characterization of the canine 3β-hydroxysteroid dehydrogenase and its expression in the corpus...
TRANSCRIPT

A
wrcrhc1
23p©
K
1
hcpa1tdod
(
0d
Journal of Steroid Biochemistry & Molecular Biology 101 (2006) 254–262
Characterization of the canine 3�-hydroxysteroid dehydrogenase and itsexpression in the corpus luteum during diestrus
M.P. Kowalewski a,∗, J.I. Mason b, A.F. Howie b, S.D. Morley b, G. Schuler a, B. Hoffmann a
a Clinic for Obstetrics, Gynecology and Andrology of Large and Small Animals, Justus-Liebig-University, Giessen,D-35392 Giessen, Germany
b Centre for Reproductive Biology, Division of Reproductive and Developmental Sciences, University of Edinburgh,Edinburgh EH16 4TJ, Scotland, UK
Received 15 March 2006; accepted 15 June 2006
bstract
3�-Hydroxysteroid dehydrogenase (3�HSD) is a key enzyme in the synthesis of bioactive steroid hormones. Objectives of the present studyere to clone canine 3�HSD and to investigate its expression in dog corpora lutea (CL) covering the periods of their formation, early and late
egression (days 5, 15, 25, 35, 45, 65 after ovulation). Complete complementary DNA sequence was amplified by RACE PCR. Subsequentloning revealed that the canine ovarian 3�HSD transcript was composed of a 5′-untranslated region (5′-UTR) of 126 nucleotides, an openeading frame (ORF) of 1122 nucleotides and a 3′-UTR of 441 nucleotides. The putative ORF encoded a 374 amino acid protein which remainsighly conserved (79–85% identity) between species. The transient expression of the cloned canine 3�HSD in a mammalian heterologousell expression system (HEK293T cells) identified the 3�HSD activity as the only activity of this canine enzyme (absence of any detectable7-hydroxysteroid dehydrogenase activity).
Qualitative RT-PCR revealed expression of 3�HSD on all days investigated and the signals were strongest on days 5 and 15, with day
5 intensity tending to decrease. However, variability between individual animals was high. The significant decrease in the expression of�HSD towards the end of diestrus as indicated by Real Time PCR (p < 0.01) and immunhistochemistry may indicate that the provision ofrogesterone is controlled by availability of the enzyme rather than the substrate.2006 Elsevier Ltd. All rights reserved.
daVesIawd
eywords: 3�-Hydroxysteroid dehydrogenase (3�HSD); Diestrus; Dog
. Introduction
A key enzyme in the steroidogenic pathway is 3�-ydroxysteroid dehydrogenase/isomerase (3�HSD). Itatalyzes the conversion of the 3�-hydroxy-�5-steroidsregnenolone, 17�-hydroxypregnenolone and dehydroepi-ndrosterone to the 3-keto-�4-steroids, progesterone (P),7�-hydroxyprogesterone and androstenedione. Apart fromhe classic steroidogenic organs, 3�HSD has also been
etected in some peripheral tissues, like sebaceous glandsf the rat and human skin [1–4]. Several isoforms have beenetected in the mouse, rat, hamster and human, showing∗ Corresponding author. Tel.: +49 641 99 38704; fax: +49 641 29328.E-mail address: [email protected]
M.P. Kowalewski).
gm
gp3r
960-0760/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.jsbmb.2006.06.029
ifferent kinetics and catalytic properties in respect to tissuend age [5]. Thus, mouse type I (adrenal/gonadal) and typeI (utero/embryonal) 3�HSD have been shown to also
xhibit a 17�HSD-like activity when using testosterone asubstrate [5] as does the rat type I (adrenal/gonadal) and typeV (skin/placenta) 3�HSD when using 5�-dihydrotesterones substrate [6–7]. On the other hand, no such featuresere observed for the human type I and -II 3�HSD [5]. Theifferent types of 3�HSD mRNA are products of distinctenes and they are not the result of alternative splicingechanisms [8–9].In the dog, the corpora lutea are the only source for pro-
esterone during the cycle and pregnancy [10–11]. In non-regnant bitches P-levels are highest between days 20 and0 after ovulation. They decrease constantly thereafter toeach anestric levels (<1 ng/ml) 80–100 days after ovulation

chemis
[teab6odlegsr
leTe
tohtCi
2
2
bt
lC
Pw−doWR(fi
2(c
(cf
RB
dscwotd
cTd
TL
P
I
35R
U
R
TR
M.P. Kowalewski et al. / Journal of Steroid Bio
12–13]. In spite of the many studies dealing with the regula-ion of the luteal function in the dog (see review by Hoffmannt al. [14]) the mechanisms allowing for this slow regressionre still obscure. The course of blood P-levels in pregnantitches exhibits an identical pattern until parturition about0–65 days after mating, when a rapid drop to basal levels isbserved. Also there, as well as in cases of spontaneous lutealysfunction leading to abortions, the mechanisms leading to aoss of steroidogenic capacity are still unclear [15–16]. How-ver, since the corpora lutea of the dog are independent ofonadotropic support during the first 20–30 days of their lifepan [17–19], any luteal failure during this period shouldesult from local dysregulations.
The conversion of pregnenolone to P by 3�HSD is theast step in the provision of P and any dysregulation in thexpression of 3�HSD should effect progesterone synthesis.he present study therefore tested for the expression of thisnzyme in dog corpora lutea in the course of diestrus.
To date, canine 3�HSD sequence has not been charac-erized. Thus, in order to obtain the respective informationn the mRNA-level and to define canine 3�HSD cDNA, weave isolated and characterized the full-length cDNA struc-ure of canine 3�HSD as expressed in the corpora lutea (CL).onsequently the catalytic properties were tested following
ts expression in transfected HEK293T cells.
. Material and methods
.1. Animals and tissue preservation
On each of the days 5, 15, 35, 45 and 65 after ovulation fiveitches (various breeds, proven health status) were ovariohys-
erectomized except that six bitches were done on day 25.Animal treatment was in accordance with animal welfareegislation (permit no II 25-3-19c20/15c GI 18/14). TheL were separated from the surrounding ovarian tissue.
SC((
able 1ist of primers used for RT-PCR, RACE PCR and Real Time (TaqMan) PCR and th
CR method Primer Primer no. Primer sequence
nitial RT-PCR Forward 1 5′-CAAGGACAGTTCTReverse 2 5′-ATTGGACTGAGCA
′-RACE PCR GSP1 3 5′-CATCAAAGCTATG′-RACE PCR GSP2 4 5′-AGGCAAGCCAGTAT-PCR of full size ORF Forward 5 5′-TTGGGACCCAGAT
Reverse 6 5′-TTGCCATCCTCAGniversal Primer Mix UPM 7 5′-CTAATACGACTCA
8 5′-CTAATACGACTCAeal Time TaqMan-PCR Forward 9 5′-GGGTACTCAGCTC
Reverse 10 5′-GCCACCTCTATGGaqMan Probe Probe 11 5′-TGCCCAGGCTAGTT-PCR; canine ORFflanked by SalI(forward) and NotI(reward) restrictionsites (underlined)
Forward 12 5′-CGGCACGTCGACTReward 13 5′-ATAAAAGCGGCCG
try & Molecular Biology 101 (2006) 254–262 255
reservation of the tissue for RNA extraction was performedith Tissue tec® O.C.T. (Vogel, Giessen) and stored at80 ◦C. For handling reasons this procedure was replaced
uring the course of the experiments by incubating the CLvernight in RNAlater® (Ambion Biotechnologie GmbH,iesbaden), an aqueous reagent for stabilization of cellularNA (ratio mass:volume = 1:6). For immunohistochemistry
IHC) and in situ hybridization (ISH) tissue samples werexed with 10% neutral phosphate buffered formaline.
.2. Cloning of canine 3βHSD by reverse transcriptionRT)- and rapid amplification of cDNA ends-polymerasehain reaction (RACE PCR)
RT-PCR and rapid amplification of 5′- and 3′-cDNA ends5′- and 3′-RACE)-PCR were performed on total RNA ofanine CL from four different bitches to determine the cDNAor canine 3�HSD.
Total RNA was isolated from canine CL using Trizol®-eagent according to the manufacturer’s protocol (Gibco-RL, Life Technologies, Karlsruhe).
For initial RT-PCR 0.2 �g of total RNA was used. Primerserived from alignment of known 3�HSD homologousequences (primers 1–2; see Table 1) served to obtain initialanine-specific PCR product. In every experiment autoclavedater was used instead of RNA as a negative control. Integrityf RNA and the assay procedure was tested by amplifica-ion of the house-keeping gene glyceraldehyde-3-phosphateehydrogenase (GAPDH) (not shown).
Using approximately 0.6 �g of total RNA, first-strandDNA synthesis was done with the PowerScript Reverseranscriptase kit (BD Biosciences Clontech GmbH, Hei-elberg, Germany). 5′-3′-RACE PCR was performed using
MART RACE cDNA Amplification kit (BD Bioscienceslontech GmbH). Employing gene-specific primers (GSP)primers 3–4) in combination with universal primer mixUPM; primers 7–8) supplied with the kit, overlapping prod-
eir binding sites in relation to the nucleic acid sequence of canine 3�HSD
Binding sites
ACTACAT-3′ 874–893GGAAG-3′ 1049–1032ATGACCTCAAT-3′ 910–933TTCCAGAGAT-3′ 1012–990TTGACC-3′ 108–126ATCAGTG-3′ 1262–1243CTATAGGGCAAGCAG TGGTATCAACGCAGAGT-3′CTATAGGGC-3′CTGTTGGAA-3′ 435–456TGCTGGTAT-3′ 512–491GTGCCGATCTT-3′ 462–485AATCTAATTTGCTCT CCAG-3′ 62–81CAACAACATCTAGGT CTACAT-3′ 1216–1235

256 M.P. Kowalewski et al. / Journal of Steroid Biochemistry & Molecular Biology 101 (2006) 254–262
F onshipp
u39oefaPalc5ittss
tic
sCL
s
2
ig. 1. Amplification of the canine 3�HSD by 3′-5′-RACE PCR. The relatiroducts visualized on a 1.5% ethidium bromide-stained gel (B).
cts of the missing cDNA coding sequences of the 5′- and′-ends were amplified (Fig. 1). After initial denaturation at4 ◦C for 1 min, the reactions were run for 35 cycles, eachne at 94 ◦C for 1 min, annealing was at 65 ◦C for 2 min,longation at 72 ◦C for 3 min, the final extension cycle lastedor 10 min at 72 ◦C. Length of the overlapping RACE PCRmplicons was 710 and 1012 bp for the 3′- and 5′-RACECR, respectively. Finally, using RT-PCR for 40 cycles atn annealing temperature of 57 ◦C and a specific primer pairocated at both ends of the open reading frame (ORF), theomplete ORF of canine 3�HSD was amplified (primers–6). All PCR products were separated on a 1.5% ethid-um bromide-stained gel, purified with Qiaex II gel extrac-
ion system (Qiagen GmbH, Hilden, Germany), ligated intohe pGEM-T vector (Promega, Heidelberg, Germany) andequenced (SRD, Oberursel, Germany) with T7 and SP6equencing primers.[Fc
of gene-specific primers (GSP1, 2) to the cDNA template (A); RACE PCR
Finally, the canine 3�HSD sequence was submitted tohe Genbank (accession number: AY739720: Canis famil-aris 3-�-hydroxysteroid dehydrogenase mRNA, completeds).
Predicted amino acids sequence and sequence compari-on with other mammalian orthologs were performed withhromasPro Version 1.2 oligo software (Technelysium Pty.td.).
The resulting nucleotide- and amino acid sequence ishown in Fig. 2; also indicated are the binding sites of primers.
.3. Real Time PCR
Real Time PCR was performed as previously described20]. For primers and TaqMan probe sequences see Table 1;ig. 2 (primers 9–10, TaqMan probe 11). Length of the ampli-ons was 78 bp.

M.P. Kowalewski et al. / Journal of Steroid Biochemistry & Molecular Biology 101 (2006) 254–262 257
F ence. Ta ame (OA
2
fpc(astpawPM
2
Eti
tetUmttld
2
26aod
ig. 2. Nucleotide sequence of canine 3�HSD and deduced amino acid seqund Real Time (TaqMan) PCR (9, 10, 11) are indicated. The open reading frY739720.
.4. Construction of a 3βHSD cDNA expression vector
The 1247 bp cDNA fragment, generated using RT-PCRor 35 cycles at an annealing temperature of 68 ◦C withrimers 12–13 (Table 1), containing the 1122 bp ORF ofanine 3�HSD flanked by restriction sites for SalI and NotINEB GmbH, Frankfurt am Main, Germany) was digestednd inserted into identical cleaved sites in the gWIZ expres-ion vector (Gene Therapy Systems, San Diego, CA, USA)o yield gWIZ-3�HSD. The structure of the recombinantlasmid was verified by control restriction enzyme analysisnd sequencing (SRD, Oberursel, Germany). Plasmidsere purified before transfection by using the PromegaureYieldTM plasmid purification system (Promega GmbHannheim, Germany).
.5. Transient expression of canine 3βHSD cDNA
HEK293T cells were grown in Dulbecco’s modifiedagle’s medium fortified with 10% fetal calf serum. Before
ransfection, cells were plated at about 100,000/24 mm2 andncubated at 37 ◦C/5% CO2 for 4–5 h. Cells were transiently
(aia
he positions of the primers used for RT-PCR (1, 2, 5, 6), RACE PCR (3, 4)RF) is shown below the nucleotide sequence. Genebank accession number:
ransfected with 1 �g expression vector containing cDNAncoding canine 3�HSD using the GeneJuice® Transfec-ion Reagent (Novagen Merck Biosciences, Nottingham,K). Transfection efficiency (up to 90%) was assessed byonitoring of �-galactosidase activity 24 h after transfec-
ion of gWIZ-�-galactosidase vector (Gene Therapy Sys-ems, San Diego, CA, USA). Non-transfected HEK293T cellines were established not to metabolize pregnenolone, dehy-roepiandrosterone, testosterone or 5�-dihydrotestosterone.
.6. Substrate specificity of 3βHSD
Following the procedures previously described [5,21],4 h after transfection HEK293T cells were incubated overh with radiolabelled substrates to test for the 3�HSDnd a likely 17�HSD-enzyme activity. The conversionsf [3H]-pregnenolone into [3H]-progesterone, of [3H]-ehydroepiandrosterone (DHEA) into [3H]-androstenedione
3�HSD activity) and of [3H]-testosterone into [3H]-ndrostenedione and of [3H]-5�-dihydrotestosterone (DHT)nto [3H]-androstane-3�,17�-diol (17�HSD-activity) weressessed following extraction of 3H-steroids from media
2 chemis
wcaRD
2
h1rayb[ca(scac
2
3tccnmql
sFm(ppabfotsoMt
2
i
mgs3i1tImanbb(ttatp(UrfhG
2
TdKDtI
3
3
r(Tp
ie
g(
58 M.P. Kowalewski et al. / Journal of Steroid Bio
ith six volumes of dichloromethane, subsequent thin-layer-hromatography (TLC) (solvent system chloroform/ethylcetate, 4:1, v/v) and quantification with a BioScanadio-Imager. Tritium-labelled steroids were obtained fromupont-NEN (Stevenage, Herts, UK).
.7. Western blot analysis
Transfected cells or tissue samples from canine CL wereomogenized in a lysis buffer (50 mM Tris, 0.1% SDS,% sodium deoxycholate; Sigma–Aldrich, Irvine, Scotland),esolved on a 10% acrylamide gel and electrobloted ontoImmobilon-P membrane (Sigma–Aldrich). Western anal-
sis was carried out using as primary antibody a rab-it polyclonal antibody against human placental 3�HSD24] at the dilution 1:2000 and horseradish peroxidase-onjugated anti-rabbit IgG at 1:20,000 dilution as secondaryntibody, both in PBS containing 3% (w/v) non-fat milkMarvel Original; Premier Beverages, Stafford, UK). Theignals were visualized using Enhanced ChemiLumines-ence (ECL) kit (Pierce, Rockford, IL, USA). A mousedrenal homogenate preparation was used as a positiveontrol.
.8. In situ hybridization (ISH)
pGEM-T plasmid clone containing the insert for canine�HSD generated with primers 1–2 were digested withhe restriction enzymes NcoI (antisense cRNA) or NotI (senseRNA) (New England Biolabs, Frankfurt, Germany) forRNA synthesis using the cRNA synthesis kit (AMS Biotech-ology, Wiesbaden, Germany) and DIG-RNA labellingix (Boehringer Mannheim). cRNA was determined semi-
uantitatively by dot blot analysis of serial dilutions of DIG-abelled cRNA.
Nonradioactive ISH was performed on paraffin-embeddedections of canine CL. Sections were mounted on Super-rost-Plus slides (Menzel Glaeser, Braunschweig, Ger-any), dewaxed, digested with 70 �g/ml proteinase K
Boehringer, Mannheim, Germany) and postfixed with 4%araformaldehyde and then processed according to therocedure by Lewis and Wells [23] and Klonisch etl. [22]. The DIG-labelled cRNA probes were detectedy alkaline phosphatase-conjugated sheep anti-DIG Fabragments (Boehringer, Mannheim) diluted 1:5000 in 1%vine serum in washing buffer, as recommended byhe supplier. The sections were incubated with the sub-trate 5-bromo-4-chloro-3-indolyl phosphate in the presencef nitroblue tetrazolium (Roche Molecular Biochemicals,annheim) yielding a dark blue precipitate at the site of
he label.
.9. Immunhistochemistry (IHC)
After deparaffinization and rehydration, sections werencubated in citrate buffer (10 nM, pH 6.0) for 15 min in
h1wo
try & Molecular Biology 101 (2006) 254–262
icrowave irradiation in an oven run at 560 W for anti-en retrival. After cooling at room temperature for 20 minections were incubated in 0.3% H2O2 in methanol for0 min to quench endogenous peroxidase and then washedn IHC-buffer/0.3% Triton X pH 7.2–7.4 (0.8 mM Na2HPO4,.47 mM KH2PO4, 2.68 mM KCl, 1.37 mM NaCl). The sec-ions were incubated in a 10% solution of goat serum inHC-buffer in order to block unspecific binding sites. Pri-ary antibody (dilution 1:3000) was a rabbit polyclonal
ntibody against human placental 3�HSD, serum from aon-immunized rabbit served for control. Sections were incu-ated at 4 ◦C overnight, they were then washed in IHC-uffer and incubated with peroxidase-conjugated goat IgGsecondary antibody) against rabbit immunoglobin (Vec-or Laboratories, Burlingame, USA) for 30 min at roomemperature, washed in IHC-buffer and incubated with thevidin–biotin–peroxidase complex (Vectastain ABC kit, Vec-or Laboratories, Burlingame, USA) for 30 min at room tem-erature. After washing they were reacted with the substrateNovaRed substrate kit, Vector Laboratories, Burlingame,SA) according to the manufacturer’s instructions. The
eactions were stopped by rinsing under running tap wateror 5 min. The sections were slightly counterstained withematoxylin and mounted in Histokit (Assistant, Osterode,ermany).
.10. Statistics
Due to the uneven distribution of the data obtained by Realime RT-PCR, results are presented as geometric means witheviation factor (Xg × DF±1). To test for the effect of time theruskal–Wallis Test (a nonparametric ANOVA) followed byunn’s Multiple Comparisons Test was applied with the sta-
istical software program, GraphPad3 (GraphPad Software,nc., San Diego, CA, USA).
. Results
.1. Cloning of cDNA from the CL of four bitches
The whole transcript is composed of a 5′-untranslatedegion (5′-UTR) of 126 nucleotides, an open reading frameORF) of 1122 nucleotides and a 3′-UTR of 441 nucleotides.he deduced canine 3�HSD ORF encodes a 374 amino acidrotein of ∼42 kDa (Figs. 2 and 9).
Canine 3�HSD is organized in four exons. The start codons localized to exon 2 indicating that the ORF is located toxons 2–4 (Fig. 3).
Alignment with the sequence from the complete dogenomic sequence, that has been available on-line since 2004http://www.ncbi.nlm.nih.gov/genome/guide/dog/index.
tml), localized the canine 3�HSD solely to chromosome7. A 100% similarity was found when the cloned 3�HSDas compared with the genomic sequence and no evidencef other 3�HSD isoform was obtained.
M.P. Kowalewski et al. / Journal of Steroid Biochemistry & Molecular Biology 101 (2006) 254–262 259
Fig. 3. Genomic organization of the canine 3�HSD gene. Exact sizes of exons and introns and their locations were obtained from the released data of thecanine genomic sequence (chromosome 17) in Genbank (accession number: NW 876264.1). Coding exons are stippled boxes, noncoding exons are boldedlines, introns are narrow solid lines; NA = not available.
Fig. 4. Amplicons of canine 3�HSD on days 5–65 after ovulation obtainedbN
3
3RE5d
tInes
FPw
a(
3
fidihydrotestosterone (Figs. 8 and 9).
y conventional RT-PCR (GSP1, GSP2): 1–12 = canine luteal samples;= negative control; M = DNA-ladder; d5–d65 = days after ovulation.
.2. Expression of 3βHSD in canine CL during diestrus
With qualitative RT-PCR, expression of mRNA for�HSD could be detected at every stage investigated (Fig. 4).eal Time PCR showed a significant effect of time (p = 0.01).xpression was highest during the early luteal phase on daysand 15 after ovulation. It decreased continuously towards
ay 65 (Fig. 5).Tissue localization of 3�HSD was performed at the pro-
ein level by IHC and at the mRNA-level by ISH. IHC andSH revealed strongest signals on days 5 and 15. Begin-
ing with day 25, signal intensity tended to decrease. How-ver, variability between individual animals was high. Theignals for 3�HSD were evenly distributed within the CLon
Fig. 6. Immunohistochemical localization of 3�HSD in canine
ig. 5. Expression of 3�HSD as determined by Real Time (TaqMan) RT-CR: RGE = relative gene expression (Xg × DF±1); bars with asterisk differith p ≤ 0.05; 5–65 = days after ovulation.
nd localized only in the cytoplasm of the luteal cells onlyFigs. 6 and 7).
.3. Substrate specificity of canine 3βHSD cDNA
Non-transfected cells or cells alternatively trans-ected with �-galactosidase showed no metabolic activ-ty towards pregnenolone, DHEA, testosterone or 5�-
Following transfection HEK293T with canine 3�HSD thenly metabolic activity observed was the conversion of preg-enolone and DHEA into progesterone and androstenedione,
corpora lutea on days 5 and 65 after ovulation (p.o.).
260 M.P. Kowalewski et al. / Journal of Steroid Biochemistry & Molecular Biology 101 (2006) 254–262
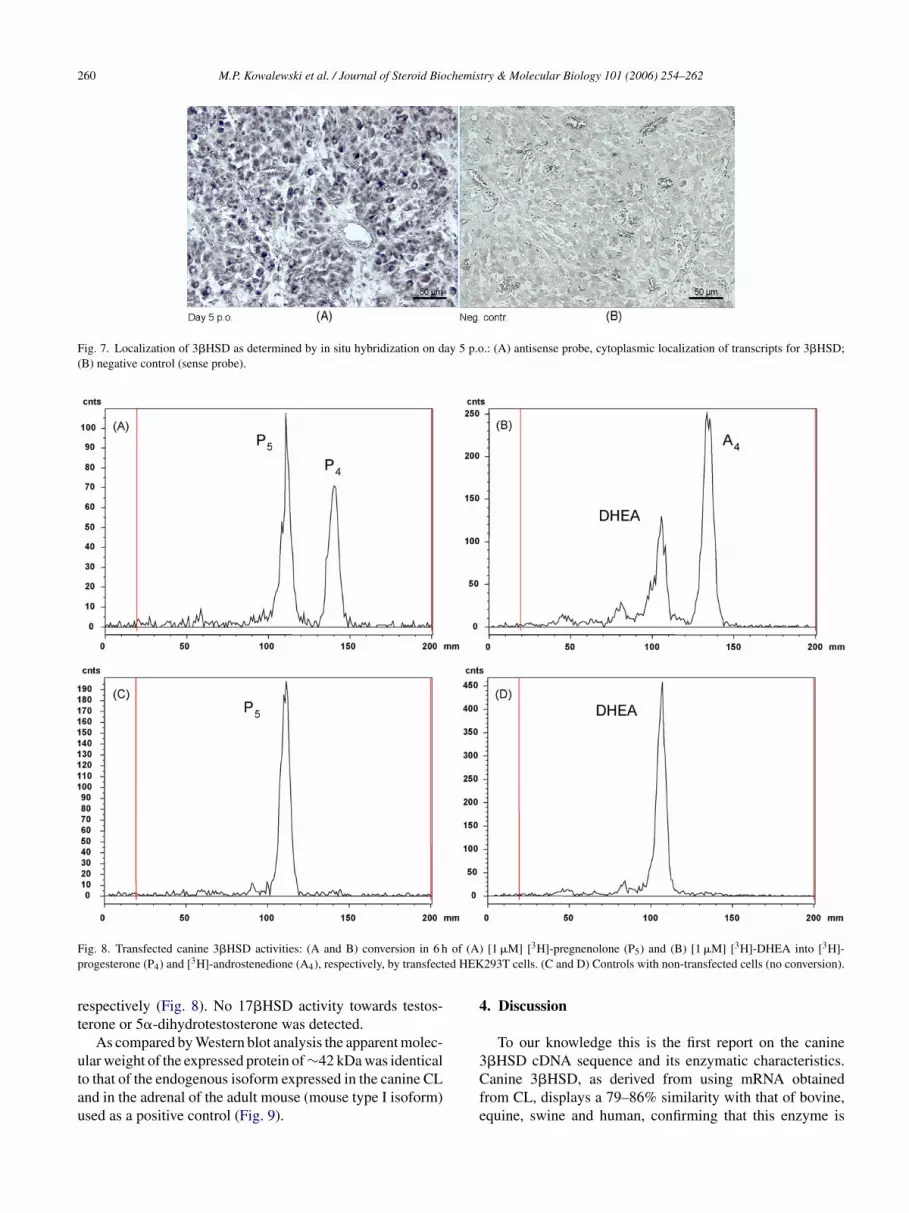
Fig. 7. Localization of 3�HSD as determined by in situ hybridization on day 5 p.o.: (A) antisense probe, cytoplasmic localization of transcripts for 3�HSD;(B) negative control (sense probe).
F of (A)p ed HEK
rt
utau
4
ig. 8. Transfected canine 3�HSD activities: (A and B) conversion in 6 hrogesterone (P4) and [3H]-androstenedione (A4), respectively, by transfect
espectively (Fig. 8). No 17�HSD activity towards testos-erone or 5�-dihydrotestosterone was detected.
As compared by Western blot analysis the apparent molec-
lar weight of the expressed protein of ∼42 kDa was identicalo that of the endogenous isoform expressed in the canine CLnd in the adrenal of the adult mouse (mouse type I isoform)sed as a positive control (Fig. 9).3Cfe
[1 �M] [3H]-pregnenolone (P5) and (B) [1 �M] [3H]-DHEA into [3H]-293T cells. (C and D) Controls with non-transfected cells (no conversion).
. Discussion
To our knowledge this is the first report on the canine
�HSD cDNA sequence and its enzymatic characteristics.anine 3�HSD, as derived from using mRNA obtainedrom CL, displays a 79–86% similarity with that of bovine,quine, swine and human, confirming that this enzyme is

M.P. Kowalewski et al. / Journal of Steroid Biochemis
Fig. 9. Western blot analysis of expressed canine 3�HSD protein in canineCL on day 5 p.o. (A) and in HEK293T cells (B). (A) 1–2 = canineluteal homogenates, P = positive control; mouse adrenal 3�HSD; (B)1HHm
wdaantgrTtemiHs
icta3ltdsaaei
t3
benciadepiaia
adco
A
S
Je
R
= homogenates of non-transfected HEK293T cells, 2 = homogenates ofEK293T cells transfected with canine 3�HSD, P = positive control,EK293T cells transfected with mouse type I 3�HSD, M = molecular weightarker.
ell conserved between species. Several isoforms have beenetected in the mouse, rat, human and in other mammalsnd all of them are products of different genes and sharevery high sequence homology (even greater than 90%);
o alternative spliced products of 3�HSD are known. Inhe present study, we have surveyed by BLAST the dogenome for evidence of other 3�HSD isoforms and noelated sequence for another possible isoform was found.herefore, it may be assumed that the cloned sequence is
he only isoform of 3�HSD in the dog. However, it cannotntirely be ruled out that other, as yet unidentified, isoformsight result from alternative splicing or that if the homology
s very low, not allowing them to be detected by BLAST.owever, in view of the observation of a high conservation
een in other species, this seems rather unlikely.Enzymatic activity of the cloned canine 3�HSD was tested
n an in vitro system using transfected HEK293T cells afteronfirming the identity of the HEK293T cells expressed withhe endogenous, luteal 3�HSD (∼42 kDa) by Western blotnalysis. Non-transfected cells were devoid of endogenous�HSD activity. Pregnenolone and DHEA were metabo-ized to progesterone and androstenedione, respectively, byransfected cells, while no activity with testosterone or 5�-ihydrotestosterone as substrates was observed. Thus, theituation is different from other species such as the mouse
nd rat, where several isoforms have been identified (type Ind -VI in the mouse and rat type I and -IV 3�HSD) alsoxhibiting a cellular 17�HSD-like activity [5–7]. Based onts substrate specificity the canine enzyme exhibited charac-try & Molecular Biology 101 (2006) 254–262 261
eristics very similar to that of human type I and rat type I�HSDs [24–25].
Expression of 3�HSD in the dog CL applying IHC haseen reported for days 40, 50 and 60 of pregnancy [10]. How-ver, so far no data were available on its expression in theon-pregnant dog and in relation to the course of diestrus. Inovering this period we could show that expression of 3�HSDn the CL of the bitch is strongest on both, the protein-(IHC)nd mRNA-levels, at the beginning of diestrus with a constantecrease towards the end of diestrus thereafter. This loss ofnzyme activity resembles the course of P concentration ineripheral plasma and may be indicate that the provision of Ps controlled by the enzyme availability rather than substratevailability. Other than described by Nishiyama et al. [10]n CL from pregnant dogs, in our study and using the samentibody, signals were evenly distributed throughout CL.
The availability of the full-length canine 3�HSD willllow further and more detailed investigations on its tissue-ependent distribution and expression. This particularly con-erns the preovulatory serum P-rise in the dog and the phasef apparent gonadotropin independence.
cknowledgments
Supported by the German Research Foundation Graduateeminar Cell–Cell-Interaction in Reproduction.
Dr. A. Taubert from the Institute of Parasitology at theLU-Giessen is thanked for assistance during Real Time PCRxperiments.
eferences
[1] A.V. Muir, M.B. Hodgins, J.A. Milne, H. Young, The presence ofhydroxysteroid dehydrogenases in mammalian sebaceous structures,Br. J. Dermatol. 83 (1970) 291–295.
[2] M.B. Simpson, W.J. Cunliffe, M.B. Hodgins, The relationship betweenthe in vitro activity of 3�-hydroxysteroid dehydrogenase, delta-5-isomerase in human sebaceous glands and their secretory activity invivo, J. Invest. Dermatol. 81 (1983) 139–144.
[3] M.E. Sawaya, N.S. Penneys, Immunohistochemical distribution of aro-matase and 3B-hydroxysteroid dehydrogenase in human hair follicleand sebaceous gland, J. Cutan. Pathol. 19 (4) (1992) 309–314.
[4] J. Simard, J. Couet, F. Durocher, Y. Labrie, R. Sanchez, N. Breton, C.Turgeon, F. Labrie, Structure and tissue-specific expression of a novelmember of the rat 3�-hydroxysteroid dehydrogenase/�5-�4 isomerase(3�HSD) family-the exclusive 3�HSD gene expressed in the skin, J.Biol. Chem. 268 (1993) 19659–19668.
[5] J.I. Mason, B.E.C. Howe, A.F. Howie, S.D. Morley, M.R. Nicol,A.H. Payne, Promiscuous 3�-hydroxysteroid dehydrogenases: testos-terone 17�-hydroxysteroid dehydrogenase activities of mouse type Iand VI 3�-hydroxysteroid dehydrogenases, Endocr. Res. 30 (4) (2004)709–714.
[6] Y. de Launoit, J. Simard, F. Durocher, F. Labrie, Androgenic 17 beta-
hydroxysteroid dehydrogenase activity of expressed rat type I 3 beta-hydroxysteroid dehydogenase/delta 5-delta 4 isomerase, Endocrinol-ogy 130 (1) (1992) 553–555.[7] R. Sanchez, Y. de Launoit, F. Durocher, A. Belanger, F. Labrie, J.Simard, Formation and degradation of dihydrotestosterone by recom-

2 chemis
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
62 M.P. Kowalewski et al. / Journal of Steroid Bio
binant members of the rat 3 beta-hydrpxysteroid dehydrogenase/delta5-delta 4 isomerase family, Mol. Cell. Endocrinol. 103 (1–2) (1994)29–38.
[8] I.G. Abbaszade, J. Arensburg, C.H. Park, J.Z. Kasa-Vubu, J. Orly, A.H.Payne, Isolation of a new mouse 3�-hydroxysteroid dehydrogenaseisoform, 3�-HSD VI, expressed during early pregnancy, Endocrinology138 (4) (1987) 1392–1399.
[9] P.A. Bain, M. Yoo, T. Clarke, S.H. Hammond, A. Payne, Multiple formsof mouse 3�-hydroxysteroid dehydrogenase/�5–�4 isomerase and dif-ferential expression in gonads, adrenal glands, liver, and kidneys of bothsexes, Proc. Natl. Acad. Sci. 88 (1991) 8870–8874.
10] T. Nishiyama, S. Tsumagari, M. Ito, G. Watanabe, K. Taya, M. Takeishi,Immunohistochemical study of steroideogenic enzymes in the ovaryand placenta during pregnancy in the dog, Anat. Histol. Embryol. 28(1999) 125–129.
11] P.W. Concannon, J.P. McCann, M. Temple, Biology end endocrinologyof ovulation, pregnancy and parturition in the dog, J. Reprod. Fertil. 39(Suppl.) (1989) 3–25.
12] P.W. Concannon, W.R. Butler, W. Hansel, P.J. Knight, J.M. Hamilton,Parturition and lactation in the bitch: Serum progesterone, cortisol andprolactin, Biol. Reprod. 19 (1978) 1113–1118.
13] B.G. Steinetz, L.T. Goldsmith, H.J. Harvey, G. Lust, Serum relaxinand progesterone concentrations in pregnant, pseudopregnant, andovariohysterectomized, progestin-treated pregnant bitches: detectionof relaxin as a marker of pregnancy, Am. J. Vet. Res. 50 (1989) 68.
14] B. Hoffmann, F. Busges, E. Engel, M.P. Kowalewski, P. Papa, Regula-tion of corpus luteum-function in the bitch, Reprod. Domest. Anim. 39(4) (2004) 2342–3240.
15] P.N. Olson, Periparturient diseases of the bitch, Proc. Soc. Theriogenol.89 (1989).
16] E.C. Feldman, R.W. Nelson, Periparturient diseases, in: Canine andFeline Endocrinology and Reproduction, third ed., Saunders, ElsevierScience, USA, 2004, p. 811.
[
try & Molecular Biology 101 (2006) 254–262
17] P.W. Concannon, Effects of hypophysectomy and of LH administra-tion on luteal phase plasma progesterone levels in the beagle bitch, J.Reprod. Fertil. 58 (1980) 407–410.
18] P.W. Concannon, P. Weinstein, S. Whaley, D. Frank, Suppressionof luteal function in dogs by luteinizing hormone antiserum and bybromocriptine, J. Reprod. Fertil. 81 (1987) 175–180.
19] A.C. Okkens, D.J. Dieleman, M.M. Bevers, A.A.M.E. Lubberink, A.H.Willemse, Influence of hypophysectomy on life span of the corpusluteum in the cyclic dog, J. Reprod. Fertil. 77 (1986) 187–192.
20] M.P. Kowalewski, G. Schuler, A. Taubert, E. Engel, B. Hoffmann,Expression of cyclooxygenase 1 and 2 in the canine corpus luteumin diestrus, Theriogenology 66 (6–7) (2006) 1423–1430.
21] T. Suzuki, H. Sasano, S. Andersson, I.J. Mason, 3�-Hydroxysteroiddehydrogenase/�5-4-isomerase activity associated with the human17�-hydroxysteroid dehydrogenase type 2 isoform, J. Clin. Endocrinol.Metab. 85 (2000) 3669–3673.
22] T. Klonisch, S. Hombach-Klonisch, C. Froehlich, J. Kauffold, K.Steger, B.G. Steinetz, B. Fischer, Canine preprorelaxin: nucleic acidsequence and localization within the canine placenta, Biol. Reprod. 60(1999) 551–557.
23] F.A. Lewis, M. Wells, Detection of virus in infected human tissue by insitu hybridization, in: D.G. Wilkinson (Ed.), In Situ Hybridization—APractical Approach, Oxford University Press, Oxford, 1992, pp.121–135.
24] M.W. Lorence, B.A. Murry, J.M. Trant, J.I. Mason, Human 3�-hydroxysteroid dehydrogenase/�5 → 4 isomerase from placenta:expression in nonsteroidogenic cells of a protein that catalyzes the dehy-drogenation/isomerization of C21 and C19 steroids, Endocrinology 126
(5) (1990) 2493–2498.25] M.W. Lorence, D. Naville, S.E. Graham-Lorence, S.O. Mack, B.A.Murry, J.M. Trant, J.I. Mason, 3�-hydroxysteroid dehydrogenase/delta5 → 4-isomerase expression in rat and characterization of the testisisoform, Mol. Cell. Endocrinol. 80 (1–3) (1991) 21–31.