interleukin-1β (il-1β) is a modulator of human luteal cell steroidogenesis: localization of the il...
TRANSCRIPT

Interleukin-1b (IL-1b) Is a Modulator of Human LutealCell Steroidogenesis: Localization of the IL Type ISystem in the Corpus Luteum*
PAULINA KOHEN†, ANDREA CASTRO†, PEDRO CABALLERO-CAMPO,OLGA CASTRO, MARGARITA VEGA, ANTONIS MAKRIGIANNAKIS,CARLOS SIMON, PILAR CARVALLO, AND LUIGI DEVOTO
Institute of Maternal and Child Research, School of Medicine and Department of Obstetrics andGynecology (P.K., A.C., O.C., M.V., L.D.), the Human Genetics Program, ICBM, School of Medicine(P.C.), University of Chile, Santiago, Chile; the Department of Obstetrics and Gynecology, University ofValencia (P.C.-C., C.S.), Valencia, Spain; and the Center for Research on Reproduction and Women’sHealth and Department of Obstetrics and Gynecology, University of Pennsylvania School of Medicine(A.M.), Philadelphia, Pennsylvania 19104
ABSTRACTThe present investigation examined the effect of interleukin-1b
(IL-1b) on progesterone production by human luteal cells and theexpression and localization of the IL-1 system in the human corpusluteum (CL). Luteal cells were isolated from corpora lutea collectedthroughout the luteal phase. After dispersion, luteal cells weretreated with a panel of monoclonal antibodies directed to leukocyte-specific molecules. The leukocytes were isolated with immunomag-netic beads. Leukocyte-free luteal cells exhibited greater steroido-genic responsiveness to hCG toward the end of the luteal phase. Thetreatment of mixed luteal cells (total luteal cells) with IL-1b inhibitedby 60% hCG-stimulated progesterone production. Interestingly, thetreatment of leukocyte-free luteal cells with IL-1b did not affect pro-gesterone production. In addition, the treatment of mixed luteal cellswith monoclonal antibodies against IL-1 receptor type I (IL-1RtI)resulted in a 2.5-fold increase in the hCG-supported progesteroneproduction. IL-1RtI and IL-1 receptor antagonist were localized by
immunohistochemistry in both somatic and immune cells of the CL.Flow cytometric analysis indicated that both nonleukocyte luteal cellsand leukocyte-luteal cells exhibited IL-1Rt-I positive cells, represent-ing 56% and 31% of the total luteal cells, respectively. However, 13%of nonleukocyte luteal cells did not express IL-1Rt-I. Northern anal-ysis demonstrated the presence of the 5.1-kb IL-1RtI messenger ri-bonucleic acid transcript in CL of different ages. RT-PCR indicatedthat both leukocyte-free luteal cells and luteal leukocytes expressIL-1RtI messenger ribonucleic acid. We conclude that 1) luteal leu-kocytes have an inhibitory effect on hCG-stimulated progesteroneproduction; 2) Il-1b inhibits hCG-stimulated progesterone productiononly in mixed luteal cell cultures, indicating that leukocytes mediatethe effect; 3) the somatic and immune cells of the CL are sites of actionand expression of the IL-1 system; and 4) interaction between thesteroidogenic and immune cells of the CL suggests a functional in-traovarian role for IL-1b in CL physiology. (J Clin Endocrinol Metab84: 4239–4245, 1999)
THE HUMAN corpus luteum (CL) undergoes remarkablechanges in structure and function from the time of
ovulation to its regression in a non-fertile cycle. During itsfunctional life the CL secretes the steroid hormone that areessential for maintenance of the secretory endometrium, im-plantation, and sustaining early pregnancy. Luteinization ofthe ovulatory follicle and steroidogenesis are dependent onpituitary-derived LH acting through a cAMP signaling cas-cade. Within luteal tissue, the steroidogenic actions of LH aremodulated by a variety of hormones, growth factors, andcytokines, including interleukin-1b (IL-1b), IL-1a, and tumornecrosis factor-a (TNFa) (1). These polypeptides are essentialto the immune cascade and stimulate or inhibit cell growthdepending on their concentrations and target cells. IL-1b isknown to inhibit luteinization of cultured murine and por-
cine granulosa cells (2, 3). Furthermore, IL-1b inhibited hCGstimulated-progesterone production by human granulosacells cocultured with white blood cells (4). It is thought thatIL-1b exerts its action through cell surface receptors. To datetwo IL-1 receptors (types I and II) have been identified. In thehuman, the type I receptor (IL-1RtI) possesses a 213-aminoacid long intracytoplasmic domain, whereas the type II re-ceptor has a short 29-residue intracytoplasmic domain (5).Current information indicates that IL-1b signaling occursexclusively via IL-1RtI (6). Cells regulated by IL-1b also pro-duce a IL-1 receptor antagonist (IL-1Ra). The complete IL-1bsystem including genes encoding IL-1b, IL-1RtI, and IL-IRahave been detected in the ovary of several species, includingthe human (7).
Interestingly, cultured human granulosa and thecal cellsdid not express messenger ribonucleic acid (mRNA) forIL-1b under basal conditions, but they did express mRNA forIL-1RtI (7). Treatment of ovarian cell cultures with forskolininduced IL-1b transcripts, but only in granulosa cells. Inaddition, IL-1-like activity has been detected in human fol-licular fluid, and the IL-1b transcript is more abundant incells of follicular aspirates than IL-1a. An intermediary rolefor IL-1 in the ovulatory process has been postulated (5).
Received May 25, 1999. Revision received August 12, 1999. AcceptedAugust 17, 1999.
Address all correspondence and requests for reprints to: Luigi DevotoM.D., University of Chile School of Medicine, P.O. Box 226–3, Santiago,Chile. E-mail: [email protected].
* This work was supported by FONDECYT Grant 196-1175 and agrant from Cooperacion Ibero-Americana.
† These authors contributed equally to this work.
0021-972X/99/$03.00/0 Vol. 84, No. 11The Journal of Clinical Endocrinology & Metabolism Printed in U.S.A.Copyright © 1999 by The Endocrine Society
4239

The cells comprising the CL have distinct morphological,endocrine, and biochemical properties. Cells present in theCL include small and large luteal cells, fibroblasts, and en-dothelial and immune cells. These cells change in number,morphology, function, and secretory capability throughoutthe life span of the CL (8, 9). Cells derived from the CLproduce IL-1b in culture, particularly immune cells (10).However, there is limited information regarding the role ofIL-1b and the cell localization of the IL-I system in the humanCL. In the present investigation we examined the ability ofIL-1b to modulate basal and hCG-stimulated progesteroneproduction by human luteal cells. Furthermore, studies werecarried out to elucidate the types of luteal cells that expressIL-1RtI and IL-1Ra in the human CL.
Materials and Methods
All chemicals, culture media, and hormones were obtained fromSigma Chemical Co. (St. Louis, MO), or from Worthington BiochemicalCorp. (Freehold, NJ). Reagents for progesterone RIA were obtained fromthe Human Reproduction Program of the WHO.
Antibodies
The following mouse monoclonal antibodies (mAb) were gifts fromDr. Maria Rosa Bono (Faculty of Sciences, University of Chile, Santiago,Chile): mAb W6/32 (IgG2a antihuman major histocompatibility com-plex class I) and mAb GAP8.3 (IgG2a anti-CD45). Dr. Francisco Sanchez-Madrid (The Princess Hospital, Madrid, Spain) provided mAb Bear-I(IgG1 anti-CD 11b) and mAb LIA 3/2 (IgG1 anti-CD 18). Mouse mAbTuk4 (IgG1 anti-CD14) was purchased from (DAKO Corp., Carpenteria,CA), the labeled F(ab9)2 antibody (antimouse IgG) was obtained fromImmunotech (Marseille, France), the mouse mAb (antihuman IL-1RtI)was purchased from Genzyme Transgenics Corp. (Cambridge, MA), andrat mAb 4 C1 (IgC2a antihuman IL-1RtI) was obtained from PharMingen(San Diego, CA).
Patients and tissues
CL were obtained from 16 eumenorrheic women, aged 27–38 yr,undergoing laparotomy for tubal sterilization (n 5 10) or myomas (n 56). at the San Borja Arriaran Clinical Hospital, University of Chile,National Health Service (Santiago, Chile). None of the patients hadexperience infertility or endometriosis and had stopped any hormonalcontraception at least 3 months before surgery. The study was approvedby the institutional review board of the University of Chile. All patientsgave their informed consent for removal of the CL.
After lutectomy, the tissue was transported to the laboratory in sterilesaline. The cycle date of each woman was confirmed by endometrialbiopsy (11) and classified as early (1–4 days; n 5 5), mid (5–9 days; n 56), or late (10–14 days; n 5 5). One piece of each CL was placed in 10%formalin for histological examination (12).
Cell dispersion
Luteal cells were dispersed as previously described (13). Briefly, CLwere enzymatically dissociated in culture medium 199/HEPES (25mmol/L) containing NaHCO3 (26 nmol/L), penicillin (50 IU/mL), col-lagenase trypsin-free (740 IU/100 mg tissue), and deoxyribonuclease (14kallikrein inhibitor units/100 mg tissue). After 90 min, luteal cells werewashed once with culture medium and twice with Dulbecco’s Ca21- andMg21-free phosphate-buffered saline (PBS) containing ethylenediaminetetraacetate (1 mmol/L). Red blood cells were removed by Histopaquecolumn (density, 1.007). The cell viability was more than 90%, as as-sessed by the trypan blue exclusion method.
Luteal leukocyte isolation
After luteal dispersion, a fraction of luteal cells was incubated witha panel of mouse mAbs (GAP 8.3, Bear-1, and LIA 3/2) directed to
leukocyte-specific molecules as previously described (10). Leukocyteswere separated from luteal cells in a magnetic concentrator (DynAl,Chantilly, VA). Subsequent to isolation of luteal leukocytes, the cellviability of luteal cells was greater than 90%, as determined by the trypanexclusion method.
Cell culture
Luteal cells were cultured as previously described (13, 14). Briefly,mixed luteal cells (total luteal cells) and leukocyte-free luteal cells (0.5–1x105/well) were cultured in serum-free medium and treated with IL-1b(25 ng/mL) in the absence and presence of hCG (10 IU/mL), for 24 h.To test the specificity of IL-1b, some cells were treated with anti IL-1RtImAb in the presence and absence of hCG. Culture medium was collectedfor determination of progesterone concentrations.
Human granulosa cells were isolated from patients undergoing invitro fertilization/embryo transfer. The cells were cultured as previouslydescribed (19) and used to detect IL-1RtI by indirect immunofluorescence.
Detection of IL-1RtI by flow cytometry
To investigate the protein expression of IL-1RtI on mixed luteal cells,flow cytometric analysis was performed as described previously withminor modifications (10). Briefly, dual color immunofluorescence stain-ing was performed to discriminate the expression level of IL-1RtI onluteal leukocytes and leukocyte-free luteal cells. In the first step, sus-pensions of mixed luteal cells were simultaneously incubated for doublestaining with the rat mAb 4C1 (anti IL-1RtI) and the panel of mouseantileukocyte mAbs for 40 min at 4 C, and then washed in PBS. Thesecond step included incubation with biotin-conjugated goat antirat Igpolyclonal antibody for 30 min at 4 C, followed by two washes. In thethird step, the cells were incubated with the conjugate streptavidin-phycoerythrin and fluorescein-F(ab)2 anti-mouse Ig for 30 min at 4 C.After two washes, the cells were resuspended at a concentration of 0.6 3106/mL in PBS and immediately analyzed or stored overnight in PBScontaining 0.5% formaldehyde in the dark until analysis. Control sam-ples included assessment of autofluorescence and negative controls(absence of primary mAbs and/or secondary mAbs). No cross-reactionwas observed between secondary antibodies, indicating highly specificbinding of the mouse or rat Ig.
RNA isolation and Northern analysis
The presence of IL-1RtI mRNA was determined in CL collectedthroughout the luteal phase. Corpora lutea were kept frozen at 270 C.Total RNA was prepared as described by Chromczynski and Sacchi (15)and was quantified by absorbance at 260 nm. Northern analysis wascarried out using Il-1RtI complementary DNA (cDNA) as a probe (Im-munex Corp., Seattle, WA). Briefly, 15 mg total RNA were resolved ona 1% formaldehyde-agarose gel, blotted onto a nylon membrane (Schlei-cher & Schuell, Inc., Nytran), and cross-linked by UV irradiation. Themembranes were prehybridized for 1 h at 65 C in a solution containing0.5 mol/L NaH2PO4 (pH 7), 0.1 mmol/L ethylenediamine tetraacetate,0.5% BSA, and 7% SDS. Hybridization was carried out overnight at 65C in the same solution after the addition of the 32P-labeled probe (106
cpm/mL solution). Membranes were washed in 2 3 SSC (standardsaline citrate)-0.1% SDS for 30 min at room temperature, followed by twowashes in 0.1 3 SSC-0.1% SDS for 30 min at 60 C. Blots were exposedto x-ray film (reflection NEF-496, DuPont/NEN, Boston, MA), for 72 h.
RT-PCR
The presence of IL-1RtI transcripts in mixed luteal cells and leuko-cyte-free luteal cells was determined by RT-PCR. Oligonucleotide prim-ers were designed according to previously published sequences of IL-1RtI and IL-1Ra, predicting PCR products of 441 and 511 bp, respectively(16, 17). Primer sequences were as follows: IL-1Rt1: forward, 59-GCCAAG AGT TCT TTA GGT GCC-39; reverse, 59-CTC ACT GCA ACC TCCGTC TC-39; and IL-1Ra: forward, 59-CAG AAG ACC TCC TGT CCTATG AGG-39; reverse, 59-GAT GAG CAG GAG GAC CTT CAT-39. TotalRNA (5 mg) was reverse transcribed in a 20-mL reaction containing 10mmol/L reverse primer, AmpliTaq buffer, 0.2 mmol/L of each deoxy-NTP, 200 U reverse transcriptase (Superscript II, Life Technologies, Inc.,
4240 KOHEN ET AL. JCE & M • 1999Vol 84 • No 11
Gaithersburg, MD), and 5 U RNasin (Promega Corp., Madison, WI) at42 C for 60 min. The PCR amplifications were performed by adding 10mmol/L forward primer, Taq buffer, 0.2 mmol/L of each deoxy-NTP,and 1 U Taq DNA polymerase (Promega Corp.) to the previous reactionmixture in a final volume of 50 mL. The cycling conditions for IL-1RtIwere as follows: initial denaturation at 95 C for 5 min, followed by 30cycles of denaturation at 95 C for 1 min, annealing at 58 C for 1 min,extension at 72 C for 1 min, and a final extension at 72 C for 10 min.Cycling conditions for IL-1Ra were the same, except for the annealingtemperature, which was 55 C. Control reactions without cDNA werecarried out in parallel and resulted in negative determination. PCRproducts were analyzed by electrophoresis in 1.25% agarose gel. PurifiedPCR products were sequenced using the DNA ds Cycle SequencingSystem (Life Technologies, Inc.).
Nested PCR for IL-1Ra
IL-1Ra product obtained from RT-PCR was submitted to a secondPCR amplification (nested), using the following specific primers: for-ward, 59-CAG AAG ACC TCC TGT CCT ATG AGG-39; and reverse,59-GAT GAG CAG GAG GAC CTT CAT-39. Cycling conditions were thesame as described for the first PCR reaction, except for the annealingtemperature, which was 60 C. Purified PCR products were sequencedusing the DNA ds Cycle Sequencing System (Life Technologies, Inc.).
Immunohistochemical staining procedures
CL were formalin-fixed and paraffin embedded. Five-micron sectionswere processed for routine hematoxylin-eosin staining and immuno-histochemistry as previously described (18). Briefly, the slides weredeparaffinized in xylene, followed by rapid rehydration through agraded series of alcohols. For the avidin-biotin-peroxidase method, en-dogenous peroxidases were blocked with 1% H2O2 in 96% methanol.Nonspecific binding was blocked with horse serum followed by incu-bation with the primary antibodies: monoclonal mouse antihuman IL-1RtI and Il-1Ra antibody, respectively (Genzyme Corp., Cambridge,MA). Control incubations included deletion of the primary antibody.After rinsing with PBS, sections were incubated with a secondary IgGantibody. Immunoreaction products were visualized by incubating sec-tions with 39-diaminobenzidine tetrahydrochloride as substratesolution.
Indirect immunofluorescence procedures for lutein andgranulosa lutein cells
Lutein or luteinized granulosa cells grown on coverslips were washedtwice in prewarmed (37 C) DMEM and twice in prewarmed PBS con-taining 1.5 mmol/L Ca21 and fixed in 100% methanol (220 C for 5 min).Cells were incubated in 10% normal goat serum for 30 min at roomtemperature and then with 10 mg/mL of the primary antibodies for 2 hat room temperature (18, 19). A fluorescein-conjugated goat antimousesecondary antibody (Jackson ImmunoResearch Laboratories, Inc., WestGrove, PA) was used at a 1:200 dilution (30 min at room temperature).Coverslips were mounted on glass slides with Fluoromount G (FisherScientific, Malvern, PA) containing 1,4-diazabicyclo-(2,2,2)octane(DABCO, Polysciences, Inc., Warrington, PA) to stabilize fluorescenceand were photographed with a Nikon microscope (Melville, NY).
Data analysis
All experiments were conducted in duplicate and repeated at leastthree times. Experimental data are presented as the mean 6 se. Thepercent increment in progesterone production was analyzed by two-tailed Student’s t test for comparison between control and treated con-ditions and between different luteal cell populations. P , 0.05 wasconsidered statistically significant.
Results
Figure 1 illustrates the effect of hCG (10 IU/mL) on pro-gesterone accumulation by cultured mixed luteal cells andleukocyte-free luteal cells. The time course of hCG stimu-
lated-progesterone accumulation was examined in lutealcells of different ages. Culture medium from mid and lateleukocyte-free luteal cells displayed higher progesterone ac-cumulation compared with that from mixed luteal cells, re-spectively (P , 0.05). No difference was noted in the pro-gesterone accumulation by mixed luteal and leukocyte-freeluteal cells from early CL. These data suggest a luteal age-dependent interaction among cAMP signaling, luteal steroi-dogenic cells, and luteal leukocytes.
Figure 2 depicts the effect of IL-1b (25 ng/mL) in thepresence and absence of hCG on progesterone accumulationby mixed and leukocyte-free early luteal cells. IL-1b had noeffect on basal progesterone production by either mixed lu-teal cells or leukocyte-free luteal cells. In contrast, treatmentwith IL-1b inhibited hCG-stimulated progesterone accumu-lation in mixed luteal cells by 60%,(P , 0.05), but did notaffect leukocyte-free luteal cells. This finding suggests thatthe action of IL-1b is leukocyte mediated.
FIG. 1. Histogram of progesterone production by human mixed andleukocyte-free luteal cells from CL of different ages. The cells weretreated with hCG for 24 h. Culture of luteal cells and progesteronedeterminations were performed as described in Materials and Meth-ods. Values for progesterone are the mean 6 SEM from four to sevenseparate CL. *, P , 0.05 compared to mixed luteal cells.
FIG. 2. Histogram of progesterone production by mixed and leuko-cyte-free luteal cells from early CL. The cells were treated for 24 hwith hCG, IL-1b, and IL-1b plus hCG. Culture of luteal cells andprogesterone determinations were performed as described in Mate-rials and Methods. Values for progesterone are the mean 6 SEM fromfive individual CL. *, P , 0.05 compared to leukocyte-free luteal cells.
IL SYSTEM, IN HUMAN CORPUS LUTEUM 4241
To verify the specific effect of IL-1b on luteal steroido-genesis, the neutralizing 4C1 mAb directed to IL-1RtI wasadded to the mixed mid luteal cells (8 mg 3 105 cells) for 24 h.Fig. 3 illustrates that hCG-stimulated progesterone accumu-lation by mixed luteal cells was amplified 2-fold in the pres-ence of IL-1RtI mAb compared with that in the presence ofhCG alone (P , 0.05). Treatment with IL-1RtI mAb had noeffect on basal progesterone accumulation or hCG-stimu-lated progesterone accumulation in leukocyte-free lutealcells (data not shown). These findings suggest that the actionof IL-1b on steroidogenesis is mediated by the leukocyteIL-1RtI.
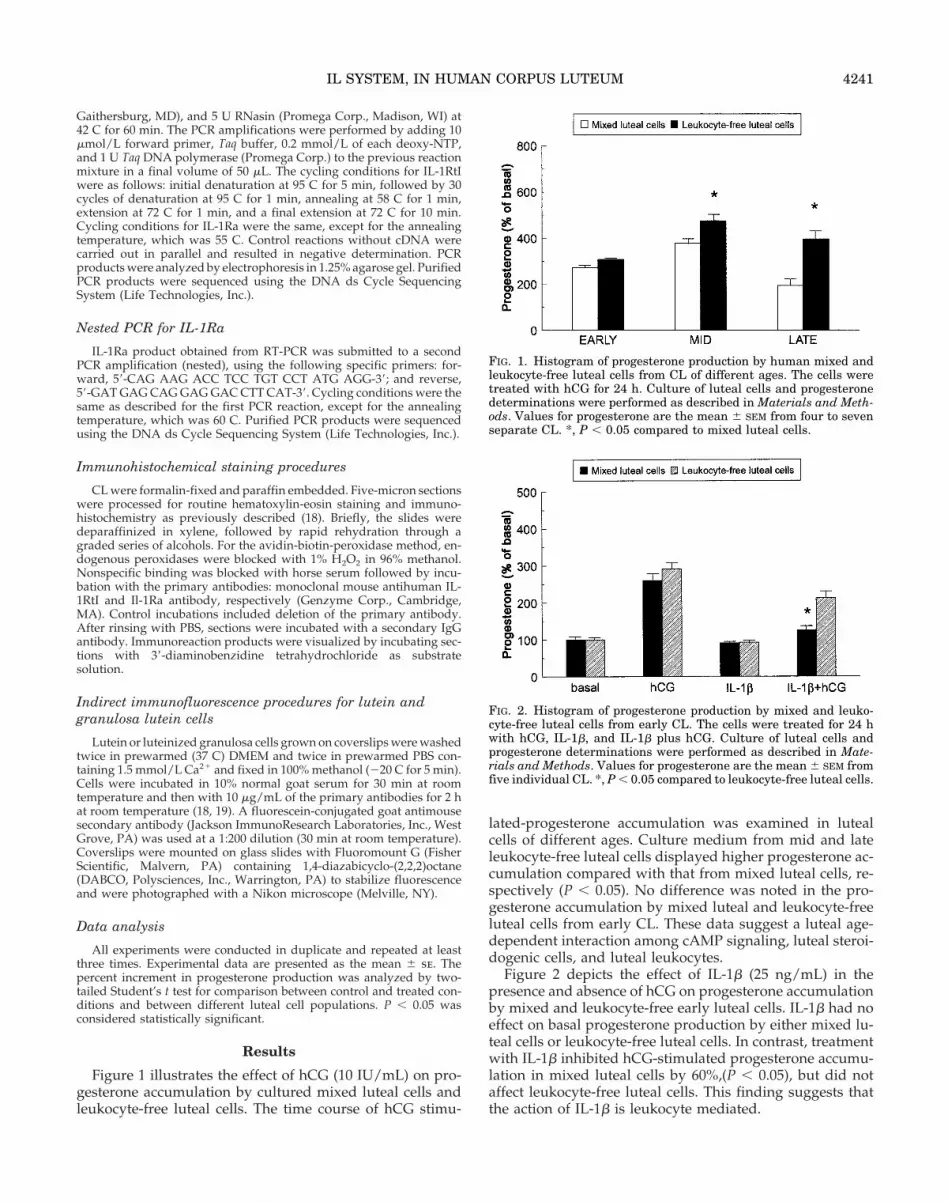
Figure 4, the upper panel shows the immunohistochemicalidentification of IL-1RtI in CL collected during the midlutealphase. Immunostaining was localized in the cytoplasmicplasma membrane of luteal cells (Fig. 4A). In control exper-iments, no immunostaining was noted when primary anti-body was omitted (Fig. 4B). The middle panel shows luteinizedgranulosa cells in primary culture (Fig. 4C, immunofluores-cence; Fig. 4D, phase contrast). The cells displayed numerouscell to cell contacts, and the cell morphology is consistentwith steroidogenic cells. The IL-1RtI protein was visualizedin leukocyte-free granulosa lutein cells by indirect immuno-fluorescence. These cells exhibited intense staining for IL-1RtI. However, we noticed that some of these cells did notstain for IL-1RtI, suggesting that IL-1RtI is not constitutivelypresent in all granulosa-lutein cells. The lower panel shows theindirect immunofluorescence for IL-1RtI in luteal cells.Mixed luteal cells (Fig. 4E) as well as leukocyte-free lutealcells (Fig. 4F) displayed intense staining for IL-1RtI. Thenegative control included the omission of the primary anti-body and showed no staining (Fig. 4G).
IL-1RtI gene expression in the corpus luteum
To determine whether the CL is a site of IL-1RtI geneexpression during the luteal phase, Northern blot was per-formed using IL-1RtI cDNA as a probe. Figure 5 is an au-toradiogram that illustrates the presence of the IL-1RtImRNA transcript in CL (total tissue) of different stages. The
cDNA probes yield a major band of 5.1 kb in the differentstages of the CL. Secretory endometrium was used as apositive control. These data indicate that IL-1RtI mRNA isexpressed during all stages of the luteal phase.
FIG. 4. A, Immunohistochemical staining of IL-1RtI is observed inthe cytoplasmic membrane of luteal cells from section of mid-CL(magnification, 3400). B, Control section from mid-CL (magnifica-tion, 3100). C, Indirect immunofluorescence identified the IL-1RtI inhuman luteinized granulosa cells in primary culture (magnification,3400). D, Phase contrast microscopy of human granulosa cells inprimary culture (magnification, 3400). E and F, Indirect immuno-fluorescence of midluteal mixed and leukocyte-free luteal cells, re-spectively (magnification, 3400). G, Control mixed luteal cells (mag-nification, 3100).
FIG. 3. Histogram of progesterone production by human mixed lutealcell from mid-CL. The cells were treated with hCG, anti-IL-1RtI mAb,and hCG plus IL-1RtI mAb for 24 h. The mAb was preincubated andadministered immediately with hCG. Values for progesterone are themean 6 SEM from three independent CL. *, P , 0.05 vs. basal; **, P ,0.05 vs. basal and hCG
4242 KOHEN ET AL. JCE & M • 1999Vol 84 • No 11

IL-1RtI in luteal leukocytes and in nonleukocyte luteal cells
RT-PCR was used to determine whether IL-1RtI is ex-pressed in both luteal leukocytes and nonleukocyte lutealcells. Figure 6 depicts a 441-bp PCR product correspondingto the expected size of IL-1RtI mRNA and was found inmixed luteal cells, leukocyte-free luteal cells, and luteal leu-kocytes. All products obtained were sequenced, confirmingindividual product identity. Secretory endometrial cell RNAwas used as a positive control, and peripheral leukocytes thatdo not express the IL-RtI mRNA were used as a negativecontrol. In all experiments b-actin was used as a control forsample to sample variation.
Immunohistochemical identification and mRNA expressionof IL-1Ra in the CL
Figure 7 shows the immunohistochemical identification ofIL-1Ra from CL collected during the midluteal phase. The
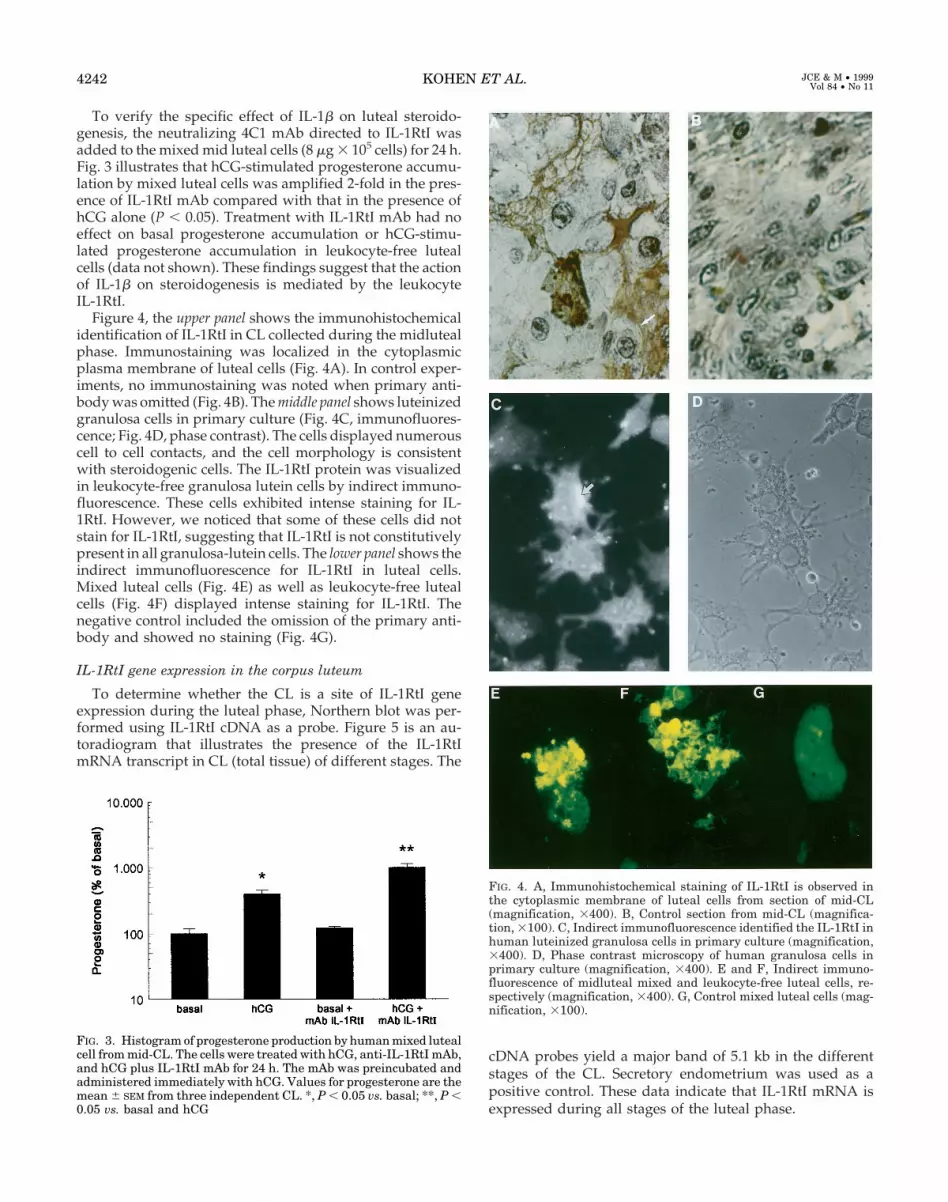
immunostaining was localized in the cytoplasmic membraneof luteal cells (Fig. 7A). In control experiments, no immu-nostaining was noted when primary antibody was omitted(Fig. 7B). The lower panel shows the indirect immunofluo-rescence for IL-1Ra in luteal cells. Mixed luteal cells (Fig. 7C)as well as leukocyte-free luteal cells (Fig. 7D) displayed in-tense staining for IL-1Ra. The negative control included theomission of the primary antibody and shows no staining (Fig.7E).
Figure 8 illustrates the RT nested-PCR of IL-1Ra mRNA inmixed luteal cells. A 551-bp PCR product corresponding tothe expected size of IL-1Ra mRNA was found in mixed lutealcells. The product obtained was sequenced, confirming prod-uct identity.
Flow cytometric analysis
We have previously validated by flow cytometry the pu-rity of the leukocyte-free luteal cell population as greaterthan 99% (10). In this study, flow cytometric analysis as-sessed the level and cellular distribution of IL-1RtI in the CL.Figure 9 shows the simultaneous detection of IL-1RtI in lutealleukocytes as well as in leukocyte-free luteal cells. The upperpanel illustrates cells that stain positive for IL-1RtI. The dotsplotted in the upper right quadrant represent leukocytes, cor-responding to 31% of the positive cells. The dots plotted in the
FIG. 5. IL-1RtI mRNA determination from human CL of differentages by Northern analysis. Fifteen micrograms of total RNA weresubmitted to electrophoresis on agarose-formaldehyde gels and an-alyzed as described in Materials and Methods. The IL-1RtI signal of5.1 kb is present in all CL analyzed, as shown in the upper panel.Secretory endometrium represents the positive control. The lowerpanel shows the 18S ribosomal RNA as an indication of the relativeamounts of RNA loaded in each lane.
FIG. 6. IL-1RtI mRNA determination from luteal cells, luteal leuko-cytes, and leukocyte-free luteal cells by RT-PCR. RT-PCR productswere submitted to electrophoresis on 1.25% agarose gels. A productof 441 bp is shown in each lane, including a positive control (1)corresponding to secretory endometrium mRNA. The negative control(control 2) corresponds to peripheral leukocytes. The lower panelshows RT-PCR products for b-actin (376 bp) obtained from the sameRNA preparations. M, pBR322 digested with HinfI and used as a sizemarker.
FIG. 7. A, Immunohistochemical staining of IL-1Ra is observed in thecytoplasmic membrane of luteal cells from a section of mid-CL (mag-nification, 3400). B, Control section from a mid-CL (magnification,3100). C and D, Indirect immunofluorescence of midluteal mixed andleukocyte-free luteal cells, respectively (magnification, 3400). E, Con-trol mixed luteal cells (magnification, 3100).
IL SYSTEM, IN HUMAN CORPUS LUTEUM 4243
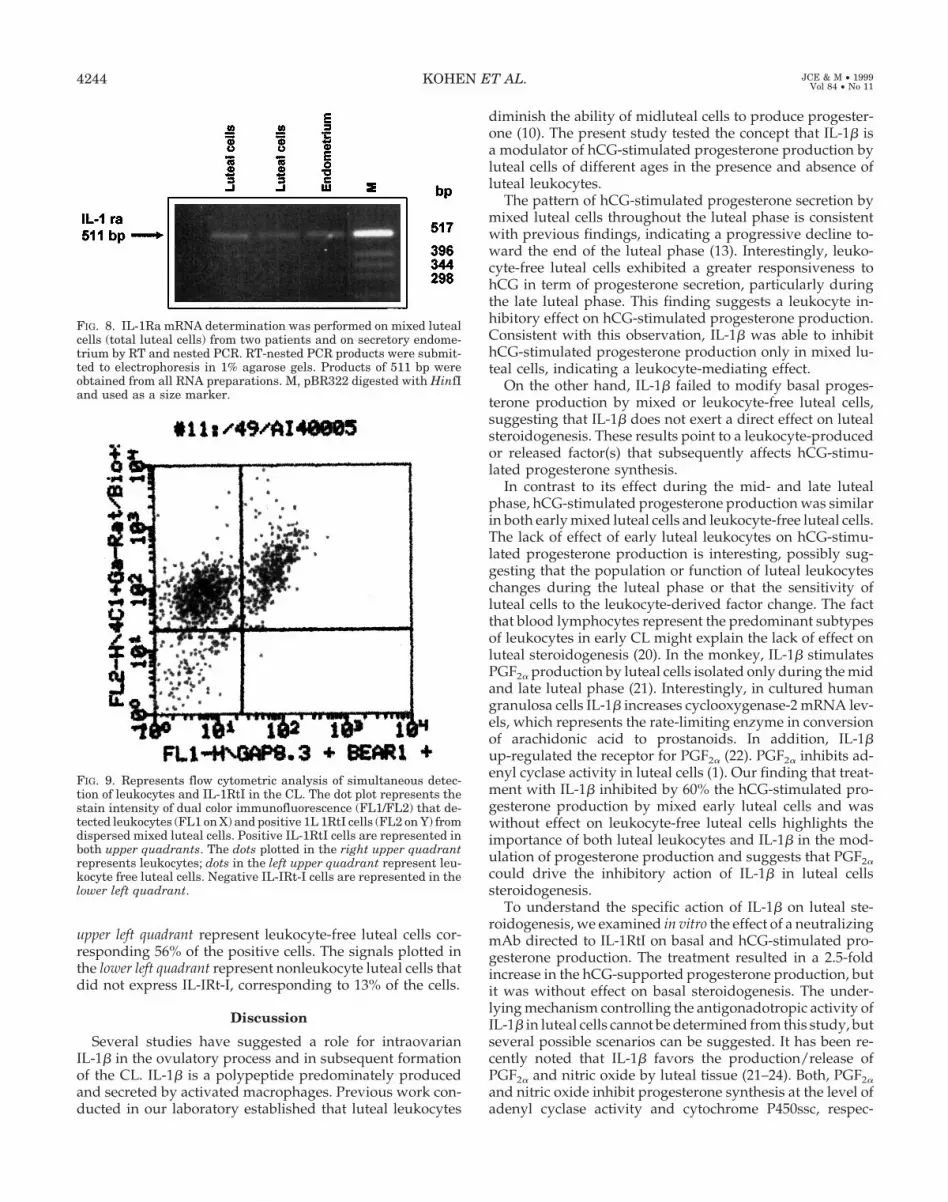
upper left quadrant represent leukocyte-free luteal cells cor-responding 56% of the positive cells. The signals plotted inthe lower left quadrant represent nonleukocyte luteal cells thatdid not express IL-IRt-I, corresponding to 13% of the cells.
Discussion
Several studies have suggested a role for intraovarianIL-1b in the ovulatory process and in subsequent formationof the CL. IL-1b is a polypeptide predominately producedand secreted by activated macrophages. Previous work con-ducted in our laboratory established that luteal leukocytes
diminish the ability of midluteal cells to produce progester-one (10). The present study tested the concept that IL-1b isa modulator of hCG-stimulated progesterone production byluteal cells of different ages in the presence and absence ofluteal leukocytes.
The pattern of hCG-stimulated progesterone secretion bymixed luteal cells throughout the luteal phase is consistentwith previous findings, indicating a progressive decline to-ward the end of the luteal phase (13). Interestingly, leuko-cyte-free luteal cells exhibited a greater responsiveness tohCG in term of progesterone secretion, particularly duringthe late luteal phase. This finding suggests a leukocyte in-hibitory effect on hCG-stimulated progesterone production.Consistent with this observation, IL-1b was able to inhibithCG-stimulated progesterone production only in mixed lu-teal cells, indicating a leukocyte-mediating effect.
On the other hand, IL-1b failed to modify basal proges-terone production by mixed or leukocyte-free luteal cells,suggesting that IL-1b does not exert a direct effect on lutealsteroidogenesis. These results point to a leukocyte-producedor released factor(s) that subsequently affects hCG-stimu-lated progesterone synthesis.
In contrast to its effect during the mid- and late lutealphase, hCG-stimulated progesterone production was similarin both early mixed luteal cells and leukocyte-free luteal cells.The lack of effect of early luteal leukocytes on hCG-stimu-lated progesterone production is interesting, possibly sug-gesting that the population or function of luteal leukocyteschanges during the luteal phase or that the sensitivity ofluteal cells to the leukocyte-derived factor change. The factthat blood lymphocytes represent the predominant subtypesof leukocytes in early CL might explain the lack of effect onluteal steroidogenesis (20). In the monkey, IL-1b stimulatesPGF2a production by luteal cells isolated only during the midand late luteal phase (21). Interestingly, in cultured humangranulosa cells IL-1b increases cyclooxygenase-2 mRNA lev-els, which represents the rate-limiting enzyme in conversionof arachidonic acid to prostanoids. In addition, IL-1bup-regulated the receptor for PGF2a (22). PGF2a inhibits ad-enyl cyclase activity in luteal cells (1). Our finding that treat-ment with IL-1b inhibited by 60% the hCG-stimulated pro-gesterone production by mixed early luteal cells and waswithout effect on leukocyte-free luteal cells highlights theimportance of both luteal leukocytes and IL-1b in the mod-ulation of progesterone production and suggests that PGF2a
could drive the inhibitory action of IL-1b in luteal cellssteroidogenesis.
To understand the specific action of IL-1b on luteal ste-roidogenesis, we examined in vitro the effect of a neutralizingmAb directed to IL-1RtI on basal and hCG-stimulated pro-gesterone production. The treatment resulted in a 2.5-foldincrease in the hCG-supported progesterone production, butit was without effect on basal steroidogenesis. The under-lying mechanism controlling the antigonadotropic activity ofIL-1b in luteal cells cannot be determined from this study, butseveral possible scenarios can be suggested. It has been re-cently noted that IL-1b favors the production/release ofPGF2a and nitric oxide by luteal tissue (21–24). Both, PGF2a
and nitric oxide inhibit progesterone synthesis at the level ofadenyl cyclase activity and cytochrome P450ssc, respec-
FIG. 8. IL-1Ra mRNA determination was performed on mixed lutealcells (total luteal cells) from two patients and on secretory endome-trium by RT and nested PCR. RT-nested PCR products were submit-ted to electrophoresis in 1% agarose gels. Products of 511 bp wereobtained from all RNA preparations. M, pBR322 digested with HinfIand used as a size marker.
FIG. 9. Represents flow cytometric analysis of simultaneous detec-tion of leukocytes and IL-1RtI in the CL. The dot plot represents thestain intensity of dual color immunofluorescence (FL1/FL2) that de-tected leukocytes (FL1 on X) and positive 1L 1RtI cells (FL2 on Y) fromdispersed mixed luteal cells. Positive IL-1RtI cells are represented inboth upper quadrants. The dots plotted in the right upper quadrantrepresents leukocytes; dots in the left upper quadrant represent leu-kocyte free luteal cells. Negative IL-IRt-I cells are represented in thelower left quadrant.
4244 KOHEN ET AL. JCE & M • 1999Vol 84 • No 11

tively. These findings are consistent with the recent report byBreard et al., who found that IL-1b diminished hCG-stimu-lated cAMP generation in rabbit granulosa cells (25). Col-lectively, these data support the idea of a functional role forIL-1b in human CL regression.
The second part of our study was designed to assess themRNA expression and cellular localization of IL-1RtI andIL-1Ra within the CL. Immunoreactive IL-1RtI, and geneexpression (mRNA) were localized in both somatic cells andimmune cells of the gland. This is consistent with our flowcytometric assessment that detected IL-1RtI protein in bothcell types. In addition, our data indicate that 56% of cells thatstain positively for IL-1RtI are nonimmunological luteal cells.These results are in agreement with those of Piquette et al.(26), who reported that human granulosa-lutein cells col-lected at the time of oocyte retrieval, and not macrophages,account for the majority of the immunohistochemical stain-ing for IL-1RtI.
Northern blot analysis documented the presence of singlemRNA transcript for IL-1RtI (5.1 kb) in the CL. Furthermore,this mRNA transcript is present in CL of different ages.Cellular localization of a fragment of the IL-1RtI transcriptwas performed by RT-PCR. The 441-bp RT-PCR productcorresponded to the amplified IL-1RtI fragment and is lo-calized in leukocyte-free luteal cells and luteal leukocytes.These data indicate that the CL is a site of IL-1RtI geneexpression and IL-1b action.
IL-1 exerts its action through cell surface receptors. Anaturally occurring antagonist, IL-1Ra, is also produced bythe cells to modulate the actions of IL-1, preventing postre-ceptor activation (20). Our observations documented thepresence of immunoreactive IL-1Ra in histological section ofthe CL throughout the luteal phase. Furthermore, immuno-fluorescence procedures identified IL-1Ra in luteal leuko-cytes and leukocyte-free luteal cells. The IL-1Ra mRNA wasonly examined in mixed luteal cells. We have previouslyshown the ability of human luteal cells to produce IL-1b (9).Taken together these findings indicate the presence of acomplete intraluteal IL-1 system in the CL, replete with li-gand, receptor, and receptor antagonist.
In summary, this study demonstrated localization of theIL-1 system in both immune and somatic cells of the humanCL. In addition, our findings reveal a functional role for IL-1bin the regulation of luteal steroidogenesis in vitro. The fac-tor(s) that mediates the antigonadotropic action of IL-1b onhCG-stimulated progesterone secretion is mainly leukocytedependent. The lack of effect of IL-1b on progesterone syn-thesis in leukocyte-free luteal cells suggests a nonsteroido-genic role for IL-1RtI localized in somatic luteal cells.
Finally, these findings highlight the importance of residentmacrophages in the regulation of CL development, function,and regression and suggest an interaction between the im-mune and steroidogenic cells within the corpus luteum.
Acknowledgments
The authors gratefully acknowledge Lane Christenson, Ph.D., andProf. J. F. Strauss III, M.D., Ph.D., from the University of Pennsylvaniafor their critical revision of the manuscript.
References
1. Terranova PF, Montgomery R. 1997 Review: cytokine involvement in ovarianprocesses. Am J Reprod Immunol. 37:50–60.
2. Gottschall PE, Aehara A, Hoffmabb ST, et al. 1987 Interleukin-1b inhibitsfollicle stimulating hormone-induced differentiation in rat granulosa cells invitro. Biochem Biophys Res Commun. 149:502–509.
3. Fukuoka MK, Yasuda S, Taiik M, et al. 1989 Interleukin-I stimulates growthand inhibits progesterone secretion in cultures of porcine granulosa cells.Endocrinology 124:884–890.
4. Best C, Hill J. 1995 Interleukin-1a and -b modulation of luteinized humangranulosa cell oestrogen and progesterone biosynthesis. Hum Reprod.10:3206–3210.
5. Adashi EY. 1996 Immune modulators in the context of ovulatory process: a rolefor interleukin-1. Am J Reprod Immunol. 35:190–199.
6. Sims JE, Gaylema, Slack JL, et al. 1993 Interleukin-1 signaling occurs exclu-sively via the type I receptor. Proc Natl Acad Sci USA. 90:5155–6159.
7. Hurwitz A, Loukides J, Ricciarelli, et al. 1992 Human intraovarian interleu-kin-1 (IL-1) system: highly compartmentalized and hormonally dependentregulation of the genes encoding IL-1, its receptor, and its receptor antagonist.J Clin Invest. 89:1746–1754.
8. Carrasco I, Troncoso JL, Devoto L, et al. 1996 Differential steroidogenic re-sponse of human luteal cell subpopulations. Hum Reprod. 11:1609–1614.
9. Retamales I, Carrasco I, Troncoso JL, et al. 1994 Morpho-functional study ofhuman luteal cells subpopulations. Hum Reprod. 9:591–596.
10. Castro A, Castro O, Troncoso JL, et al. 1998 Luteal leukocytes are modulatorsof the steroidogenic process of human mid-luteal cells. Hum Reprod.13:1584–89.
11. Noyes RW, Hertig AT, Rock J. 1950 Dating the endometrial biopsy. FertilSteril. 1:3–12.
12. Corner GW. 1956 The hitological dating of human corpus luteum of men-struation. Am J Anat. 98:377–394.
13. Devoto L, Kohen P, Castro O, et al. 1995 Multihormonal regulation of pro-gesterone synthesis in cultured human midluteal cells. J Clin EndocrinolMetab. 80:1566–1570.
14. Vega M, Devoto L, Castro O, et al. 1994 Progesterone synthesis by humanluteal cells: modulation by estradiol. J Clin Endocrinol Metab. 79:941–945.
15. Chromczynki P, Sacchi N. 1987 Single-step method of RNA isolation by acidguanidium thiocynate-phenol-cloroform extraction. Anal Biochem.162:156–159.
16. Sims JE, Acres RB, Grubin CE, et al. 1989 Cloning the interleukin 1 receptorfrom human T cells. Proc Natl Acad Sci USA. 86:8946–8950.
17. Haskill S, Martin G, Van Le L, et al. 1991 CDNA cloning of an intracellularform of the human interleukin 1 receptor antagonist associated with epithe-lium. Proc Acad Sci USA. 88:3681–3685.
18. Simon C, Frances A, Piquette G, et al. 1994 Interleukin-1 system in thematerno-trophoblast unit in human implantation: immunohistochemical ev-idence for autocrine/paracrine function. J Clin Endocrinol Metab. 78:847–854.
19. Makrigiannakis A, Coukos G, Christofidou-Solomidou M, et al. 1999 N-Cadherin mediated human granulosa cell adhesion prevents apoptosis: a rolein follicular atresia and luteolysis. Am J Pathol. 154:1391–1406.
20. Norman RJ, Brannstrom M. 1996 Cytokines in the ovary. Pathophysiology andpotential for pharmacological intervention. Pharmacol Ther. 69:219–236.
21. Young JE, Friedman CI, Danforth. 1997 Inteleukin-1b modulates prostaglan-din and progesterone production by primate luteal cell in vitro. Biol Reprod.56:663–667.
22. Narko K, Ritvos O, Ristimaki A. 1997 Induction of cyclooxygenase-2 andprostaglandin F2a receptor expression by interleukin-1b in cultures humangranulosa-luteal cells. Endocrinology. 138:3638–3644.
23. Ellman C, Corbett JA, Misho TP, et al. 1993 Nitric oxide mediates interleukin-1induced cellular cytotoxity in the rat ovary. J Clin Invest. 92:3053–3056.
24. Vega M, Johnson MC, Diaz HA, et al. 1998 Regulation of human lutealsteroidogenesis in vitro by of nitric oxide. Endocrine 8:185–191.
25. Breard E, Delarue B, Benhaim A, et al. 1998 Inhibition by gonadotropins ofinterleukin-1 production by rabbit granuosa and theca cells: effects on gona-dotropin-induced progesterone production. Eur J Endocrinol. 138:328–336.
26. Piquette G, Simon C, el Danssouri I, et al. 1994 Gene regulation of interleukin-1b, interleukin-1 receptor type I, and plasminogen activator inhibitor-1 and -2in human granulosa-luteal cells. Fertil Steril 62:760–770.
IL SYSTEM, IN HUMAN CORPUS LUTEUM 4245