characterization of cdnas encoding isoforms of hamster 3 beta-hydroxysteroid dehydrogenase/delta...
TRANSCRIPT

Characterization of cDNAs encoding isoforms of hamster3â-hydroxysteroid dehydrogenase/Ä5]4isomerase
F M Rogerson, J Courtemanche1, A Fleury1, J R Head, J-G LeHoux1
and J I MasonCecil H and Ida Green Center for Reproductive Biology Sciences, University of Texas SouthwesternMedical Center, Dallas, Texas 75235, USA
1Department of Biochemistry, University of Sherbrooke, Sherbrooke, Quebec, Canada
(Requests for offprints should be addressed to F M Rogerson, Prince Henry’s Institute of MedicalResearch, PO Box 5152, Clayton, Victoria 3168, Australia)
(F M Rogerson is now at Prince Henry’s Institute of Medical Research, Monash Medical Centre,Melbourne 3168, Australia)
(J I Mason is now at Department of Clinical Biochemistry, University of Edinburgh, EdinburghEH3 9YW, UK)
ABSTRACT
Western blot analyses of various hamster tissuesreveal high levels of expression of 3â-hydroxysteroid dehydrogenase (3â-HSD) in adrenaland liver, and moderate levels of expression inkidney. The expression in liver is sexually dimor-phic; very high levels of protein are observed inadult male liver but very low levels are seen in thefemale liver. Three distinct cDNAs encodingisoforms of 3â-HSD were isolated from hamstercDNA libraries. The type 1 isoform is a high-affinity dehydrogenase/isomerase expressed in adre-nal and male kidney. The type 2 isoform is also ahigh-affinity dehydrogenase/isomerase expressed in
kidney and male liver. The type 3 enzyme is a3-ketosteroid reductase expressed predominantly inkidney. Sequencing of the clones showed that allthree are structurally very similar, although types 1and 2 share the greatest degree of similarity.Immunohistochemical staining for 3â-HSD in theadrenal was found throughout the adrenal cortex. Inthe kidney staining was confined to tubules, and inthe liver, heavy staining was found in hepatocytes.The cloning of cDNAs for 3â-HSD from the liverand kidney should help in elucidating the functionof this enzyme in these tissues.Journal of Molecular Endocrinology (1998) 20, 99–110
INTRODUCTION
The enzyme 3â-hydroxysteroid dehydrogenase/Ä5]4isomerase (3â-HSD) is essential for theproduction of all classes of steroid hormones. It isexpressed in the adrenal cortex where it is requiredfor the synthesis of mineralocorticoids and gluco-corticoids, in the gonads where it is required for theproduction of androgens and estrogens, and in theplacenta for the production of progesterone andestrogens. In both the human (Luu-The et al. 1989,Lorence et al. 1990, Lachance et al. 1991) and rat(Lorence et al. 1991, Zhao et al. 1991, Simard et al.1993) a different isoform of 3â-HSD is expressed inthe placenta than is expressed in the adrenal andgonads. Other isoforms of 3â-HSD have beencloned and characterized from the liver and kidney
of the rat and mouse. In rat liver, two isoforms of3â-HSD have been identified; a low affinitydehydrogenase (Zhao et al. 1990, Naville et al. 1991,Simard et al. 1991) and a structurally related3-ketosteroid reductase that possesses no dehydro-genase activity (de Launoit et al. 1992a). In mouseliver both a high (Clarke et al. 1993a) and low(Keeney et al. 1993a) affinity dehydrogenase havebeen described, as well as a 3-ketosteroid reductase(Abbaszade et al. 1995). In mouse kidney, a second3-ketosteroid reductase has also been identified(Clarke et al. 1993b). Some isoforms of 3â-HSD inthe liver and kidney are expressed in a sexuallydimorphic manner (Naville et al. 1991, Couet et al.1992, Clarke et al. 1993b, Keeney et al. 1993a,b,Abbaszade et al. 1995). In both the rat (Naville et al.1991, Couet et al. 1992) and the mouse (Keeney
99
Journal of Molecular Endocrinology (1998) 20, 99–110 ? 1998 Journal of Endocrinology Ltd Printed in Great Britain0952–5041/98/020–099 $08.00/0

et al. 1993b) this sexual dimorphism has beendemonstrated to be dependent on differences in thepattern of growth hormone release between malesand females.The functions of 3â-HSD in the liver and kidney
remain unclear. Experiments in the hamster suggestthat it is a good model for the study of theregulation of steroidogenic enzymes (LeHoux et al.1992). In this study, we have isolated andcharacterized cDNAs for 3â-HSD in the hamsteradrenal, liver and kidney.
MATERIALS AND METHODS
Western blot analysis
Western blotting was performed as describedpreviously (Naville et al. 1991). Briefly, tissuewas homogenized in lyzing buffer containing 1%sodium cholate and 0·1% sodium dodecyl sulfateto solubilize membrane-bound protein. One-dimensional electrophoresis was performed in an8% polyacrylamide gel. Protein was electroblottedonto Immobilon-P nylon membranes (Millipore,Bedford, MA, USA) for 1 h at 100 V. The filterswere probed with a polyclonal antibody (5 µgIgG/ml) raised against human placental 3â-HSD.After washing, the bound antibody was detected byincubation with approximately 10 000 c.p.m./ml125I-Protein A (Dupont NEN, Boston, MA, USA)and exposing the filter to X-ray film.
Screening of cDNA libraries
ë-ZAP cDNA libraries were synthesized byStratagene Inc. (La Jolla, CA, USA) from RNAisolated from adult male hamster adrenal, kidneyand liver. The libraries were screened with afull-length cDNA encoding human type 1 3â-HSDlabeled with [á32P]dCTP by the random primermethod. Positive clones were isolated and thepBluescript plasmid, containing the cDNA ofinterest, was excised from the phagmid according tothe manufacturer’s instructions and transformed inSOLR competent E. coli cells. Both strands of DNAwere sequenced by the dideoxy chain terminationmethod (Sanger et al. 1977) using Sequenase II(United States Biochemicals, Cleveland, OH, USA)and [á35S]dATP.
Construction of 3â-HSD expression vectorsand transfection into 293 human fetal kidneycells
Types 1 and 3 hamster 3â-HSD were expressed inthe pcDNA3 vector (Invitrogen, San Diego, CA,
USA). XhoI and NotI digests were made to excisethe full-length 3â-HSD clones from the pBluescriptvector. The clones were purified from an agarose gelby the freeze–squeeze method (Tautz & Renz 1983)and ligated into the pcDNA3 vector digested thesame way. Hamster 3â-HSD type 2 constructsmade in this way possessed no activity in transienttransfection assays. It was decided to PCR amplifythe coding region of type 2 using the upstreamprimer 5*-GCCATGCCTGGATGGAGC-3*, cor-responding to nucleotides "3 to +15, and thedownstream primer 5*-TCACATTCACTGAGCCTTTGT-3*, complementary to the 3* end of thecoding region, +1108–+1128. This PCR productwas ligated directly into the expression vector pCR3(Invitrogen), and sequenced to ensure there was nomis-incorporation of nucleotides into the sequenceduring amplification.Human embryonic kidney 293 cells were grown
in Dulbecco’s modified Eagle’s high glucosemedium (Gibco-BRL, Gaithersburg, MD, USA)supplemented with Hepes (10 mM), calf bovineserum (10%) and antibiotics/antimycotics. Thecells were transfected using Lipofectin reagent(Gibco-BRL) according to the manufacturer’sprotocol. Geneticin (0·5 mg/ml) was added tothe culture medium 48 h after transfection toselect for cells expressing the neomycin resistancegene.Expression of the 3â-HSD isoforms in these cells
was confirmed by Western blotting as describedabove using a polyclonal antibody raised againsthuman type 1 3â-HSD.
Measurement of 3â-HSD activity
The kinetics of the conversion of pregnenoloneto progesterone, dehydroepiandrosterone (DHEA)to androstenedione, and 5á-dihydrotestosterone(5á-DHT) to 5á-androstanediol (5á-Adiol) weremeasured in homogenates prepared from trans-fected cells. The cells were removed from theculture dishes and sonicated in assay buffer (12 mMTris, pH 7·5 containing 1 mM EDTA and 0·25 Msucrose). The activity assays were performed withvarious concentrations of substrate including30 000 c.p.m./µl tritiated substrate (Dupont NEN).For measurement of the dehydrogenase/isomeraseactivity 1 mM NAD+ was added to the assay. Formeasurement of the reductase activity 1 mMNADH or NADPH was added to the assay.Reactions were initiated by the addition ofhomogenate, run for 10 min at 37 )C and terminatedby the addition of 12 vol methylene chloride. In allcases the amount of homogenate was adjusted sothat the conversion of substrate to product did not
and others · 3â-HSD isoforms in the hamster100
Journal of Molecular Endocrinology (1998) 20, 99–110

exceed 20%. The reactions were analyzed by thinlayer chromatography on silica plates. For theseparation of pregnenolone and progesterone, andDHEA and androstenedione, a solvent system ofchloroform:ethyl acetate (vol/vol, 4:1) was used. Forthe separation of 5á-DHT and 5á-Adiol, a solventsystem of chloroform:ethyl acetate (vol/vol, 9:1) wasused. The plates were scanned and quantified usinga radioanalyzer (BioScan, Washington DC, USA)and the results expressed as pmol product formed/min/mg protein. Double reciprocal plots weregenerated to obtain the apparent Km values for thevarious substrates.
Tissue distribution by reverse transcriptase-polymerase chain reaction (RT-PCR) analysisof mRNA transcripts
Total RNA was extracted using the Tri Reagentprotocol (Molecular Research Center, Cincinnati,OH, USA). Thirty micrograms RNA were treatedwith 15 U DNase I (Pharmacia, Uppsala, Sweden)for 30 min at 37 )C then phenol–chloroformextracted followed by ethanol precipitation withammonium acetate. A total of 5 µg of treated RNAwas used for RT using 200 U SuperScript II(Gibco BRL) for 50 min at 42 )C in the presence of0·5 mg oligo(dT)12–18 in a total volume of 20 µl. Theresulting cDNA was denatured for 15 min at 70 )Cthen treated with 1·2 U RNase H (Pharmacia) for20 min at 37 )C.Two microlitres of the above reaction were used
for the PCR using 2·5 U Taq DNA polymerase(Pharmacia) in a total volume of 100 µl. Todetermine the expression of types 1 and 3 thefollowing conditions were used: 94 )C 1 min, 65 )C1·5 min, 72 )C 1·5 min for 30 cycles. To determinethe expression of type 2 the following conditionswere used: 94 )C 1 min, 60 )C 1·5 min, 72 )C1·5 min for 30 cycles. The upstream primers for thePCR were all derived from the variable"45 to"70region of the 5* untranslated region. For type 1 theprimer sequence was 5*-GCAGCCCTTCCTAGGGTCAC-3*, for type 2 the primer sequence was5*-ATGCCATCTCCAGAATCA-3* and for type 3the primer sequence was 5*-CTGCCTTCCCCGGAGTGACA-3*. The downstream primer fortypes 1 and 3 was 5*-TGCCAGCACTGCCTTCTC-3*, corresponding to nucleotides +485–+502.The downstream primer for type 2 was 5*-TGTCATCTGAGATGTAGT-3* correspondingto nucleotides +761–+778.Ten microlitres of the above reactions were
electrophoresed on 1% agarose gel and transferredonto Zeta-Probe blotting membranes (Bio-Rad,Hercules, CA, USA) using 0·4 M NaOH. The
Southern blots were probed using the same human3â-HSD type 1 cDNA, labeled with [á32P]dCTP,as was used to isolate the three 3â-HSD clones fromthe cDNA libraries.
Immunohistochemistry
Hamster tissue was removed, cut into pieces,immediately fixed in 10% neutral buffered formalin,embedded in paraffin and 5 µm sections cut.After deparaffinization the sections were treatedwith normal blocking serum then incubated witha polyclonal antibody raised against purifiedhuman placental 3â-HSD at a dilution of1:1000. The sections were then incubated with abiotinylated goat anti-rabbit IgG followed bybiotin-linked avidin (Histostain kit for rabbitprimary antibodies, Zymed, San Francisco, CA,USA). Prior to incubation with streptavidin–peroxidase the sections were treated with 0·5%H2O2 to block endogenous peroxidase activity.Antibody binding was visualized by incubation withdiaminobenzidine. As a negative control, theantibody was pre-absorbed with membranes frombacteria over-expressing the human type 1 3â-HSDenzyme.
1. Western blot analysis of the expression of3â-HSD in male hamster adrenal, kidney and testis, andmale (M) and female (F) hamster liver. The Westernblot was probed with an antibody raised against humanplacental 3â-HSD.
3â-HSD isoforms in the hamster · and others 101
Journal of Molecular Endocrinology (1998) 20, 99–110

RESULTS
Western blot analysis
The results of the Western blot analysis are shownin Fig. 1. A high level of 3â-HSD protein is presentin the adrenal, a moderate level in the kidney andlow levels in the gonad. A very high expression ofprotein was observed in the male liver comparedwith almost undetectable levels in the female liver.The level of 3â-HSD protein in male liver is similarto that in the adrenal.
Screening of cDNA libraries
The nucleotide and deduced amino acid sequencesof the various clones are shown in Figs 2 and 3respectively. From the adrenal library, nine posi-tive clones were isolated, five of which containedthe entire coding region, as compared with thesequences of 3â-HSD from other species. All clonesgave identical sequence. This clone was designatedthe type 1 enzyme. The longest clone contains 75nucleotides of 5* untranslated sequence, an openreading frame of 1122 nucleotides, and 386nucleotides of 3* untranslated sequence. The cDNAcodes for a protein of 373 amino acids, with acalculated molecular weight of 41 753. From thekidney library, five positive clones were isolated,four of which contained the entire coding region.All clones gave identical sequence, but it wasdistinct from the adrenal sequence and this clonewas designated type 2. The longest clone contains253 nucleotides of 5* untranslated sequence, an openreading frame of 1122 nucleotides, and 370nucleotides of 3* untranslated sequence. This clonealso codes for a protein of 373 amino acids, with acalculated molecular weight of 41 903. From theliver library, 13 positive clones were isolated.Twelve of these had identical sequence to type 2.The remaining clone has sequence distinct fromboth types 1 and 2 (Fig. 3) and was designatedas type 3. This clone contains 279 nucleotides of5* untranslated sequence, an open reading frameof 1122 nucleotides, and 339 nucleotides of 3*untranslated sequence. This isoform is also 373amino acids long, with a molecular weight of 41 805.At the amino acid level, there is 95·5% sequencesimilarity between types 1 and 2, 90·4% similaritybetween types 1 and 3 and 88·2% similarity betweentypes 2 and 3.
Measurement of 3â-HSD activity
The Km values for the substrates pregnenolone,DHEA and 5á-DHT are given in Table 1. The Kmvalues calculated for type 1 and type 2 demonstrate
that they are both high affinity dehydrogenase/isomerases that are also able to function as 3-ketosteroid reductases when NADH is used ascofactor. No reductase activity was observed in thepresence of NADPH (results not shown).In contrast, type 3 possesses no dehydrogenase/
isomerase activity and acts exclusively as a 3-ketosteroid reductase when NADPH is used ascofactor. No reductase activity was observed in thepresence of NADH (results not shown).Overexpression of each of the three isoforms in
293 cells yielded products of 44 kDa that weredetectable by Western blotting (results not shown).
Tissue distribution by RT-PCR analysis ofmRNA transcripts
Figure 4 shows the results of the RT-PCR screen.Amplification products of the expected sizes, thathybridized to the 32P-labeled human type 1 cDNA,were observed for all three isoforms. Also, in theanalysis of the type 1 isoform a PCR product of thewrong molecular size, approximately 1 kb, wasobserved in the male liver. This product was notfurther characterized. Type 1 mRNA transcriptswere found in very high levels in both the male andfemale adrenal. A low level of expression was alsoobserved in the male kidney but not in the femalekidney. No transcripts were observed in the liver.High levels of type 2 mRNA transcripts wereobserved in the kidney; there appeared to be somesexual dimorphism as expression in the male kidneywas greater than in the female. Also, a high level ofexpression was observed in the male liver but notranscripts were observed in the female liver.Moderate levels of type 3 transcripts were observedin both the male and female kidney with a very lowlevel of expression observed in the male liver.
Immunohistochemical studies
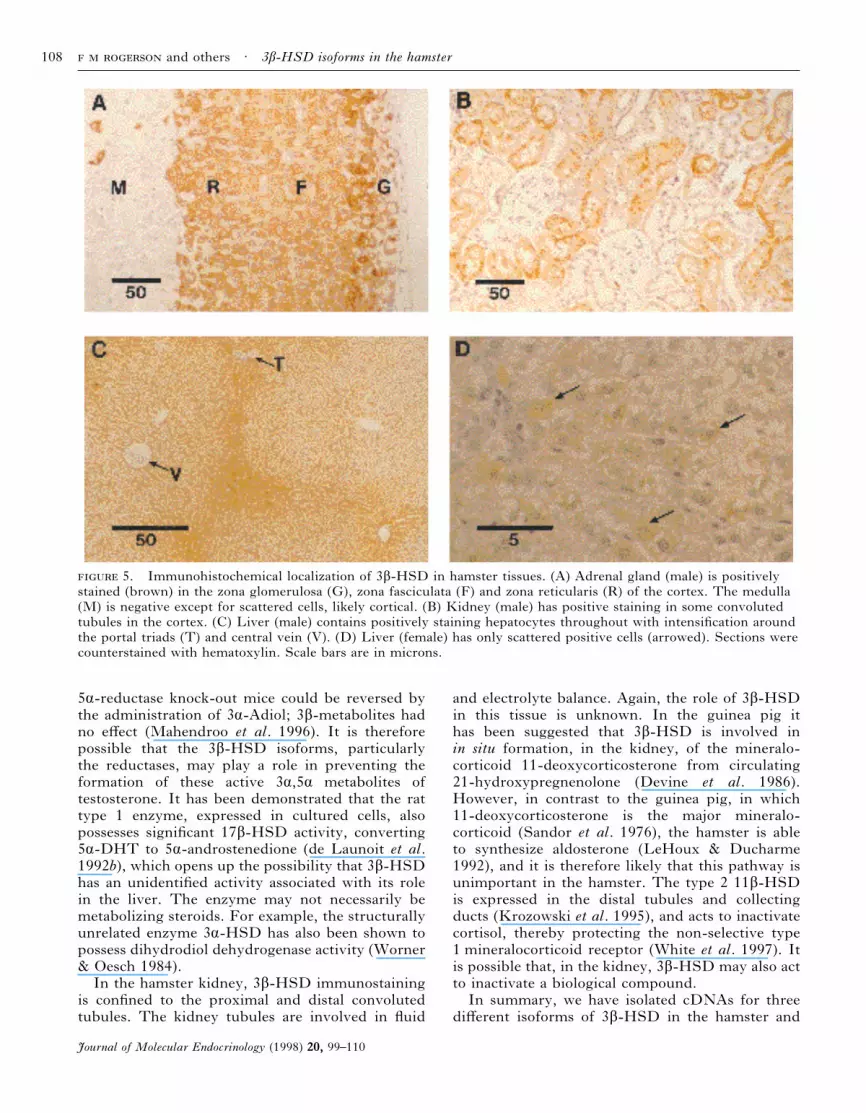
It was established that, when expressed in 293 cells,all three hamster 3â-HSD isoforms cross-reactedwith the anti-human type 1 3â-HSD polyclonalantibody used in this study (see above). In theadrenal gland intense staining was observed for3â-HSD throughout the adrenal cortex (Fig. 5A).The medulla was negative except for scatteredpositive cells that are presumed to be cortical cellstrapped during development. In the kidney (Fig.5B) staining was found predominantly in both thedistal and proximal tubules of the cortex, althoughnot all were positive and there was often hetero-geneity of staining in individual tubule profiles. Inthe male liver (Fig. 5C) positive staining wasobserved in hepatocytes throughout the tissue.
and others · 3â-HSD isoforms in the hamster102
Journal of Molecular Endocrinology (1998) 20, 99–110

Intensification of the staining, where seen, wasusually around portal triads. Only scattered positivecells were seen in the female liver (Fig. 5D). Nostaining was observed when the antibody waspre-absorbed with membranes from bacteria over-expressing the human type 1 3â-HSD enzyme(results not shown).
DISCUSSION
We report the cloning and characterization of threedifferent forms of 3â-HSD in the hamster. The type1 enzyme is a high affinity dehydrogenase/isomeraseexpressed at very high levels in both male andfemale adrenal and at low levels in male kidney. Thetype 2 enzyme is also a high affinity dehydrogenase/isomerase but is expressed at high levels in male andfemale kidney and in male liver. There is a veryhigh degree of sequence similarity between the type1 and 2 enzymes and this is paralleled by verysimilar Km values for pregnenolone, DHEA, and5á-DHT. Despite these similarities, the twoisoforms are expressed in different tissues, suggest-ing that they are the products of two genes underseparate regulatory control. Consistent with otherforms of 3â-HSD in other species, the dehydrogen-ase activity of hamster types 1 and 2 requiresNAD+, and the reductase activity requires NADH.The type 3 enzyme of the hamster is a 3-ketosteroidreductase expressed predominantly in the male andfemale kidney with very low levels of expressionin the male liver. No dehydrogenase activity wasobserved using either pregnenolone or DHEAas substrates. Comparisons of the amino acidsequences reveal that types 1 and 2 are more closelyrelated to each other than to type 3, reflecting thedifferences in enzymatic activity.The situation in the hamster is therefore similar
to other rodent species. The hamster type 13â-HSD is analogous to the high affinity type 1isoforms of 3â-HSD expressed in the adrenals andgonads in both the rat (Zhao et al. 1990, Lorenceet al. 1991, Simard et al. 1991) and mouse (Bainet al. 1991, Clarke et al. 1993a). The type 2 isoformis also a high affinity dehydrogenase but ispredominantly expressed in kidney and male liver.The tissue expression does not completely parallelany mouse 3â-HSD species but is most similar tomouse type 3 which is expressed in liver and kidneyof both sexes (Bain et al. 1991). The hamster type 2also has similar enzyme kinetics to mouse type 3although nucleotide sequence comparisons betweenhamster and mouse 3â-HSD isoforms reveal nostriking correlations (results not shown). Thehamster type 3 enzyme is a 3-ketosteroid reductase
enzyme that is expressed predominantly in kidney.Nucleotide sequence comparisons between hamsterand mouse 3â-HSD isoforms reveal nothingobvious (results not shown) but comparisons ofenzymatic activity and tissue distribution suggestthat hamster type 3 3â-HSD is most closely relatedto mouse type 4 (Clarke et al. 1993b).The type 3 enzyme requires NADPH as cofactor
for the reduction of 5á-DHT, a feature sharedby the other 3-ketosteroid reductase isoforms of3â-HSD (de Launoit et al. 1992a, Clarke et al.1993b, Abbaszade et al. 1995). A comparison ofall the known sequences of 3â-HSD reveals thatan aspartate residue is present at position 36 of allthe NAD/NADH preferring enzymes, whereasthe corresponding position of all the known3-ketosteroid reductases is either a phenylalanine ora tyrosine. It has been previously demonstrated thataspartic acid 38 determines the cofactor specificityof Drosophila alcohol dehydrogenase (Chen et al.1991). The sequences of all three hamster isoformsshare an N-terminal Gly–X–X–Gly–X–X–Glymotif, common to all known 3â-HSD enzymes, thatis thought to form part of a âáâ fold involved incofactor binding (Simard et al. 1996). From thestudy of patients with 3â-HSD deficiency a numberof amino acids have been identified that are criticalfor the function of the human type 2 enzyme,specifically Gly15, Ala82, Asn100, Leu108, Gly129,Glu142, Leu173, Tyr253 and Tyr254 (Simard et al.1996). All of these residues are present in the threehamster isoforms (bearing in mind that human type2 3â-HSD is truncated by one amino acid at theN-terminus). Glycine15 is probably involved incofactor binding. The other residues are presum-ably involved in either catalysis or substratebinding. Another critical residue in human type2, Pro186, is conserved amongst all knowndehydrogenase/isomerase isoforms but is not con-served in the reductase isoforms (Simard et al.1996). In both hamster types 1 and 2 3â-HSD thisproline is conserved but in the type 3 isoform aleucine is found in this position. Affinity labeling ofhuman type 1 3â-HSD, using radiolabeled sub-strate, has identified two residues, Cys183 andHis262, which were concluded to be involved insubstrate binding (Thomas et al. 1993). Both ofthese residues are present in all three hamsterisoforms. Within the sequences of known 3â-HSDisoforms are two Tyr–X–X–X–Lys motifs which areinvolved in the active site of short-chain alcoholdehydrogenases (Simard et al. 1996). Both of thesemotifs are present in hamster types 1 and 3, betweenamino acids 155–159 and 270–274. The type 2isoform, however, only contains the N-terminal ofthese motifs; in the C-terminal sequence the
3â-HSD isoforms in the hamster · and others 103
Journal of Molecular Endocrinology (1998) 20, 99–110

and others · 3â-HSD isoforms in the hamster104
Journal of Molecular Endocrinology (1998) 20, 99–110

2. (opposite and above). Nucleotide sequence comparisonsof the three 3â-HSD cDNA clones isolated from cDNA librariesderived from hamster adrenal, liver and kidney. The ATG startcodon of translation, the stop codon of translation (TGA) and thepolyadenylation signals are all underlined. Sequence identity isdenoted by an asterisk (*) and gaps in the sequence, placed tooptimize the sequence alignment, are denoted by a period (.).
3â-HSD isoforms in the hamster · and others 105
Journal of Molecular Endocrinology (1998) 20, 99–110

tyrosine has been altered to asparagine, suggestingthat this motif is not involved in catalysis.There is a striking sexual dimorphism in the
expression of 3â-HSD in the hamster liver. Westernblot analysis was very consistent with our previousNorthern blot analysis (Rogerson et al. 1995) andshows that isoforms of this enzyme are present invery high amounts in the male liver but that there is
very little expression in the female liver. We wereunable to detect either dehydrogenase/isomerase orreductase activity in homogenates of female liverunder conditions where both activities were observ-able in male liver (results not shown). Consistentwith these observations hamster types 2 and 33â-HSD were found by RT-PCR analyses in malebut not female liver. This is very similar to thesituation in the rat, in which both the dehydro-genase and 3-ketosteroid reductase enzymes areexpressed in much higher levels in the male liver(Naville et al. 1991, Couet et al. 1992). Bothisoforms in the rat liver appear to be regulateddirectly by the pattern of growth hormone (GH)release, and indirectly by testosterone and estrogenacting to change this pattern (Naville et al. 1991,Couet et al. 1992). A male-type pattern of GHrelease will induce expression of both enzymes,whereas a female-type pattern down-regulates bothenzymes (Naville et al. 1991, Couet et al. 1992).Sexually dimorphic expression in hamster tissues isnot confined to the liver and is also observed in thekidney; type 1 3â-HSD is expressed in male but notfemale kidney and type 2 3â-HSD is expressed at
1. Km measurements of pregnenolone,dehydroepiandrosterone (DHEA), and 5á-dihydro-testosterone (5á-DHT) for the three hamster 3â-HSDcDNAs expressed in 293 human fetal kidney cell line.Activity assays were performed in triplicate usinghomogenates prepared from transfected cells. Values aremeans&..
Km (ìM)
Type 1 Type 2 Type 3
Pregnenolone 0·29&0·09 0·13&0·03 n.d.DHEA 0·04&0·01 0·03&0·004 n.d.5á-DHT 0·90&0·06 0·66&0·13 0·41&0·09
n.d.=not detectable.
3. Deduced amino acid sequences of the three 3â-HSD cDNAclones isolated from cDNA libraries derived from hamster adrenal, liver andkidney. Sequence identity is denoted by an asterisk (*).
and others · 3â-HSD isoforms in the hamster106
Journal of Molecular Endocrinology (1998) 20, 99–110

higher levels in the male kidney. In the mouse,male-specific expression of isoforms of 3â-HSD inthe liver, kidney and gonad has been demonstratedby Western blotting (Keeney et al. 1993a), andagain it has been shown that this is influenced byGH (Keeney et al. 1993b).Western blot analysis also shows that there is a
very high level of 3â-HSD in the male liver,equivalent to that in the adrenal. Both the type 2and 3 hamster 3â-HSD isoforms isolated in thisstudy are recognized by the polyclonal antibodyraised against human type 1 3â-HSD. TheRT-PCR analysis of tissue expression, however,suggests that the levels of expression of the types 2and 3 enzymes would be insufficient to account forall the 3â-HSD-like immunoreactive protein. Thereis likely to be at least one more 3â-HSD iso-form expressed in the liver, probably a second3-ketosteroid reductase that has been described inboth rat (de Launoit et al. 1992a) and mouse(Abbaszade et al. 1995) liver. It is probable thatthere are at least six distinct 3â-HSD isoforms.In the mouse five species have been identified inthe adrenal, gonad, liver and kidney (Payne et al.1997), three of which are reversible dehydrogenasesand two of which are non-reversible reductases.Very recently a sixth isoform, a dehydrogenase, hasbeen identified in the mouse placenta and skin(Abbaszade et al. 1997). It is also probable, then,that in the hamster there are at least three moreisoforms of 3â-HSD yet to be discovered.In the adrenal, immunohistochemistry reveals that
3â-HSD is present in all three levels of the cortex,
consistent with the role of this enzyme in steroido-genesis. Of the three isoforms isolated in this studyonly the type 1 enzyme is expressed in the adrenal. Itis interesting that this isoform has a Km for DHEAsignificantly lower than for pregnenolone. This is notthe case for the corresponding isoform in the rat andmouse (Simard et al. 1996). In contrast to the twoother rodent species, the hamster adrenal expressesthe enzyme P450C17 and is therefore able to syn-thesize DHEA (LeHoux et al. 1992). The levels ofP450C17 in the hamster adrenal are low in compari-son to other steroidogenic enzymes so it is likely thatthe levels of DHEA synthesized are also low. Thismay be reflected in the very high affinity of DHEAfor hamster type 1 3â-HSD.The role of 3â-HSD in the liver and kidney is
unknown. In the liver it is possible that the roleof a 3-ketosteroid reductase is the inactivation of5á-DHT since this reaction is not reversible in thetype 3 enzyme and would lead to the inactivation ofthis androgen. The much greater expression of3â-HSD in the male liver as opposed to the femaleliver would be consistent with this hypothesis. Initself this is not proof; the expression of manydrug-metabolizing P450 s in the liver is sexuallydimorphic (Legraverend et al. 1992) and theseenzymes do not appear to be involved in themetabolism of sex steroids. It has been establishedthat 3á-reduction by the enzyme 3á-HSD is animportant step in 5á-DHT metabolism (Toscano1986). However evidence is accruing that these3á,5á metabolites are biologically active. Recently ithas been shown that parturition defects in type 1
4. Analysis of tissue expression of mRNA for the three hamster3â-HSD isoforms by RT-PCR from RNA isolated from male (M) and female(F) hamster tissues. The figure shows a Southern blot performed subsequentto the PCR using a 32P-labeled human type 1 3â-HSD cDNA.
3â-HSD isoforms in the hamster · and others 107
Journal of Molecular Endocrinology (1998) 20, 99–110
5á-reductase knock-out mice could be reversed bythe administration of 3á-Adiol; 3â-metabolites hadno effect (Mahendroo et al. 1996). It is thereforepossible that the 3â-HSD isoforms, particularlythe reductases, may play a role in preventing theformation of these active 3á,5á metabolites oftestosterone. It has been demonstrated that the rattype 1 enzyme, expressed in cultured cells, alsopossesses significant 17â-HSD activity, converting5á-DHT to 5á-androstenedione (de Launoit et al.1992b), which opens up the possibility that 3â-HSDhas an unidentified activity associated with its rolein the liver. The enzyme may not necessarily bemetabolizing steroids. For example, the structurallyunrelated enzyme 3á-HSD has also been shown topossess dihydrodiol dehydrogenase activity (Worner& Oesch 1984).In the hamster kidney, 3â-HSD immunostaining
is confined to the proximal and distal convolutedtubules. The kidney tubules are involved in fluid
and electrolyte balance. Again, the role of 3â-HSDin this tissue is unknown. In the guinea pig ithas been suggested that 3â-HSD is involved inin situ formation, in the kidney, of the mineralo-corticoid 11-deoxycorticosterone from circulating21-hydroxypregnenolone (Devine et al. 1986).However, in contrast to the guinea pig, in which11-deoxycorticosterone is the major mineralo-corticoid (Sandor et al. 1976), the hamster is ableto synthesize aldosterone (LeHoux & Ducharme1992), and it is therefore likely that this pathway isunimportant in the hamster. The type 2 11â-HSDis expressed in the distal tubules and collectingducts (Krozowski et al. 1995), and acts to inactivatecortisol, thereby protecting the non-selective type1 mineralocorticoid receptor (White et al. 1997). Itis possible that, in the kidney, 3â-HSD may also actto inactivate a biological compound.In summary, we have isolated cDNAs for three
different isoforms of 3â-HSD in the hamster and
5. Immunohistochemical localization of 3â-HSD in hamster tissues. (A) Adrenal gland (male) is positivelystained (brown) in the zona glomerulosa (G), zona fasciculata (F) and zona reticularis (R) of the cortex. The medulla(M) is negative except for scattered cells, likely cortical. (B) Kidney (male) has positive staining in some convolutedtubules in the cortex. (C) Liver (male) contains positively staining hepatocytes throughout with intensification aroundthe portal triads (T) and central vein (V). (D) Liver (female) has only scattered positive cells (arrowed). Sections werecounterstained with hematoxylin. Scale bars are in microns.
and others · 3â-HSD isoforms in the hamster108
Journal of Molecular Endocrinology (1998) 20, 99–110

characterized the corresponding proteins. Furthercharacterization of these enzymes should help inelucidating the functions of 3â-HSD in the liverand kidney.
ACKNOWLEDGEMENTS
We wish to acknowledge the technical assistanceprovided by Ms Andree Lefebvre and MsLyne Ducharme. A preliminary report of thiswork was presented at the Workshop on theMolecular and Cellular Biology of HydroxysteroidDehydrogenases held in Hanover, Germany, April1995 (Rogerson et al. 1995).
REFERENCES
Abbaszade IG, Clarke TR, Park C-HJ & Payne AH 1995 Themouse 3â-hydroxysteroid dehydrogenase multigene familyincludes two functionally distinct groups of proteins.Molecular Endocrinology 9 1214–1222.
Abbaszade IG, Arensburg J, Park C-HJ, Kasa-Vubu JZ,Orly J & Payne AH 1997 Isolation of a new mouse3â-hydroxysteroid dehydrogenase isoform, 3â-HSD VI,expressed during early pregnancy. Endocrinology 1381392–1399.
Bain PA, Yoo M, Clarke T, Hammond SH & Payne AH1991 Multiple forms of mouse 3â-hydroxysteroiddehydrogenase/Ä5]4isomerase and differential expression ingonads, adrenal glands, liver, and kidneys of both sexes.Proceedings of the National Academy of Sciences of the USA88 8870–8874.
Chen Z, Lin ZG, Lee WR & Chang SH 1991 Role of asparticacid 38 in the co-factor specificity of Drosophila alcoholdehydrogenase. European Journal of Biochemistry 202263–267.
Clarke TR, Bain PA, Sha L & Payne AH 1993a Enzymecharacteristics of two distinct forms of mouse3â-hydroxysteroid dehydrogenase/Ä5]4isomerasecomplementary deoxyribonucleic acids expressed in COS-1cells. Endocrinology 132 1971–1976.
Clarke TR, Bain PA, Greco TL & Payne AH 1993b A novelmouse kidney 3â-hydroxysteroid dehydrogenase comple-mentary DNA encodes a 3-ketosteroid reductase instead of a3â-hydroxysteroid dehydrogenase/Ä5]4isomerase. MolecularEndocrinology 7 1569–1578.
Couet J, Simard J, Martel C, Trudel C, Labrie Y & Labrie F1992 Regulation of 3-ketosteroid reductase messengerribonucleic acid levels and 3â-hydroxysteroiddehydrogenase/Ä5]4isomerase activity in rat liver by sexsteroids and pituitary hormones. Endocrinology 1313045–3050.
Devine PL, Kelly NS & Adams JB 1986 3â-Hydroxysteroidisomerase dehydrogenase in guinea pig kidney: possibleinvolvement in 11-deoxycorticosterone formation in situ.Journal of Steroid Biochemistry 25 265–270.
Keeney DS, Naville D, Milewich L, Bartke A & Mason JI1993a Multiple isoforms of 3â-hydroxysteroid dehydrogenase/Ä5]4isomerase in mouse tissues: male-specific forms areexpressed in the gonads and liver. Endocrinology 133 39–45.
Keeney DS, Murry BA, Wagner TE, Bartke A & Mason JI1993b Growth hormone transgenes regulate the expression of
sex-specific isoforms of 3â-hydroxysteroid dehydrogenase inmouse liver and gonads. Endocrinology 133 1131–1138.
Krozowski Z, Maguire JA, Stein-Oakley AN, Dowling J,Smith RE & Andrews RK 1995 Immunohistochemicallocalization of the 11â-hydroxysteroid dehydrogenase type IIenzyme in human kidney and placenta. Journal of ClinicalEndocrinology and Metabolism 80 2203–2209.
Lachance Y, Luu-The V, Verreault H, Dumont M, RheaumeE, LeBlanc G & Labrie F 1991 Structure of the human typeII 3â-hydroxysteroid dehydrogenase/Ä5]4isomerase gene:adrenal and gonadal specificity. DNA and Cell Biology 10701–711.
de Launoit Y, Zhao H-F, Belanger A, Labrie F & Simard J1992a Expression of liver-specific member of the3â-hydroxysteroid dehydrogenase family, an isoformpossessing an almost exclusive 3-ketosteroid reductaseactivity. Journal of Biological Chemistry 267 4513–4517.
de Launoit Y, Simard J, Durocher F & Labrie F 1992bAndrogenic 17â-hydroxysteroid dehydrogenase activityof expressed rat type 1 3â-hydroxysteroid dehydrogenase/Ä5]4isomerase. Endocrinology 130 553–555.
Legraverend C, Mode A, Wells T, Robinson I & GustafssonJA 1992 Hepatic steroid hydroxylating enzymes arecontrolled by the sexually dimorphic pattern of growthhormone secretion in normal and dwarf rats. FASEB Journal6 711–718.
LeHoux J-G & Ducharme L 1992 The differential regulationof aldosterone output in hamster adrenal by angiotensin IIand adrenocorticotropin. Journal of Steroid Biochemistry andMolecular Biology 41 809–814.
LeHoux J-G, Mason JI & Ducharme L 1992 In vivo effects ofadrenocorticotropin on hamster adrenal steroidogenicenzymes. Endocrinology 131 1874–1882.
Lorence MC, Murry BA, Trant JM & Mason JI 1990 Human3â-hydroxysteroid dehydrogenase/Ä5]4 isomerase fromplacenta: expression in nonsteroidogenic cells of a proteinthat catalyzes the dehydrogenation/isomerization of C21 andC19 steroids. Endocrinology 126 2493–2498.
Lorence MC, Naville D, Graham-Lorence SE, Mack SO,Murry BA, Trant JM & Mason JI 1991 3â-Hydroxysteroiddehydrogenase/Ä5]4 isomerase expression in rat andcharacterization of the testis isoform. Molecular and CellularEndocrinology 80 21–31.
Luu-The V, Lachance Y, Labrie C, Leblanc G, Thomas JL,Strickler RC & Labrie F 1989 Full length cDNA structureand deduced amino acid sequence of human 3â-hydroxy-5-ene steroid dehydrogenase. Molecular Endocrinology 31310–1312.
Mahendroo MS, Cala KM & Russell DW 1996 5á-Reducedandrogens play a key role in murine parturition. MolecularEndocrinology 10 380–392.
Naville D, Keeney DS, Jenkins G, Murry BA, Head JR &Mason JI 1991 Regulation of expression of male-specific ratliver microsomal 3â-hydroxysteroid dehydrogenase.Molecular Endocrinology 5 1090–1100.
Payne AH, Abbaszade IG, Clarke TR, Bain PA &Park C-HJ 1997 The multiple murine 3â-hydroxysteroiddehydrogenase isoforms: structure, function, and tissue-and developmentally specific expression. Steroids 62169–175.
Rogerson FM, LeHoux J-G & Mason JI 1995 Expressionand characterization of isoforms of 3â-hydroxysteroiddehydrogenase/Ä5]4isomerase in the hamster. Journalof Steroid Biochemistry and Molecular Biology 55481–487.
Sandor T, Fazekas AG & Robinson BH 1976 The biosynthesisof corticosteroids throughout the vertebrates. In General,Comparative and Clinical Endocrinology of the Adrenal
3â-HSD isoforms in the hamster · and others 109
Journal of Molecular Endocrinology (1998) 20, 99–110

Cortex, vol 1, pp 25–142. Eds I Chester Jones & IWHenderson. London: Academic Press.
Sanger F, Nicklen S & Coulsen AR 1977 DNA sequencingwith chain-terminating inhibitors. Proceedings of the NationalAcademy of Sciences of the USA 74 5463–5467.
Simard J, de Launoit Y & Labrie F 1991 Characterization ofthe structure–activity relationships of rat types I and II3â-hydroxysteroid dehydrogenase/Ä5]4isomerase bysite-directed mutagenesis and expression in HeLa cells.Journal of Biological Chemistry 266 14842–14845.
Simard J, Couet J, Durocher F, Labrie Y, Sanchez R, BretonN, Turgeon C & Labrie F 1993 Structure and tissue-specificexpression of a novel member of the rat 3â-hydroxysteroiddehydrogenase/Ä5]4isomerase (3â-HSD) family. Journal ofBiological Chemistry 268 19659–19668.
Simard J, Durocher F, Mebarki F, Turgeon C, Sanchez R,Labrie Y, Couet J, Trudel C, Rheaume E, Morel Y,Luu-The V & Labrie F 1996 Molecular biology and geneticsof the 3â-hydroxysteroid dehydrogenase/Ä5-Ä4 gene family.Journal of Endocrinology 150 S189–S207.
Tautz D & Renz M 1983 An optimal freeze–squeeze methodfor the recovery of DNA fragments from agarose gels.Analytical Biochemistry 132 16–19.
Thomas JL, Nash WE, Myers RP, Crankshaw MW & StricklerRC 1993 Affinity radiolabeling identifies peptides and amino
acids associated with substrate binding in human placental3â-hydroxy-Ä5-steroid dehydrogenase. Journal of BiologicalChemistry 268 18507–18512.
Toscano V 1986 Dihydrotestosterone metabolism. ClinicalEndocrinology and Metabolism 15 279–292.
White PC, Mune T & Agarwal AK 1997 11â-Hydroxysteroiddehydrogenase and the syndrome of apparent mineralo-corticoid excess. Endocrine Reviews 18 135–156.
Worner W & Oesch F 1984 Identity of dihydrodiol dehydro-genase and 3á-hydroxysteroid dehydrogenase in rat but notin rabbit liver cytosol. FEBS Letters 170 263–267.
Zhao H-F, Rheaume E, Trudel J, Couet J, Labrie F & SimardJ 1990 Structure and sexually dimorphic expression of aliver-specific rat 3â-hydroxysteroid dehydrogenase/isomerase.Endocrinology 127 3237–3239.
Zhao HF, Labrie C, Simard J, de Launoit Y, Trudel C,Martel C, Rheaume E, Dupont E, Luu-The V, Pelletier G& Labrie F 1991 Characterization of rat 3â-hydroxysteroiddehydrogenase/Ä5]4isomerase cDNAs and differentialtissue-specific expression of the corresponding mRNAs insteroidogenic and peripheral tissues. Journal of BiologicalChemistry 266 583–593.
9 June 1997
and others · 3â-HSD isoforms in the hamster110
Journal of Molecular Endocrinology (1998) 20, 99–110