increasing erucic acid content through combination of endogenous low polyunsaturated fatty acids...
TRANSCRIPT

Theor Appl Genet (2009) 118:765–773
DOI 10.1007/s00122-008-0936-7ORIGINAL PAPER
Increasing erucic acid content through combination of endogenous low polyunsaturated fatty acids alleles with Ld-LPAAT + Bn-fae1 transgenes in rapeseed (Brassica napus L.)
Ujjal K. Nath · Jeroen A. Wilmer · Emma J. Wallington · Heiko C. Becker · Christian Möllers
Received: 25 June 2008 / Accepted: 13 November 2008 / Published online: 3 December 2008© The Author(s) 2008. This article is published with open access at Springerlink.com
Abstract High erucic acid rapeseed (HEAR) oil is ofinterest for industrial purposes because erucic acid (22:1)and its derivatives are important renewable raw materialsfor the oleochemical industry. Currently available cultivarscontain only about 50% erucic acid in the seed oil. A sub-stantial increase in erucic acid content would signiWcantlyreduce processing costs and could increase market pros-pects of HEAR oil. It has been proposed that erucic acidcontent in rapeseed is limited because of insuYcient fattyacid elongation, lack of insertion of erucic acid into thecentral sn-2 position of the triaclyglycerol backbone anddue to competitive desaturation of the precursor oleic acid(18:1) to linoleic acid (18:2). The objective of the presentstudy was to increase erucic content of HEAR winter rape-seed through over expression of the rapeseed fatty acid
elongase gene (fae1) in combination with expression of thelysophosphatidic acid acyltransferase gene from Limnan-thes douglasii (Ld-LPAAT), which enables insertion of eru-cic acid into the sn-2 glycerol position. Furthermore,mutant alleles for low contents of polyunsaturated fattyacids (18:2 + 18:3) were combined with the transgenicmaterial. Selected transgenic lines showed up to 63% erucicacid in the seed oil in comparison to a mean of 54% erucicacid of segregating non-transgenic HEAR plants. Amongst220 F2 plants derived from the cross between a transgenicHEAR line and a non-transgenic HEAR line with a lowcontent of polyunsaturated fatty acids, recombinant F2
plants were identiWed with an erucic acid content of up to72% and a polyunsaturated fatty acid content as low as 6%.Regression analysis revealed that a reduction of 10% inpolyunsaturated fatty acids content led to a 6.5% increasein erucic acid content. Results from selected F2 plants wereconWrmed in the next generation by analysing F4 seeds har-vested from Wve F3 plants per selected F2 plant. F3 linescontained up to 72% erucic acid and as little as 4% polyun-saturated fatty acids content in the seed oil. The 72% erucicacid content of rapeseed oil achieved in the present studyrepresents a major breakthrough in breeding high erucicacid rapeseed.
Introduction
World vegetable oil markets are highly competitive requir-ing a steady improvement in oil quality to maintain orincrease market shares. Genetic improvement of oilseedshas the objective of increasing oil yields with a uniformfatty acid composition for nutritional, pharmaceutical andindustrial purposes (Roscoe 2005). Oil from traditionalrapeseed (Brassica napus L.) diVers signiWcantly from most
Dedicated to professor Gerhard Röbbelen on the occasion of his 80th birthday.
Communicated by I. Rajcan.
U. K. Nath · H. C. Becker · C. Möllers (&)Department of Crop Sciences, Plant Breeding, Georg-August-Universität Göttingen, Von-Siebold-Str. 8, 37075 Göttingen, Germanye-mail: [email protected]
J. A. Wilmer · E. J. WallingtonBIOGEMMA UK Ltd, 200, Science Park, CB4 0GZ Cambridge, UK
Present Address:J. A. WilmerBIOGEMMA, Domaine de Sandreau, Chemin de Panedautes, 31700 Mondonville, France
Present Address:E. J. WallingtonNIAB, Huntingdon Road, CB3 OLE Cambridge, UK
123

766 Theor Appl Genet (2009) 118:765–773
other vegetable oils by its high contents of erucic acid(22:1) and eicosenoic acid (20:1; Jönsson 1977; Beckeret al. 1999). They are the major very long chain fatty acids(VLCFAs) in the seed oil, together accounting for 45–60%of the total fatty acid mixture. High levels of erucic acid incooking and salad oil extracted from rapeseed have beenassociated with health problems (Beare et al. 1963). Fol-lowing identiWcation of a spontaneous rapeseed mutantwith low erucic acid content, genetic studies have shownthat this trait is under control of two loci that act in an addi-tive manner (Harvey and Downey 1964; Kondra and Ste-fansson 1965). Major eVorts in the 1960s led to thedevelopment of low erucic acid rapeseed (LEAR) varieties.Nowadays, double zero (00) or Canola quality cultivarswith a low erucic acid content in the seed oil and a lowglucosinolate content in the seeds are predominantly beinggrown in most parts of the world.
However, High Erucic Acid Rapeseed (HEAR) typesretained some importance. HEAR cultivars are presentlycultivated to a small extent in Europe (up to 40,000 hect-ares in 2006/2007) and in USA/Canada as an identity pre-served crop (Möllers 2004; Scarth and Tang 2006). Erucicacid and its derivatives are important renewable raw mate-rials used in plastic Wlm, nylon, lubricant, cosmetic andemollient industries (Leonard 1994; Sonntag 1995; Piazzaand Foglia 2001). Currently available HEAR cultivars con-tain only about 50% erucic acid in the seed oil. A substan-tial increase of the erucic acid content of the rapeseed oilwould signiWcantly reduce processing costs and couldincrease market prospects. However, possibilities toincrease erucic acid by classical breeding are limited,because in rapeseed and related Brassica species erucicacid is inserted only in the Wrst (sn-1) and third (sn-3) posi-tion of the glycerol backbone. This limits erucic acid con-tent to a maximum of 67%. The reason for this limitationlies in the speciWcity of the B. napus sn-2 acyltransferase(LPAAT—lysophosphatidic acid acyltransferase), whichdoes not accept erucoyl-CoA as a substrate (Cao et al.1990; Frentzen 1993). To overcome this limitation, thegene for an erucoyl-CoA preferring sn-2 acyltransferasewas isolated from Limnanthes species. Expression of thisgene in transgenic rapeseed altered seed oil sn-2 propor-tions of erucic acid, but did not lead to an increase in erucicacid content of the seed oil (Lassner et al. 1995; Broughet al. 1996; Weier et al. 1997).
In a next step, interest focussed on the fatty acid elonga-tion mechanism from oleic acid to erucic acid. This elonga-tion is the result of two cycles of a four-step mechanism, inwhich Wrst oleoyl-CoA and then eicosenoyl-CoA serveas substrates (Puyaubert et al. 2005). In the Wrst step, the�-ketoacyl-CoA synthase (KCS; fae1 gene) catalyses thecondensation reaction of oleoyl-CoA or eicosenoyl-CoAwith malonyl-CoA. It is believed that this initial reaction is
the rate-limiting step of the four-step mechanism (Cassagneet al. 1994). The fae1 genes from Arabidopsis and fromrapeseed were cloned and over expressed under control of aseed speciWc promoter in transgenic HEAR (Katavic et al.2001; Han et al. 2001). However, this led only to a minorincrease in erucic acid content. Even the combination withthe expression of the Ld-LPAAT gene from Limnanthesdouglasii did not result in a substantial increase of the eru-cic acid content in transgenic HEAR (Han et al. 2001).
There is some evidence that the cytosolic pool of avail-able oleoyl-CoA or malonyl-CoA may limit fatty acid elon-gation (Bao et al. 1998; Domergue et al. 1999). Crossingconventional HEAR with oilseed rape with reduced con-tents of linoleic acid (18:2) and linolenic acid (18:3) didresult in recombinant high erucic low polyunsaturated fattyacid (HELP) F3 plants which, however, did not show anincreased erucic acid content compared to the parentalHEAR genotype (50% erucic acid; Sasongko and Möllers2005). An obvious explanation for this result was that in theHELP material the activity of the �-ketoacyl-CoA synthaseactivity (fae1.1 and fae1.2 genes) was too low to allow forenhanced erucic acid synthesis.
The Wrst objective of the present study was to repeat theapproach of Han et al. (2001) to increase erucic acid contentby expressing the Ld-LPAAT gene and over expressing theBn-fae1 gene in transgenic HEAR. The second objective wasto study in this material, the eVect of genetically reducedpolyunsaturated fatty acids content on erucic acid content.
Materials and methods
Plant material
6575-1 HELP is a winter rapeseed F4 line with 27% oleicacid, 7% linoleic and linolenic acid and 50% erucic acidcontent obtained from a cross between the winter rapeseedcultivar Maplus and the high oleic acid doubled haploidwinter rapeseed line DH XXII D9 (for details see Sasongkoand Möllers 2005). BGRV2 is a UK high erucic acid winteroilseed rape breeding line from Nickerson UK Ltd withabout 52% erucic acid in the seed oil (Wilmer et al. 2003).This line was used in the transformation experiments toproduce transgenic line 361.2B (see below). F1 plants wereobtained by crossing transgenic line 361.2B with line 6575-1 HELP. Segregating F2 and F3 plant generations wereproduced by growing the plants in the greenhouse and selfpollination of F1 and F2 plants, respectively.
Performance of greenhouse experiments
In August 2005, 220 randomly selected F2 seeds along witheight seeds from each parent were sown for producing the
123

Theor Appl Genet (2009) 118:765–773 767
F2 plant population. F2 seeds were sown in a multipot traycontaining T-soil (Fruhstorfer Erde; pH 5.9). F2 plants wereallowed to grow for three weeks in the greenhouse. For ver-nalisation, plantlets were transferred to 4°C temperaturewith 8 h light for 8 weeks. Afterwards, plantlets were trans-ferred to 9 cm diameter pots containing normal compostsoil. Plantlets were cultivated in the greenhouse providing16 h light by using 400 W sodium-steam lamps. Tempera-ture varied during the day from 20 to 25°C and during thenight from 10 to 15°C. Hakaphos fertilizer containingN:P:K (15:11:15) + 0.2% trace minerals was added at twoweek intervals to the top soil (100 mg) of each pot until theplants matured. Insecticide was applied when aphid andthrips attack was recognized. Sulphur vapour supply wasconstant during the whole experiment to avoid fungal dis-eases. Self-fertilisation was ensured by covering the Xowersof the main raceme with micropore plastic bags. F3 seedswere harvested at maturity from the main raceme in Febru-ary 2006 and stored at 4°C.
During the period October 2006 to April 2007, thegreenhouse experiment was repeated with F3 seeds (fromsowing to harvest). For this experiment 41 F2 plants with anerucic acid content higher than the mean of parent 361.2B(>62.5%) were selected. Five F3 seeds from each of the F2
plants and of the parents were sown. The greenhouse exper-iment was conducted as described above, following a ran-domized complete block design. The Wve F3 plants fromeach F2 plant and the parents were considered as Wve repli-cates. Five separate tables inside the green house repre-sented the complete blocks consisting of all genotypes (F3
plants and parents). Following self-fertilisation F4 seedswere harvested from main raceme.
Methods
Binary plasmid construction
The fae1-1 (C-genome) and fae1-2 (A-genome) genes wereampliWed from BGRV2 using primers BnFAE-F (5� CCTCATGACGTCCATTAACGTAAAGCTCC 3�) and BnFAE-R (5� GTGAGCTCTTATTAGGACCGACCGTTTGGG 3�),cloned into pBluescript II (KS+) (Stratagene), andsequenced. Next, the fae genes were transferred as RcaI–EcoICRI fragment into pAR4 (Biogemma UK) NcoI–SmaIsites to place the gene between the Pnapin and CHS polyAsequences. The expression cassettes were then lifted out as aSalI–SacI fragment and ligated into pT7Blue2 SalI–SacIsites, before transfer of an EcoRI–EcoICRI fragment into thebinary vector pSCVnos144 (Biogemma UK) EcoRI–SmaIadjacent to a similar Pnapin::lat2 LPAAT::CHS poly Acassette (Brough et al. 1996) to create pEW13 and pEW14(Wilmer et al. 2001).
Agrobacterium tumefaciens transformation and generation of T1-plants
Agrobacterium tumefaciens strain C58 pMP90 carryingabove described binary plasmid pEW13 or pEW14 wereused to transform rapeseed line BGRV2 essentially follow-ing the protocol of Moloney et al. (1989). Ten transgenicplants (T1) carrying pEW13 and 18 transgenic plants withpEW14 were regenerated and oil composition was analysedfollowing self pollination of T1 and T2 plants. Erucic acidcontent in these lines varied between less than 30 and about65%. Line 361.2B, transformed with pEW14—fae1-2, wasselected as it had the highest erucic acid content in T3 seeds(Wilmer et al. 2001). T4 seeds of 361.2B were used in thecrossing experiment described above.
DNA isolation and PCR analysis to follow transgene segregation in the F2 population
DNA was isolated from leaf material frozen in liquid nitro-gen (¡196°C) following the protocol of Ishizawa et al.(1991) with some modiWcations as described in Nath et al.(2007). Multiplex-PCR was performed using speciWc prim-ers for the Ld-LPAAT gene and locus speciWc primers forthe endogenous single copy Bn-fad2 gene. AmpliWcation ofthe single copy Bn-fad2 gene served as positive control forthe presence of DNA in suYcient quantity and quality. PCRprimers, PCR conditions and gel electrophoresis were asdescribed in Nath et al. (2007).
Seed quality analysis
Trierucin analysis
Seed samples (150 mg) obtained from F2 and F3 plantswere analysed for trierucin (C69; EEE) content by high tem-perature gas liquid chromatography (HT-GLC) analysisaccording to the method described by Möllers et al. (1997).The analysis was done using silicon capillary column RTX-65TG (Restek no. 17,005) 15 m £ 0.25 mm i.d. (0.1 �mWlm thickness). Trierucin is expressed as percent of the sumof all triglycerides.
Analysis of total fatty acid composition
Following trierucin analysis the remaining part of eachsample was transferred to a new tube and left on a hot plateat 37.5°C over night to evaporate. The fatty acid proWle wasdetermined by analysis of methyl esters by gas liquid chro-matography according to Rücker and Röbbelen (1996).Fatty acids are expressed as percent of the sum of all fattyacids. The following fatty acids were determined: palmiticacid (16:0), stearic acid (18:0), oleic acid (18:1), linoleic
123

768 Theor Appl Genet (2009) 118:765–773
acid (18:2), linolenic acid (18:3), eicosenoic acid (20:1),erucic acid (22:1) and nervonic acid (24:1). Saturated fattyacids (SFA; 16:0 + 18:0), polyunsaturated fatty acids(PUFA; 18:2 + 18:3) and monounsaturated fatty acids(MUFA; 18:1 + 20:1 + 22:1) were calculated from the con-tents of individual fatty acids.
Analysis of the fatty acid compositionat the sn-2 position of triacylglycerols
A total of 15 mg mixed seed samples from 5 replicates ofthe F3 population (F4 seeds) were collected in a 5 ml plastictube. A measure of 0.5 ml iso-octane:iso-propanol (9:1)was added to each sample and seeds were homogenisedwith a steel rod. The supernatant was transferred to a newtube and the solvent evaporated by using a stream of warmair. The dried oil residue was mixed with 500 �l buVer(50 mM KH2PO4-, pH 7.2 with 0.5% Triton X-100) andkept in a supersonic bath for 2 min. The mixture was incu-bated at 30°C for 1 h by adding 25 �l (250 units) lipasefrom Rhizopus arrhizus (Sigma-Aldrich). Then 200 �lpetroleum ether-70:iso-propanol (3:2) was added and thesupernatant was collected in a 1 ml glass tube by centrifu-gation at 150 £ G (1,000 rpm) for 5 min. The extractionwas repeated twice and the supernatants were merged andevaporated by using a stream of warm air. 25 �l petroleumether-70:iso-propanol (3:2) was added and mixed well byvortex. A measure of 20 �l from this mixture was taken andtransferred to a thin layer chromatography (TLC) plate(F 1,500/LS 254, 20 £ 20 cm) using a 20 �l syringe. TLCplates were placed in the eluent diethylether/petroleumether(3:1) and allowed to run for 30 min. Afterwards dry TLCplates were stained by subjecting them to iodine vapour.Monoacylglycerol spots (representing the fatty acids at thesn-2 position) were marked with a pencil. Following evapo-ration of the iodine, the marked areas were scraped outusing a scalpel and transferred for fatty acid extraction in aseparate 1 ml glass tube. The scrape was mixed with 250 �liso-octane, incubated for 20 min and the supernatant was
collected by centrifugation in a new 1 ml glass tube. Thisprocedure was repeated twice. Iso-octane from the col-lected supernatant was evaporated and then the fatty acidswere extracted and analysed as described above.
Analysis of seed oil and protein content
Oil and protein content of seed samples were determined byNear-Infrared-ReXectance-Spectroscopy (NIRS) using theequation raps2001.eqa (Tillmann 2007). Spectra wererecorded using a standard ring cup equipped with a 14 mmPVC adapter. Values were adjusted using previously devel-oped regression equations. Oil and protein contents areexpressed on seed dry matter basis.
Statistical analysis
Spearman’s rank correlation coeYcients were calculatedusing the Plant Breeding Statistical Program PLABSTAT(Version 2N; Utz 2007). Multiple mean comparisons weremade with Fisher’s least signiWcant diVerence (LSD) proce-dure using Stat Graphics Plus for Windows 3.0 (StatisticalGraphics Corp., Rockville, USA). Hypotheses for trans-gene copy number segregation in the F2 population weretested using Chi-square (�2) test as described by Gomez andGomez (1976).
Results
Variation of seed quality traits and segregation of the transgenes in the F2 population
The F2 population derived from the cross between the trans-genic line 361.2B and the high erucic acid, low polyunsatu-rated fatty acid line 6575-1 HELP showed a largequantitative variation for erucic acid content and other traits(Table 1). Erucic acid (22:1) was the most prominent fattyacid, accounting for 58.8% of the total fatty acid content.
Table 1 Variation in oil, protein, fatty acids and trierucin content (%) in 220 F2 plants (F3 seeds) derived from the cross 361.2B £ 6575-1 HELPin comparison to the parental lines (each parent with n = 8)
SFA = 16:0 + 18:0, PUFA = 18:2 + 18:3, MUFA = 18:1 + 20:1 + 22:1
Item Oil content
Protein content
Fatty acid contents (%)
SFA 18:1 PUFA 20:1 22:1 MUFA Trierucin
361.2B Range 46–54 24–26 2–5 8–11 16–24 3–7 61–64 72–78 9–18
Mean 49.9 24.1 3.2 8.3 19.1 4.6 62.5 75.4 15.2
6575-1 HELP Range 45–51 22–27 4–6 26–31 7–9 7–11 49–52 83–89 0.0–0.0
Mean 49.5 23.9 4.9 27.8 8.1 8.9 49.4 86.1 0.0
F2 population Range 37–54 19–33 2–9 5–26 6–26 2–10 44–72 64–87 0–25
Mean 47.7 24.8 4.0 11.0 16.8 5.2 58.8 75.0 12.6
123

Theor Appl Genet (2009) 118:765–773 769
Its content ranged from 44 to 72% and showed large trans-gressive segregation compared to the higher parent 361.2B.Frequency distribution of the erucic acid content showed acontinuous variation indicating polygenic inheritance(Fig. 1). Polyunsaturated fatty acid content (PUFA;18:2 + 18:3) varied from six to 26%, in some F2 plantsthereby being as low as in the 6575-1 HELP parent. Trieru-coylglycerol (Trierucin) content showed a discontinuousvariation from 0.0 to 25% (Table 1 and Fig. 2), indicatingsegregation and functioning of the Ld-LPAAT transgene.From the 220 F2 plants analysed, 211 contained trierucin(Fig. 2), suggesting the presence of two transgene copies intransgenic parent 361.2B and a 15:1 segregation in the F2
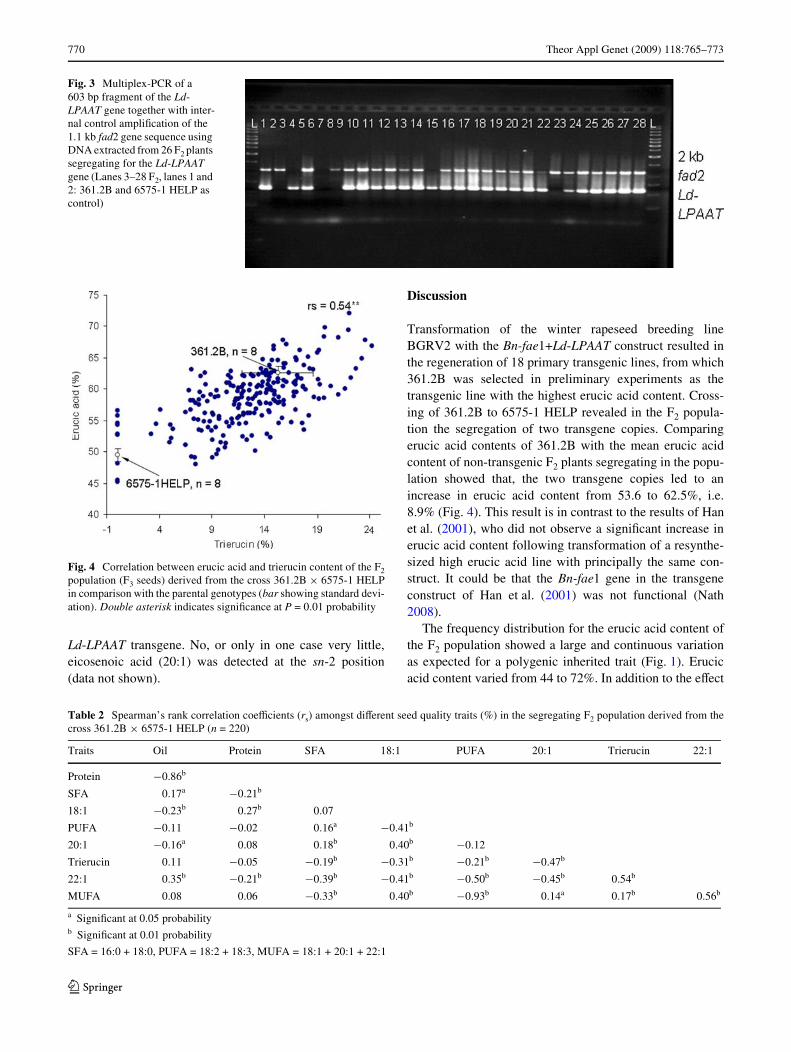
population (Chi-square �2 = 2.52). Using PCR primersdirected against the Ld-LPAAT transgene, four out of 57randomly selected F2 plants were negative in PCR (see an
example in Fig. 3), again conWrming 15:1 segregation oftwo transgene copies in the F2 population (Chi-square�2 = 0.06). All F2 plants positive in PCR for the Ld-LPAATgene contained trierucin, whereas those negative in PCRdid not contain trierucin. The clearly separated group ofsegregating non-transgenic F2 plants (Fig. 2) lacking theLd-LPAAT + Bn-fae1 transgenes contained between 45 and57% erucic acid in the seed oil (see Fig. 4), with a mean of53.6%. Parent 361.2B contained 62.5% erucic acid in theseed oil, indicating that the two transgenes in the homozy-gous state in this population increased erucic acid contentby 8.9%.
Correlations between traits in the F2 population
Spearman’s rank correlation coeYcient analysis of the F2
population (F3 seeds) showed negative correlationsbetween erucic acid and protein, SFA, oleic acid, PUFAand eicosenoic acid content (Table 2). Correlationsbetween erucic acid content and oil, trierucin and MUFAcontent were positive (see also Fig. 4). The calculation ofthe regression between erucic acid content and PUFA con-tent (y = ¡0.65x + 69.63) showed that a reduction of 10%in PUFA content led to an increase of 6.5% in erucic acidcontent.
Performance of selected F3 lines
From the F2 population, 41 plants showed a higher erucicacid content (>62.5%) than the mean of parent 361.2B.Five F3 plants from each of those 41 F2 plants were culti-vated in the greenhouse and F4 seeds were harvested. In theF3 population, erucic acid content varied from 50 to 72%(Fig. 5) with the mean of 64.8%. The scatter plot for themean erucic acid content of F3 lines (F4 seeds) versus F2
plants (F3 seeds) shows a signiWcant positive correlation(rs = 0.57**; Fig. 5), proving eVective selection of high eru-cic acid plants amongst single plants of the F2 population.The fatty acid composition of the six F3 lines (F4 seeds)with the highest erucic acid contents is shown in Table 3. F3
line III-G-7 contained with 72.3% the highest amount oferucic acid. Four of the F3 lines had an equal to or lowerPUFA content than parent 6575-1 HELP, indicating homo-zygosity of the genes causing low PUFA content. All six F3
lines showed higher trierucin contents compared to trans-genic parent 361.2B.
From the six F3 lines with the highest erucic acid contentand the parental lines, the fatty acid composition at the sn-2position of the triacylglycerols was analysed. In the F3 linesthe erucic acid content at the sn-2 position varied from 36.8to 65.3%, which compares favourably to the 31.6% oftransgenic parent 361.2B (Table 3). Erucic acid at sn-2position was only detected in case of the presence of
Fig. 1 Frequency distribution of erucic acid content in the F2 popula-tion (F3 seeds) derived from a cross between 361.2B £ 6575-1 HELPrapeseed lines
Fig. 2 Frequency distribution of trierucin (C69; EEE) content in F2population (F3 seeds) of the cross 361.2B £ 6575-1 HELP with meanof the parents (n = 8)
123
770 Theor Appl Genet (2009) 118:765–773
Ld-LPAAT transgene. No, or only in one case very little,eicosenoic acid (20:1) was detected at the sn-2 position(data not shown).
Discussion
Transformation of the winter rapeseed breeding lineBGRV2 with the Bn-fae1+Ld-LPAAT construct resulted inthe regeneration of 18 primary transgenic lines, from which361.2B was selected in preliminary experiments as thetransgenic line with the highest erucic acid content. Cross-ing of 361.2B to 6575-1 HELP revealed in the F2 popula-tion the segregation of two transgene copies. Comparingerucic acid contents of 361.2B with the mean erucic acidcontent of non-transgenic F2 plants segregating in the popu-lation showed that, the two transgene copies led to anincrease in erucic acid content from 53.6 to 62.5%, i.e.8.9% (Fig. 4). This result is in contrast to the results of Hanet al. (2001), who did not observe a signiWcant increase inerucic acid content following transformation of a resynthe-sized high erucic acid line with principally the same con-struct. It could be that the Bn-fae1 gene in the transgeneconstruct of Han et al. (2001) was not functional (Nath2008).
The frequency distribution for the erucic acid content ofthe F2 population showed a large and continuous variationas expected for a polygenic inherited trait (Fig. 1). Erucicacid content varied from 44 to 72%. In addition to the eVect
Fig. 4 Correlation between erucic acid and trierucin content of the F2population (F3 seeds) derived from the cross 361.2B £ 6575-1 HELPin comparison with the parental genotypes (bar showing standard devi-ation). Double asterisk indicates signiWcance at P = 0.01 probability
Fig. 3 Multiplex-PCR of a 603 bp fragment of the Ld-LPAAT gene together with inter-nal control ampliWcation of the 1.1 kb fad2 gene sequence using DNA extracted from 26 F2 plants segregating for the Ld-LPAAT gene (Lanes 3–28 F2, lanes 1 and 2: 361.2B and 6575-1 HELP as control)
Table 2 Spearman’s rank correlation coeYcients (rs) amongst diVerent seed quality traits (%) in the segregating F2 population derived from thecross 361.2B £ 6575-1 HELP (n = 220)
a SigniWcant at 0.05 probabilityb SigniWcant at 0.01 probability
SFA = 16:0 + 18:0, PUFA = 18:2 + 18:3, MUFA = 18:1 + 20:1 + 22:1
Traits Oil Protein SFA 18:1 PUFA 20:1 Trierucin 22:1
Protein ¡0.86b
SFA 0.17a ¡0.21b
18:1 ¡0.23b 0.27b 0.07
PUFA ¡0.11 ¡0.02 0.16a ¡0.41b
20:1 ¡0.16a 0.08 0.18b 0.40b ¡0.12
Trierucin 0.11 ¡0.05 ¡0.19b ¡0.31b ¡0.21b ¡0.47b
22:1 0.35b ¡0.21b ¡0.39b ¡0.41b ¡0.50b ¡0.45b 0.54b
MUFA 0.08 0.06 ¡0.33b 0.40b ¡0.93b 0.14a 0.17b 0.56b
123

Theor Appl Genet (2009) 118:765–773 771
caused by the two transgene copies, this variation may bedue to segregation of loci of parent 6575-1 HELP responsi-ble for low contents of polyunsaturated fatty acids (PUFA).The low PUFA content in parent 6575-1 HELP is caused bya mutation in the oleic acid desaturase fad2 gene and bytwo to three other unknown genes having minor eVects(Schierholt et al. 2001; Sasongko and Möllers 2005).According to the regression, a 10% reduction in PUFA con-tent leads to a 6.5% increase in erucic acid content. Nath(2008) reported similar results for a high erucic acid dou-bled haploid winter rapeseed population segregating forPUFA content. Furthermore, environmental eVects, diVer-ent eVective alleles at the two endogenous erucic acidloci fae1.1 (A-genome, B. rapa) and fae1.2 (C-genome,
B. oleracea) as well as other unknown genetic factors mayhave contributed to the variation found in the F2 population(Stefansson and Hougen 1964; Jönsson 1977; Ecke et al.1995; Zhao et al. 2008).
Selection for erucic acid content amongst segregating F2
plants proved to be eYcient as shown by comparing theerucic acid contents of F2 plants with those of the derivedF3 plants (Fig. 5). F2 plants with about 70% erucic acidcontent in the seed oil were conWrmed in the F3 generation(F4 seeds, Table 3). It is likely that the F3 plants are alreadyhomozygous for the two transgene copies and for theendogenous two erucic acid genes. However, F4 plant gen-eration need to be tested to conWrm this. Some of the higherucic acid lines had PUFA contents as low as in parent6575-1 HELP (Table 3), indicating homozygosity for thegenes causing low PUFA content.
The F3 line III-G-7 had with 72.3% the highest erucicacid content in the seed oil as a mean of Wve plants(Table 3). This is about 9 and 23% more than that of trans-genic parent 361.2B and non-transgenic parent 6575-1HELP. The 72.3% erucic acid content can be regarded asrelatively stable, because the two parental lines had similarerucic acid contents at two diVerent seasons in the green-house (compare results in Tables 1 and 3). Furthermore,parent 6575-1 HELP was tested previously in Weld experi-ments and likewise had about 50% erucic acid in the seedoil (Sasongko and Möllers 2005).
The formation of trierucoylglycerol (Trierucin, C69)proved that the Limnanthes douglasii lysophosphatidic acidacyltransferase (Ld-LPAAT) functioned in transgenic oil-seed rape. Amongst the six F3 lines with the highest erucicacid content, line VI-D-9 showed the highest trierucin con-tent (23.5%; Table 3). Variation amongst the lines might bedue to diVerences in homozygosity of the transgenes and inthe availability of erucoyl-CoA. Assuming random distri-bution of erucoyl-CoA to all three triacylglycerol positions,
Fig. 5 Scatter plot for erucic acid content (%) of the selected 41 F2individuals and derived F3 lines (F4 seeds; mean of Wve plants) alongwith the parents (bar showing standard deviation). Double asteriskindicates signiWcance at P = 0.01 probability
Table 3 Comparison for the traits oil, protein, fatty acids and trierucin content (%) of six selected high erucic acid F3 lines (F4 seeds) of the cross361.2B £ 6575¡1 HELP along with their parents (values are means of Wve plants)
SFA = 16:0 + 18:0, PUFA = 18:2 + 18:3, MUFA = 18:1 + 20:1 + 22:1, 22:1 (sn-2) erucic acid content at the sn-2 position
Mean values with diVerent letters indicate signiWcant diVerences at P = 0.05 (Fisher’s LSD)
Line Oil content
Protein content
Fatty acid content (%)
SFA 18:1 PUFA 20:1 22:1 MUFA Trierucin 22:1 (sn-2)
361.2B 50.2 b 24.4 c 2.9 b 8.3 f 15.3 a 4.3 de 63.2 c 75.8 d 11.4 d 31.6
HELP 51.1 b 21.4 de 3.3 a 26.5 a 5.6 c 11.2 a 49.6 d 87.3 b – 0.5
IV-D-3 54.2 a 21.9 de 2.1 cd 14.2 b 5.3 c 7.1 b 68.7 b 90.0 a 13.1 cd 38.5
II-B-2 47.9 c 28.5 a 2.3 c 10.7 de 7.7 b 4.4 de 70.5 ab 85.6 c 14.7 c 36.8
II-G-8 49.8 bc 24.4 c 2.2 cd 9.3 ef 8.5 b 3.9 e 70.7 ab 83.9 c 18.7 b 40.3
IV-F-6 53.6 a 22.9 cd 2.3 c 14.0 b 3.9 d 6.1 bc 70.9 ab 91.0 a 13.1 cd 41.3
VI-D-9 55.1 a 20.7 e 2.0 cd 13.2 bc 4.7 cd 5.3 cd 71.4 a 89.9 a 23.5 a 61.3
III-G-7 49.7 bc 26.7 b 1.9 d 12.1 cd 5.7 c 5.0 cd 72.3 a 89.4 a 22.2 a 65.3
123

772 Theor Appl Genet (2009) 118:765–773
one would expect a trierucin content of 36% for F3 lineVI-D-9 with 71.4% erucic acid content. However, thisline showed only 23.5% trierucin content, indicating thatLd-LPAAT activity may not be strong enough in compari-son to the resident rapeseed Bn-LPAAT activity, which hasa strong preference for oleic acid (Frentzen 1998). The sn-2fatty acid mixture of the seed oils of the F3 lines containedup to 65% erucic acid (Table 3). However, lines with simi-lar erucic acid content contained very diVerent amounts oferucic acid in the sn-2 position, indicating that beside theLd-LPAAT gene there may be other genes involved in theexpression of this trait. No, or very limited amounts of eru-cic acid at sn-2 position were found in non-transgenicparental line 6575-1 HELP (Table 3). Similar results werealso reported by Weier et al. (1997) and Han et al. (2001) inLd-LPAAT expressing transgenic rapeseed lines. The lackof signiWcant amounts of eicosenoic acid at the sn-2 posi-tion conWrms the pronounced preference of the Ld-LPAATenzyme for erucoyl-CoA (Han et al. 2001).
The around 72% erucic acid content achieved in thepresent experiments represents a milestone in the breedingof high erucic acid oilseed rape. Further increases in erucicacid content can be expected from progress in reducing thecontents of the remaining fatty acids, mainly oleic acid,polyunsaturated fatty acids and eicosenoic acid. Reductionin PUFA content probably could be achieved more easilyfollowing an RNAi antisense approach (Stoutjesdijk et al.2002). Integration of the antisense fad2 gene into the sameT-DNA as the Ld-LPAAT and the Bn-fae1 gene wouldgreatly simplify the genetics and breeding of high erucicacid cultivars. The material developed in the present studyshould be of interest for the oleochemical industry but alsofor further studies aimed at identifying other physiologicallimitations in erucic acid biosynthesis.
Acknowledgments UKN gratefully acknowledges German Aca-demic Exchange Service (DAAD), Bonn, Germany, for providing ascholarship, Uwe Ammermann and Rosemarie Clemens for excellenttechnical assistance. Technical assistance for B. napus plant transfor-mation was provided by Andrea Grey and Ruth Bates (BiogemmaUK).
Open Access This article is distributed under the terms of the Cre-ative Commons Attribution Noncommercial License which permitsany noncommercial use, distribution, and reproduction in any medium,provided the original author(s) and source are credited.
References
Bao X, Pollard M, Ohlrogge J (1998) The biosynthesis of erucic acidin developing embryos of Brassica rapa. Plant Physiol 118:183–190
Beare JL, Campbell JA, Youngs CG, Craig BM (1963) EVects of sat-urated fat in rats fed rapeseed oil. Can J Biochem Physiol 41:605–612
Becker HC, Löptien H, Röbbelen G (1999) Breeding: an overview. In:Gomez-Campo C (ed) Biology of Brassica coenospecies. Else-vier, Amsterdam, pp 413–460
Brough CL, Coventry JM, Christie WW, Kroon JTM, Brown AR,Barsby TL, Slabas AR (1996) Towards the genetic engineering oftriacylglycerols of deWned fatty acid composition: major changesin erucic acid content at the sn-2 position aVected by the introduc-tion of a 1-acyl-sn-glycerol-3-phosphate acyltransferase fromLimnanthes douglasii into oil seed rape. Mol Breed 2:133–142
Cao YZ, Oo KC, Huang AHC (1990) Lysophosphatidate acyl transfer-ase in the microsomes from maturing seeds of meadowfoam(Limnanthes alba). Plant Physiol 94:1199–1206
Cassagne C, Lessire R, Bessoule JJ, Moreau P, Creach A, Schneider F,Sturbois B (1994) Biosynthesis of very long chain fatty acids inhigher plants. Prog Lipid Res 33:55–69
Domergue F, Chevalier S, Santarelli X, Cassagne C, Lessire R (1999)Evidence that oleoyl-CoA and ATP-dependent elongations coex-ist in rapeseed (Brassica napus L.). Eur J Biochem 263:464–470
Ecke W, Uzunova M, Weissleder K (1995) Mapping the genome of rape-seed (Brassica napus L.) II. Localization of genes controlling erucicacid synthesis and seed oil content. Theor Appl Genet 91:972–977
Frentzen M (1993) Acyltransferases and triacylglycerols. In: MooreTS Jr (ed) Lipid metabolism in plants. CRC Press, Boca Raton,pp 195–220
Frentzen M (1998) Acyltransferases from basic science to modiWedseed oils. Fett/Lipid 100:161–166
Gomez KA, Gomez AA (1976) Statistical procedures for agriculturalresearch with emphasis on rice. IRRI, Los Banos
Han J, Lühs W, Sonntag K, Zähringer U, Borchardt DS, Wolter FP,Heinz E, Frentzen M (2001) Functional characterization of�-ketoacyl-CoA synthase genes from Brassica napus L. PlantMol Biol 46:229–239
Harvey BL, Downey RK (1964) The inheritance of erucic acid contentin rapeseed (Brassica napus). Can J Plant Sci 44:104–111
Ishizawa M, Kobayashi Y, Miyamura T, Matsuura S (1991) Simpleprocedure of DNA isolation from human serum. Nucleic AcidsRes 19:5792
Jönsson R (1977) Erucic acid heredity in rapeseed (Brassica napus L.and Brassica campestris L.). Hereditas 86:159–170
Katavic V, Friesen W, Barton DL, Gossen KK, Giblin EM, Luciw T,An J, Zou J, MacKenzie SL, Keller WA, Males D, Taylor DC(2001) Improving erucic acid content in rapeseed through bio-technology: what can the Arabidopsis FAE1 and the yeast SLC1–1 genes contribute? Crop Sci 41:739–747
Kondra ZP, Stefansson BR (1965) Inheritance of erucic and eicosenoicacid content of rapeseed oil (Brassica napus). Can J Genet Cytol7:500–510
Lassner MW, Levering CK, Davies HM, Knutzon DS (1995) Lyso-phosphatidic acid acyltransferase from meadowfoam mediatesinsertion of erucic acid at the sn-2 position of triacylglycerol intransgenic rapeseed oil. Plant Physiol 109:1389–1394
Leonard C (1994) Sources and commercial applications of high erucicvegetable oils. Lipid Technol 4:79–83
Möllers C (2004) Potential and future prospects for rapeseed oil. In:Gunstone FD (ed) Rapeseed and canola oil-production, process-ing, properties and uses. Blackwell, Oxford, pp 186–217
Möllers C, Lühs W, SchaVert E, Thies W (1997) High-temperature gaschromatography for the detection of trierucoylglycerol in the seed oilof transgenic rapeseed (Brassica napus L.). Fett/Lipid 99:356–362
Moloney MM, Walker JM, Sharma KK (1989) High eYciency trans-formation of Brassica napus using Agrobacterium vectors. PlantCell Rep 8:238–242
Nath UK, Iqbal MCM, Möllers C (2007) Early, non-destructive selec-tion of microspore-derived embryo genotypes in oilseed rape(Brassica napus L.) by molecular markers and oil quality analy-sis. Mol Breed 19:285–289
123

Theor Appl Genet (2009) 118:765–773 773
Nath UK (2008) Increasing erucic acid content in the seed oil ofrapeseed (Brassica napus L.) by combining selection for naturalvariation and transgenic approaches. PhD thesis Georg-August-Universität Göttingen, Faculty of Agriculture. http//webdoc.sub.gwdg.de/diss/2008/nath/nath.pdf
Piazza GJ, Foglia TA (2001) Rapeseed oil for oleochemical uses. EurJ Lipid Sci Technol 103:405–454
Puyaubert J, Garcia C, Chevalier S, Lessire R (2005) Acyl-CoA elong-ase, a key enzyme in the development of high-erucic acid rape-seed? Eur J Lipid Sci Technol 107:263–267
Roscoe TJ (2005) IdentiWcation of acyltransferases controlling triacyl-glycerol biosynthesis in oilseeds using a genomic based approach.Eur J Lipid Sci Technol 107:162–256
Rücker B, Röbbelen G (1996) Impact of low linolenic acid content onseed yield of winter oilseed rape (Brassica napus L.). Plant Breed115:226–230
Sasongko ND, Möllers C (2005) Toward increasing erucic acid contentin oilseed rape (Brassica napus L.) through the combination withgenes for high oleic acid. J Am Oil Chem Soc 82:445–449
Scarth R, Tang J (2006) ModiWcation of Brassica oil using conven-tional and transgenic approaches. Crop Sci 46:1225–1236
Schierholt A, Rücker B, Becker HC (2001) Inheritance of high oleicacid mutations in winter oilseed rape (Brassica napus L.). CropSci 41:1444–1449
Sonntag NOV (1995) Industrial utilization of long-chain fatty acidsand their derivatives. In: Kimber DS, McGregor DI (eds) Brassicaoilseeds. CAB International, Oxon, pp 339–352
Stoutjesdijk PA, Singh SP, Liu Q, Hurlstone CJ, Waterhouse PA,Green AG (2002) hpRNA-mediated targeting of the ArabidopsisFAD2 gene gives highly eYcient and stable silencing. Plant Phys-iol 129:1723–1731
Stefansson BR, Hougen FW (1964) Selection of rape plants (Brassicanapus) with seed oil practically free from erucic acid. Can J PlantSci 44:359–364
Tillmann P (2007) http://www.vdlufa.de/nirs. (Site visited 24 June 2008)Utz HF (2007) PLABSTAT (Version 2N), a computer program for the
computation of variances and covariances. Institute of PlantBreeding, Seed Science, and Population Genetics, University ofHohenheim, Stuttgart, Germany, (http://www.uni-hohenheim.de/ipspwww/soft.html) (Site visited 24 June 2008)
Weier D, Hanke C, Eickelkamp A, Lühs W, Dettendorfer J, SchaVertE, Möllers C, Friedt W, Wolter FP, Frentzen M (1997) Trieru-coylglycerol biosynthesis in transgenic plants of rapeseed (Bras-sica napus L.). Fett/Lipid 99:160–165
Wilmer JA, Wallington EJ, Slabas AR (2001) Oil biosynthesis—Veryhigh erucic acid rapeseed, WO 03/033713
Wilmer JA, Wallington EJ, Slabas AR (2003) Very high erucic acidrape: a dream or reality. In: Proceedings of 11th Intl RapeseedCongress, 6–10 July 2003, Copenhagen, Denmark 2:583–585
Zhao J, Dimov Z, Becker HC, Ecke W, Möllers C (2008) Mapping QTLcontrolling fatty acid composition in a doubled haploid rapeseedpopulation segregating for oil content. Mol Breed 21:115–125
123