identification and developmental expression of mrnas encoding crustacean cardioactive peptide (ccap)...
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, May 1989, p. 1875-1881 Voi. 9, No. 50270-7306/89/051875-07$02.00/0Copyright © 1989, American Society for Microbiology
Identification and Developmental Expression of a Smooth-Muscley-Actin in Postmeiotic Male Germ Cells of Mice
EUNHEE KIM, SIMON H. WATERS,t LAURA E. HAKE, AND NORMAN B. HECHT*Department of Biology, Tufts University, Medford, Massachusetts 02155
Received 7 November 1988/Accepted 24 January 1989
Mouse testis contains two size classes of actin mRNAs of 2.1 and 1.5 kiobases (kb). The 2.1-kb actin mRNAcodes for cytoplasmic ,1- and y-actin and is found throughout spermatogenesis, while the 1.5-kb actin mRNAis first detected in postmeiotic cells. Here we identify the testicular postmeiotic actin encoded by the 1.5-kbmRNA as a smooth-muscle y-actin (SMGA) and present its cDNA sequence. The amino acid sequence deducedfrom the postmeiotic actin cDNA sequence was nearly identical to that of a chicken gizzard SMGA, with oneamino acid replacement at amino acid 359, where glutamine was substituted for proline. The nucleotidesequence of the untranslated region of the SMGA differed substantially from those of other isotypes ofmammalian actins. By using the 3' untranslated region of the testicular SMGA, a highly specific probe wasobtained. The 1.5-kb mRNA was detected in RNA from mouse aorta, small intestine, and uterus, but not inRNA isolated from mouse brain, heart, and spleen. Testicular SMGA mRNA was first detected and increasedsubstantially in amount during spermiogenesis in the germ cells, in contrast to the decrease of the cytoplasmic,I- and y-actin mRNAs towards the end of spermatogenesis. Testicular SMGA mRNA was present in thepolysome fractions, indicating that it was translated. These studies demonstrate the existence of an SMGA inmale haploid germ cells. The implications of the existence of an SMGA in male germ cells are discussed.
The mammalian actin multigene family codes for at leastfour muscle actins (a-cardiac, a-skeletal, -y-enteric smooth,and a-vascular smooth) and two cytoplasmic actins (1 and -y)(49). Another actin with a more acidic charge has also beenidentified in mice (48). Although amino acid sequences ofthese genes are very homologous (>90%), their respectivemRNAs are more divergent due to many silent nucleotidereplacements in the coding region (34) and substantial differ-ences in the 3' and 5' untranslated regions. The cytoplasmic,B- and y-actins are present in varying amounts in all non-muscle cells. The muscle actins exhibit more tissue and celltype specificity.
Spermatogenesis, the differentiation of germ cells, is acomplex process (see references 1 and 18 for reviews).Diploid stem cells, the spermatogonia, proliferate by way ofa series of mitotic divisions and then begin meiosis with aprolonged prophase. Following two cell divisions, haploidcells, the spermatids, appear. The haploid spermatids differ-entiate into mature sperm during spermiogenesis, a lengthyprocess which takes 2 weeks in the mouse. To make theflagellated mature spermatozoon, reductions in ploidy anddramatic morphological changes must take place. Roundcells elongate, the nucleus condenses, sperm tails and ax-onemes form, and most of the cytoplasm is pinched off in theform of residual bodies.Mouse testis contains two size classes of actin mRNAs of
2.1 and 1.5 kilobases (kb) (51). The 1.5-kb actin mRNA ispredominantly, if not solely, expressed in postmeiotic germcells. During spermiogenesis, the 2.1-kb actin mRNAs cod-ing for the cytoplasmic ,B- and -y-actins decrease severalfold,while the 1.5-kb actin mRNA increases. The postmeiotic1.5-kb actin mRNA does not hybridize to 3' untranslated
* Corresponding author.t Present address: Department of Molecular Genetics and Micro-
biology, University of Massachusetts Medical School, Worcester,MA 01605.
(3'-UT) probes specific for cytoplasmic 1- and -y-, cardiac,and skeletal a-actins (51).To identify the actin isotype coded by the 1.5-kb mRNA in
the mouse testis, we have isolated cDNA clones coding forthe 1.5-kb actin mRNA from a mouse testicular residual-body cDNA library and a mouse total-testis cDNA library.In this report we identify the postmeiotic testicular actin asa smooth-muscle -y-actin (SMGA). To our knowledge, this isthe first nucleotide sequence presentation of an SMGA inmammals. Using the 3'-UT of the testicular SMGA as anisoactin-specific probe, we examined the developmentalexpression of the SMGA during mouse spermatogenesis.
MATERIALS AND METHODSIsolation of actin cDNA clones. A Xgt-10 cDNA library,
prepared from polyadenylated RNA isolated from purifiedresidual bodies from CD-1 mice, was screened by plaquehybridization with a mouse 1-actin coding-region probe(pAL41) (34). Eight positive clones were isolated and char-acterized by restriction mapping. None of these clones had apoly(A) tail. To complete the cDNA sequence, a Xgt-10cDNA library of mouse total testis, constructed and kindlyprovided by K. Kleene, was screened with one of the actinclones isolated from the mouse residual-body cDNA library.Four additional clones, three of which had poly(A) tails,were isolated and sequenced. A diagram for the overlappingclones is shown in Fig. 1. Appropriate fragments weresubcloned into M13 mpl8 or mpl9 vectors so that the cloneswould generate opposite strand sequences. The clones weresequenced by the dideoxy chain termination method (40, 41).
Isolation of a 3'-UT subclone of the 1.5-kb actin mRNA. A400-base-pair (bp) EcoRI fragment which includes some ofthe C-terminal amino acid coding sequence (from amino acid272 to the termination codon), the 3'-UT, and 6 bp of poly(A)tail was initially subcloned into M13 mpl9. Exonuclease IIIdigestions (Bethesda Research Laboratories, Inc.) wereused to unidirectionally delete most of the amino acid-codingregion of the clone, allowing the 3'-UT of the 1.5-kb actin
1875

1876 KIM ET AL.
I I
AUG
I
u% El ll0
UAA
ioobp
2
3
4
5
6
FIG. 1. Partial restriction map and diagram of overlappingmouse SMGA cDNA clones. Clones 1 and 2 were isolated from theresidual-body cDNA library. Clones 3, 4, 5, and 6 were isolatedfrom a total-testis cDNA library.
mRNA to be isolated (20). The clones generated afterexonuclease III digestion were screened, and a clone whichhad only 6 bp of amino acid-coding region plus the UAAtermination codon and the entire 74 bp of the 3'-UT and 6 bpof poly(A) tail was identified. The clone was used as a
specific probe for the 1.5-kb actin mRNA.Since the insert was too short to be labeled efficiently by
nick translation, the plasmid was labeled by a primer exten-sion method which is a modification of the M13 sequencingreaction (40, 41). One microgram of M13 single-strandedphage DNA and 8 ng of M13 17-bp universal sequencingprimer (Bethesda Research Laboratories) were boiled for 5min and reannealed by slow cooling at room temperature.After primer annealing, the plasmid was incubated at 30°Cwith 40 ,uM each unlabeled dATP, dGTP, and dTTP and 100,Ci of 32P-labeled dCTP (3,000 ,XCi/mmol; New EnglandNuclear Corp.) in 10 mM Tris (pH 8.0)-10 mM MgCl2-30mM NaCI-30 mM dithiothreitol-1 U of Klenow enzyme(Bethesda Research Laboratories). After 15 min, a finalconcentration of 16 ,uM unlabeled dCTP was added, and theincubation was continued for an additional 15 min at 30°C.The reaction mixture was then passed through a SephadexG-50 spin column in a 1-ml syringe and used for hybridiza-tion.RNA preparations. CD-1 mice of different ages were
obtained from Charles River Breeding Laboratories, Inc.Pachytene spermatocytes, round spermatids, and residualbodies were isolated as described previously (24, 33, 39).RNAs were purified by the guanidine isothiocyanate proce-dure as described previously (24).
Northern (RNA) blots. RNA samples were denatured withformamide, electrophoresed in 1.1% agarose gels, and trans-ferred to nitrocellulose (Schleicher & Schuell, Inc.). Thefilters were baked at 80°C for 1 to 2 h and subsequentlyprehybridized for 1 to 4 h at 65°C in 5x Denhardt solution(0.1% polyvinylpyrrolidone, 0.1% Ficoll, and 0.1% bovineserum albumin)-0.4 M sodium phosphate buffer (pH 7.0)-0.1% sodium dodecyl sulfate (SDS)-100 ,ug of sheared,denatured salmon sperm DNA (Sigma Chemical Co.) per ml.
Filters were hybridized overnight in the same buffer with a
total of 0.5 x 107 to 5 x 107 cpm of the nick-translatedpAL41 probe (34) or 2 x 107 to 5 x 107 cpm of the 3'-UT ofthe testicular SMGA labeled by primer extension. Afterhybridization, the filters were washed twice for 15 min eachat room temperature in 0.24 M sodium phosphate buffer (pH7.0)-0.1% SDS and then twice for 30 min each at 65°C in 0.4M sodium phosphate buffer (pH 7.0-0.1% SDS. The filters
were dried and autoradiographed at -70°C with Kodak XARfilm and one intensifying screen.Polysome gradients. Testes from 10 mice were decapsu-
lated, and spermatogenic cells were removed from theseminiferous tubules by successive collagen and trypsintreatments as described previously (25, 39). Cells werepelleted, washed in phosphate-buffered saline, and sus-pended in 1 nml of 0.1% diethylpyrocarbonate (DEPC)-treated 20 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid, pH 7.6) containing 100 mM KCI, 1.5mM MgCl2, and 50 ,ug of cycloheximide per ml. The deter-gent Nonidet P-40 was added to a final concentration of1.5%, the cells were homogenized at 4°C on ice with 10strokes of a Potter homogenizer, and the nuclei were pel-leted with a 1-min spin in an Eppendorf centrifuge at 4°C.The supernatant was layered over a 10-ml, 10 to 40% (wt/wt)sucrose gradient that had a'1-ml cushion of 60% sucrose. Thesucrose was dissolved in 20 mM HEPES (pH 7.4)-100 mMKCI-and contained either 1.5 mM MgCl2 or 10 mM EDTA.Gradients were cerltrifuged for 2 h at 35,000 rpm in an SW41rotor at 4°C. The bottoms of the tubes were punctured witha 28-gauge needle, and six equal fractions were collecteddirectly into a 1:1 mixture of phenol and CHCl3. Following aCHCl3 extraction, the RNA was ethanol precipitated andanalyzed as above.
RESULTSIsolation and characterization ofcDNA clones coding for the
postmeiotic 1.5-kb actin mRNA. Six cDNA clones coding fora 1.5-kb actin mRNA (Fig. 1) were isolated from a mousetesticular-residual-body cDNA library and a mouse total-testis cDNA library as described in Materials and Methods.The isolated clones were 0.8 to 1.1 kb in length. Sequenceanalysis of the six clones allowed the predicted amino acidsto be determined (Fig. 2). The amino acids were numberedaccording to the system of Vandekerckhove and Weber (49).The amino acid sequence deduced from the cDNA sequenceidentifies the testicular postmeiotic actin as an SMGA (49).The amino acid sequence of this actin was nearly identical tothat of an SMGA from chicken gizzard, with one substitutionat amino acid 359, where glutamine was substituted forproline (49).When the nucleic acid sequence of the amino acid-coding
region of the testicular SMGA was compared with that ofmouse skeletal actin (34), mouse cardiac actin (14), andmouse ,-actin (45), homologies of 84%, 84%, and 83%,respectively, were found (Fig. 2). The amino acid sequencedifferences between testicular SMGA and mouse skeletalactin (98%), mouse cardiac actin (99%), and mouse P-actin(93%) were less. It appears that sarcomeric actins have moresilent base substitutions than P-actin, leading to fewer aminoacid changes. The nucleotide usage in the third base positionof the codon in mouse actin mRNAs showed a preference foreither C or G, an observation seen with other mammalianactins (9) (Table 1). In the four mouse actins compared here,A was used least frequently.3'-UT of testicular SMGA is isotype specific. The testicular
SMGA has 74 bp of 3'-UT, which is the shortest 3'-UTamong mammalian actin sequences published to date. Whenthe 3'-UT of the testicular SMGA was compared with the3'-UTs of the mouse cytoplasmic P-actin (45), mouse cyto-plasmic ly-actin (our unpublished data), mouse skeletal actin(30), mouse cardiac actin (14), and rat aortic actin (27), nosignificant homology was found. Therefore, all of the sixmammalian actins known to date apparently have isotype-specific 3'-UTs.
MOL. CELL. BIOL.
I

VOL. 9, 1989 TESTICULAR SMOOTH-MUSCLE ACTIN 1877
5 12... 15met MM b Ala Leu Val LQysl Asp Asn Gly Ser Gly ggoj
Skeeal tcaggtgctccagccccagctcattcaccgctccaccA TOT GAA GAA GAG ACC ACC GCC CrT GM~TOT GAG AAT GOT TOT GOC CTG TOCCarletal C..C ... GAC........T ..........C........GGMCardiac ..~~~~~~~~~~~~C..CGGM........TT.G........T....C..G ..G GMis .T..C ..T.T. G.T .G ..G ..CGTC .TO..C ...C ... AXG.
20 25 30 35 40 45 50Lys Ala Gly Phe Ala Gly Asp Asp Ala Pro Arg Ala Val Phe Pro S.r Ile Met Gly Mrg Pro Arg His Gin Gly Val M4et Val Gly Met Gly Gin Lys Asp
SWh MAG OCA GOC TTT? GCA GGA OAT CAT OCO CCC AGO GOT OTC TI CCC TC ATT GTO GGC COC CCT AGA CAT CAG GOT OGOM OTOG GOA AMO GGC CAG AMA GAGSkeletal ..A ..T......C..G......C..0G....A ....C ......A..C C.T ..C ......C.....A ..T ...T.0.G...Cardiac ...C ......G..T...C ..C.A. ..C ..G ....C ...T ...A C.C ..C......C .. T ... ....G...0s ..A ..C ....C..G ..C ..C ...T ... C....A ........C ..........G..C..............T.0.G...
55 60 65 70 75 80 85Ser Tyr Val Gly Asp Glu AlaoGln Ser Lys Arg GlyIle LeumTr LeuLys Tyr Pro Ile GluHis GlyIle le Thr Asn Trp Asp Asp Met Glu Lys Ile
SMA MGC TA.T GMO GOG GAG GAG GOT CAG MGC AMG COT GOG AMC OTA ACT CIC AMGTAC COT ATT GAA CAT GOC ATC ATC ACC MAC TOO CAT GAGCAT GMG AMG ATCSkeletal TC. ..C ...T ..T ...C ........A..T ..G ..C..G . C....................C.Cardiac TO .A ..T ..T ..A..C ........A..T ..G. 0.GC . T0 TO. .T ......C.......A.A ..T.G..C..G.C..T G.T ........C..........
90 95 100 105 110 115Trp His His Ser Phie Tlyr Msn Glu Leu Mrg Val Ala Pro Giu Glu His Pro Leu Leu Thr Glu Ala Pro Leu Msn Pro Lys Ala Msn Arg Giu Lys M4et
SMrA TOO CAG CAM TOC TIC TAC AAT GMG CIT CCA GTA GCA CCA GAA GAG CAG CCC ACC CTO CTC ACA GAG GCC CCC CT AAMC CCC MAA GCA MPC MGA GM MAG AMSkeletal.......A..OG..T ..G..C ..T..G......G..T ......C......0...G..T...C.G ......Cardiac ......TA.A . :.C..T ..G..C ...G ..A ...A........T .%.....G..G ...'.'C C.T ......0 . ....A... . . ..G ..T ..G ..C ..T G.....O. TOTOGG......C...... 0.. . .T ..G....C.C.T ..A ....
120 125 130 135 140 145 150Thr Gin Ile met Phae Giu Thr Phe Msn Wa Pro Ala Met Tyr Val Ala Ile Gin Ala Val Leu Ser Leu Tyr Ala Ser Giy Mrg Th¶r Thr Giy Ile Val [l7u"
SMM ACC CMG AC AM TTC GAACMC TTC AAT GTC COT 0CC MC TAT OTT GOT AlT CMG GOT GMOTCI TOA CTC TAT GCA TOC GOC COT AM ACA GOC AlT OTT CMSkeletal ..T ..A ......T G.....O. .C..G ..........G ...C ..G..G ..C . .T ........T..C...C ..G T..Cardiac . T G.........G ..........C..C ......A..G..T ..G ..T..T..AA.A..C ..........0.~~~~~~: :::..T ..G.CA...CW. ..A ......C..A ..C ..C ........OG..C ..G ..C ..T ..T ............GA..
155 160 165 170 175 180 185Asp Ser Gly Asp Giy Val Thr His Wj Val Pro Ile Tyr Glu Giy Tyr Ala Leu Pro His Ala Ile Met Mrg Leu Asp Leu Ala Giy Mrg Asp Leu Thr Asp
SW3A OAT TOO 000 OAT GOC GTO AMC CAdAT GTC CCC ATC TAT GMG GOC TAT GCA CTG CCC CAC 0CC AMC MC COT CiT GAG LtO GOT GOA COO OAT CTO AdA GAGSkeletal ....T ..C ..T ........C..G ..............C..G ..A..........0.. . ..C ..T ..C..C ..T ...Cardiac ..T ..C ..T ..A ..T ..T....T ..........T. CT.A ..T..T ..T . .G ..T..A ..A...... C. T..T0 ..C ..C ..T ..C G.......C..G.....0. .C........T..C ..T.......C.. .G ...T.. .C.....C ..G ....
190 195 200 205 210 215 220Tyr Leu Met Lye Ilie Leu Thr Giu Mrg Giy Tyr Ser Phe Ua lhr Thir Ala Giu Mrg Giu Ile Va Mrg Asp lie LYe Giu Lye Leu Cys Tyr Val Ala Leu
SMCA TAC CIVTC AMAG ATT CTC ACA GAA AM GO CTT TCC 7IT GM MCC MAC GOT GMG AtA GAA ATT GTA COA GAG MTC AAG GAG AMG cm~7rC TAT G'TA 0CC CTGSkeletal . 0.G ..A ..C ..T ..GC.T........C..... ..A C.T ..G .G ..C . .A.G0....Cardiac ..........C..T..G C.C ..G..C . .C ..T ..A C.T ......C..T .A..A.AT.C..... T..0....... .C ..G ..C ..G C.T ...CMAG.:CACMC........0.. ..C ..G ..T.. ...A..T ..T ..A
225 230 234a 235 240 245 250Asp Phe Glu M Giu Met Ala Thr Ala Ala Ser Ser Ser Ser Leu Glu Lye Ser Tyr Glu Leu Pro Asp Giy Gin Val Ilie Thr Ilie Giy Msn Giu Mrg Phe
SMCA CAT TTC GMG MAT GMG MC GCC MCA GCA GOT TCA TOT TOT TOC CMO GMG AMAAC VW GMG TM COT CAT 000 CMG GTC MC ACT AlT GGC MAC GMG COd TTCSkeletal ..C ..............C..T ..C ..T ..C..C .0......G..T ... C.. ..C..C .C ..C ....T ..T...Cardiac ....T ..C.....T ..A ..T ..C..C.0... .T ..AC.T.C...C. :.A ............0 ..C .....C.G .... 'T ..C..A ..C...C.0...G..T ... C....C ..C..A...0.............. .
255 260 265 270 275 280 285Mrg Cye Pro Giu ER Leu Phe Gin Pro Her PheUn Giy Met Giu Ser EJ Gly Ilie His Glu Thr 1Thr fi Msn Her 1ie Met LyedeAplivs i
SMIGA COC TOC COG GMG MCC CIC TIC CMG COT TOC TIC ATC GOd A GMG TCA GOT GGCAT?C CAT GA A AM A TAMCAT TOC MC AMA MTGCAMO TdM GCAdTCSkeletal ..T .0...... ............T..C..T......T..G ..G ..C....G..C ..C..CMAG.........C ..C ....Cardiac . T ..A....T........C.T.....?. .T .....A ..T ...C .......T ..T .-CMAG............ .
is :A..A .T ... G.T ..T...........T.G ..T....A..C TG...C ..C ......T.0T...............G.290 295 300 305 310 315 320
Mrg Lye Asp Leu Tyr Ala Asn M Val MJ Ser Giy Giy [hr [hr Miet Tyr Pro Giy Ile Ala Asp Mrg Met Gin Lye Giu Ile [hr Ala Leu Ala Pro SerSmGA CGC AMA OAT TMO TAT GOT MC MAT GTO CTC TOT 000GGOC ACT MC AMC TAd COT GOC ATT GOT GAC MGO MCG CM AAG GAA MC MAO 0CC TTG GOT CCC MGCSkeletal AXG..G ..C C....C..C ... AXG..A......C..........T-..C.....C.C 6. ..0.. . .TC... . .Cardiac. .C C....C . A..A .. -C ..........T ..C ..T C.T... ..A........T..AC... . .0'.T ...CC.C. C.....C .'G'..G ..T ..T ..C ........A...................G ..T ..T ..TC.T.....T
325 330 335 340 345 350 355[hr Met Lye Ilie Lye Ilie Ilie Ala Pro Pro Glu Mrg Lye Tyr Her Val Trp Ile Giy Giy Her Ile Leu Ala Her Leu Her [hr Phe Gin Gin met Trp IleSMOA MCC AMC AAG MTC AM MCT MC GOT COT COT GMG COO AAG TAC TOA GTC TOO MCT GOC GOC TOC MC CM 0CC TOT CTC TOC AMC TIC CMG CAA AMC TMO MTC
Skeletal ............C ..C ..C ......C .0...... ..C ..T............G..G........0.. . . .Cardiac .. .T.C....T...A .T .0..0....A.. . .... . . ...... . .... .G . .... . ...... .
Is ....... . . .C ..T ..........C ......T..GO..C ..T ............A..G..........G......360 365 370
iia LYSEaIGlu Tyr Asp Glu iJGly Pro Her Ilie Val Hfis Mrg Lye Cys PheSMCA MGC AM CCA GMGTAT GAT GMOA 000 CCC TCCMCATO CAG AMO AMA C TTC MAA a9rtcagagggccttctctaggggatccccacaacttgqtcaccagccacagatcaU&Skeletal .C.M..A .C ..C ..T ..C........G... C.C .0......Cardiac ......AM.. C..A........G. C.C..G.CA...G0s .... M.C.: : 'T."C. .'C ......c..G ... C.C ..GO.0....
SMOA aaaccttcaagccttaaaaaFIG. 2. Comparison of mouse actin sequences. The predicted sequence of the amino acid-coding region of the testicular SMGA is
compared with those of mouse skeletal actin (30), mouse cardiac actin (14), and mouse 13-actin (46). A dot under a nucleotide indicates identitywith the mouse testicular SMGA sequence. Amino acids of the mouse testicular SMGA that differ from those in mouse skeletal, mousecardiac, or mouse 13-actins are boxed. The 5'-UT and 3'-UT sequences are written in lowercase letters. The poly(A) addition signal isunderlined.
1878 KIM ET AL.
TABLE 1. Nucleotide usage in the third base position of thecodon in mouse actin mRNAsa
Actin % of codons(no. of aaa) A C G T
SMGA (376) 14.4 38.0 25.5 22.1Skeletal (377) 5.6 44.8 31.6 18.0Cardiac (377) 11.4 37.1 24.7 26.8,B-Actin (375) 6.4 42.8 29.7 21.1
a aa, Amino acids.
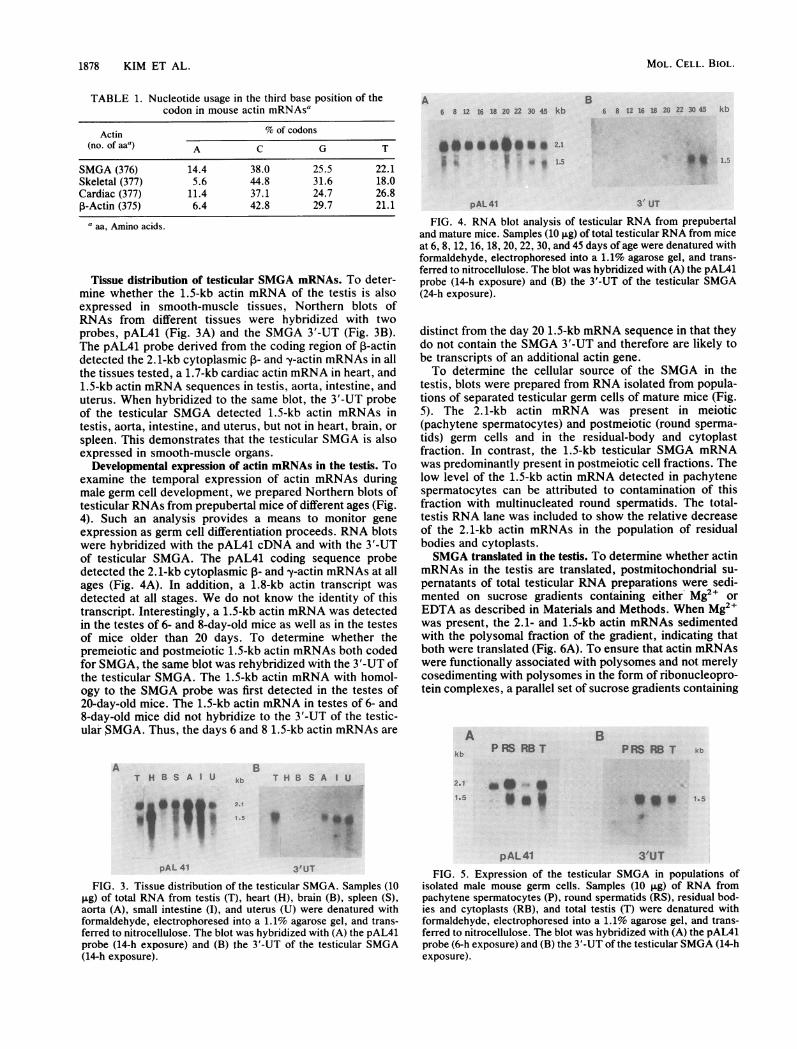
Tissue distribution of testicular SMGA mRNAs. To deter-mine whether the 1.5-kb actin mRNA of the testis is alsoexpressed in smooth-muscle tissues, Northern blots ofRNAs from different tissues were hybridized with twoprobes, pAL41 (Fig. 3A) and the SMGA 3'-UT (Fig. 3B).The pAL41 probe derived from the coding region of ,B-actindetected the 2.1-kb cytoplasmic 1- and y-actin mRNAs in allthe tissues tested, a 1.7-kb cardiac actin mRNA in heart, and1.5-kb actin mRNA sequences in testis, aorta, intestine, anduterus. When hybridized to the same blot, the 3'-UT probeof the testicular SMGA detected 1.5-kb actin mRNAs intestis, aorta, intestine, and uterus, but not in heart, brain, orspleen. This demonstrates that the testicular SMGA is alsoexpressed in smooth-muscle organs.
Developmental expression of actin mRNAs in the testis. Toexamine the temporal expression of actin mRNAs duringmale germ cell development, we prepared Northern blots oftesticular RNAs from prepubertal mice of different ages (Fig.4). Such an analysis provides a means to monitor geneexpression as germ cell differentiation proceeds. RNA blotswere hybridized with the pAL41 cDNA and with the 3'-UTof testicular SMGA. The pAL41 coding sequence probedetected the 2.1-kb cytoplasmic 1- and -y-actin mRNAs at allages (Fig. 4A). In addition, a 1.8-kb actin transcript wasdetected at all stages. We do not know the identity of thistranscript. Interestingly, a 1.5-kb actin mRNA was detectedin the testes of 6- and 8-day-old mice as well as in the testesof mice older than 20 days. To determine whether thepremeiotic and postmeiotic 1.5-kb actin mRNAs both codedfor SMGA, the same blot was rehybridized with the 3'-UT ofthe testicular SMGA. The 1.5-kb actin mRNA with homol-ogy to the SMGA probe was first detected in the testes of20-day-old mice. The 1.5-kb actin mRNA in testes of 6- and8-day-old mice did not hybridize to the 3'-UT of the testic-ular SMGA. Thus, the days 6 and 8 1.5-kb actin mRNAs are
A6 8 12 16 18 20 22 30 45 kb
B6 8 12 16 18 20 22 30 45 kb
sesOes a 2.1
iI 0 1.5
pAL 41 3' UT
FIG. 4. RNA blot analysis of testicular RNA from prepubertaland mature mice. Samples (10 ,ug) of total testicular RNA from miceat 6, 8, 12, 16, 18, 20, 22, 30, and 45 days of age were denatured withformaldehyde, electrophoresed into a 1.1% agarose gel, and trans-ferred to nitrocellulose. The blot was hybridized with (A) the pAL41probe (14-h exposure) and (B) the 3'-UT of the testicular SMGA(24-h exposure).
distinct from the day 20 1.5-kb mRNA sequence in that theydo not contain the SMGA 3'-UT and therefore are likely tobe transcripts of an additional actin gene.To determine the cellular source of the SMGA in the
testis, blots were prepared from RNA isolated from popula-tions of separated testicular germ cells of mature mice (Fig.5). The 2.1-kb actin mRNA was present in meiotic(pachytene spermatocytes) and postmeiotic (round sperma-tids) germ cells and in the residual-body and cytoplastfraction. In contrast, the 1.5-kb testicular SMGA mRNAwas predominantly present in postmeiotic cell fractions. Thelow level of the 1.5-kb actin mRNA detected in pachytenespermatocytes can be attributed to contamination of thisfraction with multinucleated round spermatids. The total-testis RNA lane was included to show the relative decreaseof the 2.1-kb actin mRNAs in the population of residualbodies and cytoplasts.SMGA translated in the testis. To determine whether actin
mRNAs in the testis are translated, postmitochondrial su-pernatants of total testicular RNA preparations were sedi-mented on sucrose gradients containing either Mg2+ orEDTA as described in Materials and Methods. When Mg2+was present, the 2.1- and 1.5-kb actin mRNAs sedimentedwith the polysomal fraction of the gradient, indicating thatboth were translated (Fig. 6A). To ensure that actin mRNAswere functionally associated with polysomes and not merelycosedimenting with polysomes in the form of ribonucleopro-tein complexes, a parallel set of sucrose gradients containing
Akb
BP RBS RB T PRS RB T kb
It 1.5
T H B S A U kb
BT H B S A I U
2.1 * *-.
1.5
0,W.
pAL 41pAL 41
3'UT
FIG. 3. Tissue distribution of the testicular SMGA. Samples (10pg) of total RNA from testis (T), heart (H), brain (B), spleen (S),aorta (A), small intestine (I), and uterus (U) were denatured withformaldehyde, electrophoresed into a 1.1% agarose gel, and trans-ferred to nitrocellulose. The blot was hybridized with (A) the pAL41probe (14-h exposure) and (B) the 3'-UT of the testicular SMGA(14-h exposure).
3'UT
FIG. 5. Expression of the testicular SMGA in populations ofisolated male mouse germ cells. Samples (10 ,ug) of RNA frompachytene spermatocytes (P), round spermatids (RS), residual bod-ies and cytoplasts (RB), and total testis (T) were denatured withformaldehyde, electrophoresed into a 1.1% agarose gel, and trans-ferred to nitrocellulose. The blot was hybridized with (A) the pAL41probe (6-h exposure) and (B) the 3'-UT of the testicular SMGA (14-hexposure).
A
MOL. CELL. BIOL.
1.50 0 0 2.1
I I i .5
1:

TESTICULAR SMOOTH-MUSCLE ACTIN 1879
A ++ B EDTAa
1 2 3 4 5 6 1 2 3 4 5 6
Ia**go,,A O
2.1
1.1
FIG. 6. RNA blot analysis of SMGA in testicular polysomes.Each lane contains 10 1Lg ofRNA isolated from a 10 to 40%o sucrose
gradient of a total-testis extract. Gradients were run in the presence
of (A) 10 mM Mg2" (24-h exposure) or (B) 10 mM EDTA (24-hexposure). Fraction 1 is the top of the gradient. The RNAs weredenatured with glyoxal, electrophoresed into a 1.1% agarose gel,and transferred to nitrocellulose. The blot was hybridized with thepAL41 probe.
EDTA in place of Mg2+ were run (Fig. 6B). The presence ofEDTA dissociated polysomes, resulting in a shift of the2.1-kb and 1.5-kb actin mRNAs from the polysome to thenonpolysomal fraction of the gradients, indicating that both2.1-kb and 1.5-kb actin mRNAs are functionally associatedwith polysomes.
DISCUSSION
We have identified from cDNA sequence analysis an
SMGA that is haploid expressed in the mouse testis. To our
knowledge, this is the first presentation of the nucleotidesequence for an SMGA in mammals. Codons for the N-terminal Met and Cys amino acids, which are thought to beremoved by posttranscriptional processing in muscle (53),are present in the nucleotide sequence. The N-terminal Metand Cys codons are also found in other mammalian andavian muscle actins (4, 12, 16, 17, 21, 47, 53). The same twocodons are also found in all six Drosophila actin genes (13),four nematode actin genes (10), and sea urchin actin genes
(6, 42). However, the same two codons are not present inmammalian and avian cytoplasmic actins (3, 9, 29, 35).The expression of an SMGA mRNA in nonmuscle germ
cells was unexpected. Because it is developmentally regu-
lated during spermiogenesis, we wished to determinewhether the same transcript is present in both haploid malegerm cells and smooth-muscle tissues. In Dictyosteliumdiscoideum, different actin genes coding for the same actinare expressed differentially (46). From the results of theNorthern blot analysis with the 3'-UT testicular SMGAprobe of RNAs from smooth-muscle tissues (Fig. 3), we
believe that the same transcript is present in haploid germ
cells and in smooth-muscle tissues. Moreover, preliminarygenomic Southern blot analysis of mouse DNA with the3'-UT of the testicular SMGA suggests that the SMGA is a
single-copy gene (data not shown). However, we cannotexclude the possibility that differential splicing or alternativetranscription initiation might generate different 5'-UT se-
quences in testis and smooth muscle.Evidence for haploid transcription during spermatogenesis
is accumulating. The two mouse protamine genes (reviewedin reference 24; N. B. Hecht, in L. Hnilica, G. Stein, and J.Stein, ed., Histones and Other Basic Proteins, in press),mouse testicular a-tubulin (8, 50), and mouse, rat, andhuman transition proteins (5, 19, 22, 23, 26, 31; P. C. Yelick,Y. K. Kwon, J. F. Flynn, K. C. Kleene, and N. B. Hecht,Mol. Reprod. Dev., in press) are first transcribed in haploidgerm cells. In addition to these genes which appear to be
testis specific, several proto-oncogenes also have a develop-mentally regulated pattern of transcription in the testis (seereference 38 for a review). A 1.7-kb c-mos transcript (15), a4.7-kb c-abl transcript (32, 37), and a 2.6-kb c-int-1 transcript(43) were detected mainly in haploid germ cells. mRNAsfrom c-mos of 6, 2.3, and 1.3 kb, c-abl of 7 and 6 kb, andc-int-1 of 2.6 kb are also detected in somatic tissues. Al-though no differences in mRNA size for SMGA actin aredetected in testicular and somatic tissues, a similar pattern ofdevelopmental regulation is seen.An inverse relationship between cell differentiation and
cytoplasmic actin expression has been observed (28, 36).The decrease of the 2.1-kb cytoplasmic actin mRNAs duringlate spermiogenesis and the predominant expression of thetesticular SMGA mRNA in haploid male germ cells show apattern analogous to those found during the in vivo differ-entiation of rat aortic smooth muscle, where a decrease incytoplasmic actin is followed by an increase in smoothmuscle a-actin (28). In both cases, cytoplasmic actin de-creases as cell differentiation proceeds. In addition, a 40-bpsequence element which is sufficient to downregulate tran-scription during myogenesis has been identified in the 3'-UTof the 1-actin gene (7). We cannot determine from our datawhether the decreased level of cytoplasmic actin mRNAsduring spermiogenesis is caused by reduced transcription orby a change in mRNA stability. However, if some level offunctional actin is required for continued germ cell differen-tiation, we speculate that the SMGA gene may be expressedin spermiogenesis to compensate for the observed downreg-ulation of the 1- and y-actins. In this case, the cytoplasmicactins and the smooth-muscle actin would be functionallyinterchangeable, and the designation of SMGA for the tes-ticular 1.5-kb mRNA might be too restrictive.
Alternatively, the smooth-muscle actin might have uniqueproperties required for the physiological needs of late sper-miogenic cells. It has been speculated that actin might beinvolved in acrosome shaping (52). It is also conceivable thatactin plays a role in sequestering the cytoplasm as residualbodies are pinched off from the elongating spermatids. Thetiming of acrosome shaping and residual-body formationcertainly coincides with the expression of the testicularSMGA. Actins are also found in mammalian spermatozoa (2,3, 11). The presence of actin and calmodulin together inspermatozoa (3) suggests that actin could also play a role incapacitation and in the acrosome reaction in the mammalianspermatozoon.The presence of a 1.5-kb actin mRNA in the testes of 6-
and 8-day-old mice is intriguing. Although its mRNA size issimilar to that of the testicular SMGA, the 3'-UT of thetesticular SMGA does not hybridize to it. A 1.7-kb actinmRNA was shown to be expressed in the type A sper-matogonia of the rat (44). The 1.7-kb rat actin mRNA wasnot detected in testicular somatic cell types or in other germcells of the rat testis. These and our results suggest thatspermatogonia may contain an additional isotype. In thetestis of 8-day-old mice, type A spermatogonia are thepredominant germ cell. If so, this 1.5-kb actin could serve asa valuable marker for type A spermatogonia. The identity ofa 1.5-kb actin mRNA in the testis of 6- and 8-day-old mice iscurrently under investigation.
ACKNOWLEDGMENTS
We thank M. Buckingham for kindly providing mouse actincDNA probe pAL41, J. Trasler for her critical comments on thismanuscript, and V. Ricciardone for her excellent typing of Fig. 2.
VOL. 9, 1989

1880 KIM ET AL.
This work was supported by Public Health Service grant GM29224 from the National Institutes of Health and by EnvironmentalProtective Agency grant CR 813481, administered by the Center forEnvironmental Management of Tufts University.
ADDENDUM IN PROOF
A SMGA cDNA clone has been isolated from a ratstomach cDNA library (K. M. McHugh, and J. L. Lessard,Mol. Cell. Biol. 8:5224-5231, 1988). The predicted aminoacid sequences of the rat SMGA and the one reported hereare identical. They share 95% nucleotide similarity in thecoding regions and 96% similarity in the 3' untranslatedregions.
LITERATURE CITED1. Bellve, A. R. 1979. The molecular biology of mammalian sper-
matogenesis, p. 151-261. In C. A. Finn (ed.), Oxford reviews ofreproductive biology. Oxford University Press, New York.
2. Camatini, M., A. Casale, and M. Cifarelli. 1987. Immunocyto-chemical identification of actin in rabbit spermatogenesis andspermatozoa. Eur. J. Cell. Biol. 45:274-281.
3. Casale, A., M. Camatini, 0. Skaili, and G. Gabbiani. 1988.Characterization of actin isoforms in ejaculated boar spermato-zoa. Gamete Res. 20:133-144.
4. Chang, K. S., K. N. Rothblum, and R. J. Schwartz. 1985. Thecomplete sequence of the chicken a-cardiac actin gene: a highlyconserved vertebrate gene. Nucleic Acids Res. 13:1223-1237.
5. Cole, K. D., and W. S. Kistler. 1987. Nuclear transition protein2 (TP2) of mammalian spermatids has a very basic carboxylterminal domain. Biochem. Biophys. Res. Commun. 147:437-442.
6. Cooper, A. D., and W. R. Crain. 1982. Complete nucleotidesequence of a sea urchin actin gene. Nucleic Acids Res.10:4081-4092.
7. DePonte-Zilli, L., A. Seiler-Tuyns, and B. M. Paterson. 1988. A40-base-pair sequence in the 3' end of the 1-actin gene regulates,-actin mRNA transcription during myogenesis. Proc. Natl.Acad. Sci. USA 85:1389-1393.
8. Distel, R. J., K. C. Kleene, and N. B. Hecht. 1984. Haploidexpression of a mouse testis a-tubulin gene. Science 224:68-70.
9. Erba, H. P., P. Gunning, and L. Kedes. 1986. Nucleotidesequence of the human -y cytoskeletal actin mRNA: anomalousevolution of vertebrate non-muscle actin genes. Nucleic AcidsRes. 14:5275-5294.
10. Files, J. G., J. Carr, and D. Hirsh. 1983. The actin gene familyof Caenorhabditis elegans. J. Mol. Biol. 164:355-375.
11. Flaherty, S. P., V. P. Winfrey, and G. E. Olson. 1986. Localiza-tion of actin in mammalian spermatozoa. Anat. Rec. 216:504-515.
12. Fornwald, J. A., G. Kunicio, I. Peng, and C. P. Ordahl. 1982.The complete nucleotide sequence of the chicken a-actin geneand its evolutionary relationship to the actin gene family.Nucleic Acids Res. 10:3861-3876.
13. Fyrberg, E. A., B. J. Bond, N. D. Hershey, K. X. Mixter, and N.Davidson. 1981. The actin genes of Drosophila: protein regionsare highly conserved but intron positions are not. Cell 24:107-116.
14. Garner, I., A. J. Minty, S. Alonso, P. J. Barton, and M.Buckingham. 1986. A 5' duplication of the a-cardiac actin genein BALB/c mice is associated with abnormal levels of a-cardiacand a-skeletal actin mRNAs in adult cardiac tissue. EMBO J.5:2559-2567.
15. Goldman, D. S., A. Kiessling, C. F. Millette, and G. M. Cooper.1987. Expression of c-mos RNA in germ cells of male andfemale mice. Proc. Natl. Acad. Sci. USA 84:4509-4513.
16. Hamada, H., M. G. Petrino, and T. Kakunaga. 1982. Molecularstructure and evolutionary origin of human cardiac muscle actingene. Proc. Natl. Acad. Sci. USA 79:5901-5905.
17. Hanauer, A., M. Levin, R. Heilig, D. Daegeln, A. Kahn, andJ. L. Mandel. 1983. Isolation and characterization of cDNAclones for human skeletal muscle a-actin. Nucleic Acids Res.
11:3503-3516.18. Hecht, N. B. 1986. Regulation of gene expression during mam-
malian spermatogenesis, p. 151-193. In J. Rossant and R.Pedersen (ed.), Experimental approaches to mammalian embry-onic development. Cambridge University Press, New York.
19. Heidaran, M. A., and W. S. Kistler. 1987. Isolation of a cDNAclone for transition protein 1 (TP1), a major chromosomalprotein of mammalian spermatids. Gene 54:281-284.
20. Henikoff, S. 1984. Unidirectional digestion with exonuclease IIIcreates targeted breakpoints for DNA sequencing. Gene 28:351-359.
21. Hu, M. C., S. B. Sharp, and N. Davidson. 1986. The completesequence of the mouse skeletal a-actin gene reveals severalconserved and inverted repeat sequences outside of the protein-coding region. Mol. Cell. Biol. 6:15-25.
22. Kistler, W. S., M. E. George, and H. G. Williams-Ashman. 1973.Specific basic proteins from mammalian testes: isolation andproperties of small basic proteins from rat testes and epididymalspermatozoa. J. Biol. Chem. 248:1847-1853.
23. Kleene, K. C., A. Bozorgzadeh, J. F. Flynn, P. C. Yelick, andN. B. Hecht. 1988. Nucleotide sequence of a cDNA encodingmouse transition protein 1. Biochim. Biophys. Acta 950:215-220.
24. Kleene, K. C., R. J. Distel, and N. B. Hecht. 1983. cDNA clonesencoding cytoplasmic poly(A)+ RNAs which first appear atdetectable levels in haploid phases of spermatogenesis in themouse. Dev. Biol. 98:455-464.
25. Kleene, K. C., R. J. Distel, and N. B. Hecht. 1984. Translationalregulation and deadenylation of a protamine mRNA duringspermiogenesis in the mouse. Dev. Biol. 105:71-79.
26. Kleene, K. C., and J. F. Flynn. 1987. Characterization of acDNA clone encoding a basic protein, TP2, involved in chro-matin condensation in the mouse. J. Biol. Chem. 262:17272-17277.
27. Kocher, O., and G. Gabbiani. 1987. Analysis of a-smooth-muscle actin mRNA expression in rat aortic smooth-musclecells using a specific cDNA probe. Differentiation 34:201-209.
28. Kocher, O., 0. Skalli, D. Cerutti, F. Gabbiani, and G. Gabbiani.1985. Cytoskeletal features of rat aortic cells during develop-ment. An electron microscopic, immunohistochemical, and bio-chemical study. Circ. Res. 56:829-838.
29. Kost, T. A., N. Theodorakis, and S. H. Hughes. 1983. Thenucleotide sequence of the chick cytoplasmic P-actin gene.Nucleic Acids Res. 11:8287-8301.
30. Leader, D. P., I. Gall, P. Campbell, and A. M. Frischauf. 1986.Isolation and characterization of cDNA clones from mouseskeletal muscle actin mRNA. DNA 5:235-238.
31. Luerssen, H., S. Hoyer-Fender, and W. Engel. 1988. The nucle-otide sequence of human transition protein 1 cDNA. NucleicAcids Res. 16:7723.
32. MeUer, D., A. Hermans, M. V. Lindern, T. V. Agthoven, A. deKlein, P. Mackenbach, A. Grootegoed, D. Talarico, G. D. Vanle,and G. Grosveld. 1987. Molecular characterization of the testisspecific c-abl mRNA in mouse. EMBO J. 6:4041-4048.
33. Meistrich, M. L., J. Longtin, W. A. Brock, S. R. Grimes, andM. L. Mace. 1981. Purification of rat spermatogenic cells andpreliminary biochemical analysis of these cells. Biol. Reprod.25:1065-1077.
34. Minty, A. J., S. Alonso, J. L. Guenet, and M. Buckingham. 1983.Number and organization of actin-related sequences in themouse genome. J. Mol. Biol. 167:77-101.
35. Nakajima-Iijima, S., H. Hamada, P. Reddy, and T. Kakunaga.1985. Molecular structure of the human cytoplasmic P-actingene: interspecies homology of sequences in the introns. Proc.Natl. Acad. Sci. USA 82:6133-6137.
36. Owens, G. K., and M. M. Thompson. 1986. Developmentalchanges in isoactin expression in rat aortic smooth muscle cellsin vivo. J. Biol. Chem. 261:13373-13380.
37. Ponzetto, C., and D. J. Wolgemuth. 1985. Haploid expression ofa unique c-abl transcript in the mouse male germ cell line. Mol.Cell. Biol. 5:1791-1794.
38. Propst, F., M. P. Rosenberg, and G. F. Vande Woude. 1988.Proto-oncogene expression in germ cell development. Trends
MOL. CELL. BIOL.

TESTICULAR SMOOTH-MUSCLE ACTIN 1881
Genet. 4:183-187.39. RomreUl, L. J., A. R. Bellve, and D. W. Fawcett. 1976. Separa-
tion of mouse spermatogenic cells by sedimentation velocity.Dev. Biol. 49:119-131.
40. Sanger, F., A. R. Coulson, B. G. Barrel, A. J. H. Smith, andB. A. Roe. 1980. Cloning in single-stranded bacteriophage as anaid to rapid DNA sequencing. J. Mol. Biol. 143:161-178.
41. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
42. Schuler, M. A., P. McOsker, and E. B. Keller. 1983. DNAsequence of two linked actin genes of sea urchin. Mol. Cell.Biol. 3:448 456.
43. Sbackleford, G. M., and H. E. Varmus. 1987. Expression of theproto-oncogene int-1 is restricted to post-meiotic male germcells and the neural tube of mid-gestational embryos. Cell50:89-95.
44. Slaughter, G. R., D. S. Needleman, and A. Means. 1987. Devel-opmental regulation of calmodulin, actin, and tubulin RNAsduring rat testis differentiation. Biol. Reprod. 37:1259-1270.
45. Tokunaga, K., H. Taniguchi, K. Yoda, M. Shimizu, and S.Sakiyama. 1986. Nucleotide sequence of a full length cDNA formouse cytoskeletal ,-actin mRNA. Nucleic Acids Res. 14:2829.
46. Tsang, A. S., H. Mabbubani, and J. G. Williams. 1982. Cell-type-specific actin mRNA populations in Dictyostelium discoi-
deum. Cell 31:375-382.47. Ueyama, H., H. Hamada, N. Battula, and T. Kakunaga. 1984.
Structure of a human smooth-muscle actin gene (aortic type)with a unique intron site. Mol. Cell. Biol. 4:1073-1078.
48. Ueyama, H., K. Kurokawa, I. Sasaki, and K. Ueda. 1987.Characterization of acidic actin in mouse sarcoma 180 cells.Cell. Struct. Funct. 12:463-470.
49. Vandekerckhove, J., and K. Weber. 1979. The complete aminoacid sequence of actins from bovine aorta, bovine heart, bovinefast skeletal muscle, and rabbit slow skeletal muscle. Differen-tiation 14:123-133.
50. Villasante, A., D. Wang, P. Dobner, P. Dolph, S. A. Lewis, andN. J. Cowan. 1986. Six mouse a-tubulin mRNAs encode fivedistinct isotypes: testis-specific expression of two sister genes.Mol. Cell. Biol. 6:2409-2419.
51. Waters, S. H., R. J. Distel, and N. B. Hecht. 1985. Mouse testiscontains two size classes of actin mRNA that are differentiallyexpressed during spermatogenesis. Mol. Cell. Biol. 5:1649-1654.
52. Welch, J. E., and M. G. O'Rand. 1985. Identification anddistribution of actin in spermatogenic cells and spermatozoa ofthe rabbit. Dev. Biol. 109:411-417.
53. Zakut, R., M. Shani, D. Givol, S. Neuman, D. Yaffe, and U.Nudel. 1982. Nucleic acid sequence of the rat skeletal muscleactin gene. Nature (London) 298:857-859.
VOL. 9, 1989