nearly so (Figure 3C and D, arrow), are quite similar comparing to M. ornata which have
median inner tepal with attenuate apex and no wing apex present in neither M. ornata nor M. acuminata (Figure
3A and B). These distinct characters were compiled in key to species as stated below.
Key to Species based on Morphology
Herbs small or large; perennial, monocarpic, monoecious. Underground stem a rhizome or corm, commonly
suckering. Pseudostems clump, erect, formed by closely clasping leaf sheaths. Leaves spirally arranged, leaf blade oblong
to lanceolate. Inflorescence erect to pendulous. Bracts spirally arranged, usually with bright color. Flowers basal
male sterile, terminal female sterile; perianth in 2 whorls with 3 outer tepals and 2 inner ones united into a compound
tepal, 5-lobed; adaxial inner tepal free. Stamens 5; Pistil 1; ovary inferior, 2 rows of ovules in each loculus. Stigma 3-lobed
to clavate, or capitate. Fruit a fleshy berry. Seed surface smooth or rough.
1. Pseudostems less than 1.5 m high, inflorescence erect, fruits in one row.
2. Rhizomatous, male bract orange red or yellow, median inner tepal apex
acuminate, seed surface smooth…………………..………………………….. M. rubra, M. siamensis

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 118
2. Suckering, male bract pink, median inner tepal apex attenuate, seed surface
rough…………………….…………………………………………………... M. ornata
1. Pseudostems more than 1.5 m high, inflorescence pendent, fruits in two rows…… M. acuminata
Anthocyanin content The analysis revealed the correlation of total anthocyanin contents and color variation of male inflorescence
bracts. The total anthocyanin level was highest in the red purple bract and, in parallel, sample of the yellow bracts
yielded low anthocyanin contents (Figure 4A).
Expression analysis of DFR gene by qRT-PCR Preliminary real-time RT-PCR analysis showed that the anthocyanin biosynthetic gene, DFR, expressed
differently in the banana bracts of distinct colors, i.e. low or undetectable in M. siamensis (Figure 4B). This low
or undetectable level of DFR gene in M. siamensis is consistent with the absence of DFR gene expression in white
pomegranate (Zhao et al. , 2015) . This result suggested that DFR may be one of the main factors responsible for
anthocyanin accumulation in the bract of banana species.
Previous researches on molecular biology of wild bananas which included M. rubra and M. siamensis
stated that they are closely related. Čížková et al. (2015) reported that the two species share the same number of
45S rDNA and 5S rDNA loci and were grouped in the same cluster. Later, Christelová et al. (2017) constructed a
dendrogram based on the results of SSR analysis and revealed that M. ornata formed a distinct separate cluster
distantly positioned from the other Rhodochlamys entries which M. rubra and M. siamensis belong to. In addition,
information on distribution areas of the two species hints their position in the classification. The fact that M. siamensis
is rare and has never been found in natural habitats by the authors, while M. rubra distributes widely from North-
East India, Myanmar to Western Thailand indicates that the former could be an unusual mutant of the latter. From
these findings on morphology and molecular biology, we propose that M. rubra and M. siamensis are conspecific
and reduce taxonomic status of M. siamensis to M. rubra Wall. ex Kurz var. siamensis (Häkkinen & Rich. H. Wallace)
Swangpol & Inta.
Conclusion
Several characters including pseudostem heights, inflorescence positions, and number of fruit row, can
be used to distinguish M. acuminata subsp. siamea, M. ornata and M. rubra. On the other hand, M. rubra and M.
siamensis are quite similar and only difference is their inflorescence bract colors. The data obtained from this
study indicated the relation of this color modification and the expression of gene involved in anthocyanin
biosynthetic pathway. Further investigations are underway to study the expressions of the other structural genes
and its regulators including in different banana species. This information will ease the understanding of molecular
genetic background of the color differences in the banana bracts and further studies on co-evolutionary aspects
with pollinators.
Acknowledgement
The projects led by S.C.S. has been supported by Mahidol University and the Thailand Research Fund
(TRF) (RSA56_80033), Biodiversity-based Economy Development Office (Public Organization- BEDO) and the
Faculty of Science, Mahidol University, Bangkok, Thailand.
References
Ahmed NU, Park JI, Jung HJ, Yang TJ, Hur Y, Nou IS. 2014. Characterization of dihydroflavonol 4- reductase
(DFR) genes and their association with cold and freezing stress in Brassica rapa. Gene 550(1): 46-55.
Cheesman, E. E. 1947. Classification of the bananas III. The genus Musa L. Kew Bulletin 106-1 17.
Christelová, P., De Langhe, E., Hřibová, E, Čížková, J., Sardos, J., Husakova, M., Van den houwe, I., Sutanto,
A., Kay Kepler, A., Swennen, R., Roux, N., Dolezel, J. 2016. Molecular and Cytological Characterization
of the Global Musa Germplasm Collection Provides Insights into the Treasure of Banana Diversity.
Biodiversity and Conservation 26 (4): 801-824.
Čížková, J., Hřibová, E., Christelová, P., Van den Houwe, I., Häkkinen, M., Roux, N., Swennen, R. and Dolezel,
J. 2015. Molecular and Cytogenetic Characterization of Wild Musa Species. PLOS ONE 10(8): e0134096.
https://doi.org/10.1371/journal.pone.0134096
Häkkinen, M. and Wallace, R. H. 2007. Musa siamensis, a new Musa species (Musaceae) from S.E. Asia. Folia
Malaysiana 8:61-70.
Hooker, J.D. 1895. Musa rubra. In. Curtis’s Botanical Magazine. V. 121, London: L. Reeve & Co., England. Table 7451.
Joe, A. , Sreejith, P.E. and Sabu, M. 2016. Notes on Musa rubra Kurz (Musaceae) and reduction of M. laterita
Cheesman as conspecific. Taiwania 61: 34-40.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 119
Kitdamrongsont, K., Pothavorn, P., Swangpol, S., Wongniam, S., Atawongsa, K., Svasti, J. and Somana, J. 2008. Anthocyanin
composition of wild bananas in Thailand. Journal of Agricultural and Food Chemistry 56: 10853-10857.
Kongsawadworakul, P. , Viboonjun, U. , Chuenwarin, P. , Traiperm, P. , & Swangpol, S. C. 2016. Expression of
anthocyanin biosynthetic genes in banana bracts. In ThammaAbstracts of the First International Symposium
on Tropical and Subtropical Ornamentals (TSO2016). 7-9 March 2016, Krabi, Thailand, p. 44.
Liew, C.-F., Loh, C.-F., Goh, C.-J. and Lim, S.-H. 1998. The isolation, molecular characterization and expression of
dihydroflavonol 4-reductase cDNA in the orchid, Bromheadia finlaysoniana. Plant Science 135(2) : 161–169,
doi:10.1016/S0168-9452(98)00071-5.
Mehrtens, F., Kranz, H., Bednarek, P. & Weisshaar, B. 2005. The Arabidopsis transcription factor MYB12 is a
flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiology 138: 1083-1096.
Nu, A. , Park, J. I. , Jung, H. J. , Yang, T. J. , Hur, Y. and Nou, I. S. 2014. Characterization of dihydroflavonol 4-
reductase (DFR) genes and their association with cold and freezing stress in Brassica rapa. Gene 550(1): 46-55.
Quattrocchio, F. ; Verweij, W. ; Kroon, A. ; Spelt, C. ; Mol, J. ; Koes, R. 2006. PH4 of Petunia is an R2R3 MYB
Protein That Activates Vacuolar Acidification through Interactions with Basic- Helix- Loop- Helix
Transcription Factors of the Anthocyanin Pathway. Plant Cell 18: 1274–1291.
Tian, J., Chen, M. C., Zhang, J., Li, K.T., Song, T. T., Zhang, X., Yao, Y.C. 2017. Characteristics of dihydroflavonol
4-reductase gene promoters from different leaf colored Malus crabapple cultivars. Horticultural Research.
4:17070. doi: 10.1038/hortres.2017.70.
Voigt, J.O. 1845. Musaceae. In: Hortus Suburbanus Calcuttensis. Calcutta: Bishop’s College Press. pp. 578–579.
Wong, C. , Kiew, R. , Argent, G., Set, O., Lee, S. K., Gan, Y. Y. 2002. Assessment of the validity of the sections
of Musa (Musaceae) using AFLP. Annals of Botany 90: 231-238.
Zhao, X., Yuan, Z., Feng, L., & Fang, Y. 2015. Cloning and expression of anthocyanin biosynthetic genes in red and
white pomegranate. Journal of Plant Research 128: 687-696.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 120
Table 1. List of banana accessions in this study.
1 Cultivated clones purchased from a plant market. 2 Specimens grown as ornamental plants in public parks.
Figure 1. Simplified scheme of the flavonoid biosynthesis pathway modified from that of petunia (Quattrocchio, 2006).
Figure 2. Male inflorescence of Musa L. (Musaceae) showing different bract colors; (A) purple, M. acuminata
Colla subsp. siamea N. W. Simmonds, (B) pink, M. ornata Roxb., (C) orange, M. rubra Wall. ex. Kurz, and (D)
yellow, M. siamensis Häkkinen & Rich. H. Wallace. Photos not to scale.
No. Acc. No.
SS & JS Species Source/Origin
1 001 Musa acuminata Colla subsp. siamea N.W. Simmonds Phetchabun
2 247 Musa acuminata Colla subsp. siamea N.W. Simmonds Kanchanaburi
3 300 Musa acuminata Colla subsp. siamea N.W. Simmonds Nakhon Nayok
4 449 Musa ornata Roxb. Bangkok 1
5 542 Musa ornata Roxb. Chiang Mai 2
6 352 Musa rubra Wall. ex Kurz Mae Hong Son
7 420 Musa rubra Wall. ex Kurz Mae Hong Son
8 516 Musa rubra Wall. ex Kurz Bangkok 1
9 524 Musa siamensis Häkkinen & Rich.H.Wallace Bangkok 1
10 624 Musa siamensis Häkkinen & Rich.H.Wallace Bangkok 2
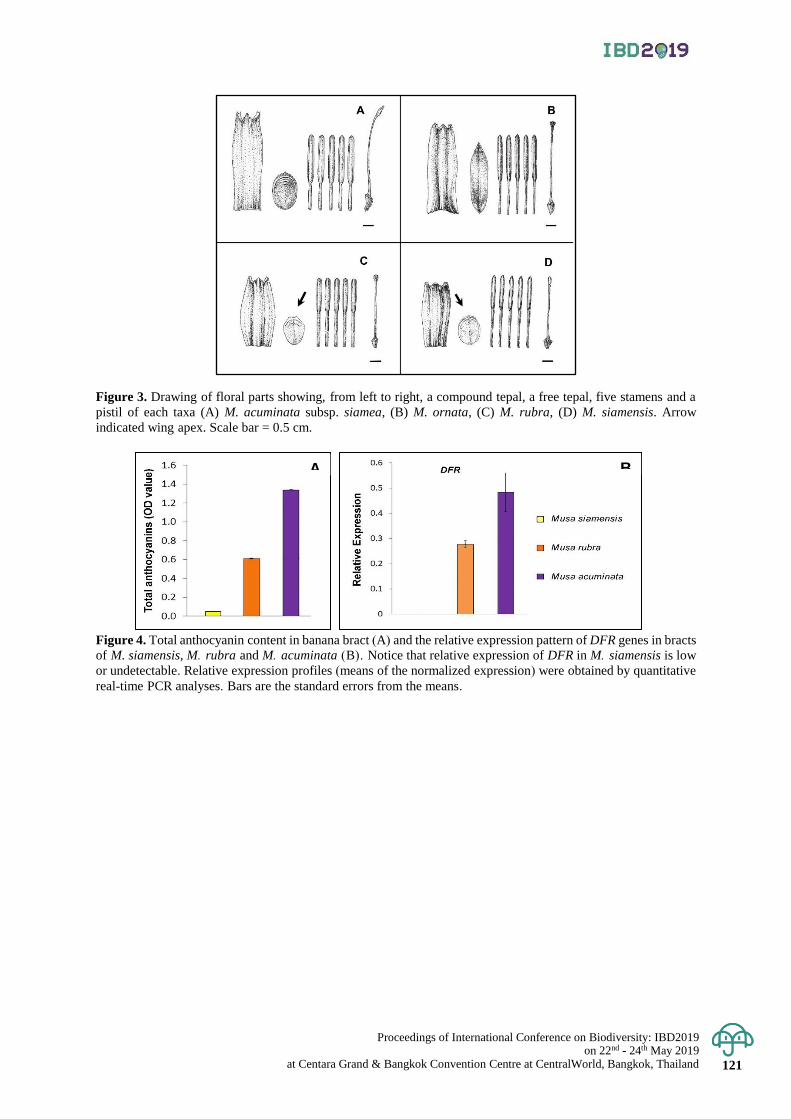
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 121
Figure 3. Drawing of floral parts showing, from left to right, a compound tepal, a free tepal, five stamens and a
pistil of each taxa (A) M. acuminata subsp. siamea, (B) M. ornata, (C) M. rubra, (D) M. siamensis. Arrow
indicated wing apex. Scale bar = 0.5 cm.
Figure 4. Total anthocyanin content in banana bract (A) and the relative expression pattern of DFR genes in bracts
of M. siamensis, M. rubra and M. acuminata (B). Notice that relative expression of DFR in M. siamensis is low
or undetectable. Relative expression profiles (means of the normalized expression) were obtained by quantitative
real-time PCR analyses. Bars are the standard errors from the means.
A B

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 122
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 122 - 128
Species richness and distribution of marine spider crabs (Majoidea) in Thailand
Kamonchanok Wongissarakul1*, Pattanee Jantrarotai2 and Puntip Wisespongpand3 1Thailand Natural History Museum, National Science Museum, Khlong Luang District, Pathum Thani, Thailand 2Department of Zoology, Faculty of Science, Kasetsart University, Chatuchak District, Bangkok, Thailand 3Department of Marine Science, Faculty of Fisheries, Kasetsart University, Chatuchak District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: The spider crabs are classified into Superfamily Majoidea. Species richness of spider crab in Thailand
were studied by collected specimens from local fishing boat along the coast of the Gulf of Thailand. Some specimens
were also examined from museums and institutes collection during September 2013 to December 2018. A total of 811
specimens were identified into 4 families, 21 genera and 40 species. Family Epialtidae were found the highest number
of spider crab species, comprising of 23 species (57.5%). The species richness of spider crab from the Andaman
Sea was higher than from the Gulf of Thailand at 35 and 19 species, respectively. There were two new records of Thailand:
Tiarinia angusta and Tylocarcinus styx. Furthermore, the results from this study indicated that the richness of marine
spider crab species was diversified due to habitat variation in marine ecosystem. Keywords: Majoidea, Marine environment, Species diversity, Spider crab, Thailand.
Introduction
Among the brachyuran superfamilies, the spider crabs (superfamily Majoidea) are one of the most diverse
group. Recently, the number of extant species of spider crabs comprised of over 800 species around the world (Ng
et al., 2008; Davie et al., 2015). Spider crabs distribute in all oceans especially in the Indo-West Pacific region.
Spider crabs are also called decorator crabs or masking crabs because they use various materials from their environment
such as pieces of algae, sponges, small pebbles and bits of shell decorate themselves with hook-setae for camouflage
and food reserve (Guinot et al., 1995; Guinot & Wicksten, 2015). Some spider crabs is popular for the live-seafood
trade such as snow crabs or queen crabs (Chionoecetes opilio) and Atlantic spider crabs (Maja brachydactyla)
(Ng, 1998; Marques et al., 2010). Most species of spider crabs play an important role as omnivorous, scavenger, consumer
and prey in marine ecosystem (Warner, 1977).
In the past, the identification key of spider crabs was complicated and confused because there were more than
ten subfamilies in family Majidae (Rathbun, 1925; Garth, 1958; Griffin & Tranter, 1986; Stevcic, 1994). Moreover,
there are widely different in morphological characters within family and subfamilies. Therefore, the genus of spider
crabs has been frequently transferred among families or subfamilies. As a result, it is a great practical problem
when using the identification keys. Later for clarifying the taxonomy of spider crabs, they were classified based
on their larval and more adult characters such as eyestalk and basal antenna characters. Finally, spider crabs are
classified in superfamily Majoidea with six families (Ng et al., 2008).
The previous studies of marine crabs in Thailand mostly focus on the commercial crabs such as Grapsid
crabs (Mutchacheep, 1973) and Portunid crabs (Naiyanetr, 1997 and Viruhpintu, 1989). For the spider crabs, there
are a few studies on spider crab diversity in Thailand with a great confusion in classification and numbers of species
because they are very diversified and the classification status have changed for a many time. The latest spider crabs
classification in Thailand (Naiyanetr, 2007), has been reported in only one family (16 genera, 32 species) which
is an outdating data and uncertainly about the species name. Therefore, the purpose of this study is to clarify and
updated classification and provide the distribution of marine spider crabs in Thailand.
Materials and Methods
Specimens collection
Specimens were collected from the gill nets, which fishing by local fishing boat of the fishery villages.
The sampling sites comprised of six stations along the Inner coast, Eastern coast and Upper coast of the Gulf of Thailand
(Figure 1, Table 1) during September 2013 to January 2015. All specimens were preserved in 70% ethanol and deposited
in the Crustacean Collection, Zoological Museum, Department of Zoology, Faculty of Science, Kasetsart University
(ZMKUcru).
The specimens in museum collection and institution collection were also examined in order to complete
overview of marine spider crabs in Thailand viz. follow; The Phuket Marine Biological Center (PMBC) where
specimens were collected in Andaman sea and from BIOSHELF project (Aungtonya et al, 2000), Marine crabs

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 123
laboratory in Marine Science Department, Faculty of Fisheries, Kasetsart University (MSCI), Thailand Natural
History Museum (THNHM) and Princess Maha Chakri Sirindhorn Natural History Museum (PSUZC).
Taxonomic identification
The spider crabs in this study were identified and described based on the mature specimens. The classification
of families and the status of species followed the checklist by Ng et al. (2008). The identification key to species were
followed Alcock (1895), Griffin (1974), Sakai (1938, 1965), Guinot & Forges (1985), Griffin & Tranter (1986),
Wagner (1986), Dai & Yang (1991), Poore (2004), Davie (2011) and Ng & Forges (2015).
Result & Discussion
Species richness of spider crabs
A total of 811 specimens of marine spider crab were identified in superfamily Majoidea with four families,
five subfamilies, 21 genera and 40 species, as present in Table 2. There were two species, Tiarinia angusta and
Tylocarcinus styx are supposed to new record in Thailand. From field collection, spider crabs were identified into
three families, six genera and 12 species; Schizophrys aspera, Prismatopus halimoides, P. aculeatus, Micippa
thalia, M. philyra, Hyastenus aries, H. hilgendorfi, H. diacanthus, Doclea armata, D. canalifera, D. rissoni and
Camposcia retusa. The spider crab species was consistent with many previous studies (Wisespongpand et al.,
2010: 2013: 2016; Kaewgunha & Tangkrock-olan, 2007; Singlaem et al., 2013 and Khanadee et al., 2014) because
we collected specimens from the same areas. However, the number of spider carb species in this study was higher than
the previous studies because we collected specimens from many areas and collected periods longer than those of studies.
The family with the highest number of spider crab species was Epialtidae which contained 23 species
(57.5% of total). Spider crabs in genus Doclea had the highest number of species which composed of six species.
We found two species, D. canalifera and D. canaliformis that have never been reported in the decapod checklist
of Thailand (Naiyanetr, 2007). In this study, D. canalifera was dominant species and its distribution was in the
Inner, Eastern and Upper coast of Gulf of Thailand, which was supported by several previous studies (Rathbun,
1910; Wagner, 1986; Griffin & Tranter, 1986; Dai & Yang, 1991). On the other hand, D. canaliformis was an
uncommon species and its distribution was only in the Gulf of Thailand (Wagner, 1986). In family Epialtidae, we
reclarified D. alcocki to D. muricata from the PMBC collection which previously were reported by Ng & Davie
(2002) and Naiyanetr (2007). D. ovis resembled to D. canaliformis in morphological characters. However, the character
of the first male gonopod and gonopore were practically used for species identification.
For the family Hymenosomatidae, we found only one genus and one species, E. magnum. This species
was known as the endemic species in the mangal area of Ranong province, Andaman Sea, which corresponded to
the report by Ng & Chuang (1996) and Naiyanetr (2007). Therefore, the management of this mangroves area is
seriously point for the long term protection and conservation of this species.
In this study, we did not found two specimens, Rochinia pulchra and Hoplophrys oatesii, which were
recorded by Promdam (2009) and Jaingam (2013) that these specimens were preserved in PMBC and MSCI,
respectively. Furthermore, we did not found Paratymolus pubescens and P. vannus (family Inachidae) which was
agreed with Singlaem et al (2013) and Khanadee et al. (2014) studies but differed from Rathbun (1910) who
recorded these two species from Chanthaburi coast, Eastern Gulf of Thailand. This may be because these two
species were restricly distributed in rocky bottom and coral reef (Griffin & Tranter 1986), where these habitats
were good refuge for spider crabs and it is the area that difficult to collect the specimens.
As result, there were the numbers of spider crab species more than Naiyanetr’s report in the decapod
checklist of Thailand (Naiyanetr, 2007). According to Naiyanetr (2007), he reported that there were 32 spider crab
species while this study found 40 species. The spider crab species that were not previously reported as follow;
Paramaja gibba, Tiarinia angusta, Micippa platipes, Tylocarcinus styx, Naxioides taurus, Hyastenus borradailei,
Phalangipus filiformis, Doclea canaliformis, D. canalifera, D. muricata, Huenia heraldica, Xenocarcinus
depressus, X. tuberculatus, X. conicus, Oncinopus araneus, Pleistacantha rubida, Platymaia alcocki and
Cystomaia suhmii. The increasing in number of spider crab species in this study was because we collected
specimens from several field areas especially in the Eastern Gulf of Thailand where there was less information
about spider crab species. Moreover, this study also reexamined spider crab specimens from several museum
collections. The MSCI, THNHM and PSUZC collections contain most of spider crab specimens collected from
various types of fishing gears throughtout the coast of the Gulf of Thailand and Andaman Sea, and including the
coral reefs. Those specimens were also collected by SCUBA diving, from the gill nets which vertically placed
near the coral reefs or rocky shores, and by the trawl nets, traps or crab dredges. On the other hand, the PMBC
collection contains most of the spider crab specimens came from the coast and the deep-water in Andaman Sea
by using traps (Aungtonya et al., 2000). Therefore, the specimens from this study came from different habitats
and from various types of fishing gears which lead to the discovery of many spider crab species including new
habitats and new localities of spider crab distribution.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 124
Geographic distribution
Within the geographic area of this study, the number of marine spider crab species that occurrence in
Andaman Sea was higher those of the Gulf of Thailand, 35 species found in the Andaman Sea and 19 species in
the Gulf of Thailand. Among these, there were 21 species only in Andaman Sea, 5 species only in Gulf of Thailand
and 14 species in both areas (Table 2.). As a result, the number of spider crab species in Andaman Sea had a considerably
more than the Gulf of Thailand due to their geographical differences (Jaingam et al., 2007). Therefore, Andaman
Sea should be promoted as Biodiversity Important Area (BIA) for further conservation and protection of marine
crab species in Thailand.
In this study, the intertidal zone had the highest number of spider crab species (22 species) due to the
diversity of habitats that suitable for spider crabs which was to many studies. For example, Doclea spp. lived on
muddy bottom of the nearshore (Wagner, 1986 and Dai & Yang, 1991), Phalangipus spp. lived on sandy to
muddy-sandy bottom of the offshore (Griffin, 1973). While Xenocarcinus depressus, X. tuberculatus, X. conicus,
Huenia heraldica and Menaethius monoceros lived in the coral reef by associated with the other marine organisms
such as sponges, gorgonians, sea whips, corallines, and algae (Griffin & Tranter, 1986; Jaingam, 2013). Moreover,
there were some spider crab species such as S. aspera, Prismatopus spp., Micippa spp. and Camposcia retusa
lived by decorating themselves with a piece of sponges, coral, algae or pebbles for camouflage, defence, shelter
and/or food supply (Jaingam et al., 2008; Wisespongpand et al., 2012). In addition, spider crabs can be used as an
indicator species of coral reef ecosystem such as Xenocarcinus spp and Huenia heraldica. Therefore, the revised
and updated classification of spider crabs in Thailand are important for the further study of spider crab diversity
due to they are species assemblages in most marine ecosystems, resulting in the complexity of the food chain and
finally lead to the stable of marine ecosystems.
Conclusion
The species richness of marine spider crab was found in the Andaman Sea higher than in the Gulf of
Thailand especially in the intertidal zone where there were the variety of habitat environments such as muddy
bottom, sandy-muddy bottom and coral reef. The highest number species was belonging to family Epialtidae by
contain 23 species. In this family, spider crab genus Doclea was highest species richness, which composed of six
species. There are two species, Tiarinia angusta and Tylocarcinus styx are supposed to be new records in Thailand.
The habitat of most spider crab species, are in marine environment except the Hymenosomatid crab (Brackish-
water environment). In order to fulfill information of the crab biodiversity in Thailand, we should be studying in
Brackish-water and freshwater areas. This study provides an updated classification of marine spider crabs in Thailand
for further spider crab identification, which it is importance for measuring the diversity of marine crabs in the future.
Acknowledgement
This study supported by Faculty of Science Kasetsart University Postgraduate Studentship (ScKUPGS)
and funding source for graduate students of Zoology Department, Faculty of Science, Kasetsart University. We
would like to thank Mr. Rueangrit Promdam of the Princess Maha Chakri Sirindhorn Natural History Museum for
lending materials and give important spider crab data for this study. We sincere thanks also goes to Prof. Peter Ng
Kee Lin of Lee Kong Chian Natural History Museum at Singapore, for his valuable suggestion and information
of the spider crab identification. We would like to express my gratitude to all curators of Reference Collection at
Phuket Marine Biological Center for provided me of this opportunity and for helping in laboratory facilities during
the examination under their care.
References
Alcock, A. 1895. Materials for a carcinological fauna of India, 1: The Brachyura Oxyrhyncha. Journal of the
Asiatic Society of Bengal 64: 157-291.
Aungtonya, C., Thaipal, S., and Tendal, O. 2000. A preliminary report on the Thai-Danish bioshelf surveys (1996–2000)
of the west coast of Thailand, Andaman sea. Research bulletin. Phuket Marine Biological Center 6: 53-76.
Dai, A., and Yang, S. 1991. Crabs of the China seas (English ed). China Ocean Press; Belin; New York: Springer-
Verlag, Beijing. 682 pp.
Davie, P. J. F. 2011. A new species and new records of Oncinopus (Crustacea: Brachyura: Inachidae) from
northern Australia. Records of the Museums and Art Galleries of the Northern Territory 27: 123-133.
Davie, P.J.F., Guinot, D. and Ng, P.K.L. 2015. Systematics and classification of Brachyura. In: Castro, P., Davie,
P.J.F., Guinot, D., Schram, F. and Von Vaupel Klein, C. (Eds.), Treatise on Zoology – Anatomy,
Taxonomy, Biology – The Crustacea, complementary to the volumes translated from the French of the
Traité de Zoologie, 9(C) (I), Decapoda: Brachyura (Part 2): 1049–1130.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 125
Garth, J. S. 1958. Brachyura of the Pacific coast of America. Oxyrhyncha. Allan Hancock Pacific Expeditions,
21, part 1: i–xii, 1–499; part 2: 677–854.
Guinot, D., and Forges, B. R. D. 1985. Crustacés Décapodes: Majidae (genres Platymaia, Cyrtomaia, Pleistacantha,
Sphenocarcinus et Naxioides). Mémoires du Muséum d'Histoire Naturelle.Série A. Zoologie, 133: 83-179.
Guinot, D., Doumenc, D., and Chintiroglou, C. C. 1995. A review of the carrying behaviour in brachyuran crabs,
with additional information on the symbioses with sea anemones. Raffles Bulletin of Zoology, 43: 377-416.
Guinot, D. and Wicksten, M.K. 2015. Camouflage: carrying behaviour, decoration behaviour, and other
modalities of concealment in Brachyura, pp. 583–638, fig. 1-11 (Chapter 71-11). In: Castro P., Davie
P.J.F., Guinot D., Schram F.R. and Von Vaupel Klein J.C. (eds.), Decapoda: Brachyura (Part 1), Treatise
on Zoology – Anatomy, Taxonomy, Biology, vol. 9C-I. Brill, Leiden and Boston.
Griffin, D. J. G. 1973. A revision of the spider crabs of the genus Phalangipus (Crustacea, Brachyura, Majidae).
Journal of Natural History 7: 165-207.
Griffin, D. J. G. 1974. Spider crabs (Crustacea: Brachyura: Majidae) from the International Indian Ocean
Expedition, 1963-1964. Smithsonian Contributions to Zoology: 1-35.
Griffin, D.J. and Tranter, H.A. 1986. The Decapoda Brachyura of the Siboga expedition. Part VIII: Majidae.
Siboga Expéditie 39(C4): 1–335, 22 plates.
Jaingam, W., Wisespongpand, P., and Thamrongnawasawat, T. 2007. Species Diversity of Crabs from Bottom
Gill Net Fisheries in Thailand, Proceedings of 45th Kasetsart University Annual Conference, Bangkok,
30 January-2nd - February 2007, Bangkok, Thailand p. 603-624.
Jaingam, W., Wisespongpand, P., and Thamrongnawasawat, T. 2008. Relationship between crabs and associated
organisms in coral reef at Mu Ko Surin National Park, Proceedings of 46th Kasetsart University Annual
Conference, Bangkok, 29 January-1nd February 2007, Bangkok, Thailand p. 492-502.
Jaingam, W. 2013. Species richness of crabs in coral reef, Mu Ko Surin National Park, Phangnga province. M. Sc
Thesis, Kasetsart University, Bangkok.
Kaewgunha, N. and Tangkrock-olan, N. 2007. Species of Marine crabs found at Ang-sila and Laem Cha-bang
Fishing Ports, Chonburi Province. Burapha Science Journal 12(1): 63-72.
Khanadee, U., Boonaum, K., Prommasuk. J., Kunsook, C. 2014. Species diversity of bycatch from the crab gill
net fishery in Kung Krabaen Bay, Chanthaburi Province, Proceeding of the 2nd PKRU conference,
Phuket, May 8th – 9th, 2014, Phuket, Thailand p. 307-319.
Mutchacheep, S. 1973. Grapsid crabs in the Gulf of Thailand. M. Sc Thesis, Chulalongkorn University, Bangkok.
Marques, A., Teixeira, B., Barrento, S., Anacleto, P., Carvalho, M. and Nunes, M. 2010. Chemical composition
of Atlantic spider crab Maja brachydactyla. Journal of Food Composition and Analysis, 23: 230-237.
Naiyanetr, P. 1997. Taxonomic study of marine crabs family Portunidae in the Gulf of Thailand. M. Sc Thesis,
Kasetsart University, Bangkok.
Naiyanetr, P. 2007. Checklist of crustacean fauna in Thailand (Decapoda and Stomatopoda). Office of environment
Policy and Planning. OEPP Biodiversity, 5: 161 pp.
Ng, P. K. L., and Chuang, C. T. N. 1996. The hymenosomatidae (crustacea: decapoda: brachyura) of Southeast
asia, with notes on other species. The Raffles Bulletin of Zoology, Supplement, 3: 1-82.
Ng, P. K. L. 1998. Crabs 1046-1156. In Carpenter, K.E. and V.H. Niem (eds). FAO species identification guide
for fishery purposes. The living marine resources of the Western Central Pacific. Vol. 2. Cephalopods,
crustaceans, holothurians and sharks. Rome, FAO. 687-1396.
Ng, P. K. L., and Davie, P. J. F. 2002. A checklist of the brachyuran crabs of phuket and western thailand. Phuket
marine biological center special publication, 23: 369-384.
Ng, P. K. L., Davie, P. J. F., and Guinot, D. 2008. Systema Brachyurorum: Part I. An annotated checklist of extant
brachyuran crabs of the world. The Raffles Bulletin of Zoology, 17: 1-286.
Ng, P. K. L., and Forges, B. R. D. 2015. Revision of the spider crab genus Maja Lamarck, 1801 (Crustacea:
Brachyura: Majoidea: Majidae), with descriptions of seven new genera and 17 new species from the
Atlantic and Indo-West Pacific. Raffles bulletin of Zoology, 63: 110-225.
Promdam, R. 2009. New records on Brachyuran crabs from the Thai-Danish BIODIVERSITY Project, in the
Andaman Sea, Thailand, Seminar of the Department of Marine and Coastal Resources Seminar,
Ratchaburi, August 23th -25th, 2009, Ratchaburi, Thailand (Poster presentation).
Poore, G. C. B. 2004. Marine Decapod Crustacea of Southern Australia: A Guide to Identification. Collingwood,
Victoria, Australia: CSIRO Pub., 617 pp.
Rathbun, M. J. 1910. The Danish Expedition to Siam 1899-1900. V. Brachyura. Konelige Danske Videnskabernes
Selskat, Naturvidenskabelige Matematiske Afhandlinger 5(4): 301–367.
Rathbun, M. J. 1925. The spider crabs of America. Bulletin of the United States National Museum. i–xx, 1-613, 153 figs, 283 pls.
Sakai, T. 1938. Studies on the crabs of Japan III. Brachygnatha, Oxyrhyncha. 193–364 pp.
Sakai, T. 1965. The crabs of Sagami Bay collected by His Majesty the Emperor of Japan. Tokyo, Maruzen Co.
Ltd. 206 pp., 100 plates.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 126
Singlaem, C., Somya, S., Surawut, S., Kunsook, C. 2013. Species diversity of marine crab in Kung krabaen bay,
Chanthaburi province, Proceedings of 6th Thai resources, Plant Genetic Conservation Project: 21-23
December 2013, Kanchanaburi, Thailand, 312-319.
Stevcic, Z., 1994. Contributions to the re-classification of the family Majidae. Periodicum Biologorum, 96(4): 419–420
Viruhpintu, S. 1989. Taxonomy of Portunid crabs in Thailand. M. Sc. Thesis, Chulalongkorn University, Bangkok.
Wagner, H. P. 1986. A revision of the genus Doclea Leach, 1815 (Crustacea, Brachyura, Majidae). Bulletin du
Muséum national d'histoire naturelle 8: 893-953.
Warner, G. F. 1977. The biology of crabs. New York: Van Nostrand Reinhold. 202 pp.
Wisespongpand, P., Jaingam, W., Poddamrong, A., and Srichomngam, W. 2010. Biodiversity of crabs on coastal
habitats of Prachuapkhirikhan province, Proceedings of the 48th Kasetsart University Annual Conference:
3-5 February 2010, Bangkok, Thailand, 435-446.
Wisespongpand, P., Aungkuldee, P., Thamrongnawasawat, T., and Khaodon, K. 2012. Distribution and life
history of rough spider crab (Schizophrys aspera) in coral reef at Mu Koh Lan, Cholburi, Proceedings
of the 50th Kasetsart University Annual Conference: 31 January-2 February 2012, Bangkok, Thailand, 525-533.
Wisespongpand, P., Sirivirintarat, T., Jaingam, W., Inbumrung, K., and Srichomngam, W. 2013. Diversity of
crabs from bottom gill net, Rayong province, Proceedings of 51st Kasetsart University Annual
Conference: 5-7 February 2013, Bangkok, Thailand, 422-429.
Wisespongpand, P., Wongissarakul, K., Jaingam, W., & Khaodon, K. 2016. Species Richness and Abundance of
Crabs from Bottom Gill Net in Cholburi Province, Proceedings of the 54th Kasetsart University Annual
Conference: 2-5 February 2016, Bangkok, Thailand., 938-946.
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 127
Figure 1. Map showing collection localities. (C = Chonburi; R = Rayong; CH = Chanthaburi; T = Trat;
P = Prachuapkhirikhan Province).
Table 1. Sampling sites for this study.
Sampling site Station Latitude Longitude
Region Province
Inner coast Chonburi North Pattaya 12º57’26N 100º53’02E
South Pattaya 12º55’30N 100º52’5E
Eastern coast Rayong Suan-son beach 12º37’58N 101º28’42E
Chanthaburi Chao-Lao beach 12º32’11N 101º55’43E
Trat Laem-Ngob 12º10’08N 102º23’55E
Upper coast Prachuapkhirikhan Bang-Kum 11º33’56N 99º39’43E
Laem-Kum beach 11º28’4N 99º37’20E
Table 2. Species richness and geographic distribution with habitat noted. (GT = Gulf of Thailand, AS = Andaman Sea).
Superfamily Majoidea (Samouelle, 1819) Geographic
distribution Habitat Noted
Family Subfamily Species GT AS
Hymenosomatidae Elamena magnum Ng & Chuang, 1996 - Mangal
Majidae Majinae Paramaja gibba (Alcock, 1895) - Deep-water
Schizophrys aspera (H. Milne Edwards, 1834) Coral reef
Prismatopus halimoides (Miers, 1879) Intertidal/Coral
reef
P. aculeatus (H. Milne Edwards, 1834) Intertidal/Coral
reef
P. longispinus (De Haan, 1839) - Offshore
Mithracinae Tiarinia angusta Dana, 1851 - Coral reef
Micippa thalia (Herbst, 1803) Intertidal/Coral
reef
M. platipes Rüppell, 1830 - Intertidal/Coral
reef
M. philyra (Herbst, 1803) - Intertidal/Coral
reef
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 128
Epialtidae Pisinae Tylocarcinus styx (Herbst, 1803) - Coral reef
Naxioides taurus (Pocock, 1890) - Offshore
Hyastenus planasius (Adams & White, 1848) - Intertidal
H. aries (Latreille, 1825) Intertidal/Coral
reef
H. borradailei (Rathbun, 1907) - Coral reef
H. hilgendorfi De Man, 1887 Intertidal/Coral
reef
H. diacanthus (De Haan, 1839) - Intertidal/Coral
reef
Phalangipus hystrix (Miers, 1886) - Offshore
P. indicus (Leach, 1815) - Offshore
P. filiformis Rathbun, 1916 Offshore
P. longipes (Linnaeus, 1758) - Offshore
Doclea armata De Haan, 1839 Intertidal
D. canaliformis Ow-Yang in Lovett, 1981 - Intertidal
D. canalifera Stimpson, 1857 - Intertidal
D. ovis (Fabricius, 1787) Intertidal
D. muricata (Herbst, 1788) - Intertidal
D. rissoni Leach, 1815 Intertidal
Tychinae Criocarcinus superciliosus (Linnaeus, 1758) - Intertidal/Coral
reef
Epialtinae Huenia heraldica (De Haan, 1837) - Coral reef
Menaethius monoceros (Latreille, 1825) Rocky
shore/Coral reef
Xenocarcinus depressus Miers, 1874 - Coral reef
X. tuberculatus White, 1847 - Coral reef
X. conicus (A. Milne-Edwards, 1865) - Coral reef
Inachidae Oncinopus aranea (De Haan, 1839) - Coral reef
Achaeus brevirostris (Haswell, 1879) Coral reef
A. lacertosus Stimpson, 1858 Coral reef
Camposcia retusa (Latreille, 1829) Coral reef
Pleistacantha rubida (Alcock, 1895) - Deep-water
Platymaia alcocki Rathbun, 1918 - Deep-water
Cystomaia suhmii Miers, 1885 - Deep-water

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 129
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 129 - 135
The effectiveness of basic bait traps for collecting adult flies
Amornrat Ninon and Taeng On Prommi* Faculty of Liberal Arts and Science, Kasetsart University, Kamphaeng Saen Campus, Kamphaeng Saen District,
Nakhon Pathom, Thailand *Corresponding author e-mail: [email protected]
Abstract: The aim of the present study is to compare the effectiveness of pork liver bait traps (PLBTs) and fish
meat bait traps (FMBTs) for collecting adult flies. Bait traps used to collect these species can be damaged by anthropogenic
or environmental effects. In this study, PLBTs and FMBTs were hung from trees in the three areas (tree near the pig
farm, tree near the dormitories and tree near the fresh market) of the Kasetsart University, Kamphaeng Saen Campus,
Nakhon Pathom Province, Thailand. This activity was carried out on the first Monday of each week for two months.
During the study, 4,252 specimens were collected. Of all these species, 2903 (68.27%) were obtained from PMBTs,
while 1,349 (31.73%) were obtained from FMBTs. Calliphoridae, belonging to three species [Chrysomya
megacephala (Fabricius), C. rufifacies (Macquart) and C. nigripes (Aubertin)] were the most abundance in this study.
The number of species collected from PMBTs was the same found in the FMBTs, but the number of each individual
in each species was different. This study provides baseline information on the necrophilous fauna for estimating
postmortem interval in cases of human death in Thailand.
Keywords: Basic meat bait trap; biodiversity; Diptera, forensic entomology.
Introduction
Fauna studies were conducted to determine bioavailability in a region. It is extremely important to understand
and evaluate processes that appear in nature as a result of gradually increasing anthropogenic effects and resultant
climatic changes. Results of fauna studies are commonly used in applied sciences including entomology, veterinary
science, public health care, agriculture, forestry and ecology, and in the protection of the environment in addition
to basic sciences such as zoology, zoogeography, population genetics, systematics, microbiology and parasitology
(Braioni, 1994; Tamutis et al., 2011). Fauna studies are also useful in elimination of the order of Diptera,
Calliphoridae, Sarcophagidae, Oestridae and Muscidae in public health and veterinary studies, which lead to myiasis
by leaving their larvae and eggs on the wounds of humans and animals (Green et al. 2005; Sankari & Ramakrishnan
2010; Avula et al. 2011; Aggarwal et al. 2014; Jervis-Bardy et al. 2014). Another applied science benefiting from
the results of fauna studies is forensic entomology, which determines postmortem interval, cause of death and whether
corpses have been moved, which contribute to solving forensic cases.
In studies in basic and applied sciences, bait traps used to attract flies to determine the fauna of a region
depend on the sense of smell and types of nutrition of flies (Cavallari et al. 2014). The most well-known traps are
carrion-baited Martín-Vega et al. 2013; Martín-Vega & Baz 2013), followed by basic meat bait traps (BMBTs),
sticky traps, synthetic smell traps and chemical (insecticide) traps (Boonchu et al. 2003; Harvey et al. 2010; Nurita
& Abu 2010). BMBTs, usually prepared with beef and beef liver, are the first traps to be preferred in that they are
inexpensive and effective (Boonchu et al. 2003). The goal of the present study is to compare the effectiveness of pork
liver bait traps (PLBTs) and fish meat bait traps (FMBTs) the collection of flies.
Materials and Methods
Insect collection
Traps were prepared as shown in Figure 1. Pork liver bait traps (PLBTs) and fish meat bait traps (FMBTs)
to serve as attractant, each 200 g, were placed in the traps. As shown in Figure 1, the upper part of each trap, raincoats
and the ropes used in hanging the traps were secured on the same place on the pipes. The traps were hung on the pipe
under the tree near the fresh market (Figure 1A), the tree near the dormitories (Figure 1AB) and the tree near the pig
farm (Figure 1C) in the Kasetsart University, Kamphaeng Saen Campus, at 08.00 h on the first Monday of each month.
Then, traps were left in that place until 17.00 h on the Friday of that same week. The traps were operated during
February and March 2019.
On the first Friday of each week, live adults in the PLBT and FMBT were killed with chloroform. All
the adult, larvae and pupae in both the PLBT and FMBT were removed. Ambient temperatures and relative
humidity were obtained from Nakhon Pathom Meteorological Station which is located near the sampling sites.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 130
Statistical analysis
In order to determine the diversity of species in PLBT and FMBT sample groups in our study, Shannon
Wiener and Simpson diversity and evenness indices were used. Diversity indices were calculated using PC-ORD ver.5.1
(McCune and Mefford, 2006).
To compare the mean temperature and relative humidity in each collection time, a t- test was used to
obtain a 95% confidence level. To evaluate the relationship between environmental factors and insect species, a
Pearson correlation coefficient was used. Statistical analyses were performed using SPSS software (version 16.0).
Result & Discussion
Average relative humidity and temperatures at the study sites are shown in Table 1. For the duration of
the experiment, the mean temperature ranged 24.99±1.21−29.33±1.73°C, whereas the relative humidity ranged
71.07±7.19−80.29±5.47. There was not a significant difference ambient temperatures measured and relative humidity
at the experimental site in each collection time (Table 1.).
A total of 4,252 specimens were collected during two months of the experiment. Of all these species,
2,903 (68.27%) were obtained from PMBTs, while 1,349 (31.73%) were obtained from FMBTs (Table 2.).
Calliphoridae, belonging to three species [Chrysomya megacephala (Fabricius) (Figure 2.), C. rufifacies (Macquart)
(Figure 3.) and C. nigripes (Aubertin) (Figure 4.)] were the most abundance in this study. The other insect group
found in these trap were Muscidae (Musca domestica), Sarcophagidae (Sarcophagida dux), Histeridae (Saprinus sp.)
and Dermestidae (Dermestes maculatus). The number of species collected from PMBTs was the same found in
the FMBTs, but the number of each individual in each species was different.
The purpose of the present study is to compare the effectiveness of pork liver bait traps (PLBTs) and fish
meat bait traps (FMBTs) in the collection of flies. Table 3 showing the diversity index results, similarities are seen
in terms of species composition in both samples. Thus, in terms of species composition of the traps, rates are
similar. The value of diversity index in both tarps was not high because of the area that expose to insect is limit.
The proper range of Shannon`s diversity index ranged 1−3 (Magurran, 1988).
According to the Pearson correlation test, the meteorological factors were associated with the Calliphoridae
species, Chrysomya megacephala and Chrysomya nigripes (Table 4.). This fact emphasizes that the bait traps
temperatures are important with respect to the rate of bait traps decomposition; relative humidity is also important
because it acts directly on the decomposition of the bait trap meat and promotes the emergence of an alternative
decomposition stage (Moura et al., 1997). Because temperature and relative humidity conditions throughout the study
exhibited was not variation, it was not possible to demonstrate the influence of these factors on the presence of
insects on the carcass or on the decomposition process itself (Rungsri et al., 2018).
Conclusion
This was a preliminary study. It is necessary to repeat and replicate it at different times of the year so as
to provide multiple sets of baseline succession data for Thailand that encompass all seasons. However, the information
obtained during this study could be useful for providing initial database information as no succession data was
previously available in central Thailand. Furthermore, these results could also possibly stimulate other entomologists
in Thailand and initiate future studies. The use of fresh meat traps can increase the number of flies collected. The fact
for this study, flies meat bait traps (FMBTs) was attached flies more than pork liver bait traps (PLBTs). Unfortunately,
fish meat bait traps was destroyed by some vertebrate in this area. Fauna studies have scientific and sociocultural
importance. Subsequently, they play an important role in raising public awareness in the determination of endemic
species, climatic change and natural history, and in the protection of nature.
Acknowledgement
This research was supported by Faculty of Liberal Arts and Science, Kasetsart University, Kamphaeng
Saen Campus.
Referenes
Aggarwal, A., Daniel, M. and Shetty, R. 2014. Oral myiasis caused by Chrysomya bezziana in anterior maxilla.
Case Reports in Dentistry 2014: 1–4.
Avula, J.K., Avula, H., Arora, N., Manchukonda, U.K. and Reddy, N.V.V. 2011. Orofacial myiasis of the gingiva
and nasal cavity: a report of two cases and general review. Journal of Periodontology 82:1383–1388.
Boonchu, N., Piangjai, S., Sukontason, K. and Sukontason, K. 2003. Comparison of the effectiveness of baits used
in traps for adult fly collection. Southeast Asian Journal of Tropical Medicine and Public Health 34: 630–633.
Braioni, M. 1994. Faunistic research in a fluvial ecosystem study: the case of the River Adige. Bolletino di
zoologia 61: 415–424.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 131
Cavallari, M., Baltazar, F., De Carvalho, E., Muñoz, D. and Tolezano, J. 2014. A modified Shannon trap for use
in forensic entomology. Advances in Entomology 2: 69–75.
Green, T., Toghill, A. and Lee, R. 2005. Thiamethoxam induced mouse liver tumors and their relevance to
humans: Part 2: Species differences in response. Toxicological Sciences 86: 48–55.
Harvey, B., Bakewell, M., Felton, T., Stafford, K., Coles, G.C. and Wall, R. 2010. Comparison of traps for the
control of sheep blowfly in the U.K. Medical and Veterinary Entomology 24: 210–213.
Jervis-Bardy, J., Fitzpatrick, N., Masood, A., Crossland, G. and Patel, H. 2014. Myiasis of the ear: a review with
entomological aspects for the otolaryngologist. Annals of Otology, Rhinology & Laryngology 124: 345–350.
Magurran, A.E. 1988. Ecological Diversity and its Management. Croom Helm Ltd., London, pp. 179.
Martín-Vega, D., Aguirre-Segura, A., Barranco, P., Baz, A. and Cifrián, B. 2013. Necrophagy in crickets, katydids
and grasshoppers? Orthoptera collected in carrion-baited traps in central Spain. Annales de la Société
entomologique de France (NS) 49: 91–99.
Martín-Vega, D. and Baz, A. 2013. Sex-biased captures of sarcosaprophagous Diptera in carrion-baited traps.
Journal of Insect Science 13: 1–12.
McCune, B. and Mefford, M.J. 2006. PC-ORD. Mutivariate analysis of ecological data. Version 5. MjM Software,
Gleneden Beach, Oregon, USA.
Moura, M.O., Carvalho, C.J.B. and Monteiro-Filho, E.L.A. 1997. A preliminary analysis of insects of medico-
legal importance in Curitiba, State of Paraná. Memórias do Instituto Oswaldo Cruz, 92, 269-274.
Nurita, A. and Abu, H. 2010. Comparative performance of two commercial neonicotinoid baits against filth flies
under field conditions. Tropical Biomedicine 27: 559–565.
Rungsri, N, Suebpongsiri, S and Prommi, T 2018. Occurrences of Calliphoridae (Diptera) on pig carcasses in the
dry season in Thailand. Journal of Food Health and Bioenvironmental Science, 11(1): 29−43.
Sankari, L. and Ramakrishnan, K. 2010. Oral myiasis caused by Chrysomya bezziana. Journal of Oral and
Maxillofacial Pathology 14: 16–18.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 132
Figure 1. Design of liver of pork meat bait traps (PMBTs) and fish meat bait traps (FMBTs) operated under the
tree near the fresh market (A), the tree near the dormitories (B) and the tree near the pig farm (C) in the Kasetsart
University, Kamphaeng Saen Campus, Nakhon Pathom Province, Thailand.
Figure 2. Adult and larvae of Chrysomya megacephala.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 133
Figure 3. Adult and larvae of Chrysomya rufifacies.
Figure 4. Adult and larvae of Chrysomya nigripes.
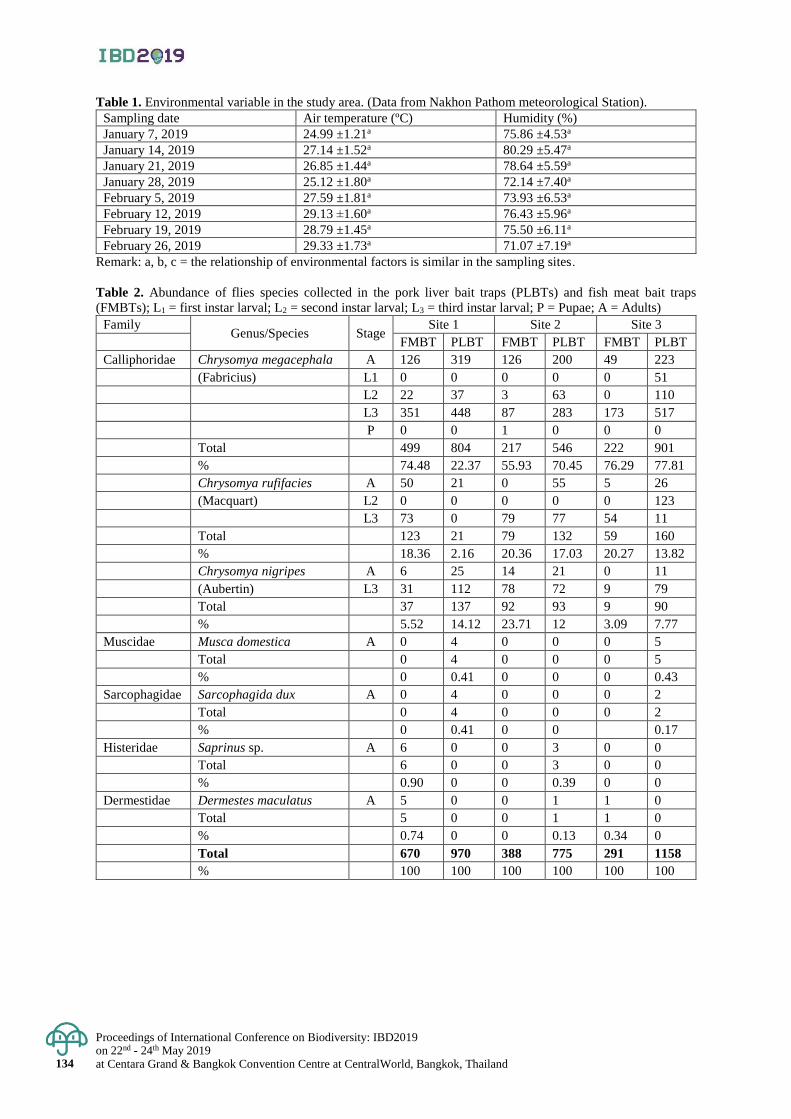
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 134
Table 1. Environmental variable in the study area. (Data from Nakhon Pathom meteorological Station).
Sampling date Air temperature (ºC) Humidity (%)
January 7, 2019 24.99 ±1.21a 75.86 ±4.53a
January 14, 2019 27.14 ±1.52a 80.29 ±5.47a
January 21, 2019 26.85 ±1.44a 78.64 ±5.59a
January 28, 2019 25.12 ±1.80a 72.14 ±7.40a
February 5, 2019 27.59 ±1.81a 73.93 ±6.53a
February 12, 2019 29.13 ±1.60a 76.43 ±5.96a
February 19, 2019 28.79 ±1.45a 75.50 ±6.11a
February 26, 2019 29.33 ±1.73a 71.07 ±7.19a
Remark: a, b, c = the relationship of environmental factors is similar in the sampling sites.
Table 2. Abundance of flies species collected in the pork liver bait traps (PLBTs) and fish meat bait traps
(FMBTs); L1 = first instar larval; L2 = second instar larval; L3 = third instar larval; P = Pupae; A = Adults)
Family Genus/Species Stage
Site 1 Site 2 Site 3
FMBT PLBT FMBT PLBT FMBT PLBT
Calliphoridae Chrysomya megacephala A 126 319 126 200 49 223 (Fabricius) L1 0 0 0 0 0 51 L2 22 37 3 63 0 110 L3 351 448 87 283 173 517 P 0 0 1 0 0 0 Total 499 804 217 546 222 901 % 74.48 22.37 55.93 70.45 76.29 77.81 Chrysomya rufifacies A 50 21 0 55 5 26 (Macquart) L2 0 0 0 0 0 123 L3 73 0 79 77 54 11 Total 123 21 79 132 59 160 % 18.36 2.16 20.36 17.03 20.27 13.82 Chrysomya nigripes A 6 25 14 21 0 11 (Aubertin) L3 31 112 78 72 9 79 Total 37 137 92 93 9 90
% 5.52 14.12 23.71 12 3.09 7.77
Muscidae Musca domestica A 0 4 0 0 0 5 Total 0 4 0 0 0 5
% 0 0.41 0 0 0 0.43
Sarcophagidae Sarcophagida dux A 0 4 0 0 0 2 Total 0 4 0 0 0 2
% 0 0.41 0 0 0.17
Histeridae Saprinus sp. A 6 0 0 3 0 0 Total 6 0 0 3 0 0
% 0.90 0 0 0.39 0 0
Dermestidae Dermestes maculatus A 5 0 0 1 1 0 Total 5 0 0 1 1 0
% 0.74 0 0 0.13 0.34 0 Total 670 970 388 775 291 1158
% 100 100 100 100 100 100
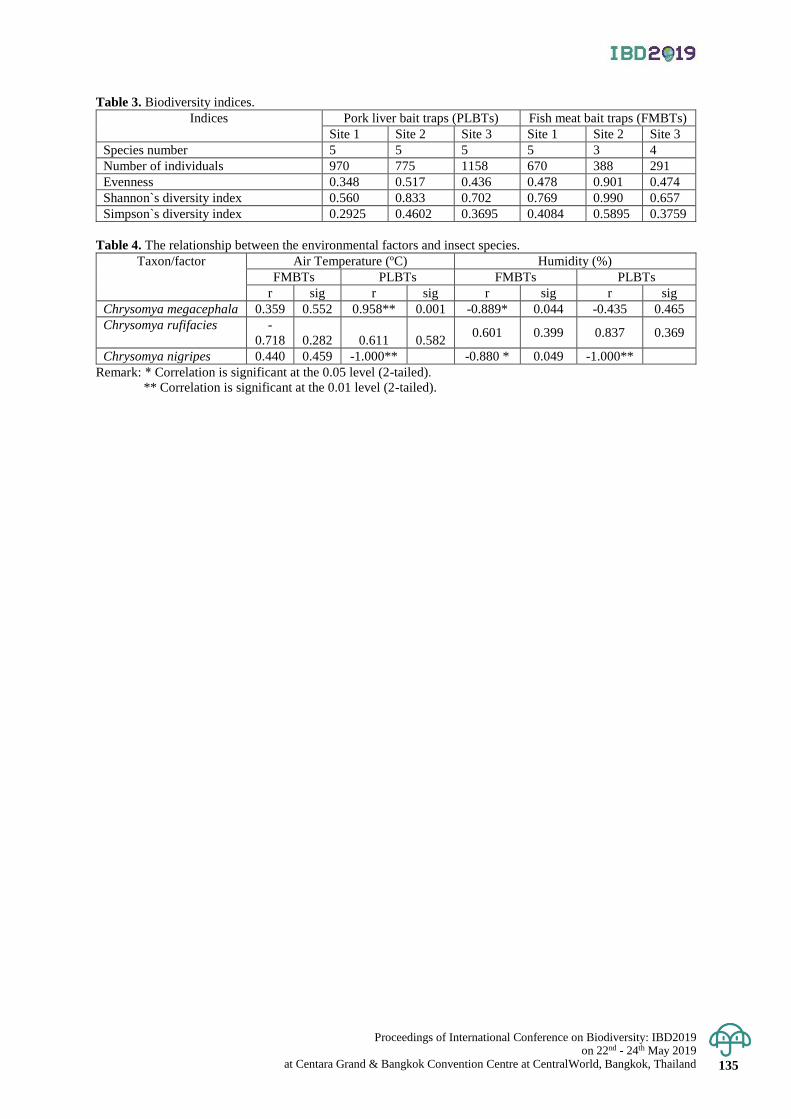
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 135
Table 3. Biodiversity indices.
Indices Pork liver bait traps (PLBTs) Fish meat bait traps (FMBTs)
Site 1 Site 2 Site 3 Site 1 Site 2 Site 3
Species number 5 5 5 5 3 4
Number of individuals 970 775 1158 670 388 291
Evenness 0.348 0.517 0.436 0.478 0.901 0.474
Shannon`s diversity index 0.560 0.833 0.702 0.769 0.990 0.657
Simpson`s diversity index 0.2925 0.4602 0.3695 0.4084 0.5895 0.3759
Table 4. The relationship between the environmental factors and insect species.
Taxon/factor Air Temperature (ºC) Humidity (%)
FMBTs PLBTs FMBTs PLBTs
r sig r sig r sig r sig
Chrysomya megacephala 0.359 0.552 0.958** 0.001 -0.889* 0.044 -0.435 0.465
Chrysomya rufifacies -
0.718 0.282 0.611 0.582 0.601 0.399 0.837 0.369
Chrysomya nigripes 0.440 0.459 -1.000** -0.880 * 0.049 -1.000**
Remark: * Correlation is significant at the 0.05 level (2-tailed).
** Correlation is significant at the 0.01 level (2-tailed).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 136
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 136 - 142
An assessment of habitat connectivity for the endangered Malayan tapir in Thailand
Damisa Kaminsin and Naparat Suttidate* Walailak University, Department of Biology, School of Science, Thasala District, Nakhon Si Thammarat, Thailand
Corresponding author e-mail: [email protected]*
Abstract: The Malayan tapir (Tapirus indicus) is threatened by habitat loss and fragmentation, causing populations
to become small and isolated. An assessment of habitat connectivity patterns is crucial for the survival of the species
because connectivity facilitates species movement and gene flow. Our goal was to identify habitat connectivity
networks for the Malayan tapir in Thailand. We first defined suitable habitat for tapirs using satellite data: evergreen
and deciduous forest patches with the minimum patch size of 13 km2 (average home range size), elevation, slope,
and mean annual precipitation. We then applied circuit theory analysis to assess potential dispersal corridors using
Circuitscape software. We identified 38 suitable habitat forest patches and 13 potential dispersal corridors for tapirs
in Thailand. We found that Khlong Saeng-Khao Sok forest complex was well connected with average habitat patch
size of 2,247.78 km2. In contrast, Chumphon forest complex was the most fragmented forest complex with the highest
number of suitable habitat patches and potential dispersal corridors. In addition, we found ten potential dispersal
corridors within the forest complexes, while there were three potential dispersal corridors between Western –
Kaeng Kra Chan, Kaeng Kra Chan – Chumphon and Chumphon – Khlong Saeng – Khao Sok. The results provide
better understanding of the current pattern of habitat connectivity networks and a guideline for priority areas for
conservation planning. We highlight that an assessment of habitat connectivity pattern can be applied as a wildlife
management tool to assist conservation efforts for the Malayan tapir and other endangered mammals in the region.
Keywords: Circuit theory, Conservation, Corridor, Habitat connectivity, Mammal
Introduction
The Malayan tapir (Tapirus indicus) is a globally endangered species (IUCN, 2016). It is also listed as
an endangered species in the Wild Animal Reservation and Protection Act, B.E. 2535 of Thailand and as Appendix
I status in the Convention on International Trade in Endangered Species (CITES) appendices (CITES, 2017). The
Malayan tapirs is a frugivore and is one of the most important seed dispersal mammals in tropical forest ecosystems.
Malayan tapirs help to decrease plant competition, increasing survival rates of seeds, and decreasing the impacts
of disease. Therefore, the Malayan tapir plays a crucial role in plant communities and tropical biodiversity. Currently,
tapir populations have declined rapidly due to anthropogenic activities, such as habitat loss and fragmentation,
caught by chance, hunting, and road accidents (Corlett, 2007; Holden et al., 2003). Specifically, habitat loss and
fragmentation can fracture a population into at-risk sub-populations. A small, isolated sub-population within a habitat
patch is more likely to become locally extinct due to limited movement between suitable habitats and low genetic
variation among sub-populations (de la Torre, et al., 2017). Thus, the survival of Malayan tapirs in fragmented
landscapes depends upon maintaining connectivity between isolated populations (Fahrig & Merriam, 1985; Noss
et al., 1996; Taylor et al., 1993).
Habitat connectivity is the connectedness between patches of suitable habitats to facilitate movement for
a given species (Lindenmayer & Fischer, 2013). The importance of habitat connectivity is to maintain a specie’s
home range, enhance gene flow among sub-populations, increase the opportunity for adaptation in response to
environmental changes, and mitigate the risk of extinction (Brodie et al., 2015; McRae et al., 2008; Sulistyawan
et al., 2017). In addition, connectivity plays a crucial role in conservation planning where the goal is often to
preserve resilient habitat networks, and design linkages of high-quality habitat (i.e., dispersal corridors) between
remnant patches or protected areas (Soule & Terborgh, 1999). Habitat connectivity has been successfully applied
in conservation planning for species in many regions, such as deer (Odocoileus hemionus) in California and
Arizona (Beier et al., 2006), jaguars (Panthera onca) in Mexico (de la Torre et al., 2017), and pandas (Ailuropoda
melanoleuca) in China (Sulistyawan et al., 2017; Wang et al., 2014). Despite the advancements in theory and application,
the assessment of habitat connectivity for Malayan tapirs has not yet been studied in Thailand which hampers conservation
efforts (Lynam et al., 2012). Broad-scale conservation planning for Malayan tapirs cannot be successful without
identifying suitable habitat patches, dispersal corridors, and an effective assessment of a connectivity network that
can maintain the viability of tapir populations and facilitate gene flow among populations (Lynam et al., 2012).
The goal of our study was to identify habitat connectivity networks for the Malayan tapir in Thailand.
First, we defined suitable habitat patches for Malayan tapirs, and then evaluated the patterns of habitat connectivity
using circuit theory.
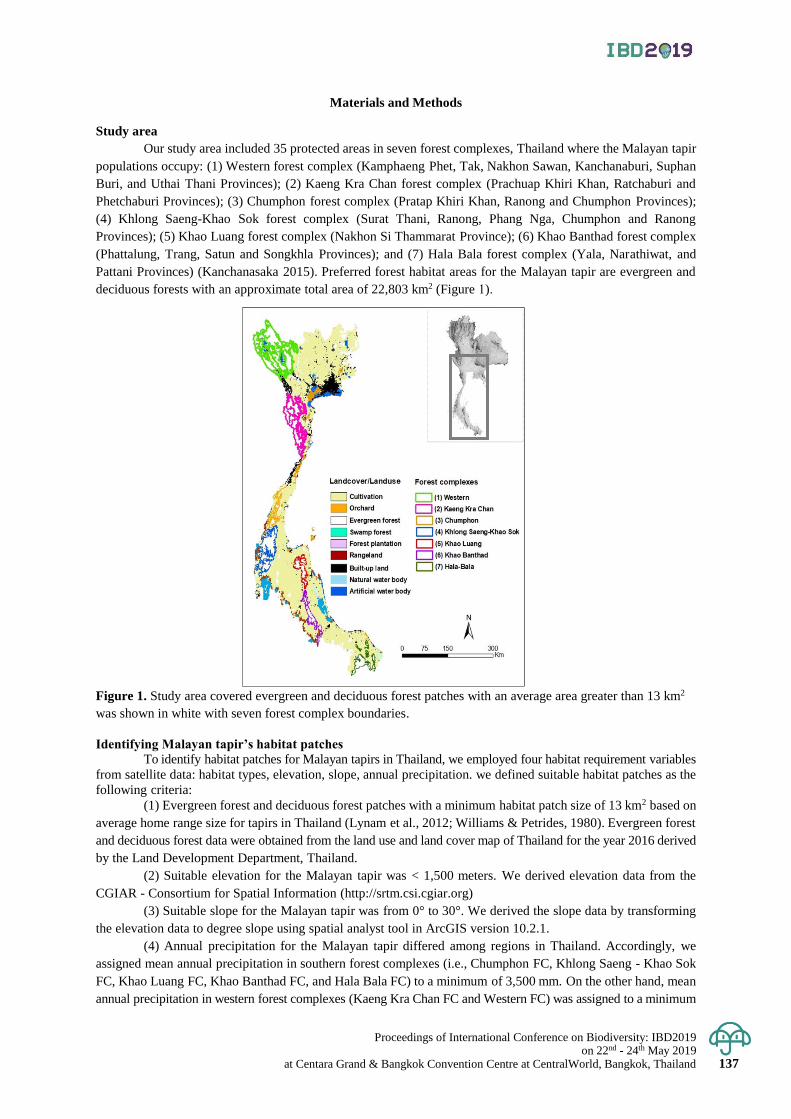
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 137
Materials and Methods
Study area
Our study area included 35 protected areas in seven forest complexes, Thailand where the Malayan tapir
populations occupy: (1) Western forest complex (Kamphaeng Phet, Tak, Nakhon Sawan, Kanchanaburi, Suphan
Buri, and Uthai Thani Provinces); (2) Kaeng Kra Chan forest complex (Prachuap Khiri Khan, Ratchaburi and
Phetchaburi Provinces); (3) Chumphon forest complex (Pratap Khiri Khan, Ranong and Chumphon Provinces);
(4) Khlong Saeng-Khao Sok forest complex (Surat Thani, Ranong, Phang Nga, Chumphon and Ranong
Provinces); (5) Khao Luang forest complex (Nakhon Si Thammarat Province); (6) Khao Banthad forest complex
(Phattalung, Trang, Satun and Songkhla Provinces); and (7) Hala Bala forest complex (Yala, Narathiwat, and
Pattani Provinces) (Kanchanasaka 2015). Preferred forest habitat areas for the Malayan tapir are evergreen and
deciduous forests with an approximate total area of 22,803 km2 (Figure 1).
Figure 1. Study area covered evergreen and deciduous forest patches with an average area greater than 13 km2
was shown in white with seven forest complex boundaries.
Identifying Malayan tapir’s habitat patches To identify habitat patches for Malayan tapirs in Thailand, we employed four habitat requirement variables
from satellite data: habitat types, elevation, slope, annual precipitation. we defined suitable habitat patches as the
following criteria:
(1) Evergreen forest and deciduous forest patches with a minimum habitat patch size of 13 km2 based on
average home range size for tapirs in Thailand (Lynam et al., 2012; Williams & Petrides, 1980). Evergreen forest
and deciduous forest data were obtained from the land use and land cover map of Thailand for the year 2016 derived
by the Land Development Department, Thailand.
(2) Suitable elevation for the Malayan tapir was < 1,500 meters. We derived elevation data from the
CGIAR - Consortium for Spatial Information (http://srtm.csi.cgiar.org)
(3) Suitable slope for the Malayan tapir was from 0° to 30°. We derived the slope data by transforming
the elevation data to degree slope using spatial analyst tool in ArcGIS version 10.2.1.
(4) Annual precipitation for the Malayan tapir differed among regions in Thailand. Accordingly, we
assigned mean annual precipitation in southern forest complexes (i.e., Chumphon FC, Khlong Saeng - Khao Sok
FC, Khao Luang FC, Khao Banthad FC, and Hala Bala FC) to a minimum of 3,500 mm. On the other hand, mean
annual precipitation in western forest complexes (Kaeng Kra Chan FC and Western FC) was assigned to a minimum

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 138
of 2,500 mm. Annual precipitation was obtained from WorldClim version 2 (http://worldclim.org/version2) (Fick
& Hijmans, 2017).
We prepared all habitat data in raster format with 1-km resolution because it is a proper resolution for
spatial analysis of mammals in Thailand (Lynam et al., 2012). We analyzed suitable habitat patches for the Malayan
tapir using ArcGIS program version 10.2.1.
Assessing habitat connectivity for Malayan tapirs
To evaluate potential dispersal corridors for the Malayan tapir, we used Circuitscape software version
4.0 (McRae et al., 2008). Circuitscape applies electrical circuit theory and random walk theory to calculate
pairwise resistances and create maps of current flowing between focal nodes (McRae et al., 2008). Circuitscape
requires two inputs: (1) a resistance map (i.e., land use and land cover with a degree of movement resistance for
Malayan tapirs); and (2) a focal node map (i.e., forest habitat patches). We prepared the resistance map by assigning
resistance values to each land use and land cover raster, ranking from 1 to 100. We assigned lowest resistance
values to forest habitat: evergreen and deciduous forest patches. Higher resistance values were assigned to other
natural habitats: swamp forest, rangeland, marsh and swamp, and small natural water body. Highest resistance
values were assigned to agricultural areas, such as forest plantation, and orchards (Table 1). For complete barriers,
we assigned them as no data areas (i.e., settlements, roads, perennial agricultural areas, and large water bodies).
Circuitscape created current maps between pairs of forest patches using pairwise analysis and iterated all pairs
into focal nodes (McRae, et al., 2013).
Table 1. Resistance values assigned to create the resistance map.
Category Resistance score
Evergreen forest 1
Deciduous forest 1
Swamp forest 20
Forest plantation 30
Rangeland 40
Marsh and swamp 50
Small natural water body 60
Agricultural areas 100
Result & Discussion
We identified 38 potential suitable forest patches in seven forest complexes with approximately an area
of 23,731 km2 and average patches size of 625 km2 (Table 2). Chumphon forest complex had the highest number
of forest habitat patches, while Khao Luang and Khlong Saeng - Khao Sok had the lowest number of forest habitat
patches. Additionally, we found large forest habitat patches (> 625 km2) in Western, Kaeng Kra Chan, Khao Luang
and Khlong Saeng-Khao Sok forest complexes. In contrast, we found many small forest habitat patches (< 625 km2)
in the Chumphon, Hala-Bala, and Khao Banthad forest complexes (Table 2). In addition, we found that Western
and Kaeng Kra Chan forest complexes had seven forest patches which included both evergreen and deciduous
forest habitat types. On the other hand, we found that suitable forest habitat patches in the south of Thailand were
only evergreen forests due to high rainfall and humidity (Kanchanasaka, 2015; Lynam et al., 2012).
We also identified 13 potential dispersal corridors: 10 dispersal corridors within the forest complexes
(Table 3), and three dispersal corridors between forest complexes: (A)Western forest complex – Kaeng Kra Chan
forest complex; (B) Kaeng Kra Chan forest complex – Chumphon forest complex; (C)Chumphon forest complex
– Khlong Saeng – Khao Sok forest complex (Figure 2).
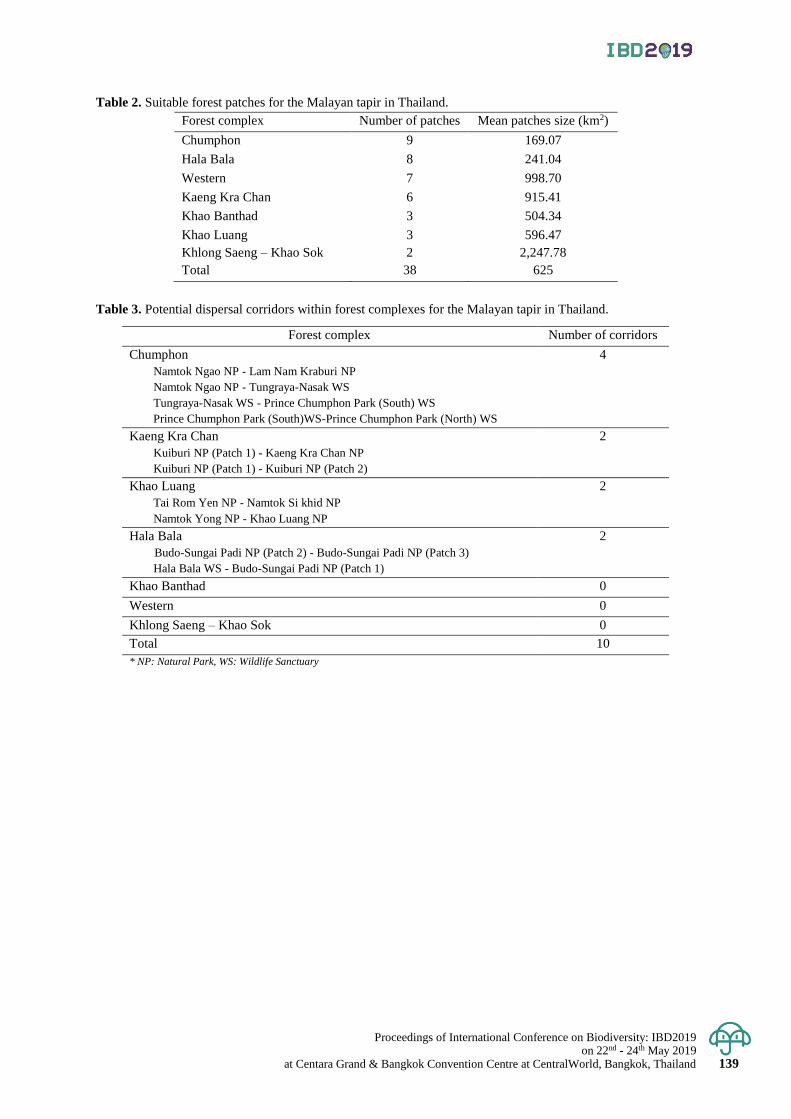
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 139
Table 2. Suitable forest patches for the Malayan tapir in Thailand.
Forest complex Number of patches Mean patches size (km2)
Chumphon 9 169.07
Hala Bala 8 241.04
Western 7 998.70
Kaeng Kra Chan 6 915.41
Khao Banthad 3 504.34
Khao Luang
Khlong Saeng – Khao Sok
3
2
596.47
2,247.78
Total 38 625
Table 3. Potential dispersal corridors within forest complexes for the Malayan tapir in Thailand.
Forest complex Number of corridors
Chumphon
Namtok Ngao NP - Lam Nam Kraburi NP
Namtok Ngao NP - Tungraya-Nasak WS
Tungraya-Nasak WS - Prince Chumphon Park (South) WS
Prince Chumphon Park (South)WS-Prince Chumphon Park (North) WS
4
Kaeng Kra Chan
Kuiburi NP (Patch 1) - Kaeng Kra Chan NP
Kuiburi NP (Patch 1) - Kuiburi NP (Patch 2)
2
Khao Luang
Tai Rom Yen NP - Namtok Si khid NP
Namtok Yong NP - Khao Luang NP
2
Hala Bala
Budo-Sungai Padi NP (Patch 2) - Budo-Sungai Padi NP (Patch 3)
Hala Bala WS - Budo-Sungai Padi NP (Patch 1)
2
Khao Banthad 0
Western 0
Khlong Saeng – Khao Sok 0
Total 10
* NP: Natural Park, WS: Wildlife Sanctuary

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 140
Figure 2. The results showed pairwise current flow density (Amps/cell) between three forest complexes: (A)
Western forest complex (1) and Kaeng Kra Chan forest complex (2); (B) Kaeng Kra Chan forest complex (2) and
Chumphon forest complex (3); (C) Chumphon forest complex (3) and Khlong Saeng – Khao Sok forest complex
(4). The highest current density (shades of yellow) indicated areas of potential dispersal corridors. Areas, where
connectivity was most difficult were shown in purple. Evergreen and deciduous forest habitat patches with an
area greater than 13 km2 were shown in green.
Discussion
Our goal was to identify habitat connectivity networks for the Malayan tapir in Thailand. We assessed
patterns of potential forest habitat patches and dispersal corridors within and between forest complexes. Our
results provide important baseline information for Malayan tapir conservation planning. Especially, the Malayan
tapir populations are at risk of locally extinct primarily due to habitat loss and fragmentation (Lynam et al., 2012).
Recent estimate Malayan tapir abundance in Thailand is approximately 538 – 720 individuals, occupying seven
forest complexes: Western, Kaeng Kra Chan, Chumphon, Khlong Saeng-Khao Sok, Khao Luang, Khao Banthad
and Hala Bala forest complexes (Kanchanasaka 2015).
Our results demonstrated that Chumphon, Hala Bala, Khao Banthad, and Khao Luang forest complexes
were more fragmented than other forest complexes. These forest complexes experience changes in land use and
land cover from anthropogenic activities, such as perennial crop, field crop, orchard, built-up areas, and artificial
water bodies. Malayan tapir populations are more likely to be separated in those forest complexes, and occupy
small proportions of the available forest habitat patches (Lynam et al., 2012; Clements et al., 2012). Malayan
Tapirs avoid human-made landscape and anthropogenic activity because they are nocturnal and solitary, and also
prefer wetter places in denser forest habitat (Clements et al., 2012). Malayan tapirs may also avoid natural water
bodies, if such areas were used by human activities (Lynam et al., 2012; Kanchanasaka 2015). In addition,
Chumphon forest complex had the lowest mean patches size, indicating that forest habitat patches in this forest
complex become more fragmented and degraded. This might due to geographical location and topography where
Chumphon forest complex is located along the western border of Thailand and potentially vulnerable to human
disturbance (Lynam et al., 2012; Kanchanasaka 2015).
We found that Chumphon forest complex had the highest number of potential dispersal corridors,
indicating habitat fragmentation and degradation. However, there was no potential dispersal corridor in Western
forest complex, and Khlong Saeng – Khao Sok forest complex because such forest complexes are well connected
and most of protected areas in each forest complex are adjacent to each other (Lynam et al., 2012; Kanchanasaka 2015).
The current flows among adjacent forest patches were substantially higher than the current flows in nonadjacent
forest patches, indicating that short distance dispersal corridors can facilitate tapirs’ movement better than the

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 141
long-distance corridors. The analyses revealed that potential dispersal corridors with low resistance mostly were
evergreen forest and deciduous forest.
The results showed that there were more potential dispersal corridors within the forest complexes than
between the forest complexes. Also, corridors within the forest complexes were connected to larger forest habitat
patches. Similarly, the study of habitat connectivity for Pumas (Puma concolor) in the southwestern United States
(Arizona and New Mexico) demonstrates that maximum current flow corridors are more likely to be located within
close proximity to high quality habitat (Dickson et al., 2013). Furthermore, we found that forest patches in Hala
Bala forest complex were more isolated with the long-distance potential corridors than other forest complexes due
to conversion of natural habitat to perennial crops. We found no potential dispersal corridors in Khlong Saeng-
Khao Sok forest complex and Western forest complex because the complexes have the largest forest patch size
with adjacent patches. Conversely, we found that forest patches in Khao Banthad forest complex were threatened
by agricultural areas that resulted in no connectedness between patches.
We suggest that Western forest complex and Khlong Saeng-Khao Sok forest complex had the lowest
habitat fragmentation with high quality habitat for Malayan tapir populations. This forest complex could
potentially be an important source for habitat connectivity network.
Conclusions
An assessment of potential suitable habitat patches and dispersal corridors for Malayan tapirs in Thailand
provides preliminary information to assist conservation planning and management for the Malayan tapir in Thailand.
Specifically, less fragmented and well-connected evergreen and deciduous forest patches can potentially be an
important refuge for the endangered Malayan tapir in Thailand. Habitat connectivity is crucial to maintain the
viability of Malayan tapirs and can help conservation management for other endangered species in the regions.
Acknowledgement
We would like to thank Walailak University and the Development and promotion of science and technology
talents project (DPST) for funding. We are thankful to the Land Development Department (Thailand) for sharing
the land use and land cover map of Thailand for the year 2016.
References
Beier, P., Majka, D.,and Jenness, J. 2006. Conceptual steps for designing wildlife corridors. Corridor Design
Arizona USA 269: 23784–23789.
Brodie, J. F., Giordano, A. J., Dickson, B. G., Hebblewhite, M., Bernard, H., Mohd-Azlan, J., Anderson, J., and
Ambu, L.2015. Evaluating multispecies landscape connectivity in a threatened tropical mammal
community. Conservation Biology 29: 122-132.
Clements, G. R., Rayan, D. M., Aziz, S. A., Kawanishi, K., Traeholt, C., Magintan, D., Yazi, M. F. A., and
Tingley, R. 2012. Predicting the distribution of the Asian tapir (Tapirus indicus) in Peninsular Malaysia
using maximum entropy modelling. Integrative zoology 7: 402-409.
Corlett, R.T. 2007. The impact of hunting on the mammalian fauna of tropical Asian forest. Biotropica 39(3): 292-303.
de la Torre, J. A., Núñez, J. M., and Medellín, R. A.2017. Habitat availability and connectivity for jaguars
(Panthera onca) in the Southern Mayan Forest: Conservation priorities for a fragmented landscape.
Biological Conservation 206:270-282.
Dickson, B. G., Roemer, G. W., McRae, B. H., and Rundall, J. M. 2013. Models of Regional Habitat Quality and
Connectivity for Pumas (Puma concolor) in the Southwestern United States. PLoS ONE 8(12): e81898.
Fahrig, L., and Merriam, G. 1985. Habitat patch connectivity and population survival. Ecology 66: 1762-1768.
Holden J, Yanuar A and Martyr D. 2003. The Asian Tapir in Kerinci Seblat National Park, Sumatra: evidence
collected through photo-trapping. Oryx 37(1): 34 -40.
IUCN. 2016. The IUCN Red List of Threatened Species: Tapirus indicus, Malay Tapir. Version 2016-1 Retrieved
12 August, 2018, from Available at: www.iucnredlist.org.
Kanchanasaka, B. 2015. Habitat Use and Population of Tapirs in Thailand. Report to at the Wildlife Research Division,
Wildlife Conservation Office, Department of National Parks,Wildlife and Plant Conservation, Bangkok.
Lindenmayer, D. B., and Fischer, J. 2013. Landscape modification and habitat fragmentation: a synthesis. Global
Ecology and Biogeography 16: 265–280.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 142
Lynam, A. j., Tantipisanuh, N., Chutipong, W., Ngoprasert, D., Baker, M. c., Cutter, P., Kitamura, S., Steinmetz,
R., Sukmasuang, R., and Thunhikorn, S. 2012. Comparative sensitivity to environmental variation and
human disturbance of Asian tapirs (Tapirus indicus) and other wild ungulates in Thailand. Integrative
zoology 7(4): 389-399.
McRae, B. H., Dickson, B. G., Keitt, T. H., and Shah, V. B. 2008. Using circuit theory to model connectivity in
ecology, evolution, and conservation. Ecology 89: 2712-2724.
Noss, R. F., Quigley, H. B., Hornocker, M. G., Merrill, T., and Paquet, P. C. 1996. Conservation biology and
carnivore conservation in the Rocky Mountains. Conservation Biology 10: 949-963.
Soule, M. E., and Terborgh, J. 1999. Conserving nature at regional and continental scales - a scientific program
for North America. Bioscience 49: 809-817.
Steinmetz, R., Chutipong, W., Seuaturien, N. and Cheungsa-ad, E.2008. Community structure of large mammals
in tropical montane and lowland forest in the Tenasserim-Dawna mountains, Thailand. Biotropica 40: 344-353.
Sulistyawan, B. S., Eichelberger, B. A., Verweij, P., Boot, R. G. A., Hardian, O., Adzan, G., and Sukmantoro, W.
2017. Connecting the fragmented habitat of endangered mammals in the landscape of Riau–Jambi–
Sumatera Barat (RIMBA), central Sumatra, Indonesia (connecting the fragmented habitat due to road
development). Global Ecology and Conservation 9: 116–130.
Taylor, P. D., Fahrig, L., Henein, K., and Merriam, G. 1993. Connectivity is a vital element of landscape structure.
Oikos 68: 571-573.
Wang, F., McShea, W. J., Wang, D., Li, S., Zhao, Q., Wang, H., and Lu, Z. 2014. Evaluating Landscape Options
for Corridor Restoration between Giant Panda Reserves. PLoS One 9(8): e105086.
Williams, K. D., and Petrides, G. A. 1980. Browse Use, Feeding Behavior, and Management of the Malayan
Tapir. The Journal of Wildlife Management 44(2): 489-494.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 143
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 143 - 147
Satun UNESCO Global Geopark, the readiness for sustainable tourism
Suwimon Hengpatana*, Phannipha Anuruksakornkul, Apinya Wanaset,
Taweesin Tangseng and Danuvas Suwanwong
Faculty of Economics, Srinakharinwirot University, Wattana District, Bangkok, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Satun Geopark is recently UNESCO Global Geopark creating pride for the people in the country. The area
covers four of seven districts of Satun Province namely Thungwa, Manang, La-Ngu, and Muang Satun. The Geopark
attraction points are categorized into four types. It consists of ecotourism and adventure attraction; sea – islands
attraction; attraction for learning; and intangible/ cultural heritage. The famous tourist attractions are Li-Pe Island,
Hin Ngam Island, Adang-Ra Wi Islands, Tarutao National Park, Petra National Park, Pak Bara Beach, Satun Karst
Zone includes Le- Stegodon Sea Cave, Chet Khot Cave, and Phu Pha Phet Cave. Being a global geopark can
attract eco-tourists from all over the world. Therefore, in the Satun geopark, there must be efficient and proper tourism
management not only to preserve the existing ecosystem but also impress the tourists. Logistics management is consequently
applied to sustainable tourism. Satun Geopark logistics management can be categorized into four components which
consist of geological resources and landscape; transportation; amenities; and environmental management as well as
creating the value of tourism products using three supporting factors: information and communication technology;
innovation; and sustainable management provide tourism value for the tourists in terms of flexible, convenient, fast,
and safe. The objectives of this paper are to discover the component of logistics management and to evaluate the logistics
management potential for sustainable tourism for Satun UNESCO Geopark. The results could be used for government
agencies, and the private sector involved such as Satun Provincial Administrative Organization, Sub-district Administrative
Organization in the area as well as travel service operators.
Keywords: Satun UNESCO Global Geopark, logistics management, sustainable tourism, geotourism
Introduction
In Thailand, tourism has become a pioneer sector and made a significant contribution to the country’s
gross domestic product (GDP). It generates income and creating employment for local people. During January to
August 2018, there are 25,886,325 foreign tourists. Compared to the same period of last year tourism expanded by 9.94%,
in line with the expansion of tourists in almost all regions and generated total revenue of 1,350,317.90 million
baht, expanding by 12.85% from the same period of last year. From the large amount of tourists, the management
of sustainable tourism is needed.
Thailand is a country with abundant of natural resources, including cultural and historical attractions.
Southern Thailand contains many islands and beautiful beaches. As UNESCO announced to Satun geopark is
UNESCO global geopark on April 17, 2018, creating pride for the people in the country. Being a global geological
park can attract eco-tourists from all over the world. Satun Geopark covered four districts of Satun Province
namely; Muang, La Ngu, Tungwa, and Manang located along the Coast of Andaman Sea. It possesses many
beautiful islands such as Li-Pe Island, Hin Ngam Island, Adang-Ra Wi Islands, and Dong Islands located in Tarutao
National Park. Moreover, Satun Geopark also features some attractions about culture and the local way of lives.
The Geopark attraction points can be categorized into four types; (1) ecotourism and adventure attraction such as
Le-Stagodon Cave, Puphapet Cave, Chet Chod Cave, Thanpliew Waterfall; (2) sea – islands attraction such as
Adang - Ra Wi Island, Li-Pe Island, Tarutao Island, Petra Island. There are also dunes called Dragon’s back at
Tan-Yong-Po and many diving sites such as Jabang diving site; (3) attraction for learning such as Satun Geopark
Museum, many fossil sites such as Khao Noi stone hierarchy, trilobite fossil at Tarutao Island, nautiloid at Khao
Dang. There are also many Karst in Thungwa and Manang Districts; (4) intangible/ cultural heritage such as Ma-Ni,
a native Nigrito at Manung District, floating boat tradition of Orang Laut, the indigenous sea people at Adang and
Li-Pe Island. Since there are many types of attractions in the area, and some area is famous for foreign tourists,
there must be attracted abundant of tourists both Thai and foreigner tourists. Therefore, in the Satun geopark, there
must be efficient and proper tourism management not only to preserve the existing ecosystem but also impress
the tourists. Logistics management is the way to manage sustainable tourism. By managing the tourism logistics,
it is the planning of the administration of tourism products and services before tourists travel until tourists come
to travel conveniently throughout the journey.
This article, therefore, is the studied the logistics management components for tourism and to evaluate
the logistics management potential for sustainable tourism of Satun UNESCO Global Geopark. The government

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 144
sector can apply the findings to prepare for the infrastructure, logistics system, and facilities to develop logistics
management for sustainable tourism to facilitate the tourists and to create a balance in the economy, society, and environment.
Materials and Methods
The area of the study is Satun Geopark (as known as Satun UNESCO Global Geopark) which composed of four
of seven desticts of Satun Province namely; Muang, Thungwa, La-Ngu, and Manang District. The area covers four types
of geopark attraction points; ecotourism and adventure attraction; sea – islands attraction; attraction for learning;
and intangible/ cultural heritage.
Geotourism is a form of tourism that sustains or enhances the geographical characters of a location such
as its geology, environment, culture, heritage, aesthetics, and the well-being of the communities. (Tourtellot, 2011).
There are several works of literature about geotourism management in different principles. Valiakos (2010)
mentioned five components of geological park management. It composes of 1) geology and landscape which consist
of three elements; territory; geography conservation; and natural and cultural heritage 2) management structure
3) interpretation and environmental study 4) geotourism 5) sustainable economic management in the area. Lewis
T.O. Cheung (2014) explained the factors that affect geopark. There are the external and the internal value of the park.
The external value of the park consists of five components. There are convenient public transportation, easy access
to the bathroom, visitor guides and useful maps, adequate leisure facilities, and clear road signs. Also, the internal
value of the park consists of four components which are outstanding scenery and landscape; beautiful mountain
and attractive area; coast, seashore, and beautiful beach; and interesting geology and ecology. He also analyzed
the three main components of motivation in tourism. It includes 1) travel for novelty which are five elements that
tourist have; enjoyment and fun; freedom; new experiences in different lifestyles; tried exotic foods; and seen as
much as can be seen during the free time. 2) Leisure travel which includes what the tourists want to have in travel;
body movements and participation in sports; positive attitude about the trip after returning home; and have been
changed from busy jobs. 3) Tourism for escape from something which is an escape from the need at home;
searching for excitement; meet people with similar interests. Moreover, Patrick (2014) described how to develop
ideas and activities in geological park. He mentions four categories. 1) Sustainable mobility: the concept of public
transport or bicycle access 2) Sustainable hotel business 3) Geothermal energy from Geological Park 4) Energy and
climate strategy.
Result & Discussion
The integrity of tourism resources in ecology, terrain, and nature, Satun Geopark create various types
of natural tourism activities. The activity has been classified into various types (and still counting). The tourism
sector today aims to cater to the needs and preferences of all types of tourists, and thus, seems to take into consideration
specific areas of their interest. In present, there are many tour operators provide variety of tourism activities for
the tourist. Their offices located near Pak Bara Port. The tourists are able to choose the activities or the trip that they prefer
from the tour operator in the area or book it from internet.
The tourism in Satun Geopark can be categorized as following:
1) Adventure Tourism (as known as adventure travel). This kind of tourism has been famous among adventure seeker
in Satun Geopark. The adventure tourists indulge in challenging activities such as water rafting, scuba diving, caving,
hiking, rock climbing, and so on. There are several adventure points in Satun Geopark such as Le-Stegodon Cave,
Chet Chod Cave, Wangsaithong Waterfall, and so on. The diving sites in the area are Jabang Pinnacle, Adang-
Rawi, Li-Pe Island, Tarutao Islands, and Petra Islands. The tourists can go diving by buying a package tour from tour
operator at Pak-Bara Port and nearby and speed boat rental is also available.
2) Cultural Tourism. The concept of cultural tourism encompasses things, such as history of a given region, the lifestyle
of people in a particular geographical locale, architecture, oral traditions, religions, festivals, cuisine, and so on. Tourists
can visit the floating boat which is the native fishery tradition.
3) Ecotourism. Ecotourism also encompasses the concepts of geotourism. It involves the traveling to primary
attractions which are natural beauty or settlements of indigenous communities. The tourists can learn the lifestyle
of Mani (the native Nigrito) in Thungwa, Manang, and La-Ngu Districts.
4) Educational Tourism. There are many fossil sites in Satun Geopark for geo-scientist, geologist, students, and
other people who interested in studying variety of fossil. The Satun Geopark terrain originated as a sea floor in the Cambrian
Period more than 500 million years ago. The oldest fossils of primitive sea life in Thailand were found in the red
sandstones on Tarutao Island.
As various types of attraction points and variety activities provided in Satun Geopark, the logistics
management is consequently applied to sustainable tourism. From the review literature, logistic management of sustainable
tourism of geopark can be categorized into four components and applied to Satun UNESCO Global Geopark as
in Table 1. The components of logistics management for sustainable tourism are 1) transportation 2) amenities 3)

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 145
geological resources and landscape, and 4) environmental management as well as creating value of tourism
products using three supporting factors: information and communication technology; innovation; and sustainable
management provide tourism value for the tourists in terms of flexible, convenient, fast, and safe.
The evaluation of the particular component of logistics management for sustainable tourism of Satun
Geopark is as following:
1) Transportation. There are three sub-component combining to Satun Geopark. A) The transportation from the
visitor’s origin to Satun Geopark is essential for the visitor to decide whether they travel to travel site or not. The transportation
information should inform the visitor how to go to the main attraction points such as the type of vehicle, the time
table of the train from the nearby airport or train station to Satun Geopark and back. B) The transportation network
among geo-point is significant. In the present, the visitor or the tourist would like to travel by public transportation.
The local transit in Satun Geopark has to be improved to support the coming visitors travel from site to site. There
is still a lack of local transportation. The best way for the visitor is to rent a car or a motorcycle, but there is an insufficient
service provider in the area. C) Transportation from Satun Geopark to Langkawi Island, Malaysia. Because it is
convenient for the visitor to travel from Langkawi Island to Satun Geopark, it takes about one hour by speed boat
from Langkawi Island to Li-Pe Island. Also, there is an international airport in Langkawi Island. The transportation
in the area is the obstacle for Satun Geopark tourism development, and it has to be improved shortly.
2) Amenities. It composes of accommodation, restaurant, primary healthcare and hospital, public service, and
travel service provider. The accommodation in Satun Geopark is hotel and homestay. There are many hotels and
resorts in Li-Pe Island and land, but there is still a lack of homestay in the area. For primary public healthcare and
hospital, there are emergency medical service ( EMS) system provided by every Sub-District Administrative
Organization (SAO) in Satun Geopark, but in some SAOs have to improve their services. There is also a marine
emergency service from Tarutao National Park located at Rawi Island. The public service in the area is security
checkpoints for the police. The travel service providers are private businesses mostly located near Pak Bara Port.
3) Geological Resources and Landscape. The sub-component of this part are a territory and natural beauty,
ecological value, nature, cultural heritage, and landscape design around the geopark site. There are abundant
resources of attraction in Satun Geopark. The landscape around geopark must have well design and fit the nature.
4) Environmental management. There is four sub-component consist of water supply, electricity supply, waste
disposal, and air pollution control. In the present, there are wastewater and waste disposal in some areas. This
problem has to be appropriately diminished. The Chief of SAOs and stakeholders in the area have to solve these
problems. The obstacle is political in the local areas.
Conclusions
Satun Geopark is recently UNESCO Global Geopark. Being a global geological park can attract eco-tourists
from all over the world. Therefore, in the Satun geopark, there must be efficient and proper tourism management
not only to preserve the existing ecosystem but also impress the tourists. Logistics management is consequently
applied to sustainable tourism. Satun Geopark logistics management can be categorized into four components. It
consists of geological resources and landscape; transportation; amenities; and environmental management as well
as creating the value of tourism products using three supporting factors: information and communication technology;
innovation; and sustainable management provide tourism value for the tourists in terms of flexible, convenient, fast, and safe.
The logistics management for sustainable tourism needs a lot of support from both local and central
government. The government has not only promoted and supported the tourism industry, but also requiring the
tourism industry to be a part of the country’s strategic plans such as National Economic and Social Development Plan,
National Tourism Development Strategy, and Service sector development strategy of Thailand. The strategies
focus on driving policies to promote, support and develop tourism, both products and service systems, to be able
to facilitate tourists for maximum efficiency along with sustainable development for tourism which will lead to
income generation in the country and increase the competitiveness of the country.
Acknowledgement
The authors acknowledge the financial support provided by The Thailand Research Fund (TRF) for
research project “Logistics Management for Sustainable Tourism of Satun UNESCO Global Geopark”.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 146
References
Cheung, TOL; Fok, L. 2014. The motivations and environmental attitudes of nature-based visitors to protected
areas in Hong Kong. Available on: https://mailattachment.googleusercontent.com/attachment/u/0/?ui=2
&ik=8381694a82&view=att&th=164b059fb9885723&attid=0.5&disp=inline&safe=1&zw&saddbat=A
NGjdJ9 (16 July 2018)
Lewis T.O. Cheung, Lincoln Fok & Wei Wei Fang. 2014. Understanding geopark visitors' preferences and
willingness to pay for global geopark management and conservation. Available on:
https://www.tandfonline.com /doi/abs/10.1080/14724049.2014.941848 (16 July 2018)
Petrick. 2014. Genetics, neuroscience, and psychopathology: Clothing the emperor. Available on:
http://globalgeoparksnetwork.org/wp-content/uploads/2015/09/6th-International-UNESCO Conference-
on-Global-Geoparks_Sept_13.pdf (16 July 2018)
Piboonrungroj and Disney. 2012. Tourism supply chains: a conceptual framework. PhD Networking Conference.
Tourtellot Jonathan. 2011. UNESCO’s geoparks “Clarify” geotourism. Available on:
https://blog.nationalgeographic.org/2011/11/16/unescos-geoparks-clarify-geotourism/ (2 July 2019)
Zouros N.,Valiakos I. 2010. Geoparks management and assessment. Available on
https://ejournals.epublishing.ekt.gr/index.php/geosociety/article/view/11262 (16 July 2018)
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 147
Table 1. The Components of Logistics Management of Sustainable Tourism of Geopark of Satun UNESCO
Global Geopark.
Component Sub-Component Source
1. Transportation - Transportation from origin to Satun
Geopark
- Transportation network in Satun
Province and nearby
- Transportation from Satun Geopark to
Langawi Island, Malaysia
Zouros N.,Valiakos I.
(2010),Lewis T.O. Cheung
(2014), Piboonrungroj and
Disney ( 2012(
2. Amenities - Accommodation
- Restaurant
- Primary healthcare and hospital
- Public service
- Travel service provider
Zouros N.,Valiakos I.
(2010 ,Lewis T.O. Cheung
(2014)
3. Geological Resources and
Landscape
- Territory and natural beauty and
ecological value
- Nature
- Cultural heritage
- Landscape design around geopark site
Zouros N.,Valiakos I.
(2010), Petrick (2014)
4. Environmental management - Water supply
- Electricity supply
- Waste disposal
- Air pollution control
Zouros N.,Valiakos I.
(2010), Petrick (2014)

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 148
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 148 - 155
Analysis of factors related to bamboo structure for preventing erosion and restoration
of coastal area on the Upper Gulf of Thailand
Patcharaporn Yaowasooth Marine and Coastal Resources Research and Development Center, the Upper Gulf of Thailand, Muang District,
Samut Sakhon, Thailand Corresponding author e-mail: [email protected]
Abstract: Coastal erosion caused by anthropogenic, as well as natural processes in the coastal region of Thailand
and other countries is becoming a big issue at present days, because coastal erosion have negative impact on
human and coastal ecosystem. To prevent coastal erosion in the Upper Gulf of Thailand, the Department of Marine
and Coastal Resources (DMCR) carried out a project for the installation of bamboo structures and planting of trees
in the coastal area. At the end of the project, it has been found that the restoration of coastal ecosystem and
prevention from further coastal erosion were succeeded in some areas, on the other hand different result shown in
other areas. Based on the DMCR project outcome, this study analyzed the factors related to prevention of erosion
and the restoration of the coastal zone structure, in the Upper Gulf of Thailand. This study found that the success
of the project that is the prevention of erosion and restoration of the coastal area related with people’s participation,
strong leadership, government support and specification of bamboo. From focus group discussion, it was found
that strong policies for the protection and maintenance of installed bamboo structures and thereby appropriate
revision of policy are also an important factor for sustainability of the project. Moreover, devoted local leader,
and active committee members are needed to maintain, and monitor the project. This study results are applicable
for other coastal regions of different countries, with similar soil characteristics, to prevent erosion and restoration
of coastal areas. Keywords: Coastal erosion, bamboo structure, Upper Gulf of Thailand
Introduction
Thailand has a coastline measuring of 3,148 km, divided into the Gulf of Thailand with a length of 2,055
km and rest 1,093 km with the Andaman Sea. Coastal erosion mainly occurs along the coast of the Gulf of Thailand.
In seventeen provinces of Thailand 830 km along the coastline has been eroded, where 730 km belong to the Gulf
of Thailand and rest only 100 km coastline belong to Andaman Sea (Suraswadi, 2016). It also observed that the path
of erosion goes from the east to west coast of the Gulf of Thailand. Considering the erosion rate, some areas are declared
as critical area, where the erosion rate is over 5 meters per year, in other areas rate of erosion is 1-5 meters per year.
The Upper Gulf of Thailand, which started from the Bang Pa Kong River mouth to the Tha Chin River mouth
with a coastline of about 41.8 km has suffered the most severe erosion. From 2009, the rate of mangrove forest
deforestation in the Upper Gulf of Thailand is high and there has been a shift of livelihood along the coast to shrimp
farms. This trend increased the shrimp farms from 64,336 hectares to 237,220 hectares within the year of 2009 to
2016, it also damaged mangrove forest higher than 64 % (DMCR, 2011). Besides that, growing salt farms,
industry, agricultural and infrastructure make the deforestation problem more critical. The effect of coastal
erosion, Thamma-apipon, (2012) found coastal erosion affected the local communities in three aspects; there was
an economic impact, social impact, and environmental impact. Coastal erosion has effects to ecosystems and natural
resources. Mangrove forests, coral reefs, sea grass and land will be affected directly, ultimately, it affects the ecological
balance in the marine and coastal area. Coastal erosion affects the environment including; loss of land, mangrove
deterioration, marine animal reduction, loss of coastal scenery, adaption of house structure and prevention of erosion.
These directly affect the coastal community. Coastal erosion affected the economic on fisheries, aquaculture,
housing, tourism, and industry. After the effect of an economic system, people lose their resources, land, jobs, and
income etc. Tourism is the main source of income for Thailand. Besides that, industry of fisheries can create jobs
and increase income for the community as well as the country. Almost one-fourth of Thai people live in the 22
coastal provinces. That coastal erosion has an effect on the economic sector by various ways. It reduces the value
of land, results in income loss, and increases the retraction of the house and coastal defence structure costs. Dean
(2009) found that the cost of different erosion control structures is especially pertinent given the current recession.
The division of coastal management, in all states, reduces its budget by 7% (Gregson, 2009) and this reduction in
budget made the impact on enforcement and permitting personnel for local erosion control structures. And coastal
erosion has the impact on the social life of the people who live in coastal areas, and the number of affected people
are more than 12 million. Communities affected by erosion, lose their land and property, must change lifestyle by
migration. It is also the cause of changing the folkway of humans in coastal areas. Coastal communities migrated
to other areas which resulted in loss of life and cultural traditions of the communities; left some communities

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 149
homeless and resulted in the unavailability of work which normally has effects to other sectors of the economy.
Anxiety in a new career that may affect the state of mind and family relationships. Moreover, it is the cause of declining
living standards or not as good as the original. (DMCR, 2011B)
Community leader Khok Kham and Phanthai Norasing, Samut Sakhon Province of Thailand organized
a meeting with the local people for finding the solution of coastal erosion by bringing the folk wisdom. Local
fishermen are using bamboo for make fish trap and stable for shell aquatic. Fishermen noticed that after installation
of bamboo for aquaculture mussels, sediment accumulated near the area of bamboo streak. Observing that, they
installed bamboo streak adjacent each other without keeping any space between the streaks, which generate more
sediment accumulation easily and reduced the wave action. Local people used this technology to prevent coastal
erosion and increase the sediment along the coastline, and also planted mangrove tree to restore mangrove forest.
Coastal erosion issues are recognized to be solved urgently. The technology of using bamboo streak to
prevent erosion in the coastal areas by community are now proven. DMCR with community in coastal areas started
DMCR project to prevent coastal erosion by installation of bamboo structure, which reduced the weave energy,
generate sediment accumulate and helps for reforestation. The project started in 2011 on the Upper Gulf of
Thailand and includes eight areas: Khok Kham Moo 3, Khok Kham Moo 8, Bang Ya Praek Moo 6, Phanthai
Norasing Moo 6, Bang Krachao Moo 1 Sub-district, Samut Sakhron Province, Bang Kaeo Moo 10 Sub-district,
Samut Song Kram Province, Laem Fa Pha Moo 11 Sub-district, Samut Prakan Province and Song Khlong Moo 9
Sub-district, Chachoengsao Province. Communities with DMCR improved the design of the bamboo structure.
The summary of the design was bamboo size at least 3 inches width with 5 meters long, installed in 5 rows, depth
of about 2 meters emerged into the ground and 3 meters kept above from the ground, installed row alternating
zigzag design with the bamboo stick and at least 35 stick per 1meter bamboo structure combined with 5 row. The
ways for boat is 10 meters long and had a bamboo streak to cover the boat ways about 10 meters (Figure 2). This
project is a good example for prevent erosion, restoration of coastal area and local people are willing to prevent
erosion to make their life stable. In 2013 Intatha studied about factors influencing the capacity of communities to
respond to coastal erosion in the Upper Gulf of Thailand. This study shows. In the past most common method
used to protect the coast was rock placements (hard structure). But later soft structure such as bamboo structure
used to protect high coastal erosion. Moreover, the government and other stakeholders had applied a combination
of protection and restoration by installation bamboo structure and planting trees. This combined method shows
better results.
DMCR project found some areas were succeeded to prevent coastal erosion and restoration of coastal
area by using bamboo structure, but some areas were failed. This experience make an interest to researchers to
find the factors related to success of bamboo structure to prevent erosion and restoration of coastal area on the
Upper Gulf of Thailand and to determine the findings to be incorporated in existing guidelines and policies for
review as well as successful implementation by using bamboo structures to prevent erosion and restoration of
coastal areas. Besides that, this study results will be useful for policy modification and implementation to prevent
erosion and restoration of coastal area.
Materials and Methods
Study area
The study focused on three areas of the Upper Gulf of Thailand where DMCR project implemented. The areas
are; Bang Ya Phraek Sub-district, Samut Sakhon Province, Bang Kaeo Sub-district, Samut Songkhram Province,
and Song Khlong Sub-district, Chachoengsao Province, Thailand.
Data collection and sampling methods
Primary and secondary data both are collected for this research which is particularly relevant to the study.
Primary data was collect from questionnaires (Quantitative survey) a sample population of 333 people from 1973
people (National statistical office Thailand, 2016) as well as from focus group discussion (Qualitative survey).
This research conducted in coastal areas of Bang Ya Phraek sub-district, Samut Sakhon province, Bang Kaeo sub-
district, Samut Songkhram province, and Song Khlong sub-district, Chachoengsao province, of Thailand. Secondary
data was collected from various sources such as articles published in journals and books etc.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 150
Sample size: Theoretical aspects, formulas. Simplified formula for proportions (Taro Yamane, 1967)
𝑛 =𝑁
1+𝑁(𝑒)2
𝑛 =1973
1+1973(0.05)2
𝑛 = 333 people
n = the sample size
N = the population size
e= the acceptable sampling error
*95% confidence level and p = 0.5 are assume
Data analysis
Data analysis is one of the most sophisticated activities of the study to produce the correct results. In this
study, quantitative and qualitative data were used for analysis. One of the best software for analyzing statistical
data Statistical Package for Social Sciences (SPSS 15.0 for windows evaluation version) was used in this study.
Quantitative data analysis
The data collected through a personal interview by using pre-designed questionnaire and collected data
analyzed by SPSS. Inferential statistics and descriptive both types of data were collected for quantitative data
analysis. Collected data were coded, sorted, and categorized for analyzing by software. Analyzed data used to
prepare Multinominal logistic regression to analyze the factors related to a bamboo structure to prevent erosion
and restoration of coastal area on the Upper Gulf of Thailand.
Multinominal logistic regression
In logistic regression analysis, there is only one independent variable, the logistic regression model can
be written as
Pr𝑜𝑏 (𝑒𝑣𝑒𝑛𝑡) =𝑒𝛽0+𝛽1𝑋
1 + 𝑒−(𝛽0+𝛽1𝑋)
Or Pr𝑜𝑏 (𝑒𝑣𝑒𝑛𝑡) =1
1+𝑒−(𝛽0+𝛽1𝑋)
β0 and β1 Coefficients were estimated from the data
X is an independent variable
E is natural logarithm with a value of 2.718
From the above equation, we derive a new equation as
Pr𝑜𝑏 (𝑒𝑣𝑒𝑛𝑡) =𝑒𝑧
1 + 𝑒𝑧
Or Pr𝑜𝑏 (𝑒𝑣𝑒𝑛𝑡) =1𝑒𝑧
1 + 𝑒−𝑧
By Z= β0+β1X1+ β2X2+……………+ βpXp and the event not as incident from the equation.
Prob (no event) = 1-Prob (event)
Qualitative data analysis
Focus group discussion and interview with community leader was done to accumulate qualitative data.
In focus group, discussion representative from local dwellers, local government officer, and a representative from
different occupation of that area were confirmed.
Result & Discussion
Socio-economic characteristics of the respondents The number of population in the study areas were 1,973 people. For personal interview 333 respondents
were chosen randomly. Almost 90% of respondents were local dwellers and their life is dependent on marine and
coastal ecosystem for livelihoods. Most of the respondents age limit were in the range of 41-60 years. The main
occupation of the dwellers in the study area was fishery and their education level was primary 71%, Major part of
the respondents 66%, monthly income was in the class of $142- $425 (5,001 – 15,000 Baht) the result is similar
to Thai people’s average income 3,976-12,275 Baht (World Bank, 2016).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 151
Awareness about coastal erosion issue of the respondents The study found that most of the respondents in the study area were aware of coastal erosion, and the value
was higher than 85%. The awareness of respondents about social (migration, change occupation etc.), economic
(loss of income, loss of land, spend money for protecting land and house etc.), and environmental (loss of mangrove
forest, coastal resources degraded etc.) effect due to erosion also analyzed in the study. The results of the study
showed that the respondents are firstly conscious about the social effect, then secondly the economic effect, and
thirdly the environmental effect.
Respondent’s opinion about the impact of bamboo structures About the impact of bamboo structures in preventing erosion and restoration of coastal areas as well as
in socio-economic factors a significant portion of the respondents had a positive view. 34% respondents thought
the impact of bamboo structures on social benefit is high, where 37% respondents thought the impact bamboo
structures of economic benefit is high, and highest 63% respondents thought bamboo structures have high positive
impact on environmental benefit. On the other hand, for social benefits perspective, 39% respondents thought the
impact of bamboo structure in social benefit is low, where 38% and 25% respondents thought the impact of bamboo
structure in economic and environmental benefit is low respectively.
Respondent’s opinion about factors of success Budget
Budget is the main driving force in success to prevent erosion and restoration of the coastal area.
Respondent’s first preference was the government fund collaboration with the community. Their second choice
was NGO fund collaboration with community and the third was only community funded project.
Participation of stakeholders For success of the project, participation from different stakeholders is another important factor. Among
all stakeholders, respondent's first priority was participation of leader, the second priority was participation of the
community, and third was government official participation.
Miscellaneous factors Comparing various factors together, the study results found shows that specification of bamboo (type, species,
age, and thickness etc.) was the first preference from the respondent. Environmental (area, wave, and wind etc.)
condition was in the second position and in the third position the species of tree used for restoration of the coastal area.
Respondent’s opinion about policy and regulations For management of coastal area "The strategies for erosion control" policy of Thailand was established
in 2012. However, the contents of the policy is not enough to describe the management of bamboo structure for
erosion control in coastal areas. This study found that most of the respondent’s suggestion is to modify the current
policy without delay for providing more emphasis on using bamboo structure to prevent erosion and restoration
of coastal area. Moreover, almost 65% respondents suggested that policy should be modified after discussion with
the community of the coastal area, as well as policy should be modified as per suggestion of the community. For
the success of the policy implementation, 43% respondents recommended that collaboration of government and
community will be the main factor.
Participation of the Respondents in the Project Considering the full study area, the study found that participation in the project activity of respondents
was significantly high. 75% of the respondents were present in clarification meeting about use of bamboo structure
to prevent erosion and restoration of the coastal area. On the other hand, considering individual study area
participation rate was different, and values were 92%, 65% and 46% in Bang Ya Phreak, Bang Kaeo and Song
Khlong sub-district respectively. Similar results found for installation of bamboo structures and planting mangrove trees. 56% respondents
participated in the installation of bamboo structures and planting mangrove trees. But for individual area,
participation rate was 79%, 45%, and 17% in Bang Ya Phreak, Bang Kaeo and Song Khlong Sub-district respectively. Participation for monitoring after installation and planting for full study area was 50%. Where, for individual
study area participation values were 63%, 49% and 27% in Bang Ya Phreak, Bang Kaeo and Song Khlong Sub-district
respectively. 77% of all respondents, recommended that the maintenance activity should be continued after installation
of bamboo structures, and planting mangrove trees to prevent erosion and to restore the coastal area. Besides that,
respondents demand to continue maintenance activities were recorded at 94%, 71%, and 49% in Bang Ya Phreak,
Bang Kaeo and Song Khlong Sub-district respectively. Data related to participation in project activity shows, in Khlong Sub-district the participation rate was
very low compared with Bang Ya Phreak and Bang Kaeo Sub-district.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 152
Statistical analysis 33 factors were included in the questionnaire survey to find the most effective factors for using bamboo
structures to prevent coastal erosion and restoration of the study area. The collected data was analyzed by
Multinominal regression model of SPSS program. The output shows the factors that are significant with project success.
In this study, failed and partially successful area’s data was analyzed through comparing the data of successful area. Multinominal logistic analysis showed among the 33 factors 4 factors were highly significance for project
success and the significance results between project success and these four factors at 0.01 significance level (Table 1).
Four significant factors are leadership, participation in project activities (includes meeting, installation of bamboo
structures, planting trees, monitoring and maintenance), government support, and the specification of bamboo.
It indicates, to make a project success for prevent coastal erosion and restoration of coastal area strong leadership,
active participation of the beneficiary in project activity, government positive support, and maintain the quality of
bamboo by following the specification is essential.
Project leader’s opinion The researcher also carried out personal interviews with the project leader of the 3 study area. All leaders
came from the local community, but they have different views and different level of involvement in the project activity.
Bang Ya Phreak Leader’s opinion Leader of the Bang Ya Phreak was the initiator of using bamboo structures to protect coastal erosion in
his area. Bang Ya Phreak area facing the coastal erosion problem for a long period of time, and it destroyed their
land and property. Rock protection work, for controlling erosion, was not working properly in that area. Besides
that, rock protection created more erosion to the adjacent area after construction. Another problem was the very
clayey soil of Bang Ya Phreak area, which are not suitable for rock protection. At that time, he observed the
success of protecting coastal erosion in Khok Kham Sub-district by using bamboo structures. This observation
encouraged him to initiate bamboo structure installation in Bang Ya Phreak area to prevent erosion, and restores
more area by increasing sedimentation. To start that activity, he spend his personal income, as well as convinced
the community to organize a fund for installing bamboo structure to prevent erosion. He identified erosion as an important issue for coastal ecosystem. He address erosion issues in the order
of the economic effect, the social effect and the environmental effect. He has been highly involved with these
activities for more than 11 years. The people who were not capable or had no interest in bamboo installation
activity, he involved them in other supporting works like establishing a mangrove tree nursery by donating mangrove
trees for restoration of the coastal area. He also invited the children in the area and took them to learn folkways
and planting of mangrove trees. Moreover, he communicated with the universities, colleges, and schools to send
their students for participating in bamboo installation and mangrove tree planting. Among the factors related with the success of the prevention of erosion and restoration by using bamboo
structures, he prioritized community participation with self-willingness is the most important factor for success,
followed by the specification of bamboo and lastly, the involvement of stakeholders from outside of community
such as government, NGOs for support knowledge and budget. His opinion about the future of the project was
that, these activities should be continued to prevent erosion and restorations of the coastal area. Restore land and
mangrove forest will enhance the aquatic life as well as natural resources of the area. Proper and sustainable use
of those resources in the future, will enhance the community people's quality of life and help them to enjoy a
comfortable and happier life.
Bang Kaeo Leader’s opinion The project starting story was in Bang Kaeo area almost the same to Bang Ya Phreak area. Bang Kaeo
area also suffers coastal erosion for many years. The leader of Khok Kham project area known about the problem
of Bang Kaeo. The leader with people from his community visit Khok Kham. After visiting, Bang Kaeo leader
realized that, to prevent coastal erosion and restoration installation of bamboo structures should be start in Bang
Kaeo area. To implement the same project, he talked with the local community of Bang Kaeo area, but unfortunately
most of them were not agreed to do. At that stage, he decided to start the project by spending his own money for
set an example in front of local community with the help of agreed people. He was highly involved for more than 11 years with these projects as well as highly aware about coastal
erosion issue and the importance of the coastal ecosystem. He chose to address these issues in the following order:
social effect, economic effect and finally the environmental effect. He also communicated with some NGOs and
organizations that could provide funds. He found that, if they can show some success NGOs and other organizations
would support them because of their corporate social responsibility (CSR). He discussed all the findings with the
community and finally, the community agreed to start to install bamboo structures to prevent coastal erosion and
restoration of the area. The leader of Bang Kaeo had a view that, the factors related with the success to prevent erosion and
restoration by using bamboo structure are interconnected. He prioritized community participation are the most
important factor for success. Quality of bamboo and monitoring the repair work were his second priority. His third
priority was the approach of leader. He believes that a leader should maintain the approach of ‘not give up on a task

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 153
until you succeed’. His opinion about the future of the project was that, these activities should be continued for a better
future of the community. He believes that this project will be the best example for green technology to prevent
coastal erosion and restoration. Besides that, from the success of the project, future generations will get a better
quality life.
Song Khlong leader opinion The project opening story in Song Khlong area was different from the other two study areas. Before starting
the project activities, officers of the project organized a meeting with the leader and the community of Song Khlong
area. In the meeting the number of participants from the community was low. After that, officers arranged more
meetings with the leader and gave him instructions to ensure the participation of the community in the project activities.
However, later on, without ensuring community participation, project officers installed bamboo structures with
the help of laborers. Due to the lack of monitoring, after a few years they found all the bamboo structures were damaged. He was involved in 5 years in project activities, He considered economic effect is the first, social effect
is the second, and finally the environmental effect is the third important issue for erosion in the coastal ecosystem.
He believes that, in Song Khlong area, rock structures will be effective for preventing coastal erosion instead of
bamboo structure, but for restoration of that area planting mangrove trees will be the good solution. He also
suggested that before starting the project, the government should make some focused group discussions with the community
to check the willingness of the people to participate in the project. Among the various factors, the participation of the community was chosen by him as the most important
factor for successful prevention of coastal erosion and restoration of the coastal area by using bamboo structures.
The natural environment in the coastal area (strong winds and high sea waves, soil characteristics, etc.) was
selected by his as the next important parameter for success of the project. Beside these, he recognized specification
of bamboo, and policies of the government are also important factors for success of the project.
Focus group discussion In focus group discussion, the main topic was "Guidelines for policy modification on use of bamboo
structures to prevent coastal erosion and restoration of the coastal areas successfully by the community ". The guidelines
for policy modification and implementation formulated by the focused group discussions were community involvement
should be the first priority at the design stage of the project. Participation of local community should be ensured
from the beginning of the project and continue until the end. At the start, this includes looking for the project site or
area, planning for the project, analysis of the problem in the area, evaluation of the damaged resources. During
implementation stage, it includes installation of the structures and restoration activities, monitoring activities, and finally
evaluation of the project. Sustainability of the structure depends on regular monitoring and maintenance, for this reason, sufficient
budget have to insure for maintenance program. Knowledge should be shared with the community people as well
as the officers of the project. Besides this, training about implementation, monitoring, and maintenance activities,
as well as visits in success areas to get acquainted with their experiences should be organized. Ownership of the newly restored areas should be belongs to the government. Restored area should not
be sold or rented by the government. Moreover, the government must include instructions on the policy about the
use of restored lands for community as public goods with sustainable resource management. The policy should
emphasize for increase awareness about economic benefits, environmental benefits, and social benefits of the
community by using bamboo structures for prevention and restoration of the coastal area. Penalty impose methodology to a person or a group who destroy the bamboo structures or hampers project
activities should also be included in policy. To select the leader with high morality and awareness on environmental,
policy should provide a proper guideline through community discussions.
Conclusions
The Upper Gulf of Thailand facing severe coastal erosion because of natural process and human
activities. The DMCR has the project to prevent and restoration of the coastal area on the Upper Gulf of Thailand
by using bamboo structures. The bamboo structures suitable sediment characteristics in that areas. Sediment
characteristic on the Upper Gulf of Thailand is sandy clay loam (Sawangarreruks at all, 2013). After implementation,
DMCR found the bamboo structures can prevention of erosion were succeed in many areas, but in some areas it
failed. This study was done to find the factors related to the success of using a bamboo structure for preventing
erosion and restoration of coastal area in the Upper Gulf of Thailand. A pre-designed questionnaire survey, consisting of 33 factors was done to collect data from 333 respondents
about the success of the project using the bamboo structure for preventing erosion and restoration of coastal area.
The data was analyzed by Multinominal regression model of SPSS program and found that among the thirty-three
factors, four factors were highly significant with the significant value at 0.01 and had relation to the success of
DMCR project. Four significant factors for success of the project were people’s participation in the project
(installation of bamboo, planting trees, meetings, and maintenance), strong leadership, government support for various

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 154
activities, and the specification of bamboo. The result from questionnaire survey shows that 75% of total respondents
participated in the project activity, where the percentage of participation was higher than 90% in the successful
area. Strong leadership is essential for project success was chosen by 60% of the respondents. Where government
support on different activities is an essential factor for success was chosen by 84% of the respondents. Comparing with
various quality issues and environment factors, specification of the bamboo was chosen by 90% of the respondents. (Appendices) Three focus group discussions were carried out in the study area to get some guidelines for policy
modification and implementation about using bamboo structures to prevent erosion and restoration of coastal
areas. The findings from the focus group discussion were, policy should be more specific and details about using
bamboo structure and current policy should be modified without delay for successful implementation of the
project. Penalty impose methodology should be included in the policy for destroying the bamboo structures and
hampering the project activity. Moreover, leader with high morality should be chosen by the community.
Policy recommendations 1. Do not carry out activities or do any actions that may result in or cause problems in increasing coastal
erosion.
2. Establish rules for the construction process to prevent coastal erosion.
3. Continuing the project to prevent coastal erosion in the area.
Acknowledgement
Above all, I would like to profoundly thank the Korea Environment and Technology Institute (KEITI)
of the Government of the Republic of Korea for tuition and subsistence allowances. Sincere thanks to the Ministry
of Natural Resources and Environment (MNRE) of Thailand for granting me this opportunity to acquire more
knowledge in my field of interest and for their cooperation. I am grateful and very thankful to my advisor,
Professor Jin-Young Jung Ph.D., and Dusit Wechakit Ph.D. from suggestions, teachings, imparting of knowledge
and data to the unconditional contribution directives that greatly improved this paper. I would like to extend my
gratitude to Professor Yohan Lee Ph.D. and Ferdous Ahmed for their assistance to improve this paper, the support,
encouragement and care rendered to me.
References
Department of marine and coastal. 2011. “Prevention of coastal erosion in Thailand”. Strategic Management
Prevention of coastal erosion. Marine and Coastal Resources Journal, 20 December 2018. 70 – 78.
Department of marine and coastal. 2011. Monitoring and evaluation result of monitoring and evaluation the
restoration of coastal ecosystems. By planting bamboo slow wave Samut Prakan and Chachoengsao
province case study. Chulalongkorn University. Bangkok: GRT.
Department of marine and coastal. 2013. “Coastal areas of Thailand”. Accessed February 25. 2016.
http://marinegiscenter.dmcr.go.th/km/coastal-zone-management01 /#.V12 fs 7t97IU.
Department of marine and coastal 2015. Bamboo slow wave manual to prevent coastal erosion. DMCR, Bangkok: GRT.
Gregson, J. 2009. Executive Secretary notes. Paper presented at the NC Coastal Resources Committee Meeting
February 2009, Morehead City, NC.
Intatha, C. 2013. Factors influencing the capacity of communities to respond to coastal erosion in the Upper Gulf
of Thailand. Mahidol University, Thailand. National statistical office Thailand. 2016. “Number of
population”. Accessed May 8. 2016. http://service.nso.go.th/nso/nsopublish/districtList/page1.htm
Statistics Solutions. 2016. “Multinomial Logistic Regression.” Accessed December 23. 2016.
http://www.statisticssolutions.com/mlr/.
Suchart Sawangarreruks. S., lertkasetvittaya. N., Yaowasooth. P., luangkhamin. C., Passada and Nikomjit. S.
2013. “After effect of flooding on the Macro benthic community in the Upper Gulf of Thailand.” Key
challenge of marine Biodiversity Conservation in our Changing Climate 9-11 September 2013,
Department of Marine and Coastal Resources Research and Development Center Upper Gulf of Thailand.
Suraswadi Pinsak. 2016. “The new coastal resources law, Marine protected areas and nature-based solutions for
coastal erosion in Thailand.” Accessed December 20. 2018.
https://www.iucn.org/sites/dev/files/import/downloads/1431_1443_the_new_coastal_resources_law__
marine_protected.pdf
Thama-apipon Sanawaya. 2012. Studied the effects of the problems of coastal erosion on coastal communities.
Silpakorn University, Thailand.
World Bank. 2016. “Thai people average income.” Accessed December 20. 2018.
http://www.worldbank.org/th/news/press-release/2011/08/02/thailand-now-upper-middle-income-economy.
Yamane, Taro. 1967. Statistics: An Introductory Analysis, 2nd Ed., New York: Harper and Row.
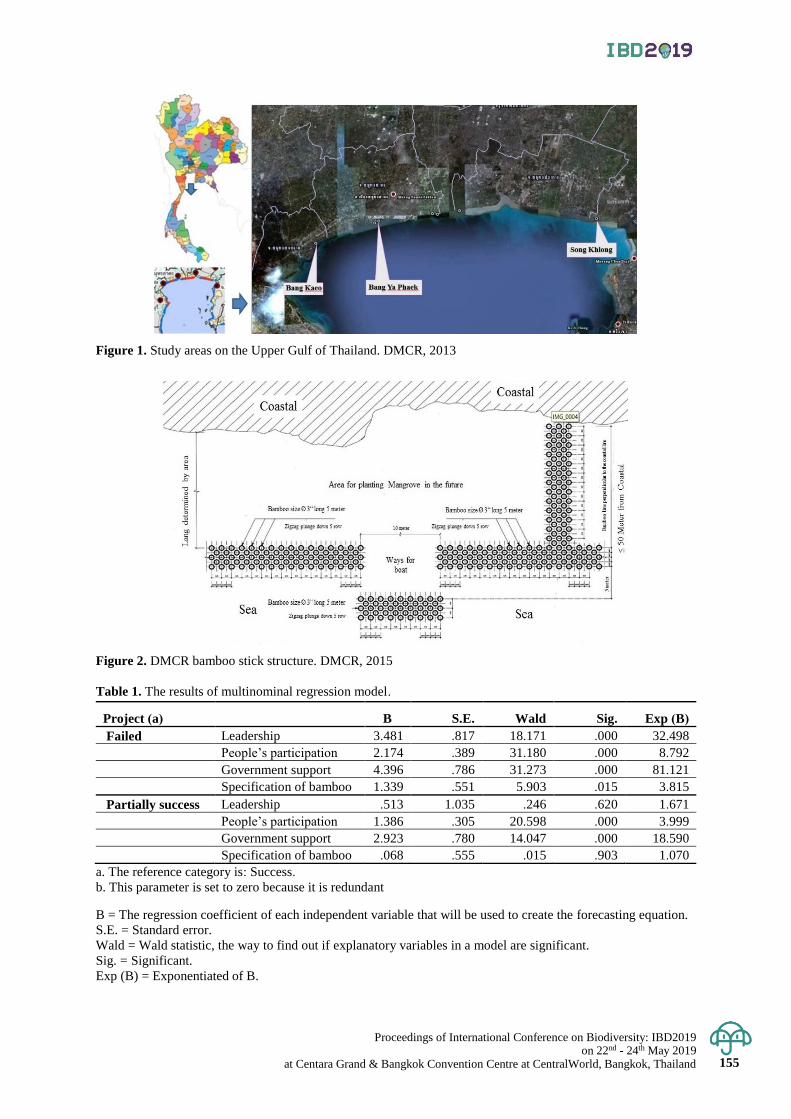
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 155
Figure 1. Study areas on the Upper Gulf of Thailand. DMCR, 2013
Figure 2. DMCR bamboo stick structure. DMCR, 2015
Table 1. The results of multinominal regression model.
Project (a) B S.E. Wald Sig. Exp (B)
Failed Leadership 3.481 .817 18.171 .000 32.498
People’s participation 2.174 .389 31.180 .000 8.792
Government support 4.396 .786 31.273 .000 81.121
Specification of bamboo 1.339 .551 5.903 .015 3.815
Partially success Leadership .513 1.035 .246 .620 1.671
People’s participation 1.386 .305 20.598 .000 3.999
Government support 2.923 .780 14.047 .000 18.590
Specification of bamboo .068 .555 .015 .903 1.070
a. The reference category is: Success.
b. This parameter is set to zero because it is redundant
B = The regression coefficient of each independent variable that will be used to create the forecasting equation.
S.E. = Standard error.
Wald = Wald statistic, the way to find out if explanatory variables in a model are significant.
Sig. = Significant.
Exp (B) = Exponentiated of B.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 156
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 156 - 162
Exploring the suitability map of wild banana (Musa serpentina Swangpol & Somana)
in Thailand using species distribution models with the limited occurrence data
Thanayut Changruenngam and Jantrararuk Tovaranonte* School of Science, Mae Fah Luang University, Muang District, Chiang Rai, Thailand
*Corresponding author e-mail: [email protected]
Abstract: Species distribution model (SDMs) is one of the powerful tools to predict the suitability map to address
the ecology and conservation approaches. However, the limited number of occurrence data has been a problem in
model performance. Here, the three algorithms (MaxEnt, generalized linear model: GLM, and random Forest: RF)
were selected to project the suitable map with eight occurrences of Musa serpentina Swangpol & Somana wild
banana which endemic to the west of northern and central Thailand and eleven environmental variables. Due to a limitation
of occurrence data, the fuzzy logic had been applied to GLM and RF model to enhance the occurrence data.
MaxEnt is presence-only data, it could not use the fuzzy logic to create pseudo absence data. The results showed
that three climatic variables had affected all three models especially precipitation of warmest quarter (BIO8) and
precipitation of coldest quarter (BI019). All algorithms could predict the suitable map well with high AUC values
(> 0.9). GLM had the highest performance with AUC value of 0.991.
Keywords: MaxEnt, Random forest, GLM, Fuzzy logic, Musa serpentine
Introduction
Species distribution models (SDMs) play an important role for conservation of endangered species. It can
predict future results under climate change and generate species distribution models under geospatially explicit
layers of abiotic or biotic data which defines the ecological requirements of species under study (Franklin, 2010).
SDMs consist of 2 categories according to the characteristics of data. First, presence data-only algorithms such as
genetic algorithm for ruleset prediction (GARP) and maximum entropy (MaxEnt) (Phillips, 2008). MaxEnt is
commonly shown accurate prediction capabilities of these models. Second, binary presence/absence data
algorithms such as classical modelling approach: the generalized linear model (GLM) and modern method:
random forest (RF) and boosted regression trees (BRT). From previous studies, consideration of the actual values
of the predictions which emphasize more clearly found that MaxEnt, BRT and GLM performed well, followed by
RF then GARP (Elith & Graham, 2009). The applications of SDMs are widely available, such as predicting
distribution of disease vectors. (Khatchikian et al., 2011). The other applications include conservation of the
suitable habitat of species (Tovaranonte et al., 2015) and the evaluation of species vulnerability to climate change
(Trisurat, Shrestha, & Kjelgren, 2011).
Musa serpentina Swangpol & Somana (‘Nakkharat’ or ‘Naga’) is a rare wild banana in Musaceae. It is
endemic to the west of northern and central Thailand such as Tak, Mae Hong Son and Kanchanaburi Provinces.
It commonly grows in the open mixed deciduous forest by stream banks or low slopes by roadsides. The altitude
of habitat is approximately 240-570 meters above sea level. Due to a small seed set and drastic fragmentation, its
conservation status is considered as endangered species based on IUCN Red List Categories and Criteria: Version 3.1
(Criteria D in Section V; IUCN, 2001)(Swangpol & Somana, 2011).
SDMs should provide conservation practitioners with the estimated spatial distributions of species
requiring attention. These species are often rare and have a small sample size, posing challenges for creating
accurate species distribution models. The studies indicated that the number of known occurrences had greatly
affected the accuracy of SDMs whereas MaxEnt always had the highest accurate predictive ability (Hernandez,
Graham, Master, & Albert, 2006). Wisz et al. (2008) compared AUC form 14 SDMs and found MaxEnt and GLM
had higher performance. GLM gave the highest AUC compared to RF and MaxEnt in different species distribution
(Elith & Graham, 2009). However, RF algorithm can also generate better species predictive distributions for a
habitat prediction of small sample size. while MaxEnt had lower performance (Mi, Huettmann, Guo, Han, & Wen,
2017). Since the sample size of Musa serpentina is small. Hence, the suitability map of Musa serpentine should
be predicted by MaxEnt, RF and GLM algorithm.
GLM and RF are algorithms for binary presence/absence data. The methods of pseudo-absence selection
influence a performance of species distribution models which is binary presence/absence data (Mary S. Wisz &
Guisan, 2009). The pseudo-absence data was established by using opposite condition with environment of Musa
serpentina. Generally, the boundaries of suitable environment cannot be clearly defined. Therefore fuzzy logic is
a tool for increasing flexibility of boundaries of suitable environment. It is used to handle the concept of partial
truth where the true values may range between completely true and completely false that is these values of variables

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 157
are any real numbers in closed interval 0 to 1 (0 = false, 1= truth). These are a feature of the membership function
of fuzzy set.
Here, this study aimed to compare the predicted performance between GLM and RF which developed
by Fuzzy pseudo-absence data with MaxEnt for the small sample size condition. And there is establish the
suitability map for the conservation of the habitat of Musa serpentina.
Materials and Methods
Study area and the occurrence data
Thailand is located between 5°45′ and 20°30′N and 97°30′ and 105°45′E. The total area is 511,731 square
kilometers or around 200,000 square miles. There are eight localities of Musa serpentina were obtained from
published data (Swangpol & Somana, 2011), herbarium data and unpublished data
Figure 1. (a). Distribution map of Musa serpentina in the west side of northern and central regions of Thailand
(Swangpol & Somana, 2011) and (b). suitability map of Musa serpentina overlaid by the area of Band 4 and 5 of three models
Environmental variables
Twenty environmental variables were used at 30 seconds (1km2) resolution. Twenty climate variables
were downloaded from the WorldClim website (http://www.worldclim.org, 11th March 2017, Hijmans et al., 2005).
These variables are BIO1 (annual mean temperature), BIO2 (mean diurnal range), BIO3 (Isothermality), BIO4
(temperature seasonality), BIO5(Max temperature of warmest month), BIO6 (min temperature of coldest Month),
BIO7 (temperature annual range), BIO8 (mean temperature of wettest quarter), BIO9 (mean temperature of driest
quarter), BIO10 (mean temperature of warmest quarter), BIO11 (mean temperature of coldest quarter), BIO12
(annual precipitation) BIO13 (precipitation of wettest month), BIO14 (precipitation of driest month), BIO15
(precipitation seasonality), BIO16 (precipitation of wettest quarter), BIO17 (precipitation of driest quarter),
BIO18 (precipitation of warmest quarter), BIO19 (precipitation of coldest quarter) and Alt (altitude).
Fuzzy Logic
Fuzzy Logic is a computer system that works by fuzzy set invented by Zadeh (Zadeh, 1965). It is used
to handle the concept of partial truth where the truth values may range between completely true and completely
false (0 = false, 1= truth). These are a feature of the membership function of fuzzy set. It was created by the degree
of truth as an extension of valuation. The membership functions were used as input and output variables of the
rule-based system (IF-THEN rules) derived from knowledge. Fuzzy logic was used to create pseudo-absence data
for identifying suitable habitat of Musa serpentine. After that, areas with probability level of less 0.6 as area for
1,000 randomly sampled was used as pseudo-absence data or Fuzzy pseudo-absence data.
Fuzzy pseudo-absence data
A suitable habitat of Musa serpentina is in highly disturbed habitats of open mixed deciduous forest by
stream banks or low slopes by roadsides and altitude 240–570 meters (Swangpol & Somana, 2011). The open
mixed deciduous forest often found at height of 50-800 meters, rainfall range from 1,200-1,600 mm per year and
not found in the South of Thailand (Marod, Pinyo, Duengkae, & Hiroshi, 2010). The presence data and absence
data were defined as 1 and 0 respectively. A membership function of fuzzy logic was created with Triangular
function and the rule-based system as follows.
Input
Altitude (0 -1): low (less 300 m), appropriate (200 - 600 m), high (more than 500)
Rainfall (0-1): low (less 1300 mm), appropriate (1,100 - 1,700 mm), high (more than 1500 mm)
Legend
") serpentina
1
2
3
4
5
b a

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 158
Output
Minimum of the Fuzzy membership from the input (0 - 1)
Rule-based system 1. If altitude = low then 0
2. If altitude = High and then 0
3. If altitude = appropriate and rainfall = low then 1
4. If altitude = appropriate and rainfall = appropriate then 1
5. If altitude = appropriate and rainfall = high then 0
The map outputs were overlaid type "and" are shown in Figure 2. We used areas with a level of less 0.6
as an area for 1,000 randomly sampled to use as pseudo-absence data (Morgane, Frederic, Jiguet, Cecile Helene,
& Wilfried, 2013).
Figure 2. The fuzzy map in a fuzzy logic system to create pseudo-absence data from (a) altitude data, (b) rainfall
data and (c) The overlaid map
Modelling and evaluation methods
The program named MaxEnt version. 3.2.1 was used to project eight samples and 1000 background
samples were created randomly. The function random forest was used to build an ensemble of classification trees.
The model generated 500 trees from both binary presence data and Fuzzy pseudo-absence data. We used function
GLM to create all possible subsets of models with the options for each variable from presence data and Fuzzy
pseudo-absence data. Area Under the ROC Curve (AUC) was used to evaluate the performance of the models
(Liu, White, & Newell, 2011). The AUC is the current best practice for evaluating the success of a model for
binary presence/absence data (Rushton et.al, 2004). Therefore, the value of AUC is an efficient tool for testing
the performance of SDMs. For example, in 2013, the value of AUC was used to evaluate the distribution model
of the particle to find the most effective model (Tovaranonte et al., 2015). The criteria for the AUC values that
are frequently seen are 0.90-1.00: excellent, 0.80-0.90: good, 0.70-0.80: fair, 0.60-0.70: bad and 0.50-0.60: failed
(Swets, 2015). In addition, each model different in prediction which can be observed from the Pearson product-
moment correlation coefficient and percentage of duplicate area. Summarizing the modelling and evaluation
methods are shown in Figure 3.
a b c
Probability
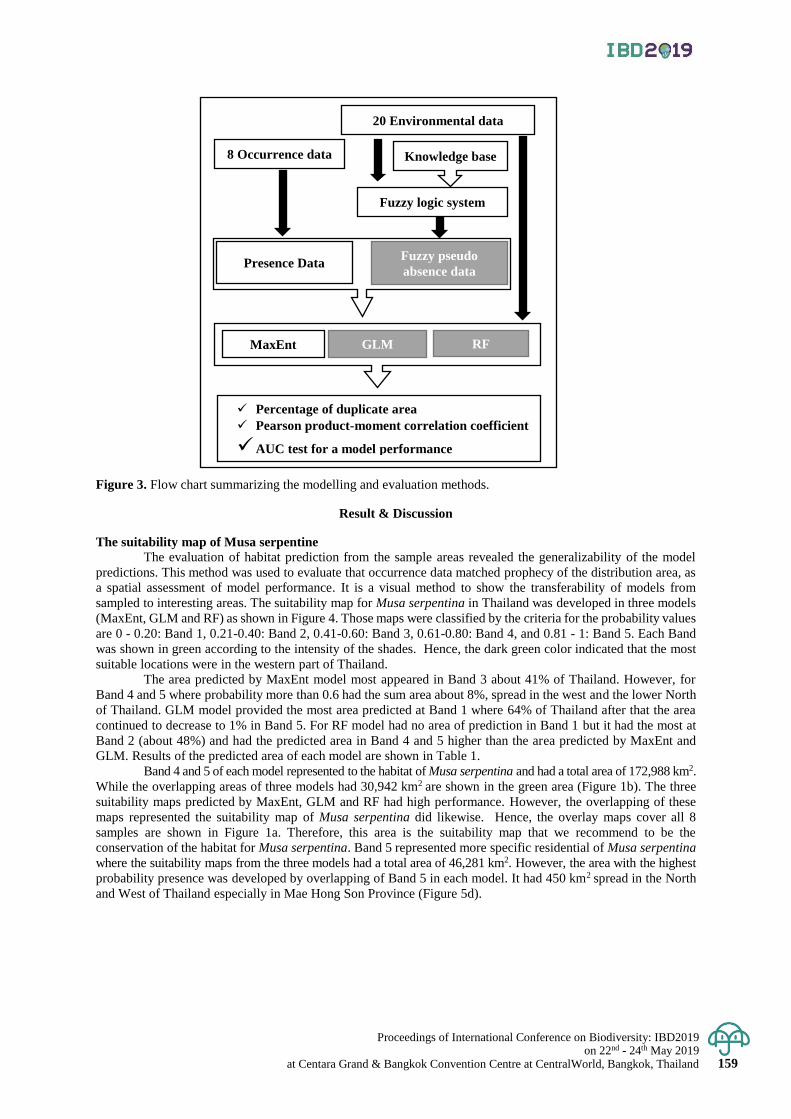
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 159
Figure 3. Flow chart summarizing the modelling and evaluation methods.
Result & Discussion
The suitability map of Musa serpentine
The evaluation of habitat prediction from the sample areas revealed the generalizability of the model
predictions. This method was used to evaluate that occurrence data matched prophecy of the distribution area, as
a spatial assessment of model performance. It is a visual method to show the transferability of models from
sampled to interesting areas. The suitability map for Musa serpentina in Thailand was developed in three models
(MaxEnt, GLM and RF) as shown in Figure 4. Those maps were classified by the criteria for the probability values
are 0 - 0.20: Band 1, 0.21-0.40: Band 2, 0.41-0.60: Band 3, 0.61-0.80: Band 4, and 0.81 - 1: Band 5. Each Band
was shown in green according to the intensity of the shades. Hence, the dark green color indicated that the most
suitable locations were in the western part of Thailand.
The area predicted by MaxEnt model most appeared in Band 3 about 41% of Thailand. However, for
Band 4 and 5 where probability more than 0.6 had the sum area about 8%, spread in the west and the lower North
of Thailand. GLM model provided the most area predicted at Band 1 where 64% of Thailand after that the area
continued to decrease to 1% in Band 5. For RF model had no area of prediction in Band 1 but it had the most at
Band 2 (about 48%) and had the predicted area in Band 4 and 5 higher than the area predicted by MaxEnt and
GLM. Results of the predicted area of each model are shown in Table 1.
Band 4 and 5 of each model represented to the habitat of Musa serpentina and had a total area of 172,988 km2.
While the overlapping areas of three models had 30,942 km2 are shown in the green area (Figure 1b). The three
suitability maps predicted by MaxEnt, GLM and RF had high performance. However, the overlapping of these
maps represented the suitability map of Musa serpentina did likewise. Hence, the overlay maps cover all 8
samples are shown in Figure 1a. Therefore, this area is the suitability map that we recommend to be the
conservation of the habitat for Musa serpentina. Band 5 represented more specific residential of Musa serpentina
where the suitability maps from the three models had a total area of 46,281 km2. However, the area with the highest
probability presence was developed by overlapping of Band 5 in each model. It had 450 km2 spread in the North
and West of Thailand especially in Mae Hong Son Province (Figure 5d).
Fuzzy logic system
20 Environmental data
8 Occurrence data Knowledge base
Fuzzy pseudo
absence data Presence Data
MaxEnt GLM RF
Percentage of duplicate area
Pearson product-moment correlation coefficient
AUC test for a model performance

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 160
Figure 4. Suitability map of Musa serpentina in Thailand created from (a) MAXENT, (b) Random Forest and (c)
GLM.
Species distribution models MaxEnt, GLM and RF had high performance for creating the suitability map of Musa serpentina. Those
maps can provide conservation practitioners to estimate the accurate spatial distributions of Musa serpentina.
However, each algorithm had estimated in difference size of the area in the map. Therefore, the percentage of
duplicate area, Pearson product-moment correlation coefficient and AUC from each model were compared.
Figure 5. The same area predicted from MaxEnt, RF and GLM (a) Band 2, (b) Band 3, (c) Band 4, (d) Band 5
Percentage of the duplicate area represented the prediction area of each algorithm in the same band. However, the
predicted size of areas from three models were different. Band 1 had no duplicate area as the RF's map had a
probability of not less than 0.2. Range 0.21 - 0.40 (Band 2) had duplicated area for 52% which is close to GLM's
predicted area (Figure 5a) and Band 3 had 27% of duplicated area (Figure 5b). Band 4 and Band 5 were used to define
the suitability habitat because of the probability > 0.6. However, the predicted area of each model is different.
Therefore, predicted for the same area of Band 4 found 13 % (Figure 5c). Band 5 is the model's predicted area that
has the highest probability (>0.80), represented to the suitability map of Musa serpentina where all models shown
a duplicate area of 9 %.
Table 1. Percentage of predicting area by MAXENT, GLM, RF of Musa serpentine in Thailand.
Band Probability
Model’s predicted area of
Musa serpentine in Thailand (%) Percentage of duplicate
area (%) MAXENT GLM RF
1 0.00 – 0.20 28 64 0 0
2 0.21 – 0.40 24 24 48 52
3 0.41 – 0.60 41 6 25 27
4 0.61 – 0.80 7 4 21 13
5 0.81 – 1.00 0.06 1 7 9
The r-value of MaxEnt and GLM had a strong positive relationship (0.564) but the relationship between
RF with the remaining two models had a moderate positive relationship (Table 2). However, the reliability of
the correlation coefficient depends on hypothesis test of the significance of the correlation coefficient where H0:
ρ = 0, H1: ρ ≠ 0 represented no significant correlation exists and a significant correlation exists. Testing
the significance of the correlation coefficient of every pair of model's predicted maps had p-value ≤ 0.001 (Table 2).
Therefore, there is sufficient evidence to conclude that there is a significant linear positive relationship between
a
Legend
0-0.20
0.21-0.40
0.41-0.60
0.61-0.80
0.81-1
b c
Probability
b c d a
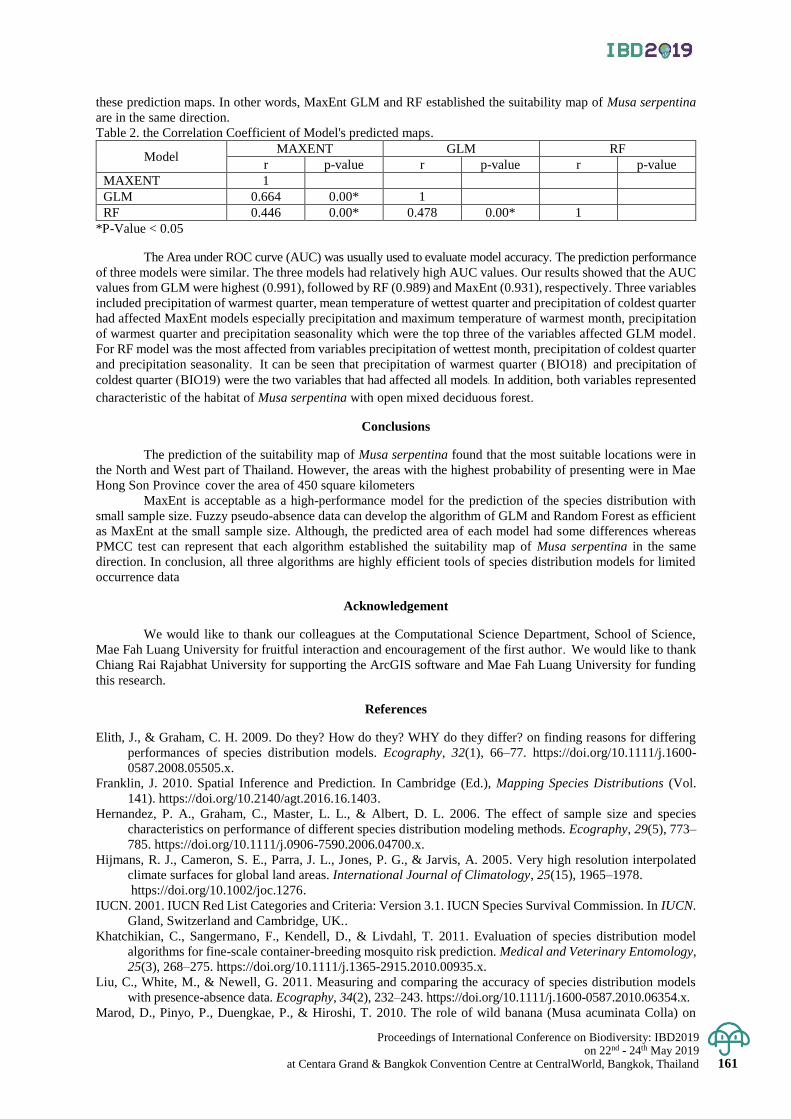
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 161
these prediction maps. In other words, MaxEnt GLM and RF established the suitability map of Musa serpentina
are in the same direction.
Table 2. the Correlation Coefficient of Model's predicted maps.
Model MAXENT GLM RF
r p-value r p-value r p-value
MAXENT 1
GLM 0.664 0.00* 1
RF 0.446 0.00* 0.478 0.00* 1
*P-Value < 0.05
The Area under ROC curve (AUC) was usually used to evaluate model accuracy. The prediction performance
of three models were similar. The three models had relatively high AUC values. Our results showed that the AUC
values from GLM were highest (0.991), followed by RF (0.989) and MaxEnt (0.931), respectively. Three variables included precipitation of warmest quarter, mean temperature of wettest quarter and precipitation of coldest quarter
had affected MaxEnt models especially precipitation and maximum temperature of warmest month, precipitation
of warmest quarter and precipitation seasonality which were the top three of the variables affected GLM model.
For RF model was the most affected from variables precipitation of wettest month, precipitation of coldest quarter
and precipitation seasonality. It can be seen that precipitation of warmest quarter (BIO18) and precipitation of
coldest quarter (BIO19) were the two variables that had affected all models. In addition, both variables represented
characteristic of the habitat of Musa serpentina with open mixed deciduous forest.
Conclusions
The prediction of the suitability map of Musa serpentina found that the most suitable locations were in
the North and West part of Thailand. However, the areas with the highest probability of presenting were in Mae
Hong Son Province cover the area of 450 square kilometers
MaxEnt is acceptable as a high-performance model for the prediction of the species distribution with
small sample size. Fuzzy pseudo-absence data can develop the algorithm of GLM and Random Forest as efficient
as MaxEnt at the small sample size. Although, the predicted area of each model had some differences whereas
PMCC test can represent that each algorithm established the suitability map of Musa serpentina in the same
direction. In conclusion, all three algorithms are highly efficient tools of species distribution models for limited
occurrence data
Acknowledgement
We would like to thank our colleagues at the Computational Science Department, School of Science,
Mae Fah Luang University for fruitful interaction and encouragement of the first author. We would like to thank
Chiang Rai Rajabhat University for supporting the ArcGIS software and Mae Fah Luang University for funding
this research.
References
Elith, J., & Graham, C. H. 2009. Do they? How do they? WHY do they differ? on finding reasons for differing
performances of species distribution models. Ecography, 32(1), 66–77. https://doi.org/10.1111/j.1600-
0587.2008.05505.x.
Franklin, J. 2010. Spatial Inference and Prediction. In Cambridge (Ed.), Mapping Species Distributions (Vol.
141). https://doi.org/10.2140/agt.2016.16.1403.
Hernandez, P. A., Graham, C., Master, L. L., & Albert, D. L. 2006. The effect of sample size and species
characteristics on performance of different species distribution modeling methods. Ecography, 29(5), 773–
785. https://doi.org/10.1111/j.0906-7590.2006.04700.x.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., & Jarvis, A. 2005. Very high resolution interpolated
climate surfaces for global land areas. International Journal of Climatology, 25(15), 1965–1978.
https://doi.org/10.1002/joc.1276.
IUCN. 2001. IUCN Red List Categories and Criteria: Version 3.1. IUCN Species Survival Commission. In IUCN.
Gland, Switzerland and Cambridge, UK..
Khatchikian, C., Sangermano, F., Kendell, D., & Livdahl, T. 2011. Evaluation of species distribution model
algorithms for fine-scale container-breeding mosquito risk prediction. Medical and Veterinary Entomology,
25(3), 268–275. https://doi.org/10.1111/j.1365-2915.2010.00935.x.
Liu, C., White, M., & Newell, G. 2011. Measuring and comparing the accuracy of species distribution models
with presence-absence data. Ecography, 34(2), 232–243. https://doi.org/10.1111/j.1600-0587.2010.06354.x.
Marod, D., Pinyo, P., Duengkae, P., & Hiroshi, T. 2010. The role of wild banana (Musa acuminata Colla) on

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 162
wildlife diversity in mixed deciduous forest, Kanchanaburi Province, Western Thailand. Kasetsart Journal
- Natural Science, 44(1), 35–43.
Morgane, B.-M., Frederic, Jiguet, C., Cecile Helene, A., & Wilfried, T. 2013. Selecting pseudo-absences for
species distribution models: how, where and how many? Methods in Ecology and Evolution 2012, 3, 327–
338. https://doi.org/10.1111/j.2041-210X.2011.00172.x.
Phillips, S. 2008. A Brief Tutorial on Maxent. AT&T Research, 1–38. https://doi.org/10.4016/33172.01.
RUSHTON, S. . P., ORMEROD, S. J., & KERBY, G. 2004. New paradigms for modeling species distributions?
Journal of Applied Ecology, 41(April 2004), 193–200. https://doi.org/10.1111/j.0021-8901.2004.00903.x.
Swangpol, S., & Somana, J. 2011. Musa serpentina. THAI FOREST BULLETIN (BOTANY), 39, 31–36.
Swets, J. A. 2015. Measuring the Accuracy of Diagnostic. American Association for the Advancement of Science,
240(4857), 1285–1293. Retrieved from http://www.jstor.org/stable/1701052?seq=1&cid=pdf-
reference#references_tab_contents.
Tovaranonte, J., Blach-Overgaard, A., Pongsattayapipat, R., Svenning, J. C., & Barfod, A. S. 2015. Distribution
and diversity of palms in a tropical biodiversity hotspot (Thailand) assessed by species distribution
modeling. Nordic Journal of Botany, 33(2), 214–224. https://doi.org/10.1111/j.1756-1051.2013.00217.x.
Trisurat, Y., Shrestha, R. P., & Kjelgren, R. 2011. Plant species vulnerability to climate change in Peninsular
Thailand. Applied Geography, 31(3), 1106–1114. https://doi.org/10.1016/j.apgeog.2011.02.007.
Wisz, M. S., Hijmans, R. J., Li, J., Peterson, A. T., Graham, C. H., Guisan, A., … Zimmermann, N. E. 2008.
Effects of sample size on the performance of species distribution models. Diversity and Distributions, 14(5),
763–773. https://doi.org/10.1111/j.1472-4642.2008.00482.x.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 163
Proceedings of International Conference on Biodiversity: IBD2019 (2019); 163 - 173
Stimulating the access to biodiversity and technologies to combat climate change
Fabrice Mattei
Rouse & Co. International (Thailand) Ltd, Klongtoey District, Bangkok, Thailand
Corresponding author e-mail: [email protected], [email protected]
Abstract: On 13 November 2017, in a joint statement issued by the Executive Secretaries at the UN Climate
Change Conference 2017 ("COP 23") taking place in Bonn, Germany1, the Executive Secretaries of respectively
the Convention on Biological Diversity ("CBD"), the United Nations Convention to Combat Desertification
("UNCCD") and the United Nations Framework Convention on Climate Change ("UNFCCC") called for
establishing a financial mechanism, called Facility, to secure finance for large projects that will help to address
common issues relating to climate change:“We are calling for the establishment of a new Project Preparation
Facility to bridge this gap and promote an integrated, coherent and multi-disciplinary approach to these related
issues while supporting the respective mandates of the three Rio Conventions.”2 This declaration officially
recognizes what was widely known; biodiversity and climate change are interconnected. Biodiversity is affected
by climate change, and equally the conservation of biodiversity contributes to mitigate climate change impacts3.
The promises made under the CBD and UNFCCC to conserve biodiversity and to combat climate change both rely
on "access". Facilitating the access to Genetic Resources ("GR") under the CBD while facilitating the access to
technologies under the UNFCCC and Paris Agreement ("PA"). This paper challenges the assumptions behind those
promises (1) before it evaluates the financial mechanisms which are supposed to stimulate the access to
biodiversity by developed countries and to climate change technologies by developing countries (2). It then
investigates what are the legal mechanisms which could further stimulate the access to biodiversity and climate
change technologies including empowering the Green Climate Fund ("GCF") to own and license climate change
technologies (3).
Keywords: Biodiversity, Technology Transfer, Climate Change, Green Climate Fund, Paris Agreement on Climate Change.
Introduction
On 13 November 2017, in a joint statement issued by the Executive Secretaries at the UN Climate Change
Conference 2017 ("COP 23") taking place in Bonn, Germany , the Executive Secretaries of respectively the
Convention on Biological Diversity ("CBD"), the United Nations Convention to Combat Desertification ("UNCCD") and
the United Nations Framework Convention on Climate Change ("UNFCCC") called for establishing a financial
mechanism, called Facility, to secure finance for large projects that will help to address common issues relating to
climate change:
“We are calling for the establishment of a new Project Preparation Facility to bridge this gap and
promote an integrated, coherent and multi-disciplinary approach to these related issues while supporting
the respective mandates of the three Rio Conventions.”
This declaration officially recognizes what was widely known; biodiversity and climate change are interconnected.
Biodiversity is affected by climate change, and equally the conservation of biodiversity contributes to mitigate
climate change impacts. The promises made under the CBD and UNFCCC to conserve biodiversity and to combat
climate change both rely on "access". Facilitating the access to Genetic Resources ("GR") under the CBD while
facilitating the access to technologies under the UNFCCC and Paris Agreement ("PA"). This paper challenges the
assumptions behind those promises (1) before it evaluates the financial mechanisms which are supposed to
stimulate the access to biodiversity by developed countries and to climate change technologies by developing
countries (2). It then investigates what are the legal mechanisms which could further stimulate the access to
biodiversity and climate change technologies including empowering the Green Climate Fund ("GCF") to own and
license climate change technologies (3).

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 164
Materials and Methods
1. Assumptions behind stimulating the access to biodiversity and technologies
1.1 Assumptions behind stimulating the access to biodiversity
Under the CBD, one way of ensuring the conservation of biodiversity is to facilitate its access and use for
commercial or research purposes by third parties through the Access and Benefit Sharing ("ABS")4. The promotion
of access to biodiversity is based on several assumptions, three of them. First, GR from biodiversity-rich countries
are ‘green gold’5. Second, limiting access to GR will inhibit the already failing bio-prospecting activity and any
financial and non financial benefits to share so that the conservation of biodiversity will be at risk6. Third, the
access to GR is mostly between resource-rich to biodiversity rich countries also known as North-South7. These
assumptions are challengeable. Studies reveal that access to GR by developed countries is less appealing than it
first appears, due in particular to the development of biotechnology and options for off-site access to GR. There is
no clear study confirming that access to biodiversity contributes to its conservation. There is a debate between
conservation and preservation of biodiversity. It contends that the access and sustainable use of biodiversity don't
necessarily ensure its preservation. Finally, the access and use of GR is not always north-south driven8. Since the
assumptions behind the access to biodiversity are not fully grounded, the CBD's access mechanism is weakened.
1.2 Assumptions behind stimulating the access to technologies
(i) Technologies' coverage
The access to technologies for mitigation of, and adaptation to, climate change has been much debated
especially at the Conferences of the Parties ("COP") of the UNFCCC. There is debate on what the term
“technology” covers and what are the critical technologies to give access to9? Chapter 24 of Agenda 21 under the
UNFCCC refers to "Environmentally Sound Technologies" (“ESTs”), which are intended to encompass the
following technologies: technologies protecting the environment, less polluting technologies, technologies using
resources in a more sustainable manner, technologies aiming at recycling of waste and products, and technologies
handling residual wastes.
The Intergovernmental Panel for Climate Change (“IPCC”) further distinguishes between “Climate
Change Mitigation Technology” ("CCMT") and “Climate Change Adaptation Technology” ("CCAT"). CCMT
covers technological change and substitution that reduce energy resource inputs and emissions, while CCAT
covers technologies intended to reduce the harmful effects arising from expected climate change. Finally, the PA
further specifies the stage of technology cycle which matter to give access to, it should be "early stages of the
technology cycle"10, rather than mature technologies.
To sum up, the term “technology” should cover environmentally friendly technologies at early stages of
the technology cycle that involve energy efficiency, alternatives to fossil fuel and carbon generation, pollution and
toxic remediation, water purification, recycling, safety and health concerns, renewable resources.
(ii) Access scope to technologies
Under the UNFCCC and the PA, the access to and transfer of technologies is an encompassing notion in
climate change adaptation and mitigation. The first assumption under the UNFCCC is that climate change
technologies are under developed countries’ ownerships and their transfer to developing countries is also under
their control:
Article 4, paragraph 5 of the UNFCCC reads: “The developed country Parties shall take all practicable
steps to promote, facilitate and finance, as appropriate, the transfer of, or access to, environmentally
sound technologies and know-how to other Parties, particularly developing country Parties, to enable
them to implement the provisions of the Convention.”
Chapter 34 of Agenda 21 of Transfer Of Environmentally Sound Technology, Cooperation And Capacity-
building11 further suggests that the access and transfer of “ESTs” should be promoted in a balanced manner taking
into account the interests of developed and developing countries:
"To promote, facilitate, and finance, as appropriate, the access to and the transfer of environmentally
sound technologies and corresponding know-how, in particular to developing countries, on favourable

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 165
terms, including on concessional and preferential terms, as mutually agreed, taking into account the need
to protect intellectual property rights as well as the special needs of developing countries for the
implementation of Agenda 21;"12
The Kyoto Protocol also promotes the access to climate change technology in its Article 10 by repeating
UNFCCC’s paradigm according to which developed countries shall “take all practical steps” to facilitate the
transfer of, or access to, transfer ETSs to developed countries:
“All Parties...shall: (c) Cooperate in the promotion of effective modalities for the development,
application and diffusion of, and take all practicable steps to promote, facilitate and finance, as
appropriate, the transfer of, or access to, environmentally sound technologies…in particular to
developing countries…”13
More recently, the PA sets the stage for urgently needed climate change technologies’ access,
development and transfer. Notably, countries anchored the Technology Mechanism (“TM”) within the PA and
decided that it would be a key element in implementing the PA. They strengthened the TM, requesting further
work on technology access, research, development and demonstration, as well as enhancing endogenous capacities
and technologies (e.g. locally created and developed technologies).
The assumptions under the UNFCCC and PA are challengeable. First, there is a growing number of
developing countries which are heavily engaged in climate change technologies, developing and patenting
inventions in this area. For example, the Thai company Siam Cement is heavily engaged in environmental friendly
paper and cement.
The Siam Cement Public Company Limited ("SCG") is a Thai company which was established over 100 years
ago in Thailand in 1913, following a Royal Decree of His Majesty King Rama VI in order to eliminate reliance
on imports of cement, as well as fully using own country natural resource14. The goal is to diversify and meet
the economic growing in Thailand.
SCG established and participated in new industries under SCG. SCG comprises over 100 companies under 5
business groups (Cement, paper, chemical, building material, distribution) employing approximately 40,000
employees, handling more than 80,000 product items and filing over 150 patents a year. In 2005, SCG established
a working committee dedicated specifically to innovation.
In 2017, SCG had nearly 1,700 staff members in the R&d department. This included over 100 PhD graduates
and it has a research budget of more than 424,240,40.00 USD per year, again resulting in over USD
4,250,400,250.40 of sales from high value-added products.
A wide range of environmental friendly products have been invented, produced and distributed worldwide by
SCG. For example, products that use minimal natural resources such as "Idea Green paper"15, which reduces the
use of trees by 30 %. Another innovative product is "EL Green plastic pellets"16 (used for plastic packaging);
this is biodegradable under certain temperatures and moisture levels. Another green invention is SCG Marine
Cement17. It is sea water resistant and the greenhouse gas emissions resulting from its production have been
reduced by at least 350 kilograms per ton of cement. These locally made and developed technologies effectively
reduces greenhouse gas emissions and contribute to mitigate climate change impacts18
Second, early stage technologies are not generally opened to access and transfer to developing countries, due to
their confidentiality, owners’ high return on investment expectations, and delays in patenting them especially in
developing countries. There is also a significant part of climate change technology which is mature technology.
One example is solar technology. China has engaged into this area becoming the world’s largest developer,
manufacturer and exporter of solar cells. This was due partially because the technology used was several
generations old and many patents were in public domain19.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 166
Since the assumptions for facilitating access to technologies are not fully well grounded, the UNFCC and PA
access facilitation mechanisms are also weakened.
2. Financial mechanisms to stimulate the accesses to biodiversity and technologies
The financial mechanisms to stimulate the access to biodiversity and climate change technologies under
the CBD, UNFCCC and PA are of various kinds; public, private and hybrid.
2.1 Financial mechanisms to stimulate the accesses to biodiversity
The financial mechanisms to stimulate the access to biodiversity are dominantly (i) private and to some
extend (ii) hybrid between public and private financing sources. There is a consensus that the conservation of
biodiversity cannot be fully achieved through private financial stimulants. Although the participation of the private
sector in promoting the access to biodiversity through the Access and Benefit Sharing (“ABS”) scheme is
encouraged under the CBD and related COPs, its real impact on the conservation of biodiversity is negligible.
(i) Private financial mechanisms
The externalization of biodiversity conservation costs is possible with the help of payments from
industrialized nations' companies to developing countries for their access and use of GR and TK. Upon reviewing
the COPs decisions on financial resources, they are pushing the GEF to take into account the potential of private
sector involvement. However, the private sector has little or no incentive to collaborate with GEF’s activities. In
1996 and in 1999, the Secretariat of the CBD submitted two proposals for Council consideration aimed at
increasing the role and importance of the private sector in the GEF20. However, no measures have taken place
pursuant to these two proposals. The problem of biodiversity lies in the external uncompensated benefits that
diverse resources render to the global community.
There is a specific mechanism under the CBD to allow private financing of biodiversity conservation
known as the Access and Benefit Sharing (“ABS”) promoting the access to GR and TK and their use for
commercial or non commercial purposes.
Private law contracts regulating Prior Inform Consent ("PIC") and Material Transfer Agreement ("MTA")
are the core instruments of ABS. Conservation of biodiversity is supposed to be linked to monetary and non-
monetary benefits shared by the users with the providers of the GR. According to the literature there was over-
expectation on the capacity to extract value from GR especially from bio-prospecting for new industrial products,
such as pharmaceuticals. These activities are unlikely to generate significant funds to conserve biodiversity. One
reason for this is that a considerable percentage of the added value in bio-prospecting projects accrues outside the
country where it takes place. In addition, the assumption that private companies were prepared to subject
themselves to stringent conditions and complex procedures to get access to GR and associated TK has proven
wrong. Companies are also unwilling to incur significant up-front costs in bio-prospecting, especially if samples
of the same resources are available in other countries without or with less stringent regulations.
(ii) Hybrid financial mechanisms
Debt-for-nature swaps are financial transactions that reduce a developing country’s debt stock or service
in exchange for a commitment to protect nature from the debtor-¬government. It is a voluntary transaction whereby
the donor(s) cancels the debt owned by a developing country’s government. The savings from the reduced debt
service are invested in conservation projects. Several organizations such as the Conservation International, The
Nature Conservancy, and the World Wildlife Fund have participated in debt-for-nature swaps. The first debt-for-
nature-swap took place in 1987. It was between Conservation International and the Bolivian government according
to which Conservation International purchased the Bolivian debt having of an original value of USD 650,000 for
the amount of USD 100,000. In return, the Bolivian government agreed to expand an environmentally protected
area by 1.2 million hectares where 13 of the 18 threatened species in this country live. When looking at the
conservation needs, total debt of developing countries and rarity of debt-for-nature swaps agreements this financial
mechanism is of little significance to conserve biodiversity through accessing it.
2.2 Financial mechanism to stimulate the access to climate change technologies
The PA refers to "Financial Mechanism" of the UNFCCC to strengthen cooperative action on technology
transfers by facilitating the access to technologiwa, especially to "early stages of technology cycle". Article 10.5
of the PA reads as follows:
“Accelerating, encouraging and enabling innovation is critical for an effective, long-term global
response to climate change and promoting economic growth and sustainable development. Such effort
shall be, as appropriate, supported, including by the Technology Mechanism and, through financial
means, by the Financial Mechanism of the Convention, for collaborative approaches to research and
development, and facilitating access to technology, in particular for early stages of the technology cycle,
to developing country Parties.”

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 167
The financial support to accessing to technologies is provided by the GCF. It cooperates with the
Technology Mechanism that was first mentioned in the Copenhagen Accord in 2009 and was formally established
one year later during the COP 16 held in Cancun, Mexico and is reiterated in the PA. The GCF, together with the
GEF, are the two operating entities of the Financial Mechanism and as such represent the main channels through
which sources of international climate finance are flowing.
The Paris Decision 1/CP.2121 serving as guidance for the implementation of the PA and pre-2020 action,
"strongly urges developed country Parties to scale up their level of financial support, with a concrete roadmap to
achieve the goal of jointly providing USD 100 billion annually by 2020 for mitigation and adaptation"22 .
Furthermore, the Decision adds that prior to 2025 the COP shall set a new ‘collective quantified goal from a floor
of USD 100 billion per year’23.
Most climate change technologies relate to CCMT rather than CCAT24. Since its establishment in 1991,
the GEF has been funding projects on CCMT in developing countries. As at June 30, 2017, the GEF has supported
867 projects on CCMT with over USD 5.3 billion GEF funding in more than 165 countries25. Out of 867 projects
that were implemented in developing countries, 31.1 % in Asia, 25.3 % were in Africa, 18.5 % in LAC, and 17.2
% in Eastern Europe and Central Asia. In addition, there were 69 global and regional projects that account for
eight per cent of the overall CCM portfolio. The table below categorizes these 867 projects in the areas of low
carbon energy, energy efficiency, renewable energy, sustainable transport and urban systems. However out of the
867 projects it appears that only 67 relate to technology transfers which is approximately 7.7 %. It indicates that
access to technologies by developing countries is very limited.
3. Legal mechanisms to stimulate the access to biodiversity and climate change technologies
3.1 Legal stimulants for accessing to biodiversity
The CBD relies on a contractual model to grant access to genetic resources and ensure benefit sharing in
case of exploitation thereof. National policy makers determine criteria for ensuring benefits sharing. Bio-rich
countries' national access regimes to biodiversity were adopted in a context of concerns which are still valid about
the unauthorized access and misappropriation of GR and the lack of compensation for the benefits their commercial
exploitation could generate. Access is generally regarded as a threat rather than an opportunity. Consequently,
such regimes have been mostly defensive, aimed at controlling rather than facilitating the access and use of GR
for research and commercial purposes26. Those stringent regimes have had limited or no effects on conservation
since, as we mentioned earlier, ABS mechanism has not proven to contribute to the conservation of biodiversity.
The following are some examples.
South Africa’s ABS regulations27 require that the applicant for a permit be either a corporation registered
under South African law, or a natural person who is a citizen or a permanent resident of South Africa. If the
applicant is a foreign natural or corporation, they must apply jointly with a South African corporation or citizen or
permanent resident. Therefore, foreign individuals, corporations or institutions cannot apply for a permit alone.
In Costa Rica28, if the applicant lives in a foreign country, he or she must assign a legal representative
who is a resident of Costa Rica.
India also has a differential procedure to handle foreign applications, as does the Andean Community,
Brazil and others. Some countries like India have created a separate procedure depending on nationality, or if the
materials accessed or research results will be sent abroad29.
The Philippines has a relatively long history of regulating access to biodiversity and impose stringent
conditions against bio-prospectors which deter most access to biodiversity30.
(I) Legal stimulants for accessing to technologies
The challenge faced in climate change sector is to encourage innovations in this field while at the same
time promoting the access, diffusion and the transfer of these technologies to developing countries. Technology
transfer is crucial and is seen by developing countries as a prerequisite to adhere to treaties like the UNFCCC. For
example, at the 1992 Rio Earth Summit, China and India insisted that developed countries commit themselves to
technology transfer as a requirement for developing country support for the proposed agreements. Discussions in
general about technology transfer for climate change mitigation have been affected by deep divides between
developed and developing countries. Most developing countries see patents as a barrier to access to technologies

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 168
and their transfer, while developed countries see patents as a necessary mechanism to incentivize and license
climate change technologies. Consequently, like with the access regime to biodiversity under CBD, developing
countries' legislations on patent have been mostly defensive, aiming even at imposing compulsory licenses against
developed countries' inventions having the potential to reduce greenhouse gas emissions. The cleavage is strong
enough to explain why the PA is silent on intellectual property, including patent.
Stimulating access to technologies requires strong incentives. Most climate technologies are owned and
developed by commercial companies. Giving access to their technologies by developing countries implies risking
intellectual property rights and empowering existing or potential competitors. Some negotiations on climate
change technology transfer have not identified this problem. At the Second COPs to the UNFCCC in 1996, the
Chinese government published a booklet "The List of Chinese Government Needed Technologies" in which it
demanded climate change technology equipments including integrated gasification combined cycles, fuel cells,
and rice husk energy transfer instruments. However, the booklet remains silent on any compensation for
technology owners. The demands of China and other developing countries to access to technologies can be driven
by their desire to enhance their competitiveness explaining why such demands are usually resisted by developed
countries, creating impasses about how to stimulate the access to ESTs.
GCF's juridical personality and legal capacity
Due to the difficulties to incentivize companies to give access and transfer their technologies and know how to
developing countries, we should investigate whether the GCF could actually own and license technologies which
would allow it to invest into R&D projects and facilitate the implementation of climate change mitigating
approaches in developing countries where they are most needed. Legally speaking, there are no reasons why the
GCF could not own green technologies and patents. The GCF is not only a juristic person but it also has the legal
capacity to own and license patents. The Governing instrument for the GCF, decision 3/CP.17, 11 December 2011,
article 7 reads as follows:
“In order to operate effectively internationally, the Fund will possess juridical personality and will have
such legal capacity as is necessary for the exercise of its functions and the protection of its interests.”
GCF's mandate to own and license technologies
The next question is whether it is the GCF's mandate to own and license technologies? Under the PA, the mandate
of the GCF is to facilitate the access and transfer of technologies and R&D cooperation. Articles 10, 10.5 and
10.6 of the PA refer to the Technology Mechanism and Financial Mechanism as key instruments to allow the
access and transfer of technologies to developing countries.
The COPs Draft Decision no. 7/CP.21 gives to the GCF some specific tasks for its future actions including
"undertaking R&D activities":
"Also invites the Board of the Green Climate Fund, in line with paragraph 38 of the governing instrument
of the Green Climate Fund, to consider ways to provide support, pursuant to the modalities of the Green
Climate Fund, for facilitating access to environmentally sound technologies in developing country
Parties, and for undertaking collaborative research and development for enabling developing country
Parties to enhance their mitigation and adaptation action;"
The COPs decision 3/CP.17 at its paragraph 38 entrusts the GCF to "ensure adequate resources for […] technology
development and transfer […]".
The 2016 Green Climate Fund's publication entitled “Support for facilitating access to environmentally
sound technologies and for collaborative research and development” at paragraphs 23-28 refer to options for R&D
collaboration which the GCF could undertake.
Paragraph 24 reads:
"The benefits of research and development can help countries to address their climate and sustainable
development in the longer term. Even though research and development as an activity poses many risks,
it is only through this process that new technologies are developed and brought to the market."
The GCF solemnly declares the need to invest in R&D and is willing to devote investment, despite the risks that
are inherent.

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 169
Paragraph 25 is even more specific on the possibilities for GCF to finance R&D activities:
"GCF could provide financing for research and development activities and help countries tackle the he
challenges faced in the early stages of the technology development cycle […]"
It further mentions in its Paragraph 26 possible R&D activities which it could finance as follows:
"(a) Competitive GCF innovation funding to stimulate local/regional interest and investment through the
tailored request for proposals;
(b) Promoting micro-finance for research and development projects in developing countries;
(c) Working with governments to strengthen national innovation systems;
(d) Facilitating the establishment of research center networks on environmentally sound technologies;
and
(e) Catalyzing research partnerships with relevant stakeholders, including collaboration with the private
sector.
The Board may wish to consider the following options for providing such support for research and
development:
(a) Business incubation and financial support for viable, new technologies to be deployed in developing
countries; or
(b)Capacity-building programmes/request for proposals for developing countries to enhance endogenous
capacities related to climate technologies."
Paragraph 27 further explains:
"The first option [means a) above] would focus on bringing new, viable technologies to market through
business incubation, and early and growth stage financing. The Private Sector Facility could potentially
play an important role in this area, attracting venture capital and angel investors to deploy their capital
in developing countries."
The GCF plans so explicitly to finance R&D activities in order to bring technologies to market. Certainly, the GCF
is not mandated to fund basic research, which is wise, because the cycle to produce a workable technology is long
and uncertain. However, the phase of industrialization of technology is often a time that patents are filed. The issue
of industrial property will arise so necessarily for incubation projects in which the GCF would invest.
To summarize, the GCF has legal personality and capacity, as it may perform all legal operations that
allow it to achieve its missions. The GCF should be able to own technologies and license them. This would also
mean that owners of patented technologies could donate a license to GCF which may then manage a patent pool.
The GCF could use these licenses for its beneficiaries, mainly developing countries.
Result & Discussion
From the review above, key findings are that facilitating the access to climate change technologies and
the access to GR have shown little impacts on mitigating or adapting to climate change. This result ties well with
previous studies wherein access to biodiversity hasn’t necessarily resulted in improved biodiversity conservation.
One concern about the findings of access to biodiversity was that limited information is available on bio-
prospecting activities and how the access to biodiversity is usually retributed. Likewise, there is lack of information
on technology transfers of EST, especially between private entities.
Conclusion
The conservation of biodiversity by facilitating its access to developed countries has shown little results.
Most rich bio-resources countries have enacted defensive legislations which deter bio-prospectors from entering
into MTAs to access to GR and TK. Furthermore, the development of biotechnology offers viable alternatives to
GR. It can be concluded that continuing promoting the access to biodiversity under the CBD won't offset the fast
decline of GR. Emphasizes should be placed on other mechanisms that access, for example public financial
instruments. Promoting the access to climate change technologies is also challenging. However, the option for the
GCF to invest in green technologies including through incubator whose mission is to support, including financially,
the development of existing technologies is promising. R&D in the field of adaptation of existing technologies is
conducive to collaboration, because it requires double competencies, both technological and knowledge of the

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 170
conditions specific to each country or region. In addition, it is often considered that technology held by western
companies need to be adapted to the specific conditions of other countries. Integrating a licensing system into the
GCF could better ensure developing countries gain easier and faster access to climate change technologies. The
GCF and the Technology Mechanism are exciting new institutions offering the possibility of a global license
system that ensures that intellectual property, royalties, investment etc are not barriers to technology access, and
can become the missing link between innovating and giving accessing to technologies.
References
1 Joint Statement by the Executive Secretaries of the Rio Conventions calling for the establishment of a Project
Preparation Facility (PPF) to increase financing for large-scale, transformative projects which integrate
action on land degradation, biodiversity loss, and global warming. https://www.cbd.int/cooperation/joint-
statement-rio-convention-2017-en.pdf 2 Supra no. 2 3 The CBD Secretariat has published two technical series on biodiversity and climate change to support the
implementation of relevant adaptation activities. Technical Series No. 10 – Interlinkages between
Biological Diversity and Climate Change; https://www.cbd.int/doc/publications/cbd-ts-10.pdf
Technical Series No. 25 – Guidance for Promoting Synergy among Activities Addressing Biological
Diversity, Desertification, Land Degradation and Climate Change;
https://www.cbd.int/doc/publications/cbd-ts-25.pdf 4Joint Statement by the Executive Secretaries of the Rio Conventions calling for the establishment of a Project
Preparation Facility (PPF) to increase financing for large-scale, transformative projects which integrate
action on land degradation, biodiversity loss, and global warming. https://www.cbd.int/cooperation/joint-
statement-rio-convention-2017-en.pdf 5Supra no. 2 6The CBD Secretariat has published two technical series on biodiversity and climate change to support the
implementation of relevant adaptation activities. Technical Series No. 10 – Interlinkages between
Biological Diversity and Climate Change; https://www.cbd.int/doc/publications/cbd-ts-10.pdf
Technical Series No. 25 – Guidance for Promoting Synergy among Activities Addressing Biological
Diversity, Desertification, Land Degradation and Climate Change;
https://www.cbd.int/doc/publications/cbd-ts-25.pdf 7Achim Seiler and Graham Dutfield, Regulating Access and Benefit
Sharing, German Federal Agency for Nature Conservation 2001 Sarah Laird and Rachel Wynberg Access
and Benefit-Sharing in practice: Trends in Partnerships across Sectors, CBD Technical Series No. 38. 8Nicolas Pauchard, Access and Benefit Sharing under the Convention on Biological Diversity and Its Protocol:
What Can Some Numbers Tell Us about the Effectiveness of the Regulatory Regime? Resources 2017,
6, 11, pp. 1-15. 9Carlos M. Correa, Do national access regimes promote the use of genetic resources and benefit sharing?, Int. J.
Environment and Sustainable Development, Vol. 4, No. 4, 2005, pp. 444-463. 10South-South Cooperation as a tool for the implementation of the new CBD Strategic Plan for 2011–2020 Volume
1 Issue 1 · October 2010, https://www.cbd.int/cooperation/ssc/ssc-newsletter-01-01-en.pdf; Malgozata
Blicharska, Steps to overcome the North-South divide in research to climate change policy and practice,
https://www.nature.com/articles/nclimate3163?WT.feed_name=subjects_developing-world 11M.D. Madhusudan and T.R. Shankar Raman, Conservation as if Biological Diversity Matters: Preservation
versus Sustainable Use in India; Monika Böhm and Ben Collen, Toward equality of biodiversity
knowledge through technology transfer, Centre for Biodiversity & Environment Research, University College London;
http://discovery.ucl.ac.uk/1467059/5/Bohm%26Collen_Toward%20equality%20of%20biodiversity%20
technology%20transfer.pdf
“However, the traditional view of north-south technology transfer (e.g., from resource-rich to
biodiversity-rich countries in the original context of the CBD) is being challenged increasingly because
south-south technology transfer is growing in importance”;
“It is likely that, in future, south-south transfer will require greater attention as more agreements are made
to foster these interactions”
“It is likely that technology transfer is more frequent among countries with economic, geographic,
cultural, or historic ties”. 12Heleen de Coninck and Shikha Bhasin, Meaningful technology development and transfer: A necessary condition
for a viable climate regime, Towards a Workable and Effective Climate Regime, Radboud University;
Energy research Centre of the Netherlands (ECN) pp. 451-464 13Article 10.5 of the Paris Agreement "Accelerating, encouraging and enabling innovation is critical for an
effective, long-term global response to climate change and promoting economic growth and sustainable
development. Such effort shall be, as appropriate, supported, including by the Technology Mechanism

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 171
and, through financial means, by the Financial Mechanism of the Convention, for collaborative
approaches to research and development, and facilitating access to technology, in particular for early
stages of the technology cycle, to developing country Parties." 14Transfer Of Environmentally Sound Technology, Cooperation And Capacity-building, http://www.un-
documents.net/a21-34.htm 15Paragraph 14 b) http://www.un-documents.net/a21-34.htm 16Kyoto Protocol to the United Nations Framework Convention on Climate Change of 1998
http://unfccc.int/resource/docs/convkp/kpeng.pdf 17http://www.scg.com/en/01corporate_profile/ 18http://www.scgpackaging.com/others/home-office-paper/idea-green/EN 19http://www.scg.com/id/05sustainability_development/eco-value.html 20http://www.scg.com/en/08news_release/01_news/detail.php?ContentId=2164 21Orose Leelakulthanit, Sustainability: The Case Of Siam Cement Group (SCG), Journal of Business Case Studies
– Fourth Quarter 2014 Volume 10, Number 4; Innovative IP development, Siam Cement Group
http://www.wipo.int/ipadvantage/en/articles/article_0054.html 22Cynthia Cannady, Access to Climate Change Technology by Developing Countries, ICTSD Global Platform on
Climate Change, Trade Policies and Sustainable Energy, Issue Paper 25, September 2009, pp. 1-44. 23Porter Gareth, Raymond Clemencon, Waafas Ofosu-Amaah, and Michael Philips, Communicative Structure and
Governance of the Global Environment
Through Policy Networks of International Aid Organizations, 1998. 24Shah Md. Mushfiqur Rahman, A critical examination of the Financial Mechanism as adopted in the Convention
on Biological Diversity (CBD) from a developing country’s perspective, The Northern University Journal
of Law, Volume I (2010). 25Elisa Morgera,The Evolution of Benefit Sharing: Linking Biodiversity and Community Livelihoods, 26Gordon C. Rausser University of California, Berkeley Arthur A. Small Columbia University, Valuing Research
Leads: Bioprospecting and the Conservation of Genetic Resources, Journal of Political Economy, 2000,
vol. 108, no. 1. Nicolas Pauchard, Access and Benefit Sharing under the Convention on Biological
Diversity and Its Protocol: What Can Some Numbers Tell Us about the Effectiveness of the Regulatory
Regime? Resources 2017, 6, 11; doi:103 390/resources6010011 pp.1-15, Barett, C.B.; Lybbert, T.J. Is
bioprospecting a viable strategy for conserving tropical ecosystems? Ecol. Econ. 2000, 34, 293–300,
Moretti, C.; Aubertin, C. Stratégies des firmes pharmaceutiques: La bioprospection en question. In Les
Marchés de la Biodiversité; Aubertin, C., Pinton, F., Boisvert, V., Eds.; Institut de Recherche Pour le
Développement (IRD): Paris, France, 2007., Morgera, E.; Tsioumani, E.; Buck, M. Unraveling the
Nagoya Protocol: A Commentary on the Nagoya Protocol on Access and Benefit-sharing to the
Convention on Biological Diversity.
http://booksandjournals.brillonline.com/content/books/9789004217188
“Key publications on the ABS regime implemented through the CBD and its protocol note its difficulties,
so far, in achieving its objectives, as very few ABS agreements have been concluded and consequently
very few benefits have been shared."
“However, to our knowledge, there is not any accurate counting of either access permits granted or ABS
agreements concluded since the entry into force of the CBD or the NP. I found piecemeal information
indicating that the number is small but no systematic inventory. I consider that providing reliable data
about access permits and ABS agreements is essential to analyze the concrete functioning of the regime.
This is the core aim of our contribution.” 27Cabrera, M.J. (2003) ‘Bioprospecting: policy, regulatory and market incentives’, paper presented at the
Megadiverse Countries Meeting on Environmental Legislation on Access to Genetic Resources,
Protection of Traditional Knowledge and Intellectual Property Rights, Cusco, 27–29 November, pp. 246-247 28Cox, P.A. 2001. Ensuring equitable benefits: The Falealupo Convenant and the isolation of anti-viral drug
prostatin from a Samoan Medicinal Plant. Pharmaceutical Biol. 39, 33-40. 29Debt for Nature Swaps, Financing Solutions for Sustainable Development (2017) 30James P. Resor, Debt-for-nature swaps: a decade of experience and new directions for the future,
http://www.fao.org/docrep/w3247e/w3247e06.ht 31http://unfccc.int/meetings/copenhagen_dec_2009/items/5262.php 32http://unfccc.int/resource/docs/2015/cop21/eng/10a01.pdf 33Paragraph 15 of the Paris Decision 34Paragraph 54 of the Paris Decision 35Antoine Dechezleprêtre, Matthieu Glachant, Ivan Hascic, Nick Johnstone, Yann Ménière, nvention and Transfer
of Climate Change Mitigation Technologies on a Global Scale: A Study Drawing on Patent Data (2009)
Mines paris Tech http://personal.lse.ac.uk/dechezle/Innovation_diffusion_climate_techs_AFD.pdf

Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019 at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 172
United Nations Environment Programme (UNEP)/European Patent Office (EPO) study Patents and clean
energy technologies in Africa, 2013, www.epo.org/clean-energy-africa; 36Report of the Global Environment Facility to the Twenty-third Session of the Conference of the Parties to the
United Nations Framework Convention on Climate Change, August 2017
https://www.thegef.org/sites/default/files/documents/Final%20COP%2023%20Report%20August%203.pdf 37Carlos M. Correa, Do national access regimes promote the use of genetic resources and benefit sharing?, Int. J.
Environment and Sustainable Development, Vol. 4, No. 4, 2005, pp. 444-463. Jorge Cabrera Medaglia,
Frederic Perron-Welch And Freedom-Kai Phillips, Overview Of National And Regional Measures On
Access And Benefit Sharing (2004) Centre for International Sustainable Development Law (CISDL). 38Neil R. Crouch South Africa’s bioprospecting, access and benefit-sharing legislation: current realities, future
complications, and a proposed alternative South African Journal of Science 104, September/October
2008, South Africa’s legislative framework on bioprospecting, access and benefit sharing, 7th Pan-
African ABS Workshop Hans Merensky Hotel, Phalaborwa 25 February-1 March 2013,
http://www.abs-initiative.info/uploads/media/Lactitia_Tshitwamulomoni_-_DEA_-
_South_Africa_s_legislative_framework.pdf 39Carment Richerzhagen , The effectiveness of access and benefit sharing in Costa Rica: implications for national
and international regimes, Ecological Economics, Volume 53, Issue 4, 2005 pp.445-460 40Indian ABS guidelines http://nbaindia.org/uploaded/pdf/Gazette_Notification_of_ABS_Guidlines.pdf 41Carlos M. Correa, Do national access regimes promote the use of genetic resources and benefit sharing?, Int. J.
Environment and Sustainable Development, Vol. 4, No. 4, 2005, pp. 444-463. 42Tim Forsyth, Climate change investment and technology transfer in Southeast Asia,
http://personal.lse.ac.uk/FORSYTHT/Harr-ch13.pdf 43Legal Personality and the Green Climate Fund (2011) http://legalresponseinitiative.org/wp-
content/uploads/2013/09/BP38E-Briefing-Paper-Legal-Personality-of-the-Green-Climate-Fund-8-
December-2011.pdf 44"Accelerating, encouraging and enabling innovation is critical for an effective, long-term global response to
climate change and promoting economic growth and sustainable development. Such effort shall be, as
appropriate, supported, including by the Technology Mechanism and, through financial means, by the
Financial Mechanism of the Convention, for collaborative approaches to research and development, and
facilitating access to technology, in particular for early stages of the technology cycle, to developing country Parties." 45"Support, including financial support, shall be provided to developing country Parties for the implementation of
this Article, including for strengthening cooperative action on technology development and transfer at
different stages of the technology cycle, with a view to achieving a balance between support for mitigation
and adaptation. The global stocktake referred to in Article 14 shall take into account available information
on efforts related to support on technology development and transfer for developing country Parties." 46http://unfccc.int/resource/docs/2015/cop21/eng/l07.pdf 47Paragraph 22 of the COPs Decision no. 7/CP.21 48http://unfccc.int/resource/docs/2011/cop17/eng/09a01.pdf#page=55 "The Board shall also ensure adequate
resources for capacity-building and technology development and transfer. The Fund will also provide
resources for innovative and replicable approaches." 49GCF/B.14/02, 30 September 2016 https://www.greenclimate.fund/documents/20182/409835/GCF_B.14_02_-
_Support_for_facilitating_access_to_environmentally_sound_technologies_and_for_collaborative_rese
arch_and_development.pdf/410006c7-c3f6-4abc-96fe-cfa9280994e0 50Menno van der Veen Department of Biotechnology, Section Biotechnology and Society, Delft University of
Technology, Delft, The Netherlands. The Green Climate Fund as a Patent Pool for Innovations, Nature
Biotechnology (2012) 51Report Of The Global Environment Facility To The Twenty-Fourth Session Of The Conference Of The Parties
To The United Nations Framework Convention On Climate Change, 55th GEF Council Meeting
December 17 – 20, 2018 Washington, D.C, available at :
http://www.thegef.org/sites/default/files/council-meeting-
documents/EN_GEF.C.55.Inf_.XX_UNFCCC_CoP_Report.pdf
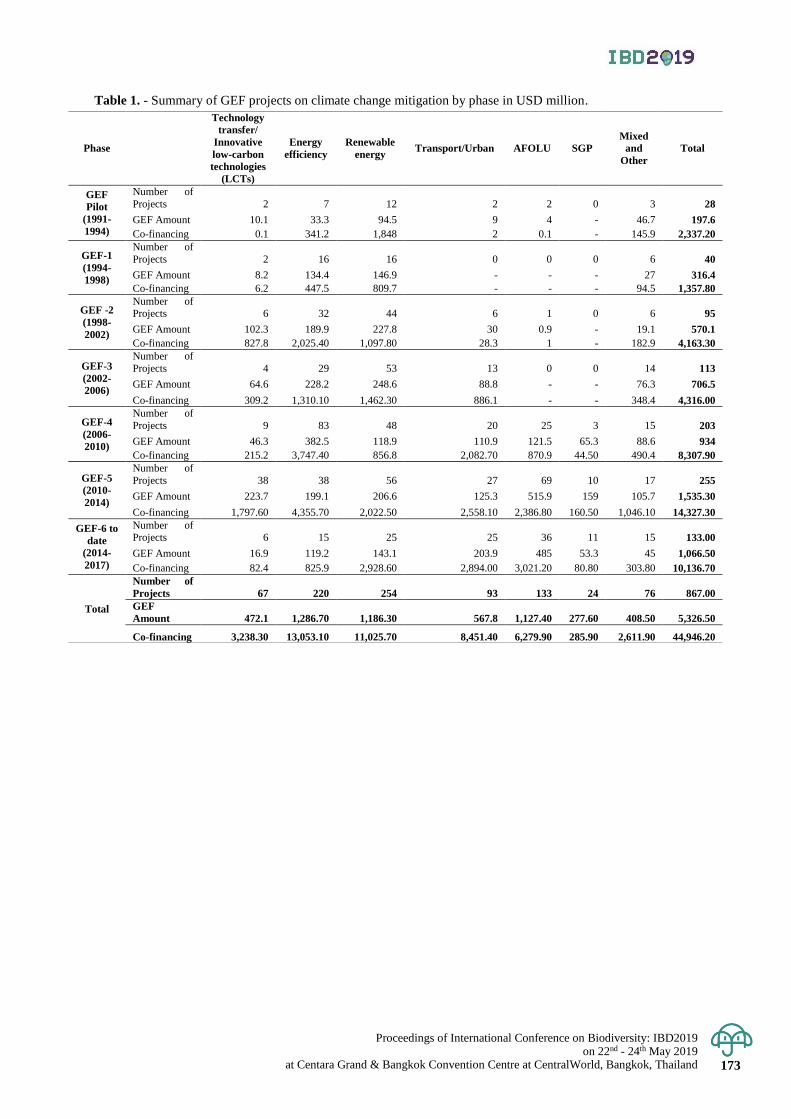
Proceedings of International Conference on Biodiversity: IBD2019 on 22nd - 24th May 2019
at Centara Grand & Bangkok Convention Centre at CentralWorld, Bangkok, Thailand 173
Table 1. - Summary of GEF projects on climate change mitigation by phase in USD million.
Phase
Technology
transfer/
Innovative
low-carbon
technologies
(LCTs)
Energy
efficiency
Renewable
energy Transport/Urban AFOLU SGP
Mixed
and
Other
Total
GEF
Pilot
(1991-
1994)
Number of
Projects 2 7 12 2 2 0 3 28
GEF Amount 10.1 33.3 94.5 9 4 - 46.7 197.6
Co-financing 0.1 341.2 1,848 2 0.1 - 145.9 2,337.20
GEF-1
(1994-
1998)
Number of
Projects 2 16 16 0 0 0 6 40
GEF Amount 8.2 134.4 146.9 - - - 27 316.4
Co-financing 6.2 447.5 809.7 - - - 94.5 1,357.80
GEF -2
(1998-
2002)
Number of Projects 6 32 44 6 1 0 6 95
GEF Amount 102.3 189.9 227.8 30 0.9 - 19.1 570.1
Co-financing 827.8 2,025.40 1,097.80 28.3 1 - 182.9 4,163.30
GEF-3
(2002-
2006)
Number of
Projects 4 29 53 13 0 0 14 113
GEF Amount 64.6 228.2 248.6 88.8 - - 76.3 706.5
Co-financing 309.2 1,310.10 1,462.30 886.1 - - 348.4 4,316.00
GEF-4
(2006-
2010)
Number of
Projects 9 83 48 20 25 3 15 203
GEF Amount 46.3 382.5 118.9 110.9 121.5 65.3 88.6 934
Co-financing 215.2 3,747.40 856.8 2,082.70 870.9 44.50 490.4 8,307.90
GEF-5
(2010-
2014)
Number of
Projects 38 38 56 27 69 10 17 255
GEF Amount 223.7 199.1 206.6 125.3 515.9 159 105.7 1,535.30
Co-financing 1,797.60 4,355.70 2,022.50 2,558.10 2,386.80 160.50 1,046.10 14,327.30
GEF-6 to
date
(2014-
2017)
Number of Projects 6 15 25 25 36 11 15 133.00
GEF Amount 16.9 119.2 143.1 203.9 485 53.3 45 1,066.50
Co-financing 82.4 825.9 2,928.60 2,894.00 3,021.20 80.80 303.80 10,136.70
Total
Number of
Projects 67 220 254 93 133 24 76 867.00
GEF
Amount 472.1 1,286.70 1,186.30 567.8 1,127.40 277.60 408.50 5,326.50
Co-financing 3,238.30 13,053.10 11,025.70 8,451.40 6,279.90 285.90 2,611.90 44,946.20

www.ibd2019exhibition.org
)