conditional mutagenesis in mice with heat shock promoter-driven cre transgenes
TRANSCRIPT

Conditional mutagenesis in mice with heat shock promoter-drivencre transgenes
Paula Dietrich,1 Ioannis Dragatsis,1 Shouhong Xuan,1 Scott Zeitlin,2 Argiris Efstratiadis 1
1Department of Genetics and Development, Columbia University, Russ Berrie Medical Sciences Pavilion, 1150 St. Nicholas Ave.,New York, New York 10032, USA2Department of Pathology, Columbia University, 1150 St. Nicholas Ave., New York, New York 10032, USA
Received: 23 September 1999 / Accepted: 4 November 1999
Abstract. To explore the potential of a simple and rapid approachfor ubiquitous conditional gene disruption, we have generated Cre-producer mouse transgenic lines (Hs-cre1, 6and7) expressing arecombinase transgene (cre) from a heat shock gene promoter andtested their performance in Cre-mediated excision of target DNAin crosses with Cre-responder strains carryingloxP-modified al-leles of the genes encoding the Huntington’s disease gene homolog(Hdh), the epidermal growth factor receptor (Egfr), and the type 1insulin-like growth factor receptor (Igf1r). Analyses of progenypossessing various transgene/reporter combinations showed thatcre expression can occur without heat shock in early embryos, butthis constitutive transcription is stochastic and transgene depen-dent. Thus,Hs-cre1behaves predominantly as a “deleter” strain,since the majority of progeny (°70–85%) exhibit complete recom-bination, regardless of reporter locus. LinesHs-cre6andHs-cre7,however, function successfully as “mosaicking” strains because, inaddition to two extreme classes of progeny with 0% or 100%recombination, they generate an intermediate class of mosaics ex-hibiting various degrees of partial DNA excision. Notably, thefrequency of offspring in each class varies between reporters, butmosaic embryos are consistently obtained in adequate numbers(°30–60%). TheHs-cre6 transgene is also inducible and can beused to introduce mosaicism into adult tissues at preselected de-velopmental times by heat shock treatment of mice with 0% re-combination in tail DNA. By bypassing the lethality resulting fromsome gene knockouts, mosaic embryos and mice make particularmutational analyses possible and are also very useful for the iden-tification of cell lineage-specific gene functions.
Introduction
Targeted mutagenesis in mice (reviewed by Mu¨ller 1999) allowsthe study of mammalian gene functions in the context of the entireexperimental organism by establishing causal relationships be-tween defined null mutations and their phenotypic consequences.This powerful approach, however, reveals only indispensablefunctions because apparently opportunistic compensation by othergenes with overlapping activities sometimes precludes the appear-ance of a phenotype (see Doetschman 1999; Mu¨ ller 1999).Whereas this inherent problem is intractable, advances in method-ology could make manageable other limitations of gene knockouts.For example, a potential drawback, especially for genes partici-pating in interacting developmental pathways, is the manifestationof complex pleiotropic effects that are difficult to analyze. More-over, whenever nullizygosity results in embryonic lethality, astudy of gene function at more advanced developmental stages is
precluded (Copp 1995). To obviate such limitations, the develop-ment of various strategies for spatial or temporal conditional mu-tagenesis is continuing (Rajewski et al. 1996; Marth 1996; Lobeand Nagy 1998; Rossant and McMahon 1999), by using predomi-nantly the cre/loxP site-specific recombination system (Sauer1998).
The cre gene of phage P1 encodes a recombinase (Cre) thatcatalyzes DNA recombination between pairs of specific 34-bp se-quences (loxP sites). Intramolecular Cre-mediated recombinationbetween twoloxP sites in direct (“head-to-tail”) orientation resultsin excision of the intervening DNA segment. In practice, therefore,conditional mutagenesis requires crosses between Cre-producingand Cre-responding strains of mice. Producers carry a fusion trans-gene expressing Cre recombinase under the direction of a suitablepromoter, whereas in responders a critical DNA segment of anendogenous locus (to be deleted for elimination of function) hasbeen modified by conventional gene targeting and is flanked bydirect loxP sites (“floxed”), which, as positioned, do not interferewith normal gene activity. Accordingly, the type of conditionalmutagenesis that is desirable in any particular case is determinedby the choice of the promoter driving thecre transgene.
As in the case of a conventional gene knockout, conditionalmutagenesis results in irreversible loss-of-function by generating anull allele. However, the phenotypic manifestations are differentand depend on the controlled mutagenic event that can be plannedto occur with tissue specificity or at a preselected developmentaltime or both. The utility of this approach can be further enhancedby using inducible promoters to drivecre expression. A crucialelement in this regard is the environmental stimulus or signalingmolecule which, acting as a transcriptional switch, can controlpromoter function appropriately.
To increase the options that are currently available (see Dis-cussion), we decided to explore the use of heat shock as a potentialalternative for inducible expression of Cre. In mice, two of the heatshock proteins of the HSP70 family are encoded by loci (Hsp70-1and Hsp70-3) that are ubiquitously induced by stress (Morange1999). These two linked genes (Gu¨nther and Walter 1994) are 99%identical in sequence for a stretch of 2483 bp that includes apromoter region extending for 289 bp upstream from the site oftranscriptional initiation (GenBank Accession No. AF109906).Thus, we used theHsp70-1gene promoter for our purposes, de-spite uncertainties about the feasibility of this approach stemmingfrom the information that there is constitutive expression ofHsp70in two-cell stage mouse embryos, which disappears at the four-cellstage and reappears at a low level at the eight-cell stage (Hahnel etal. 1986; Mezger et al. 1994; Bevilacqua et al. 1995; Christians etal. 1995). After this time, however, only expression inducible byheat shock can occur (Christians et al. 1995). Regardless of themechanism of constitutive promoter activation, our primary con-cern was the response of floxed reporter loci. According to oneCorrespondence to:A. Efstratiadis
Mammalian Genome 11, 196–205 (2000).
© Springer-Verlag New York Inc. 2000
Incorporating Mouse Genome

report (Kothary et al. 1989), inducible expression of alacZ trans-gene driven by theHsp70-3promoter was observed postimplan-tation, but constitutive transcription at the two- or four-cell stageor in blastocysts was not detected. On the other hand, eliminationof some prokaryotic sequences present in the initial construct re-sulted in constitutive expression of a modifiedlacZ transgene atthe two- and eight-cell stages (Bevilacqua et al. 1995). Similarresults were obtained with in vitro cultured embryos carrying aluciferase transgene driven by theHsp70-1promoter, but eight-cellembryos were not examined (Christians et al. 1995). Culturing,however, apparently influenced the transcriptional performance ina non-physiological manner, because luciferase assays with em-bryos that were not exposed to the in vitro conditions showed alevel of transgene expression that was only 5.5%, 9%, or 20% ofthe in vitro value at the two-cell stage, depending on genetic back-ground (Christians et al. 1995, 1997). Because these and otheranalogous data are still heterogeneous and fragmented, we decidedto test, first, whether it is at all feasible to use theHsp70-1pro-moter for induciblecre expression, without making attempts tooptimize the system. Here we present results demonstrating that,within some limitations, transgenic lines expressingcre under thedirection of heat shock regulatory elements can be used success-fully for generation and mutational analysis of mosaic embryosand mice.
Materials and methods
Generation of cre transgenic mice.Founders ofcre transgenic lineswere generated by standard procedures, after injecting into fertilized eggs(derived from C57BL/6J×CBA crosses) a∼2.4-kb NotI-SalI DNA frag-ment (Fig. 1A). The injected DNA consisted of a promoter-containingsegment from a mouse heat-shock gene (655 bp) and a modifiedcre genesequence (1.6 kb). The heat-shock component represented 536 bp of 58flanking region and the first 119 (of 225) bp of the 58 noncoding region ofthe mouseHsp70-1gene (Hunt and Calderwood 1990) that was obtainedby PCR amplification of 129/Sv/Ev mouse genomic DNA with the primers58-GGAGACATGGACAAGCAAGCATTC-38 (forward) and 58-TGGAT-GCTCTGGGGAAGGCTGGT-38 (reverse). The source of the modifiedcrecoding region, which included a nuclear localization signal and poly(A)addition sequence, was plasmid pMC-Cre (Gu et al. 1993). In most of ourexperiments, we used three of the transgenic lines that we generated, des-ignatedHs-cre1, 6and 7. Transgenic mice were identified by Southernanalysis ofEcoRI-digested DNA from tail biopsies, by using as a probe a0.8-kb EcoRI-ClaI fragment from the transgenic construct correspondingto a portion of thecre coding sequence (Fig. 1A). In linesHs-cre6and7,each containing only one copy of the transgene, single hybridizing frag-ments of 8.5 and 5.5 kb, respectively, were identified, whereas the presenceof two tandem integrations of the transgene in lineHs-cre1was revealedfrom the detection of two fragments (2.4 and 7.5 kb) recognized by thecreprobe.
Determination of Cre-mediated recombination.To ascertain the ef-ficiency of Cre-mediated recombination, theHs-cre transgenic lines werecrossed with three reporter strains that will be described in detail else-where. These Cre-responding mice carryloxP-modified alleles of the genesencoding the Huntington’s disease gene homolog (Hdh), the epidermalgrowth factor receptor (Egfr), and the type 1 insulin-like growth factorreceptor (Igf1r). The floxed segments of the reporter loci are theHdhpromoter and exon 1, theEgfr exons 9 and 10, and theIgf1r exon 3 (Fig.1B–D). A third loxP site located next to the 38 end of the neomycinselection cassette (neo) is present in theIgf1r reporter allele (Fig. 1D). Inthis particular case, “marker eviction” (see Discussion) was unnecessarybecause the selectable marker did not interfere with expression (that is, thefloxed allele exhibited wild-type behavior). Thus, in contrast to the neo-natal lethality ofIgf1r nullizygous mutants (Liu et al. 1993),Igf1r(flox/−)mice were viable. The floxedHdh allele was also unaffected by the pres-ence of theneocassette, whereas floxing at theEgfr locus resulted in thegeneration of a null allele.
The degree of Cre-mediated DNA excision (“excision index”; EI) wasquantitated by Southern analysis of DNA that was isolated from micecarrying Hdh, Egfr, and Igf1r floxed alleles and digested withBamHI,EcoRV, andKpnI, respectively (see Fig. 1B–D). The level of radioactivityin diagnostic restriction fragments of the floxed (“flox”) and recombined(“Dflox”) alleles that were hybridized with appropriate probes was deter-mined with a PhosphorImager (Molecular Dynamics). After correction forbackground hybridization, the percentage of recombination observed withthe Hdh andEgfr reporter alleles was calculated as: EI4 [Dflox/(flox +Dflox)] × 100. For specimens withIgf1r(lox/−) genotype, our calculationswere based on the consideration that, if recombination had not occurred,the radioactivity of the fragments representing the null (n) and floxed (f)alleles should be equal. Thus, the percentage of recombination was calcu-lated from the reduction of the level of hybridization in the floxed fragmentas: EI4 [(n − f)/n] × 100.
To examine histochemically the results of recombination monitoredthrough the expression of alacZ sequence activated in theEgfr reporterupon DNA excision, we stained frozen tissue sections forb-galactosidaseactivity, as described (Hogan et al. 1994).
PCR analyses.For routine genotyping of progeny, thecre transgene wasdetected by PCR (45 s at 94°C, 45 s at 61°C, and 1 min at 72°C, for 30cycles) with the primers 58-CTGCCACGACCAAGTGACAGC-38 (for-ward) and 58-CTTCTCTACACCTGCGGTGCT-38 (reverse) to generate a324-bp product corresponding to a portion of thecre coding region. Thesame primers and conditions were used for amplification of cDNA inRT-PCR analyses. The sources of total RNA template for these assays werenine pooled two-cell stage embryos, individual embryos (embryonic day14.5; E14.5) or dissected adult tissue. RNA was isolated with the Trizolreagent (Gibco-BRL) according to the manufacturer’s instructions and thentreated with RQ1 RNase-free DNase I (Promega), to eliminate contami-nating DNA. cDNA was synthesized from the entire RNA preparationderived from the pooled two-cell embryos or from 2mg aliquots of RNAprepared from advanced embryos or tissues using random hexamer primersand Superscript II reverse transcriptase (Gibco-BRL), and then treated withE. coli RNase H. In control reactions, addition of reverse transcriptase wasomitted. Aliquots of the cDNA were then amplified in PCR reactions eitherfor cre,as described above, or for actin (1 min at 94°C and 2 min at 72°C,for 25 cycles) with the primers 58-GACAACGGCTCCGGCATGTG-CAAAG-38 (forward) and 58-TTCACGGTTGGCCTTAGGGTTCAGGG-38 (reverse) to generate a 320-bp product. One-third of each PCR reactionwas fractionated on a 3% NuSieve agarose (FMC Corporation) gel.
For lacZ andneosequence amplification, individual E3.5 blastocystswere lysed by boiling for 1 min in 10 mM Tris pH 8.0, 1 mM EDTAcontaining 0.4% NP40, and approximately one half of the lysate was usedfor each PCR reaction (45 s at 94°C, 45 s at 61°C, and 1 min at 72°C, for30 cycles) by simultaneous use of two pairs of primers: 58-CGGC-GGTGAAATTATCGATGAG-38 (lacZ forward) and 58-ATCAGCA-GGATATCCTGCACCA-38 (lacZ reverse), generating a 315-bp product;58-GCCCGGCATTCTGCACGCTT-38 (neo forward; Pgk promoter se-quence), and 58-AGAGCAGCCGATTGTCTGTTGT-38 (neo reverse),generating a 192-bp product. Half of each PCR reaction was fractionatedon a 3% NuSieve agarose gel.
For semiquantitative PCR analysis of blastocyst DNA with 20% ofeach lysate, the number of cycles was limited to 23 (i.e., kept within thelogarithmic amplification phase). One-third of each PCR reaction wasfractionated on a 3% NuSieve agarose gel and analyzed by Southern blot-ting by using as probes the 315-bplacZ and the 192-bpneoPCR products.The degree of recombination was calculated from the ratio of radioactivity(counted with a PhosphorImager) that was hybridized to thelacZ andneoPCR products.
Heat shock of mice.An empty mouse cage, together with a tray filledwith water, was placed inside a hybridization oven (Model 400, RobbinsScientific) adjusted to 42°C for at least 2 h prior to treatment (a high levelof humidity is required for efficient heat shock; Fujio et al. 1987; Nowaket al. 1990). At the end of this time period, a relative humidity level of 75%was achieved, as monitored with a hygrometer. For heat shock treatment,adult mice (25–35 g) were placed in the prewarmed cage inside the hu-midified oven for 17 min, and then transferred to a clean cage and allowedto recover at room temperature. Pregnant females (E14.5) received thesame treatment for 13–15 min, to apply heat shock to embryos.
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes 197

The conditions of this protocol were determined empirically by varyingboth the time and temperature of heat shock (data not shown). Prolongingthe heat shock treatment or using temperatures above 42°C were found tobe unsuitable conditions, as they affected variably the viability of adultmice. In contrast, although the efficiency of induction is not the highest
attainable, the mice respond well to our standardized protocol (they appearto be lethargic for∼20 min or occasionally longer, but return to normalafter this recovery period). Moreover, abnormalities in embryos were neverobserved by exposing pregnant females to the chosen experimental con-ditions.
<
Fig. 1. Restriction maps of thecre transgenic construct and floxed reporter loci. Therestriction sites are:NotI (N), EcoRI (R), BamHI (B), ClaI (C), SalI (S), EcoRV(RV), andKpnI (K). (A) Diagram of the DNA fragment that was injected to generatemouse lines carrying acre transgene driven by a heat shock promoter. The compo-nents of this construct are described in Materials and methods. The transcriptionalorientation (arrow) and the probe used for Southern analyses (black rectangle) areindicated.(B–D) The diagrams represent regions of the wild-type (wt), floxed (flox),and recombined (D) alleles of theHdh, Egfr and Igf1r reporter loci. The exonspresent in the displayed regions (open numbered boxes), the selection marker cas-settes (neo; dark gray rectangles), the transcriptional orientations (arrows), and thepositions and orientations ofloxP sites (gray triangles) are indicated. The sizes ofdiagnostic restriction fragments and the probes used for Southern analyses (blackrectangles) are shown. In(C), the CMV IE1 promoter/enhancer and the lacZ reporter(modified to include a nuclear localization signal; nls) that are present in the floxedEgfr locus are shown (light gray rectangles). Also shown is a transcriptional/translational STOP sequence (crossed rectangle) consisting of the SV40 early poly-adenylation signal, a false translational initiation signal, and a 58 splice site (Laksoet al. 1992). In(D), the different products of Cre-mediated DNA excision [D(1), D(2)and D(3)] generated from all possible pairwise combinations of threeloxP sitespresent in the floxedIgf1r allele are indicated. A null allele (ko) generated by directIgf1r gene knockout is also shown.
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes198

Results
Expression of cre in the absence of heat shock.In the breedingprogram that we pursued, we crossed mice from one of the Cre-producing transgenic linesHs-cre1, 6,or 7 with animals from oneof the three reporter strains with floxedHdh, Egfror Igf1r alleles(see Materials and methods), and examined progeny carrying par-ticular transgene/reporter combinations. Preliminary analyses ofDNA excision indices in tail biopsies showed various degrees ofrecombination in the reporter loci, from which we inferred thatcreexpression was leaky and had occurred without heat shock.
To confirm that the observed recombination was due to con-stitutive expression of thecre transgene during very early embryo-genesis, as might have been expected (see Introduction), we per-formed PCR assays (Fig. 2). First, we usedcre-specific primersand RNA template from two-cell embryos (of theHS-cre1 trans-genic line, in the example shown in Fig. 2A), and synthesizedRT-PCR products that were analyzed by Southern blotting with acre cDNA probe. The results showed thatcre transcripts wereindeed present at this early stage (Fig. 2A).
Action of Cre in the absence of heat shock.Next, we sought todemonstrate directly the functional consequences of the presum-ably constitutive early transcription ofcre transgenes, that is, theoccurrence of Cre-mediated recombination. For this purpose, how-ever, instead of collecting the required large number of two-cellembryos to obtain amounts of DNA template adequate for PCRanalysis, we used DNA extracted from blastocysts. In the dis-
played examples, for whichEgfr was used as the reporter locus(Fig. 2B and C), aliquots of DNA from specimens genotyped inadvance were selected for simultaneous amplification oflacZ andneo sequences (located outside and inside the floxed region, re-spectively; Fig. 1C) with two sets of corresponding primers. In acontrolEgfr(lox/+) blastocyst lacking Cre activity, which was de-rived by crossing anEgfr(lox/+) female with a wild-type male,PCR products were obtained as expected from both thelacZ andthe floxed, but non-recombinedneosequence of the targeted allele(Fig. 2B, lane 4). In contrast, in twoEgfr(lox/+) blastocysts ob-tained from anEgfr(lox/+) female that was mated with a malehomozygous for theHs-cre1 transgene, only thelacZ sequencewas amplified (Fig. 2B, lanes 2 and 3). From these results, weinferred that complete (100%) Cre-mediated excision of the floxedneo segment had occurred, thus precluding amplification of thissequence. The same result (PCR amplification oflacZ, but not ofneo sequence) was obtained by using, as an additional control,DNA from a tail biopsy (Fig. 2B, lane 5), in which 100% recom-bination had occurred, as previously shown by Southern analysis.
Analogous results, which, however, differed in the level ofrecombination attained, were obtained with blastocysts from fe-males of the same genotype [Egfr(lox/+)] that were crossed withmales homozygous for theHs-cre6 transgene. In this case, PCRproducts obtained after a limited number of amplification cycleswere analyzed by Southern blotting withlacZandneoprobes (Fig.2C), and then quantitated using a PhosphorImager. After normal-ization of theneo readings by considering that thelacZ resultsshould be invariable between samples, we showed that the calcu-lated excision indices were 51% (lane 3), 67% (lane 4), and 100%(lane 5), whereas no recombination had occurred (0%; lane 2)when the blastocyst was derived after mating with a wild-typemale partner (absence ofcre expression). When genomic (tail)DNA, already known by direct Southern analysis to be recombinedto a level of 50% (lane 6), was used as a control, the results wereidentical to those in Fig. 2C, lane 3.
Strain-dependent DNA excision.For a more detailed overview ofthe behavior of particular transgene/reporter combinations in re-gard to recombination, we determined by Southern analysis theDNA excision indices in a large number of E12.5-E16.5 embryosand in tail biopsies examined predominantly at postnatal day 10(P10). We then classified the specimens into three categories, asshown in Table 1, by placing those that exhibited either 0% or100% recombination in two extreme classes, while lumping theremaining specimens with any degree of partial recombination ina third (intermediate) class.
From these results (Table 1), it became immediately obviousthat HS-cre1 had behaved as a “deleter” strain (see Discussion),consistent with the PCR data from the analysis of blastocysts de-
Fig. 2. RT-PCR assays in early embryos.(A) Analysis ofcre expressionat the two-cell stage. Total RNA was extracted from nine pooled two-cellstage embryos carrying theHs-cre1transgene and used as a template forcDNA synthesis by reverse transcriptase (RT) in the presence of randomhexamer primers. In a parallel control reaction, the enzyme was omitted.PCR amplification products of the cDNA and control reactions (+RT and−RT, respectively) withcre-specific primers were analyzed by Southernblotting with acreprobe (see Materials and methods and Fig. 1A). The sizeof the cre PCR product (in bp) is indicated.(B) Cre-mediated DNA exci-sion assayed in blastocysts. DNA from blastocysts with anEgfr(flox/+)/Hs-cre1genotype was amplified by PCR with simultaneously two sets ofprimers specific forneo and lacZ (lanes 2 and 3). The control reactionswere: no DNA (lane 1), DNA from anEgfr(flox/+) blastocyst (lane 4; 0%recombination); and tail DNA control preselected for 100% recombination(lane 5). The size markers (lane M) areHaeIII fragments offX174 DNA.The positions of theneoandlacZ PCR products are indicated.(C) Differ-ent levels of Cre-mediated DNA excision assayed at the blastocyst stage.DNA was prepared from anEgfr(flox/+) blastocyst (lane 2, control; 0%recombination) or from blastocysts with anEgfr(flox/+)/Hs-cre6genotype(lanes 3–5) and used for semiquantitative PCR performed for 23 cyclesonly in the presence ofneoandlacZ primers (as described in B). Reactionproducts were analyzed by Southern blotting withneoandlacZprobes. Forcontrols, PCR reactions were performed without DNA template (lane 1)and with tail DNA (50% recombination, preselected; lane 6). The sizemarkers (lane M) are32P-end-labeledMspI fragments of pBR322 DNA.The positions of theneoand lacZ PCR products are indicated.
Table 1. Number (%) of specimens in each of three classes exhibiting the indicateddegree of reporter constructcre-mediated recombination.
HScreline
Reporterconstruct
Specimensexamined
% Recombination
0% Intermediatea 100%
6 Egfr embryos (E13.5) 3 (17.7) 10 (58.8) 4 (23.5)6 Hdh embryos (E16.5) 0 (0) 11 (40) 16 (59.3)7 Hdh embryos (E12.5) 4 (13.8) 16 (55.2) 9 (31.0)1 Egfr tails (P10) 0 (0) 6 (27.3) 16 (72.7)1 Hdh tails (P10) 0 (0) 9 (15.0) 51 (85.0)1 Igf1r tails (P10) 3 (9.7) 7 (22.6) 21 (67.7)6 Egfr tails (P10) 8 (18.2) 21 (47.7) 15 (34.1)6 Hdh tails (P10) 0 (0) 20 (31.7) 43 (68.3)6 Igf1r tails (P10) 20 (37.0) 24 (44.4) 10 (18.5)7 Hdh tails (P10) 8 (26.6) 11 (36.7) 11 (36.7)
a % recombination in the range 5–88%.Genotypes wereflox/+, but some of theIgf1r animals wereflox/−. Among the latter,those with 100% recombination died at birth (genotyped at P0).
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes 199

scribed above (Fig. 2B). In this regard, all three reporter lociapparently responded in a very similar manner to the recombinaseproduced by theHs-cre1 transgene. Thus, escape from recombi-nation either was not observed (Egfr and Hdh reporters) or oc-curred in very few animals (∼10%; Igf1r reporter), whereas thevast majority of specimens (2/3 or more) exhibited complete ex-cision (100% category), with the remaining falling into the inter-mediate class.
The results with theHs-cre6line, on the other hand, revealedan interesting interplay between this particular Cre-producingtransgene and each responding floxed locus. Thus, less than 1/5 ofthe Igf1r tails exhibited an excision index of 100%, while themajority of specimens were almost equally divided between the0% class and the intermediate class. In contrast, the response of thefloxed Hdh locus to theHs-cre6transgene did not differ signifi-cantly from that to theHs-cre1transgene (complete escape fromrecombination was not observed in embryos or tails). Finally, withthe Egfr reporter, the frequency distribution of specimens (em-bryos and tails) in the three recombination categories was inter-mediate between theIgf1r and Hdh patterns. Thus, escape fromrecombination was observed, but in∼twofold fewer specimensthan in theIgf1r case, while the occurrence of partial rather thancomplete recombination was found to be more frequent. The con-clusion that the frequency distributions depend on both compo-nents of a transgene/reporter combination was further supported bythe values of excision indices detected inHdhspecimens (embryosand tails) in the presence of theHs-cre7transgene. In contrast tothe other two transgenic lines, escape of floxedHdh from recom-bination did occur in this case, albeit in small numbers, whereasthe 100% class ceased to predominate.
Generation of mosaic progeny.The most important observation ofpractical significance (Table 1) is that, with all reporters examined,theHs-cre6line can yield a variable (∼30–60%), but neverthelesssignificant fraction of animals with partial excision indices cover-ing almost the entire range between 0% and 100% recombination,which are of course the most useful for mosaic analysis (see Dis-cussion). TheHs-cre7line is equally useful for the same purpose,but has other limitations (see below).
Tissue mosaicism in regard to recombination levels was vali-dated by determining DNA excision indices in various organsdissected from∼P30 animals. As the presented examples of thisanalysis indicate (Fig. 3), there is variability in excision indicesbetween animals carrying the same transgene/reporter combina-tion, not only in the tail, but in all examined tissues. When eachanimal is considered individually, however, the tail index appearsto be quite representative of the indices in other tissues, althoughan exact matching of values was not observed. In this regard, theaverage deviation (expressed as % increase or decrease) from theexcision index of the tail, considered as a reference value, is∼16%for all animals and tissues shown (Fig. 3). Accordingly, the tailindex, which is easily determined without an invasive procedure,can be used for selection of experimental animals.
For direct visualization of mosaicism, we examined heart andkidney sections (Fig. 4) from preselected animals carrying theHs-cre6/Egfr transgene/reporter combination, which exhibited50% and 100% recombination levels in their tail DNA. The floxedEgfr locus is a convenient Cre-responder for this type of analysisbecause, upon Cre-mediated excision of the floxed DNA segment,a CMV promoter becomes juxtaposed to alacZ reporter sequencemodified to include a nuclear localization signal (see Fig. 1C) and,by driving its expression, allows histochemical detection ofb-ga-lactosidase activity (blue staining of cell nuclei). By counting num-bers of stained and unstained nuclei in different optical fields, wecalculated average percentages of stained nuclei in the tissue sec-tions and found that their ratio between the 50% and 100% speci-mens was close to 1:2 in each case (see Fig. 4). Thus, there is a
correlation between the percentage of stained cells and the exci-sion index in each tissue.
Heat shock-inducible cre expression.For inducible expression ofCre recombinase, we first used wild-type mice to establish appro-priate conditions for heat shock treatment (see Materials and meth-ods), and then applied a standardized protocol to animals carryingcre transgenes. Examples of our results from such experiments arepresented in Fig. 5. As expected, the heat shock response wasrapid, and transcription from the endogenousHsp70 loci, whichcould be easily monitored by Northern analysis, was induced inembryos after a 15-min application of thermal stress to pregnantfemales (Fig. 5A). Analogous specific induction of the exogenousHsp70-1promoter of theHs-cre1transgene was also observed inembryos (Fig. 5B), butcre transcription occurred at a level thatwas significantly lower in comparison with the endogenous locus.Thus, although it was detectable by Northern analysis, monitoring
Fig. 3. Assays of Cre-mediated recombination levels in mouse organs.(A)The DNA excision indices in various organs of mice carrying the indicatedcre transgene/reporter combinations are shown. Each symbol correspondsto one individual.(B, C) Examples of the data from Southern analyses ofDNA prepared from organs, to determine excision indices. The results in(B) are from a mosaicHdh(flox/+) female carrying theHs-cre6transgene,whereas those in(C) are from a mosaicHdh(flox/+) male carrying theHs-cre7 transgene. The positions of theBamHI DNA fragments corre-sponding to the floxed (flox) and recombined (D) alleles (see Fig. 1B) areindicated.
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes200

by RT-PCR was more convenient. A similar difference in amountsbetween endogenous and exogenous induced transcripts, estimatedfrom the results of Northern analysis to be as high as 30-fold, wasdetected in adult tissues ofHs-cre6mice after heat shock (Fig.5C). Moreover, despite some correlations (both types of transcriptswere most abundant in the lung, for example), there was differen-tial variability in the relative levels ofHsp70andcre transcriptionproducts between the examined tissues. Nevertheless, RT-PCRassays demonstrated thatcre was expressed in all of these tissuesafter heat shock (Fig. 5D).
It was previously shown that the transient heat shock-inducedtranscriptional output from the loci encoding Hsp70 proteins ortheir transactivators differs in magnitude and kinetics between tis-sues (Nowak et al. 1990; Blake et al. 1990; Fiorenza et al. 1995;Flanagan et al. 1995). Thus, variability in the response of theexogenous promoter is not surprising, although the exact reasonsfor an overall lack of correspondence to the behavior of the en-dogenous loci are unclear. It is likely that transgene position ef-fects contribute to this difference (see also Discussion). In addi-tion, however, we think that our results are by necessity unable torepresent accurately the peaks of expression known to occur asyn-chronously in different tissues, as the mice were sacrificed 1 h afterheat shock, to ensure that the earliest transcriptional responses willnot be missed. We note that during the course of these experimentsaiming to the characterization of the system, we also demonstratedthat, in contrast to the transgenes in linesHs-cre1andHs-cre6,thetransgene in theHs-cre7line is not inducible (not shown).
Heat shock-inducible Cre-mediated recombination.Next, we useda functional assay to determine whether heat shock-induced Creexpression were mediating recombination of a reporter allele, andalso if it were feasible to apply multiple treatments to increase theefficiency of DNA excision. To address these questions, we firstperformed a pilot experiment using an animal at P30 that wascarrying theHs-cre6/floxed Hdh transgene/reporter combination.This mouse was heat shocked for 17 min at 42°C, and this treat-ment was repeated 24 h later. A small portion of tail was removed
prior to heat shock (control) and 24 h after each of the heat shocktreatments. Southern analysis of DNA isolated from the tail speci-mens showed that, in the absence of heat shock, the excision indexof the reporter allele was 36% and was increased to 58% and 73%after the first and second rounds of heat shock, respectively (datanot shown).
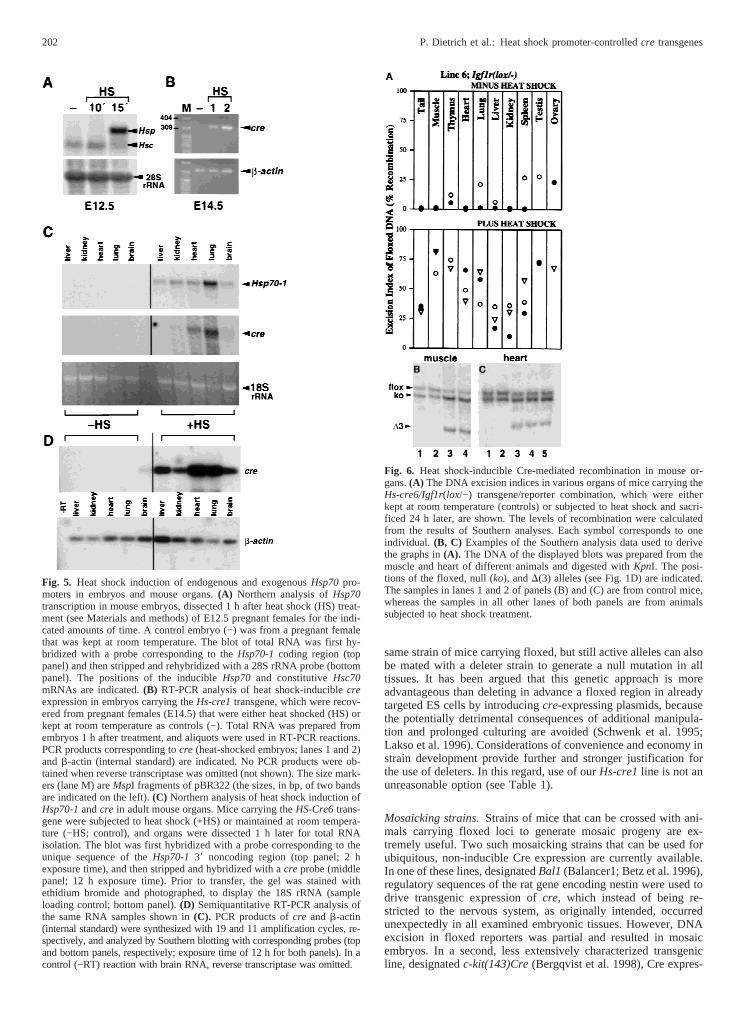
Analogous results demonstrating that the efficiency of recom-bination increases with multiple heat shock treatments were alsoobtained with many mice carrying theHs-cre6/floxed Igf1r trans-gene/reporter combination (not shown). Thus, two consecutiveheat shock treatments separated by a 24-h recovery period wereadministered to such mice at P30, to determine the level of induc-ible recombination in various tissues. These animals, and alsocontrol mice that were not subjected to treatment, had exhibited norecombination in tail DNA when previously examined at P10. Theresults of these experiments, including examples of Southernanalyses of DNA prepared from two of the examined tissues(muscle and heart), are shown in Fig. 6. As the data from controlanimals indicate, recombination in the absence of heat shock, ap-parently due to stochastic leakiness of thecre transgene, occurredonly occasionally in some of the tissues, and the excision index,whenever it was significant, did not exceed a level of 25%. Incontrast, after the application of heat shock treatment, inefficientrecombination (excision index#25%) was a rare occurrence, andmoderate or high excision indices were observed that differedbetween tissues, although they frequently tended to fall within arelatively narrow range between animals.
Discussion
Advances and problems in conditional mutagenesis.Despite sev-eral successful applications, streamlined protocols for Cre-mediated conditional mutagenesis in mice are still largely lacking,and efforts for further refinements are continuing (Rossant andMcMahon 1999). Most of the problems that remain unsolved toone degree or another are related to Cre-expression, whereas thefloxing of responder loci is a rather straightforward process (seeSauer 1998).
Depending on the pattern ofcre expression, conditional muta-genesis can be either ubiquitous or tissue-specific, and each ofthese types can be either inducible or non-inducible. If the systemallows for induction, a developmental time for the occurrence ofgene inactivation can also be selected. In this regard, a transcrip-tional switch used for induction should ideally be able to conferprecise, rapid, and complete promoter activation (Spencer 1996;Porter 1998).
Unfortunately, not all of these criteria are fulfilled with thecurrently available inducible systems (see below). Thus far, thebest reported results of conditional mutagenesis were obtainedwithout induction, by driving Cre expression with tissue-specificpromoters (see, for example, Kulkarni et al. 1999). Nevertheless,even in these cases, the frequency of in vivo excision, althoughhigh at times (∼80% or more), tended to be variable.
Deleter strains.In addition to purposely derived deleter strains ofmice (Lakso et al. 1996), other ubiquitous deleters were generatedfortuitously from cre transgenes that were designed to be ex-pressed with tissue specificity, but were transcribed against expec-tation in early embryos (Schwenk et al. 1995).
A useful application of deleter strains is, for example, theeviction of floxed selection markers from mutated loci targeted forintroduction of subtle changes in coding regions, to prevent po-tential interference with normal transcription (Lakso et al. 1996;Sauer 1998). It is expected, however, that deleters will be usedmore often in the generation of conventional gene knockouts fromcorresponding floxed versions of various loci (Schwenk et al.1995). Thus, in addition to conditional mutagenesis of choice, the
Fig. 4. X-gal staining of cell nuclei in frozen sections of the hearts andkidneys ofEgfr(flox/+) mice carrying theHs-cre6 transgene, which hadexhibited 50% or 100% recombination of the reporter in tail DNA. Thepercent values presented at the bottom are averages (± standard error) ofratios (×100) of stained nuclei (black color in the photographs) over thetotal number of nuclei (stained and unstained) that were counted in threeoptical fields from each section. The ratios of percentages in correspondingsections from the specimens with 50% and 100% tail excision indices arealso shown.
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes 201
same strain of mice carrying floxed, but still active alleles can alsobe mated with a deleter strain to generate a null mutation in alltissues. It has been argued that this genetic approach is moreadvantageous than deleting in advance a floxed region in alreadytargeted ES cells by introducingcre-expressing plasmids, becausethe potentially detrimental consequences of additional manipula-tion and prolonged culturing are avoided (Schwenk et al. 1995;Lakso et al. 1996). Considerations of convenience and economy instrain development provide further and stronger justification forthe use of deleters. In this regard, use of ourHs-cre1line is not anunreasonable option (see Table 1).
Mosaicking strains.Strains of mice that can be crossed with ani-mals carrying floxed loci to generate mosaic progeny are ex-tremely useful. Two such mosaicking strains that can be used forubiquitous, non-inducible Cre expression are currently available.In one of these lines, designatedBal1 (Balancer1; Betz et al. 1996),regulatory sequences of the rat gene encoding nestin were used todrive transgenic expression ofcre, which instead of being re-stricted to the nervous system, as originally intended, occurredunexpectedly in all examined embryonic tissues. However, DNAexcision in floxed reporters was partial and resulted in mosaicembryos. In a second, less extensively characterized transgenicline, designatedc-kit(143)Cre(Bergqvist et al. 1998), Cre expres-
Fig. 5. Heat shock induction of endogenous and exogenousHsp70pro-moters in embryos and mouse organs.(A) Northern analysis ofHsp70transcription in mouse embryos, dissected 1 h after heat shock (HS) treat-ment (see Materials and methods) of E12.5 pregnant females for the indi-cated amounts of time. A control embryo (−) was from a pregnant femalethat was kept at room temperature. The blot of total RNA was first hy-bridized with a probe corresponding to theHsp70-1coding region (toppanel) and then stripped and rehybridized with a 28S rRNA probe (bottompanel). The positions of the inducibleHsp70 and constitutiveHsc70mRNAs are indicated.(B) RT-PCR analysis of heat shock-induciblecreexpression in embryos carrying theHs-cre1transgene, which were recov-ered from pregnant females (E14.5) that were either heat shocked (HS) orkept at room temperature as controls (−). Total RNA was prepared fromembryos 1 h after treatment, and aliquots were used in RT-PCR reactions.PCR products corresponding tocre (heat-shocked embryos; lanes 1 and 2)and b-actin (internal standard) are indicated. No PCR products were ob-tained when reverse transcriptase was omitted (not shown). The size mark-ers (lane M) areMspI fragments of pBR322 (the sizes, in bp, of two bandsare indicated on the left).(C) Northern analysis of heat shock induction ofHsp70-1andcre in adult mouse organs. Mice carrying theHS-Cre6trans-gene were subjected to heat shock (+HS) or maintained at room tempera-ture (−HS; control), and organs were dissected 1 h later for total RNAisolation. The blot was first hybridized with a probe corresponding to theunique sequence of theHsp70-1 38 noncoding region (top panel; 2 hexposure time), and then stripped and hybridized with acre probe (middlepanel; 12 h exposure time). Prior to transfer, the gel was stained withethidium bromide and photographed, to display the 18S rRNA (sampleloading control; bottom panel).(D) Semiquantitative RT-PCR analysis ofthe same RNA samples shown in(C). PCR products ofcre and b-actin(internal standard) were synthesized with 19 and 11 amplification cycles, re-spectively, and analyzed by Southern blotting with corresponding probes (topand bottom panels, respectively; exposure time of 12 h for both panels). In acontrol (−RT) reaction with brain RNA, reverse transcriptase was omitted.
Fig. 6. Heat shock-inducible Cre-mediated recombination in mouse or-gans.(A) The DNA excision indices in various organs of mice carrying theHs-cre6/Igf1r(lox/−) transgene/reporter combination, which were eitherkept at room temperature (controls) or subjected to heat shock and sacri-ficed 24 h later, are shown. The levels of recombination were calculatedfrom the results of Southern analyses. Each symbol corresponds to oneindividual. (B, C) Examples of the Southern analysis data used to derivethe graphs in(A). The DNA of the displayed blots was prepared from themuscle and heart of different animals and digested withKpnI. The posi-tions of the floxed, null (ko), andD(3) alleles (see Fig. 1D) are indicated.The samples in lanes 1 and 2 of panels (B) and (C) are from control mice,whereas the samples in all other lanes of both panels are from animalssubjected to heat shock treatment.
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes202

sion from the c-kit promoter presumably occurring early in em-bryogenesis resulted in reporter mosaicism in various tissues. OurHs-cre6line is a third mosaicking strain, sharing a common featurewith the other two: excision indices vary considerably betweenindividuals, while less variability is detected between tissues of aparticular animal.
Constitutive and inducible cre expression with the Hs-cre6 trans-genic line. Despite some limitations, theHs-cre6 line offers aunique advantage, as it can be used for inducible expression ofcreat a preselected prenatal or postnatal developmental time.
If need arises, the potential exists to induce the expression ofcre in Hs-cre6transgenic embryos, by subjecting pregnant femalesto heat shock at least as early as E12.5, since we did not observeany deleterious effects of hyperthermia (Edwards et al. 1995;Mirkes et al. 1999) by application of our protocol at this age.However, a fraction of embryos are destined to exhibit gradedlevels of DNA excision because of expression of Cre without heatshock. Thus, a spectrum of mosaic embryos, which can be ana-lyzed by terminating a number of pregnancies at chosen stages, isprovided automatically.
The inducibility of theHs-cre6 line is especially useful formosaic analysis postnatally, by applying heat shock at varioustimes forcre expression in animals with 0% excision indices ac-cording to tail tests. Depending on the particular floxed responderunder study, it may be useful on occasion to apply a second or eventhird round of heat shock, which, according to our experience, isfeasible (it does not jeopardize viability) and can increase signifi-cantly tissue excision indices.
Comparison of available strains for inducible cre expression.Al-though it cannot be used for tissue-specificcre expression, theHs-cre6 transgenic line offers several advantages for ubiquitousconditional mutagenesis, in comparison with analogous induciblesystems. Among them, the tetracycline-controlled system (Gossenand Bujard 1992; Gossen et al. 1995; Kistner et al. 1996), stillbeing modified for optimization (Baron et al. 1997; Mansuy et al.1998), can be used to inducecre expression at a preselected time.Depending on the experimental design, this can be achieved eitherby withdrawal (tTA system) or by administration (rtTA system) oftetracycline (tet) or its analog doxycycline (dox). There are alsosome alternative approaches. For example, theMx1 promoter canbe used to drive expression ofcre in the presence of interferon(Kuhn et al. 1995), while in other systems induction of recombi-nation is based upon the activation of chimeric proteins, which arecomposed of a fusion between Cre and a mutant form of theligand-binding domain of the estrogen or progesterone receptoractivated by the addition of tamoxifen (Feil et al. 1996; Danielianet al. 1998) or RU486 (Kellendonk et al. 1999), respectively.
Although the tet system is currently at the most advanced stageof development, its use with mice has not been consistently suc-cessful (see St-Onge et al. 1996). In fact, an ideal system does notexist at present and the goal of generating conditionally a truly nullmutation in all tissues, that is, altering the genotype in exactly thesame way as with a conventional gene knockout but at a particulardevelopmental time, has not been accomplished yet. Nevertheless,as a variation on a common principle, each of the available pro-cedures, including the heat shock system presented here, offers analternative strategy that can be chosen whenever it fits a particularexperimental design.
An advantage of the heat shock approach is that it makespossible the ubiquitous expression of Cre in very early embryos.This has not been achieved yet with the tet system, because the useof different promoters to drive the expression of tTA or rtTA hasnot been explored extensively. Another difference between theheat shock and other inducible systems is that, in comparison withthe considerable delay for in vivo response after removal or ad-
ministration of dox or other drugs (Mansuy et al. 1998), the effectof heat shock is very rapid. Furthermore, the problem of poten-tially deleterious effects on cells exerted, for example, by excesstTA and rtTA (Baron et al. 1997) or high doses of tamoxifenanalogs (Danielian et al. 1998; Vasioukhin et al. 1999) continue togenerate concern. Finally, unless the control elements of the tetsystem are integrated on a single plasmid (Schultze et al. 1996),the breeding of animals, to bring together with the responder com-ponents the tTA regulator (binding to the tet operator; tetO) andthe tetO/Cre sequence, is more complicated than crosses betweenHs-cre6and responder loci.
A common disadvantage of the tet and heat shock systems isleakiness, but the respective mechanisms and consequences differ.In the tet system, leakiness is defined as the intrinsic transcrip-tional activity of the tetO/minimal promoter element that is drivingcre. In this regard, unregulated (basal) levels ofcre expressionmay be sufficient to provide some level of enzymatic activity withundesirable recombinogenic action (St-Onge et al. 1996). In theheat shock system, leakiness corresponds to constitutive expres-sion of cre before the blastocyst stage. This feature cannot becontrolled, but nevertheless is well suited for the purpose of mo-saic analysis, as evidenced by the fact that a spectrum of mosaicswas generated simultaneously with all three reporters that wetested, although differences in particular frequencies of recombi-nation events were observed. We attribute this to the interestinginterplay between producer and responder loci that was revealedduring this study.
From the point of view of producers, variegation incre ex-pression was not unexpected, since it is a usual occurrence withtransgenes affected differentially in their transcriptional perfor-mance by integration sites (Wilson et al. 1990). Interestingly, fromthe point of view of responder loci, the efficiency of recombinationappeared to depend both on the sequence context ofloxP siteintegration and presumably on Cre dosage, despite the fact that Creis an enzyme and has the ability to gain access to chromatin andfunction even in postmitotic cells (Ku¨hn et al. 1995). Some evi-dence that the degree of mosaicism decreases with increasing Credosage has been recently provided (Zhang et al. 1998; Bo¨ger andGruss 1999). On the other hand, considered altogether, our datawith three lines of Cre producers and three reporter loci demon-strate, albeit indirectly, that the chromatin structure around theloxP sites must also be an important determinant of recombinationlevel. The fact that certain chromosomal loci are more accessiblethan others to Cre-mediated recombination was previously docu-mented in cell culture experiments (Baubonis and Sauer 1993).
The significance of mosaic analysis.The generation of chimeras,usually achieved today by injection of nullizygous ES cells intowild-type host blastocycts and vice versa, and also the generationof mosaics by means of conditional mutagenesis have enhancedsignificantly the power of mutational analysis (Rossant and Spence1998). Mosaics and chimeras are derived from one or more thanone zygote, respectively, but in both cases the organism consists ofa mixture of cell populations with different genotypes. Either ap-proach permits a genetic analysis, whenever a null mutation resultsin embryonic or early postnatal lethality. Thus, evidence is pro-vided whether the mutated gene has a function that is cell autono-mous or non-autonomous.
Despite such common analytical potential, mosaics and chi-meras have their own advantages and disadvantages. For example,because of a developmental bias of ES cells to colonize preferen-tially the epiblast (Beddington and Robertson 1989), the impact ofa mutation on embryonic or extraembryonic tissues can be ana-lyzed in injection chimeras, but not in mosaics. However, whenembryonic tissues at more advanced developmental stages are con-sidered, the initial advantage of chimeras turns into a disadvantagebecause ES cells tend not to make equal contributions to all lin-
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes 203

eages, apparently in a strain-dependent manner (Berger et al. 1995;Ioffe et al. 1995). Thus, mosaics are more advantageous, sincetheir mutant and wild-type cells have the same genetic back-ground. Moreover, whenever nullizygous ES cells necessary forthe generation of chimeras cannot be obtained because the muta-tion affects their viability, mosaic analysis becomes the only op-tion available. Finally, it is feasible to perform mutational analysesat chosen developmental times with mosaics that result from in-ducible cre expression, whereas the consequences of chimerismare strictly deterministic. We think, therefore, that our mosaickingHs-cre6transgenic line is a particularly useful tool.
Although the performance of the heat shock system could beoptimized further to increase its utility for induciblecre expres-sion, its availability already increases the flexibility in choosingamong existing alternatives a suitable strategy for the design andapplication of conditional mutagenesis, especially if the analyticalpower of mosaic studies can benefit the aims of a particular re-search program.
Acknowledgments.We thank Rick Morimoto for invaluable advice at thebeginning of this project, Monica Mendelsohn and Zaiqi Wu for help ingenerating transgenic and reporter mouse lines, and Krishna Ratnam fortechnical assistance. This work was supported by National Institutes ofHealth grants HD34526 and MH50733 (Project 2), and by National CancerInstitute grant CA75553 (Project 3) to A. Efstratiadis.
References
Baron U, Gossen M, Bujard H (1997) Tetracycline-controlled transcriptionin eukaryotes: novel transactivators with graded transactivation poten-tial. Nucleic Acids Res 25, 2723–2729
Baubonis W, Sauer B (1993) Genomic targeting with purified Cre recom-binase. Nucleic Acids Res 21, 2025–2029
Beddington RSP, Robertson EJ (1989) An assessment of the developmen-tal potential of embryonic stem cells in the midgestation embryo. De-velopment 105, 733–737
Berger CN, Tam PPL, Sturm KS (1995) The development of haematopoi-etic cells is biased in embryonic stem cell chimaeras. Dev Biol 170,651–663
Bergqvist I, Eriksson B, Eriksson M, Holmberg D (1998) Transgenic Crerecombinase expression in germ cells and early embryogenesis directshomogeneous and ubiquitous deletion of loxP-flanked gene segments.FEBS Lett 438, 76–80
Betz UA, Vosshenrich CAJ, Rajewsky K, Mu¨ller W (1996) Bypass oflethality with mosaic mice generated by Cre-loxP-mediated recombina-tion. Curr Biol 6, 1307–1316
Bevilacqua A, Kinnunen LH, Bevilacqua S, Mangia F (1995) Stage-specific regulation of murineHsp68gene promoter in preimplantationmouse embryos. Dev Biol 170, 467–478
Blake MJ, Gershon D, Fargnoli J, Holbrook NJ (1990) Discordant expres-sion of heat shock protein mRNAs in tissues of heat-stressed rats. J BiolChem 265, 15275–15279
Boger H, Gruss P (1999) Functional determinants for the tetracycline-dependent transactivator tTA in transgenic mouse embryos. Mech Dev83, 141–153
Christians E, Campion E, Thompson EM, Renard JP (1995) Expression ofthe HSP 70.1gene, a landmark of early zygotic activity in the mouseembryo, is restricted to the first burst of transcription. Development 121,113–122
Christians E, Michel E, Renard JP (1997) Hsp70 genes and heat shockfactors during preimplantation phase of mouse development. Cell MolLife Sci 53, 168–178
Copp AJ (1995) Death before birth: clues from gene knockouts and mu-tations. Trends Genet 11, 87–93
Danielian PS, Muccino D, Rowitch DH, Michael SK, McMahon AP (1998)Modification of gene activity in mouse embryos in utero by a tamoxifen-inducible form of Cre recombinase. Curr Biol 8, 1323–1326
Doetschman T (1999) Interpretation of phenotype in genetically engi-neered mice. Lab Anim Sci 49, 137–143
Edwards MJ , Shiota K, Smith MSR, Walsh D (1995) Hyperthermia andbirth defects. Reprod Toxicol 9, 411–425
Feil R, Brocard J, Mascrez B, LeMeur M, Metzger D et al. (1996) Ligand-
activated site-specific recombination in mice. Proc Natl Acad Sci USA93, 10887–10890
Fiorenza MT, Farkas T, Dissing M, Kolding D, Zimarino V (1995) Com-plex expression of murine heat shock transcription factors. Nucleic Ac-ids Res 23, 467–474
Flanagan SW, Ryan AJ, Gisolfi CV, Moseley PL (1995) Tissue-specificHSP70 response in animals undergoing heat stress. Am J Physiol 268,R28–R32
Fujio N, Hatayama T, Kinoshita H, Yukioka M (1987) Induction of fourheat-shock proteins and their mRNAs in rat after whole-body hyperther-mia. J Biochem (Tokyo) 101, 181–187
Gossen M, Bujard H (1992) Tight control of gene expression in mamma-lian cells by tetracycline reponsive promoters. Proc Natl Acad Sci USA89, 5547–5551
Gossen M, Freundlieb S, Bender G, Mu¨ller G, Hillen W et al. (1995)Transcriptional activation by tetracycline in mammalian cells. Science268, 1766–1769
Gu H, Zou Y-R, Rajewsky K (1993) Independent control of immunoglob-ulin switch recombination at individual switch regions evidencedthrough Cre-loxP-mediated gene targeting. Cell 73, 1155–1164
Gunther E, Walter L (1994) Genetic aspects of the hsp70 multigene familyin vertebrates. Experientia 50, 987–1001
Hahnel AC, Gifford DJ, Heikkila JJ, Schultz GA (1986) Expression of themajor heat shock protein (hsp 70) family during early mouse embryodevelopment. Teratog Carcinog Mutagen 6, 493–510
Hogan B, Beddington R, Costantini F, Lacy E (1994)Manipulating themouse embryo: A laboratory manual.(Cold Spring Harbor, N.Y.: ColdSpring Harbor Laboratory Press)
Hunt C, Calderwood S (1990) Characterization and sequence of the mouseHsp70gene and its expression in mouse cell lines. Gene 87, 199–204
Ioffe E, Liu Y, Bhaumik M, Poirier F, Factor SM, Stanley P (1995) WW6:an embryonic stem cell line with an inert genetic marker that can betraced in chimeras. Proc Natl Acad Sci USA 92, 7357–7361
Kellendonk C, Tronche F, Casanova E, Anlag K, Opherk C et al. (1999)Inducible site-specific recombination in the brain. J Mol Biol 285, 175–182
Kistner A, Gossen M, Zimmermann F, Jerecic J, Ullmer C et al. (1996)Doxycycline-mediated quantitative and tissue-specific control of geneexpression in transgenic mice. Proc Natl Acad Sci USA 93, 10933–10938
Kothary R, Clapoff S, Darling S, Perry MD, Moran LA et al. (1989)Inducible expression of anhsp68–lacZhybrid gene in transgenic mice.Development 105, 707–714
Kuhn R, Schwenk F, Aguet M, Rajewsky K (1995) Inducible gene target-ing in mice. Science 269, 1427–1429
Kulkarni RN, Bruning JC, Winnay JN, Postic C, Magnuson MA et al.(1999) Tissue-specific knockout of the insulin receptor in pancreaticbcells creates an insulin secretory defect similar to that in type 2 diabetes.Cell 96, 329–339
Lakso M, Sauer B, Lee E, Manning RW, Yu S-H et al. (1992) Targetedoncogene activation by site-specific recombination in transgenic mice.Proc Natl Acad Sci USA 89, 6232–6236
Lakso M, Pichel JG, Gorman JR, Sauer B, Okamoto Y et al. (1996)Efficient in vivomanipulation of mouse genomic sequences at the zygotestage. Proc Natl Acad Sci USA 93, 5860–5865
Liu J-P, Baker J, Perkins AS, Robertson EJ, Efstratiadis A (1993) Micecarrying null mutations of the genes encoding insulin-like growth factorI (Igf-1) and type 1 IGF receptor (Igf1r). Cell 75, 59–72
Lobe CG, Nagy A (1998) Conditional genome alteration in mice. Bioes-says 20, 200–208
Mansuy IM, Winder DG, Moallem TM, Osman M, Mayford M et al.(1998) Inducible and reversible gene expression with the rtTA systemfor the study of memory. Neuron 21, 257–265
Marth JD (1996) Recent advances in gene mutagenesis by site-directedrecombination. J Clin Invest 97, 1999–2002
Mezger V, Renard J-P, Christians E, Morange M (1994) Detection of heatshock element-binding activities by gel shift assay during mouse pre-implantation development. Dev Biol 165, 627–638
Mirkes PE, Cornel LM, Wilson KL, Dilmann WH (1999) Heat shockprotein 70 (Hsp70) protects postimplantation murine embryos from theembryolethal effects of hyperthermia. Dev Dyn 214, 159–170
Morange M (1999) Heat shock proteins in embryonic development. InHandbook of Experimental Pharmacology,vol. 136, Latchman DS, ed.(New York: Springer), pp 305–327
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes204

Muller U (1999) Ten years of gene targeting: targeted mouse mutants, fromvector design to phenotype analysis. Mech Dev 82, 3–21
Nowak TS, Bond U, Schlesinger MJ (1990) Heat shock RNA levels inbrain and other tissues after hyperthermia and transient ischemia. J Neu-rochem 54, 451–458
Porter A (1998) Controlling your losses: conditional gene silencing inmammals. Trends Genet 14, 73–79
Rajewsky K, Gu H, Kühn R, Betz UAK, Mu¨ller W et al. (1996) Condi-tional gene targeting. J Clin Invest 98, 600–603
Rossant J, McMahon A (1999) “Cre”-ating mouse mutants—a meetingreview on conditional mouse genetics. Genes Dev 13, 142–145
Rossant J, Spence A (1998) Chimeras and mosaics in mouse mutant analy-sis. Trends Genet 14, 358–363
Sauer B (1998) Inducible gene targeting in mice using the Cre/lox system.Methods 14, 381–392
Schultze N, Burki Y, Lang Y, Certa U, Bluethmann H (1996) Efficientcontrol of gene expression by single step integration of the tetracyclinesystem in transgenic mice. Nat Biotechnol 14, 499–503
Schwenk F, Baron U, Rajewsky K (1995) A cre-transgenic mouse strainfor the ubiquitous deletion ofloxP-flanked gene segments includingdeletion in germ cells. Nucleic Acids Res 23, 5080–5081
Spencer DM (1996) Creating conditional mutations in mammals. TrendsGenet 12, 181–187
St-Onge L, Furth PA, Gruss P (1996) Temporal control of the Cre recom-binase in transgenic mice by a tetracycline responsive promoter. NucleicAcids Res 24, 3875–3877
Vasioukhin V, Degenstein L, Wise B, Fuchs E (1999) The magical touch:genome targeting in epidermal stem cells induced by tamoxifen appli-cation to mouse skin. Proc Natl Acad Sci USA 96, 8551–8556
Wilson C, Bellen H, Gehring W (1990) Position effects on eukaryotic geneexpression. Annu Rev Cell Biol 6, 679–714
Zhang Y, Wienands J, Zu¨rn C, Reth M (1998) Induction of the antigenreceptor expression on B lymphocytes results in rapid competence forsignaling of SLP-65 and Syk. EMBO J 17, 7304–7310
P. Dietrich et al.: Heat shock promoter-controlledcre transgenes 205