in vitro evaluation of dew-retting of flax by fungi from southern europe
TRANSCRIPT

*Corresponding Author E-mail: [email protected]
© 2001 Association of Applied Biologists
Ann. appl. Biol. (2001), 138:343-351Printed in Great Britain 343
In vitro evaluation of dew-retting of flax by fungi from southern Europe
By G FILA*, L M MANICI and F CAPUTO
ISCI - Istituto Sperimentale per le Colture Industriali,Via di Corticella, 133 � I-40128 Bologna, Italy
(Accepted 14 February 2001; Received 11 October 2000)
Summary
Flax dew-retting is widely adopted in most flax-growing countries, but it does not represent a practicalsolution where dry weather conditions occur after harvest. A study of the local microbiological aspectswas undertaken as a contribution to improve field-retting of flax under southern European climates.Fungi were isolated from soil and dew-retted flax in northern Italy, and 23 representative strains werechosen to test their ability to ret flax stems. Experiments were performed in vitro on flax stem piecesartificially inoculated with single fungal strains. Retting degree was assessed with a mechanical test,to evaluate the ease with which the bast was detached from the wood core, and by the analysis of theresidual fibre pectins using uronic acid. Uronic acid dosage provided a better differentiation of thestrains than the mechanical test. There was a large variability in retting ability among the speciesassayed and even among strains of the same species. The best results were obtained with all Aspergillusand Penicillium strains, while Mucor and Rhizopus strains showed a variable retting ability. Fusarium,Trichoderma strains and Epicoccum nigrum had the poorest retting abilities among all the fungalstrains assayed.
Key words: Flax, fungi, in vitro screening, retting, galacturonic acid, fibre
Introduction
Fibre-flax (Linum usitatissimum L.) was widelycultivated in Italy until the beginning of the twentiethcentury, when the advent of cotton and of syntheticfibres brought about a progressive decline of thiscrop. In spite of this, Italy remains one of the mostimportant producers of linen products, importing allraw materials entirely from abroad. Today, given thehigh internal demand, the revitalisation of flaxgrowing is an attractive opportunity for Italianfarmers, at a time when the profitability of food cropshas been curtailed by lower prices and lower grants.Moreover, agriculture would benefit from theintroduction of flax, which would make it possibleto widen rotations and reduce energy inputs. A recentrevival in the research about flax-growing in Italy(Cremaschi & Maestrini, 1996; Reyneri & Cavallero,1997) has demonstrated the suitability of this cropto many environments in northern and southern Italy.
The main obstacle to increased flax cultivation inItaly is the absence of industrial facilities for rettingand processing the straw into spinnable fibre. In thepast, the fibre was extracted by water-retting, aprocess which is labour-intensive, has high energyrequirements, and produces huge amounts ofpolluting waste water. This is why in the last fourdecades water-retting has been almost completelysupplanted by dew-retting in most flax-producing
countries (Sharma & Van Sumere, 1992; Meijer,Vertregt, Rutgers & van der Waart, 1995). Thistechnique consists of spreading out the straw in thefield to expose it to the degrading action of aerobicmicro-organisms such as fungi and yeasts (Wieringa,1956; Frederiksen, 1967). Although dew-retting isgreatly weather-dependent, it has many importantadvantages, such as low energy requirements,suitability for mechanisation, and absence ofenvironmental drawbacks.
Several studies done in the past in northernEuropean countries reported that the most frequentfungi involved in flax dew-retting wereCladosporium herbarum, Alternaria spp.,Stemphylium spp., Epicoccum nigrum, Botrytiscinerea, Fusarium culmorum, Mucor spp., Rhizopusspp., Penicillium spp. and Aspergillus spp.(Wieringa, 1956; Frederiksen, 1967; Brown, 1984;Brown & Sharma, 1984). However, more recentstudies showed that retting efficiency of fungiisolated from dew-retted flax varies widely(Henriksson et al., 1997; Akin, Rigsby, Henriksson& Eriksson, 1998).
In this study an analysis was made of the rettingability of several wild fungal strains isolated innorthern Italy. The knowledge acquired should helpevaluate the retting potential of the natural mycofloraand contribute to the optimisation of a dew-rettingprotocol for southern European environments.

344 G FILA, L M MANICI & F CAPUTO
Materials and Methods
Plant materialThe flax plants used (cv. �Alba�) were cultivated
at the ISCI experimental farm (Budrio, northernItaly; 44.50°N; 11.50°E). Plants were sown on 23March 1999 at 1800 seeds m-2 and harvested at green-yellow maturity on 6 July 1999. After harvest, plantswere allowed to dry for 2 days in the field, thenstored in a ventilated storeroom.
The experimental material consisted of 12 cm-longpieces, cut from the middle part of stems withuniform diameter of about 2 mm, autoclaved at120°C, 105 kPa, for 30 min.
Isolation and conservation of fungiThe fungal isolates were obtained mostly from
samples of soil and dew-retted flax collected fromJune to October 1997 in flax-cultivated fields at ourexperimental farm.
Soil samples were taken from the top layerremoving the first 5 cm; they were partially dried atroom temperature for 3 days, then they were sievedthrough a 5-mm mesh screen and homogenised byhand. Twenty-five g of the powdered soil was mixedin 225 ml of sterile water with a magnetic stirrer for10 min to obtain a 10-1 diluted soil solution. In asimilar way, retted flax samples were cut into 1-2cm pieces, 20 g of which was mixed in 200 ml ofsterile water with a magnetic stirrer for 10 min toobtain a 10-1 diluted solution.
1 ml aliquots of 10-2 and 10-3 soil dilutions weremixed with 49 ml of water agar containing 3 g litre-1
bovine bile (Ox-gall Powder, Sigma) and 200 mglitre-1 streptomycin sulphate (Sigma). The mixturewas finally distributed on five Petri dishes andincubated for 48 h at 24 ± 2°C under natural light.Colonies were counted and expressed as CFU(Colony Forming Unit) g-1 soil or flax. Fungalisolates of different morphologies were transferredto potato-dextrose agar (PDA) containing 200 mglitre-1 streptomycin sulphate. Species were identifiedby macroscopic and microscopic morphologicalcharacteristics using relevant taxonomic keys(Nelson, Tousson & Marasas, 1983; Samson & vanReenen-Hoekstra, 1988). In some cases (isolatesISCI f1, ISCI f2, ISCI f6, ISCI f10, ISCI f16, ISCIf29) the fungal identification was done or confirmedby Centraalbureau voor Schimmelcultures (Baarn,the Netherlands) (Table 1). Fungal strains weremaintained on PDA in the dark at 8 ± 2°C. Botrytiscinerea and Trichoderma spp. strains came from theISCI collection. Trichoderma strains were previouslyisolated from soil in northern Italy according to themethod of Elad, Chet & Henis (1981) and identifiedby the International Mycological Institute (Egham,Surrey, UK) (Table 1).
Choice of fungal strainsTable 1 reports the isolates chosen for in vitro
screening for flax-retting ability. Each isolate waschosen according to one or more of four criteria:documented flax-retting ability within the genus;high frequency of isolation from flax samples; highfrequency of isolation from soil samples; and knownability to produce pectinases or plant cell wall-degrading enzymes.
Alternaria spp. were also isolated with highfrequency from our dew-retted flax samples and haveflax retting ability (Sharma, 1986; Henriksson et al.,1997). However, these species were not included inthe screening because some Dematiaceous fungiwere previously indicated as responsible for fibrediscoloration in France (Anselme, 1960). Thisproblem was also observed recently in NorthernItaly.
Retting experimentsSeveral sporulating colonies of each strain were
obtained by incubating them on PDA for 2 wk undernear UV light or under natural light, depending onthe species. Inoculum was obtained by collectingspores with a sterile spatula and suspending them ina sterile water solution. E. nigrum was inoculatedas a mycelial fragment suspension. Flax stemsamples were inoculated with each strain by dippingthem for ten minutes in the spore suspension, with aconcentration varying from 1.5 to 2 ´ 106 spores ml-1.Each inoculated sample was laid in a Petri dish (3 gper plate) on a wetted sterile filter paper and sealedwith Parafilm. The trial was arranged in a completelyrandomised design with three replicates. The plateswere incubated at 24 ± 2°C under 12 h white lightphotoperiod. At the end of the incubation time, theretted stems were collected from each dish, dried inan oven at 40°C for 48 h, then stored in a plastic bagat room temperature until the evaluation tests.
In a preliminary experiment, carried out toestablish the optimum incubation time, nine isolateswere evaluated after 7, 19 and 38 days of incubation.On the basis of these first results, a secondexperiment with 23 isolates was performed with anincubation time of 15 days.
Retting degree evaluationMechanical test
The retting degree was evaluated first with a two-step mechanical test, designed to mimic as closelyas possible the breaking and scutching processesused industrially.
The first step (breaking) was carried out with abreaking-machine assembled in our laboratory,reproducing the model described by Bargale (1990).The machine consisted of two hand-driven pairs ofcounter-rotating fluted rollers. When stems passedthrough the rollers, the wooden pith and the bark

345Selection of flax-retting fungi
Fungal species Isolate code Origin References Aspergillus flavus Link c,d (ISCI f2) soil Samson & van Reenen-Hoekstra, 1988Aspergillus flavus Link c,d (ISCI f15) dew-retted flax Samson & van Reenen-Hoekstra, 1988Aspergillus niger v. Tieghemb,d (ISCI f1) dew-retted flax Annis & Goodwin, 1997Aspergillus niger v. Tieghem b,d (ISCI f18) dew-retted flax Annis & Goodwin, 1997Aspergillus niger v. Tieghem b,d (ISCI f28) soil Annis & Goodwin, 1997Botrytis cinerea Pers. a (ISCI f26) diseased plant tissues Bratt, Mercer & Brown, 1988Epicoccum nigrum Link a (ISCI f8) dew-retted flax Brown, 1984Fusarium culmorum (W.G. Smith) Sacc. a,d (ISCI f5) soil Sharma, 1986; Urbanek, Zalewska-
Sobczac & Krzechowska, 1976Fusarium oxysporum Schelect.b (ISCI f12) soilFusarium oxysporum Schelect. b (ISCI f13) dew-retted flaxFusarium semitectum Berk. & Rav. c,d (ISCI f11) soil Wood, 1971Fusarium solani (Mart.) Sacc. c,d (ISCI f4) soil Wood, 1971Myrothecium verrucaria (Albert. c
& Schw:Fries) (ISCI f16) soilMucor hiemalis Wehmer b (ISCI f6) dew-retted flaxMucor hiemalis Wehmer b (ISCI f10) dew-retted flaxMucor hiemalis Wehmer b (ISCI f27) soilPenicillium semplicissimum(Oudemans) Thom c,d (ISCI f29) dew-retted flax Yao, Conway & Sams, 1996Penicillium verrucosum complex c,d (ISCI f3) soil Yao et al., 1996Rhizopus stolonifer (Ehrenb.) Lind d (ISCI f7) soil Samson & van Reenen-Hoekstra, 1988Rhizopus stolonifer (Ehrenb.) Lind d (ISCI f8) dew-retted flax Samson & van Reenen-Hoekstra, 1988Trichoderma harzianum Rifai a (ISCI f21) soil Henriksson et al., 1997Trichoderma harzianum Rifai a (ISCI f20) soil Henriksson et al., 1997Trichoderma longibrachiatum Rifai a (ISCI f24) soil Henriksson et al., 1997a Documented flax-retting ability within the genus; bIsolated with relatively high frequency from flax samples; cIsolated with relativelyhigh frequency from soil samples; dWell known ability to produce pectinases or cell wall-degrading enzymes.
Table 1. Fungal strains screened for flax retting ability
were crushed into 2-3 mm pieces and removed, whilethe fibre remained intact. In the second step(scutching), the wood and bark pieces still attachedto the fibre were manually detached by scraping thefibre bundle with a knife-shaped woodenbladeboard, thus obtaining a clean fibre bundle.
The evaluation was preceded by a conditioningof the stem samples at 22°C and 60% r.h. The stemswere then weighed (total weight, WT) and passedthree times through the breaking machine.Subsequently, the samples including the fibre andthe residual part of the shive were weighed again(partially de-shived weight, W1). The fibre from thecrushed sample was then manually freed of theremaining cortex and woody parts still attached toit, and weighed again (fibre weight, W2).
Finally, the relative amount of shive detached bythe breaking action was calculated, and expressedas percentage of shive removal (SR):
SR (%) = [(WT � W1)/(WT � W2)] ´ 100
It was assumed that the more advanced rettingwas, the easier was removing shive by breaking. Theshive removal percentage was then taken as ameasure of degree of retting.
Fibre yield and lengthDuring breaking, fibres are subjected to a
mechanical stress, which causes many disruptions,thus reducing long-fibre yield and the average length.Fibre disruptions are likely to occur more frequently
when fibre bundles are excessively split into thinnerbundles or elementary fibres, or damaged by cellulaseactivity, which lowers their mechanical resistance.Long-fibre yield and length may then be consideredas a rough index of fibre degradation from over-retting. A high fibre yield and average lengthassociated with a high retting degree are indicativeof a good retting.
For fibre yield and length assessment, the scutchedfibre was bundled and hackled manually with a 1mm comb, from the central part towards each endof the bundle, to eliminate all the fibre shorter thanhalf the original length. Fibre yield was calculatedas follows:
Fibre yield = (long-fibre weight/total fibre weight)´ 100.
Average fibre length (AFL) was measured on thecombed bundle with a cutting-and-weighing method,based on van Langenhove & Bruggeman (1992). Thefibres were aligned at one end and paralleled, andthen they were cut into 20 mm sections with ascalpel. Each section was weighed on a precisionbalance.
The average fibre length was then calculated withthe following expression:
AFL (mm) = (åWi ´ 20)/WMAX
Where: Wi = weight of the ith section;WMAX = weight of the section with the maximum weight.

346 G FILA, L M MANICI & F CAPUTO
Uronic acid analysisThe retting degree determined mechanically was
associated with the dosage of the residual fibrepectins, evaluated as galacturonic acid, which is theprincipal component of pectic polymers.
A 200 mg fibre sample was finely cut into 0.5-1mm length pieces, that were first extracted in anEppendorf tube with 2 ml of chloroform/methanol(1:1) by shaking 24 h at room temperature, to removelipid components. After drying with supervacuum,total pectins were extracted with 200 ml of 5 g litre-1
Na2 EDTA for 2 h at 100°C. The product of extraction
was then analysed with the hydroxydiphenyl methodof Blumenkrantz & Asboe-Hansen (1973) asmodified by van den Hoogen et al. (1998), whoadjusted it for a 96 well microtitre plate. Thismodification was particularly useful for analysing alarge number of samples.
To each well of the plate was added 40 µl of theextraction solution and 200 µl of concentratedH2SO4. The plate was then incubated at 80°C for 60min. After cooling, the absorbance was read in amicrotitre-plate spectrophotometer at 540 nm.Subsequently, 40 µl of m-hydroxydiphenyl reagent(100 µl of 100 mg ml-1 m-hydroxydiphenyl indimethyl sulphoxide, mixed with 4.9 ml 80%sulphuric acid) were added, and after 15 min theabsorbance at 540nm was read again. Quantificationof uronic acid was made against a galacturonic acidstandard. The uronic acid content was expressed asmg of galacturonic acid g-1 of fibre.
The breaking test was done in both experimentsand uronic acid analysis in the second experimentonly.
The percentage of shive removal, after arcsinetransformation, and uronic acid data were subjectedto one-way analysis of variance and to Fisher�s LeastSignificant Difference (LSD) test using Statgraphics2.1 (Manugistics Inc., Rockville, MD, USA). To aidinterpretation, the percentage of shive removal andthe uronic acid values were subjected to Skott-Knott
cluster analysis (Gates & Bilbro, 1978), as suggestedby Madden, Knoke & Louie (1982).
The fibre yield and length values were clusteredusing the Nearest Neighbour clustering method onsquare Euclidean distances using the MultivariateMethods module of Statgraphics Plus version 1(Manugistics Inc., Rockville, MD, USA).
Results
Isolation of fungal strainsThe fungal populations in soil samples varied from
3 ´104 to 1.4 ´ 105 CFU g-1 soil; in flax samplesthey varied from 9 ´105 to 2.5 ́ 106 CFU g-1 fibre. F.solani, F. semitectum, Aspergillus spp. and M.verrucaria were the most frequent isolates from soilsamples while Alternaria spp. were the most frequentfungi (around 50%) found on dew-retted flax,followed by F. oxysporum, M. hiemalis and A. niger.
Preliminary experimentAll the fungi tested were able to colonise flax stem
pieces under controlled conditions. This preliminaryexperiment provided a first sorting of the rettingability of nine fungi (Table 2). After 7 days ofincubation the SR was significantly higher than thenon-inoculated control for all fungi. SR increasedat 19 days, and little or no further increment wasrecorded at 38 days. A period of two to three weekswas therefore considered sufficient to ret samplesin vitro. All of these fungi were included in thesecond experiment, except for Mucor sp. (ISCI f30)which was relatively ineffective at 38 days, andAspergillus flavus (ISCI f32), which was replacedby other Aspergillus strains.
Second experimentAll fungi were able to colonise flax samples in
vitro. Significant differences among treatments (P£ 0.01) were found both for SR and uronic acidcontent. Mean separation by LSD test (P = 0.05)
Table 2. Retting degree assessed with the breaking test (Shive Removal %) on flax samples retted in vitro after three incubationtimes. Each value is the mean of three replicates (SE = Standard Error)
Fungus Isolate code Retting time 7 days 19 days 38 days SR (%) SE SR (%) SE SR (%) SE
Control 47.5 4.92 51.2 3.20 48.6 2.75Aspergillus flavus (ISCI f2) 83.5 5.55 89.7 3.11 90.1 3.34Aspergillus flavus (ISCI f32) 64.7 6.78 90.3 5.62 90.6 1.54Trichoderma harzianum (ISCI f21) 80.0 4.58 94.1 1.48 95.9 1.54Penicillium semplicissimum (ISCI f29) 86.7 1.61 95.6 1.09 89.1 1.76Mucor hiemalis (ISCI f6) 79.4 5.07 87.7 0.74 89.9 4.19Mucor sp. (ISCI f30) 67.5 3.47 69.9 0.88 53.7 9.74Penicillium verrucosum (ISCI f3) 76.9 8.89 93.2 1.46 93.1 2.28Aspergillus niger (ISCI f1) 91.0 4.13 98.0 0.14 96.6 0.62Mucor hiemalis (ISCI f27) 82.6 9.34 90.3 1.33 84.3 1.86Mean (excluding control) 79.1 89.9 87.0

347Selection of flax-retting fungi
showed that the water-treated control differed fromall fungal treatments; there were also significantdifferences among the 23 isolates (data not shown).
Mean separation by Skott-Knott analysis of flax-retting efficiency as measured by SR and uronic acidcontent data allowed a clustering of strains. Thewater treatment in both cases fell in a cluster
separated from all the fungal treatments (Tables 3and 4).
All the A. niger and Penicillium spp. strains scoredthe highest values at the mechanical test, and fell incluster A, whereas A. flavus strains were in bothcluster A (ISCI f2) and in cluster B (ISCI f17) (Table3). The highest ranking position was also shared by
Fungus Isolate code SR (%) SE Clusters
A. niger (ISCI f1) 96.9 1.78 AA. flavus (ISCI f15) 96.9 1.28 AA. niger (ISCI f28) 96.6 0.99 AT. harzianum (ISCI f21) 95.9 0.20 AP. semplicissimum (ISCI f29) 95.9 0.70 AP. verrucosum (ISCI f3) 95.0 0.17 AR. stolonifer (ISCI f8) 95.0 0.02 AA. niger (ISCI f18) 95.0 0.28 AM. hiemalis (ISCI f6) 94.2 1.12 AA. flavus (ISCI f2) 93.7 0.44 BT. harzianum (ISCI f20) 93.3 1.76 BF. oxysporum (ISCI f12) 92.2 0.55 BF. oxysporum (ISCI f13) 90.6 0.85 BT. longibrachiatum (ISCI f24) 90.2 0.31 BM. hiemalis (ISCI f27) 89.9 3.78 BF. solani (ISCI f4) 87.7 1.50 CB. cinerea (ISCI f26) 89.9 1.26 BM. hiemalis (ISCI f10) 88.1 0.08 CR. stolonifer (ISCI f7) 83.4 1.73 CF. semitectum (ISCI f11) 81.4 1.17 CF. culmorum (ISCI f5) 81.3 1.59 CE. nigrum (ISCI f9) 79.3 1.31 DM. verrucaria (ISCI f16) 68.4 7.79 DControl 52.9 3.28 E
Table 3. Retting degree of flax samples retted in vitro with 23 fungal isolates. Each value, expressed as Shive-Removal percentage (see text), is the mean of three replicates (SE = Standard Error). Means were grouped with
Scott-Knott cluster analysis
Gal. acidFungus Isolate code (mg g-1 fibre) SE Clusters
Control 20.26 0.52 AT. longibrachiatum (ISCI f24) 17.26 0.26 BT. harzianum (ISCI f21) 17.26 0.15 BE. nigrum (ISCI f9) 13.56 0.37 CT. harzianum (ISCI f20) 13.27 0.07 CR. stolonifer (ISCI f7) 12.36 0.19 DM. hiemalis (ISCI f27) 12.27 0.11 DM. hiemalis (ISCI f10) 10.09 0.23 EM. verrucaria (ISCI f16) 9.54 0.00 FF. oxysporum (ISCI f13) 9.13 0.27 FF. oxysporum (ISCI f12) 8.61 0.71 FF. solani (ISCI f4) 7.76 0.25 GF. semitectum (ISCI f11) 5.73 0.47 HF. culmorum (ISCI f5) 5.4 0.36 HB. cinerea (ISCI f26) 4.93 0.11 IA. niger (ISCI f18) 4.83 0.38 IP. semplicissimum (ISCI f29) 4.41 0.17 IM. hiemalis (ISCI f6) 3.76 0.10 JP. verrucosum (ISCI f3) 3.43 0.18 JA. flavus (ISCI f2) 3.33 0.21 JA. flavus (ISCI f15) 2.8 0.18 KA. niger (ISCI f1) 2.63 0.06 KA. niger (ISCI f28) 2.55 0.06 KR. stolonifer (ISCI f8) 2.00 0.22 K
Table 4. Residual fibre pectins, expressed as galacturonic acid, of flax samples retted in vitro by 23 fungalisolates. Each value is the mean of three replicates (SE = Standard Error). Means were grouped with Scott-Knott
cluster analysis

348 G FILA, L M MANICI & F CAPUTO
T. harzianum (ISCI f21), R. stolonifer (ISCI f8)and M. hiemalis (ISCI f6) (Table 3). The stemsretted with Fusarium spp. strains showed a medium-low SR. F. oxysporum, which was the most frequentFusarium found on flax, showed the highest valueswith no difference between the soil and the flaxisolate. The other Fusarium strains showed amedium-low SR (cluster C). Finally, E. nigrum andM. verrucaria sp. treatments produced the worstresults at the mechanical test (Table 3).
Apart from A. flavus, significant differencesamong strains belonging to the same species werealso found in T. harzianum, R. stolonifer and M.hiemalis. All these species had one strain in clusterA, with others in cluster B (T. harzianum and M.hiemalis) or cluster C (R. stolonifer, M. hiemalis)(Table 3).
Uronic acid dosage provided a greaterdifferentiation among the assayed strains, whichwere separated into 11 clusters by Scott-Knottanalysis (Table 4). All Penicillium and Aspergillusstrains showed the lowest residual pectin values(Cluster K, J and I, Table 4), along with R. stolonifer(ISCI f8) and M. hiemalis (ISCI f6).
In the pectin-based sorting, the strains belongingto the same genus showed similar values. AllPenicillium and Aspergillus strains fell in the lowestclusters (I, J, K), Fusarium strains situatedthemselves at the intermediate positions (F, G, H),while the three Trichoderma and E. nigrum showedthe highest residual pectin values apart from thecontrol (Clusters B and C, Table 4).
As expected, an easier mechanical decorticationtended to be associated with low levels of residualpectins. This was particularly evident for Aspergillusspp., Penicillium spp., R. stolonifer (ISCI f8) andM. hiemalis (ISCI f6); while Fusarium showedintermediate values among the assayed strains inboth tests (Tables 3 and 4). Nevertheless, there wasa poor correlation (R = -0.55) between SR and uronicacid, since some strains did not share this tendency.In particular, two Trichoderma strains (ISCI f21;ISCI f20) ranked high at the breaking test, but had ahigh content of residual pectins (Tables 3 and 4).
Fibre degradation from over-retting wasrepresented by the combined reduction in long-fibreyield and length (Fig. 1). After having chosen a five-clusters data separation, it was possible to identifya group containing the majority of strains with littleor no degradation, where fibre yield was higher than83% and length was higher than 85% of the originallength (Fig. 1). Only four strains fell outside thisgroup; E. nigrum, M. verrucaria and F. semitectum,which were also low-retting strains, and P.verrucosum which had a higher retting ability.
Discussion
Most of the fungal species isolated in this studyfrom flax and soil (Table 1) belonged to generaalready observed on dew-retted flax by otherresearchers, who indicated them as dew-rettingagents.
The retting action of Aspergillus sp., Penicilliumsp., Mucor spp., Rhizopus spp., F. culmorum, E.nigrum and B. cinerea has been reported in NorthernEurope (Arthaud, 1965; Brown & Sharma, 1984;Rynduch, 1989). Trichoderma sp., Fusarium spp.and E. nigrum isolated from dew-retted flax inEurope and the USA showed variable ability to retflax in vitro (Brown, 1984; Henriksson et al., 1997).
A crucial step in the screening for retting abilitywas the evaluation of degree of retting, which is notstraightforward. Over the years several methods havebeen proposed, none of which has reached the statusof standard or widespread use. The variousapproaches were based on a more or less empiricalsubjective assessment (Greenhill & Conchman,1947; Dujardin, 1948), or evaluated the ease withwhich fibres were detached from the wood core(Seaby & Mercer, 1984). Fischer & Topf (1988)analysed the firmness of stems. Other methodsconsidered chemical parameters, such as the causticweight loss test (Brown, Sharma & Black, 1986),the cation exchange capacity (Morvan, Morvan,Jauneau & Demarty, 1985) or the residual pectincontent (Meijer et al., 1995). In our approach, wepreferred to adopt a conventional breaking test, withthe aim of simulating as closely as possible the workeffectively done at an industrial level to extract thefibres.
This method has some important advantages. It isin itself a direct measurement of retting and has anoperational meaning. The rapidity and ease ofexecution allow processing of a great number ofsamples in a short time. The greatest limit is the lackof linearity. The wood core represents the largestmass fraction of the flax stem. Its removal bymechanical breaking immediately raises the SR, andeven a poorly retted stem shows values around 70%.The retting process also consists of the separationof the fibres from the other bast tissues. This phasetakes more time than the detachment of the bast fromthe wood core, but has little effect on SR. This partlyexplains the low correlation coefficient between thisparameter and pectin content. However, a selectionof the best retting fungi on the basis of mechanicaldata alone, by taking for example the cluster A inTable 3, would have chosen eight out of 10 of thehighest (uronic acid lower than 5 mg g-1 fibre) pectin-degrading strains, clusters K, J, I of Table 4. Thisdemonstrates that from a practical point of view, thebreaking test could be useful for a first screening ofa wide range of samples, to eliminate the ineffective
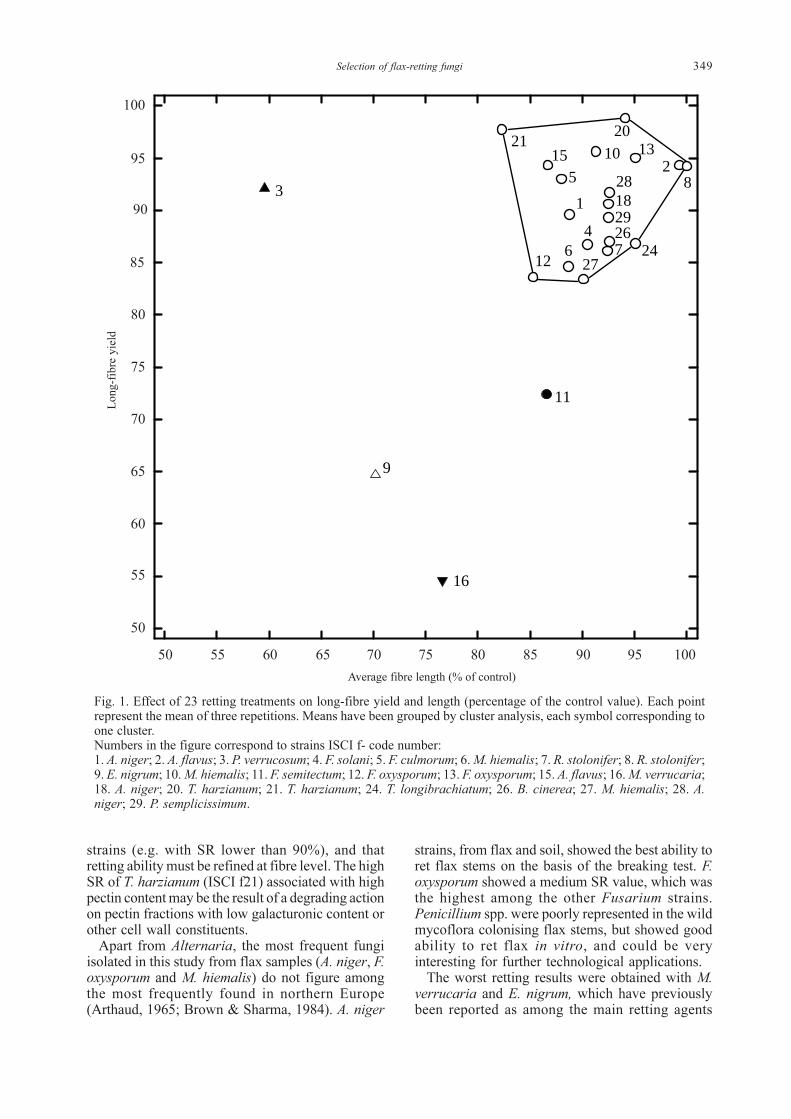
349Selection of flax-retting fungi
strains (e.g. with SR lower than 90%), and thatretting ability must be refined at fibre level. The highSR of T. harzianum (ISCI f21) associated with highpectin content may be the result of a degrading actionon pectin fractions with low galacturonic content orother cell wall constituents.
Apart from Alternaria, the most frequent fungiisolated in this study from flax samples (A. niger, F.oxysporum and M. hiemalis) do not figure amongthe most frequently found in northern Europe(Arthaud, 1965; Brown & Sharma, 1984). A. niger
strains, from flax and soil, showed the best ability toret flax stems on the basis of the breaking test. F.oxysporum showed a medium SR value, which wasthe highest among the other Fusarium strains.Penicillium spp. were poorly represented in the wildmycoflora colonising flax stems, but showed goodability to ret flax in vitro, and could be veryinteresting for further technological applications.
The worst retting results were obtained with M.verrucaria and E. nigrum, which have previouslybeen reported as among the main retting agents
50 55 60 65 70 75 80 85 90 95 100
Lon
g-Fi
bre
yiel
d
50
55
60
65
70
75
80
85
90
95
10020
28
1310
281829
24267
155
1
46
2712
21
11
3
9
16
50 55 60 65 70 75 80 85 90 95 100
100
95
90
85
80
75
70
65
60
55
50
Average fibre length (% of control)
Lon
g-fi
bre
yiel
d
Fig. 1. Effect of 23 retting treatments on long-fibre yield and length (percentage of the control value). Each pointrepresent the mean of three repetitions. Means have been grouped by cluster analysis, each symbol corresponding toone cluster.Numbers in the figure correspond to strains ISCI f- code number:1. A. niger; 2. A. flavus; 3. P. verrucosum; 4. F. solani; 5. F. culmorum; 6. M. hiemalis; 7. R. stolonifer; 8. R. stolonifer;9. E. nigrum; 10. M. hiemalis; 11. F. semitectum; 12. F. oxysporum; 13. F. oxysporum; 15. A. flavus; 16. M. verrucaria;18. A. niger; 20. T. harzianum; 21. T. harzianum; 24. T. longibrachiatum; 26. B. cinerea; 27. M. hiemalis; 28. A.niger; 29. P. semplicissimum.

350 G FILA, L M MANICI & F CAPUTO
(Brown, 1984). The weakness of the fibres obtainedwith E. nigrum treatment confirmed that it attackscellulosic fibres causing over-retting and reducingfibre quality, as observed also in other studies(Henriksson et al., 1997; Brown, 1984). Among thelow retting performances, the treatments withFusarium from soil (F. culmorum, F. semitectum andF. solani) cannot be included in the list of fungi withgood retting performance, even if they, together withother fungi not considered in our study, may play asecondary role in field retting.
In our model, a good quality retting process ischaracterised by easy extraction of fibres and lowresidual pectins, along with low fibre degradation.According to the results, this condition is satisfiedby the three strains of A. niger (ISCI f1, ISCI f18and ISCI f28), one A. flavus strain (ISCI f15) and R.stolonifer (ISCI f8). A. niger in particular, ischaracterised by a well-known pectinolytic activity(Thibault, 1983; Annis & Goodwin, 1997) and haslong been recognised as a retting agent (De Franca,Rosemberg & De Jesus, 1969). Well-retted sampleswere also obtained with the other Penicillium andAspergillus strains (ISCI f29, ISCI f3, ISCI f2) andM. hiemalis (ISCI f6).
It is interesting to point out the widely differentresults observed between M. hiemalis and R.stolonifer, two Zygomycetes with world-widedistribution that are frequently isolated from soil,grain, fruits, etc. Several species of Mucor andRhizopus are industrially important (Samson & vanReenen-Hoekstra, 1988; Akin et al., 1998), and thissuggests that these two good-retting strains couldbe used as fermenting inoculum to produce rettingenzymes. The variability observed in this screeningindicates that a larger survey within those twospecies may find strains with a retting efficiencysimilar to or higher than that observed for A. niger.
In conclusion, the in vitro screening reported inthis study made it possible to identify some strainswith high ability to ret flax among a wide set ofisolates from the wild mycoflora of northern Italy.The high flax retting activity of the A. niger, M.hiemalis and R. stolonifer strains naturally presenton dew retted flax suggests the possibility of rettingflax directly in the field. In unfavourableMediterranean climates, this could be achievedthrough the control of some environmentalconditions, for example by retting at different timesof the year, or with artificial watering (Cavallero,Reyneri & Rondi, 1997), or pre-harvest retting withherbicide treatment (Sharma, 1986). The presencein northern Italy of filamentous fungi with goodretting ability such as Penicillium spp, A. flavus,Mucor hiemalis, Rhizopus stolonifer, T. harzianumISCI f21 and others (Table 3) suggests the possibilityof using these as artificial inoculum to accelerateand enhance dew-retting in the field. Moreover, this
screening widens the availability of retting strainsfor flax-specific enzyme production to be used inhigh-quality warm water retting (Henriksson, Akin,Slomczynski & Eriksson, 1999).
Acknowledgements
We wish to thank Dr C Cerato, of the Crop Biology& Protection section of ISCI, who kindly suppliedall the Trichoderma strains utilised in this work.
References
Akin D E, Rigsby L L, Henriksson G, Eriksson K E L. 1998.Structural effects on flax stem of three potential retting fungi.Textile Research Journal 68:515-519.
Annis S L, Goodwin P H. 1997. Recent advances in themolecular genetics of plant wall-degrading enzymes producedby plant pathogenic fungi. European Journal of PlantPathology 103:1-14.
Anselme I C. 1960. Problemes liés au rouissage à terre du linen France. Étude preliminaire. Fibra 5:83-87.
Arthaud J. 1965. Le rouissage du lin. In Institute TecniqueAgricole du Lin, pp. 1-21. Paris, France: INRA.
Bargale P C. 1990. Development and testing of a manual flax-scutching machine. Biological Wastes 32:29-37.
Blumenkrantz N, Asboe-Hansen G. 1973. New Method forQuantitative Determination of Uronic Acids. AnalyticalBiochemistry 54:484-489.
Bratt R P, Mercer P C, Brown A E. 1988. Degradation of flaxstem by Botrytis cinerea. Transactions of the BritishMycological Society 90:537-544.
Brown A E. 1984. Epicoccum nigrum, a primary saprophyteinvolved in the retting of flax. Transactions of the BritishMycological Society 83:29-35.
Brown A E, Sharma H S S. 1984. Production of polysaccharide-degrading enzymes by saprophytic fungi from glyphosate-treated flax and their involvement in retting. Annals of AppliedBiology 105:64-74.
Brown A E, Sharma H S S, Black D L R. 1986. Relationshipbetween pectin content of stems of flax cultivars, fungal cellwall-degrading enzymes and pre-harvest retting. Annals ofApplied Biology 109:345-351.
Cavallero A, Reyneri A, Rondi G. 1997. Fibre flax productionand quality in Northern Italy. II. Comparison between dewretting and retting methods controlled by field irrigation.Italian Journal of Agronomy 1:81-87.
Cremaschi D, Maestrini C. 1996. Varietà di lino da fibra inambiente padano. L�Informatore Agrario 45:48-52.
De Franca, F P, Rosemberg J A, De Jesus A M. 1969. Rettingof flax by Aspergillus niger. Applied Microbiology 17:7-9.
Dujardin A. 1948. The Retting of Flax. Flax DevelopmentCommittee. Belfast, UK: Carswell and Son. 194 pp.
Elad Y, Chet I, Henis J. 1981. A selective medium for improvingquantitative isolation of Trichoderma spp. from soil.Phytopathology 74:1171-1175.
Fischer K, Topf W. 1988. Entwicklung objektiverQualitätskriterien fur Flachs. Teil 2:TextiltechnologischeUntersuchungen. Melliand Textilberichte 12:858-861.
Frederiksen P S. 1967. Practical investigation of field rettingof fibre flax, yield and quality problems. Fibra 12:3-17.
Gates C E, Bilbro J D. 1978. Illustration of cluster analysismethod for mean separation. Agronomy Journal 70:795-797.
Greenhill W L, Conchman J F. 1947. The water-retting of flax.Résumé of investigations from 1940 to 1945. Commonwealthof Australia � Council for Scientific and Industrial Research.Bulletin 211.

351Selection of flax-retting fungi
Seaby D A, Mercer P C. 1984. Development of a hand tool totest the degree of retting of flax-straw. Annals of AppliedBiology 104:567-573.
Sharma H S S. 1986. An alternative method of flax rettingduring dry weather. Annals of Applied Biology 109:605-611.
Sharma H S S, van Sumere C F. 1992. Enzyme treatment offlax. The Genetic Engineer and Biotechnologist 12:19-23.
Thibault J F. 1983. Études structurale de substances peptiquesà l�aide d�une endopolygalacturonase d�Aspergillus niger:purification, proprieté et possibilités d�utilisation de cetteenzyme. Thèse Doctorat, Nantes.
Urbanek H, Zalewska-Sobczac J, Krzechowska M. 1976.Exo-polygalacturonate lyase of Fusarium culmorum. Bulletinde l�Academie Polonaise de Sciences 24:635-639.
van den Hoogen B M, van Weeren P R, Lopes-Cardozo M,van Golde L M G, Barneveld A, van de Lest C H A. 1998.A microtiter plate assay for the determination of uronic acids.Analytical Biochemistry 257:107-111.
van Langenhove L, Bruggeman J P. 1992. Methods of fibreanalysis. In The Biology and Processing of Flax, pp. 311-328. Eds H S Sharma and C F Van Sumere. Belfast, UK: MPublications.
Wieringa K T. 1956. The micro-organisms decomposing pecticsubstances in the dew retting process of flax. NetherlandsJournal of Agricultural Science 4:16-21.
Wood T M. 1971. The cellulase of Fusarium solani. Purificationand specificity of the (1-4)-glucanase and the ß-D-glucosidasecomponents. Biochemical Journal 121:353-362.
Yao C, Conway W S, Sams C E. 1996. Purification andcharacterization of a polygalacturonase produced byPenicillium expansum in apple fruit. Phytopathology 86:1160-1166.
Henriksson G, Akin D E, Slomczynski D, Eriksson K-E L.1999. Production of highly efficient enzymes for flax rettingby Rhizomucor pusillus. Journal of Biotechnology 68:115-123.
Henriksson G, Akin D E, Hanlin R T, Rodriguez C, ArchibaldD D, Rigsby L L, Eriksson K-E L. 1997. Identification andretting efficiencies of fungi isolated from dew-retted flax inthe United States and Europe. Applied and EnvironmentalMicrobiology 63:3950-3956.
Madden L V, Knoke J K, Louie R. 1982. Considerations forthe use of multiple comparison procedures inphytopathological investigations. Phytopathology 72:1015-1017.
Meijer W J M, Vertregt N, Rutgers B, van de Waart M. 1995.The pectin content as a measure of the retting and rettabilityof flax. Industrial Crops and Products 4:273-284.
Morvan C, Morvan O, Jauneau A, Demarty M. 1985.Capacité d�échange cationique des fibres de lin. ComptesRendus de l�Academie d�Agriculture de France 71:837-841.
Nelson P E, Tousson TA, Marasas W F O. 1983. Fusariumspecies. An illustrated manual for identification. USA: ThePennsylvania State University Press. 190 pp.
Reyneri A, Cavallero C. 1997. Fibre flax production and qualityin Northern Italy: I. Effects of environment, seeding rate andnitrogen fertilization. Italian Journal of Agronomy 1:73-79.
Rynduch W. 1989. Accelerating of flax dew retting. In Flax inEurope. Production and processing, Proceedings of theEuropean Regional Workshop on Flax. Poznan, Poland, 19-21 June 1989, pp. 133-135.
Samson R A, van Reenen-Hoekstra E S. 1988. Introductionto food-borne pathogens. 3rd Edn. Centraalbureau vorSchimmelcultures. 299 pp.

352 G FILA, L M MANICI & F CAPUTO