endogenous interleukin (il)–1α and il‐1β are crucial for host defense against disseminated...
TRANSCRIPT

IL-1 in Disseminated Candidiasis • JID 2006:193 (15 May) • 1419
M A J O R A R T I C L E
Endogenous Interleukin (IL)–1a and IL-1b AreCrucial for Host Defense against DisseminatedCandidiasis
Alieke G. Vonk,1,3 Mihai G. Netea,1,3 Johan H. van Krieken,2 Yoichiro Iwakura,4 Jos W. M. van der Meer,1,3
and Bart Jan Kullberg1,3
Departments of 1Medicine and 2Pathology, Radboud University Nijmegen Medical Center, and 3Nijmegen University Center for Infectious Diseases,Nijmegen, The Netherlands; 4Laboratory Animal Research Center, Institute of Medical Science, University of Tokyo, Tokyo, Japan
Background. Interleukin (IL)–1a and IL-1b are protective proinflammatory cytokines involved in host defenseagainst Candida albicans. It is, however, unknown whether they provide protection through similar mechanisms.We investigated the effect of endogenous IL-1a and IL-1b on disseminated C. albicans infection.
Methods. Mice deficient in the genes encoding IL-1a (IL-1a�/�), IL-1b (IL-1b�/�), or both molecules (IL-1a�/�b�/�) were used. Survival and C. albicans outgrowth in the kidneys was assessed after intravenous injectionof C. albicans.
Results. Both mortality and C. albicans outgrowth in the kidneys were significantly increased in IL-1a�/� andIL-1b�/� mice, compared with those in control mice, with the IL-1a�/�b�/� mice being most susceptible todisseminated candidiasis. The host defense mechanisms triggered by IL-1a and IL-1b differed from one another.IL-1b�/� mice showed decreased recruitment of granulocytes in response to an intraperitoneal C. albicans challenge,and generation of superoxide production was diminished in IL-1b�/� granulocytes. IL-1a�/� mice had a reducedcapacity to damage C. albicans pseudohyphae. Protective type 1 responses were deficient in both IL-1a�/� and IL-1b�/� mice, as assessed by production of interferon-g by splenocytes in response to heat-killed C. albicans.
Conclusion. Although IL-1a and IL-1b have differential effects on the various arms of host defense, bothcytokines are essential for mounting a protective host response against invasive C. albicans infection.
Despite the availability of potent antifungal agents,
acute disseminated candidiasis remains a life-threat-
ening disease that occurs mainly in immunocompro-
mised patients [1]. Immunotherapies with cytokines
have great potential to augment host resistance and as
adjunctive treatment for invasive candidiasis. For fur-
ther development of these strategies, a better under-
standing of the protective immune mechanisms against
invasive candidiasis is needed.
Interleukin (IL)–1a and IL-1b are proinflammatory
cytokines that exert similar biological activities after
Received 5 July 2005; accepted 29 December 2005; electronically published 4April 2006.
Potential conflicts of interest: none reported.Financial support: Netherlands Organisation for Scientific Research (VIDI grant
to M.G.N.).Reprints or correspondence: Dr. Bart Jan Kullberg, Dept. of Medicine (463),
Radboud University Nijmegen Medical Center, P.O. Box 9101, 6500 HB Nijmegen,The Netherlands ([email protected]).
The Journal of Infectious Diseases 2006; 193:1419–26� 2006 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2006/19310-0012$15.00
interaction with the IL-1 type I receptor (IL-1RI) and
the IL-1R accessory protein [2]. Exogenous recombi-
nant human IL-1a or IL-1b has been administered in
studies of disseminated murine candidiasis, and these
studies have clearly indicated a protective role for IL-
1 in this infection model [3, 4]. The mechanisms of
this beneficial effect have been only partly elucidated.
IL-1 has no direct antifungal effect, and the protective
effect of IL-1 in host defense against Candida albicans
does not depend on the presence of granulocytes or
humoral factors, such as acute-phase proteins [3, 4].
To characterize the role of endogenous IL-1a and IL-
1b in disseminated candidiasis and to gain further in-
sight into the mechanisms through which both IL-1
molecules confer protection against disseminated can-
didiasis, mice in which the genes encoding IL-1a (IL-
1a�/�), IL-1b (IL-1b�/�), or both (IL-1a�/�b�/�) had
been disrupted were used in the present study. The IL-
1–deficient mice and their immunocompetent litter-
mates were subjected to experimental disseminated C.
albicans infection.
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

1420 • JID 2006:193 (15 May) • Vonk et al.
MATERIALS AND METHODS
Mice. IL-1a�/�, IL-1b�/�, and IL-1a�/�b�/� mice were pro-
duced as described elsewhere [5]. As control mice for the IL-
1–deficient mice, age-matched C57Bl/6 (IL-1a+/+b+/+) mice were
used. The mice were allowed to become accustomed to laboratory
conditions for 1 week before experimental use as described below.
Infection model. Mice were injected intravenously with
C. albicans (ATCC MYA-3573; UC820) blastoconidia,51 � 10
and survival was assessed. For measurement of circulating gran-
ulocytes and cytokines, mice were bled from the retroorbital
plexus on day 1, 3, or 7 of infection. To quantify fungal out-
growth, the kidneys were removed aseptically, weighed, and ho-
mogenized, and serial dilutions were plated on Sabouraud agar,
as described elsewhere [3]. Colony-forming units were counted,
and results were expressed as log colony-forming units per kid-
ney. For histologic analysis, kidneys of subgroups of mice (5
mice/group) were fixed in buffered formaldehyde (4%). Paraffin-
embedded sections were stained with periodic acid–Schiff or he-
matoxylin-eosin.
To investigate whether the role of endogenous IL-1 in candi-
diasis is mediated by polymorphonuclear neutrophils (PMNs),
mice were rendered granulocytopenic by use of cyclophospha-
mide (Bristol-Myers Squibb), administered subcutaneously at a
dose of 150 mg/kg on day �4 and at a dose of 100 mg/kg on
days �1 and 2 of infection with C. albicans blastoconidia41 � 10
[3]. Daily differential counts in peripheral blood smears con-
firmed granulocytopenia (! cells/L; data not shown).6100 � 10
Hyperuricemia-induced tumor necrosis factor (TNF) production
was prevented by gastric instillation of sodium bicarbonate at a
dose of 100 mg/kg twice per day, starting on day �4 and con-
tinuing until the end of the experiment [6]. C. albicans outgrowth
in the kidneys on day 1 or 3 of infection was determined as
described elsewhere [3].
PMN recruitment. PMN recruitment to an infection site
was determined after intraperitoneal injection of cfu of71 � 10
heat-killed C. albicans in uninfected mice. After 4 h, mice were
killed by CO2 asphyxiation, and peritoneal exudates were ob-
tained by washing with ice-cold PBS. The numbers of PMNs
were assessed in Giemsa-stained cytocentrifuge preparations.
Superoxide production. Superoxide production was studied
in a luminol-enhanced peroxidase-catalyzed chemiluminescence
assay [7]. Briefly, peritoneal exudate PMNs were suspended at a
concentration of cells/mL of Hanks’ balanced salt so-62 � 10
lution without phenol red (Gibco), supplemented with 0.25%
human serum albumin. Cells ( /well) were incubated in52 � 10
96-well microtiter plates (Costar Corning) with 50 mmol/L lu-
minol and 4.5 U/mL horseradish peroxidase (Sigma) and were
stimulated with either medium (as a negative control) or 2�
108 heat-killed blastoconidia and PMA (50 ng/mL). Chemilu-
minescence was measured on a Victor2 1420 counter and ex-
pressed as the total amount of superoxide produced in 35 min,
by integrating the area under the curve per PMN.
Phagocytosis and intracellular killing of C. albicans. Exu-
date peritoneal phagocytes were collected 4 h (PMN) or 72 h
(macrophages) after intraperitoneal injection of 10% proteose
peptone. Cells were centrifuged at 550 g, counted, and resus-
pended in RPMI 1640 Dutch modification (with 20 mmol/L
HEPES, without glutamine; RPMI-dm; ICN Biomedicals) sup-
plemented with 5% heat-inactivated fetal calf serum.
Phagocytosis and intracellular killing were studied in an ad-
herent monolayer of phagocytes, as described elsewhere [8].
Briefly, monolayers were incubated with opsonized blasto-
conidia (ATCC 10261) in modified Eagle’s medium (Gibco)
(effector-to-target cell [E:T] ratio, 40:1). After 15 min, super-
natants containing noningested blastoconidia were plated on
Sabouraud agar. The percentage of phagocytosed blastoconidia
was calculated as [1 � (number of uningested cfu/cfu at the
start of incubation)] � 100. To assess the percentages of C. al-
bicans blastoconidia internalized versus those only attached to
the membrane, fluorescein isothiocyanate–labeled C. albicans
was opsonized and incubated for 15 min on a macrophage
monolayer (E:T ratio, 40:1, as described above). After washing
with sterile medium to remove the extracellular nonadherent
yeasts, the total fluorescence was measured in the monolayer
to enumerate the total of internalized and adherent C. albicans
cells. The fluorescence of the extracellular, membrane-adhered
C. albicans was quenched by adding methylene blue, the cells
were washed, and the number of fluorescent internalized blas-
toconidia was assessed. In 3 separate experiments, 90%–97%
of the total number of internalized and adherent blastoconidia
were shown to be intracellular, and no differences in the ratios
that were adherent to phagocytosed C. albicans cells between
wild-type and IL-1–deficient mice were apparent.
After removal of the nonphagocytosed blastoconidia, killing
of blastoconidia by PMNs was assessed in the same monolayers
in fresh medium. After 3 h of incubation at 37�C and 5% CO2,
the wells were gently scraped with a plastic paddle and washed
with 200 mL of distilled H2O to achieve lysis of macrophages.
This procedure was repeated 3 times, after which the pooled
washes were adjusted to a final volume of 1 mL with distilled
water. Microscopic examination of the culture plates showed
that there was a complete removal of phagocytes. Viable in-
tracellular blastoconidia were quantified as described elsewhere
[8]. The percentage of phagocyte-killed yeasts was determined
as [1�(cfu after incubation/number of phagocytized cfu)] �
100. Phagocyte-free incubations of blastoconidia were included
as controls for yeast viability.
Assessment of PMN-mediated pseudohyphal damage. PMN-
mediated pseudohyphal damage was determined by the XTT
dye assay, as described elsewhere [9]. ATCC MYA-3573 blas-
toconidia were suspended at cfu/mL of RPMI-dm (pH61 � 10
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

IL-1 in Disseminated Candidiasis • JID 2006:193 (15 May) • 1421
Figure 1. The survival of interleukin (IL)–1–deficient mice after an in-travenous injection of cfu of Candida albicans. Survival of IL-51 � 101a�/� mice and IL-1b�/� mice was significantly impaired, compared withthat of IL-1a+/+b+/+ mice ( ). IL-1a�/�b�/� mice showed significantlyP ! .05impaired survival, compared with that of IL-1a+/+b+/+ mice ( ) andP ! .001IL-1a�/� mice ( ). Data are the cumulative results of experimentsP ! .05performed in quadruplicate for at least 25 mice/group and were analyzedusing the Kaplan Meier log rank test.
Figure 2. Outgrowth of Candida albicans in the kidneys 7 days afteran intravenous injection of cfu of C. albicans. The log-transformed51 � 10data represent the cumulative results of experiments performed twiceand were analyzed using 1-way analysis of variance and the Mann-Whitney U test for posttest comparisons. Horizontal bars indicate themeans. a , vs. interleukin (IL)–1a+/+b+/+ mice; b , vs. IL-1b�/�P ! .001 P ! .01mice; c , vs. IL-1a+/+b+/+mice; d , vs. IL-1b�/� mice.P ! .01 P ! .05
6.4). After incubation at 37�C for 24 h, pseudohyphae were
obtained and resuspended in RPMI 1640 without phenol red
and l-glutamine (RPMI-wp; ICN Biomedicals). Pseudohyphae
( ) and PMNs in RPMI-wp were added to the wells in51 � 10
the presence of 10% fresh IL-1a+/+b+/+ serum (E:T ratio, 8:1).
Control wells contained pseudohyphae or PMNs only. After
incubation for 2 h, PMNs were lysed with sterile H2O. After
15 min, sterile XTT (400 mg/mL; Sigma Chemical) and coen-
zyme Q0 (50 mg/mL; Sigma) were added. After 1 h of incubation
at 37�C, the plate was centrifuged (770 g), the supernatants were
transferred to a microtiter plate, and the absorbance was mea-
sured in a spectrophotometer at 450 nm. The percentage of fungal
damage was calculated as 1� [(A450 hyphae and PMNs�A450
PMNs)/A450 hyphae] � 100.
Production of cytokines and NO. Resident peritoneal mac-
rophages were obtained aseptically with ice-cold PBS. Cells were
resuspended in RPMI-dm in a round-bottom 96-well plate (1
� 105 cells/well). For cytokine production, macrophages were
stimulated with culture medium as a negative control, heat-
killed blastoconidia (ATCC MYA-3573; ), or pseudo-71 � 10
hyphae (1 � 106). For cell-associated IL-1a, fresh RPMI was
added to the remaining macrophages, which were subsequently
disrupted by 3 freeze-thaw cycles. Supernatants collected after
24 h of incubation and cell lysates were stored at �80�C until
assay.
For NO production, macrophages ( cells/mL) were65 � 10
stimulated with heat-killed C. albicans ( cfu/mL) and71 � 10
interferon (IFN)–g (100 U/mL), or lipopolysaccharide (LPS)
(1 mg/mL) for 24 h at 37�C. The nitrite concentration in the
supernatants was determined by the Griess reaction [10].
Splenocyte stimulation. Fourteen days after an intravenous
injection of C. albicans, spleens were removed and gent-41 � 10
ly squeezed in sterile 200-mm filter chambers. Of the spleno-
cytes, 95% were lymphocytes, 2% were monocytes, and 3%
were granulocytes. Splenocytes ( cells/mL) were stim-65 � 10
ulated with control medium or heat-killed C. albicans71 � 10
(ATCC MYA-3573) blastoconidia (E:T ratio, 2:1). IFN-g and
IL-10 concentrations were measured in supernatants after 48
h of incubation at 37�C in 5% CO2 in 24-well plates (Greiner).
Cytokine assays. TNF-a, IL-1a, and IL-1b concentrations
were determined using specific radioimmunoassays, as described
elsewhere [11]. The detection limit was 40 pg/mL for TNF-a
and 20 pg/mL for IL-1a and IL-1b. IL-10, IFN-g, and IL-6
concentrations were determined by ELISA (Biosource), and the
detection limits were 8, 15.6, and 150 pg/mL, respectively. Mu-
rine keratinocyte-derived chemokine (KC), macrophage inhib-
itory protein (MIP)–2, and monocyte chemotactic protein–1
(MCP-1) were measured by ELISA (R&D Systems).
Statistical analysis. Parametric data are expressed as mean
�SD, and data that showed normal distribution after log trans-
formation are expressed as means and 95% confidence intervals
(CIs). Nonparametric values are expressed as medians. Since
�3 groups were compared, parametric data were analyzed using
1-way analysis of variance (ANOVA), and nonparametric data
were analyzed using the Kruskal-Wallis 1-way ANOVA. For post-
test comparisons of nonparametric data, the Mann-Whitney U
test was applied. The Kaplan-Meier log rank test was used to
analyze survival data. The data represent the pooled results of
all experiments performed.
RESULTS
Disseminated candidiasis in nonneutropenic mice. Whereas
72% of the IL-1a+/+b+/+ mice survived infection with 51 � 10
cfu of C. albicans, only 41% of IL-1a�/� and 38% of IL-1b�/�
mice survived ( , control mice vs. IL-1a�/� or IL-1b�/�P ! .05
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from
1422 • JID 2006:193 (15 May) • Vonk et al.
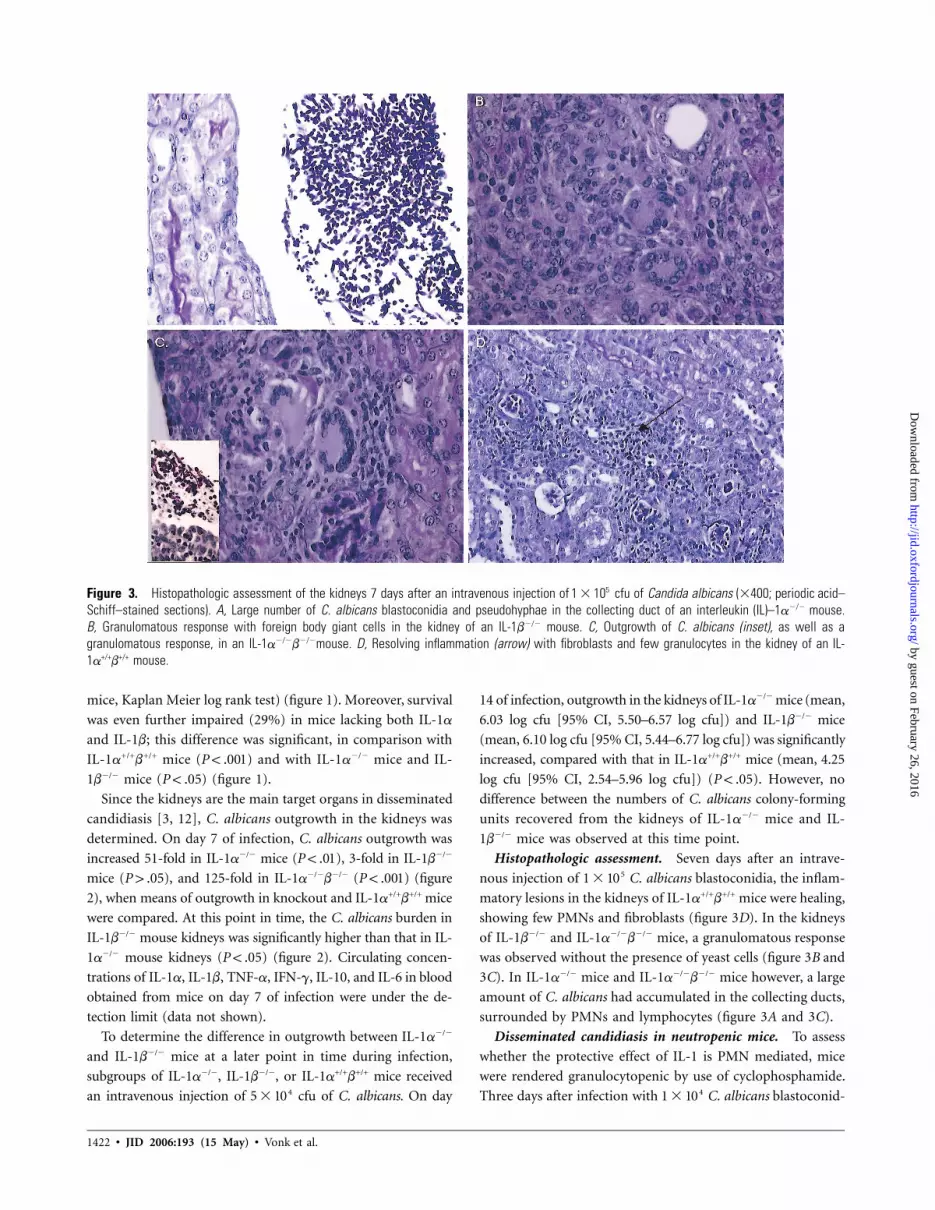
Figure 3. Histopathologic assessment of the kidneys 7 days after an intravenous injection of cfu of Candida albicans (�400; periodic acid–51 � 10Schiff–stained sections). A, Large number of C. albicans blastoconidia and pseudohyphae in the collecting duct of an interleukin (IL)–1a�/� mouse.B, Granulomatous response with foreign body giant cells in the kidney of an IL-1b�/� mouse. C, Outgrowth of C. albicans (inset), as well as agranulomatous response, in an IL-1a�/�b�/�mouse. D, Resolving inflammation (arrow) with fibroblasts and few granulocytes in the kidney of an IL-1a+/+b+/+ mouse.
mice, Kaplan Meier log rank test) (figure 1). Moreover, survival
was even further impaired (29%) in mice lacking both IL-1a
and IL-1b; this difference was significant, in comparison with
IL-1a+/+b+/+ mice ( ) and with IL-1a�/� mice and IL-P ! .001
1b�/� mice ( ) (figure 1).P ! .05
Since the kidneys are the main target organs in disseminated
candidiasis [3, 12], C. albicans outgrowth in the kidneys was
determined. On day 7 of infection, C. albicans outgrowth was
increased 51-fold in IL-1a�/� mice ( ), 3-fold in IL-1b�/�P ! .01
mice ( ), and 125-fold in IL-1a�/�b�/� ( ) (figureP 1 .05 P ! .001
2), when means of outgrowth in knockout and IL-1a+/+b+/+ mice
were compared. At this point in time, the C. albicans burden in
IL-1b�/� mouse kidneys was significantly higher than that in IL-
1a�/� mouse kidneys ( ) (figure 2). Circulating concen-P ! .05
trations of IL-1a, IL-1b, TNF-a, IFN-g, IL-10, and IL-6 in blood
obtained from mice on day 7 of infection were under the de-
tection limit (data not shown).
To determine the difference in outgrowth between IL-1a�/�
and IL-1b�/� mice at a later point in time during infection,
subgroups of IL-1a�/�, IL-1b�/�, or IL-1a+/+b+/+ mice received
an intravenous injection of cfu of C. albicans. On day45 � 10
14 of infection, outgrowth in the kidneys of IL-1a�/� mice (mean,
6.03 log cfu [95% CI, 5.50–6.57 log cfu]) and IL-1b�/� mice
(mean, 6.10 log cfu [95% CI, 5.44–6.77 log cfu]) was significantly
increased, compared with that in IL-1a+/+b+/+ mice (mean, 4.25
log cfu [95% CI, 2.54–5.96 log cfu]) ( ). However, noP ! .05
difference between the numbers of C. albicans colony-forming
units recovered from the kidneys of IL-1a�/� mice and IL-
1b�/� mice was observed at this time point.
Histopathologic assessment. Seven days after an intrave-
nous injection of C. albicans blastoconidia, the inflam-51 � 10
matory lesions in the kidneys of IL-1a+/+b+/+ mice were healing,
showing few PMNs and fibroblasts (figure 3D). In the kidneys
of IL-1b�/� and IL-1a�/�b�/� mice, a granulomatous response
was observed without the presence of yeast cells (figure 3B and
3C). In IL-1a�/� mice and IL-1a�/�b�/� mice however, a large
amount of C. albicans had accumulated in the collecting ducts,
surrounded by PMNs and lymphocytes (figure 3A and 3C).
Disseminated candidiasis in neutropenic mice. To assess
whether the protective effect of IL-1 is PMN mediated, mice
were rendered granulocytopenic by use of cyclophosphamide.
Three days after infection with C. albicans blastoconid-41 � 10
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

IL-1 in Disseminated Candidiasis • JID 2006:193 (15 May) • 1423
Table 1. No. of circulating polymorphonuclear neutrophils(PMNs) before or on day 3 of infection with Candida albicansblastoconidia ( cfu administered intravenously), and re-51 � 10cruitment of PMNs 4 h after intraperitoneal injection of heat-killed C. albicans blastoconidia ( cfu).71 � 10
Strain
Circulating PMNs,no. �109/L
IntraperitonealPMN recruitment
Beforeinfection
On day 3of infection %a No. �106
IL-1a+/+b+/+ 0.89 � 0.31 0.84 � 0.71 63 � 13 5.81 � 3.26IL-1a�/� 0.42 � 0.10 0.86 � 0.30 54 � 6 5.56 � 1.62IL-1b�/� 1.38 � 0.47 3.20 � 0.99 37 � 12b 1.85 � 0.84c
IL-1a�/�b�/� 0.67 � 0.35 2.95 � 0.52 39 � 8b 3.21 � 0.70
NOTE. Data are the of at least 3 mice/group, were obtainedmean � SDfrom 1 experiment, and were analyzed using 1-way analysis of variance andthe Mann-Whitney U test for posttest comparisons. IL, interleukin.
a Percentage of the total no. of peritoneal exudate cellsb , vs. control or IL-1a�/� mice.P ! .005c , vs. control or IL-1a�/� mice.P ! .01
Figure 4. Percentage of exudate polymorphonuclear neutrophil (PMN)–mediated pseudohyphal damage. Pseudohyphae ( ) were incubated51 � 10in the presence of PMNs for 2 h. Data are the cumulative results58 � 10of 2 separate experiments and were analyzed using the Kruskal-Wallis1-way analysis of variance. For posttest comparisons, the Mann-WhitneyU test was applied. Horizontal bars indicate medians. a , vs. in-P ! .001terleukin (IL)–1a+/+b+/+ or IL-1b�/� mice.
ia, the largest quantity of C. albicans was recovered from the
kidneys of IL-1a�/�b�/� mice ( , logmean � SD 5.23 � 0.37
cfu), and the difference was significant in comparison with
IL-1a+/+b+/+ mice ( , log cfu) ( ).mean � SD 3.64 � 0.51 P ! .01
Outgrowth in the kidneys of IL-1a�/� mice ( ,mean � SD
4.27�1.31 log cfu) was similar to that in the kidneys of IL-
1b�/� mice ( , log cfu. (Data are cu-mean � SD 4.13 � 0.67
mulative results of 2 experiments for at least 6 mice/group.)
PMN circulating numbers and recruitment. To study the
role of endogenous IL-1a and IL-1b in the recruitment of
PMNs to the site of a localized C. albicans infection, peritoneal
PMNs were collected after intraperitoneal injection of heat-
killed C. albicans (table 1). Whereas the recruitment of PMNs
in IL-1a�/� mice did not differ from that in IL-1a+/+b+/+ mice,
significantly lower numbers of PMNs were recruited to the site
of infection in IL-1b�/� mice, indicating that IL-1b is most
important for PMN recruitment to the site of infection. In
contrast, during the early days of infection, increased numbers
of circulating PMNs were observed in IL-1b�/� mice and IL-
1a�/�b�/� mice, compared with those in IL-1a�/� and IL-
1a+/+b+/+ mice (table 1), underscoring the importance of IL-1b
in the migration of PMNs from the bloodstream to the site of
infection. The mean numbers of peripheral blood PMNs were
similar for all groups on day 7 of infection (data not shown).
Anticandidal response of PMNs against C. albicans blas-
toconidia and pseudohyphae. To further characterize the in-
fluence of endogenous IL-1a and IL-1b on the activity of PMNs
against C. albicans, the fungicidal capacity of PMNs was de-
termined in vitro. Whereas IL-1a�/� PMNs showed a tenden-
cy only toward reduced superoxide production ( ,mean � SD
58.3 � 103 �15.4 � 103 cps), IL-b�/� PMNs produced signifi-
cantly less superoxide than did control PMNs ( ,mean � SD
30.6 � 103 � 15.9 � 103 vs. cps) (3 371.5 � 10 � 10.5 � 10 P !
). However, this observation was not accompanied by an.01
altered capacity of IL-1–deficient PMNs to phagocytose or kill
internalized C. albicans blastoconidia; all groups phagocytosed
∼25% of the blastoconidia and killed ∼75% of internalized
blastoconidia.
In contrast, IL-1 modulated the capacity of PMNs to damage
pseudohyphae (figure 4). Pseudohyphal damage caused by IL-
1a�/� or IL-1a�/�b�/� PMNs was significantly less efficient than
that caused by IL-1a+/+b+/+ or IL-1b�/� PMNs ( ) (figureP ! .001
4). Deficiency of both IL-1 molecules showed a tendency to
further impair PMN antipseudohyphal resistance, compared
with IL-1a deficiency alone. Deficiency of IL-1b alone did
not diminish the capacity of PMNs to damage C. albicans
pseudohyphae.
Macrophage function and stimulation. Macrophage re-
cruitment was determined by harvesting peritoneal exudate
cells 72 h after an intraperitoneal injection of cfu of71 � 10
heat-killed C. albicans. No differences in the num-mean � SD
bers of macrophages that were recruited into the peritoneal
cavity were observed, compared with those in control mice
(IL-1a+/+b+/+, ; IL-1a�/�,9 9 91.01 � 10 � 0.12 � 10 1.10 � 10 �
; IL-1b�/�, ; IL-1a�/�b�/�,9 9 90.02 � 10 0.96 � 10 � 0.19 � 10
). IL-1 influenced neither the9 91.18 � 10 � 0.15 � 10 mean �
capacity of exudate peritoneal macrophages to phagocytoseSD
C. albicans blastoconidia (IL-1a+/+b+/+, ; IL-1a�/�,41% � 5%
; IL-1b�/�, ; IL-1a�/�b�/�, )50% � 4% 56% � 3% 41% � 16%
nor their capacity to damage C. albicans pseudohyphae (data
not shown). No differences between NO production by mac-
rophages from the mouse strains were observed in response to
either heat-killed C. albicans (all around the detection limit) or
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

1424 • JID 2006:193 (15 May) • Vonk et al.
Table 2. In vitro cytokine and chemokine production by resident peritoneal macrophages( ) stimulated with either heat-killed Candida albicans blastoconidia ( ) or heat-killed5 71 � 10 1 � 10pseudohyphae ( ).61 � 10
Stimulation, strain TNF-a, ng/mL IL-6, pg/mL MCP-1, pg/mL MIP-2, pg/mL KC, pg/mL
BlastoconidiaIL-1a+/+b+/+ 0.56 � 0.14 926 � 547 87 � 45 2190 � 1633 1842 � 880IL-1a�/� 0.88 � 0.41 154 � 9a 40 � 1 2660 � 1630 380 � 197IL-1b�/� 0.46 � 0.22 329 � 235 50 � 22 2166 � 1769 308 � 345a
IL-1a�/�b�/� 0.72 � 0.41 159 � 21a 1674 � 753 265 � 254a
PseudohyphaeIL-1a+/+b+/+ 1.32 � 0.82 1378 � 796 221 � 135 3350 � 2699 1920 � 879IL-1a�/� 0.95 � 0.42 366 � 203 152 � 154 2550 � 1656 435 � 196IL-1b�/� 0.74 � 0.29 417 � 268 98 � 56 2495 � 1860 99 � 87a
IL-1a�/�b�/� 0.98 � 0.39 233 � 187a 1295 � 510 303 � 326a
NOTE. Data are the production in 5 mice/group, obtained from 1 experiment, and were analyzedmean � SDusing 1-way analysis of variance and the Mann-Whitney U test for posttest comparisons. IL, interleukin; KC, keratin-ocyte-derived chemokine; MCP, monocyte chemotactic protein; MIP, macrophage inhibitory protein; TNF, tumor ne-crosis factor
a , vs. IL-1a+/+b+/+ mice.P ! .05
LPS (control mice, ng/mL; IL-1b�/� mice, 0.78�1.37 � 0.54
1.35 ng/mL; IL-a�/�b�/� mice, ng/mL; ).5.28 � 0.2 P 1 .05
IL-1–deficient resident peritoneal macrophages showed im-
paired production of IL-6 and of the chemokine KC in response
to heat-killed C. albicans blastoconidia or pseudohyphae. The
difference in IL-6 production was significant for IL-1a�/� mac-
rophages and IL-1a�/�b�/� macrophages, in comparison with
IL-1a+/+b+/+ macrophages, and the difference in KC production
was significant for IL-1b�/� macrophages and IL-1a�/�b�/� mac-
rophages, in comparison with IL-1a+/+b+/+ macrophages (P !
) (table 2). No differences between groups were observed in.05
the production of TNF-a, MCP-1, or MIP-2.
Stimulation of splenic lymphocytes. To assess whether IL-
1 induces a type 1 or a type 2 immune response, splenocytes
were stimulated with heat-killed C. albicans. Whereas IL-
1a+/+b+/+ splenocytes produced IFN-g, neither IL-1a�/� nor IL-
1b�/� splenocytes did ( ) (table 3), indicating that bothP ! .05
IL-1a and IL-1b are important for protective IFN-g produc-
tion. Whereas IL-1b�/� and IL-1a�/�b�/� splenocytes did not
produce any IL-10, both IL-1a+/+b+/+ splenocytes and IL-1a�/�
splenocytes did ( ) (table 3), indicating that both IL-1aP ! .05
and IL-1b are important for protective IFN-g production,
whereas the residual IL-10 production in IL-1a�/� mice further
contributed to a type 2 response.
DISCUSSION
The results of the present study indicate that deficiency of
endogenous IL-1a or IL-1b has deleterious effects on the out-
come of disseminated candidiasis. Both IL-1a�/� mice and IL-
1b�/� mice showed increased mortality associated with an in-
creased outgrowth of C. albicans in the kidneys. IL-1b proved
to be important for PMN recruitment and generation of su-
peroxide production. IL-1a was essential for the capacity of
PMNs to damage C. albicans pseudohyphae, and both IL-1a
and IL-1b were required for the induction of protective Th1
responses.
One of the mechanisms through which IL-1 has been sug-
gested to confer protection is the enhancement of granulo-
poiesis and influx of PMNs to the site of infection. Although
IL-1a�/� mice had 50% fewer circulating granulocytes at the
time of infection than control mice, their number in the cir-
culation on day 3 of infection and PMN recruitment to the
site of infection did not differ from that of control mice. In
IL-1b�/� mice, however, a trend toward an increased number
of circulating granulocytes during infection was observed, and
this coincided with significantly decreased PMN migration to
the site of infection in a standardized model of intraperitoneal
C. albicans challenge. This is in line with earlier evidence that
exogenous administration of IL-1a or IL-1b induces peripheral
blood granulocytosis [13, 14]. Our data on PMN recruitment
in response to heat-killed C. albicans also suggest an important
role for endogenous IL-1b in the early PMN influx, in line with
studies showing that exogenous administration of IL-1 induces
PMN accumulation [15, 16]. Moreover, PMN recruitment in IL-
1–deficient mice may be further impaired as a result of decreased
production of neutrophil chemoattractants. Whereas IL-1a�/�
mice showed a tendency toward reduced production of the CXC
chemokine KC, KC production by IL-1b�/� macrophages was
significantly reduced, thus likely contributing to impaired gran-
ulocyte recruitment in IL-1b�/� mice. Whereas the differences
in chemokine production between IL-1a�/� and IL-1b�/� mice
may not be impressive, the major difference between the mouse
strains was the complete absence of IL-1b in the IL-1b�/� mice.
Since IL-1b has been shown to be an important mediator of
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

IL-1 in Disseminated Candidiasis • JID 2006:193 (15 May) • 1425
Table 3. In vitro cytokine production bysplenocytes ( cells/mL) stimulated with65 � 10heat-killed Candida albicans blastoconidia(107/mL).
Strain IFN-g, pg/mL IL-10, pg/mL
IL-1a+/+b+/+ 47.4 � 30.7 32.3 � 8.8IL-1a�/� !15.6a 30.8 � 12.2IL-1b�/� !15.6a
!8b
IL-1a�/�b�/� !15.6a!8b
NOTE. Data were obtained from 1 experiment andare expressed as the production in 5 mice/mean � SDgroup. IFN, interferon; IL, interleukin.
a , vs. IL-1a+/+b+/+ mice.P ! .05b , vs. IL-1a+/+b+/+ and IL-1a�/� mice.P ! .05
granulocyte recruitment [17, 18], this effect most likely is in-
strumental in explaining the difference in recruitment between
the IL-1a�/� and IL-1b�/� mice. However, it has to be taken into
account that assessment of PMN recruitment to a peritoneal
inflammatory stimulus is, at best, an approximation of early
parenchymal PMN recruitment at the various sites of infection.
Transcription of the genes encoding MIP-2 and KC is in-
duced through signals mediated by Toll-like receptor 4 [19],
and IL-1 has been shown to selectively stabilize KC mRNA
[20]. Hence, it is hypothesized that the normal production of
MIP-2—in contrast to the 80% reduced production of KC by
IL-1–deficient macrophages—is due to the selective effect of
IL-1 on KC mRNA.
The effect of endogenous IL-1 on PMN function has not
been investigated previously. The results of the present study
indicate that IL-1 affects PMN function and that the modes
of action differ for IL-1a and IL-1b. Whereas IL-1b�/� PMNs
showed impaired superoxide production, IL-1a�/� PMNs
showed a decreased capacity to damage C. albicans pseudo-
hyphae, indicating that IL-1a is important for PMN degranu-
lation. In addition, production of IL-6, which is induced by IL-
1 and known to stimulate PMN function [21–23], was reduced
only in IL-1b�/� mice, and this may further contribute to im-
paired PMN function and reduced anticandidal defense. Since
the C. albicans burden in IL-1a�/� kidneys was significantly
higher than that in IL-1b�/� kidneys on day 7 of infection, it is
suggested that the initially reduced PMN recruitment in IL-
1b�/� mice is overridden by the effect of the reduced capacity
to kill C. albicans pseudohyphae in IL-1a�/� mice. However,
despite the difference in outgrowth between the 2 mouse strains
on day 7, IL-1b�/� mice showed susceptibility to infection similar
to that of IL-1a�/� mice. Most of the apparent discrepancy is
explained by the fact that, on day 14 of infection, the fungal
burden in both IL-1a�/� mice and IL-1b�/� mice was equally
increased. Another possible explanation for this observation is
that, in addition to fungal outgrowth, the additional inflam-
matory damage in the organs of IL-1b�/� mice may also have
contributed to mortality.
If the beneficial effect of endogenous IL-1 is exerted only
through PMNs, the differences in outgrowth between the groups,
as observed in nongranulocytopenic mice, should disappear in
granulocytopenic mice. However, the observed differences per-
sisted, indicating that the effect of endogenous IL-1 is at least
partly mediated through cells or mechanisms other than mod-
ulation of PMN recruitment and function, which is in line with
previous findings [3, 4]. Nevertheless, because cyclophospha-
mide also reduces lymphocyte numbers, it cannot be excluded
that a minor beneficial effect of IL-1 on C. albicans outgrowth
may be mediated through PMNs.
Macrophages and lymphocytes are other cells that are likely
to mediate the protective effect of IL-1 [24, 25]. Most impor-
tantly, IFN-g production was found to be absent in IL-1–de-
ficient splenocytes. This is in agreement with previous data
showing that endogenous IL-1 is important for the production
IFN-g during C. albicans stimulation of whole blood [26]. The
crucial role of IFN-g in host defense has become apparent from
studies showing that IFN-g–deficient mice are highly suscep-
tible to disseminated candidiasis and that administration of re-
combinant IFN-g reduces outgrowth [12, 27, 28]. Recently, we
observed that delayed development of a type 1 response—that
is, delayed IFN-g production—contributed to increased out-
growth in localized C. albicans infection [9]. Furthermore, we
observed foreign body giant cells (FBGCs) in the kidneys of
IL-1–deficient mice. The transition of macrophages to func-
tionally inactive FBGCs is favored in the presence of type 2
cytokines [29]. Therefore, the presence of FBGCs points to an
anti-inflammatory type 2 response, which has been shown to
be detrimental to anticandidal host defense [30].
In conclusion, the present study has further clarified the
mechanisms through which IL-1a and IL-1b enhance host re-
sistance against candidiasis. Interestingly, the absence of either
IL-1a or IL-1b had divergent consequences. Although they are
products of different genes, IL-1a and IL-1b are highly ho-
mologous and have similar tridimensional structures and com-
mon receptors. It is unclear at this point what determines the
functional differences in IL-1a�/� and IL-1b�/� mice—a dif-
ferent distribution of cytokines in the organs, the known di-
chotomy between cell-bound IL-1a and secreted IL-1b [31],
or differential receptor affinity. Although a functional difference
between IL-1a and IL-1b was observed in this study, both IL-
1a and IL-1b were shown to be essential for anticandidal host
defense. The complementary roles of IL-1a and IL-1b in host
defense against C. albicans infection are further underscored
by the even higher susceptibility to disseminated candidiasis of
mice lacking both IL-1a and IL-1b. These data constitute an
additional body of evidence that IL-1–dependent mechanisms
play an important role in innate host defense against C. albicans.
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

1426 • JID 2006:193 (15 May) • Vonk et al.
Acknowledgments
We are greatly indebted to Ineke Verschueren, Debby Smits, and Maichelvan Riel, for their help with the experiments.
References
1. Blumberg HM, Jarvis WR, Soucie JM, et al. Risk factors for candidalbloodstream infections in surgical intensive care unit patients: theNEMIS prospective multicenter study. The National Epidemiology ofMycosis Survey. Clin Infect Dis 2001; 33:177–86.
2. Dinarello CA. Interleukin-1. Cytokine Growth Factor Rev 1997; 8:253–65.
3. Kullberg BJ, van ’t Wout JW, van Furth R. Role of granulocytes inincreased host resistance to Candida albicans induced by recombinantinterleukin-1. Infect Immun 1990; 58:3319–24.
4. van ’t Wout JW, van der Meer JW, Barza M, Dinarello CA. Protectionof neutropenic mice from lethal Candida albicans infection by recom-binant interleukin 1. Eur J Immunol 1988; 18:1143–6.
5. Horai R, Asano M, Sudo K et al. Production of mice deficient in genesfor interleukin (IL)-1alpha, IL-1beta, IL-1alpha/beta, and IL-1 receptorantagonist shows that IL-1beta is crucial in turpentine-induced feverdevelopment and glucocorticoid secretion. J Exp Med 1998; 187:1463–75.
6. Netea MG, Kullberg BJ, Blok WL, Netea RT, van der Meer JW. The roleof hyperuricemia in the increased cytokine production after lipopoly-saccharide challenge in neutropenic mice. Blood 1997; 89:577–82.
7. Lundqvist H, Dahlgren C. Isoluminol-enhanced chemiluminescence:a sensitive method to study the release of superoxide anion from humanneutrophils. Free Radic Biol Med 1996; 20:785–92.
8. Vonk AG, Wieland CW, Netea MG, Kullberg BJ. Phagocytosis andintracellular killing of Candida albicans blastoconidia by neutrophilsand macrophages: a comparison of different microbiological test sys-tems. J Microbiol Methods 2002; 49:55–62.
9. Vonk AG, Netea MG, Van Krieken JH, van der Meer JW, Kullberg BJ.Delayed clearance of intraabdominal abscesses caused by Candida al-bicans in tumor necrosis factor–a– and lymphotoxin-a–deficient mice.J Infect Dis 2002; 186:1815–22.
10. Ding AH, Nathan CF, Stuehr DJ. Release of reactive nitrogen inter-mediates and reactive oxygen intermediates from mouse peritonealmacrophages: comparison of activating cytokines and evidence for in-dependent production. J Immunol 1988; 141:2407–12.
11. Netea MG, Demacker PN, Kullberg BJ, et al. Low-density lipoproteinreceptor-deficient mice are protected against lethal endotoxemia andsevere gram-negative infections. J Clin Invest 1996; 97:1366–72.
12. Kullberg BJ, van’t Wout JW, Hoogstraten C, van Furth R. Recombinantinterferon-g enhances resistance to acute disseminated Candida albi-cans infection in mice. J Infect Dis 1993; 168:436–43.
13. Annema A, Sluiter W, van Furth R. Effect of interleukin 1, macrophagecolony-stimulating factor, and factor increasing monocytopoiesis onthe production of leukocytes in mice. Exp Hematol 1992; 20:69–74.
14. Nakai S, Hirai Y. The therapeutic potential of interleukin-1 beta in the
treatment of chemotherapy- or radiation-induced myelosuppressionand in tumor therapy. Biotherapy 1989; 1:339–54.
15. Tessier PA, Naccache PH, Clark-Lewis I, Gladue RP, Neote KS, McCollSR. Chemokine networks in vivo: involvement of C-X-C and C-Cchemokines in neutrophil extravasation in vivo in response to TNF-alpha. J Immunol 1997; 159:3595–602.
16. Czuprynski CJ, Brown JF. Purified human and recombinant murineinterleukin-1 alpha induced accumulation of inflammatory peritonealneutrophils and mononuclear phagocytes: possible contributions toantibacterial resistance. Microb Pathog 1987; 3:377–86.
17. Patton LM, Saggart BS, Ahmed NK, Leff JA, Repine JE. Interleukin-1beta-induced neutrophil recruitment and acute lung injury in hamsters.Inflammation 1995; 19:23–9.
18. Mizgerd JP. Molecular mechanisms of neutrophil recruitment elicitedby bacteria in the lungs. Semin Immunol 2002; 14:123–32.
19. Netea MG, Van Der Graaf CA, Vonk AG, Verschueren I, van der MeerJW, Kullberg BJ. The role of Toll-like receptor (TLR) 2 and TLR4 inthe host defense against disseminated candidiasis. J Infect Dis 2002;185:1483–9.
20. Tebo JM, Datta S, Kishore R, et al. Interleukin-1-mediated stabilizationof mouse KC mRNA depends on sequences in both 5′- and 3′-un-translated regions. J Biol Chem 2000; 275:12987–93.
21. Huang Q, Yang J, Lin Y, et al. Differential regulation of interleukin 1receptor and Toll-like receptor signaling by MEKK3. Nat Immunol 2004;5:98–103.
22. Borish L, Rosenbaum R, Albury L, Clark S. Activation of neutrophilsby recombinant interleukin 6. Cell Immunol 1989; 121:280–9.
23. van Enckevort FH, Netea MG, Hermus AR, et al. Increased suscep-tibility to systemic candidiasis in interleukin-6 deficient mice. Med Mycol1999; 37:419–26.
24. Ashman RB, Papadimitriou JM. Production and function of cytokinesin natural and acquired immunity to Candida albicans infection. Mi-crobiol Rev 1995; 59:646–72.
25. Vazquez Torres A, Balish E. Macrophages in resistance to candidiasis.Microbiol Mol Biol Rev 1997; 61:170–92.
26. Netea MG, Stuyt RJ, Kim SH, van der Meer JW, Kullberg BJ, DinarelloCA. The role of endogenous interleukin (IL)–18, IL-12, IL-1b, andtumor necrosis factor–a in the production of interferon-g induced byCandida albicans in human whole-blood cultures. J Infect Dis 2002;185:963–70.
27. Balish E, Wagner RD, Vazquez Torres A, Pierson C, Warner T. Can-didiasis in interferon-g knockout (IFN-g�/�) mice. J Infect Dis 1998;178:478–87.
28. Stuyt RJ, Netea MG, Verschueren, I et al. Role of interleukin-18 in hostdefense against disseminated Candida albicans infection. Infect Immun2002; 70:3284–6.
29. Anderson JM. Multinucleated giant cells. Curr Opin Hematol 2000;7:40–7.
30. Romani L, Puccetti P, Bistoni F. Biological role of Th cell subsets incandidiasis. Chem Immunol 1996; 63:115–37.
31. Dinarello CA. Biologic basis for interleukin-1 in disease. Blood 1996;87:2095–147.
by guest on February 26, 2016http://jid.oxfordjournals.org/
Dow
nloaded from