immunohistochemical co-localization of placental lactogen ii and relaxin in the golden hamster...
TRANSCRIPT

1 Supported by NIH grant HD.14966 (FT).
2 Correspondence to: Dr. RH. Renegar. Department of Anatomy and
Cell Biology, School ofMedicine, East Carolina University, Greenville, NC
27858.
935
0022-1554/90/$3.30
The Journal of Histochemistry and CytochemistryCopyright © 1990 by The Histochemical Society, Inc.
Vol. 38, No. 7, pp. 935-940, 1990
Printed in USA.
Original Article
Immunohistochemical Co-localization of Placental
Lactogen II and Relaxin in the Golden Hamster
(Mesocricetus a ura tus)’
RANDALL H. RENEGAR,2 JONATHAN N. SOUTHARD, and FRANK TALAMANTES
Department of Anatomy and Cell Biology, School of Medicine, East Carolina University, Greenville, North Carolina (RHR,);
and Department ofBiology, University of California, Santa Cruz, California (JNS,FT).
Received for publicationjuly 31, 1989 and in revised formJanuary 31, 1990; accepted February 2, 1990 (9A1749).
Two hormones with lactogenic activity are produced by thehamster placenta during the second halfofpregnancy. One
of these hormones, hamster placental lactogen II (haPL-II),has been well characterized; however, its cellular source is
not known. In the present study, haPLrII was localized inplacental tissues using a specific antibody and the avidin-bio-tin-peroxidase immunohistochemical technique. Becauserelaxin has been localized in the hamster placenta, it was
of interest to determine if haPL-II and relaxin are localizedin the same cells. haPL-II immunoactivity was observed in
primary and secondary giant trophoblast cells ofthe placentaon Days 12, 14, and 15 of pregnancy. On Day 15 positive
Introduction
Lactogenic activity in placental tissue extracts and blood from preg-
nant hamsters was first reported by Kelly et al. (1976). Serum (Kelly
et al. , 1976) and tissue (Soares and Talamantes, 1982) lactogenic
activity in the hamster is detectable beginning at midpregnancy
(Day 8). Serum concentrations increase to term (Day 16) and placen-
tal tissue concentrations increase to Day 14 but decrease thereafter
(Soares and Talamantes, 1982). Two proteins designated hamster
placental lactogen I (haPL-I, MW 37,000) and haPL-II (MW 22,000)
account for most of the lactogenic activity in serum and placental
extracts during the latter halfofpregnancy in this species (Southard
et al., 1987; Southard et al., 1986). The haPL are so designated
because of their similarity in structure to PL found in the mouse
(Jackson et al. , 1986; Soares et al. , 1983) and rat (Duckworth et
al., 1982; Robertson et al., 1982). Placental lactogen II is the most
extensively characterized ofthe hamster hormones (Southard et al.,
1986), and recently a specific antibody and a homologous radio-
immunoassay were developed (Southard and Talamantes, 1987).
Hamster placental lactogen II is first detected in serum on Day 10
staining was also observed in large cells located withinmesometrial arteries and in eosinophilic bodies associatedwith degenerating sheathed arteries of the decidua basalis.haPL-II-positive staining was not observed in placentae fromDays 8 or 10 of pregnancy. On Day 14, haPL-II was co-localized with relaxin in 75#{176}/oof the giant trophoblast cells
observed. Therefore, it is probable that these hormones are
synthesized and secreted by the same cell. (J HiswchemCyrochem 38:935-940, 1990)
KEY WORDS: Placenta; Placental lactogen; Hamster; Immuno-
histochemistry; Relaxin.
ofgestation, and concentrations increase to term (Southard et al.,
1987).
The objective ofthe present study was to immunolocalize haPL.II
in the hamster placenta throughout the latter half of gestation.
Because the placenta is the source ofrelaxin in the pregnant ham-
ster (Steinetz et al. , 1988; Renegar et al. , 1987), we also examined
whether haPL-II and relaxin are localized within the same placen-
tal cells.
Materials and Methods
Animals and Tissue Preparation. Pnimiparous female Golden (Syrian)
hamsters (120-150 g; Charles River, Kingston, NY) were maintained in the
animal housing facility of the Department of Comparative Medicine on
a 12L:12D (lights on at 0700 hn) schedule. Animals were checked daily (be-
tween 2000 and 2200 hr) for estrus, as determined by expression of lordosis
in the presence of a male. Females in estrus were housed overnight with
a male and the following day was designated Day 1 of gestation. Animal
maintenance and handling were performed in accordance with the institu-
tion’s guidelines for care and use oflaboratory animals. Animals were anesthe-
tized with a combination ofxylazine (Haven-Lockhart; Shawnee, KS) andketamine HCI (Bristol Laboratories; Syracuse, NY) and the reproductive
tract exposed through a midventral incision. Placentae were recovered onDays 8, 10, 12, 14, and 15 ofgestation (n = 3 or 4 animals/day) between
0800 and 1200 hr, except on Day 15 when tissues were removed between
1900 and 2100 hr to obtain tissues immediately before partunition (Day
by guest on March 16, 2016jhc.sagepub.comDownloaded from

936 RENEGAR, SOUTHARD, TALAMANTES
Hamster placental lactogen Ii immunoreactivity was not observed
Figure 1. Cross-section ofan implantation site on Day 12 ofgestation. The chonioallantoic placenta is well developed. The trophospongium (ts), which is composedof fetal trophoblast cells, including the secondary giant trophoblasts, has invaded the decidua basalis (db). The placental labyrinth (Ib), where nutrient exchange
between mother and fetus occurs, develops by extension of fetal vessels into the trophospongium. Primary giant trophoblast cells are located within the reticularmatrix (.) at the peripheral margins of the discoid placenta. A central placental artery (arrowheads) and peripheral venous channels course through the deciduaand mesometnium (mm). m, muscularis; ys, yolk sac; f, fetus. (Inset) High magnification of the decidual portion of a central artery. These arteries are characterizedby their sheath of connective tissue cells and minimal smooth muscle. A sheath forms around mesometnial arteries as pregnancy progresses. Eosin and hematoxy-in stained. Bar � 200 am; inset � 50 �tm.
16 = day ofpantunition). Tissues were fixed by immersion in Bouin’s fluid,
dehydrated, and embedded in paraffin. Tissues from Days 10 and 14 were
also fixed in Zamboni’s, Bouin-Hollande’s, or Carnoy’s solution to exam-inc the influence of fixation on immunostaining for haPL-II. Seven-�tm
sections were placed on glass slides for immunohistochemistry or for he-
matoxylin and eosin staining.
lmmunohistochemistry. Hamster placental lactogen II and relaxin were
localized in placental tissue sections using the avidin-biotin-peroxidase
technique (Vectastain ABC kit; Vector Laboratories, Burlingame, CA) as
previously described (Renegar et al., 1987). The antiserum to haPL-Il has
been previously used to develop a homologous haPL.II radioimmunoassay
and does not crossreact significantly with hamster prolactin or growth hon.
mone (Southard and Talamantes, 1987). The rabbit antiserum to porcine
relaxin (UF-1M.; kindly provided by Dr. Mj. Fields. University of Florida)
has been previously used to localize relaxin in the hamster placenta (Rene-
garet al., 1987). Diluted antisera to haPL.Il (U10,000) and relaxin (12500)
were incubated on tissue sections for 24 hr at 4C. Bound avidin-biotin-per-
oxidase complex was localized by incubating sections in a solution of 0.5%
(w/v) 3,3’-diaminobenzidine, 0.02% hydrogen peroxide in 0.1 M Tnis (hy-
droxymethyl) amino methane (Tnis) buffer (pH 7.2). Completed slides were
counterstained with hematoxylin.
Method specificity for haPL.Il staining was assessed (a) by treating ad-
jacent sections with specific antiserum and non-immune rabbit serum and
(b) by omission of individual components of the staining procedure. Posi-
tive staining was present only when specific antiserum and all components
of the staining procedure were used. Specificity of the antiserum for its
antigen was assessed by (a) exhaustive dilution to demonstrate the pres-
ence ofnonspecific low-titer antibodies and (b) absorption with each anti-
gen. Absorption ofhaPL.II-specific antiserum overnight with purified haPIAI
(0.15 riM) (Southard et al., 1986) but not porcine relaxin (NIH.RXN-P1;
2.1 saM) eliminated all positive staining. Likewise, absorption of relaxin.
specific antisera with porcine relaxin (1.0 �sM) but not haPL-II (1.69 �tM)
eliminated all positive staining.
Estimate of Co-localization. To evaluate haPL-II and relaxin co-local-ization in the hamster placenta, identical areas of adjacent tissue sections
from Day 14 ofgestation were stained for haPL.II or relaxin, photographed,
and printed (total magnification x 254). Immunostained cells (haPL.II
or relaxin) with the nucleus present in both sections were counted. Cells
identified in photomicrographs were confirmed by direct microscopic ob-
servation of tissue slides. Three or four microscopic fields (189 cm2) from
each of four different placentae were examined.
Results
by guest on March 16, 2016jhc.sagepub.comDownloaded from
..� .
.�.... 4 ij�
� ..
� ‘� � . .,it1�Z�Vf. . � � ,
.- �
. .5
- � .-.-.- ‘..,� .� -v,. � � - �
, � V �.�
; �
haN1-II IMMUNOLOCALIZATION
s.s):;,.. , lb ,
�
;:� � . a .
,. -...,.; . -. ..
‘5 a _ ‘-
fi .::� �- � �
. - #{149}�;�.#{149} .,._
I #{149}4 W4.’..’, ‘.. �#{149}�.4f�. -
� ,;,,,� � #{149}
. ‘,,##{149} �,- ,.Op_. .4�
.‘. � . . F k ‘
.�. ._.4. p�.#{248}ay ‘
a � ‘5 ‘#{149}‘“� ..,.
a � . . .
937
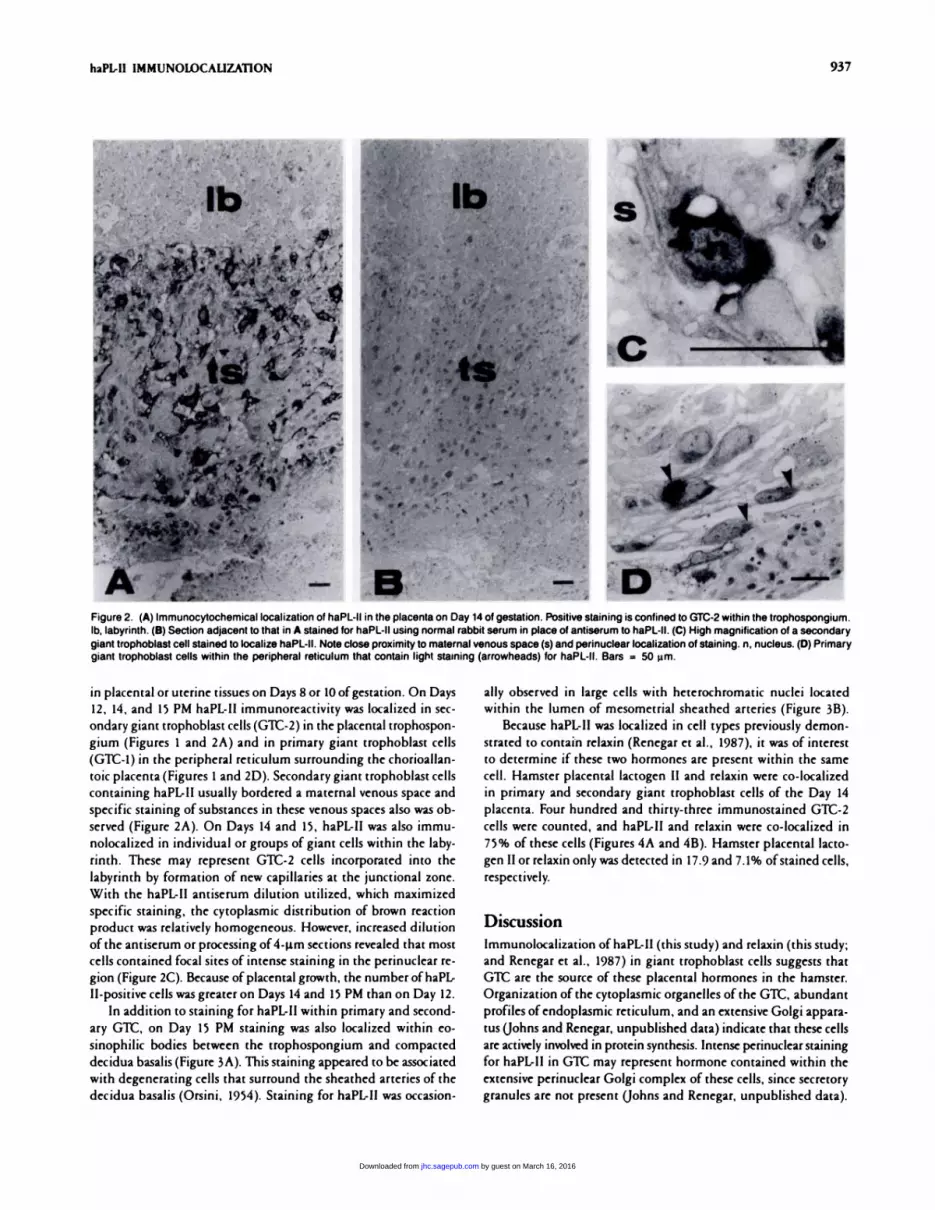
Figure 2. (A) Immunocytochemical localization of haPL-lI in the placenta on Day 14 of gestation. Positive staining is confined to GTC-2 within the trophospongium.Ib, labyrinth. (B) Section adjacent to that in A stained for haPL-II using normal rabbit serum in place of antiserum to haPL-Il. (C) High magnification of a secondary
giant trophoblast cell stained to localize haPL-II. Note close proximity to maternal venous space (s) and perinuclear localization of staining. n, nucleus. (D) Primarygiant trophoblast cells within the peripheral reticulum that contain light staining (arrowheads) for haPL-Il. Bars = 50 �m.
in placental or uterine tissues on Days 8 or 10 ofgestation. On Days
12, 14, and 15 PM haPL-Il immunoreactivity was localized in sec-
ondary giant trophoblast cells (G’Lt-2) in the placental trophospon-
gium (Figures 1 and 2A) and in primary giant trophoblast cells
(GTC-1) in the peripheral reticulum surrounding the chonioallan-
toic placenta (Figures 1 and 2D). Secondary giant trophoblast cells
containing haPL-II usually bordered a maternal venous space and
specific staining ofsubstances in these venous spaces also was ob-
served (Figure 2A). On Days 14 and 15, haPL-II was also immu-
nolocalized in individual or groups of giant cells within the laby-
ninth. These may represent Glt-2 cells incorporated into the
labyrinth by formation of new capillaries at the junctional zone.
With the haPL-II antiserum dilution utilized, which maximized
specific staining, the cytoplasmic distribution of brown reaction
product was relatively homogeneous. However, increased dilution
ofthe antiserum or processing of4-�.tm sections revealed that most
cells contained focal sites of intense staining in the peninuclear ne-
gion (Figure 2C). Because of placental growth, the number of haPL-
Il-positive cells was greater on Days 14 and 15 PM than on Day 12.
In addition to staining for haPL-II within primary and second-
ary GTC, on Day 15 PM staining was also localized within eo-
sinophilic bodies between the trophospongium and compacted
decidua basalis (Figure 3A). This staining appeared to be associated
with degenerating cells that surround the sheathed arteries of the
decidua basalis (Orsini, 1954). Staining for haPL-II was occasion-
ally observed in large cells with hetenochromatic nuclei located
within the lumen of mesometnial sheathed arteries (Figure 3B).
Because haPL-II was localized in cell types previously demon-
strated to contain relaxin (Renegar et al. , 1987), it was of interest
to determine if these two hormones are present within the same
cell. Hamster placental lactogen II and relaxin were co-localized
in primary and secondary giant trophoblast cells of the Day 14
placenta. Four hundred and thirty-three immunostained GTC-2
cells were counted, and haPL-II and relaxin were co-localized in
75% of these cells (Figures 4A and 4B). Hamster placental lacto-
gen II or relaxin only was detected in 17.9 and 7.1% ofstained cells,
respectively.
Discussion
Immunolocalization of haPL-II (this study) and relaxin (this study;
and Renegar et al. , 1987) in giant trophoblast cells suggests that
GTC are the source of these placental hormones in the hamster.
Organization of the cytoplasmic organdIes of the GTC, abundant
profiles ofendoplasmic reticulum, and an extensive Golgi appara-
tus Uohns and Renegar, unpublished data) indicate that these cells
are actively involved in protein synthesis. Intense peninuclear staining
for haPL.lI in Git may represent hormone contained within the
extensive peninuclear Golgi complex of these cells, since secretory
granules are not present Uohns and Renegar, unpublished data).
by guest on March 16, 2016jhc.sagepub.comDownloaded from

C.
‘1 v�’�4�
�e:i., �
4
‘5
A
SI
� �
.� .,,
�
V,. � �-4. _
Figure 4. (A) Day 14 placental tissue stained to localize haPL-Il. (B) Section adjacent to that in A stained to localize relaxin. Note co-localization in cells (arrow-heads) adjacent to the maternal venous space (s). Bars = 50 �m.
938 RENEGAR, SOUTHARD, TALAMANTES
,� 101” # �: � -� #{149}
1r, � � , �.,;.., � “;
4 �. �ti � , �,‘t ;;��‘ ,
... .� t � ‘-.- : � � r� ‘-
��:�:; ,�, , � � �
k,� � b 1� � #{149}
.4% ,,#{149} � �; #{149}( S , #{149}, � � ‘
.., � ,jt -�. �, � iq-. ‘ .‘ ,.v�, ;-� �%- �
‘‘�. � f -.� .- � . p� � .�. . . #{149}: ,�,#{149} d.,’�5�t’ �
�W;4#{149} � �s . #{149}::Sd�” .a� �
Figure 3. Portion of a Day 15 placenta immunostained to localize haPL-ll. (A) Note staining within cells and maternal venous spaces of the trophospongium (ts).
db, decidua basalis. Staining is also associated with eosinophilic bodies in degenerating sheath cells of a decidual artery (a). (B) Portion of a mesometrial sheathedartery with staining located in large cells adjacent to the vessel lumen (.). Bars = 50 �tm.
Giant trophoblast cells of the mouse placenta also contain immu-
noactive placental lactogen (Hall and Talamantes, 1984). Placental
lactogen has been localized in binucleate cells of the fetal choni-
onic epithelium and the syncytial layer ofthe placentomes of sheep
(Wooding, 1981; Reddy and Watkins, 1980; Watkins and Reddy,
1980; Carnegie et al., 1978; Martal et al., 1977). Immunoreactive
human placental lactogen (Watkins, 1978; de Ikonicoffand Cedard,
1973) and mRNA for human placental lactogen (Hoshina et al.,
1985; Boime et al., 1982; Hoshina et al., 1982; McWilliams and
Boime, 1980) have been demonstrated in the syncytium ofthe hu-
man placental villus.
Hamster placental lactogen II is first detectable in serum and
placental tissue extracts on Day 10 of gestation (Southard et al.,
1987). The reason that haPL-II immunoreactivity in placental tis-
sue sections was not detected on Day 10 is unknown. Watkins (1978)
reported that formalin fixation reduced the antigenicity of PL in
human villus samples but that fixation by dehydration using Car-
noy’s solution allowed for successful immunolocalization. Demon-
stration of haPL-II in tissue sections on Day 10 was unsuccessful
regardless of the fixative. Low serum hormone concentrations
(Southard et al., 1987) may reflect low rates of hormone synthesis
in trophoblast cells on Day 10. It is possible that the quantity of
.w
by guest on March 16, 2016jhc.sagepub.comDownloaded from

haPL-II IMMUNOLOCALIZATION 939
hormone within individual cells on Day 10 is below the sensitivity
of the methodology used.
Onsini (1954) concluded that primary and secondary giant
trophoblast cells of the hamster placenta are phagocytic and that
these cells may function in penetration ofthe uterine epithelium,
enlargement of the decidual cavity, and degeneration of maternal
tissue during implantation and placental growth. Association of
haPL-II with these cells may indicate a role for this hormone in
placental growth and development. Hamster placental lactogen II
binds to prolactin receptors in the mouse mammary gland and
stimulates a-lactalbumin synthesis by these same tissues (Southard
et al. , 1986). Therefore, haPL-II may have a role in lactogenesis in
the hamster.
Arteries that supply the hamster placenta are distinguished by
their prominent sheaths (Orsini, 1954). These arteries have an en-
dothelium and a single layer ofmuscle surrounded by layers of hyper-tnophied connective tissue cells. Sheathed arteries are located within
the decidua basalis early in pregnancy, and mesometnial arteries
undergo this transformation later in pregnancy. Near the time of
partunition, sheath cells surrounding arteries ofthe decidua basalis
undergo degeneration to form eosinophilic bodies. Although these
structures contained haPL-II immunoreactivity, the significance of
this observation is unknown.
Orsini (1954) described two types oflarge cells, the tertiary gi-
ant trophoblast cells and the endovascular cells, which reside within
the lumen ofmesometrial arteries during late pregnancy. Both cells
are of fetal origin; however, their function is not known. Endovas-
culan cells predominate at this stage of pregnancy and are probably
those stained for haPL-II in this study. The significance of haPL-II
staining in these cells is unknown.
Specificity control experiments demonstrated that staining with
each antiserum was specific for the antigen in question. Additional
evidence ofantibody specificity is implied by the fact that haPL-II
positive staining was not observed on Days 8 and 10 when relaxin
immunoreactivity is present (Renegar et al. , 1987). Data from this
study indicate that haPL-II and relaxin are localized within and
probably synthesized and secreted by the same cell. Localization
of a single hormone in any given cell is likely owing to limitations
of the techniques used to determine co-localization. Staining in-
tensity for haPL-Il was greater than that for relaxin and may ac-
count for the detection ofcells stained only for haPL-II. This differ-
ence in staining intensity may be due to haPL-II localization using
a homologous system (antibody to the native hormone) and relaxin
localization using a heterologous system (antibody to porcine
relaxin). Relaxin is detectable in GTC on Days 8 and 10 of gesta-
tion in the hamster (Renegar et al. , 1987), whereas haPL-II was not
detected in the present study until Day 12. Temporal differences
in the appearance of haPL-II and relaxin in the Git suggests that
different factors regulate synthesis and secretion of these hormones
on that hormone expression evolves as Git differentiation proceeds.
Prolactin and relaxin C-peptide have been immunolocalized in
cells of the panietal decidua adherent to fetal membranes and in
decidua-like cells of the placental basal plate of the human (Sak-
bun et al., 1987). Although, the significance ofthis co-localization
in the placenta is unknown, Sortino et al. (1989) recently reported
that human relaxin stimulated prolactin secretion from dispersed
pituitary cells. Therefore, a direct paracnine effect ofhuman placental
relaxin on decidual prolactin secretion is possible. Additional study
will be required to determine ifsuch a relationship exists between
hamster relaxin and haPL-II.
Literature Cited
Boime I, Boothby M, Hoshima M, Daniels-McQueen 5, Darnell R (1982):
Expression and structure ofhuman placental hormone genes as a function
of placental development. Biol Reprod 26:73
Carnegie JA, McCully ME, Robertson HA (1978): Localization of ovine
placental lactogen by immunofluorescence using sections of tissue embed-
ded in glycolmethacrylate. J Histochem Cytochem 26:223
de lkonicoffLK, Cedard L (1973): Localization ofhuman chorionic gonad-
otropic and somatomamotropic hormones by the peroxidase immunohisto
enzymes method in villi and amniotic epithelium ofhuman placentas (from
six weeks to term). Am J Obstet Gynecol 116:1124
Duckworth ML, Kirk KL, Fniesen HG (1986): Isolation and identification
of a cDNA clone of rat placental lactogen II. J Biol Chem 21:10871
Hall J, Talamantes F (1984): Immunocytochemical localization of mouse
placental lactogen in the mouse placenta. J Histochem Cytochem 32:379
Hoshina M, Boothby M, Hussa R, Pattillo R, Camel HM, Boime I (1985):
Linkage of human chonionic gonadotrophin and placental lactogen bio-
synthesis to trophoblast differentiation and tumorigenesis. Placenta 6:163
Hoshina M, Hussa R, Pattillo R, Camel HM, Boime I (1982)-� The role of
trophoblast differentiation in the control of the hCG and hPL genes. AdvExp Med Biol 176:299
Jackson U, Colosi P, Talamantes F, Linzer DIH (1986): Molecular cloning
of mouse placental lactogen cDNA, Proc NatI Acad Sci USA 83:8496
Kelly PA, Tsushima T, Shiu RPC, Fniesen HG(1976): Lactogenic and growth
hormone-like activities in pregnancy determined by radioreceptor assays.Endocrinology 99:765
MartalJ, DijianeJ, Dubois MP (1977): Immunofluorescence localizationof ovine placental lactogen. Cell Tissue Res 184: 427
McWilliams D, Boime I (1980): Cytological localization ofplacental lacto-
gen messenger nibonucleic acid in syncytiotrophoblast layers of humanplacenta. Endocrinology 107:761
Orsini MW (1954): The trophoblastic giant cells and endovascular cells as-
sociated with pregnancy in the hamster, Cricetus auratus. AmJ Anat 94:275
Reddy 5, Watkins WB (1978): Immunofluorescent localisation of ovine
placenta lactogen. J Reprod Fertil 52:173
Renegar RH, Cobb AD, Leavitt WW (1987): Immunocytochemical local-
ization of nelaxin in the golden hamster (Mesocricetus auratus) during the
last half of gestation. Biol Reprod 37:925
Robertson MC, Gillespie B, Fniesen HG (1982): Characterization ofthe two
forms ofrat placental lactogen (rPL): rPL.I and rPL-II. Endocrinology 111:1862
Sakbun V. Koay ESC, Bryant-Greenwood GD (1987): Immunocytochemi.
cal localization of prolactin and relaxin C.peptide in human decidua andplacenta. J Clin Endocninol Metab 65:339
Sortino MA, Cronin MJ, Wise PM (1989): Relaxin stimulates prolactin scene-
tion from anterior pituitary cells. Endocrinology 124:2013
Soares MJ, Colosi P. Ogren L, Llamantes F(1983): Identification and par-
tial characterization ofa lactogen from the midpregnant mouse conceptus.
Endocrinology 112:1313
Soares MJ, i�lamantes F(1982): Placental and serum hormone changes during
the second half of pregnancy in the hamster. Biol Reprod 7:523
SouthardJN, Campbell GT, ‘Illamantes F(1987): Hamster placental lacto-gen: gestational profiles and high molecular weight forms. Endocrinology121:900
by guest on March 16, 2016jhc.sagepub.comDownloaded from

940 RENEGAR, SOUTHARD, TALAMANTES
SouthardJN, l#{224}lamantesF(1987): Immunological studies ofrodent placental Watkins WB (1978): Use ofimmunocytochemical techniques for the local-
lactogens. Mol Cell Endocninol 50:29 ization of human placental lactogen. J Histochem Cytochem 26:288
SouthardJN, Thoudarson G, Talamantes F (1986): Purification and partial Watkins WB, Reddy S (1980): Ovine placental lactogen in the cotyledonary
characterization of hamster placental lactogen. Endocrinology 119:508 and intercotyledonary placenta of the ewe. J Reprod Fentil 58:411
Steinetz BG, O’Byrne EM, Goldsmith LT, Anderson MB (1988): The source Wooding FBP(1981): Localization ofovine placental lactogen in sheep placen-
of relaxin in pregnant Syrian hamsters. Endocrinology 122:795 tomes by electron microscope immunocytochemistry.J Reprod Fertil 62:15
by guest on March 16, 2016jhc.sagepub.comDownloaded from