evolution of interleukin-1β
TRANSCRIPT

Cytokine & Growth Factor Reviews 13 (2002) 483–502
Survey
Evolution of interleukin-1�
Steve Birda, Jun Zoua, Tiehui Wanga, Barry Mundayb,Charlie Cunninghamc, Chris J. Secombesa,∗
a Department of Zoology, University of Aberdeen, Tillydrone Avenue, Aberdeen AB24 2TZ, UKb School of Human Life Sciences, University of Tasmania, Locked Bag 1-320, Launceston, Tasmania 7250, Australia
c Sars International Center for Marine Molecular Biology, 5008 Bergen, Norway
Abstract
All jawed vertebrates possess a complex immune system, which is capable of anticipatory and innate immune responses. Jawlessvertebrates posses an equally complex immune system but with no evidence of an anticipatory immune response. From these findings it hasbeen speculated that the initiation and regulation of the immune system within vertebrates will be equally complex, although very little hasbeen done to look at the evolution of cytokine genes, despite well-known biological activities within vertebrates. In recent years, cytokines,which have been well characterised within mammals, have begun to be cloned and sequenced within non-mammalian vertebrates, with thenumber of cytokine sequences available from primitive vertebrates growing rapidly. The identification of cytokines, which are mammalianhomologues, will give a better insight into where immune system communicators arose and may also reveal molecules, which are uniqueto certain organisms. Work has focussed on interleukin-1 (IL-1), a major mediator of inflammation which initiates and/or increases a widevariety of non-structural, function associated genes that are characteristically expressed during inflammation. Other than mammalian IL-1�sequences there are now full cDNA sequences and genomic organisations available from bird, amphibian, bony fish and cartilaginous fish,with many of these genes having been obtained using an homology cloning approach. This review considers how the IL-1� gene haschanged through vertebrate evolution and whether its role and regulation are conserved within selected non-mammalian vertebrates.© 2002 Elsevier Science Ltd. All rights reserved.
Keywords:Interleukin-1; Comparative immunology; Vertebrate; Gene evolution; ICE cut site
Contents
1. Evolution of immune system communication. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4832. Interleukin-1 family. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4853. IL-1 in vertebrates. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4874. Known vertebrate IL-1� sequences. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4885. Processing of IL-1� in vertebrates. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4906. Vertebrate IL-1� gene organisation. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4927. Vertebrate IL-1� expression and regulation. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4928. Vertebrate IL-1 receptors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4949. Interleukin-18. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 494
10. Cytokine and IL-1 evolution. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49511. Conclusion. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 496Acknowledgements. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 497References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 497
∗ Corresponding author. Tel.:+44-1224-272872;fax: +44-1224-272396.E-mail address:[email protected] (C.J. Secombes).
1. Evolution of immune system communication
The study of the immune system of non-mammalian ver-tebrates has become increasingly important in organismsthat have commercial value such as birds and fish. Due tothis work, the origin of immune system components havealso become the focus of investigation. It is now accepted,
1359-6101/02/$ – see front matter © 2002 Elsevier Science Ltd. All rights reserved.PII: S1359-6101(02)00028-X

484 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
Table 1Cytokines cloned and sequenced from non-mammalian vertebrates (excluding IL-1�)
Organism Gene GenBank accession number Reference
BirdsGallus gallus(Chicken) IL-2 AF000631 [10]
IL-6 AJ309540 [11]IL-8 AJ009800 [12]IL-15 AF139097 [13]IL-18 AJ277865 [14]
IFN-� X92476 [15]IFN-� X92479 [15]IFN-� X99774 [16]
FGF-2 M95707 [17]FGF-3 Z47555 [18]FGF-4 U14654 [19]FGF-8 U55189 [20]FGF-10 D86333 [21]FGF-12 AF108754 [22]FGF-13 AF108755 [22]FGF-18 AB030229 [23]FGF-19 AF315355 [24]
TGF-�2 X58071 [25]TGF-�3 M31154 [26]TGF-�4 M31160 [27]
MIF M95776 [28]
K60 (CXC chemokine) AF277660 [29]K203 (CC chemokine) Y18692 [29]
Stem cell factor D13516 [30]
cMGF X14477 [31]
Meleagris gallopavo(Turkey) IL-2 AJ007463 [32]
Type I interferon U28140 [33]IFN-� AJ000725 [34]
Coturnix japonica(Japanese Quail) IFN-� AJ001678 [34]
Phasianus colchicus(Ring-Necked Pheasant) IFN-� AJ001289 [34]
AmphibiansXenopus laevis(African Clawed Frog) Embryonic FGF X62593 [35]
FGF-2 M18067 [36]FGF-3 Z25539 [37]FGF-8 Y10312 [38]FGF-9 U47622 [39]FGF-10 AB073747 [40]
TGF-�2 X51817 [41]TGF-�5 J05180 [42]
Bony fishOncorhynchus mykiss(Rainbow Trout) IL-8 AJ279069
TNF-� AJ277604 [43]
TGF-�1 X99303 [44]TGF-�2 AJ318935TGF-�3 AJ318928 [45]
FGF6 Y16850 [46]
CK-1 (CXC chemokine) AF093802 [47]Putative chemokine (CXC chemokine) AJ300835Putative chemokine (CC chemokine) AJ315149
Cyprinus carpio(Common Carp) Allograft inflammatory factor 1 AB012309 [48]
Chemokine-1 (CC chemokine) AB010469 [48]

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 485
Table 1 (Continued)
Organism Gene GenBank accession number Reference
NK cell enhancing factor AB010959 [48]
Danio rerio (Zebrafish) FGF8 AF034264 [49]
Scyba (CXC chemokine) AF279919 [50]CCL1 (CC chemokine) AF201450
Paralichthys olivaceus(Bastard Halibut) IL-8 AF216646 [51]
TNF-� AB040448 [52]
Acipenser bari(Siberian Sturgeon) TGF-�3 AJ318926 [45]
Anguilla anguilla (European Eel) TGF-�1 AJ318934TGF-�3 AJ318927 [45]
Pleuronectes platessa(Plaice) TGF-�1 AJ318929 [53]TGF-�2 AJ318930 [53]TGF-�3 AJ318931 [53]
Cartilaginous fishTriakis scyllium(Banded Dogfish) IL-8 AB063299
Jawless fishLampetra fluviatilis(Lamprey) LFCA-1 (CXC chemokine) AJ231072 [54]
Publications cited where available.
with the sequencing of various immune genes, that all jawedvertebrates (Gnathostomes) possess a complex immune sys-tem, which is capable of anticipatory and innate immuneresponses[1,2]. Jawless vertebrates posses an equally com-plex immune system but no evidence has been presentedto prove the presence of an anticipatory immune response.However, innate immunity is found and is shown to havea more ancient origin. From these studies it can be specu-lated that the initiation and regulation of the immune systemwithin vertebrates will be equally complex.
The evolution of the immune system has awarded cy-tokines a key role as co-ordinators of the immune system.The important role of cytokines in fine-tuning and control-ling immune responses[3,4] raises the question of whetheror not these molecules have been highly conserved throughevolution. Very little has been done to look at the evolution ofcytokine genes within vertebrates, despite well-known bio-logical activities throughout the vertebrates and invertebrates[5]. Which cytokines will be found within primitive verte-brates and how far back in the vertebrate phylogeny thesemolecules go is only beginning to be addressed. It may bethat cytokines important in anticipatory responses will proveto be unique to jawed vertebrates whereas cytokines impor-tant in non-specific defences may be more universal. Re-cently, cytokines which have been well characterised withinmammals[6] have begun to be cloned and sequenced withinnon-mammalian vertebrates. Cytokines have now been se-quenced within amphibians, birds, bony fish, cartilaginousfish and jawless fish (Tables 1 and 2), with the numberof cytokine sequences available from primitive vertebratesgrowing rapidly. Molecules related to vertebrate cytokinereceptors are also being cloned in many vertebrate groups[5,7–9].
2. Interleukin-1 family
Interleukin-1� was initially discovered within mice andhumans and over the last 10 years has been characterisedwithin a wide variety of mammalian species (Table 2). Theinterleukin-1 (IL-1) family of cytokines has four main mem-bers: IL-1�, IL-1�, IL-1 receptor antagonist (IL-1ra) andIL-18, with a number of newly discovered molecules show-ing clear homology to this group[81–87]. IL-1 is a majormediator of inflammation and in general initiates and/or in-creases a wide variety of non-structural, function associatedgenes that are characteristically expressed during inflamma-tion, particularly other cytokines. It is one of the key media-tors of the body’s response to microbial invasion, inflamma-tion, immunological reactions and tissue injury. Both in vivoand in vitro experiments have shown that IL-1� and IL-1�have similar if not identical multiple biological effects whichhave been well catalogued[81,88–91]and both forms affectnearly every cell type and share a common receptor on tar-get cells. Recent work however has shown that the endoge-nous roles of IL-1� and IL-1� are different where IL-1� butnot IL-1� is a potent activator of the humoral immune re-sponse[92]. IL-1� and IL-1� are produced by many differ-ent cell types[88] including neutrophils, natural killer cells,B-lymphocytes, T-lymphocytes and cells of the central ner-vous system, but are made mainly by blood monocytes andtissue macrophages[93–96]. The monocyte/macrophage isan important source because of its strategic locations, itsability to synthesise large amounts of IL-1 and to processthe IL-1 precursor more effectively than other cells.
Members of the IL-1 family belong to what is nowknown as the�-trefoil superfamily due to the presence of12 �-sheets in their mature protein structure which fold

486 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
Table 2Table of known IL-1� sequences
Organism Gene GenBank accession number Reference
Homo sapien(Human) IL-1� mRNA M15330 [55]IL-1� gene M15840 [56]
Cercocebus torquatus(Red-Crowned Mangabey) IL-1� mRNA U19837 [57]Macaca fascicularis(Crab-Eating Macaque) IL-1� mRNA D63353 [58]Macaca mulatta(Rhesus Monkey) IL-1� mRNA U19845 [57]Macaca nemestrina(Pig-Tailed Macaque) IL-1� mRNA U19853 [57]
Mus musculus(Mouse) IL-1� mRNA M15131 [59]IL-1� gene X04964 [60]
Rattus norvegicus(Norway Rat) IL-1� mRNA M98820Sigmodon hispidus(Hispid Cotton Rat) IL-1� mRNA AF421387Equus callus(Horse) IL-1� mRNA D42147 [61]Bos taurus(Cow) IL-1� mRNA M35589 [62]Cervus elaphus(Red Deer) IL-1� mRNA U20500Ovis aries(Sheep) IL-1� mRNA X54796 [63]
Sus scrofa(Pig) IL-1� mRNA M86725 [64]IL-1� gene X74568 [65]
Capra hircus(Goat) IL-1� mRNA D63351Felis domesticus(Cat) IL-1� mRNA M92060Oryctolagus cuniculus(Rabbit) IL-1� mRNA D21835 [66]Tursiops truncatus(Bottle-Nosed Dolphin) IL-1� mRNA AB028216 [67]Trichosurus vulpecula(Brush-Tailed Possum) IL-1� mRNA AF071539 [68]Ornithorhynchus anatinus(Duck-Billed Platypus) IL-1� gene AJ239048 [69]Mesocricetus auratus(Golden Hamster) IL-1� mRNA, partial AB028497Canis familiaris(Dog) IL-1� mRNA, partial Z70047
Gallus gallus(Chicken) IL-1� mRNA Y15006 [70]IL-1� gene AJ245728
Xenopus laevis(African Clawed Frog) IL-1� mRNA AJ010497 [71]IL-1� gene AJ314758 [69]
Dicentrarchus labrax(European Seabass) IL-1� mRNA AJ269472 [72]IL-1� gene AJ311925 [73]
Oncorhynchus mykiss(Rainbow Trout) IL-1� 1 mRNA AJ223954 [74]IL-1� 1 gene AJ004821 [75]IL-1� 2 mRNA AJ245925 [76]IL-1� 2 gene AJ245925 [76]
Sparus aurata(Gilthead Seabream) IL-1� mRNA AJ277166 [77]IL-1� gene AJ419178 [77]
Carrassius auratus(Goldfish) IL-1� 1 mRNA AJ249136 [69]IL-1� 1 gene AJ419848 [69]IL-1� 2 mRNA AJ249137 [69]IL-1� 2 gene AJ419849 [69]
Cyprinus carpio(Common Carp) IL-1� 1 mRNA AB010701 [78]IL-1� 1 gene AJ245635 [79]IL-1� 2-1 mRNA AJ401030IL-1� 2-2 mRNA AJ401031
Scophthalmus maximus(Turbot) IL-1� mRNA, partial AJ295836Pleuronectes platessa(Plaice) IL-1� mRNA, partial AJ010640 [69]Danio rerio (Zebrafish) IL-1� mRNA, partial AI330756Scyliorhinus canicula(Small Spotted Catshark) IL-1� mRNA AJ251201 [80]
IL-1� gene AJ295947 [80]
Publications cited where available.

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 487
to form a trefoil-like structure[97,98]. Other members ofthis superfamily include the fibroblast growth factors[82].IL-1�, IL-1� and IL-1ra contain the IL-1 family signaturepattern or motif, taken from a selected conserved region inthe C-terminal section with the following consensus pattern;[FC]-x-S-[ASLV]-x(2)-P-x(2)-[FYLIV]-[LI]-[SCA]-T-x(7)-[LIVM] as found in the PROSITE database[99]. IL-1�and IL-1� differ substantially in relation to localisation,maturation and secretion. Both forms are translated fromthe messenger RNA (mRNA) into the immature forms(31 kDa), pro-IL-1� and pro-IL-1�, which due to the lackof a hydrophobic leader peptide sequence remain in thecystol and do not accumulate in any organelles[100]. Bothforms can undergo cleavage by enzymes, to produce ma-ture forms, which augments the activity of these peptides.Pro-IL-1� but not pro-IL-1� can be cleaved by calciumdependent membrane proteases called calpains[101,102]to give a 17 kDa mature peptide. IL-1� is rarely found inthe circulation or extracellular biological fluids and only incases of serious illness when it is thought to have origi-nated from lysed cells[103] and cleavage by extracellularproteases[104]. Pro-IL-1� is just as active as the matureform, and appears to remain intracellular and act at thislevel [105,106]. Pro-IL-1� can also be found on the surfaceof several cells, especially monocytes and B-cells, where itremains biologically active.
Pro-IL-1� remains primarily cytosolic and is not fullyactive until it is proteolytically cleaved and transported outof the cell. A cysteine protease, IL-1� converting enzyme(ICE), also known as caspase-1, has been identified as thespecific enzyme responsible for this cleavage of pro-IL-1�[107–109]. Caspases are primarily involved in apopto-sis [110,111], and over expression of ICE itself can leadto increased apoptosis in cells transfected with this gene[112,113]. The ability of both ICE and pro-IL-1� to coexistwithout mature IL-1� being formed suggests ICE activitymust be regulated[114–116]. Also, it has been shown thatICE has five isoforms generated by alternative splicing ofthe mRNA[117], each with varying degrees of activity. Togenerate mature IL-1�, ICE specifically cleaves the aspar-tate (amino acid position 116)–alanine (amino acid position117) bond[118]. It is generally thought that cleavage ofpro-IL-1� is an obligatory step for IL-1� release, sincetreatment with most ICE inhibitors fails to cause detectablelevels of extracellular pro-IL-1� [118]. This is not absoluteand some ICE inhibitors can cause pro-IL-1� to be se-creted instead of the mature form[118,119]. The processesinvolved in secretion of the mature form of IL-1� are illdefined. It has been suggested that pro-ICE present in thecytoplasm is activated, assembled and inserted into the cellmembrane where it would cleave pro-IL-1� and release themature form into the extracellular environment[120]. It isthought that mature IL-1� is released by a novel secretorypathway as it has been shown that IL-1� is contained, in part,within intracellular vesicles that protect it from proteasedigestion and active secretion of IL-1� requires its translo-
cation across the membrane[121]. More recently it hasbeen shown that release of IL-1� is mediated by adenosine5′-triphosphate (ATP) and requires a priming step by LPSwhich is independent of caspase-1 cleavage[122]. In addi-tion, alternative mechanisms are present for the processingof the pro-IL-1� [123]. There are several sites in pro-IL-1�which have been shown to be vulnerable to cleavage byenzymes in the vicinity of alanine 117. Proteolytic enzymesincluding chymase, elastase, cathepsin G, collagenase andmatrix metalloproteinases have all been shown to cleavepro-IL-1� into a form similar in size and specific activityas the ICE processed IL-1� [123,124]. These proteases arereleased at inflammatory and tissue damage sites by a va-riety of immune cells[125,126]. Crystal structure analysishas confirmed site directed mutagenesis studies, showingthat both mature IL-1� and IL-1� bind to the IL-1 receptor(IL-1RI) at two sites, contacting the first two domains ofthe receptor (site A) or the third domain (site B)[127].
The IL-1ra is a specific antagonist of IL-1, in that it bindsthe IL-1 receptor without transmitting any signal into thecell [128]. Directed mutagenesis results have shown that,unlike IL-1�/�, IL-1ra has only one binding site for theIL-1 receptor, explaining the absence of signal transmission[129]. The IL-1ra cDNA encodes a 177 amino acid proteinwhich contained a 25 amino acid signal peptide and allowedthe protein to be secreted into the extracellular environment,named secretory IL-1ra (sIL-1ra). Upon stimulation withLPS, human blood monocytes initially express this gene[130], but after 24 h the primary transcript in these cells is anintracellular form of IL-1ra (icIL-1ra) which lacks a leaderpeptide[100]. This form arises due to alternative splicingof the sIL-1ra mRNA which modifies the exon encoding forthe signal sequence[131,132]. A second form of icIL-1rais also generated by alternative splicing of the mRNA andhas been termed icIL-1ra type II[133–135]. It is thoughtthat these intracellular isoforms may represent a reservoir ofIL-1ra, released upon cell death, whose function is to limitthe pro-inflammatory action of cell debris.
3. IL-1 in vertebrates
Due to the importance of IL-1 in the immune systemof mammals and evidence to suggest its biological activ-ity in the immune system of non-mammalian vertebrates,work has focussed on isolating this gene in selected species.Other than mammalian IL-1� sequences there are now fullcDNA sequences available from chicken,Xenopus, trout,carp, goldfish, seabass and catshark (Table 2). Many of thesegenes have been obtained using an homology cloning ap-proach. The genomic organisation for these IL-1� geneshave also been obtained within selected species. In the caseof Ornithorhynchus anatinus(Duck-Billed Platypus), onlythe genomic sequence is available, due to this organism be-ing a protected species, with the predicted platypus IL-1�transcript being obtained with the aid of software, which

488 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
searches for intron–exon boundaries[69]. To date, no se-quence data is available for IL-1� or IL-1ra, other than inmammals whilst IL-18 is known in birds as well as mam-mals (see later).
4. Known vertebrate IL-1� sequences
IL-1� has been characterised within a large number ofmammals, including a representative from the marsupials[68] and monotremes (GenBank accession no. AJ239048,[69]), which are known to be the most primitive mammalgroups. More recently IL-1� has been cloned from selectednon-mammalian vertebrates, which include chicken[70],Xenopus[71], trout[74], carp[78], seabass[72] and catshark[80]. When these genes are aligned amino acid homologyis found to be highest in regions containing the secondarystructure of the IL-1� molecule. This suggests the presenceof 12 �-sheets in each of these genes giving the proteinssimilar folding patterns and making each a member of the�-trefoil family [98]. The secondary structure dictates howthe molecule will fold, which gives the molecule its tertiarystructure. The correct folding of the IL-1� molecule is im-portant if it is to interact with the receptor and induce a signalinto the target cell. An early study showed that a histidineresidue, (position 146 in the human precursor protein) wascritical for this molecule binding to its receptor[136]. Thisamino acid residue appears to be conserved in the platypusand chicken molecules. The use of site specific mutagene-sis allowed the identification of another seven amino acids(Arg4, Leu6, Phe46, Ile56, Lys93 Lys103 and Glu105) in hu-man mature IL-1� that are essential for binding to the type IIL-1 receptor[137]. On the three-dimensional structure, theyform a cluster localised in one region of the IL-1� molecule.These amino acids are strongly conserved in other placen-tal mammal IL-1� proteins, partially conserved in platypusand possum IL-1� [68] and not conserved in the knownnon-mammalian sequences. In both the possum and platy-pus IL-1� five of the seven positions contain identical or re-lated amino acids. These amino acid differences may explainwhy placental mammal IL-1� recombinant protein does notcross-react well in possum and vice versa[68,138]. A morerecent comparison has identified two specific receptor bind-ing sites (sites A and B) which have been determined in hu-man IL-1� using the crystal structure of IL-1� complexedwith its receptor[139]. These regions in the non-mammaliansequences show little conservation in the alignment.
The IL-1 family signature, which spans�-sheets 9–11of IL-1�, is reasonably well conserved within each of thenon-mammalian genes (Fig. 1). All mammalian sequences,including the possum and platypus, contain this consensussequence. Both the chicken and theXenopussequences alsohave this signature pattern, but whether this will be true forall bird and amphibian species remains to be seen. Differ-ences begin to be found in the motif within the bony and car-tilaginous fish and slight changes are required to the motif
to allow the inclusion of these sequences to the IL-1 family(Fig. 1). In common, as well, is the presence of numerouscopies of an mRNA instability motif (attta) in the 3′ UTRof each IL-1� gene, which is typical of genes coding forinflammatory mediators[140].
Although the IL-1� sequences share the IL-1 family sig-nature and show the presence of possible�-sheets the over-all nucleotide and amino acid identities of these moleculeswithin vertebrates is quite low.Fig. 2shows that some of thehighest homologies are found between the mammals (hu-man, possum and platypus) whereas the lowest values arefound between the catshark and other vertebrates. This is notunexpected as this sequence represents the earliest knownIL-1� sequence isolated to date. All known IL-1� aminoacid and nucleotide sequences have also been comparedusing different methods of phylogenetic comparison to con-struct trees to look at the relationship of this molecule withinvertebrates. Methods used included neighbour joining[141],maximum parsimony[142] and maximum likelihood[143],although only the tree from neighbour joining is shown as thetrees constructed were fairly congruent (Fig. 3, [69]). In allcases the mammalian and bony fish form two separate cladeswith the relationship between them unresolved. Wheremethods attempted to resolve the relationship between thevertebrates, the data was not supported (for example themaximum likelihood tree obtained from amino acid datahas a bootstrap value of 55%). In each tree, the relation-ships of the bony fish are well supported, with the carp andgoldfish genes forming a separate group to the trout andseabass. The placement of the chicken and theXenopuschanges within the different trees, but in the majority ofcases they branch with the mammals. The bootstrap val-ues in the maximum parsimony tree and neighbour-joiningtrees obtained from amino acid data did not support thisrelationship. Increasing the representatives from each ofthese groups would probably help in resolving the group-ing of these organisms. Including all the IL-1� genesindicated that the platypus and possum appear to be an-cestral to all the mammals. The genetic distance betweenthe bony fish sequences from the catshark was less thanthe mammals, indicating a closer relationship with thissequence.
Within the trout a second IL-1� gene (IL-1�2) has beenfound [76]. It has a similar intron–exon organisation to thefirst trout IL-1� gene sequenced (IL-1�1) [75], except thatthere are small differences in the sizes of the introns and ex-ons. Multiple copies of IL-1� genes have also been foundin the carp (GenBank accession nos. AJ401030, AJ401031)and in the goldfish[69]. The presence of multiple copiesof IL-1 family genes is not just restricted to fish. Multiplegene have been found within various mammals. In humans,a number of novel homologue members of the IL-1 fam-ily have been found[83,144]. The roles of these at presentare speculative, with three being similar to IL-1ra and onebeing more related to either IL-1� or IL-18. The receptors,signalling pathways, biological activities and tissue expres-

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 489
Fig. 1. Alignment of the IL-1 family signatures found to date in mammals, birds, amphibians, bony fish and cartilaginous fish IL-1� sequences. Abovethe alignment is the signature, which is found within all mammalian sequences. Below are the changes (shown in red) required to incorporate all thenon-mammalian sequences.
sion of these homologues have yet to be investigated. It isthought that these homologues may bind to some IL-1R-likereceptors that have yet to be paired with ligands. In pig twogenes for IL-1� have also been identified which share 86%amino acid identity[145]. The findings of two IL-1� genesin trout, carp and goldfish are not totally unexpected as thesefish have a tetraploid ancestry and are quite likely to havemultiples of certain genes. What will be of interest is to de-termine how these genes are regulated and what role, if any,these genes play in the fishes immune system.
What remains to be determined is whether polymorphismexists for IL-1� within non-mammalian vertebrates. In mam-
Fig. 2. Amino acid (yellow) and nucleotide (white) identities of IL-1� sequences from selected mammals and non-mammals.
mals, a certain amount of polymorphism exists for IL-1members since two allelic forms have been described forIL-1� [146], IL-1ra [147] and several for IL-1� [148]. Thesepolymorphisms may be of clinical importance since a spe-cific IL-1� allele is associated with juvenile polyarthritis[149], a rare IL-1� allele is related to the severity of rheuma-toid arthritis erosions[150] or certain types of diabetes[151]and a IL-1ra allele may contribute to susceptibility to sepsis[152]. A recent study has also shown that the IL-1 genotypeis significantly associated with the severity of adult peri-odonitis[153]. These finding could prove particularly usefulfor commercially important species, such as trout or seabass,

490 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
Fig. 3. Rooted phylogram showing the phylogenetic relationships between all known IL-1� genes that have been sequenced within the vertebrates.Phylogeny was estimated using amino acid sequences with the neighbour-joining method and the resulting tree was bootstrapped 1000 times. Bootstrapvalues are shown as a percentage. Scale bar represents an estimate of the number of amino acid substitutions per site.
where a certain genotype could prove advantageous againsta particular disease or infection.
5. Processing of IL-1� in vertebrates
All vertebrate IL-1� genes found contain no apparentsignal peptide, as deduced by plotting hydrophobicity ofthe amino acids and using signal P v1.1[154], indicatingthat these molecules are secreted through a non-classicalpathway not involving the golgi/endoplasmic reticulumroute. Differences are found in how the mammalian andnon-mammalian pro-IL-1� molecules are processed intotheir mature form. The ICE cut site is present within allmammalian IL-1� sequences found to date, including theplatypus, although it remains to be tested experimentallythat this is the actual cut site within this primitive mammal.Throughout all other vertebrate groups in which IL-1� has
been cloned and sequenced, no ICE cut site is present.Fig. 4depicts the possible cut site within these peptides based onan alignment of all known non-mammalian vertebrates withselected mammalian sequences. The cut site for the maturepeptide has been predicted within the chicken[70], carp[155], and trout[156,157]and active recombinant peptidesproduced (Fig. 5). The existence in non-mammalian verte-brates of an ICE homologue and whether IL-1� requiresenzymatic cleavage to become biologically is under inves-tigation. ICE enzymes are likely to be present in lower ver-tebrates, since apoptosis is a well characterised event in fish[158] and related genes have been found in invertebrate’s[159]. Recent isolation of a caspase-1 enzyme sequencein chicken[160], numerous caspase homologs in zebrafish[161] and caspase-6 in trout[162] indicate that a homologfor caspase-1 could be present within all vertebrates. Studieshave also shown that the mammalian precursor IL-1� canbe cleaved at different amino acid residues and by different

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 491
Fig. 4. Alignment of the cut site region of IL-1� sequences. The ICE cut site (aspartic acid) seen in all the mammalian sequences is in red. The potentialcut site regions in non-mammalian sequences are highlighted white. Amino acids in green relate to�-sheets.
cleavage enzymes to obtain a biologically active molecule.Granzyme A but not granzyme B, which are released fromcytotoxic T-lymphocytes, cleaves at Arg120 [163]. Elastase,trypsin and chymotrypsin, with chymotrypsin producing themost biologically active IL-1�, can cleave precursor IL-1�[164]. A mast cell chymase[126] and various extracellu-lar matrix metalloproteinases (MMP)[124] have also beenshown to produce biologically active IL-1�. These proteasesare found in inflammatory fluids, at sites of neutrophil,lymphocyte, or macrophage infiltration, and can therefore
Fig. 5. Northern blot showing the effect of LPS and rainbow trout recombinant IL-1� on IL-1� gene expression in rainbow trout macrophages.
participate in the generation of active IL-1�. As pro-IL-1�can be released from live cells or passively from deadcells [165], it is possible that extracellular processing ofpro-IL-1� by inflammatory proteases takes place. Evidencehas also been found in human plantar stratum and psoriaticscales where biologically active IL-1� has been activatedin a process not involving ICE[166]. Thus non-mammalianvertebrate IL-1� may be activated by cleavage by an enzymedistinct from an ICE homologue. The exact cut site, how thepro-peptides are processed and what protease is involved in

492 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
non-mammalian vertebrates remains to be determined. Thequestion also remains as to the origin of the ICE cut site inIL-1� and due to the unavailability of reptilian sequencesfor IL-1� it is unknown if the ICE cut site is present withinthis group of vertebrates. If not then the use of ICE forthis purpose would appear to be an exclusively mammalianevent.
6. Vertebrate IL-1� gene organisation
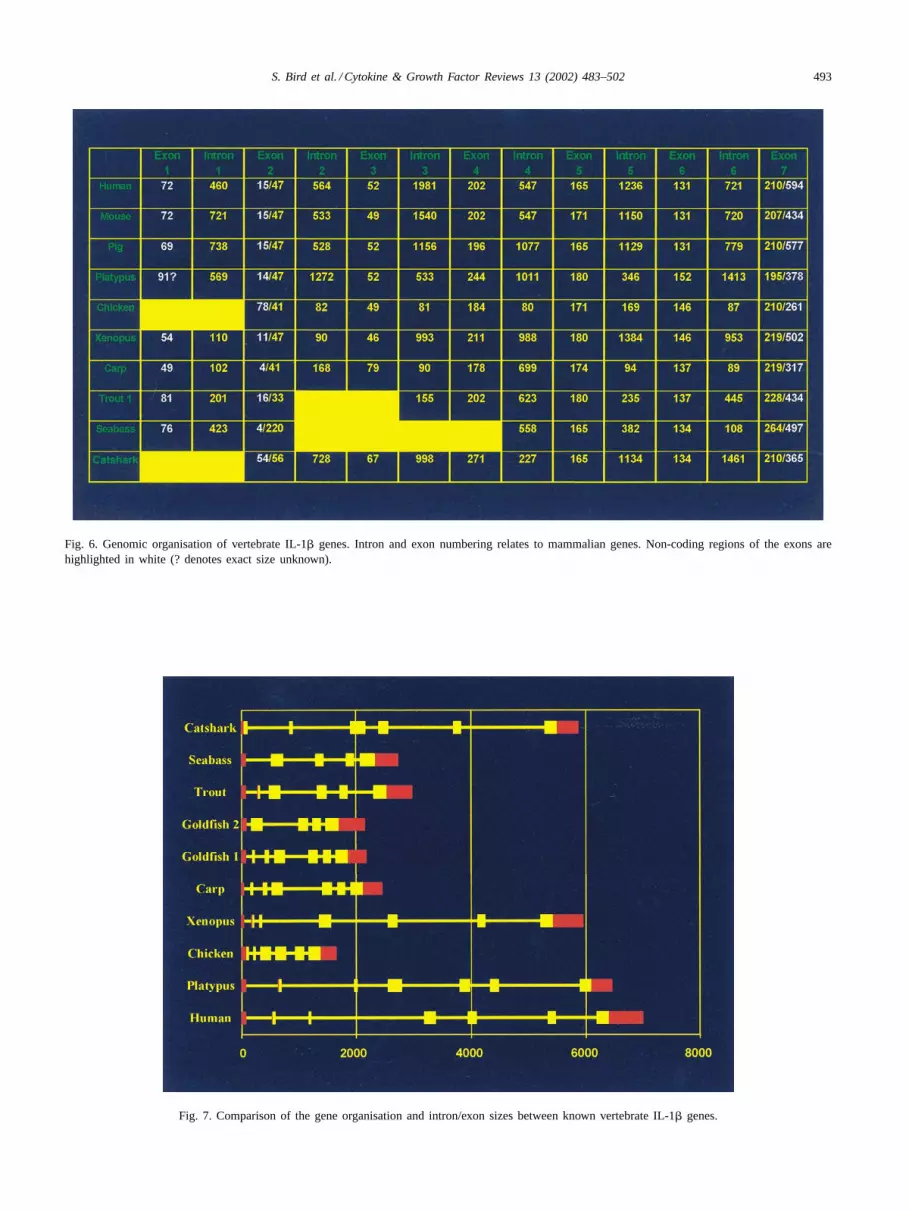
The intron and exon arrangement of IL-1� genes havebeen determined within selected vertebrate species andcompared (Figs. 6 and 7). The precursor sequence of hu-man, mouse and pig IL-1� were the first to be sequencedand are found to span seven exons separated by six introns[56,60,65]. The first non-mammalian vertebrates to havetheir IL-1� genomic sequence discovered were trout, carpand chicken ([75,79]; GenBank accession no. AJ245728),followed by seabass[73] and Xenopus(GenBank acces-sion no. AJ314758,[69]). A monotreme mammal, theDuck-Billed Platypus (GenBank accession no. AJ239048,[69]), is also known.
Similar to the known mammalian IL-1� genes, carp con-sists of six introns and seven exons, with exon sizes similarto those seen in the human IL-1� gene. The chicken, troutand catshark IL-1� genes possess a different organisationcontaining only five introns, although trout appears to havelost a coding exon where chicken and catshark have lost a5′ UTR exon. The introns for the carp, trout and chickengenes are considerably smaller than the mammalian genes.In contrast, the catshark introns are much larger. The platy-pus andXenopussequence has the same intron/exon organi-sation to mammals, although the sizes of the introns do vary.The recently sequenced seabass gene organisation has a dif-ferent organisation again, containing four introns and fiveexons, and like trout appears to be missing an exon at the 5′end. Exons 4–7 appear to be relatively conserved betweenspecies whereas exons 1–3 appear in some organisations tobe spliced together or completely missing. On discovery ofthe trout IL-1� gene it was hypothesised tentatively that themammalian IL-1� had evolved by the introduction of extrasequence at the N-terminus[75]. With the introduction ofother non-mammalian sequences this still may hold true, al-though in view of the larger size of the coding region of thecatshark gene, the ancestral IL-1� gene may have lost cod-ing sequence as it evolved within some vertebrate groups.
There seems to be a lot of variation in the number ofintrons and where they are placed in the genomic organi-sation of IL-1�, which is especially seen in the bony fishwhere each species studied to date shows a different or-ganisation. However, this is not an unusual event as intronshave been shown to frequently occur in different positionsand/or different numbers in homologous genes of differ-ent species[167]. This is thought to be due to either gains(“introns-late”) or losses (“introns-early”) of introns, or per-
haps both. Evidence exists to support both the introns-earlyand introns-late hypotheses[168–170]. Furthermore, it hasbeen proposed that introns may speed up the evolution ofgenes by increasing recombination events between exons[168]. The sequence and size of introns may be evolutionaryimportant in directing the molecular organisation of genes.
7. Vertebrate IL-1� expression and regulation
The involvement of IL-1� in the non-mammalian verte-brate immune response is indicated by expression studies.In mammals, IL-1� is produced in response to many stimuliwhich include bacterial LPS, numerous microbial products,cytokines (TNF, IFN-�, GM-CSF and IL-2), T-cell/antigenpresenting cell interactions and immune complexes[171].Similar patterns of expression have also been seen in tissuesfound to contain lymphomyeloid tissue in chicken,Xeno-pus, bony fish and cartilaginous fish. In chicken, IL-1� isquickly induced in blood monocyte derived macrophagesreaching optimal levels within 1 h after LPS treatment[70].The XenopusIL-1� transcript was inducible in vivo fol-lowing injection with LPS[71]. Expression was clearlydetectable by RT-PCR in the kidney, liver and spleen, withthe brain showing the lowest expression levels. Northernblot analysis confirmed that the transcript was not constitu-tively expressed in the spleen and that there was a three-foldincrease post injection. Stimulation of splenocytes in vitrowith LPS also produced a significant increase in transcriptexpression. In carp, administration of PMA and LPS tophagocytes isolated from the head kidney induced expres-sion of IL-1� in these cells after 2 h stimulation[79]. Also,fish injected with immunopotentiators, sodium alginate andscleroglucan 48 h earlier showed increased IL-1� mRNAlevels in peritoneal and head kidney cells when comparedto saline injected fish[78]. In trout, head kidney leucocytesand isolated macrophages express IL-1� after stimulationwith LPS for 4 h ([74,172], Fig. 5). A total of 5�g/ml LPSis shown to produce maximal expression and was first de-tectable 1–2 h post stimulation with maximal induction at4 h. Culture temperature has been shown to have a markedeffect on trout IL-1� expression[172]. Using LPS a cleardose dependent induction of the trout IL-1�2 is shown incultured trout leucocytes[76]. Also, a trout mononuclearphagocyte cell line, RTS-11 cells express IL-1�. Stimu-lating these cells with LPS led to increased expression ofIL-1� [173]. More recently, LPS stimulation of seabassgill and head kidney cells in vitro for 4 h has also re-sulted in an increase in IL-1� expression, as seen usingRT-PCR and Northern blotting[72]. Expression studies inS. caniculusindicate that the IL-1� molecule is biologi-cally relevant to cartilaginous fish immune responses toGram-negative bacteria[80]. IL-1� was shown to be in-ducible in vivo following injection with LPS, 24 h earlier.Expression was clearly detectable by RT-PCR in the spleenand testes, tissues known to contain a variety of leucocytes
S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 493
Fig. 6. Genomic organisation of vertebrate IL-1� genes. Intron and exon numbering relates to mammalian genes. Non-coding regions of the exons arehighlighted in white (? denotes exact size unknown).
Fig. 7. Comparison of the gene organisation and intron/exon sizes between known vertebrate IL-1� genes.

494 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
Fig. 8. Comparison of mammalian and non-mammalian vertebrate IL-1� promoters (? denotes exact size unknown).
[174]. Stimulation of splenocytes in vitro with LPS from0 to 8 h also resulted in a significant increase in transcriptexpression with maximal expression seen after 5 h. North-ern blot analysis confirmed that there was a seven-foldincrease in the transcript after 5 h incubation with LPS[69,80].
Within the seabass[72], trout [172] and carp[79] the ex-pression of incompletely spliced variants of the IL-1� genehas been shown. They appear to be expressed at lower levelsin these fish relative to the mature transcript and are not uni-versally present in all tissues that express IL-1�. It is thoughtthat the incompletely spliced forms of these cytokines maybe involved in the regulation of the mature transcript, as hasbeen seen in mammals in certain situations[175].
The regulation of the non-mammalian IL-1� genes hasalso been investigated with the sequencing of promotersfrom selected species, which include trout[176] and cat-shark (GenBank accession no. AJ295947). The IL-1� pro-moter has been examined extensively in humans and mice[177–185]and homologous positive and negative regulatoryelements for transcription factors have been found in thenon-mammalian IL-1� promoters. A TATA box is present24–27 bp upstream of the transcription start site in all speciesexamined. An NF-�B element, which is known to be cru-cial for activity within the immediate upstream promoter ofmammalian IL-1� [186] and the more upstream enhancer[184], has also been identified (Fig. 8). In trout an NF-�Bsite is located 19–21 bp (trout IL-1�1 and IL-1�2) upstreamof the TATA box whereas in the catshark it is 91 bp up-stream. It is known that IL-1� is a strong inducer of NF-�Band is thought to positively autoregulate its own synthesis,which appears to hold true in fish as the addition of troutrecombinant IL-1� to trout macrophages induces IL-1� ex-pression (Fig. 5). Preliminary studies using the trout IL-1�1promoter confirm that the NF-�B transcription factor site
is required for expression of the trout IL-1�1 gene[176].Other potential elements for transcription activators in thetrout and catshark promoters include sites for AP1, SP1 andCEBP.
8. Vertebrate IL-1 receptors
IL-1 receptors have been isolated within a few selectnon-mammalian species. Within trout a homologue of themammalian IL-1 type II receptor has been isolated[187],which is known as a ‘decoy’ receptor and thought to rep-resent a way of negatively regulating IL-1 activity. This re-ceptor gene is induced in trout head kidney leukocytes afterstimulation by LPS and TNF-�, consists of a signal peptide,transmembrane region, extracellular region with three Ig-likedomains and a short cytoplasmic tail which is characteris-tic of the mammalian IL-1 type II receptor. Homologues ofthe mammalian IL-1 type I receptor gene have been clonedin chicken[188] and salmon[189]. In the chicken the im-portance of this receptor has been shown as IL-1 activityis blocked when this receptor is produced in soluble form[190]. This receptor has also been shown to transduce a sig-nal into target cells as IL-1 activity is decreased when thisreceptor is blocked using an antibody.
9. Interleukin-18
IL-18 or IFN-� inducing factor (IGIF) was cloned in micefrom partial amino acid sequences deduced from the purifiedproteins[191] and later in humans[192]. Analysis of the pri-mary amino acid sequence of both human and mouse IL-18indicated the presence of an IL-1 signature-like sequence andit was proposed that IL-18 contained 12 strands of�-sheets

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 495
Fig. 9. Amino acid alignment of the IL-18 cut site region from mammalian and chicken sequences with the aspartic acid coloured red. Identical (∗) andsimilar (: or .) residues are indicated.
forming the�-trefoil fold, typical of the IL-1 family[193].IL-18 exists as an inactive pro-IL-18 molecule until it iscleaved by ICE to generate the mature active form of IL-18[194,195]. It appears that the correct folding of IL-18 oc-curs prior to processing with ICE, at the level of pro-IL-18[196]. The structure and processing of this molecule suggestthat IL-18 and IL-1 originate from a common ancestor, al-though the target specificity’s and the receptor systems arecompletely different. It has also been reported that the IL-1receptor related protein is the functional component of theIL-18 receptor[197].
In contrast to what is observed with IL-1�, constitutivegene expression of IL-18 has been seen within various celltypes including unstimulated, freshly isolated human PBMCand murine splenocytes[198], and haemopoietic cell lines ofmyelomonocytic origin[199]. It has been shown that the pro-moter for IL-18 does not contain a TATA box and that pro-moter activity upstream of exon 2 acts constitutively[200].It is also found that the 3′ UTR of human IL-18 lacks theAUUUA destabilisation sequence, allowing for more sus-tained levels of the polyadenylated species and translationinto protein. The presence of preformed pro-IL-18 in thecell would allow for a very rapid production of the mature,active molecule once the cell is activated.
Recently, IL-18 was cloned within chicken[11], the onlynon-mammalian vertebrate to date known to have this cy-tokine. Sequence comparison reveals a putative ICE cleav-age site at aspartic acid 29 (Fig. 9). Synthesis of IFN-� incultured primary chicken spleen cells was detected when in-cubated with the chicken mature recombinant IL-18 usingthe predicted cut site. Thus the presence of ICE in chickenswould appear crucial for IL-18 processing in this species.
10. Cytokine and IL-1 evolution
The evolution of the genes that encode the current spec-trum of known mammalian cytokines and receptor com-plexes involved multiple duplications from a smaller set ofgenes, followed by the divergence of sequence and product
function [201]. To date, very little in fact is known aboutthe evolution of cytokine genes and much is inferred frommammalian sequences due to the lack of data from othervertebrate groups. A number of studies have attempted topredict the possible evolutionary history of several cytokinesusing mammalian nucleotide data.
In general cytokines have been found to be some of themost rapidly evolving genes known. A survey of 1196 or-thologous human and mouse genes found that 7 out of the 25most divergent genes were cytokines and cytokine receptors(Fas, IFN-�, IFN-�R, IL-4, IL-4R, IL-6, IL-6R), sharing aslittle as 51% nucleotide sequence identity and 41% aminoacid identity[202]. Using knockout mice a number of geneshave been termed essential and non-essential genes depend-ing on the survivability of these mice. It was found that alarge proportion of the non-essential genes are immune sys-tem genes, including cytokines, which appear to be evolvingfaster than the essential ones[203]. This rapid evolution ofcytokine genes is reflected in the finding that cross-reactivityof IL-1� between marsupial and placental mammals is low.IL-1 in marsupials was first demonstrated when a moleculewith molecular weight of 15–17 kDa was obtained fromopposum LPS stimulated macrophages and keratinocytes[138]. This IL-1 seemed to be non-stimulatory to mouse thy-mocytes and mouse and human IL-1 did not stimulate oppo-sum thymocytes. Recent work has isolated the IL-1� genefrom possum LPS stimulated lung alveolar macrophages[68]. The deduced amino acid sequence has 43–46% iden-tity to placental mammals and contained an ICE cut site. Thepossum recombinant IL-1� when given intravenously hada profound effect on the haematology of possums. A largetransient decrease in circulating leucocytes and primary neu-trophils followed by a large increase in numbers of both neu-trophils and lymphocytes was found. Also when used as anadjuvant with KLH, serum antibody levels were increasedsix-fold at 6 weeks after immunisation relative to unadju-vanted, immunised animals[204]. In agreement with earlierwork, there was little cross-reactivity between possum IL-1�and placental mammal leucocytes. These cytokine genesmay be under directional selection because of host-parasite

496 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
co-evolution. One reason put forward for this high rate ofevolution for antimicrobial host defence proteins, such ascytokines is said to be due to pathogenic microorganisms us-ing molecular mimicry[205–208]. For example, numerousherpes viruses and pox viruses encode chemokine mimicsable to block chemokine action. Mutations in these immuneregulatory genes become favoured because they result in atemporary advantage until it is counteracted by mutations inthe pathogen genome.
Knowledge on the structure and function of cytokinesfrom different species is beginning to accumulate withthe use of such recombinant cytokines. Preliminary resultsfrom cross-reacting cytokines from different mammalianspecies has shown that there is an apparent threshold ofaround 60% amino acid identity[209]. Below this iden-tity, it is unlikely that cross-reactivity between cytokinesfrom different species will be found. Cross-reactivity ofdifferent cytokines can also be correlated to the foldingfamilies that the cytokines belong to. Cytokines belongingto the short 4-�-helical bundle cytokine family are the leastcross-reactive whereas cytokines belonging to the�-sheetbased folds are the most cross-reactive and also the mostconserved amongst the species studied. This may show thatcertain families are able to accumulate more differences intheir amino acid composition and still produce a proteinable to fold in the desired way.
A number of ideas have been formulated on the evolutionof IL-1 with the accumulation of sequence data and theircomparisons with closely related genes which share the same�-sheet structure. The IL-1 family belongs to the�-trefoilfamily, which also includes molecules, such as IL-18 and theFGF family of proteins. Although the FGF and IL-1 familieswere considered homologous, there was limited evidencebeyond similarity in three-dimensional structures to suggesta common ancestor for any pair of�-trefoil families [210].Each family possesses distinct functions, occur in extra- andintracellular contexts and lack similarities in either the typeof molecules with which they interact or in the location oftheir binding sites on the tertiary structure[211]. It was un-known if the fold arose independently several times withindifferent molecules during evolution, meaning that�-trefoilproteins could be examples of analogous or homologousgenes[212]. More recent work has analysed the significanceof sequence identities observed after structure alignment,which has provided evidence that�-trefoil proteins are ho-mologous, having arisen from a common ancestor[210]. Itis now thought that the�-trefoil fold arose from the dupli-cation of an ancestral gene encoding a homotrimeric singlerepeat protein.
Until recently, only mammalian sequences were availableto infer the possible evolution of the interleukin-1 familymembers. It was initially found that genes for human, mouseand rat IL-1ra, IL-1� and IL-1� have similar intron–exon or-ganisation, indicating that gene duplication events may havebeen important in the creation of this gene family[60,213]. Ithas been suggested that the IL-1ra duplicated from an ances-
tral IL-1 gene first, corresponding to the time of the appear-ance of amphibians (320–400 million years ago). IL-1� andIL-1� separation is thought to have occurred more recently(270–300 million years ago). An alternative hypothesis isthat IL-1� duplicated first from the ancestral IL-1, favouredby the fact that IL-1� has diverged more from IL-1ra andIL-1� than these two proteins have from each other. Whichever duplication took place first, the dates where duplicationsare thought to have taken place may suggest that bony andcartilaginous fish contain only one IL-1 gene. It has been fur-ther proposed that the common ancestor of IL-1� and IL-1rawas an alternatively spliced gene[214]. One transcript couldhave encoded the ancestral IL-1� while the other transcriptencoded the IL-1ra. After gene duplication, one copy of thegene would be freed to encode only the receptor antagonist.With the discovery of IL-1� sequences within vertebrates asancient as cartilaginous fish it is certainly clear that IL-1�had an early origin. However, it remains to be discoveredat what stage other IL-1 family members evolved.
11. Conclusion
The discovery of IL-1� genes in mammals, amphibians,bony fish and cartilaginous fish has major implications forinvestigation into the immune systems of vertebrates. Theidentification of the coding sequences for IL-1� within thesevertebrate groups is only the beginning and studies into thefunctional characteristics of the proteins encoded by thesegenes must now be investigated. Ultimately the productionof recombinant peptides for all non-mammalian genes willallow the bioactivity to be determined and compared be-tween vertebrate groups. The effect of molecules, such asIL-1 on the immune system is in its early stages and requiresintense investigation. Once a better idea of IL-1 mediatedimmune cell activity is established, the use of IL-1 or IL-1inhibitors may be realistically considered as a way to modu-late the immune response possibly at the time of vaccinationin commercially important vertebrate species. Whatever theoutcome, it is likely that IL-1 will be a major regulator ofthe immune responses in these other vertebrates as it is inmammals. In addition to analysing IL-1 ligands and theiractivities in vertebrates it will also be of interest to inves-tigate the signalling mechanisms in some species. Furtherwork needs to be done to detect or isolate IL-1 receptorgenes from tissues within non-mammalian vertebrates anddetermine expression patterns/regulations for each of thesereceptors, in addition to assessing ligand-binding kinetics.
Homology cloning has proved to be a useful tool withininvestigations for isolating and cloning cytokine sequencesfrom a wide variety of vertebrates[43,45,71,74,80,204]. Thecontinuing discovery of more cytokines has been improvedmore recently, using methods such as subtractive hybridi-sation [48,78]. Techniques, like this, are especially usefulwhen looking for molecules that may have diverged dur-ing evolution, which is not uncommon for cytokines. Also,

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 497
with major advances within technology, the sequencing ofcomplete genomes is now becoming a possibility within arelatively short time[215,216]. The identification of allgenes within selected vertebrates other than mammals willimpact research into the evolution of cytokines dramati-cally. It will allow the identification of all cytokines, whichare mammalian homologues, may reveal molecules, whichare unique to certain organisms and will also give a betterinsight into where immune system communicators arose.
With the discovery of IL-1� within cartilaginous fish andthe presence in these vertebrates of components required fora functional innate and adaptive immune system, it givesgood reason to suspect that other cytokines are present at thislevel in vertebrate phylogeny. What is unknown is at whatlevel within evolution did these immune regulators arise,since it is now generally considered unlikely that cytokinesfrom vertebrates will have a homologue within invertebrates[5,217]. Whether IL-1 will be found within an organismmore primitive than a cartilaginous fish is under study withinjawless vertebrates. To date, little success has been had inidentifying cytokines within hagfish and lamprey, althougha homologue of IL-8 has been sequenced within the lam-prey,Lampetra fluviatilis[54]. It may be that cytokines suchas IL-1 arose within the jawed vertebrate lineage. It also re-mains to be determined where IL-1 family members such asIL-1� and IL-1ra arose during vertebrate evolution. It hasbeen speculated that these family members arose during theemergence of the amphibians[213]. There is still much thatremains to be discovered about the evolution of cytokineswithin vertebrates and it will be the focus of many studiesfor years to come.
Acknowledgements
Most of the non-mammalian sequences and the Duck-Billed Platypus sequence presented in this review wereobtained during a BBSRC programme (1/S09641) and aBBSRC studentship to S.B. In the case of the platypus se-quence, many thanks go to Niall Stewart in the collectingand curating of the monotreme tissues. The platypus mate-rial was collected under a permit (to B.M.) provided by theTasmanian Parks and Wildlife Service.
References
[1] Klein J. Homology between immune responses in vertebrates andinvertebrates: does it exist? Scand J Immunol 1997;46:558–64.
[2] Du Pasquier L. The immune system of invertebrates and vertebrates.Comp Biochem Phys 2001;129B:1–15.
[3] Balkwill F, Burke F. The cytokine network. Immunol Today1989;10:299–304.
[4] Vilcek J, The cytokines: an overview. In: Thomson A, editor. Thecytokine handbook. London: Academic Press, 1998. p. 1–20.
[5] Scapigliati G, Bird S, Secombes C. Invertebrate and fish cytokines.Eur Cytokine Netw 2000;11:354–61.
[6] Thomson A. The cytokine handbook. London: Academic Press,1998.
[7] Secombes C. The phylogeny of cytokines. In: Thomson A, editor.The cytokine handbook. London: Academic Press, 1998.
[8] Secombes C, Zou J, Daniels G, Cunningham C, Koussounadis A,Kemp G. Rainbow trout cytokine and cytokine receptor genes.Immunol Rev 1998;166:333–40.
[9] Secombes C, Bird S, Hong S, Laing K, Zou J. Phylogeny ofvertebrate cytokines. Adv Exp Med Biol 2001;484:89–94.
[10] Sundick R, Gill-Dixon C. A cloned chicken lymphokinehomologous to both mammalian IL-2 and IL-15. J Immunol1997;159:720–5.
[11] Schneider K, Klaas R, Kaspers B, Staeheli P. Chicken interleukin-6—cDNA structure and biological properties. Eur J Biochem2001;268:4200–6.
[12] Kaiser P, Hughes S, Bumstead N. The chicken 9E3/CEF4 CXCchemokine is the avian orthologue of IL8 and maps to chickenchromosome 4 syntenic with genes flanking the mammalian chemo-kine cluster. Immunogenetics 1999;49:673–84.
[13] Lillehoj H, Burnside J, Choi K. Molecular characterization andtissue distribution of chicken cytokines homologous to mammalianIL-15 and IL-2. FASEB J 2000;14(Suppl):A1125.
[14] Schneider K, Puehler F, Baeuerle D, Elvers S, Staeheli P, KaspersB, et al. cDNA cloning of biologically active chicken interleukin-18.J Interferon Cytokine Res 2000;20:879–83.
[15] Sick C, Schultz U, Staeheli P. A family of genes coding for twoserologically distinct chicken interferons. J Biol Chem 1996;271:7635–9.
[16] Weining K, Schultz U, Munster U, Kaspers B, Staeheli P.Biological properties of recombinant chicken interferon-gamma. EurJ Immunol 1996;26:2440–7.
[17] Zuniga A, Borja M, Meijers C, Zeller R. Expression of alternativelyspliced BFGF 1st coding exons and antisense messenger RNAsduring chicken embryogenesis. Dev Biol 1993;157:110–8.
[18] Mahmood R, Kiefer P, Guthrie S, Dickson C, Mason I. Multipleroles for FGF-3 during cranial neural development in the chicken.Development 1995;121:1399–410.
[19] Niswander L, Jeffrey S, Martin G, Tickle C. A positive feedbackloop coordinates growth and patterning in the vertebrate limb.Nature 1994;371:609–12.
[20] Vogel A, Rodriguez C, Izpisua-Belmonte J. Involvement of FGF-8in initiation, outgrowth and patterning of the vertebrate limb.Development 1996;122:1737–50.
[21] Ohuchi H, Nakagawa T, Yamamoto A, Araga A, ] T, Ishimaru Y,et al. The mesenchymal factor, FGF-10, initiates and maintains theoutgrowth of the chick limb bud through interaction with FGF-8,an apical ectodermal factor. Development 1997;124:2235–44.
[22] Munoz-Sanjuan I, Simandl B, Fallon J, Nathans J. Expression ofchicken fibroblast growth factor homologous factor (FHF)-1 and ofdifferentially spliced isoforms of FHF-2 during development andinvolvement of FHF-2 in chicken limb development. Development1999;126:409–21.
[23] Ohuchi H, Kimura S, Watamoto M, Itoh N. Involvement of fibroblastgrowth factor (FGF)-18- FGF8 signaling in specification of left-rightasymmetry and brain and limb development of the chick embryo.Mech Dev 2000;95:55–66.
[24] Ladher R, Anakwe K, Gurney A, Schoenwolf G, Francis-West P.Identification of synergistic signals initiating inner ear development.Science 2000;290:1965–8.
[25] Burt D, Paton I. Molecular cloning and primary structure of thechicken transforming growth factor beta 2 gene. DNA Cell Biol1991;10:723–34.
[26] Jakowlew S, Dillard P, Kondaiah P, Sporn M, Roberts A. Comple-mentary deoxyribonucleic acid cloning of a novel transforminggrowth factor-beta messenger ribonucleic acid from chick embryochondrocytes. Mol Endocrinol 1988;2:747–55.

498 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
[27] Burt D, Jakowlew S. Correction: a new interpretation of a chickentransforming growth factor beta 4 complementary DNA. MolEndocrinol 1992;6:989–92.
[28] Wistow G, Shaughnessy M, Lee D, Hodin J, Zelenka P. A macro-phage migration inhibitory factor is expressed in the differentiatingcells of the eye lens. Proc Natl Acad Sci USA 1993;90:1272–5.
[29] Sick C, Schneider K, Staeheli P, Weining K. Novel chicken CXCand CC chemokines. Cytokine 2000;12:181–6.
[30] Zhou J, Ohtaki M, Sakurai M. Sequence of a cDNA encodingchicken stem cell factor. Gene 1993;127:269–70.
[31] Leutz A, Damm K, Sterneck E, Kowenz E, Ness S, Frank R, et al.Molecular cloning of the chicken myelomonocytic growth factor(cMGF) reveals relationship to interleukin-6 and granulocyte colonystimulating factor. EMBO J 1989;8:175–81.
[32] Lawson S, Rothwell L, Kaiser P. Turkey and chicken interleukin-2cross react in in vitro proliferation assays despite limited aminoacid sequence identity. J Interferon Cytokine Res 2000;20:161–70.
[33] Suresh M, Karaca K, Foster D, Sharma J. Molecular and functionalcharacterization of turkey interferon. J Virol 1995;69:8159–63.
[34] Kaiser P, Sonnemans D, Smith L. Avian interferon-� genes:sequence analysis suggests probable cross-species reactivity among-st galliforms. J Interferon Cytokine Res 1998;18:711–9.
[35] Isaacs H, Tannahill D, Slack J. Expression of a novel FGFin the Xenopus embryo. A new candidate inducing factor formesoderm formation and anteroposterior specification. Development1992;114:711–20.
[36] Kimelman D, Kirschner M. Synergistic induction of mesoderm byFGF and TGF-beta and the identification of an messenger RNAcoding for FGF in the early Xenopus embryo. Cell 1987;51:869–77.
[37] Kiefer P, Mathieu M, Close J, Peters G, Dickson C. FGF3 fromXenopus laevis. EMBO J 1993;12:4159–68.
[38] Christen B, Slack J. FGF-8 is Associated with anteroposteriorpatterning and limb regeneration in Xenopus. Dev Biol 1997;192:455–66.
[39] Song J, Slack J. XFGF-9: a new fibroblast growth factor fromXenopus embryos. Dev Dyn 1996;206:427–36.
[40] Yokoyama H, Yonei-Tamura S, Endo T, Izpisua Belmonte J,Tamura K, Ide H. Mesenchyme with FGF-10 expression isresponsible for regenerative capacity in Xenopus limb buds. DevBiol 2000;219:18–29.
[41] Rebbet M, Bhatia-Dey N, Dawid I. The sequence of TGF-�2 fromXenopus laevis. Nucleic Acids Res 1990;18:2185.
[42] Kondaiah P, Sands M, Smith J, Fields A, Roberts A, Sporn M, et al.Identification of a novel transforming growth factor beta (TGF-beta5) mRNA in Xenopus laevis. J Biol Chem 1990;265:1089–93.
[43] Laing K, Wang T, Zou J, Holland J, Hong S, Bols N, et al. Cloningand expression analysis of rainbow troutOncorhynchus mykisstumour necrosis factor alpha. Eur J Biochem 2001;268:1315–22.
[44] Hardie L, Laing K, Daniels G, Grabowski P, Cunningham C,Secombes C. Isolation of the first piscine transforming growthfactor beta gene: analysis reveals tissue specific expression anda potential regulatory sequence in rainbow trout (Oncorhynchusmykiss). Cytokine 1998;10:555–63.
[45] Laing K, Pilstrom L, Cunningham C, Secombes C. TGF-beta3exists in bony fish. Vet Immunol Immunopathol 1999;72:45–53.
[46] Rescan P. Identification of a fibroblast growth factor 6 (FGF6)in a non-mammalian vertebrate: continuous expression of FGF6accompanies muscle fiber hyperplasia. Biochem Biophys Acta1998;1443:305–14.
[47] Dixon B, Shum B, Adams E, Magor K, Hedrick R, Muir D, etal. CK-1, a putative chemokine of rainbow trout (Oncorhynchusmykiss). Immunol Rev 1998;166:341–8.
[48] Fujiki K, Shin D, Nakao M, Yano T. Molecular cloning of carp(Cyprinus carpio) CC chemokine, CXC chemokine receptors,allograft inflammatory factor-1, and natural killer cell enhancingfactor by use of suppression subtractive hybridization. Immuno-genetics 1999;49:909–14.
[49] Fuerthauer M, Thisse C, Thisse B. A role for FGF-8 in thedorsoventral patterning of the zebrafish gastrula. Development1997;124:4253–64.
[50] Long Q, Quint E, Lin S, Ekker M. The zebrafish scyba geneencodes a novel CXC-type chemokine with distinctive expressionpatterns in the vestibulo-acoustic system during embryogenesis.Mech Dev 2000;97:183–6.
[51] Lee E, Park H, Kim Y, Chung J, Choi T. Cloning and sequenceanalysis of the interleukin-8 gene from flounder (Paralichthysolivaceous). Gene 2001;274:237–43.
[52] Hirono I, Nam B, Kurobe T, Aoki T. Molecular cloning,characterization, and expression of TNF cDNA and gene fromJapanese flounderParalychthys olivaceus. J Immunol 2000;165:4423–7.
[53] Laing K, Cunningham C, Secombes C. Genes for three differentisoforms of transforming growth factor beta are present inplaice (Pleuronectes platessa) DNA. Fish Shellfish Immunol2000;10:261–71.
[54] Najakshin A, Mechetina L, Alabyev B, Taranin A. Identificationof an IL-8 homolog in lamprey (Lampetra fluviatilis): earlyevolutionary divergence of chemokines. Eur J Immunol 1999;29:375–82.
[55] Nishida T, Nishino N, Takano M, Kawai K, Bando K, Masui Y,et al. cDNA cloning of IL-1 alpha and IL-1 beta from mRNA ofU937 cell line. Biochem Biophys Res Commun 1987;143:345–52.
[56] Bensi G, Raugei G, Palla E, Carinci V, Buonamassa D, Melli M.Human interleukin-1 beta gene. Gene 1987;52:95–101.
[57] Villinger F, Brar S, Mayne A, Chikkala N, Ansari A. Comparativesequence analysis of cytokine genes from human and nonhumanprimates. J Immunol 1995;155:3946–54.
[58] Totsuka K, Takakura H, Hashimoto O, Tatsumi M. Molecularcloning and expression of cynomolgus monkey interleukin-1 betaCdna. Int Arch Allergy Immunol 1998;116:40–8.
[59] Gray P, Glaister D, Chen E, Goeddel D, Pennica D. Twointerleukin-1 genes in the mouse—cloning and expression of thecDNA for murine interleukin-1 beta. J Immunol 1986;137:3644–8.
[60] Telford J, Macchia G, Massone A, Carinci V, Palla E, MelliM. The murine interleukin-1 beta gene—structure and evolution.Nucleic Acids Res 1986;14:9955–63.
[61] Kato H, Ohashi T, Nakamura N, Nishimura Y, Watari T, GoitsukaR, et al. Molecular cloning of equine interleukin-1 alpha andinterleukin-1 beta cDNAs. Vet Immunol Immunopathol 1995;48:221–31.
[62] Leong S, Flaggs G, Lawman M, Gray P. The nucleotide sequencefor the cDNA of bovine interleukin-1 beta. Nucleic Acids Res1988;16:9054.
[63] Fiskerstrand C, Sargan D. Nucleotide sequence of ovine inter-leukin-1 beta. Nucleic Acids Res 1990;18:7165.
[64] Huether M, Lin G, Smith D, Murtaugh M, Molitor T. Cloning,sequencing and regulation of an messenger RNA encoding porcineinterleukin-1 beta. Gene 1993;129:285–9.
[65] Vandenbroeck K, Fiten P, Beuken E, Martens E, Janssen A, VanDamme J, et al. Gene sequence, cDNA construction, expression inEscherichia coliand genetically approached purification of porcineinterleukin-l beta. Eur J Biochem 1993;217:45–52.
[66] Mori S, Goto F, Goto K, Ohkawara S, Maeda S, Shimada K,et al. Cloning and sequence analysis of a cDNA for lymphocyteproliferation potentiating factor of rabbit polymorphonuclear leuko-cytes—identification as rabbit interleukin-1 beta. Biochem BiophysRes Commun 1988;150:1237–43.
[67] Inoue Y, Itou T, Ueda K, Oike T, Sakai T. Cloning and sequencingof a bottle-nosed dolphin (Tursiops truncatus) interleukin-1 alphaand -1 beta complementary DNAs. J Vet Med Sci 1999;61:1317–21.
[68] Wedlock D, Goh L, Parlane N, Buddle B. Molecular cloningand physiological effects of brushtail possum interleukin-1�. VetImmunol Immunopathol 1999;67:359–72.
[69] Bird S. Molecular evolution of IL-1� within vertebrates. Ph.D.Thesis, University of Aberdeen, Scotland, UK, 2002.

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 499
[70] Weining K, Sick C, Kaspers B, Staeheli P. A chicken homolog ofmammalian interleukin-1 beta: cDNA cloning and purification ofactive recombinant protein. Eur J Biochem 1998;258:994–1000.
[71] Zou J, Bird S, Minter R, Horton J, Cunningham C, Secombes C.Molecular cloning of the gene for interleukin-1� from Xenopuslaevis and analysis of expression in vivo and in vitro. Immuno-genetics 2000;49:332–8.
[72] Scapigliati G, Buonocore F, Bird S, Zou J, Pelegrin P, Falasca C,et al. Phylogeny of cytokines: molecular cloning and expressionanalysis of sea bassDicentrarchus labraxinterleukin-1 beta. FishShellfish Immunol 2001;11:711–26.
[73] Buonocore F, Prugnoli D, Falasca C, Secombes C, Constantini S,Facchiano A, et al. Phylogeny of cytokines II: gene organisationand protein structure of IL-1� in seabass,Dicentrarchus labraxL,in press, 2002.
[74] Zou J, Grabowski P, Cunningham C, Secombes C. Molecularcloning of interleukin 1 beta from rainbow trout,Oncorhynchusmykiss, reveals no evidence of an ice cut site. Cytokine 1999;11:552–60.
[75] Zou J, Cunningham C, Secombes C. The rainbow troutOncorhyn-chus mykissinterleukin-1� gene has a different organisation tomammals and undergoes incomplete splicing. Eur J Biochem1999;259:901–8.
[76] Pleguezuelos O, Zou J, Cunningham C, Secombes C. Cloning,sequencing and analysis of expression of a second IL-1� gene inrainbow trout (Oncorhynchus mykiss). Immunogenetics 2000;51:1002–11.
[77] Pelegrin P, Garcia-Castillo J, Mulero V, Meseguer J. Interleukin-1�
isolated from a marine fish reveals up regulated expression inmacrophages following activation with lipopolysaccharide andlymphokines. Cytokine 2001;16:67–72.
[78] Fujiki K, Shin D, Nakao M, Yano T. Molecular cloning andexpression analysis of carp (Cyprinus carpio) interleukin-1�,high affinity immunoglobulin E Fc receptor� subunit and serumamyloid A. Fish Shellfish Immunol 2000;10:229–42.
[79] Engelsma M, Stet R, Schipper H, Verburg van Kemenade B.Regulation of interleukin-1 beta RNA expression in the commoncarp,Cyprinus carpioL. Dev Comp Immunol 2001;25:195–203.
[80] Bird S, Wang T, Zou J, Cunningham C, Secombes C. The firstcytokine sequence within cartilaginous fish: interleukin-1� inthe small spotted catshark (Scyliorhinus canicula). J Immunol2000;168(7):3329–40.
[81] Dinarello C. Interleukin-1. Cytokine Growth Factor Rev1997;8:253–65.
[82] Dinarello C. Interleukin-18. Methods 1999;19:121–32.[83] Mulero J, Pace A, Nelken S, Loeb D, Correa T, Drmanac R, et al.
IL1HY1: a novel interleukin-1 receptor antagonist gene. BiochemBiophys Res Commun 1999;263:702–6.
[84] Busfield S, Comrack C, Yu G, Chickering T, Smutko J, Zhou H,et al. Identification and gene organization of three novel membersof the IL-1 family on human chromosome 2. Genomics 2000;66:213–6.
[85] Smith D, Renshaw B, Ketchem R, Kubin M, Garka K, Sims J.Four new members expand the interleukin-1 superfamily. J BiolChem 2000;275:1169–75.
[86] Lin H, Ho A, Haley-Vicente D, Zhang J, Bernal-Fussell J, PaceA, et al. Cloning and characterization of IL-1HY2, a novelinterleukin-1 family member. J Biol Chem 2001;276:20597–602.
[87] Pan G, Risser P, Mao W, Baldwin D, Zhong A, Filvaroff E,et al. IL-1H, an interleukin 1-related protein that binds IL-18receptor/IL-1Rrp. Cytokine 2001;13:1–7.
[88] Oppenheim J, Kovacs E, Matsushima K, Durum S. There is morethan one interleukin-1. Immunol Today 1986;7:45–56.
[89] Dinarello C. Interleukin-1 and interleukin-1 antagonism. Blood1991;77:1627–52.
[90] Dinarello C. The biological properties of interleukin-1. EurCytokine Netw 1994;5:517–31.
[91] Dinarello C. Biological basis for interleukin-1 in disease. Blood1996;87:2095–147.
[92] Nakae S, Asano M, Horai R, Iwakura Y. Interleukin-1 beta, butnot interleukin-1 alpha, is required for T-cell dependent antibodyproduction. Immunology 2001;104:402–9.
[93] Lepe-Zuniga J, Gery I. Production of intracellular and extracellularinterleukin-1 (IL-1) by human monocytes. Clin Immunol Immuno-pathol 1984;31:222–30.
[94] Dinarello C, Cannon J, Wolff S, Bernheim H, Beutler B, CeramiA, et al. Tumour necrosis factor (cachectin) is an endogenouspyrogen and induces production of interleukin-1A. J Exp Med1986;163:1433–50.
[95] Dinarello C. Biology of interleukin-1. FASEB J 1988;2:108–15.[96] Arend W, Gordon D, Wood W, Janson R, Joslin F, Jameel S. IL-1�
production in cultured human monocytes is regulated at multiplelevels. J Immunol 1989;143:118–26.
[97] Murzin A, Lesk A, Chothia C. Beta-trefoil fold. Patterns of structureand sequence in the Kunitz inhibitors interleukins-1 beta and -1alpha and fibroblast growth factors. J Mol Biol 1992;223:531–43.
[98] Nicola N, Guidebook to cytokines and their receptors. Oxford:Sambrook and Tooze, 1994.
[99] Hofmann K, Bucher P, Falquet L, Bairoch A. The PROSITEdatabase, its status in 1999. Nucleic Acids Res 1999;27:215–9.
[100] Andersson J, Bjork L, Dinarello C, Towbin H, Andersson U.Lipopolysaccharide induces human interleukin-1 receptor antagonistand interleukin-1 production in the same cell. Eur J Immunol1992;22:2617–23.
[101] Kobayashi Y, Yamamoto K, Saido T, Kawasaki H, Oppenheim J,Matsushima K. Identification of calcium-activated neutral proteaseas a processing enzyme of human interleukin-1 alpha. Proc NatlAcad Sci USA 1990;87:5548–52.
[102] Kavita U, Mizel S. Differential sensitivity of interleukin-1 alphaand interleukin-1 beta precursor proteins to cleavage by calpain, acalcium dependent protease. J Biol Chem 1995;270:27758–65.
[103] Wakabayashi G, Gelfand J, Jung W, Connolly R, Burke J, DinarelloC. Staphylococcus epidermidisinduces complement activation,tumor necrosis factor and interleukin-1, a shock-like state and tissueinjury in rabbits without endotoxemia—comparison toEscherichiacoli. J Clin Invest 1991;87:1925–35.
[104] Kobayashi Y, Appella E, Yamada M, Copeland T, OppenheimJ, Matsushima K. Phosphorylation of intracellular precursors ofhuman IL-1. J Immunol 1988;140:2279–87.
[105] Maier J, Voulalas P, Roeder D, Maciag T. Extension of the lifespan of human endothelial cells by an interleukin-1 alpha antisenseoligomer. Science 1990;249:1570–4.
[106] Weitzmann M, Savage N. Nuclear internalization and DNAbinding activities of interleukin-1, interleukin-1 receptor and inter-leukin-1 receptor complexes. Biochem Biophys Res Commun1992;187:1166–71.
[107] Cerretti D, Kozlosky C, Mosley B, Nelson N, Vanness K,Greenstreet T, et al. Molecular cloning of the interleukin-1 betaconverting enzyme. Science 1992;256:97–100.
[108] Thornberry N, Bull H, Calaycay J, Chapman K, Howard A,Kostura M, et al. A novel heterodimeric cysteine protease isrequired for interleukin-1 beta processing in monocytes. Nature1992;356:768–74.
[109] Tocci M. Structure and function of interleukin-1� convertingenzyme. Vitamin Horm 1997;53:27–63.
[110] Nicholson D, Thornberry N. Caspases: killer proteases. TrendsBiochem Sci 1997;22:299–306.
[111] Takahashi A. Caspase: executioner and undertaker of apoptosis.Int J Hematol 1999;70:226–32.
[112] Faucheu C, Diu A, Chan A, Blanchet A, Miossec C, Herve F,et al. A novel human protease similar to the interleukin-1 betaconverting enzyme induces apoptosis in transfected cells. EMBOJ 1995;14:1914–22.

500 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
[113] Miossec C, Dutilleul V, Durand L, Faucheu C, Pilatre L, GolecJ, et al. Evidence for an interleukin-1 beta converting enzyme(ICE)-like activity distinct from ICE and responsible for ICE andCPP32 cleavage during apoptosis. Apoptosis 1997;2:125–35.
[114] Laliberte R, Perregaux D, Svensson L, Pazoles C, Gabel C. Tenidapmodulates cytoplasmic pH and inhibits anion transport in vitro.2. Inhibition of IL-1 beta production from ATP-treated monocytesand macrophages. J Immunol 1994;153:2168–79.
[115] Schonbeck U, Herzberg M, Petersen A, Wohlenberg C, GerdesJ, Flad H, et al. Human vascular smooth muscle cells expressinterleukin-1 beta converting enzyme (ICE), but inhibit processingof the interleukin-1 beta precursor by ICE. J Exp Med 1997;185:1287–94.
[116] Young J, Sukhova G, Foster D, Kisiel W, Libby P, Schonbeck U.The serpin proteinase inhibitor 9 is an endogenous inhibitor ofinterleukin 1 beta converting enzyme (caspase-1) activity in humanvascular smooth muscle cells. J Exp Med 2000;191:1535–44.
[117] Alnemri E, Fernandesalnemri T, Litwack G. Cloning and expressionof 4 novel isoforms of human interleukin-1 beta converting enzymewith different apoptotic activities. J Biol Chem 1995;270:4312–7.
[118] Miller D, Calaycay J, Chapman K, Howard A, Kostura M,Molineaux S, et al. The IL-1 beta converting enzyme as atherapeutic target. Ann N Y Acad Sci 1993;696:133–48.
[119] Chin J, Kostura M. Dissociation of IL-1 beta synthesis andsecretion in human blood monocytes stimulated with bacterial cellwall products. J Immunol 1993;151:5574–85.
[120] Singer I, Scott S, Chin J, Bayne E, Limjuco G, Weidner J, et al.The interleukin-1 beta converting enzyme (ICE) is localized onthe external cell surface membranes and in the cytoplasmic groundsubstance of human monocytes by immuno-electron microscopy. JExp Med 1995;182:1447–59.
[121] Rubartelli A, Cozzolino F, Talio M, Sitia R. A novel secretorypathway for interleukin-1 beta, a protein lacking a signal sequence.EMBO J 1990;9:1503–10.
[122] Mehta V, Hart J, Wewers M. ATP-stimulated release of interleukin(IL)-1 beta and IL-18 requires priming by lipopolysaccharide and isindependent of caspase-1 cleavage. J Biol Chem 2001;276:3820–6.
[123] Fantuzzi G, Ku G, Harding M, Livingston D, Sipe J, Kuida K, etal. Response to local inflammation of IL-1 beta converting enzymedeficient mice. J Immunol 1997;158:1818–24.
[124] Schonbeck U, Mach F, Libby P. Generation of biologically activeIL-1� by matrix metalloproteinases: a novel caspase-1 independentpathway of IL-1� processing. J Immunol 1998;161:3340–6.
[125] Hazuda D, Strickler J, Kueppers F, Simon P, Young P. Processingof precursor interleukin-1 beta and inflammatory disease. J BiolChem 1990;265:6318–22.
[126] Mizutani H, Schechter N, Lazarus G, Black R, Kupper T. Rapidand specific conversion of precursor interleukin-1 beta (IL-1 beta)to an active IL-1 species by human mast cell chymase. J Exp Med1991;174:821–5.
[127] Vigers G, Caffes P, Evans R, Thompson R, Eisenberg S,Brandhuber B. X-ray structure of interleukin-1 receptor antagonistat 2.0-angstrom resolution. J Biol Chem 1994;269:12874–9.
[128] Hannum C, Wilcox C, Arend W, Joslin F, Dripps D, HeimdalP, et al. Interleukin-1 receptor antagonist activity of a humaninterleukin-1 inhibitor. Nature 1990;343:336–40.
[129] Evans R, Bray J, Childs J, Vigers G, Brandhuber B, SkalickyJ, et al. Mapping receptor-binding sites in interleukin (IL)-1receptor antagonist and IL-1 beta by site directed mutagenesis—identification of a single site in IL-1ra and 2 sites in IL-1-beta. JBiol Chem 1995;270:11477–83.
[130] Arend W. Interleukin-1 receptor antagonist. Adv Immunol 1993;54:167–227.
[131] Haskill S, Martin G, Vanle L, Morris J, Peace A, Bigler C, et al.cDNA cloning of an intracellular form of the human interleukin-1receptor antagonist associated with epithelium. Proc Natl Acad SciUSA 1991;88:3681–5.
[132] Butcher C, Steinkasserer A, Tejura S, Lennard A. Comparison of2 promoters controlling expression of secreted or intracellular IL-1receptor antagonist. J Immunol 1994;153:701–11.
[133] Muzio M, Polentarutti N, Sironi M, Poli G, Degioia L, IntronaM, et al. Cloning and characterization of a new isoform of theinterleukin-1 receptor antagonist. J Exp Med 1995;182:623–8.
[134] Muzio M, Polentarutti N, Facchetti F, Peri G, Doni A, Sironi M, etal. Characterization of type II intracellular IL-1 receptor antagonist(IL-1ra3): a depot IL-1ra. Eur J Immunol 1999;29:781–8.
[135] Malyak M, Smith M, Abel A, Hance K, Arend W. The differentialproduction of three forms of IL-1 receptor antagonist by humanneutrophils and monocytes. J Immunol 1998;161:2004–10.
[136] MacDonald H, Wingfield P, Schmeissner U, Shaw A, Clore G,Gronenborn A. Point mutations of human interleukin-1 withdecreased receptor binding affinity. FEBS Lett 1986;209:295–8.
[137] Labriola-Tompkins E, Chandran C, Kaffka K, Biondi D, GravesB, Hatada M, et al. Identification of the discontinuous binding sitein human interleukin-1� for the type I interleukin-1 receptor. ProcNatl Acad Sci USA 1991;88:11182–6.
[138] Brozek C, Ley R. Production of Interleukin-1 in a southamerican opposum (Monodelphus domestica). Dev Comp Immunol1991;15:401–12.
[139] Vigers G, Anderson L, Caffes P, Brandhuber B. Crystal structureof the type-1 interleukin-1 receptor complexed with interleukin-1�.Nature 1997;386:190–4.
[140] Caput D, Beutler B, Hartog K, Thayer R, Brown-Shimer S,Cermi A. Identification of a common nucleotide sequence in the3′-untranslated region of mRNA molecules specifying inflammatorymediators. Proc Natl Acad Sci USA 1986;83:1670–4.
[141] Saitou N, Nei M. The neighbor-joining method—a new method forreconstructing phylogenetic trees. Mol Biol Evol 1987;4:406–25.
[142] Swofford D. PAUP—a computer-program for phylogeneticinference using maximum parsimony. J Gen Physiol 1993;102:A9.
[143] Strimmer K, Von Haeseler A. Quartet puzzling: a quartet maximumlikelihood method for reconstructing tree topologies. Mol BiolEvol 1996;13:964–9.
[144] Kumar S, McDonnell P, Lehr R, Tierney L, Tzimas M, GriswoldD, et al. Identification and initial characterisation of four novelmembers of the interleukin-1 family. J Biol Chem 2000;275:10308–14.
[145] Vandenbroeck K, Fiten P, Opdenakker G, Billiau A. Two distinctinterleukin-1� genes in the pig genome. Immunogenetics 1994;40:82.
[146] Di Giovine F, Takhsh E, Blakemore A, Duff G. Single basepolymorphism at -511 in the human interleukin-1 beta gene (IL-1beta). Hum Mol Genet 1992;1:450.
[147] Clay F, Tarlow J, Cork M, Cox A, Nicklin M, Duff G. Novelinterleukin-1 receptor antagonist exon polymorphisms and their usein allele specific mRNA assessment. Hum Genet 1996;97:723–6.
[148] Bailley S, Di Giovine F, Blakemore A, Duff G. Geneticpolymorphism of human interleukin-1 alpha. Eur J Immunol1993;23:1240–5.
[149] McDowell T, Symons J, Ploski R, Forre O, Duff G. A geneticassociation between juvenile rheumatoid arthritis and a novelinterleukin-1 alpha polymorphism. Arthritis Rheum 1995;38:221–8.
[150] Di Giovine F, Crane A, Gough A, De Vries N, Van de PutteL, Emery P, et al. A genetic polymorphism in the IL-1� geneis associated with join erosions in rheumatoid arthritis. Cytokine1994;6:545.
[151] Pociot F, Molvig J, Wogensen L, Worsaae H, Nerup J. A taqIpolymorphism in the human interleukin-1 beta (IL-1 beta) genecorrelates with IL-1 beta secretion in vitro. Eur J Clin Invest1992;22:396–402.
[152] Fang X, Schroder S, Hoeft A, Stuber F. Comparison of twopolymorphisms of the interleukin-1 gene family: interleukin-1receptor antagonist polymorphism contributes to susceptibility tosevere sepsis. Crit Care Med 1999;27:1330–4.

S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502 501
[153] McDevitt M, Wang H, Knobelman C, Newman M, Di Giovine F,Timms J, et al. Interleukin-1 genetic association with periodontitisin clinical practice. J Periodontol 2000;71:156–63.
[154] Nielsen H, Engelbrecht J, Brunak S, von Heijne G. Identificationof prokaryotic and eukaryotic signal peptides and prediction oftheir cleavage sites. Protein Eng 1997;10:1–6.
[155] Yin Z, Kwang J. Carp interleukin-1� in the role of an immuno-adjuvant. Fish Shellfish Immunol 2000;10:375–8.
[156] Hong S, Zou J, Crampe M, Peddie S, Scapigliati G, Bols N, etal. The production and bioactivity of rainbow trout (Oncorhynchusmykiss) recombinant IL-1 beta. Vet Immunol Immunopathol2001;81:1–14.
[157] Peddie S, Zou J, Cunningham C, Secombes C. Rainbow trout(Oncorhynchus mykiss) recombinant IL-1 beta and derived peptidesinduce migration of head kidney leucocytes in vitro. Fish ShellfishImmunol 2001;11:697–709.
[158] Weyts F, Verburg van Kemenade B, Flik G, Lambert J, WendelaarBonga S. Conservation of apoptosis as an immune regulatorymechanism: effects of cortisol and cortisone on carp lymphocytes.Brain Behav Immun 1997;11:95–105.
[159] Fraser A, Evan G. A license to kill. Cell 1996;85:781–4.[160] Johnson A, Bridgham J, Munks M, Witty J. Characterization of the
chicken interleukin-1 beta converting enzyme (caspase-1) cDNA andexpression of caspase-1 mRNA in the hen. Gene 1998;219:55–62.
[161] Inohara N, Nunez G. Genes with homology to mammalian apoptosisregulators identified in zebrafish. Cell Death Differ 2000;7:509–10.
[162] Laing K, Holland J, Bonilla S, Cunningham C, Secombes C. Cloningand sequencing of caspase-6 in rainbow trout,Oncorhynchusmykiss, and analysis of its expression under conditions known toinduce apoptosis. Dev Comp Immunol 2001;25:303–12.
[163] Irmler M, Hertig S, MacDonald H, Sadoul R, Bercherer J,Proudfoot A, et al. Granzyme A is an interleukin-1� convertingenzyme. J Exp Med 1995;181:1917–22.
[164] Black R, Kronheim S, Cantrell M, Deeley M, March C, Prickett K,et al. Generation of biologically active interleukin-1� by proteolyticcleavage of the inactive precursor. J Biol Chem 1988;263:9437–42.
[165] Beuscher H, Gunther C, Rollinghoff M. IL-1 beta is secreted byactivated murine macrophages as biologically inactive precursor. JImmunol 1990;144:2179–83.
[166] Lundqvist E, Companjen A, Prens E, Egelrud T. Biological activityof human interleukin-1 beta: comparison with recombinant humaninterleukin-1 beta. Eur Cytokine Netw 1998;9:41–6.
[167] Trotman C. Introns early: slipping lately? Trends Genet 1998;14:132–4.
[168] Gilbert W, De Souza S, Long M. Origin of genes. Proc Natl AcadSci USA 1997;94:7698–703.
[169] O’Neill R, Brennan F, Delbridge M, Crozier R, Graves J.De novoinsertion of an intron into the mammalian sex determining gene,SRY. Proc Natl Acad Sci USA 1998;95:1653–7.
[170] Tarrio R, Rodriguez-Trelles F, Ayala F. New Drosophila intronsoriginate by duplication. Proc Natl Acad Sci USA 1998;95:1658–62.
[171] Stylianou E, Saklatvala J. Interleukin-1. Int J Biochem Cell Biol1998;30:1075–9.
[172] Zou J, Holland J, Pleguezuelos O, Cunningham C, SecombesCJ. Factors influencing the expression of interleukin-1 beta incultured rainbow trout (Oncorhynchus mykiss) leucocytes. DevComp Immunol 2000;24:575–82.
[173] Brubacher J, Secombes C, Zou J, Bols N. Constitutive and LPSinduced gene expression in a macrophage like cell line fromthe rainbow trout (Oncorhynchus mykiss). Dev Comp Immunol2000;24:565–74.
[174] Zapata A, Torroba M, Sacedon R, Varas A, Vicente A. Structure ofthe lymphoid organs of elasmobranchs. J Exp Zool 1996;275:125–43.
[175] Jarrous N, Kaempfer R. Induction of human interleukin-1 geneexpression by retinoic acid and its regulation at processing ofprecursor transcripts. J Biol Chem 1994;269:23141–9.
[176] Wang T, Zou J, Cunningham C, Secombes C. Cloning andfunctional characterisation of the interleukin-1�1 promoter ofrainbow trout (Oncorhynchus mykiss). Biochem Biophys Acta2002;1575(1–3):108–16.
[177] Bensi G, Mora M, Raugei G, Buonamassa D, Rossini M, MelliM. An inducible enhancer controls the expression of the humaninterleukin-1 beta gene. Cell Growth Differ 1990;1:491–7.
[178] Buras J, Monks B, Fenton M. The NF-beta A binding element, notan overlapping NF-IL-6 binding element, is required for maximalIL-1 beta gene expression. J Immunol 1994;152:4444–54.
[179] Cogswell J, Godlevski M, Wisely G, Clay W, Leesnitzer L, WaysJ, et al. NF-kappa B regulates IL-1 beta transcription through aconsensus NF-kappa B binding site and a nonconsensus CRE-likesite. J Immunol 1994;153:712–23.
[180] Kominato Y, Galson D, Waterman W, Webb A, Auron P. Monocyteexpression of the human prointerleukin-1 beta gene (IL1�) isdependent on promoter sequences which bind the hematopoietictranscription factor Spi-1/PU.1. Mol Cell Biol 1995;15:58–68.
[181] Cahill C, Waterman W, Xie Y, Auron P, Calderwood S.Transcriptional repression of the pro-interleukin-1 beta gene byheat shock factor 1a. J Biol Chem 1996;271:24874–9.
[182] Husmann M, Jehnichen P, Jahn B, Schlosshan D, Romahn E, MarxJ. A novel SP-1 site in the human interleukin-1 beta promoterconfers preferential transcriptional activity in keratinocytes. Eur JImmunol 1996;26:3008–14.
[183] Tseng Y, Schuler L. Transcriptional regulation of interleukin-1beta gene by interleukin-1 beta itself is mediated in part by Oct-1in thymic stromal cells. J Biol Chem 1998;273:12633–41.
[184] Goto M, Katayama K, Shirakawa F, Tanaka I. Involvement ofNF-kappa B p50/p65 heterodimer in activation of the humanpro-interleukin-1 beta gene at two subregions of the upstreamenhancer element. Cytokine 1999;11:16–28.
[185] Roman J, Ritzenthaler J, Fenton M, Roser S, Schuyler W.Transcriptional regulation of the human interleukin-1 beta geneby fibronectin: Role of protein kinase C and activator protein 1(AP-1). Cytokine 2000;12:1581–96.
[186] Hiscott J, Marois J, Garoufalis J, Daddario M, Roulston A, KwanI, et al. Characterization of a functional NF-kappa-B site inthe human interleukin-1 beta promoter—evidence for a positiveautoregulatory loop. Mol Cell Biol 1993;13:6231–40.
[187] Sangrador-Vegas A, Martin S, O’Dea P, Smith T. Cloning andcharacterization of the Rainbow trout (Oncorhynchus mykiss) typeII interleukin-1 receptor Cdna. Eur J Biochem 2000;267:7031–7.
[188] Guida S, Heguy A, Melli M. The chicken IL-1 receptor—differential evolution of the cytoplasmic and extracellular domains.Gene 1992;111:239–43.
[189] Subramaniam S, Stansberg C, Olsen L, Zou J, Secombes C,Cunningham C. Cloning of aSalmo salarinterleukin-1 receptor-likecDNA. Dev Comp Immunol 2002;26(5):415–31.
[190] Klasing K, Peng R. Soluble type-I interleukin-1 receptor blockschicken IL-1 activity. Dev Comp Immunol 2001;25:345–52.
[191] Okamura H, Tsutsui H, Komatsu T, Yutsudo M, Hakura A,Tanimoto T, et al. Cloning of a new cytokine that inducesIFN-gamma production by T-cells. Nature 1995;378:88–91.
[192] Ushio S, Namba M, Okura T, Hattori K, Nukada Y, Akita K, etal. Cloning of the cDNA for human IFN-gamma lnducing factor,expression inEscherichia coli, and studies on the biologic activitiesof the protein. J Immunol 1996;156:4274–9.
[193] Bazan J, Timans J, Kastelein R. A newly defined interleukin-1?Nature 1996;379:591.
[194] Gu Y, Kuida K, Tsutsui H, Ku G, Hsiao K, Fleming M, etal. Activation of interferon-gamma inducing factor mediated byinterleukin-1 beta converting enzyme. Science 1997;275:206–9.
[195] Ghayur T, Banerjee S, Hugunin M, Butler D, Herzog L, Carter A, etal. Caspase-1 processes IFN-gamma inducing factor and regulatesLPS induced IFN-gamma production. Nature 1997;386:619–23.

502 S. Bird et al. / Cytokine & Growth Factor Reviews 13 (2002) 483–502
[196] Liu B, Novick D, Kim S, Rubinstein M. Production of abiologically active human interleukin-18 requires its prior synthesisas pro-IL-18. Cytokine 2000;12:1519–25.
[197] Torigoe K, Ushio S, Okura T, Kobayashi S, Taniai M, Kunikata T,et al. Purification and characterization of the human interleukin-18receptor. J Biol Chem 1997;272:25737–42.
[198] Puren A, Fantuzzi G, Dinarello C. Gene expression, synthesis, andsecretion of interleukin-18 and interleukin-1 beta are differentiallyregulated in human blood mononuclear cells and mouse spleencells. Proc Natl Acad Sci USA 1999;96:2256–61.
[199] Akita K, Ohtsuki T, Nukada Y, Tanimoto T, Namba M, Okura T, etal. Involvement of caspase-1 and caspase-3 in the production andprocessing of mature human interleukin-18 in monocytic THP.1cells. J Biol Chem 1997;272:26595–603.
[200] Tone M, Thompson S, Tone Y, Fairchild P, Waldmann H.Regulation of IL-18 (IFN-gamma inducing factor) gene expression.J Immunol 1997;159:6156–63.
[201] Shields D, Harmon D, Nunez F, Whitehead A. The evolution of he-matopoietic cytokine/receptor complexes. Cytokine 1995;7:679–88.
[202] Makalowski W, Zhang J, Boguski M. Comparative analysis of1196 orthologous mouse and human full length mRNA and proteinsequences. Genome Res 1996;6:846–57.
[203] Hurst L, Smith N. Do essential genes evolve slowly? Curr Biol1999;9:747–50.
[204] Harrison G, Wedlock D. Marsupial cytokines: structure, functionand evolution. Dev Comp Immunol 2000;24:473–84.
[205] Murphy P. Molecular mimicry and the generation of host defenseprotein diversity. Cell 1993;72:823–6.
[206] McFadden G, Lalani A, Everett H, Nash P, Xu X. Virus encodedreceptors for cytokines and chemokines. Semin Cell Dev Biol1998;9:359–68.
[207] Pease J, Murphy P. Microbial corruption of the chemokine system:An expanding paradigm. Semin Immunol 1998;10:169–78.
[208] Murphy P. Viral exploitation and subversion of the immune systemthrough chemokine mimicry. Nat Immunol 2001;2:116–22.
[209] Scheerlinck J. Functional and structural comparison of cytokinesin different species. Vet Immunol Immunopathol 1999;72:39–44.
[210] Ponting C, Russell R. Identification of distant homologues offibroblast growth factors suggests a common ancestor for allbeta-trefoil proteins. J Mol Biol 2000;302:1041–7.
[211] Russell R, Sasieni P, Sternberg M. Supersites within superfolds.Binding site similarity in the absence of homology. J Mol Biol1998;282:903–18.
[212] Russell R, Saqi M, Bates P, Sternberg M. Recognition of analogousand homologous protein folds—assessment of prediction successand associated alignment accuracy using empirical substitutionmatrices. Protein Eng 1997;11:1–9.
[213] Eisenberg S, Brewer M, Verderber E, Heimdal P, Brandhuber B,Thompson R. Interleukin-1 receptor antagonist is a member ofthe interleukin-1 gene family—evolution of a cytokine controlmechanism. Proc Natl Acad Sci USA 1991;88:5232–6.
[214] Hughes A. Evolution of the interleukin-1 gene family in mammals.J Mol Evol 1994;39:6–12.
[215] Green E. Strategies for the systematic sequencing of complexgenomes. Nat Rev Genet 2001;2:573–83.
[216] Krin E, Hommais F, Soutourina O, Ngo S, Danchin A, Bertin P.Description and application of a rapid method for genomic DNAdirect sequencing. FEMS Microbiol Lett 2001;199:229–33.
[217] Beschin A, Bilej M, Torreele E, De Baetselier P. On the existenceof cytokines in invertebrates. Cell Mol Life Sci 2001;58:801–14.