the hamster (mesocricetus auratus) as experimental model in chagas' diseases: parasitological...
TRANSCRIPT

ORIGINAL PAPER
The hamster (Mesocricetus auratus) as an experimental modelof toxocariasis: histopathological, immunohistochemical,and immunoelectron microscopic findings
Ana Maria Gonçalves da Silva & Pedro Paulo Chieffi & Wellington Luiz Ferreira da Silva &
Edite Hatsumi Yamashiro Kanashiro & Guita Rubinsky-Elefant & Edécio Cunha-Neto &
Eliane Conti Mairena & Thales De Brito
Received: 10 June 2014 /Accepted: 26 August 2014 /Published online: 19 December 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Toxocariasis is a globally distributed parasitic infec-tion caused by the larval stage of Toxocara spp. The typicalnatural hosts of the parasite are dogs and cats, but humans canbe infected by the larval stage of the parasite after ingestingembryonated eggs in soil or from contaminated hands orfomites. The migrating larvae are not adapted to complete theirlife cycle within accidental or paratenic hosts like humans andlaboratory animals, respectively, but they are capable of invad-ing viscera or other tissues where they may survive and inducedisease. In order to characterize hamsters (Mesocricetusauratus) as a model for Toxocara canis infection, histopatho-logical and immunohistochemistry procedures were used todetect pathological lesions and the distribution of toxocaralantigens in the liver, lungs, and kidneys of experimentallyinfected animals. We also attempted to characterize the immu-nological parameters of the inflammatory response and corre-late them with the histopathological findings. In the kidney, a
correlation between glomerular changes and antigen depositswas evaluated using immunoelectronmicroscopy. The hamsteris an adequate model of experimental toxocariasis for short-term investigations and has a good immunological and patho-logical response to the infection. Lung and liver manifestationsof toxocariasis in hamsters approximated those in humans andother experimental animal models. A mixed Th2 immunolog-ical response to T. canis infection was predominant. The ham-ster model displayed a progressive rise of anti-toxocaral anti-bodies with the formation of immune complexes. Circulatingantigens, immunoglobulin, and complement deposits weredetected in the kidney without the development of a definiteimmune complex nephropathy.
Keywords Toxocariasis . Hamster . Nephropathy
Introduction
Toxocariasis is a globally prevalent parasitic infection causedby the larval stage of Toxocara spp. The predominant naturalhosts of Toxocara canis and Toxocara cati (both members ofthe family Ascaridoidea) are canides and felines, respectively.T. canis is a common intestinal nematode parasite of dogs(Akao and Ohta 2007; Despommier 2003). The parasite isvery common in puppies, and a strong immunity begins inmost dogs by 2–3 months of age.
Consequently, among well-kept dogs that were dewormedas young puppies, infection is uncommon. Disease typicallyoccurs under conditions of suppressed immunity due tomalnourishment or some other concurrent disease or by beingexposed to reinfection with a low infective dose, as recentlyreported by Fahrion et al. (2008). Following infection, larvaeundergo extensive migration through the liver and lungs
A. M. G. da Silva (*) : P. P. Chieffi : E. H. Y. Kanashiro :G. Rubinsky-Elefant : T. De BritoInstitute of Tropical Medicine, University of São Paulo,São Paulo, SP, Brazile-mail: [email protected]
A. M. G. da Silva :W. L. F. da Silva : T. De BritoDepartment of Pathology, Medical School, University of São Paulo,São Paulo, SP, Brazil
P. P. ChieffiSanta Casa de São Paulo School of Medical Sciences,São Paulo, Brazil
E. Cunha-Neto : E. C. MairenaLaboratory of Immunology, Heart Institute (InCor), Medical School,São Paulo University, São Paulo, Brazil
A. M. G. da Silva : P. P. Chieffi : T. De BritoLaboratory of Medical Investigation (LIM 06), São Paulo, Brazil
Parasitol Res (2015) 114:809–821DOI 10.1007/s00436-014-4246-5

before migrating back to the intestine where they completetheir development and begin to shed eggs into the host’s feces(Despommier 2003).
Humans can be infected by T. canis, in its larval stage, byingesting embryonated eggs in soil, from contaminated handsor fomites, or consuming raw viscera and muscles of infectedparatenic hosts. The migrating larvae are not adapted to com-plete their life cycle within accidental or paratenic hosts likehumans and common laboratory animals, but they are capableof invading the viscera where they may survive for manymonths or years (Glickman and Schantz 1981).
Clinical manifestations of toxocariasis vary from asymp-tomatic to severe organ injury. The most common clinicalforms are visceral and ocular toxocariasis (Despommier2003). Other minor forms of the disease include chronicweakness, digestive pain, headache, cutaneous allergic signs(itchy rash, pruritus, and/or cough), and diffuse myalgia.These syndromes have been described as “coverttoxocariasis” (in Irish children) or “common toxocariasis”(in French adults) (Despommier 2003; Fillaux and Magnaval2013; Rubinsky-Elefant et al. 2010). Visceral larva migrans(VLM), initially defined by Beaver et al. (1952), is a syn-drome caused by Toxocara larvae that is characterized byextreme chronic eosinophilia accompanied by eosinophil-rich granulomatous lesions in an enlarged liver together withpulmonary infiltration. Patients with VLM may display fever,hepatosplenomegaly, hypergammaglobulinemia, pruriticrashes, and bronchospasm, but most have mild or no symp-toms. For this reason, Toxocara infection is suspected onlyafter incidental findings, such as sustained eosinophilia.
Ocular larva migrans (OLM) first was described by Wilderin 1950 (Wilder 1950) after the identification of nematodelarvae in eosinophilic granulomas from the enucleated eyes ofchildren with retinoblastoma. OLM is a self-limiting diseaseusually caused by the migration of Toxocara spp. larvae to theeye.
The degree of tissue damage in Toxocara spp. infectionsand the concomitant signs and symptoms vary with regard tothe severity of infection, the organs invaded, and the immu-nological response of the host. However, the liver, lungs, andcentral nervous system, including the eyes, appear to be mostsensitive. Renal manifestations, such as nephrotic syndromein humans and glomerular pathology, have been described asoutcomes of T. canis infection; this is similar to other parasiticinfections in humans and experimental animals (Casarosaet al. 1992; Shetty and Aviles 1999). Mice usually are themodel animals of choice for studying experimentaltoxocariasis.
The aim of the current study was to characterize hamsters(Mesocricetus auratus) as a model for T. canis infection usingstandard histopathological procedures and immunohisto-chemistry to detect the distribution of toxocaral antigens inthe lung, liver, and kidney. An attempt to characterize the
immunological parameters of the inflammatory response ofthe experimental host also was made. Finally, we evaluatedthe correlation between glomerular manifestations and localantigen deposits using immunoelectron microscopy.
Methods
Histopathology T. canis eggs were obtained from adult fe-male worms, and embryonated eggs were prepared as de-scribed previously (Glickman and Summers 1983). Hamsterswere housed in the animal facility of the Institute of TropicalMedicine, S. Paulo University (IMT), and were maintained oncommercial pellet food and water ad libitum. Six groups offive male hamsters each (mean weight, 180 g) were orallyexposed to 750 embryonated eggs by gavage (Ollero et al.2008).Hamsterswere sacrificed at 4, 6, 10, 15, 30, and 45 dayspostinfection, and liver, lung, and kidney fragments wereroutinely fixed in 10 % buffered formalin (pH 7.2), processed,embedded in paraffin, sectioned at 3–4-μm thickness, andstained with hematoxylin and eosin. For each group, fivenormal, uninfected hamsters were sacrificed and used as con-trols. Another experiment was performed with sacrifice of theanimals 5, 15, and 30 days after infection. All experimentswere performed in accordance with the recommendations forthe care and use of laboratory animals of the Institute ofTropical Medicine. The protocol was approved by the Com-mittee on the Ethics of Animal Experiments (Permit #009/08).
Immunohistochemistry assay Two antisera, one polyclonaland one monoclonal, were used. Both antibodies were pro-duced in the Laboratory of Seroepidemiology andImmunobiology of the IMT. Polyclonal antibody to larvalT. canis excretory/secretory (TES) antigens was raised inrabbits using a standard procedure (Bowman et al. 1987a). Amonoclonal antibody to larval TES antigens (designatedB6C3) first was obtained by immunizing BALB/c mice usinga previously described protocol (Bowman et al. 1987b). Sub-sequently, monoclonal antibodies were raised using a methoddescribed previously (de St Groth and Scheidegger 1980). Anenzyme-linked immunosorbent assay (ELISA) with TES an-tigens and an indirect fluorescent antibody assay were used forthe detection of specific immunoglobulin (Ig)-producing hy-brids and larval surfaces. Both antibodies were purified usingProtein A Sepharose CL-4B (Thermo Scientific, USA). Lung,liver, and kidney sections (3–4 μm thick) were analyzed usingEnVision (Dako, USA) immunohistochemistry methods. Forantigen retrieval, sections were microwaved in 10 mM citratebuffer (pH 6.0). The polyclonal antibody was used at a 1:150–1:200 dilution, and the monoclonal antibody at a 1:100 dilu-tion and an appropriate secondary antibody, a Novolink poly-mer—horseradish peroxidase (HRP) (Novocastra, UK). Stain-ing was completed using the chromogen solution, 3,3′-
810 Parasitol Res (2015) 114:809–821

diaminobenzidine (DAB), or alkaline phosphatase as sub-strates. All specimens then were lightly counterstained withHarris hematoxylin.
Immunofluorescence (IF) IF reactions were performed to de-tect Igs and complement C3 deposits in hamster kidneys.Snap-frozen tissue was cut into 4-μm-thick sections. Fromthe 15 hamsters infected, kidneys from 5 hamsters at 5, 15,and 30 days postinfection and 8 control kidneys from normal(noninfected) hamsters were used. The sections were fixed inacetone, washed with phosphate-buffered saline (PBS, pH7.2), and incubated in a moist chamber with antisera. IndirectIF was performed to detect Ig deposits in hamster kidneys. Acommercial monoclonal anti-hamster Ig (IgM and IgG cloneHG-31; Sigma Chemical, USA) was used at an optimal dilu-tion of 1:2000. The secondary antibody (Sigma Chemical,USA) was a goat anti-mouse IgG fluorescein isothiocyanate(FITC)-labeled antibody at an optimal dilution of 1:200. Nor-mal hamster kidney cryostat sections were used as negativecontrols. Liver tissue sections from infected hamsters wereused as positive controls because Igs could be detected in theinflamed connective tissue of the portal spaces. C3 deposits inglomeruli were detected using direct IF. An anti-mouse C3,kindly supplied by Dr. M. Laurenti, also detected hamster C3and was used at an optimal dilution of 1:75. The positivecontrol for C3 deposits consisted of fragments of mouse andhamster hind footpad skin that had been locally inoculatedwith Leishmania amazonensis promastigotes (Laurenti et al.1996, 2004). Fragments of normal hamster kidneys and ofnormal hind footpad skin from hamsters and mice were usedas negative controls.
Immunoelectron microscopy Immunoelectron microscopywas performed essentially as described elsewhere (De Britoet al. 1998, 1999) using the monoclonal antibody, B6C3. Thekidneys of 17 hamsters were used, corresponding to 4 days (2kidneys), 6 days (2 kidneys), 10 days (3 kidneys), 15 days (5kidneys), and 30 days (5 kidneys) after infection. To controlfor potential nonspecific binding of the secondary antibody,the reaction was performed with the primary antibody re-placed with normal hamster serum. The experimental modelwas not evaluated beyond 30 days postinfection because renalamyloidosis is prone to develop 30–40 days postinfection.
ELISA for the detection of IgG anti-T. canis Serum samplesfrom all 15 hamsters sacrificed before infection or 5, 15, or30 days after oral infection with 750 embryonated T. caniseggs were used. The control group consisted of 15 normalhamsters. TES antigens were obtained as described previously(De Savigny 1975), with some modifications (Elefant et al.2006). Ascaris suum adult worm extract (AWE) was producedas described previously (Kanamura et al. 1981), with somemodifications (Elefant et al. 2006). Anti-Toxocara IgG
antibodies were detected by ELISA based on the methoddescribed by De Savigny et al. (1979), with some modifica-tions (Elefant et al. 2006). Polystyrene plates were sensitizedwith TES antigen solution (1.9 μg/mL in 0.1 M carbonate-bicarbonate buffer [pH 9.6], 100 μL/well) at 37 °C for 1 h andthen at 4 °C for 18 h. A blocking solution of 5 % skim milk(Molico, Nestlé) in 0.01 M PBS (pH 7.2) containing 0.05 %Tween 20 (PBS-T) (200 μL/well) then was added for 2 h at37 °C. All sera were pre-absorbed with AWE (25 μg/mL) in asolution of 1 % skim milk in PBS-T to remove nonspecificantibodies. Serum samples were processed by twofold serialdilutions in PBS-T from 1:50 to 1:6400 (100 μL/well) andwere incubated at 37 °C for 30 min. An anti-hamster (IgG andIgM) monoclonal antibody (Sigma Chemical, USA), diluted1:10,000 in PBS-T (100 μL/well), was added, and sampleswere incubated at 37 °C for 40 min. Samples then wereincubated with a HRP-conjugated anti-mouse Ig (SigmaChemical, USA), diluted 1:1000 in PBS-T (100 μL/well), at37 °C for 40 min. Washes with PBS-T were performed intriplicate after each incubation. The chromogen substrate,ortho-phenylenediamine (OPD; 100 mL/well, 0.4 mg/mL,Sigma Chemical, USA), in 0.05 M citrate buffer (pH 5.0)and 30 % hydrogen peroxide (0.4 μL/mL) then was added.The reaction proceeded for 20 min at 37 °C and was termi-nated with 50 μL of 2 N H2SO4. The assay was monitoredwith negative and positive serum samples and a blank (with-out serum). The absorbance was determined at 492 nm usingan automatic 7 microplate reader (Titertek Multiskan MCC/340, Labsystems, Finland). Cutoff values were calculatedbased on the mean absorbance of negative control serumsamples±2 standard deviations (0.106).
Detection of circulating TES antigens by sandwichELISA Polystyrene plates were sensitized with rabbit anti-TES polyclonal antibody (15 μg/mL in 0.1 M carbonate-bicarbonate buffer, pH 9.6, 50 μL/well; Laboratory ofSeroepidemiology and Immunobiology of the IMT) at 37 °Cfor 2 h. Subsequently, samples were incubated at 4 °C for 18 h.A blocking solution of 5 % fetal bovine serum (100 μL;Cultilab, Brazil) in PBS-Twas added at 37 °C for 1 h. Serumsamples then were treated with ethylenediamine tetraaceticacid (EDTA) to dissociate antigen complexes (Bowmanet al. 1987). Briefly, sera were mixed with an equal volumeof 0.1 M EDTA (pH 7.5) and heated at 100 °C for 10 min.Samples then were centrifuged at 10,800×g at 4 °C for10 min. The resulting supernatants were used for ELISA(50 μL/well), and samples were incubated for 2 h at 37 °Cand subsequently for 18 h at 4 °C. Different dilutions of TESalso were incubated to generate a standard curve. After threewashing cycles, plates were incubated with 50 μL/well ofmouse anti-TES monoclonal antibody (130 μg/mL; Labora-tory of Seroepidemiology and Immunobiology of IMT-SP) at37 °C for 1 h. The mouse anti-TES monoclonal antibody had
Parasitol Res (2015) 114:809–821 811

been pre-absorbed with 10% normal rabbit serum in PBS-Tat4 °C for 24 h. After three washing cycles with PBS-T, HRP-conjugated anti-mouse Ig (Sigma Chemical, USA) diluted1:1000 in PBS-T plus 10 % normal rabbit serum (50 μL/well)was added, and samples were incubated at 37 °C for 40 min.All washes were performed in triplicate with PBS-T. As achromogen substrate (50 μL/well), OPD in 0.05 M (pH 5.0)citrate buffer (0.5 mg/mL) and 30 % H2O2 (0.5 μL/mL) wereadded, and samples were incubated for 30 min at 37 °C. Thereaction was stopped with 25 μL of 2 N H2SO4, and theabsorbance was determined at 492 nm.
Between-group comparisons of specific anti-TES antibod-ies and circulating antigens were made using the Mann-Whitney test (for two groups) and the Kruskal-Wallis test(for three or more groups) with Sigma Stat 3.5 software(USA). The level of significance was set accepting a type 1error of 5 % (α=0.05 %).
RNA isolation, reverse transcription, and real-time reversetranscriptase polymerase chain reaction (RT-PCR) RNApreparation and processing techniques were essentially asdescribed by Chomczynsky and Sacchi (1987). In brief, totalRNA was isolated from liver samples using TRIzol reagent(Invitrogen, USA). Contaminating DNA was removed bytreatment with ribonuclease-free deoxyribonuclease I. Real-time RT-PCR was performed as described by Bilate et al.(2008). Briefly, cDNA was reverse-transcribed from 5 μg oftotal RNA using SuperScript I RT (Invitrogen, USA). Subse-quently, messenger RNA (mRNA) expression was analyzedby real-time quantitative PCR using SYBR Green I PCRMaster Mix (Applied Biosystems, USA) and 250 nmol/L
Real-Time PCR Systems. The following primers were de-signed using Primer Express v.3.0 software (AppliedBiosystems, USA): glyceraldehyde phosphate dehydrogenase(GAPDH) (DQ403055), 5′-CTGACATGCCGCCTGGAG-3′and 5 ′ -TCAGTGTAGCCCAGGATGCC-3 ′ ; IFNγ(AF034482), 5′-GAAGCTCACCAAGATTCCGGTAA-3′and 5 ′-TTTTCGTGACAGGTGAGGCAT-3 ′ ; IL10(AF046210), 5′-GGCAACTGCAGCGCTGTC-3′ and 5′-AGACGCCTTTCTCTTGGAGCTTAT-3 ′ ; and IL4(AF046213.1), 5′-CTCCACGGAGAAAGACCTCATCT-3′and 5′-TGGAATTGTTCTTCAAGCACAGG-3′. After everyPCR cycle, an amplicon melting point curve was obtained.This process yielded a single peak when the annealing tem-perature recommended by Primer Express was used,confirming the specificity of the reaction. GAPDH mRNAexpression was measured as a normalization control. Thechoice of endogenous genes is of paramount importance inquantitative PCR (qPCR), and the best candidate is that whichdisplays little variation among samples. One alternative that isused when no single gene is stable enough is the use of severalendogenous housekeeping genes.
In the case of hamster tissues, one shortcoming is that since itsgenome has not been sequenced yet, we only have the se-quence of GAPDH as endogenous housekeeping gene. Wehave followed the procedure to certify whether the GAPDHendogenous gene is adequate for the qPCR reaction in thehamster tissue in the current manuscript. We found that theaverage GAPDH level in all samples (control + experimentalsamples in all time points) was 17.4 with a standard deviationof 1.1, a satisfactory number, showing that GAPDH expres-sion does not change significantly upon disease induction. Wehave previously validated the GAPDH gene as a single reli-able endogenous housekeeping gene marker in the hamster(Bilate et al. 2007, 2008)
The relative amount of mRNA was calculated using the2−ΔΔCt method.
Statistical analysis was performed by unpaired Student’s ttest. Results were expressed as mean±standard error of themean.
Results
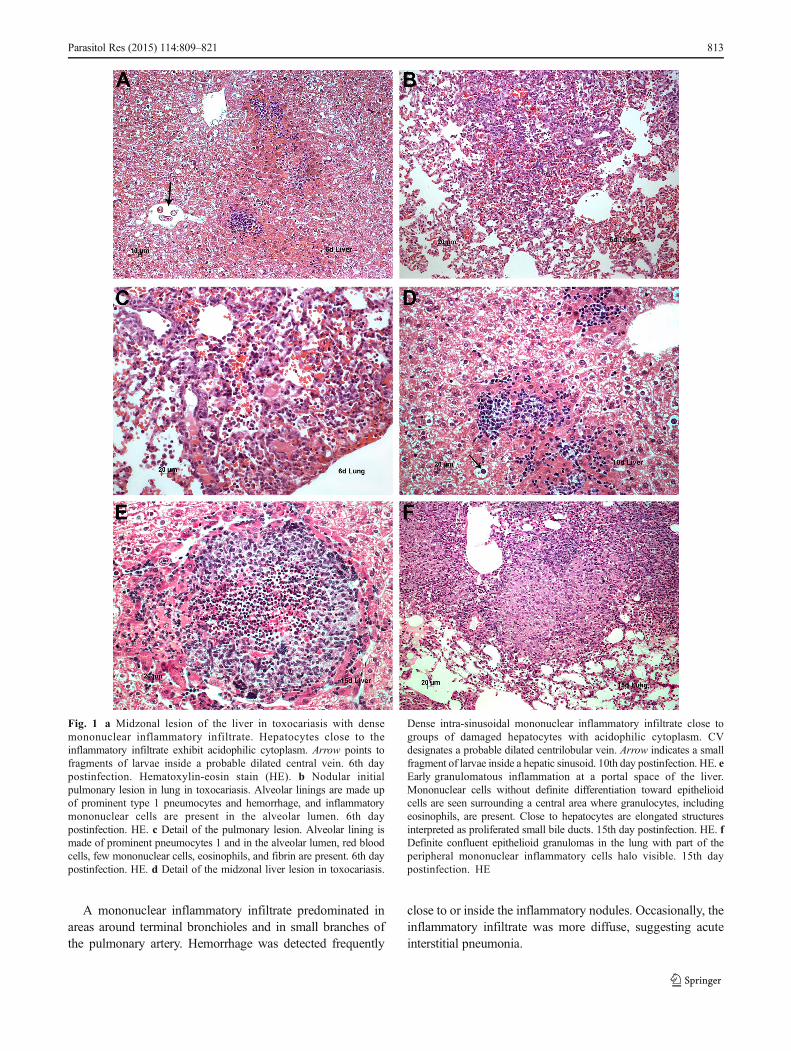
Histopathology and immunohistochemistry Larvae at 4–10 days postinfection were detected in the liver(Fig. 3b) and were reactive to polyclonal and to mono-clonal sera. The portal spaces occasionally displayedslight focal inflammatory infiltrates corresponding tomacrophages and large lymphocytes intermingled with afew neutrophils. The polyclonal antibody reacted withTES antigens in the cytoplasm of enlarged Kupffer cellsin the sinusoidal lining. Macrophages of the portalspaces also harbored TES antigens, as demonstrated bytheir reaction with polyclonal and monoclonal antibodies.The main histopathological finding was groups of mono-nuclear cells and a few granulocytes scattered along theliver sinusoids at the midzonal region of the liver lobulesurrounding either isolated or groups of hepatocytes;these cells exhibited mild regressive changes particularlyrepresented by acidophilic cytoplasm (Fig. 1a, d).
In the lungs, TES antigens were present in a few intra-alveolar macrophages and infrequently in cells of thebronchial lining (Fig. 3a). TES antigens also were de-tected in peribronchial spaces and lymphatic nodules. At4 and 6 days postinfection, we observed a focal reactiv-ity of cells of the alveolar lining and detected an inter-stitial and alveolar nodular infiltration of macrophages,lymphocytes, and a few granulocytes in the lung paren-chyma; this could be approximated as the pulmonarycounterpart of the midzonal lesion in the lobule of theliver (Fig. 1b, c). A few remnants of the bronchiolarepithelial lining may be present at the central part ofthe inflammatory pulmonary nodule.
812 Parasitol Res (2015) 114:809–821
sense and antisense primers on Applied Biosystems® 7500
A mononuclear inflammatory infiltrate predominated inareas around terminal bronchioles and in small branches ofthe pulmonary artery. Hemorrhage was detected frequently
close to or inside the inflammatory nodules. Occasionally, theinflammatory infiltrate was more diffuse, suggesting acuteinterstitial pneumonia.
Fig. 1 a Midzonal lesion of the liver in toxocariasis with densemononuclear inflammatory infiltrate. Hepatocytes close to theinflammatory infiltrate exhibit acidophilic cytoplasm. Arrow points tofragments of larvae inside a probable dilated central vein. 6th daypostinfection. Hematoxylin-eosin stain (HE). b Nodular initialpulmonary lesion in lung in toxocariasis. Alveolar linings are made upof prominent type 1 pneumocytes and hemorrhage, and inflammatorymononuclear cells are present in the alveolar lumen. 6th daypostinfection. HE. c Detail of the pulmonary lesion. Alveolar lining ismade of prominent pneumocytes 1 and in the alveolar lumen, red bloodcells, few mononuclear cells, eosinophils, and fibrin are present. 6th daypostinfection. HE. d Detail of the midzonal liver lesion in toxocariasis.
Dense intra-sinusoidal mononuclear inflammatory infiltrate close togroups of damaged hepatocytes with acidophilic cytoplasm. CVdesignates a probable dilated centrilobular vein. Arrow indicates a smallfragment of larvae inside a hepatic sinusoid. 10th day postinfection. HE. eEarly granulomatous inflammation at a portal space of the liver.Mononuclear cells without definite differentiation toward epithelioidcells are seen surrounding a central area where granulocytes, includingeosinophils, are present. Close to hepatocytes are elongated structuresinterpreted as proliferated small bile ducts. 15th day postinfection. HE. fDefinite confluent epithelioid granulomas in the lung with part of theperipheral mononuclear inflammatory cells halo visible. 15th daypostinfection. HE
Parasitol Res (2015) 114:809–821 813
The kidneys were unremarkable at 4 days postinfectionexcept for one larva detected in the renal interstitium in oneanimal.
At 10 days postinfection, portal inflammation was morefrequent, with occasional small areas of piecemeal necrosis(interface hepatitis) and focal progression of the inflammatory
infiltrate into the liver lobule. However, the midzonal inflam-matory areas remained the predominant finding with macro-phages, lymphocytes, neutrophils, and eosinophils present.TES antigens, detected by both sera, were present in macro-phages, including Kupffer cells. Again, the detection of TESantigens by the monoclonal antibody was more predominant
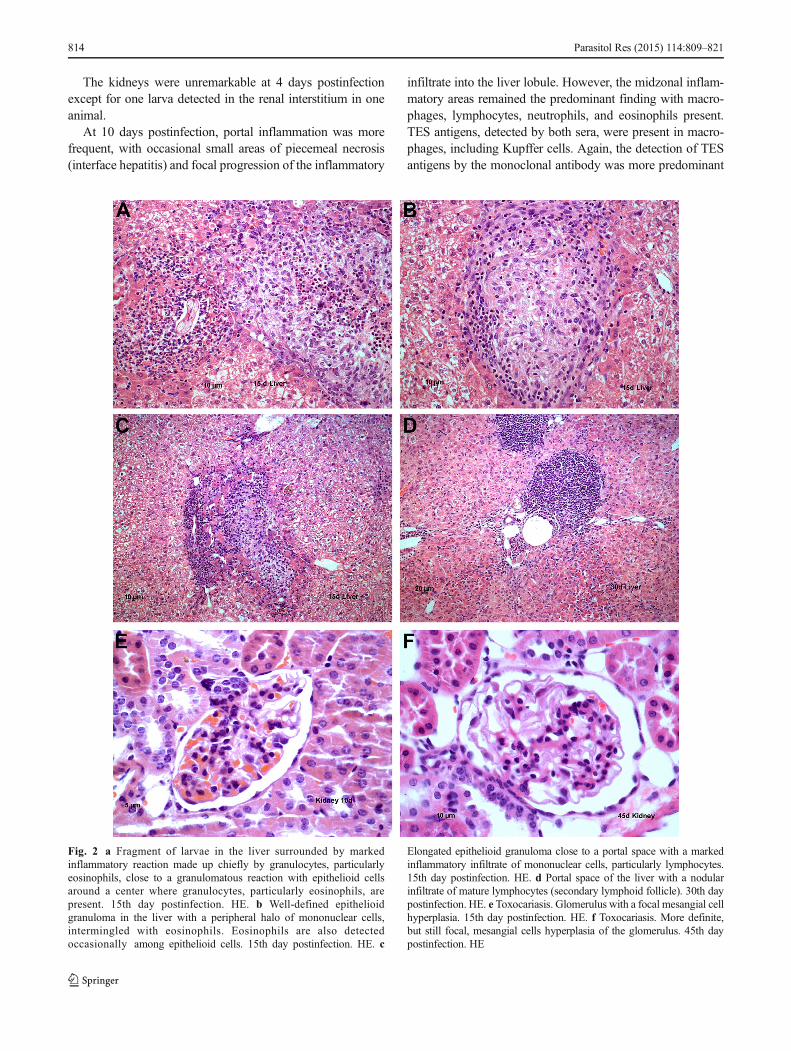
Fig. 2 a Fragment of larvae in the liver surrounded by markedinflammatory reaction made up chiefly by granulocytes, particularlyeosinophils, close to a granulomatous reaction with epithelioid cellsaround a center where granulocytes, particularly eosinophils, arepresent. 15th day postinfection. HE. b Well-defined epithelioidgranuloma in the liver with a peripheral halo of mononuclear cells,intermingled with eosinophils. Eosinophils are also detectedoccasionally among epithelioid cells. 15th day postinfection. HE. c
Elongated epithelioid granuloma close to a portal space with a markedinflammatory infiltrate of mononuclear cells, particularly lymphocytes.15th day postinfection. HE. d Portal space of the liver with a nodularinfiltrate of mature lymphocytes (secondary lymphoid follicle). 30th daypostinfection. HE. e Toxocariasis. Glomerulus with a focal mesangial cellhyperplasia. 15th day postinfection. HE. f Toxocariasis. More definite,but still focal, mesangial cells hyperplasia of the glomerulus. 45th daypostinfection. HE
814 Parasitol Res (2015) 114:809–821
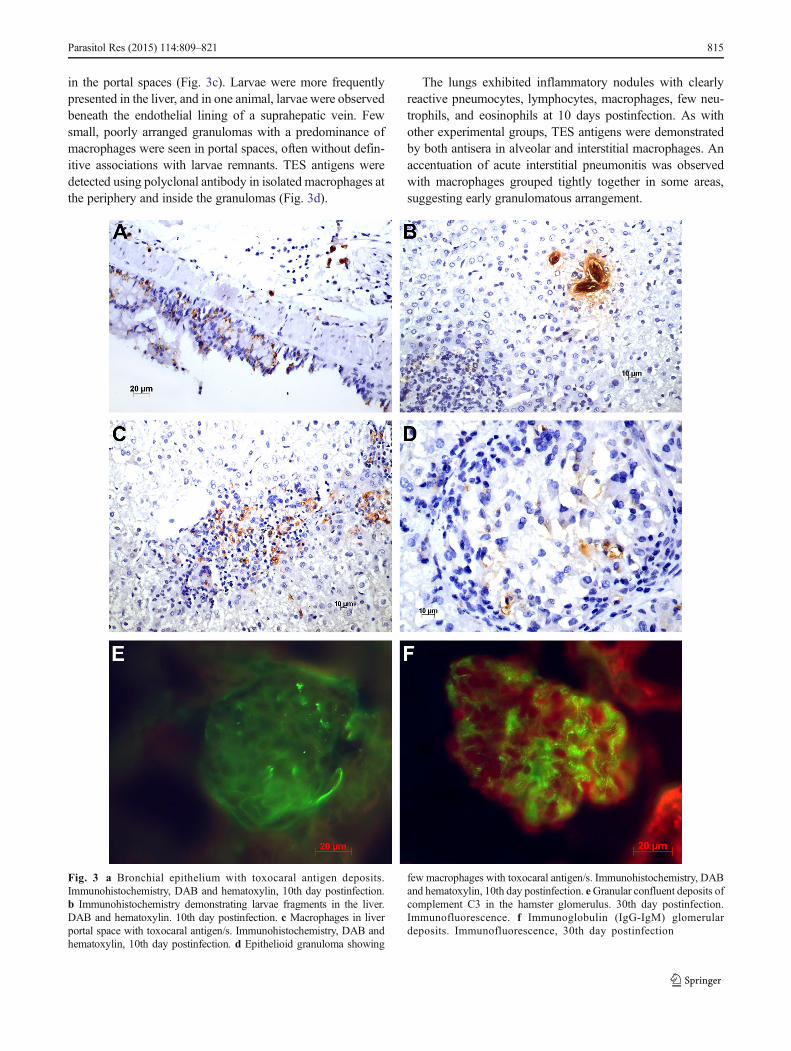
in the portal spaces (Fig. 3c). Larvae were more frequentlypresented in the liver, and in one animal, larvae were observedbeneath the endothelial lining of a suprahepatic vein. Fewsmall, poorly arranged granulomas with a predominance ofmacrophages were seen in portal spaces, often without defin-itive associations with larvae remnants. TES antigens weredetected using polyclonal antibody in isolated macrophages atthe periphery and inside the granulomas (Fig. 3d).
The lungs exhibited inflammatory nodules with clearlyreactive pneumocytes, lymphocytes, macrophages, few neu-trophils, and eosinophils at 10 days postinfection. As withother experimental groups, TES antigens were demonstratedby both antisera in alveolar and interstitial macrophages. Anaccentuation of acute interstitial pneumonitis was observedwith macrophages grouped tightly together in some areas,suggesting early granulomatous arrangement.
Fig. 3 a Bronchial epithelium with toxocaral antigen deposits.Immunohistochemistry, DAB and hematoxylin, 10th day postinfection.b Immunohistochemistry demonstrating larvae fragments in the liver.DAB and hematoxylin. 10th day postinfection. c Macrophages in liverportal space with toxocaral antigen/s. Immunohistochemistry, DAB andhematoxylin, 10th day postinfection. d Epithelioid granuloma showing
few macrophages with toxocaral antigen/s. Immunohistochemistry, DABand hematoxylin, 10th day postinfection. eGranular confluent deposits ofcomplement C3 in the hamster glomerulus. 30th day postinfection.Immunofluorescence. f Immunoglobulin (IgG-IgM) glomerulardeposits. Immunofluorescence, 30th day postinfection
Parasitol Res (2015) 114:809–821 815
816 Parasitol Res (2015) 114:809–821
The kidneys displayed groups of glomeruli with a modestmesangial expansion (Fig. 2e). Few interstitial small, focalTES antigen deposits were observed with polyclonal antibodystaining in the interstitium of the upper medullary zone.
Fifteen days postinfection, macrophages predominated inliver midzonal inflammatory nodules; these occasionallyprogressed to epithelioid granulomas (Fig. 2c) with or withoutsmall central areas of necrosis composed mostly of cellulardebris (Figs. 1e and 2a, b). The inflammatory nodules werecloser to the portal spaces but still occurred at the midzonalregion of the lobule. Larvae presented less frequently at15 days postinfection, but portal granulomatous inflammationwas increased with macrophages occasionally predominatingover epithelioid cells and with isolated giant cells observed.
Notably, we observed the ectopic formation of lymphoid-like aggregates in one animal. These aggregates contained afew macrophages and plasma cells at the portal spaces andresembled a pseudolymphoma infiltration (Fig. 2d).
Lung tissue exhibited areas of acute interstitial pneumonitisand the presence of TES antigens in macrophages, as notedprimarily by polyclonal antisera. Obvious epithelioid granu-lomas were observed in a few cases (Fig. 1f).
Larvae presented in the kidney interstitium at 15 dayspostinfection and glomerular mesangial changes were compa-rable to those observed at previous time points.
Histopathological findings were essentially similar at30 and 45 days postinfection but declining when com-pared with previous time points. Larvae were detectedless frequently, and their remnants could be observed insmall portal liver granulomas with few giant and epithe-lioid cells. However, macrophage activity persisted withengulfed TES antigens observable by both antisera. Por-tal formation of ectopic lymphoid aggregates was seen ina few cases and was intermingled with macrophagesharboring TES antigens.
Interstitial pneumoniawas slight or absent at 30 and 45 dayspostinfection. In one case, larval remnants were observed inthe pulmonary interstitium. Small interstitial inflammatorynodules presented rarely with deposits of ferric pigment inmacrophages.
Kidney tissue displayed groups of glomeruli with mildmesangial expansion, as noted for previous time points(Fig. 2f). In one case, larvae were identified in the interstitium.
IF IgG and IgM deposits were observed in 6 of 15 inoculatedanimals using indirect IF. Mesangial deposits usually werefocal in the glomerulus and affected few glomeruli in thekidney (Fig. 3f). Complement deposits, observed in five inoc-ulated animals, presented as small granules apparently locatedin the mesangial region (Fig. 3e). In four animals, Igs andcomplement were present, and in one animal, only comple-ment was detected. In four hamsters, Igs and complementwere absent from the glomerulus.
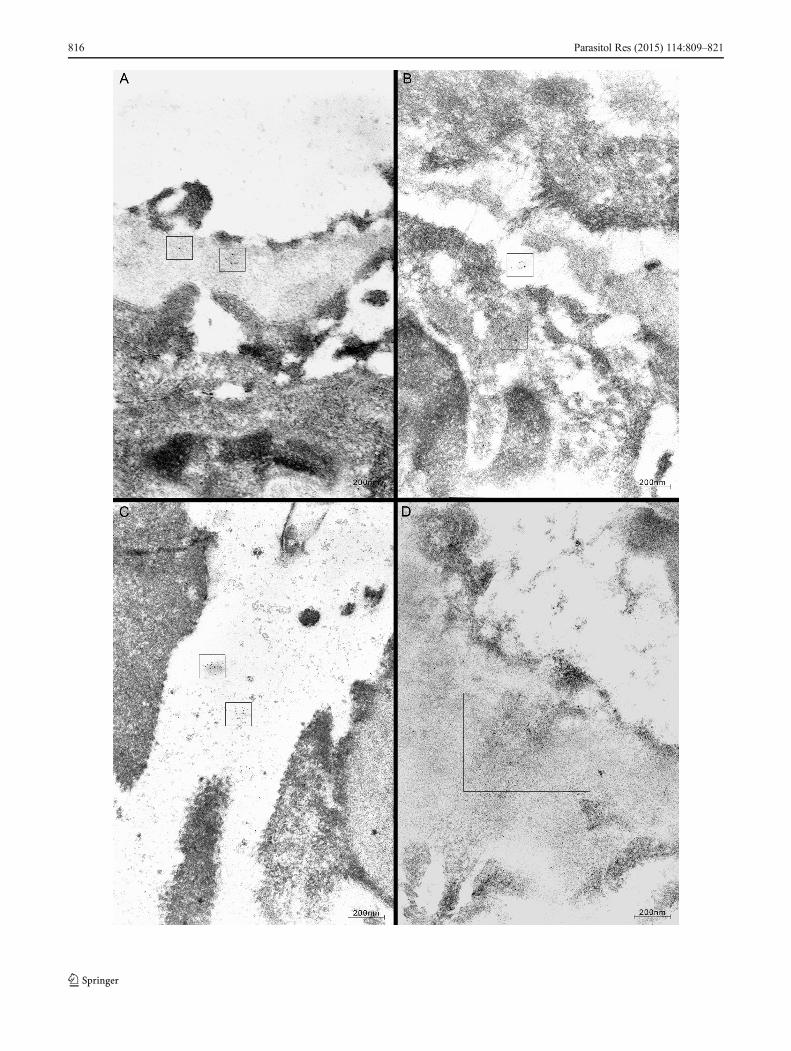
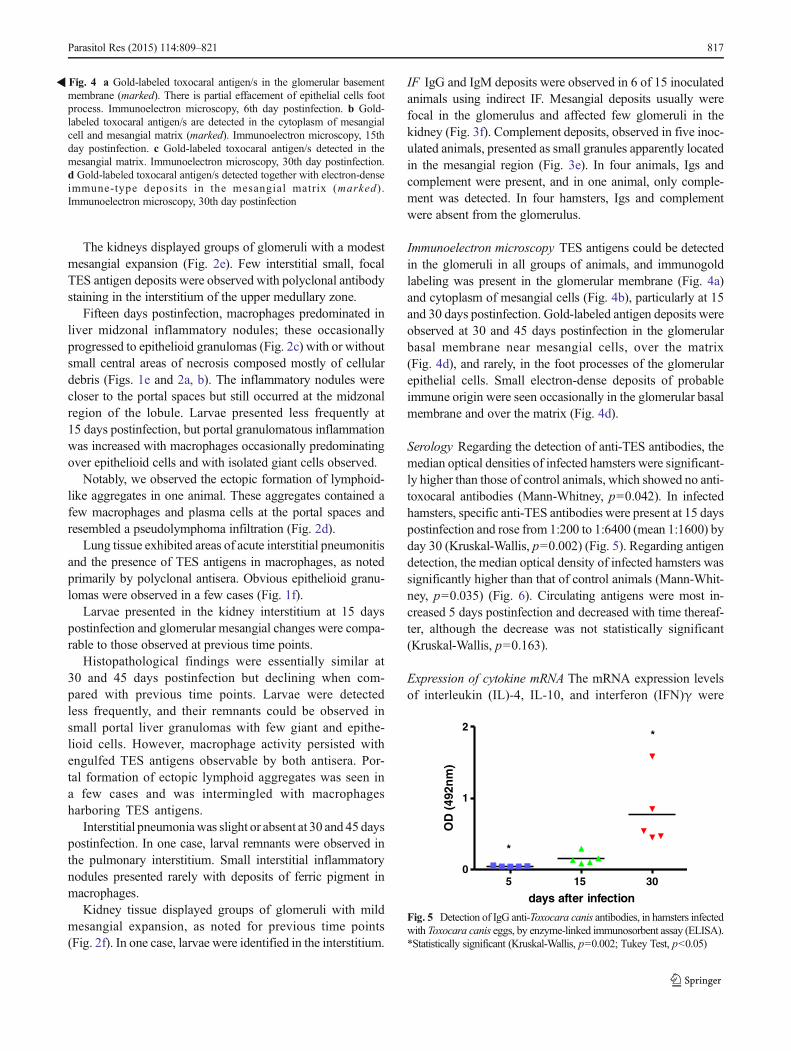
Immunoelectron microscopy TES antigens could be detectedin the glomeruli in all groups of animals, and immunogoldlabeling was present in the glomerular membrane (Fig. 4a)and cytoplasm of mesangial cells (Fig. 4b), particularly at 15and 30 days postinfection. Gold-labeled antigen deposits wereobserved at 30 and 45 days postinfection in the glomerularbasal membrane near mesangial cells, over the matrix(Fig. 4d), and rarely, in the foot processes of the glomerularepithelial cells. Small electron-dense deposits of probableimmune origin were seen occasionally in the glomerular basalmembrane and over the matrix (Fig. 4d).
Serology Regarding the detection of anti-TES antibodies, themedian optical densities of infected hamsters were significant-ly higher than those of control animals, which showed no anti-toxocaral antibodies (Mann-Whitney, p=0.042). In infectedhamsters, specific anti-TES antibodies were present at 15 dayspostinfection and rose from 1:200 to 1:6400 (mean 1:1600) byday 30 (Kruskal-Wallis, p=0.002) (Fig. 5). Regarding antigendetection, the median optical density of infected hamsters wassignificantly higher than that of control animals (Mann-Whit-ney, p=0.035) (Fig. 6). Circulating antigens were most in-creased 5 days postinfection and decreased with time thereaf-ter, although the decrease was not statistically significant(Kruskal-Wallis, p=0.163).
Expression of cytokine mRNA The mRNA expression levelsof interleukin (IL)-4, IL-10, and interferon (IFN)γ were
�Fig. 4 a Gold-labeled toxocaral antigen/s in the glomerular basementmembrane (marked). There is partial effacement of epithelial cells footprocess. Immunoelectron microscopy, 6th day postinfection. b Gold-labeled toxocaral antigen/s are detected in the cytoplasm of mesangialcell and mesangial matrix (marked). Immunoelectron microscopy, 15thday postinfection. c Gold-labeled toxocaral antigen/s detected in themesangial matrix. Immunoelectron microscopy, 30th day postinfection.d Gold-labeled toxocaral antigen/s detected together with electron-denseimmune-type deposits in the mesangial matrix (marked) .Immunoelectron microscopy, 30th day postinfection
5 15 300
1
2*
*
days after infection
OD
(49
2nm
)
Fig. 5 Detection of IgG anti-Toxocara canis antibodies, in hamsters infectedwith Toxocara canis eggs, by enzyme-linked immunosorbent assay (ELISA).*Statistically significant (Kruskal-Wallis, p=0.002; Tukey Test, p<0.05)
Parasitol Res (2015) 114:809–821 817
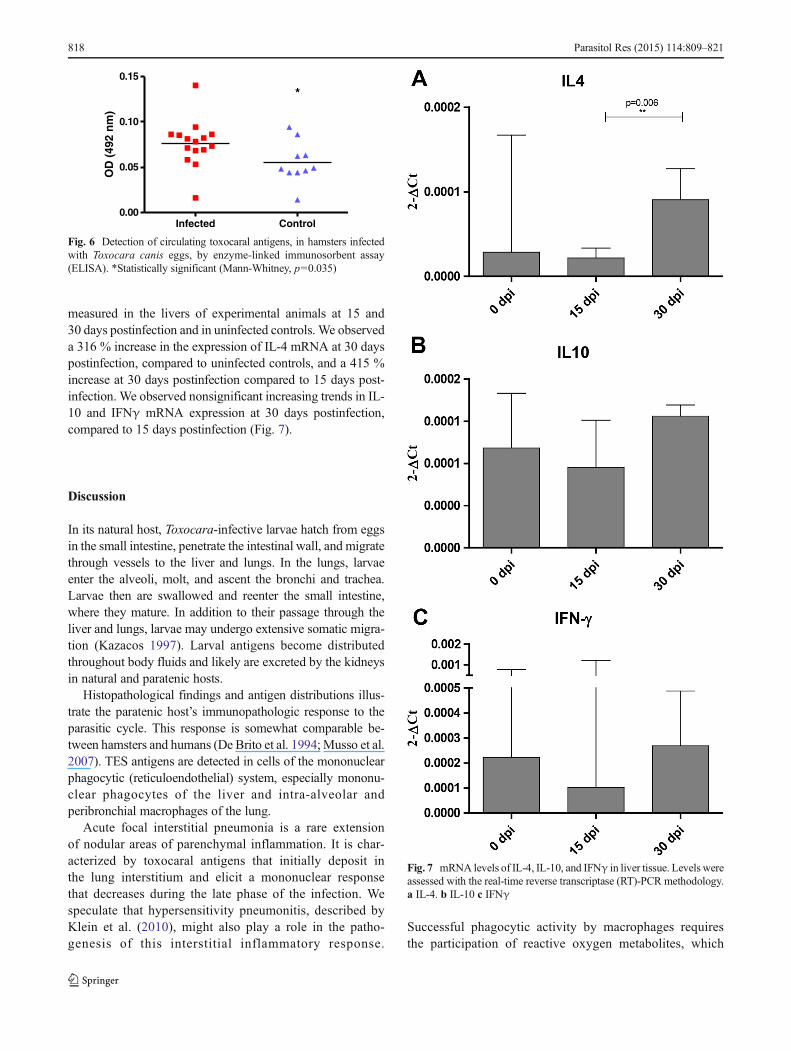
measured in the livers of experimental animals at 15 and30 days postinfection and in uninfected controls. We observeda 316 % increase in the expression of IL-4 mRNA at 30 dayspostinfection, compared to uninfected controls, and a 415 %increase at 30 days postinfection compared to 15 days post-infection. We observed nonsignificant increasing trends in IL-10 and IFNγ mRNA expression at 30 days postinfection,compared to 15 days postinfection (Fig. 7).
Discussion
In its natural host, Toxocara-infective larvae hatch from eggsin the small intestine, penetrate the intestinal wall, and migratethrough vessels to the liver and lungs. In the lungs, larvaeenter the alveoli, molt, and ascent the bronchi and trachea.Larvae then are swallowed and reenter the small intestine,where they mature. In addition to their passage through theliver and lungs, larvae may undergo extensive somatic migra-tion (Kazacos 1997). Larval antigens become distributedthroughout body fluids and likely are excreted by the kidneysin natural and paratenic hosts.
Histopathological findings and antigen distributions illus-trate the paratenic host’s immunopathologic response to theparasitic cycle. This response is somewhat comparable be-tween hamsters and humans (De Brito et al. 1994;Musso et al.2007). TES antigens are detected in cells of the mononuclearphagocytic (reticuloendothelial) system, especially mononu-clear phagocytes of the liver and intra-alveolar andperibronchial macrophages of the lung.
Acute focal interstitial pneumonia is a rare extensionof nodular areas of parenchymal inflammation. It is char-acterized by toxocaral antigens that initially deposit inthe lung interstitium and elicit a mononuclear responsethat decreases during the late phase of the infection. Wespeculate that hypersensitivity pneumonitis, described byKlein et al. (2010), might also play a role in the patho-genesis of this interstitial inflammatory response.
Successful phagocytic activity by macrophages requiresthe participation of reactive oxygen metabolites, which
Infected Control0.00
0.05
0.10
0.15
*O
D (
492
nm
)
Fig. 6 Detection of circulating toxocaral antigens, in hamsters infectedwith Toxocara canis eggs, by enzyme-linked immunosorbent assay(ELISA). *Statistically significant (Mann-Whitney, p=0.035)
Fig. 7 mRNA levels of IL-4, IL-10, and IFNγ in liver tissue. Levels wereassessed with the real-time reverse transcriptase (RT)-PCR methodology.a IL-4. b IL-10 c IFNγ
818 Parasitol Res (2015) 114:809–821

are generated during the oxidative metabolic burst understimulatory conditions (Fan et al. 2004).
The detection of TES antigens in the bronchiolar liningindicates the migration of larvae up the bronchus. We specu-late that this finding, if present in humans, might contribute tothe pathogenesis of bronchospasm.
During the initial phase of the infection, innate immunitypredominates in the host with a marked presence of mononu-clear phagocytes. As the infection progresses, the adaptiveimmune response is expressed chiefly as granulomatous in-flammation, but the mononuclear phagocytic response is main-tained to some extent. Granulomas appear to express both Th1-and Th2-mediated immunological responses during this phase.Th1 coincides with IgE production, and eosinophilia is associ-ated with the Th2-mediated response (Fan et al. 2013).
In humans and experimental helminth models, TES anti-gens are exemplary Th2-cell inducers that are associated withhigh-level tissue eosinophilia, mucosal mastocytosis, and IgEproduction. However, several interesting exceptions to thisrule exist, including initial Th1 stimulation (Maizels andYazdanbakhsh 2003).
The importance of IL-10 as a regulatory mediator has longbeen recognized. The participation of regulatory T cells in thehost response to helminths has been reported recently(Maizels 2013) and has been described specifically for T. canisinfection in mice (Othman et al. 2011).
A recent analysis of serum cytokines in children withpulmonary manifestations associated with T. canis infection(Nagy et al. 2012) indicated that TES products predominantlyactivate the Th2 immune response, as seen in our experimen-tal model, but TES antigens also significantly increased IFNγ,a typical Th1-type cytokine that is important in the formationof granulomas.
The liver granulomas detected in our experiment at 10 dayspostinfection typically were poorly organized with few epi-thelioid cells and with intermingled inflammatory cells, suchas macrophages. These inflammatory cells occasionally har-bored phagocytized antigens. Lymphocytes, plasma cells, andgranulocytes are also present. These findings support possiblya Th2 response, as described in an experimental T. canisinfection in mice (Cuéllar et al. 2001; Fan et al. 2003).
More obvious “tuberculoid” epithelioid granulomas, asexpected in a Th1 response, were observed at 15 days postin-fection. However, eosinophils present chiefly at the peripheryof the granulomatous response in these cases implicate still anassociated Th2 response. Notably, IgE and eosinophilia areimmune hallmarks to worm infections.
As with schistosomiasis, trichinelosis, or filarial wormdisease, antibody-dependent cell-mediated cytotoxicity ispresent in toxocariasis. During late phases of infection,epithelioid granulomas persist, owing to the continuoussupply of toxocaral antigens from encysted larva duringthe infection.
Elevated IL-4 mRNA expression, which we detected inliver tissue at 30 days postinfection, is consistent with asignificant Th2 response, which may be accompanied by atrend toward increased IFNγ and IL-10 mRNA levels. Theseconditions are suggestive of Th2 predominance with a mixed-type local immune reaction in the liver. Our findings areconsistent with those of Pinelli et al. (2007) who reportednearly the same levels of IL-4 mRNA in pulmonary tissue at30 days postinfection. The nonsignificant trends of increasedIFNγ and IL-10 that we observed may be related to theinoculum size because smaller inocula elicited lower re-sponses. IL-10 was only detectable with an inoculum of1000 eggs in the report by Pinelli et al. (2007).
Our results are also in accordance with those of Torinaet al. (2005) who found increased levels of IFNγ and IL-10 in plasma and peripheral blood mononuclear cells ofinfected dogs.
We observed the formation of large ectopic portal lym-phoid aggregates during the late phase of infection, with fewmacrophages and plasma cells present. This intriguing patternis reminiscent of a pseudolymphoma and most likely resultsfrom an inflammatory continuum that begins with diffusemononuclear infiltrates and progresses toward the develop-ment of lymphocytes into nodular secondary lymphoid folli-cles. Portal lymphoid follicle-like structures are nonspecificbut have been linked to chronic inflammatory diseases with animmunologic background. This might be the status in ourexperimental infection and could be related to toxocaralantigens.
In toxocaral infection of the paratenic host, most of thecirculating antigens are derived from larvae and are present invarious host fluids. Host kidneys play an important role in thedisposal of circulating antigens from the peripheral blood. Aclose relationship exists between the supply of parasite anti-gens and possible glomerular lesions, but the detailed patho-genic mechanism controlling lesion development is poorlyunderstood.
In chronic parasitic diseases, the host immune responsereacts to different antigens produced during the different phasesof the parasitic cycle (van Velthuysen and Florquin 2000). Theestablishment of a glomerulopathy associated with a parasiticinfection requires persistent antigenic presentation to the kid-neys, usually over a period of years, as in schistosomiasis (DeBrito et al. 1998, 1999; Nussenzveig et al. 2002) and lessfrequently in visceral leishmaniasis (Sartori et al. 1987) andother parasitic diseases (van Velthuysen and Florquin 2000).Nonspecific mesangioproliferative glomerulonephritis, withglomerular IgM-IgG and C3 usually detected by IF, is probablythe most frequent pattern of parasitic-associated glomerulopa-thy. In humans and various experimental animal models, themesangial area has been suggested as the first and principaltarget of parasitic antigens (Casarosa et al. 1992; De Brito et al.1998; Nussenzveig et al. 2002).
Parasitol Res (2015) 114:809–821 819

Our hamster model proved to be a good model as far asantibody response is concerned. It exhibited a progressive risein anti-toxocaral antibodies, possibly produced by the “classi-cal” pathway or in the context of polyclonal B cell activationand circulating antigens. In the detection of circulating anti-gens by sandwich ELISA, levels vary according to differentprocedures, such as the use of polyclonal and/or monoclonalantibodies, and the treatment of the samples to dissociateantigen-antibody complexes, and also according to theanimal species. Bowman et al. (1987) in a study of infectedmice with T. canis reported that they found 1000 times lesslevels of circulating antigens than another study byMatsumura et al. (1984) in dogs. The decrease in circulatingantigens that we observed during the late phases of infectionlikely resulted from clearance and/or metabolic changes, asdescribed previously by Bowman et al. (1987) and Robertsonet al. (1988). The development of immune complexes and thedetection of circulating antigens may reflect active metabo-lism of the parasite (Aguila et al. 1987). In humantoxocariasis, the high burden of infective larvae leads toelevated antibody production, and the probable developmentof immune complexes could be related to renal involvement.Several investigators have described this relationship betweenToxocara spp. and renal disease (Nada et al. 1996; Shetty andAviles 1999; Zotos et al. 2006).
The infected hamsters developed a modest, focal, nonspe-cific mesangioproliferative glomerulonephritis, chiefly duringthe late phase of infection. This condition was characterizedby the presence of glomerular antigen deposits, as seen byimmunoelectron microscopy, and by localized deposits of Igsand complement, as detected by IF. Antigen deposits wereobserved in all groups of infected animals, suggesting a bind-ing of antibody to antigen that could have been “planted”previously in glomeruli during the physiological process ofantigen clearance. Overall, our model suggests that conditionsexist for the development of preformed immune complexesmediating glomerular disease over time.
The deposition of Igs and complement in the glomerularmesangium and capillary walls is necessary to induce anephropathy-like membranoproliferative glomerulonephritis.This can be associated with chronic infectious diseases in-volving persistent antigen presentation to the kidneys, such asschistosomiasis and hepatitis C (Sethi and Fervenza 2012).Unlike chronic diseases like schistosomiasis or visceral leish-maniasis, the mesangioproliferative glomerulonephritis in ourmodel was focal, rather than diffuse, in the kidneys, and Igdeposits generally were segmental in the glomerulus, suggest-ing mild renal involvement.
Shetty and Aviles (1999) described a nephrotic syndromein a 7-year-old boy coincident with T. canis infection. Treat-ment with corticosteroids resulted in remission of renal symp-toms and abatement of T. canis infection. However, idiopathicminimal change disease, which poses a differential diagnosis
and might be present in a child with toxocariasis, needed to bedefinitively ruled out in this clinical presentation. In an exper-imental murine model of toxocariasis, Hassan and El-Manawaty (1994) described marked mesangioproliferativeglomerulonephritis with mesangial expansion in most of theglomeruli, hyalinosis, and fibrotic adhesions to the Bowman’scapsule. No serological studies were reported. The findings ofthese authors suggested a chronification of the experimentalglomerular disease, a finding that we did not observe in ourhamster model.
Although speculative and potentially of limited relevancefor humans, the present study raises the possibility thatmesangioproliferative glomerulonephritis with nephrotic syn-drome in humans could be an alternative manifestation oftoxocariasis. Consistent with this suggestion, some authorshave speculated that toxocariasis should be considered in thedifferential diagnosis of renal diseases, especially in patientswho do not respond to traditional treatments (Nada et al.1996). Our finding of glomerular TES antigen depositionoccasionally complicated with a mild immune complexmesangioproliferative glomerulonephritis in a hamster modelof toxocariasis supports this possibility.
Acknowledgments We thank Dr. Carmen Arroyo Sanchez for helpingin the statistical analysis. English revision was done by BioMed Proof-reading LLC.
References
Aguila C, Cuéllar C, Fenoy S, Guillén JL (1987) Comparative study ofassays detecting circulating immune complexes and specific anti-bodies in patients infected with Toxocara canis. J Helminthol 61:196–202
Akao N, Ohta N (2007) Toxocariasis in Japan. Parasitol Int 56:87–93Beaver PC, SnyderH CG, Dent J, Laffery J (1952) Chronic eosinophilia
due to visceral larvamigrans: report of three cases. Pediatrics 9:7–19Bilate AMB, Salemi VM, Ramires FJ, de Brito T, Russo M, Fonseca SG,
Faé KC, Martins DG, Silva AM, Mady C, Kalil J, Cunha-Neto E(2007) TNF aggravates experimental chronic Chagas disease car-diomyopathy. Microbes Infect 9:1104–1113
Bilate AM, Teixeira PC, Ribeiro SP, de Brito T, Silva AM, RussoM,KalilJ, Cunha-Neto E (2008) Distinct outcomes of Trypanosoma cruziinfection in hamsters are related to myocardial parasitism, cytokine/chemokine gene expression, and protein expression profile. J InfectDis 198:614–623
Bowman DD,Mika-GrieveM, Grieve RB (1987a) Circulating excretory-secretory antigen levels and specific antibody responses in miceinfected with Toxocara canis. Am J Trop Med Hyg 36:75–82
Bowman DD, Mika-Grieve M, Grieve RB (1987b) Toxocara canis:monoclonal antibodies to larval excretory antigens that bind withgenus and species specificity to the surface of infective larvae. ExpParasitol 64:458–465
Casarosa L, Papini R, Mancianti F, Abramo F, Poli A (1992) Renalinvolvement in mice experimentally infected with Toxocara canisembryonated eggs. Vet Parasitol 42:265–272
820 Parasitol Res (2015) 114:809–821

Chomczynski P, Sacchi N (1987) Single-step method of RNA isolationby acid guanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem 162:156–159
Cuéllar C, Fenoy S, Del Àguila C, Guillén JL (2001) Isotype specificimmune responses in murine experimental toxocariasis. Mem InstOswaldo Cruz 96:549–553
De Brito T, Chieffi PP, Peres BA, Santos RT, Gayotto LCC, Vianna MR,Porta G, Alves VAF (1994) Immunohistochemical detection oftoxocaral antigens in human liver biopsies. Int J Surg Pathol 2:117–124
De Brito T, Carneiro RWC, Nakhle MC, Lima DMC, Abrantes-LemosCP, Sandoval M, Silva AMG (1998) Localization byimmunoelectron microscopy of Schistosoma mansoni antigens inthe glomerulus of the hamster (Mesocricetus auratus) kidney. ExpNephrol 6:368–376
DeBrito T, Nussenzveig I, Carneiro CRW, NakhleMC, Carvalho S, SilvaAMG, Sandoval M, Saldanha LB (1999) Glomerular detection ofschistosomal antigen by immunoelectron microscopy in humanMansonian schistosomiasis. Int J Surg Pathol 7:217–225
De Savigny DH (1975) In vitro maintenance of Toxocara canis larvae anda simple method for production of Toxocara ES antigen for use inserodiagnostic tests for visceral larva migrans. J Parasitol 61:81–782
De Savigny DH, Voller A, Woodruff AW (1979) Toxocariasis: serolog-ical diagnosis by enzyme immunoassay. J Clin Pathol 32:284–288
de St Groth F, Scheidegger D (1980) Production of monoclonal antibodystrategy and tactics. J Immunol Methods 35:1–21
Despommier D (2003) Toxocariasis: clinical aspects, epidemiology, med-ical ecology, and molecular aspects. Clin Microbiol Rev 16:265–272
Elefant GR, Shimizu SH, Sanchez MCA, Jacob CMA, Ferreira AW(2006) A serological follow-up of toxocariasis patients after chemo-therapy based on the detection of IgG, IgA and IgE antibodies byenzyme-linked immunosorbent assay. J Clin Lab Anal 20:164–172
Fahrion AS, Staebler S, Deplazes P (2008) Patent Toxocara canis infec-tions in previously exposed and in helminth-free dogs after infectionwith low numbers of embryonated eggs. Vet Parasitol 152:108–115
Fan C-K, Lin Y-H, DuW-Y, Su K-E (2003) Infectivity and pathogenicityof 14-month-cultured embryonated eggs of Toxocara canis in mice.Vet Parasitol 113:145–155
Fan C-K, Lin Y-H, Hung C-C, Chang S-F, Su K-E (2004) Enhancedinducible nitric oxide synthase expression and nitrotyrosine accu-mulation in experimental granulomatous hepatitis caused byToxocara canis in mice. Parasite Immunol 26:273–281
Fan C-K, Liao C-W, Cheng Y-C (2013) Factors affecting disease mani-festation of toxocarosis in humans: genetics and environment. VetParasitol 193:342–352
Fillaux J, Magnaval JF (2013) Laboratory diagnosis of humantoxocariasis. Vet Parasitol 193:327–336
Glickman LT, Schantz PM (1981) Epidemiology and pathogenesis ofzoonotic toxocariasis. Epidemiol Rev 3:230–250
Glickman LT, Summers BA (1983) Experimental Toxocara canis infec-tion in cynomolgus macaques (Macaca fascicularis). Am J Vet Res44(12):2347–2354
Hassan AT, El-Manawaty NHA (1994) Experimental murine toxocariasishistopathological study of chronic renal infection, transplacentaltransmission and ultrastructural study of egg shell. J Egypt SocParasitol 24:333–340
Kanamura HY, Hoshino-Shimizu S, Silva LC (1981) Solubilization ofantigen S. mansoni adult worms for the passive hemagglutinationtest. Rev Inst Med Trop Sao Paulo 23:92–95
Kazacos KR (1997) Visceral, ocular and neural larva migrans. InPathology of Infectious Diseases - Daniel H Connor, Francis W.Chandler, David A. Schwartz, Herbert J. Manz, Ernst E. Lack,p.1459-1473, Appleton&Lange Ed
Klein JRH, Tazelaar HD, Leslie KO, Colby TV (2010) One hundredconsecutive granulomas in a pulmonary pathology consultationpractice. Am J Surg Pathol 34:1456–1464
Laurenti MD, Corbett CE, Sotto MN, Sinhorini IL, Goto H (1996) Therole of complement in the acute inflammatory process in the skinand in host–parasite interaction in hamsters inoculated withLeishmania (Leishmania) chagasi. Int J Exp Pathol 77:15–24
Laurenti MD, Orn A, Sinhorini IL, Corbett CE (2004) The role ofcomplement in the early phase of Leishmania (Leishmania)amazonensis infection in BALB/c mice. Braz J Med Biol Res 37:427–434
Maizels RM (2013) Toxocara canis: molecular basis of immune recog-nition and evasion. Vet Parasitol 193:365–374. doi:10.1016/j.vetpar.2012.12.032
Maizels RM, Yazdanbakhsh M (2003) Immune regulation by helminthparasites: cellular and molecular mechanisms. Nat Rev Immunol 3:733–744. doi:10.1038/nri1183
Matsumura K, Kazuta Y, Endo R, Tanaka K (1984) Detection of circu-lating antigens in dogs by sandwich enzyme-immunoassay.Immunol 51:609–613
Musso C, Castelo JS, Tsanaclis A, Pereira FEL (2007) Prevalence ofToxocara-induced liver granulomas, detected by immunohisto-chemistry, in a series of autopsies at a children’s reference hospitalin Vitoria, ES, Brazil. Virchows Arch 450:411–417
Nada SM, Abazza BE, Mahmoud LA, Habeeb YS, Hussein HF, AmerOT (1996) Toxocariasis as a cause of renal disease in children inSharkia Governorate, Egypt. J Egypt Soc Parasitol 26(3):709–717
Nagy D, Bede O, Danka J, Szénási Z, Sipka S (2012) Analysis of serumcytokine levels in children with chronic cough associated withToxocara canis infection. Parasite Immunol 34:581–588
Nussenzveig I, Brito T, Carneiro CRW, Silva AMG (2002) HumanSchistosoma mansoni associated glomerulopathy in Brazil.Nephrol Dial Transplant 17:4–7
Ollero MD, Fenoy S, Cuéllar C, Guillén JL, Aguila C (2008)Experimental toxocarosis in BALB/c mice: effect of the inoculationdose on brain and eye involvement. Acta Trop 105:124–130
Othman AA, El-Shourbagy SH, Soliman RH (2011) Kinetics of Foxp-3-expressing regulatory cells in experimental T. canis infection. ExpParasitol 127:454–459
Pinelli E, Brandes S, Dormans J, FonvilleM,HamiltonCM, der Giessen J(2007) Toxocara canis: effect of inoculum size on pulmonary pa-thology and cytokine expression in BALB/c mice. Exp Parasitol115(1):76–82
Robertson BD, Burkot TR, Gillespie SH, KennedyMW,WanbaiF MRM(1988) Detection of circulating parasite antigen and specific anti-body in Toxocara canis infections. Clin Exp Immunol 74:236–241
Rubinsky-Elefant G, Hirata CE, Yamamoto JH, Ferreira MU (2010)Human toxocariasis: diagnosis, worldwide seroprevalences andclinical expression of the systemic and ocular forms. Ann TropMed Parasitol 104(1):3–23
Sartori A, Oliveira AV, Roque-Barreira C, Rossi MA, Campos-Neto A(1987) Immune complex glomerulonephritis in experimental kala–azar. Parasite Immunol 9:93–103
Sethi S, Fervenza FC (2012) Membranoproliferative glomerulonephri-tis—a new look at an old entity. N Engl J Med 366:1119–1131
Shetty AK, Aviles DH (1999) Nephrotic syndrome associated withToxocara canis infection. Ann Trop Paediatr 19:297–300
Torina A, Caracappa S, Barera A, Dieli F, Sireci G, Genchi C, Deplazes P,Salerno A (2005) Toxocara canis infection induces antigen-specificIL-10 and IFNγ production in pregnant dogs and their puppies. VetImmunol Immunopathol 108(1–2):247–251
van Velthuysen MLF, Florquin S (2000) Glomerulopathy associated withparasitic infections. Clin Microbiol Rev 13:55–66
Wilder HC (1950) Nematode endophthalmitis. Trans Am AcadOphthalmol Otolaryngol 55:99–104
Zotos PG, Psimenou E, Roussou M, Kontogiannis S, PanoutsopoulusA, Dimopoulus AM (2006) Nephrotic syndrome as a manifes-tation of Toxocara canis infection. Nephrol Dial Transplant21(9):2675–2676
Parasitol Res (2015) 114:809–821 821