increased allergen-specific th2 responses in vitro in atopic subjects receiving subclinical allergen...
TRANSCRIPT

Atlergy 1997:52:860-865Printed in UK - all rights reserved
Copvrigttt © Munksgaard 1997
ALLERGYISSN 0105-4538
Short communication
Increased allergen-specific Th2 responses invitro in atopic subjects receiving subclinicalallergen challenge
Gabrielsson S, Paulie S, Roquet A, Ihre E, Lagging E, van Hage-Hamsten M, Harfast B, Troye-Blotnberg M. Increased allergen-specificTh2 responses in vitro in atopic subjects receiving subclinieal allergenchallenge.Allergy 1997: 52: 860-865. © Munksgaard 1997.
The study aimed to deterrnine whether inhalation of subclinical allergendoses leads to a shift in the balance between T helper (Th) 1 and Th2 cellsin asthmatic patients. Elevated IgE requires allergen-specific T eellsproducing eytokines such as interleukin (IL)-4 or IL-13. Interferon-gamma(IFN-y) produced by Till eells counteracts the effects of IL-4. In nature,allergic persotis are often exposed to low levels of allergen, leading tohyperreactivity, but not to acute allergic reactions. In this study, nine allergiepersons inhaled low doses of allergen or placebo in a double-blind mannerover seven consecutive weekdays. During the study, the bronchialresponsiveness to histamine challenge increased, but no subject exhibitedasthmatic symptoms. Blood was drawn on days 0,1, 4, and 9, and the numberof IL-4- and IFN-y-producing cells was measured by enzyme-linkedimmunospot (ELISPOT) assay after in vitro stimulation with a low-dosephytohemagglutinin (PHA) mixed with the relevant allergen or with PHAalone. In three of the four subjects receiving allergen, the 1L-4/IFN-Y ratioincreased during the time of the study. No increase was seen in the placebogroup. No increase was seen in serum IgE levels in any of the groups. Weconclude that a shift in the balance between Thi and Tli2 cells can bedetected in subjects exposed to subclinical allergen doses.
S. Gabrielsson\ S. Paulie\A. Roquet̂ , E. lhre^ E. Lagging^M. van Hage-Hamsten^ B. Harfast ,̂M. Troye-Blomberg^'Department of Immunology, StockholmUniversity, Stocl<holm; ^Division of RespiratoryMedicine, and ''Clinical Immunology, KarolinskaHospital, Stockholm, Sweden
Keywords: atopy; ELISPOT; interferon-gamma;interleukin-4; low-dose provocation.
Susanne GabrielssonDepartment of ImmunologyStockholm UniversityS-106 91 StockholmSweden
Accepted for publication 1 April 1997
Atopic patients exhibit elevated levels of both totaland allergen-specific IgE in serum (1). Develop-ment of IgE-producing B cells requires the eyto-kines interleukin (IL)-4 or IL-13 (2-5), producedby a subset of CD4-I- T-helper (Th) eells commonlyreferred to as Th2-cells (6). On the other hand,interferon-gamma (IFN-y), produced by Thi-cells,inhibits IL-4-induced IgE production (3,7). Allergen-specific T-cell clones obtained from atopic patientshave a Th2-like cytokine profile (8, 9), and peri-pheral blood mononuclear cell (PBMC) culturesfrom allergic patients respond with a Th2-likecytokine profile when activated in vitro with allergen(10, 11).
When atopic asthma patients are challenged withhigh doses of allergen, an immediate asthmatic
response is detectable within minutes. In somecases, the immediate response is followed by anonspecific bronchial hyperreactivity that can per-sist for several days (12). During this response, ili2-like eytokines can be detected in bronchoalveolarlavage (BAL) and lung biopsies (13), as well as inperipheral blood (14).
Recruitment of inflammatory cells starts longbefore patients have clinical symptoms. The expo-sure of low doses of allergen usually leads tosubchnical inflammation and is characterized byunspecific airway hyperreactivity detected as in-creased sensitivity to histamine (15, 16). In thisstudy, we wanted to investigate whether low dosesof allergen, which did not evoke an acute asthmaticresponse but inereased bronchial reactivity, would
860

IL-4/IFN-y ratio after allergen exposure
induce a Th2-response detected as an increasedratio of IL-4- to IFN-y-producing cells after invitro activation with allergen. In this double-blind,placebo-controlled study, we used the highly sensi-tive enzyme-linked immunospot (ELISPOT) assayto detect individual cytokine-producing cells.
patients investigated for eosinophil count, serum andintracellular eosinophil cationic protein (ECP), CD23,and HLA-DR, Tlie only significant difference seenbetween the placebo and the allergen-exposedgroups was an increase in bronchial hyperrespon-siveness (BHR) in the allergen-exposed group (16).
Material and methodsReagents
The monoclonal antibodies used in the ELISPOTwere mAb 1-DIK and 7B6-1 to human IFN-y andmAb 82-4 (IL-4-1) and 12-1 (IL-4-2) to humanIL-4 (all mAbs were obtained from MABTECH,Stockholm, Sweden). Tlie specific allergens usedwere Aquagen-SO bireh, timothy, Artemisia vulgaris,and cat, 1x10*̂ SO-U/ml (ALK, Denmark), Tliemitogen was phytohemagglutinin (PHA) (MurexDiagnostics, Dartford, UK).
Subjects and study design
Tlie atopic group comprised nine patients with ahistory of mild allergic asthma to pollen and/oranimal dander positive in skin prick test and radio-allergosorbent test (RAST > class 2), The trialstarted with a histamine ehallenge, after which thesubjects were challenged with either allergen orplacebo in a double-blind, randomized manner byinhaling low doses of allergen daily for 5 days, witlian interruption for 2 days and then continued foranother 2 days. Four of the patients receivedallergen and five of them received placebo. Dif-ferent allergen doses (Table 1) were selected bybasal PD20 values, skin prick test, and specific IgEtiters, and were chosen to correspond to 1/100 ofthe dose that caused an acute allergic reaction (15),Blood samples were drawn just before inhalationon days 0, 1, 4, and 9, The patients included in thisstudy were part of a larger study, including 14
Table 1. Total and allergen-specific IgE before and after inhalation of subclinical
allergen doses
Donorno.
123456789
Inhalation dose(SQ-U/day)
35,0
0,735,0
14,0—_-_-
Total
Day 0
951701B02630
24035025
280
IgE (kU/mll
Day 9
1101301802530
20026028
290
Specific
Day 0
46.0
2.73.67.4
19.0
68.0
71.0
3.362.0
IgE (kU/mll
Day 9
43.0
2.53.B8.3
18.0
58.0
71.0
3.364.0
IgE detection
Specific serum IgE was measured by RAST andtotal IgE by PRIST (Pharmacia-Upjohn, Sweden)according to the instructions of the manufacturers.
Preparation of PBMC
Mononuclear cells from heparinized peripheralblood were isolated by Ficoll-Paque (Pharmacia,Stockholm, Sweden) gradient centrifugation. Thecells were frozen as follows, PBMC were suspended(10^-10» cells/ml) in tissue-culture media (TCM,RPMI-Hepes supplemented with gentamicin[25 |ig/ml], 2 niM L-glutamine, and 10% heat-inactivated fetal calf serum), containing 10% dime-thyl sulfoxide (DMSO), and frozen gradually atl°C/min to -70°C in a freezing container (NalgeneCryo 1°C, Nalge Company, USA), Tliereafter, thetubes were stored in liquid nitrogen. Before use,the cells were thawed quickly in a 37°C water bath,diluted, and washed in TCM, Samples from thedifferent time points were analyzed in the sameELISPOT experiment for each subject.
ELISPOT assay
Tlie reverse ELISPOT assay was performed mainlyas described before (10), Briefly, the anti-human-IFN-Y mAb 1-DIK or anti-human-IL-4 mAb 82-4(MABTECH AB, Stockholm, Sweden) was dilutedin a concentration of 15 |ig/ml in sterile, filtered(0,45-|j.m) phosphate-buffered saline (PBS), and100 10,1 per well was added to nitrocellulose plates(Millititer, Millipore Corp,, Bedford, MA, USA),Plates were incubated overnight at 4°C, After eachstep, the plate was washed with filtered PBS, Halfor 1 ml cell suspension (2 million cells/ml) wastransferred to 5-ml culture tubes (Falcon, Becton-Dickinson, NJ, USA), and allergens at differentconcentrations and/or a suboptimal dose of PHAwas added, Tlie use of a suboptimal dose of PHAhas been shown to induce a synergistic effect onthe number of IL-4-producing cells in allergic, butnot healthy, subjects (10), The tubes were incu-bated at 37°C in a humid atmosphere containing5% CO2 for 4h, Thereafter, the cells were resus-pended, and 100 p,l of 2x10^ cells/ml for IL-4 and2x10"̂ cells/ml for IFN-ywere added to each coatedELISPOT well in duplicates, Tlie plates were then
861
Gabrielsson et al.
incubated for 40-42 h at 37°C, after which 100 lillwell of the biotinylated IFN-y mAb 7B6-1 orIL-4 mAb 12-1 (1 mg/ml) was added and incubatedfor 2 h at room temperature. Then, the plateswere incubated for 90 min at room tetnperaturewith 100 )il of streptavidin-alkaline-phosphatase(MABTECH AB, Stockholm, Sweden) diluted1/1000. A volume of 100 |il/well of BCIP/NBTsubstrate solution (Bio-Rad, Richmond, CA, USA)was added and incubated at room temperatureuntil dark spots emerged (approximately 1 h). Tliecolor development was stopped by washing withtap water. After drying, the spots were countedunder a dissection microscope (x40) where onespot indicates one cytokine-producing cell. Themedium control background, ranging from 0-2spots/200000 cells to 0-12 spots/20000 cells forIL-4 and IFN-y, respectively, was subtracted fromthe values.
Analysis of data
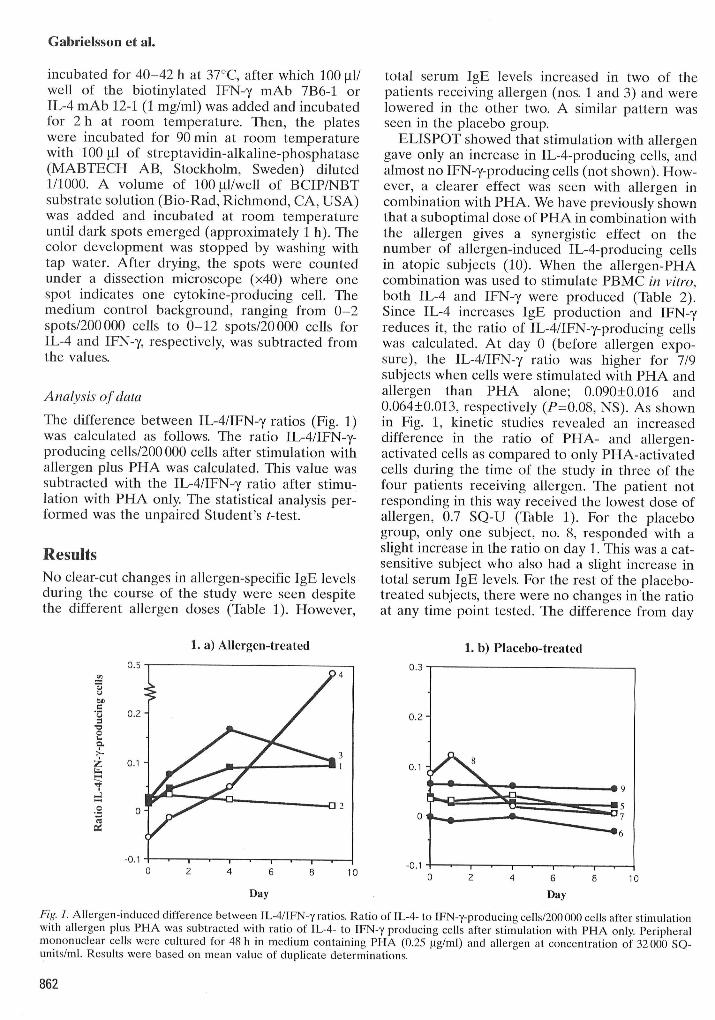
The difference between IL-4/IFN-y ratios (Fig. 1)was calculated as follows. The ratio IL-4/IFN-y-producing cells/200 000 cells after stimulation withallergen plus PHA was calculated. This value wassubtracted with the IL-4/IFN-y ratio after stimu-lation with PHA only. The statistical analysis per-formed was the unpaired Student's t-test.
ResultsNo clear-cut changes in allergen-specific IgE levelsduring the course of the study were seen despitethe different allergen doses (Table 1). However,
1. a) Allergen-treated
total serum IgE levels increased in two of thepatients receiving allergen (nos. 1 and 3) and werelowered in the other two. A similar pattern wasseen in the placebo group.
ELISPOT showed that stimulation with allergengave only an increase in IL-4-producing cells, andalmost no IFN-y-producing cells (not shown). How-ever, a clearer effect was seen with allergen incombination with PHA. We have previously shownthat a suboptimal dose of PHA in combination withthe allergen gives a synergistic effect on thenumber of allergen-itiduced IL-4-producing cellsin atopic subjects (10). When the allergen-PHAcotTibination was used to stitnulate PBMC in vitro,both IL-4 and IFN-y were produced (Table 2).Since IL-4 increases IgE production and IFN-yreduces it, the ratio of IL-4/IFN-y-producing cellswas calculated. At day 0 (before allergen expo-sure), the IL-4/IFN-y ratio was higher for 7/9subjects when cells were stirnulated with PHA andallergen than PHA alone; 0.090+0.016 and0.064±0.013, respectively (P^0.08, NS). As shownin Fig. 1, kinetic studies revealed an increaseddifference in the ratio of PHA- and allergen-activated cells as compared to only PHA-activatedcells during the time of the study in three of thefour patients receiving allergen. The patient notresponding in this way received the lowest dose ofallergen, 0.7 SQ-U (Table 1). For the placebogroup, only one subject, no. 8, responded with aslight increase in the ratio on day 1. Tliis was a cat-sensitive subject who also had a slight increase ititotal serum IgE levels. For the rest of the placebo-treated subjects, there were no changes in the ratioat any time point tested. The difference from day
1. b) Placebo-treated
-0.1
Day
Fig. 1. Allergen-induced difference between IL-4/IFN-y ratios. Ratio of IL-4- to IFN-y-producing cells/200 000 cells after stimulationwith allergen plus PHA was subtracted with ratio of lL-4- to IFN-y producing cells after stimulation with PHA only. Peripheralmononuclear cells were cultured for 48 h in medium containing PHA (0.25 |Xg/ml) and allergen at concentration of 32 000 SQ-units/ml. Results were based on mean value of dupheate determinations.
862

IL-4/IFN-Y ratio after allergen exposure
Table 2. In vitro cytokine responses after stimulation with allergen combined with suboptimal dose of PHA before and during inhalation of subclinical allergen doses
Donor
1234
CJl
6789
IL-4"
1446642132205015189297
Day 0
iFN-Y"
263179262127811916459229
IL-4
172106441982126218072239
Day 1
IFN-Y
197176207525910815830152
lL-4
13311616123322914230666195
Day 4
IFN-Y
9622153168247182263134175
IL-4
24379852261806579B3240
Day 9
IFN-Y
171340362416671120271210
" IL-4-producing ceiis/200 000 cells.'̂ IFN-Y-producing ceiis/20 000 cells.
1 to 9 was not statistically different between thegroups, although a clear tendency could be seen(f^O.ll , unpaired Student's /-test) despite the lownumber of patients.
The absolute numbers of IL-4-producing cells inresponse to allergen and PHA also increased in thepatients inhaling allergen (Table 1). However,these levels varied more from day to day than theratios in both groups. In contrast to what has beendescribed before (10), the IL-4 response was notcorrelated to the specific serum IgE levels, probablydue to the low number of patients included in thisstudy.
Clinical evaluation of 14 patients undergoing thesame treatment as here (16), nine of whom areincluded in this study, showed a significant increasein bronchial responsiveness during the study periodin all patients receiving allergen. None of thepatients experienced any immediate allergic symp-toms or any delayed symptoms.
DiscussionSpecific antigen stimulation in vitro induces verylow frequencies of IL-4-producing cells comparedto several other cytokines; e.g., IL-2 (17). In thisstudy, we used a system where the allergen wascombined with a suboptimal dose of PHA, whichinduces different responses in atopic subjects (Tli2)from those of healthy subjects (Thl) (18). Usingthis protocol, we have previously shown (10) thatthe allergen-specific IL-4 response is amplified ina synergistic way. This is in contrast to what hasbeen described in patients with psoriasis, a Thldisease, where a synergistic increase in the numberof IFN-"/-producing cells was seen (19). Takentogether, these data clearly indicate that PHA doesnot direct the response in any particular direction,but rather reflects the frequencies of cytokine-producing cells.
Type I allergic responses are of the Th2-type (20,21). lL-4 produced by Tli2-cells induces switchingof IgM to IgE, while IFN-y produced by Thl cellsinhibits the IgE-promoting effects of IL-4. Hence,the ratio of IL-4/IFN-y-producing cells should givebetter information on the long-term IgE effectsthan only measurement of IL-4. Moreover, esti-mation of the ratio of IL-4/IFN-'Y-producing cellshas been shown to be more reproducible than theabsolute numbers of IL-4- and IFN-y-producingcells (10).
Previous studies have reported an increased Th2response in patients inhaling high doses of allergenduring the pollen season (14). In the present study,we used an experimental allergen challenge model,which probably more resembles the normal levelin the environment of cat, dog, or other nonseasonalallergens. Minute amounts of allergen were inhaledfor 7 days by the patients. Even though no clinicalsymptoms occurred, this procedure resulted in anincreased ratio of IL-4/IFN-Y-producing cells afterin vitro allergen exposure in 3/4 patients. Tlienonresponding patient received the lowest dose ofallergen challenge. No spontaneous IL-4 produc-tion after inhalation could be seen in any of thesubjects. Tlie reason that no spontaneous but onlyin vitro activated IL-4-producing cells could bevisualized is not known. Tliis observation mightindicate that the low allergen dose given in vivowas not enough to stimulate cells to produce IL-4.It is well established that cells stimulated to proli-ferate do not always produce cytokines (22).
The shift to the Tli2 type response after lowdoses of allergen challenge suggests a proliferationor activation of Th2 cells which, when exposed tohigher levels of allergen, start to produce IL-4. Tliiswould lead to a greater IgE production than insubjects not preexposed to low allergen levels.However, it remains to be established whether theincreased number of IL-4-producing cells seen in
863

Gabrielsson et al.
our study was due to clonal expansion or realloca-tion from other organs.
No clear-cut changes in total or allergen-specificIgE levels could be seen in either the allergen orthe placebo groups. This was expected since nospontaneous IL-4 production could be detected.Moreover, the time point for IgE detection in thisstudy might be too early. IgE increase after allergenexposure in vivo has been reported after 14 days(23).
Our observation that Th2 cells are induced afterlow allergen exposure contrasts with what has beenseen when low doses of allergen are administeredsubcutaneously during specific immunotherapy. Inthe latter case, IL-4 production is reduced (24). Thediscrepancy in the induction of functionally distinctimmune responses might be that different allergendoses or that different routes of administration areused. This could lead to uptake of the allergen bydifferent types of antigen-presenting cells (25).Other differences in the microenvironment betweenthe two sites could also affect the development ofThO cells into Thl or Th2 cells (26).
Taken together, our results show that, despitelack of evident clinical symptoms or signs of acti-vation of inflammatory cells in peripheral blood,an increase in the allergen-induced ratio of IL-4/IFN-y-producing cells can be seen in subjectsexposed to repeated low doses of allergen.
AcknowledgmentsThis work was supported by the King Gustaf the Fifth 80-yearJubilee Foundation, the Swedish Asthma and Allergy Associ-ation, the Swedish National Board for Laboratory Animals,and ihc VSrdal Foundation.
References1. Johansson SGO, Bennich M, Berg T. In vitro diagnosis of
atopic allergy. III. Quantitative estimation of circulatingIgE antibodies by the radioallergosorbent test. Int ArchAllergy Appl Immunol 1971;41:443-51.
2. Del Prete G, Maggi E, Parronchi P, et al. IL-4 is an essentialfactor for the IgE synthesis induced in vitro by human Tcell clones and their supernatants. J Immunol f988;140:4193-8.
3. Pene J, Roussel F, Briere F, et al. IgE production by normalhuman lymphocytes is induced by interleukin 4 and sup-pressed by interferons y and a and prostaglandin Ej. ProcNatl Acad Sci U S A 1988;85:6880-4.
4. Vercelh D, Geha RS. Regulation of isotype switching. CurrOpin Immunol 1992:4:794-7.
5. Coffman R, Carty J. A T cell activity that enhancespolyelonal IgE production and its inhibition by interleron-y.J Immunol f986;l36:949-54.
6. Mosmann TR, Cherwinski H, Bond MW, Giedlin MA,Coffman RL. Two types of murine helper T cell clone.J Immunol 1986;f36:2348-57.
7. Chretien I, Pene J, Briere F, Malefijt RDW, Rousset F,De Vries JE. Regulation of human IgE synthesis. I. Human
IgE synthesis in vitro is determined by the reciprocalantagonistic effects of interleukin 4 and interferon-y. EurJ fmmunol 1990;20:243-5L
8. Wierenga EA, Snoek M, deGroot C, et al. Evidence forcompartmentalization of functional subsets of CD4* Tlymphocytes in atopic patients. J Immunol 1990;144:4651-6.
9. Romagnani S, Maggi E, Parronchi P, Macchia D,Piccinni MP, Ricci M. Increased numbers of Th2-like CD4-FT cells in target organs and in the allergen-specific reper-toire of allergic patients. Int Arch Allergy Appl Immunol1991 ;94:133-6.
10. Gabrielsson S, Paulie S, Rak S, et al. Specific inductionof interleukin-4-producing cells in response to in vitroallergen stimulation in atopic individuals. Clin Exp Allergy1997;27: in press.
11. Imada M, Estelle F, Simons R, Jay F, Hayglass K. Allergen-stimulated interleukin-4 and interferon-y production inprimary culture: responses of subjects with allergic rhinitisand normal controls. Immunology 1995;85:373-80.
12. Cartier A, Thomson N, Frith P, Roberts R, Hargreave F.Allergen-induced increase in bronchial responsiveness tohistamine: relation to the late asthmatic response andchange in airway calibre. J Allergy Clin Immunol 1982;70:170-7.
13. Robinson D, Hamid O, Bentley A, Ying S, Kay A,Durham S. Activation of CD4+ T cells, increased Tli2-typecytokine niRNA expression, and eosinophil recruitment inbronchoalveolar lavage after allergen inhalation challengein patients with atopic asthma. J Allergy Clin Immunol1993:92:313-24.
14. Lagier B, Pons N, Rivier A, et al. Seasonal variations ofinterleukin-4 and interferon-y release by peripheral bloodmononuclear cells from atopic subjects stimulated bypolyclonal activators. J Allergy Clin Immunol 1995:96:932-40.
15. Ihre E, Zetterstrom O. Increase in non-specific tironchialresponsiveness alter repeated inhalation of low doses ofallergen. Clin Exp Allergy 1993:23:298-305.
16. Roquet A. The contribution of eosinophils and mediatorsin allergic inflammation. Stockholm: Reproprint, 1996.
17. Powers GD, Abbas AK, Miller RA. Frequencies of IL-2-and lL-4-secreting T cells in naive and antigen-stimulatedlymphocyte populations. J Immunol 1988:140:3352-7.
18. Piccinni M-P, Beloni L, Giannarini L, et al. Abnormalproduction of the T helper 2 cytokines interleukin-4 andinterleukin-5 by T cells from newborns with atopic parents.Eur J Immunol 1996:26:2293-8.
19. Sigmundsdottir H, Sigurgeirsson B, Troye-Blomberg M,Good MF, Valdimarsson H, Jonsdottir I. Circulating T-cellsof patients with active psoriasis respond to streptococcalM-peptides sharing sequences with human epidermalkeratins. Scand J Immunol 1997: in press.
20. Romagnani S. Lymphokine production tiy human T cellsin disease states. Ann Rev Immunol 1994:12:227-57.
21. Del Prete GF, de Carli M, D'Elios MM, et al. Allergenexposure induces the activation of allergen-specific Th2cells in the airway mucosa of patients with allergic respi-ratory disorders. Eur J Immunol 1993:23:1445-9.
22. Troye-Blomberg M, Riley EM, Kabilan L, et al. Productionby activated human T cells of interleukin 4 but notinterferon-gamma is associated with elevated levels ofserum antibodies to activating malaria antigens. Proc NatlAcad Sci U S A 1990:87:5484-8.
23. Weller FR, Kallenberg CGM, Jansen HM, et al.Tlie primary immune response in bronchial asthma. I. Akinetic study of Helix pomatia hemocyanin-specific IgE,IgG, IgA, and IgM antibody responses in patients withasthma and in matched controls. J Allergy Clin Immunol1985:76:29-34.
864

IL-4/IFN-Y ratio after allergen exposure
24, Secrist H, Chelen CI, Wen Y, Marshall JD, Umetsu DT, lymphocyte differentiation in vivo. J Exp Med 1995;Allergen immunotherapy decreases interleukin 4 produc- 181:1569-74,tion in CD4+ T cells from allergic individuals, J Exp Med 26, Seder R, Paul WE, Dvorak AM, et al. Mouse splenic and1993;178:2123-30, bone marrow cell populations that express high-affinity FCj.
25, Pfeiffer C, Stein J, Southwood S, Ketelaar H, Sette A, receptors and produce interleukin 4 are highly enriched inBottomly K, Altered peptide ligands can control CD4 T basophils, Proc Natl Acad Sci U S A 1991;88:2835-9,
865
