phyllocarid crustaceans from the upper devonian gogo formation, western australia
TRANSCRIPT

This article was downloaded by: [Yale University]On: 14 September 2011, At: 05:56Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Systematic PalaeontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tjsp20
Phyllocarid crustaceans from the Upper Devonian GogoFormation, Western AustraliaDerek E. G. Briggs a , W. D. Ian Rolfe b , Piers D. Butler c , Jeff J. Liston d e & J. KeithIngham e fa Department of Geology & Geophysics and Yale Peabody Museum of Natural History, YaleUniversity, New Haven, CT, 06520-8109, USAb 4A Randolph Crescent, Edinburgh EH3 7TH, UKc Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queen's Road,Bristol, BS8 1RJ, UKd Division of Environmental and Evolutionary Biology, Faculty of Biomedical and LifeSciences, University of Glasgow, G12 8QQ, UKe Hunterian Museum, University of Glasgow, G12 8QQ, UKf Department of Geographical and Earth Sciences, University of Glasgow, Glasgow, G12 8QQ,UK
Available online: 06 Sep 2011
To cite this article: Derek E. G. Briggs, W. D. Ian Rolfe, Piers D. Butler, Jeff J. Liston & J. Keith Ingham (2011): Phyllocaridcrustaceans from the Upper Devonian Gogo Formation, Western Australia, Journal of Systematic Palaeontology, 9:3, 399-424
To link to this article: http://dx.doi.org/10.1080/14772019.2010.493050
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching and private study purposes. Any substantial or systematicreproduction, re-distribution, re-selling, loan, sub-licensing, systematic supply or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Journal of Systematic Palaeontology, Vol. 9, Issue 3, September 2011, 399–424
Phyllocarid crustaceans from the Upper Devonian Gogo Formation,Western Australia
Derek E. G. Briggsa∗, W. D. Ian Rolfeb, Piers D. Butlerc, Jeff J. Listonde and J. Keith Inghamef
aDepartment of Geology & Geophysics and Yale Peabody Museum of Natural History, Yale University, New Haven, CT 06520-8109,USA; b4A Randolph Crescent, Edinburgh EH3 7TH, UK; cDepartment of Earth Sciences, University of Bristol, Wills Memorial Building,
Queen’s Road, Bristol BS8 1RJ, UK; dDivision of Environmental and Evolutionary Biology, Faculty of Biomedical and Life Sciences,University of Glasgow G12 8QQ, UK; eHunterian Museum, University of Glasgow G12 8QQ, UK; fDepartment of Geographical and
Earth Sciences, University of Glasgow, Glasgow G12 8QQ, UK
(Received 24 November 2009; accepted 13 April 2010; printed 15 September 2011)
Phyllocarids (Crustacea: Malacostraca) from the Upper Devonian (Givetian-Frasnian) Gogo Formation of Western Australiaare described and two new species are reconstructed. The mineralized exoskeleton is well preserved in concretions, but theappendages, apart from the mandible, are unknown. Montecaris gogoensis sp. nov. (Echinocarididae) is represented by >600specimens; a possible second species of Montecaris is represented only by ∼14 specimens of antero-dorsal fragments of thevalves, which are highly tuberculate. Schugurocaris wami sp. nov. (Ceratiocarididae) is represented by >300 specimens; apossible second species with a very elongate telson is represented by four specimens. An undetermined species of Dithyrocaris(Dithyrocarididae) is represented by >80 specimens.
Keywords: Montecaris; Schugurocaris; Dithyrocaris; Malacostraca
Introduction
Phyllocarids are an important subclass of Malacostraca(Rolfe 1969) distinguished by a bivalved carapace reminis-cent of that in other non-malacostracan crustaceans suchas some branchiopods and ostracodes. Living representa-tives are known as leptostracans. Phyllocarids have receivedrelatively little attention despite the importance placed onthem as a possible sister group to Eumalacostraca (Rolfe1981; Dahl 1992; but contrast Schram 1986). This is dueto their rarity and generally poor preservation (Pratt 1987;Sepkoski 2000), as many taxa probably lacked a biomineral-ized exoskeleton. Phyllocarids were particularly importantduring the Devonian: Rolfe & Edwards (1979) reviewedtheir stratigraphical distribution during this period, andRode & Lieberman (2002) their phylogeny and biogeogra-phy. The phyllocarids described here from the Upper Devo-nian Gogo Formation of Western Australia, however, areboth abundant and well preserved.
Phyllocarids, together with the enigmatic bivalvedcrustaceans known as Concavicarida (Brunton et al. 1969;Briggs & Rolfe 1983), constitute almost a quarter of fossilsfound in the Gogo Formation (Brunton et al. l969). Thisdiverse fauna also includes radiolarians (Nazarov et al.1982; Nazarov & Ormiston 1983; Won 1997a, b), sponges,brachiopods, ammonoids (Glenister 1958), a eurypterid
∗Corresponding author. Email: [email protected]
(Playford & Lowry 1966; Rolfe 1966; Tetlie et al. 2004),conodonts (Glenister & Klapper 1966; Seddon 1970;Druce 1976; Nicoll 1977; Choo et al. 2009), a shark,placoderms (arthrodires, ptyctodontids and the antiarchBothriolepis), and osteichthyes (actinopterygians,onychodontids, dipnoans and the tetrapodomorph Gogona-sus) (e.g. Gardiner & Miles 1975; Rich et al. 1985; Long1988a, b, 1995a, b, 2006; Campbell & Barwick 2002;Choo et al. 2009; Long and Trinajstic 2010; Young et al.2010). Although the occurrence of phyllocarid crustaceanshas been reported (Rolfe 1966; Brunton et al. 1969), theyhave not been described in detail previously.
Rolfe (1966) recognized two species of phyllocaridsfrom the Gogo Formation, identifying the more abun-dant provisionally as Montecaris lehmanni and the rareras Schugurocaris sp. Rolfe (1966) first considered that anadditional, rare, heavily tuberculate form of Montecarismight represent a new species, but subsequently these verylarge specimens (“about two feet long”: Brunton et al.1969, p. 81) were interpreted as older individuals of thesame species, listed as Montecaris aff. lehmanni. Bruntonet al. (1969) noted that the fauna may include an additionalceratiocaridid. Here we describe these taxa formally and,while retaining Rolfe’s (1966) generic identifications, erectnew species for the two most abundant Gogo Formationphyllocarids.
ISSN 1477-2019 print / 1478-0941 onlineCopyright C© 2011 The Natural History MuseumDOI: 10.1080/14772019.2010.493050http://www.informaworld.com
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

400 D. E. G. Briggs et al.
Geological setting
The Gogo Formation was deposited within and around theSadler Limestone, the ‘Great Devonian Barrier Reef’ of thenorth-eastern Canning Basin of Western Australia (Play-ford 1980; Playford et al. 1989, 2009). The Formationcomprises mainly dark grey to black shales and siltstones,with some limestone lenses and bedded silty micritic lime-stone concretions (Playford & Lowry 1966; Long 1988a)that yield the well known Gogo fossils (Long 2006; Long& Trinajstic 2010). It is the basinal equivalent of the SadlerLimestone Formation marginal-slope facies, and the PillaraLimestone platform facies. The Gogo Formation is datedas Late Givetian to early Frasnian, based on conodontsand radiolarians (Glenister & Klapper 1966; Seddon 1970;Druce 1976; Nazarov et al. 1982; Nazarov & Ormiston1983; Playford et al. 2009).
Concretions occur at several levels through the shale(Campbell & Barwick 1999). However, they were collectedfrom soil, some in shallow valleys to which theyhad been transported. Thus the precise location of theconcretions is of limited value in interpreting taxonassociations.
Material and methods
The first major collection of Gogo fossils was made in1963 by an expedition led by H. A. Toombs of the NaturalHistory Museum, London in conjunction with the West-ern Australian Museum (Rolfe 1966; Long 2006). Thecollection yielded mainly phyllocarids and some fishes.A further expedition in 1967 yielded much more materialfrom ∼100 localities (Miles 1971, fig. 1), which forms themain basis of this study. That joint expedition was mountedby the following museums (abbreviations used in cataloguenumbers in parentheses): Natural History Museum, London(NHM), the Western Australian Museum, Perth (WAM),and the Hunterian Museum, Glasgow (GLAHM) (Bruntonet al. 1969). The specimens are divided between these threeinstitutions and the National Museum of Natural History,Washington, DC (USNM). Unlike the fishes, the phyllo-carids cannot be acid prepared. The most successful methodproved to be bold splitting of the concretions using hammerand chisels, followed by excavation of unexposed surfacesusing a vibrotool. Specimens collected prior to 1963, held inadditional repositories recorded in Briggs & Rolfe (1983),have also been studied. Historical background to the Gogodiscoveries, together with accounts of the main collectingexpeditions up to 2001 and the scientific significance of thefishes, is given by Long (2006). Terminology follows thatof Rolfe (1969, 1981).
Preservation
A sample of 1442 phyllocarid specimens representingthe entire 1967 Gogo collection was surveyed to providean indication of completeness of individual specimens(the other bivalved arthropods, the concavicarids, weretreated by Briggs & Rolfe 1983): 149 (10%) are unde-termined fragments, 208 (14%) are mandibles and 32 (2%)tails; 935 (65%) are isolated elements (574, Montecarisgogoensis; 301, Schugurocaris; 60, Dithyrocaris)—singlevalves, abdominal segments, telson, or furca; 110 (8%)are partial combinations (70, Montecaris gogoensis; 16,Schugurocaris; 24, Dithyrocaris), probably the result ofmoulting—mainly the joined valves, the exposed poste-rior segments of the abdomen with the telson and furca,or just the telson and furca; and eight (<1%) specimensof Montecaris show a complete carapace and abdominalregion (Figs 1, 2). Trunk limbs are not preserved on anyspecimens. Isolated mandibles are common in the GogoFormation and at some other localities worldwide (Dzik1980), but several examples of concretions containing justthe opposed mandibles of a pair, in approximately theirposition in life, are an unusual feature of this formation(Fig. 5). Such examples are presumably a product of amuch higher decay resistance of the mandibles comparedto other appendages, reflecting strengthening to equip themto process hard food.
Concretions containing isolated body parts tend to splitaround the fossil; more complete individuals tend tobreak along the different surfaces of the constituent parts.Complete carapaces have collapsed, presumably after decayof the adductor muscles, and the valves lie parallel to thebedding plane (Figs 1, 2). Single valves, complete cara-paces, and tails (telson and furca) normally lie nearly paral-lel to bedding, as do abdominal segments that bear spines.Isolated mandibles and abdominal segments without spinesdo not show a preferred orientation. Carapaces, in particu-lar, often display fractures resulting from compaction (Figs1, 2). This indicates that the cuticle was brittle and mayhave been biomineralized.
The cuticle of the Gogo phyllocarids is white to grey incolour, in striking contrast to the pale brown matrix of theconcretions. It is often both over- and underlain by a layer ofcrystal bundles of calcite. Structures similar to the crystalbundles have been reported in some Gogo fishes (Campbell& Barwick 1988, 1999). The cuticle composition (analysedby XRD) is dominated by calcite, with variable amountsof apatite, the latter probably a diagenetic introduction.Silicified phyllocarids are rare: Fig. 22B illustrates anisolated Dithyrocaris sp. tail (NHM IC 499) that emergedduring acid extraction of a left posterior ventrolateralplate of the Gogo fish Holonema from locality 42 (NHMP.50987, Miles 1971, p. 170; Gardiner & Miles 1975; Long1995b, previously incorrectly reported as from locality 67,which does not exist).
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
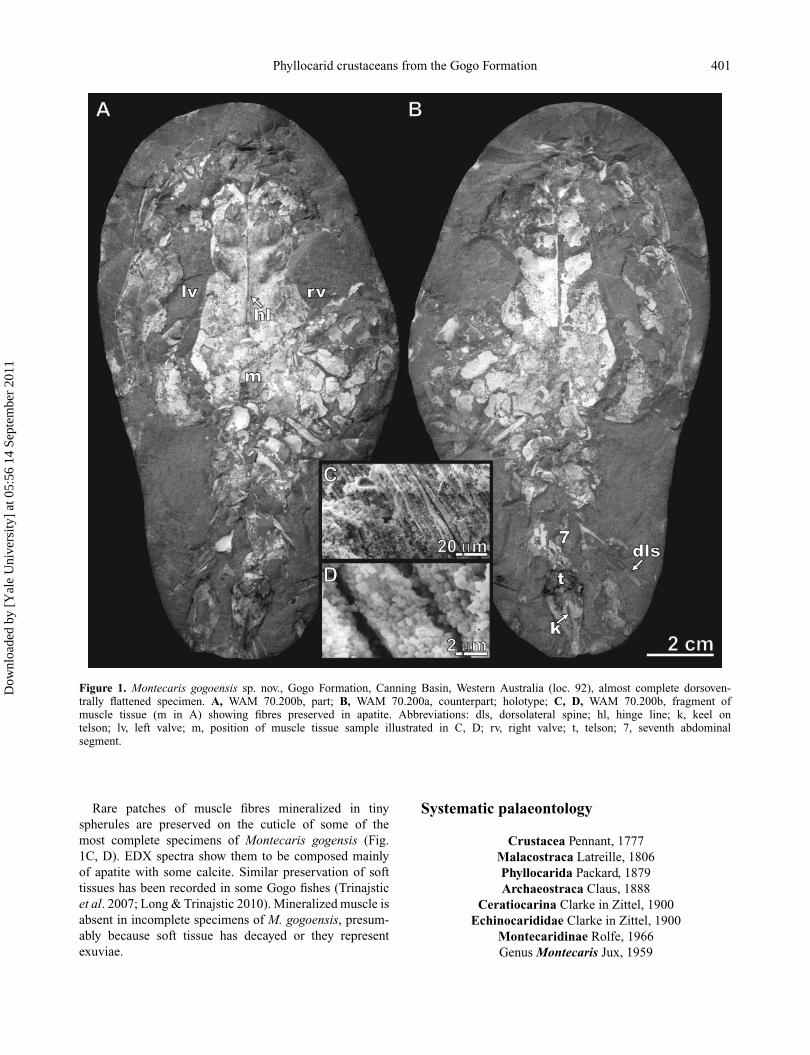
Phyllocarid crustaceans from the Gogo Formation 401
Figure 1. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia (loc. 92), almost complete dorsoven-trally flattened specimen. A, WAM 70.200b, part; B, WAM 70.200a, counterpart; holotype; C, D, WAM 70.200b, fragment ofmuscle tissue (m in A) showing fibres preserved in apatite. Abbreviations: dls, dorsolateral spine; hl, hinge line; k, keel ontelson; lv, left valve; m, position of muscle tissue sample illustrated in C, D; rv, right valve; t, telson; 7, seventh abdominalsegment.
Rare patches of muscle fibres mineralized in tinyspherules are preserved on the cuticle of some of themost complete specimens of Montecaris gogensis (Fig.1C, D). EDX spectra show them to be composed mainlyof apatite with some calcite. Similar preservation of softtissues has been recorded in some Gogo fishes (Trinajsticet al. 2007; Long & Trinajstic 2010). Mineralized muscle isabsent in incomplete specimens of M. gogoensis, presum-ably because soft tissue has decayed or they representexuviae.
Systematic palaeontology
Crustacea Pennant, 1777Malacostraca Latreille, 1806Phyllocarida Packard, 1879Archaeostraca Claus, 1888
Ceratiocarina Clarke in Zittel, 1900Echinocarididae Clarke in Zittel, 1900
Montecaridinae Rolfe, 1966Genus Montecaris Jux, 1959
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
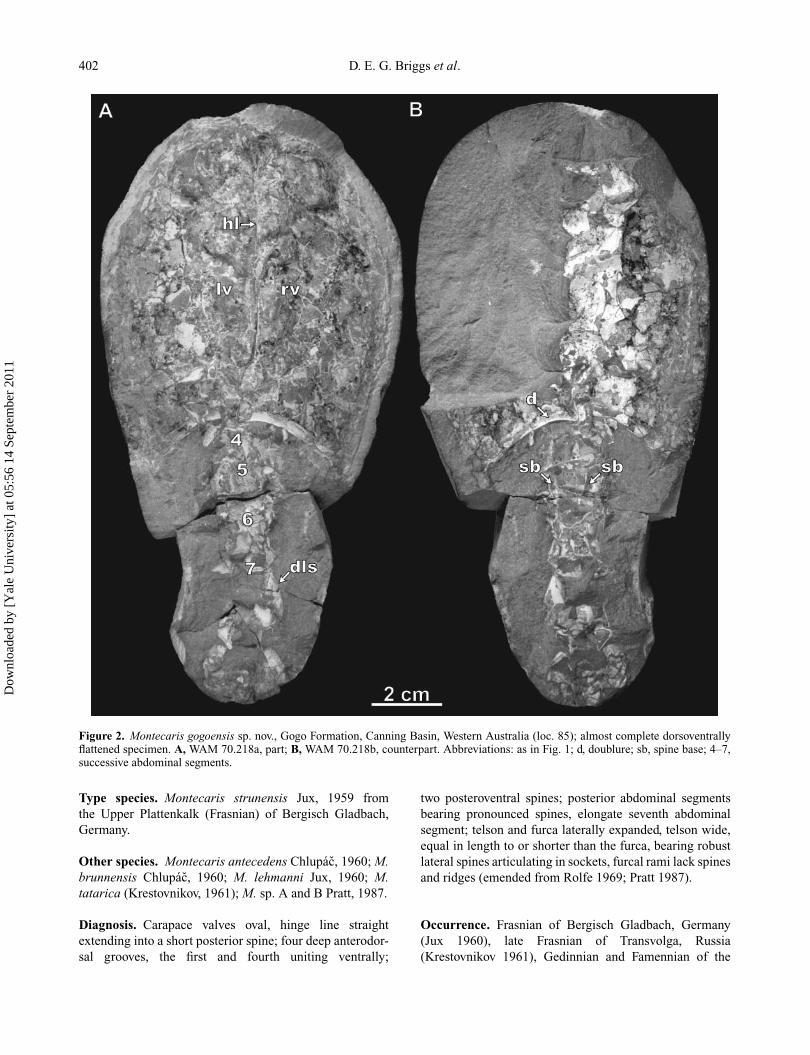
402 D. E. G. Briggs et al.
Figure 2. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia (loc. 85); almost complete dorsoventrallyflattened specimen. A, WAM 70.218a, part; B, WAM 70.218b, counterpart. Abbreviations: as in Fig. 1; d, doublure; sb, spine base; 4–7,successive abdominal segments.
Type species. Montecaris strunensis Jux, 1959 fromthe Upper Plattenkalk (Frasnian) of Bergisch Gladbach,Germany.
Other species. Montecaris antecedens Chlupac, 1960; M.brunnensis Chlupac, 1960; M. lehmanni Jux, 1960; M.tatarica (Krestovnikov, 1961); M. sp. A and B Pratt, 1987.
Diagnosis. Carapace valves oval, hinge line straightextending into a short posterior spine; four deep anterodor-sal grooves, the first and fourth uniting ventrally;
two posteroventral spines; posterior abdominal segmentsbearing pronounced spines, elongate seventh abdominalsegment; telson and furca laterally expanded, telson wide,equal in length to or shorter than the furca, bearing robustlateral spines articulating in sockets, furcal rami lack spinesand ridges (emended from Rolfe 1969; Pratt 1987).
Occurrence. Frasnian of Bergisch Gladbach, Germany(Jux 1960), late Frasnian of Transvolga, Russia(Krestovnikov 1961), Gedinnian and Famennian of the
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
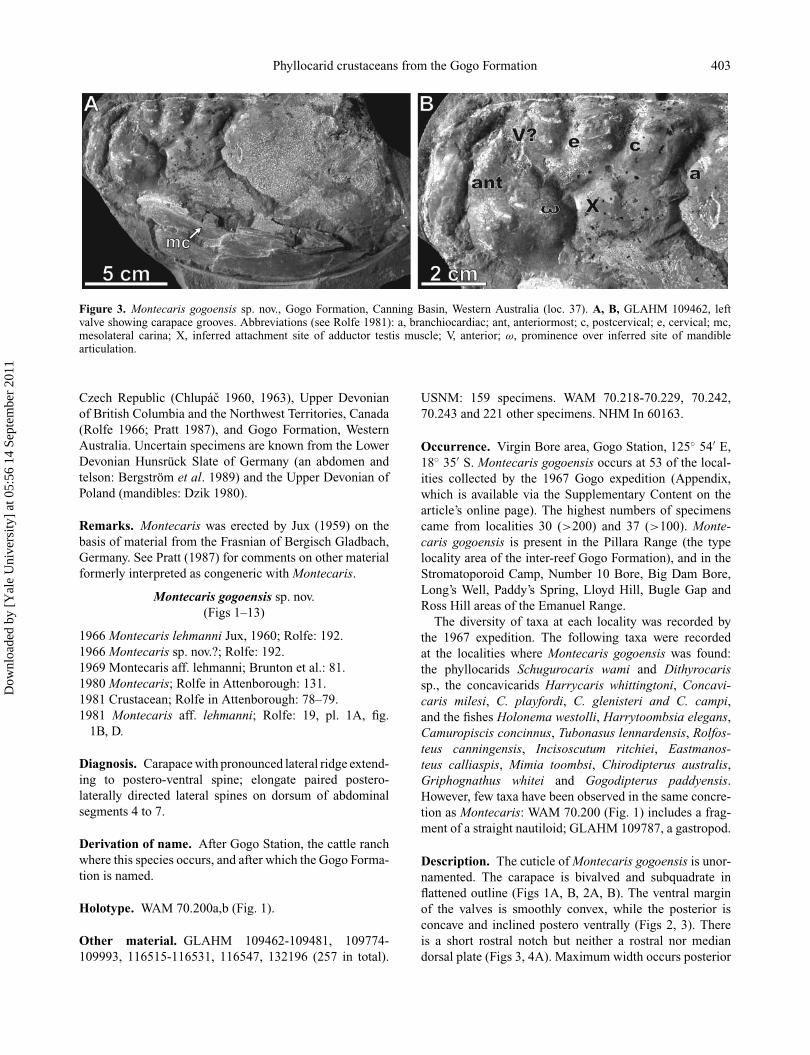
Phyllocarid crustaceans from the Gogo Formation 403
Figure 3. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia (loc. 37). A, B, GLAHM 109462, leftvalve showing carapace grooves. Abbreviations (see Rolfe 1981): a, branchiocardiac; ant, anteriormost; c, postcervical; e, cervical; mc,mesolateral carina; X, inferred attachment site of adductor testis muscle; V, anterior; ω, prominence over inferred site of mandiblearticulation.
Czech Republic (Chlupac 1960, 1963), Upper Devonianof British Columbia and the Northwest Territories, Canada(Rolfe 1966; Pratt 1987), and Gogo Formation, WesternAustralia. Uncertain specimens are known from the LowerDevonian Hunsruck Slate of Germany (an abdomen andtelson: Bergstrom et al. 1989) and the Upper Devonian ofPoland (mandibles: Dzik 1980).
Remarks. Montecaris was erected by Jux (1959) on thebasis of material from the Frasnian of Bergisch Gladbach,Germany. See Pratt (1987) for comments on other materialformerly interpreted as congeneric with Montecaris.
Montecaris gogoensis sp. nov.(Figs 1–13)
1966 Montecaris lehmanni Jux, 1960; Rolfe: 192.1966 Montecaris sp. nov.?; Rolfe: 192.1969 Montecaris aff. lehmanni; Brunton et al.: 81.1980 Montecaris; Rolfe in Attenborough: 131.1981 Crustacean; Rolfe in Attenborough: 78–79.1981 Montecaris aff. lehmanni; Rolfe: 19, pl. 1A, fig.
1B, D.
Diagnosis. Carapace with pronounced lateral ridge extend-ing to postero-ventral spine; elongate paired postero-laterally directed lateral spines on dorsum of abdominalsegments 4 to 7.
Derivation of name. After Gogo Station, the cattle ranchwhere this species occurs, and after which the Gogo Forma-tion is named.
Holotype. WAM 70.200a,b (Fig. 1).
Other material. GLAHM 109462-109481, 109774-109993, 116515-116531, 116547, 132196 (257 in total).
USNM: 159 specimens. WAM 70.218-70.229, 70.242,70.243 and 221 other specimens. NHM In 60163.
Occurrence. Virgin Bore area, Gogo Station, 125◦ 54′ E,18◦ 35′ S. Montecaris gogoensis occurs at 53 of the local-ities collected by the 1967 Gogo expedition (Appendix,which is available via the Supplementary Content on thearticle’s online page). The highest numbers of specimenscame from localities 30 (>200) and 37 (>100). Monte-caris gogoensis is present in the Pillara Range (the typelocality area of the inter-reef Gogo Formation), and in theStromatoporoid Camp, Number 10 Bore, Big Dam Bore,Long’s Well, Paddy’s Spring, Lloyd Hill, Bugle Gap andRoss Hill areas of the Emanuel Range.
The diversity of taxa at each locality was recorded bythe 1967 expedition. The following taxa were recordedat the localities where Montecaris gogoensis was found:the phyllocarids Schugurocaris wami and Dithyrocarissp., the concavicarids Harrycaris whittingtoni, Concavi-caris milesi, C. playfordi, C. glenisteri and C. campi,and the fishes Holonema westolli, Harrytoombsia elegans,Camuropiscis concinnus, Tubonasus lennardensis, Rolfos-teus canningensis, Incisoscutum ritchiei, Eastmanos-teus calliaspis, Mimia toombsi, Chirodipterus australis,Griphognathus whitei and Gogodipterus paddyensis.However, few taxa have been observed in the same concre-tion as Montecaris: WAM 70.200 (Fig. 1) includes a frag-ment of a straight nautiloid; GLAHM 109787, a gastropod.
Description. The cuticle of Montecaris gogoensis is unor-namented. The carapace is bivalved and subquadrate inflattened outline (Figs 1A, B, 2A, B). The ventral marginof the valves is smoothly convex, while the posterior isconcave and inclined postero ventrally (Figs 2, 3). Thereis a short rostral notch but neither a rostral nor mediandorsal plate (Figs 3, 4A). Maximum width occurs posterior
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
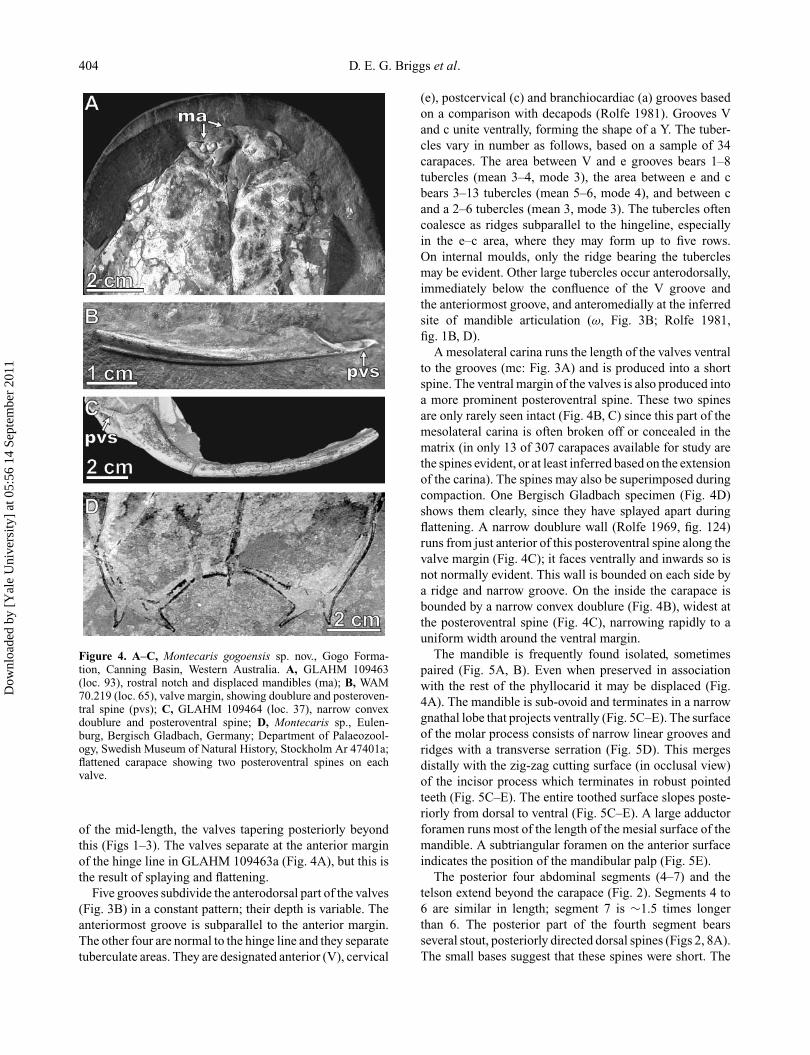
404 D. E. G. Briggs et al.
Figure 4. A–C, Montecaris gogoensis sp. nov., Gogo Forma-tion, Canning Basin, Western Australia. A, GLAHM 109463(loc. 93), rostral notch and displaced mandibles (ma); B, WAM70.219 (loc. 65), valve margin, showing doublure and posteroven-tral spine (pvs); C, GLAHM 109464 (loc. 37), narrow convexdoublure and posteroventral spine; D, Montecaris sp., Eulen-burg, Bergisch Gladbach, Germany; Department of Palaeozool-ogy, Swedish Museum of Natural History, Stockholm Ar 47401a;flattened carapace showing two posteroventral spines on eachvalve.
of the mid-length, the valves tapering posteriorly beyondthis (Figs 1–3). The valves separate at the anterior marginof the hinge line in GLAHM 109463a (Fig. 4A), but this isthe result of splaying and flattening.
Five grooves subdivide the anterodorsal part of the valves(Fig. 3B) in a constant pattern; their depth is variable. Theanteriormost groove is subparallel to the anterior margin.The other four are normal to the hinge line and they separatetuberculate areas. They are designated anterior (V), cervical
(e), postcervical (c) and branchiocardiac (a) grooves basedon a comparison with decapods (Rolfe 1981). Grooves Vand c unite ventrally, forming the shape of a Y. The tuber-cles vary in number as follows, based on a sample of 34carapaces. The area between V and e grooves bears 1–8tubercles (mean 3–4, mode 3), the area between e and cbears 3–13 tubercles (mean 5–6, mode 4), and between cand a 2–6 tubercles (mean 3, mode 3). The tubercles oftencoalesce as ridges subparallel to the hingeline, especiallyin the e–c area, where they may form up to five rows.On internal moulds, only the ridge bearing the tuberclesmay be evident. Other large tubercles occur anterodorsally,immediately below the confluence of the V groove andthe anteriormost groove, and anteromedially at the inferredsite of mandible articulation (ω, Fig. 3B; Rolfe 1981,fig. 1B, D).
A mesolateral carina runs the length of the valves ventralto the grooves (mc: Fig. 3A) and is produced into a shortspine. The ventral margin of the valves is also produced intoa more prominent posteroventral spine. These two spinesare only rarely seen intact (Fig. 4B, C) since this part of themesolateral carina is often broken off or concealed in thematrix (in only 13 of 307 carapaces available for study arethe spines evident, or at least inferred based on the extensionof the carina). The spines may also be superimposed duringcompaction. One Bergisch Gladbach specimen (Fig. 4D)shows them clearly, since they have splayed apart duringflattening. A narrow doublure wall (Rolfe 1969, fig. 124)runs from just anterior of this posteroventral spine along thevalve margin (Fig. 4C); it faces ventrally and inwards so isnot normally evident. This wall is bounded on each side bya ridge and narrow groove. On the inside the carapace isbounded by a narrow convex doublure (Fig. 4B), widest atthe posteroventral spine (Fig. 4C), narrowing rapidly to auniform width around the ventral margin.
The mandible is frequently found isolated, sometimespaired (Fig. 5A, B). Even when preserved in associationwith the rest of the phyllocarid it may be displaced (Fig.4A). The mandible is sub-ovoid and terminates in a narrowgnathal lobe that projects ventrally (Fig. 5C–E). The surfaceof the molar process consists of narrow linear grooves andridges with a transverse serration (Fig. 5D). This mergesdistally with the zig-zag cutting surface (in occlusal view)of the incisor process which terminates in robust pointedteeth (Fig. 5C–E). The entire toothed surface slopes poste-riorly from dorsal to ventral (Fig. 5C–E). A large adductorforamen runs most of the length of the mesial surface of themandible. A subtriangular foramen on the anterior surfaceindicates the position of the mandibular palp (Fig. 5E).
The posterior four abdominal segments (4–7) and thetelson extend beyond the carapace (Fig. 2). Segments 4 to6 are similar in length; segment 7 is ∼1.5 times longerthan 6. The posterior part of the fourth segment bearsseveral stout, posteriorly directed dorsal spines (Figs 2, 8A).The small bases suggest that these spines were short. The
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
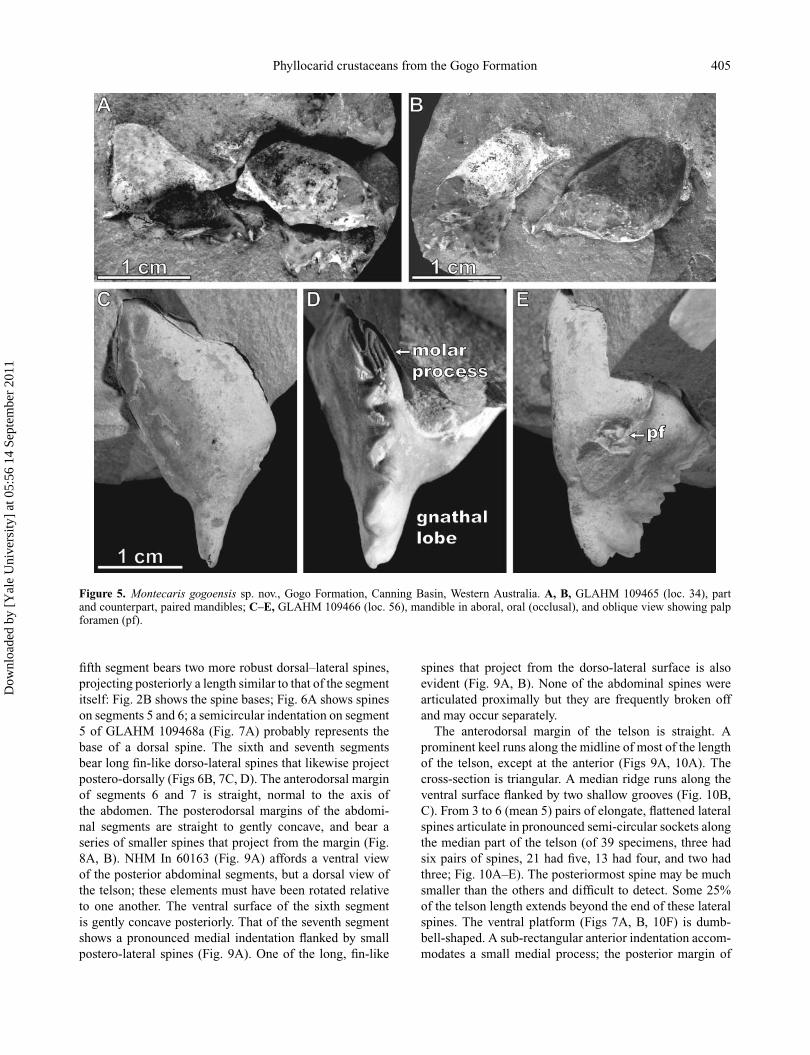
Phyllocarid crustaceans from the Gogo Formation 405
Figure 5. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia. A, B, GLAHM 109465 (loc. 34), partand counterpart, paired mandibles; C–E, GLAHM 109466 (loc. 56), mandible in aboral, oral (occlusal), and oblique view showing palpforamen (pf).
fifth segment bears two more robust dorsal–lateral spines,projecting posteriorly a length similar to that of the segmentitself: Fig. 2B shows the spine bases; Fig. 6A shows spineson segments 5 and 6; a semicircular indentation on segment5 of GLAHM 109468a (Fig. 7A) probably represents thebase of a dorsal spine. The sixth and seventh segmentsbear long fin-like dorso-lateral spines that likewise projectpostero-dorsally (Figs 6B, 7C, D). The anterodorsal marginof segments 6 and 7 is straight, normal to the axis ofthe abdomen. The posterodorsal margins of the abdomi-nal segments are straight to gently concave, and bear aseries of smaller spines that project from the margin (Fig.8A, B). NHM In 60163 (Fig. 9A) affords a ventral viewof the posterior abdominal segments, but a dorsal view ofthe telson; these elements must have been rotated relativeto one another. The ventral surface of the sixth segmentis gently concave posteriorly. That of the seventh segmentshows a pronounced medial indentation flanked by smallpostero-lateral spines (Fig. 9A). One of the long, fin-like
spines that project from the dorso-lateral surface is alsoevident (Fig. 9A, B). None of the abdominal spines werearticulated proximally but they are frequently broken offand may occur separately.
The anterodorsal margin of the telson is straight. Aprominent keel runs along the midline of most of the lengthof the telson, except at the anterior (Figs 9A, 10A). Thecross-section is triangular. A median ridge runs along theventral surface flanked by two shallow grooves (Fig. 10B,C). From 3 to 6 (mean 5) pairs of elongate, flattened lateralspines articulate in pronounced semi-circular sockets alongthe median part of the telson (of 39 specimens, three hadsix pairs of spines, 21 had five, 13 had four, and two hadthree; Fig. 10A–E). The posteriormost spine may be muchsmaller than the others and difficult to detect. Some 25%of the telson length extends beyond the end of these lateralspines. The ventral platform (Figs 7A, B, 10F) is dumb-bell-shaped. A sub-rectangular anterior indentation accom-modates a small medial process; the posterior margin of
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
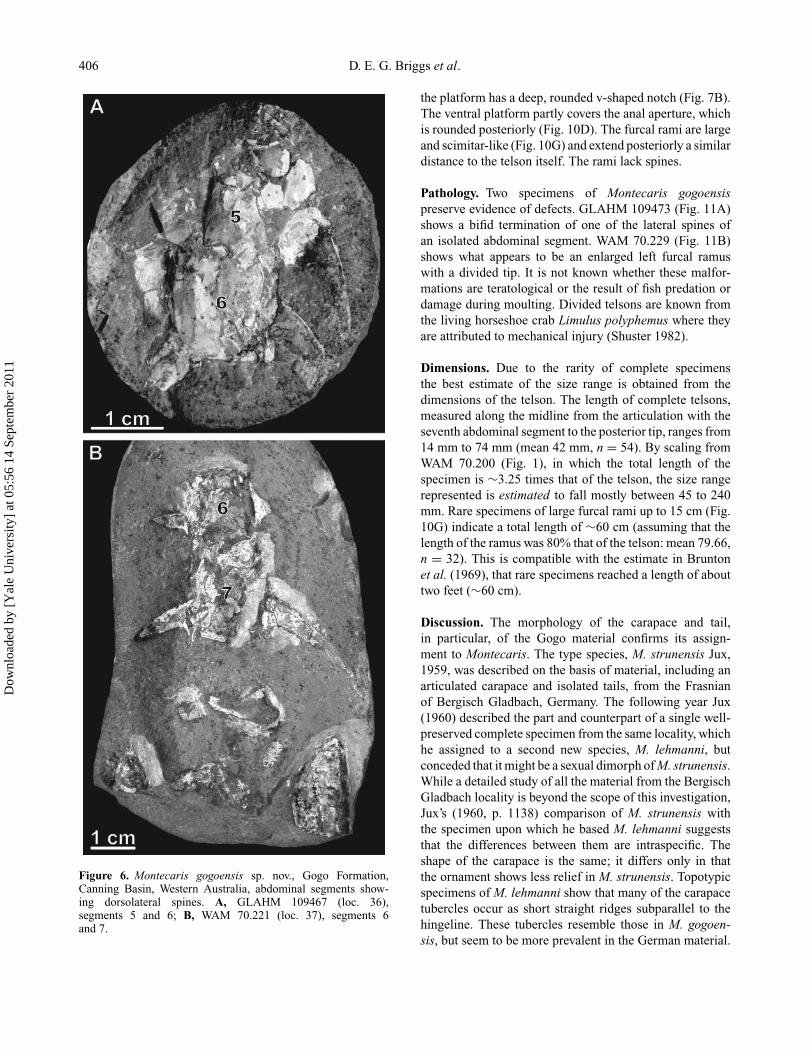
406 D. E. G. Briggs et al.
Figure 6. Montecaris gogoensis sp. nov., Gogo Formation,Canning Basin, Western Australia, abdominal segments show-ing dorsolateral spines. A, GLAHM 109467 (loc. 36),segments 5 and 6; B, WAM 70.221 (loc. 37), segments 6and 7.
the platform has a deep, rounded v-shaped notch (Fig. 7B).The ventral platform partly covers the anal aperture, whichis rounded posteriorly (Fig. 10D). The furcal rami are largeand scimitar-like (Fig. 10G) and extend posteriorly a similardistance to the telson itself. The rami lack spines.
Pathology. Two specimens of Montecaris gogoensispreserve evidence of defects. GLAHM 109473 (Fig. 11A)shows a bifid termination of one of the lateral spines ofan isolated abdominal segment. WAM 70.229 (Fig. 11B)shows what appears to be an enlarged left furcal ramuswith a divided tip. It is not known whether these malfor-mations are teratological or the result of fish predation ordamage during moulting. Divided telsons are known fromthe living horseshoe crab Limulus polyphemus where theyare attributed to mechanical injury (Shuster 1982).
Dimensions. Due to the rarity of complete specimensthe best estimate of the size range is obtained from thedimensions of the telson. The length of complete telsons,measured along the midline from the articulation with theseventh abdominal segment to the posterior tip, ranges from14 mm to 74 mm (mean 42 mm, n = 54). By scaling fromWAM 70.200 (Fig. 1), in which the total length of thespecimen is ∼3.25 times that of the telson, the size rangerepresented is estimated to fall mostly between 45 to 240mm. Rare specimens of large furcal rami up to 15 cm (Fig.10G) indicate a total length of ∼60 cm (assuming that thelength of the ramus was 80% that of the telson: mean 79.66,n = 32). This is compatible with the estimate in Bruntonet al. (1969), that rare specimens reached a length of abouttwo feet (∼60 cm).
Discussion. The morphology of the carapace and tail,in particular, of the Gogo material confirms its assign-ment to Montecaris. The type species, M. strunensis Jux,1959, was described on the basis of material, including anarticulated carapace and isolated tails, from the Frasnianof Bergisch Gladbach, Germany. The following year Jux(1960) described the part and counterpart of a single well-preserved complete specimen from the same locality, whichhe assigned to a second new species, M. lehmanni, butconceded that it might be a sexual dimorph of M. strunensis.While a detailed study of all the material from the BergischGladbach locality is beyond the scope of this investigation,Jux’s (1960, p. 1138) comparison of M. strunensis withthe specimen upon which he based M. lehmanni suggeststhat the differences between them are intraspecific. Theshape of the carapace is the same; it differs only in thatthe ornament shows less relief in M. strunensis. Topotypicspecimens of M. lehmanni show that many of the carapacetubercles occur as short straight ridges subparallel to thehingeline. These tubercles resemble those in M. gogoen-sis, but seem to be more prevalent in the German material.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
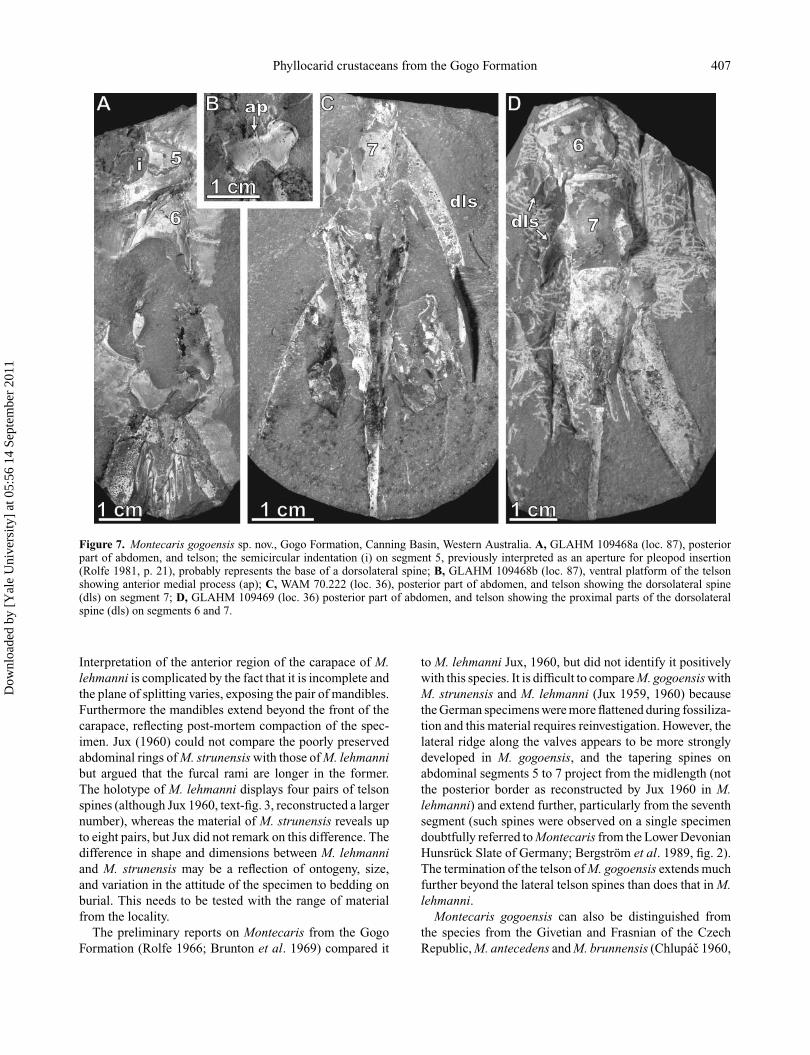
Phyllocarid crustaceans from the Gogo Formation 407
Figure 7. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia. A, GLAHM 109468a (loc. 87), posteriorpart of abdomen, and telson; the semicircular indentation (i) on segment 5, previously interpreted as an aperture for pleopod insertion(Rolfe 1981, p. 21), probably represents the base of a dorsolateral spine; B, GLAHM 109468b (loc. 87), ventral platform of the telsonshowing anterior medial process (ap); C, WAM 70.222 (loc. 36), posterior part of abdomen, and telson showing the dorsolateral spine(dls) on segment 7; D, GLAHM 109469 (loc. 36) posterior part of abdomen, and telson showing the proximal parts of the dorsolateralspine (dls) on segments 6 and 7.
Interpretation of the anterior region of the carapace of M.lehmanni is complicated by the fact that it is incomplete andthe plane of splitting varies, exposing the pair of mandibles.Furthermore the mandibles extend beyond the front of thecarapace, reflecting post-mortem compaction of the spec-imen. Jux (1960) could not compare the poorly preservedabdominal rings of M. strunensis with those of M. lehmannibut argued that the furcal rami are longer in the former.The holotype of M. lehmanni displays four pairs of telsonspines (although Jux 1960, text-fig. 3, reconstructed a largernumber), whereas the material of M. strunensis reveals upto eight pairs, but Jux did not remark on this difference. Thedifference in shape and dimensions between M. lehmanniand M. strunensis may be a reflection of ontogeny, size,and variation in the attitude of the specimen to bedding onburial. This needs to be tested with the range of materialfrom the locality.
The preliminary reports on Montecaris from the GogoFormation (Rolfe 1966; Brunton et al. 1969) compared it
to M. lehmanni Jux, 1960, but did not identify it positivelywith this species. It is difficult to compare M. gogoensis withM. strunensis and M. lehmanni (Jux 1959, 1960) becausethe German specimens were more flattened during fossiliza-tion and this material requires reinvestigation. However, thelateral ridge along the valves appears to be more stronglydeveloped in M. gogoensis, and the tapering spines onabdominal segments 5 to 7 project from the midlength (notthe posterior border as reconstructed by Jux 1960 in M.lehmanni) and extend further, particularly from the seventhsegment (such spines were observed on a single specimendoubtfully referred to Montecaris from the Lower DevonianHunsruck Slate of Germany; Bergstrom et al. 1989, fig. 2).The termination of the telson of M. gogoensis extends muchfurther beyond the lateral telson spines than does that in M.lehmanni.
Montecaris gogoensis can also be distinguished fromthe species from the Givetian and Frasnian of the CzechRepublic, M. antecedens and M. brunnensis (Chlupac 1960,
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

408 D. E. G. Briggs et al.
Figure 8. Montecaris gogoensis sp. nov., Gogo Formation,Canning Basin, Western Australia (loc. 37), abdominal segmentsshowing spines (s) along posterodorsal margin. A, WAM 70.223;B, GLAHM 109470a.
1963). Montecaris antecedens is known only from thetail, so the comparison is incomplete. However, the telsonof M. antecedens, unlike that of M. gogoensis, lacks a
median ridge, and both telson and the furcal rami displaya strong sculpture. Montecaris brunnensis is much morecompletely known. The carapace lacks the anterior nodesand lateral ridge so characteristic of M. gogoensis. However,as Chlupac (1963, p. 116) pointed out, “a comparison of thelocation of nodes in the antero-dorsal part cannot be madeowing to the flattening” of the material from the CzechRepublic. This likely reflects taphonomic conditions, butmay indicate that the cuticle was thinner. The spines on theposterior margin of the abdominal segments of M. brun-nensis are much less pronounced than in M. gogoensis, andlateral abdominal spines are absent. The sixth abdominalsegment is clearly longer than those anterior to it. The furcalrami in M. brunnensis extend much further posteriorly thanthe telson, which lacks a median ridge, whereas they extenda similar distance in M. gogoensis.
M. gogoensis also differs from M. tatarica (Krestovnikov,1961), from the Transvolga of Russia. The carapace of M.tatarica lacks well developed grooves and nodes anteri-orly, and the postero-ventral spine is not pronounced. Thereare no lateral spines on the abdomen. The furcal rami arestrongly curved and longer than the telson.
Montecaris gogoensis is clearly different from all otherdescribed species of the genus, justifying its assignmentto a new species. The reconstruction (Fig. 13) adds somedetail to that in Attenborough (1980, p. 131; 1981, pp.78–79), which was based on knowledge of the crustaceanavailable at the time. The pre-telson abdomen, for example,was depicted upside-down based on NHM In 60163 (Fig.9A)! The present more comprehensive study has shownthat the abdomen was rotated ∼180◦ relative to the telsonin this specimen (as reported by Bergstrom et al. 1989, p.325).
The generalized mandible of M. gogoensis is quite differ-ent from that referred to this genus by Dzik (1980), whichmust therefore belong to another taxon.
The discovery of the elongate spines that project fromthe midlength of the abdominal segments on M. gogoensissupports its assignment to Echinocarididae Clarke in Zittel,1900. Such a placement is consistent with cladistic analy-ses by Rode & Lieberman (2002) and Briggs et al. (2003).Similar spines occur in the genus Pephricaris Clarke, 1898(Pephricarididae Van Straelen, 1933) which, along withthe enigmatic Ohiocaris Rolfe, 1962 (Ohiocarididae Rolfe,1962), may also belong to the Echinocarididae Clarke inZittel, 1900 (see Feldmann et al. 2004; Hannibal 2009)rather than a separate higher taxon (Rode & Lieberman2002; Briggs et al. 2003).
Montecaris sp. indet.(Fig. 14)
1966 Montecaris sp. nov.? Rolfe: 192.1969 Montecaris aff. lehmanni; Brunton et al.: 81, partim.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
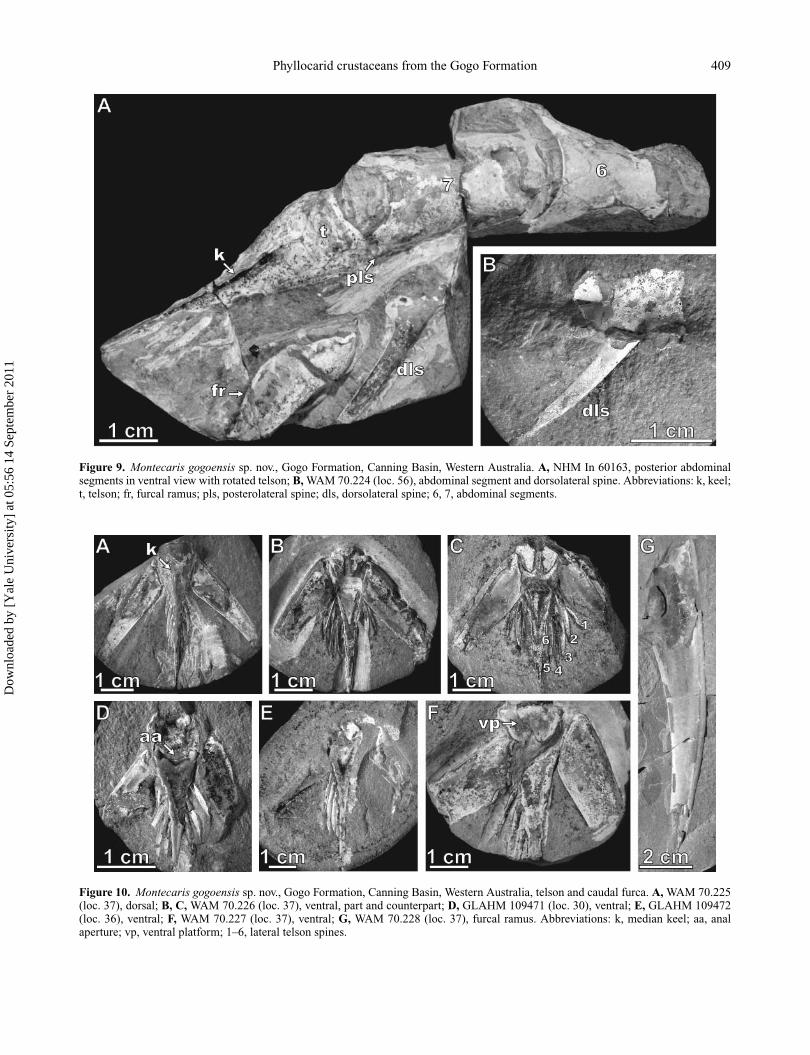
Phyllocarid crustaceans from the Gogo Formation 409
Figure 9. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia. A, NHM In 60163, posterior abdominalsegments in ventral view with rotated telson; B, WAM 70.224 (loc. 56), abdominal segment and dorsolateral spine. Abbreviations: k, keel;t, telson; fr, furcal ramus; pls, posterolateral spine; dls, dorsolateral spine; 6, 7, abdominal segments.
Figure 10. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia, telson and caudal furca. A, WAM 70.225(loc. 37), dorsal; B, C, WAM 70.226 (loc. 37), ventral, part and counterpart; D, GLAHM 109471 (loc. 30), ventral; E, GLAHM 109472(loc. 36), ventral; F, WAM 70.227 (loc. 37), ventral; G, WAM 70.228 (loc. 37), furcal ramus. Abbreviations: k, median keel; aa, analaperture; vp, ventral platform; 1–6, lateral telson spines.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

410 D. E. G. Briggs et al.
Figure 11. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia, pathological specimens. A, GLAHM109473 (loc. 23), bifid termination of a dorsolateral spine (dls) on an abdominal segment; B, WAM 70.229 (loc. 37), left furcal ramuswith divided tip.
Figure 12. Montecaris gogoensis sp. nov., Gogo Formation,Canning Basin, Western Australia. Anterodorsal valve fragments,resulting from predator attacks. A, WAM 70.242 (loc. 37); B,WAM 70.243 (loc. 30).
Material. GLAHM 109476–109480; USNM 527376–527377; WAM 70.203, 70.205, 70.206, 70.207,70.208–70.210.
Occurrence. Virgin Bore area, Gogo Station, 125◦ 54′
E, 18◦ 35′ S. Montecaris sp. indet. occurs at four ofthe localities collected by the 1967 Gogo expedition(Appendix), most specimens coming from locality 37,which also yields the second highest number of Montecarisgogoensis. Montecaris sp. indet. is present in the Stromato-poroid Camp and Number 10 Bore areas.
Description. This form is known only from anterodorsalfragments of the carapace. The carapace is distinguishedby the presence of many tubercles, from ∼0.5 to 2 mm indiameter at the base, concentrated in a 1 cm band that runsadjacent to and parallel to the hinge line from a point nearits anterior extremity (Fig. 14A, B). The tubercles are vari-ably developed, both in number (up to ∼90 – much greaterthan the ∼15 of M. gogoensis) and arrangement, from spec-imen to specimen. It is not known whether they differ onthe opposing valves of a pair. In addition to the presenceof the tubercles, this type of Montecaris carapace differs inthe presence of a more pronounced lateral projection (‘cara-pace horn’) at the anterior extremity of the hinge line whichreaches up to 5 mm across and has the appearance of a largetubercle flattened parallel to the hinge (Fig. 14A). Behind
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

Phyllocarid crustaceans from the Gogo Formation 411
Figure 13. Montecaris gogoensis sp. nov., Gogo Formation, Canning Basin, Western Australia, reconstruction (drawn by J. K. Ingham).
this tubercle lies a clearly developed anteriormost groove,which runs parallel to the anterior margin of the valve, likethat in M. gogoensis. The other grooves on the carapace ofM. gogoensis, except for the posteriormost (Fig. 3B), whichmay define the extent of these fragments, are not evident orare only faintly discernible.
Dimensions. The valve fragments reach lengths up to 120mm (WAM 70.203a) and 123 mm (WAM 70.205a) (notfigured). If the grooves on WAM 70.203a are homologouswith the anterior and posteriormost grooves in M. gogoen-sis (Fig. 3), they indicate a carapace length of ∼11 cm,
Figure 14. Montecaris sp. indet., anterodorsal fragments of cara-pace, showing extensive tuberculation. A, WAM 70.206a (loc.37) showing carapace horn (h) and anteriormost carapace groove(ant); B, WAM 70.207a (loc. 37).
and a total length of perhaps 22 cm. WAM 70.205a yieldsestimates twice this value.
Discussion. Rolfe (in Brunton 1969) considered this formto represent fragments of large specimens of Montecarisgogoensis, and estimates of the size of the specimens (whereextrapolation is possible) indicate that they fall withinthe appropriate size range. The distribution of the cara-pace tubercles suggests that these features coincide withthe ‘suppression’ of grooves (compare Fig. 3). This formmay represent intraspecific variation but the possibility thatit is a different species cannot be ruled out.
Ceratiocarididae Salter, 1860Genus Schugurocaris Krestovnikov, 1961
Type species. Schugurocaris magnifica Krestovnikov,1961 from the Upper Devonian of the Volga region of theSouthern Urals of Russia.
Other species. Schugurocaris coherbaria (Chlupac,1963), S. cornwallisensis (Copeland, 1960) including S. c.cornwallisensis (Chlupac, 1963) and S. c. damesi (Chlupac,1963), S. grata (Barrande in Novak, 1885), S. ignotaKrestovnikov, 1961, S. libelluliformis (Krestovnikov, 1961),S. magnifica sargaica Krestovnikov, 1961, S. m. tenuilin-eata Krestovnikov, 1961, S. sinuosa (Krestovnikov, 1961).
Diagnosis. Elongate pointed telson triangular in crosssection, elevated into a sharp angular ridge along the mid-line; long narrow furcal rami; abdomen, telson and furcaornamented with cusps and oblique striae.
Occurrence. Upper Silurian of the Canadian Arctic(Copeland 1960, 1962, 1971), Silurian and Upper
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
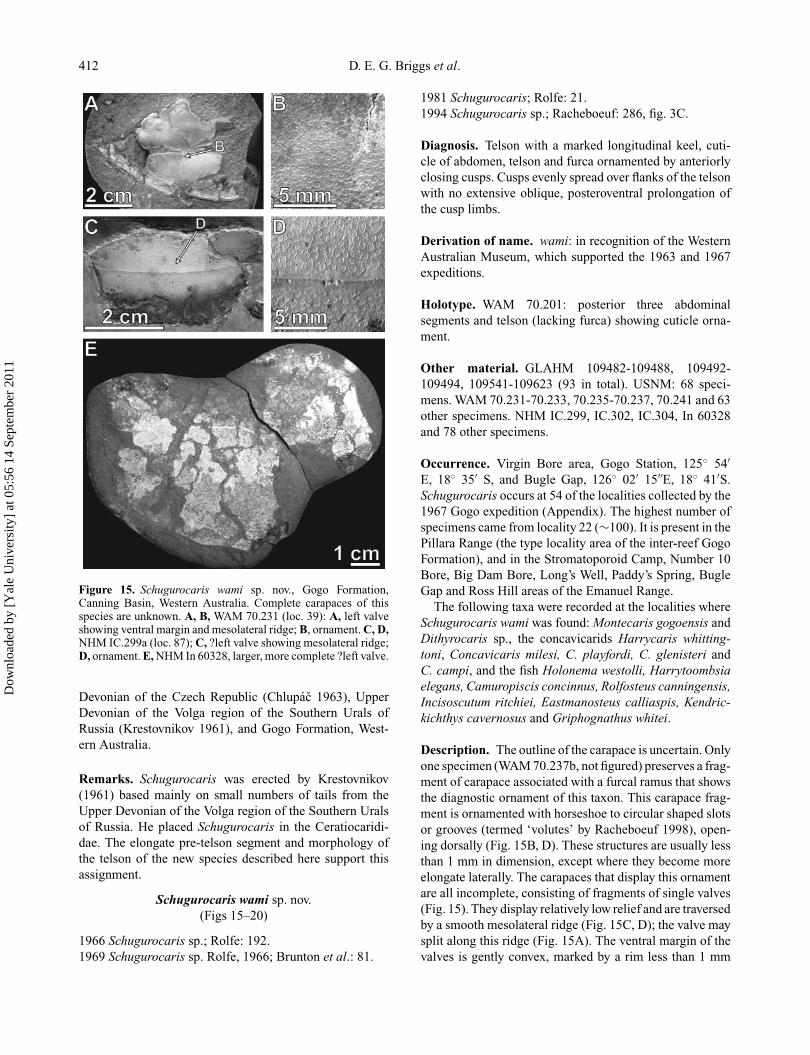
412 D. E. G. Briggs et al.
Figure 15. Schugurocaris wami sp. nov., Gogo Formation,Canning Basin, Western Australia. Complete carapaces of thisspecies are unknown. A, B, WAM 70.231 (loc. 39): A, left valveshowing ventral margin and mesolateral ridge; B, ornament. C, D,NHM IC.299a (loc. 87); C, ?left valve showing mesolateral ridge;D, ornament. E, NHM In 60328, larger, more complete ?left valve.
Devonian of the Czech Republic (Chlupac 1963), UpperDevonian of the Volga region of the Southern Urals ofRussia (Krestovnikov 1961), and Gogo Formation, West-ern Australia.
Remarks. Schugurocaris was erected by Krestovnikov(1961) based mainly on small numbers of tails from theUpper Devonian of the Volga region of the Southern Uralsof Russia. He placed Schugurocaris in the Ceratiocaridi-dae. The elongate pre-telson segment and morphology ofthe telson of the new species described here support thisassignment.
Schugurocaris wami sp. nov.(Figs 15–20)
1966 Schugurocaris sp.; Rolfe: 192.1969 Schugurocaris sp. Rolfe, 1966; Brunton et al.: 81.
1981 Schugurocaris; Rolfe: 21.1994 Schugurocaris sp.; Racheboeuf: 286, fig. 3C.
Diagnosis. Telson with a marked longitudinal keel, cuti-cle of abdomen, telson and furca ornamented by anteriorlyclosing cusps. Cusps evenly spread over flanks of the telsonwith no extensive oblique, posteroventral prolongation ofthe cusp limbs.
Derivation of name. wami: in recognition of the WesternAustralian Museum, which supported the 1963 and 1967expeditions.
Holotype. WAM 70.201: posterior three abdominalsegments and telson (lacking furca) showing cuticle orna-ment.
Other material. GLAHM 109482-109488, 109492-109494, 109541-109623 (93 in total). USNM: 68 speci-mens. WAM 70.231-70.233, 70.235-70.237, 70.241 and 63other specimens. NHM IC.299, IC.302, IC.304, In 60328and 78 other specimens.
Occurrence. Virgin Bore area, Gogo Station, 125◦ 54′
E, 18◦ 35′ S, and Bugle Gap, 126◦ 02′ 15′′E, 18◦ 41′S.Schugurocaris occurs at 54 of the localities collected by the1967 Gogo expedition (Appendix). The highest number ofspecimens came from locality 22 (∼100). It is present in thePillara Range (the type locality area of the inter-reef GogoFormation), and in the Stromatoporoid Camp, Number 10Bore, Big Dam Bore, Long’s Well, Paddy’s Spring, BugleGap and Ross Hill areas of the Emanuel Range.
The following taxa were recorded at the localities whereSchugurocaris wami was found: Montecaris gogoensis andDithyrocaris sp., the concavicarids Harrycaris whitting-toni, Concavicaris milesi, C. playfordi, C. glenisteri andC. campi, and the fish Holonema westolli, Harrytoombsiaelegans, Camuropiscis concinnus, Rolfosteus canningensis,Incisoscutum ritchiei, Eastmanosteus calliaspis, Kendric-kichthys cavernosus and Griphognathus whitei.
Description. The outline of the carapace is uncertain. Onlyone specimen (WAM 70.237b, not figured) preserves a frag-ment of carapace associated with a furcal ramus that showsthe diagnostic ornament of this taxon. This carapace frag-ment is ornamented with horseshoe to circular shaped slotsor grooves (termed ‘volutes’ by Racheboeuf 1998), open-ing dorsally (Fig. 15B, D). These structures are usually lessthan 1 mm in dimension, except where they become moreelongate laterally. The carapaces that display this ornamentare all incomplete, consisting of fragments of single valves(Fig. 15). They display relatively low relief and are traversedby a smooth mesolateral ridge (Fig. 15C, D); the valve maysplit along this ridge (Fig. 15A). The ventral margin of thevalves is gently convex, marked by a rim less than 1 mm
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
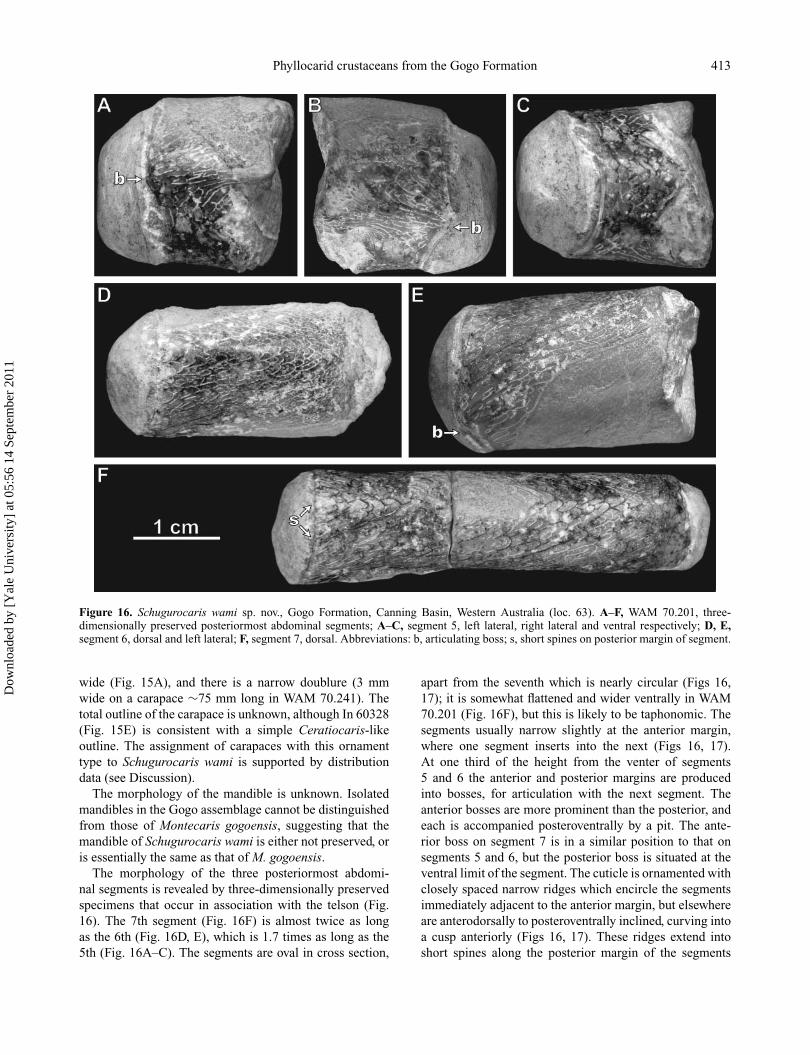
Phyllocarid crustaceans from the Gogo Formation 413
Figure 16. Schugurocaris wami sp. nov., Gogo Formation, Canning Basin, Western Australia (loc. 63). A–F, WAM 70.201, three-dimensionally preserved posteriormost abdominal segments; A–C, segment 5, left lateral, right lateral and ventral respectively; D, E,segment 6, dorsal and left lateral; F, segment 7, dorsal. Abbreviations: b, articulating boss; s, short spines on posterior margin of segment.
wide (Fig. 15A), and there is a narrow doublure (3 mmwide on a carapace ∼75 mm long in WAM 70.241). Thetotal outline of the carapace is unknown, although In 60328(Fig. 15E) is consistent with a simple Ceratiocaris-likeoutline. The assignment of carapaces with this ornamenttype to Schugurocaris wami is supported by distributiondata (see Discussion).
The morphology of the mandible is unknown. Isolatedmandibles in the Gogo assemblage cannot be distinguishedfrom those of Montecaris gogoensis, suggesting that themandible of Schugurocaris wami is either not preserved, oris essentially the same as that of M. gogoensis.
The morphology of the three posteriormost abdomi-nal segments is revealed by three-dimensionally preservedspecimens that occur in association with the telson (Fig.16). The 7th segment (Fig. 16F) is almost twice as longas the 6th (Fig. 16D, E), which is 1.7 times as long as the5th (Fig. 16A–C). The segments are oval in cross section,
apart from the seventh which is nearly circular (Figs 16,17); it is somewhat flattened and wider ventrally in WAM70.201 (Fig. 16F), but this is likely to be taphonomic. Thesegments usually narrow slightly at the anterior margin,where one segment inserts into the next (Figs 16, 17).At one third of the height from the venter of segments5 and 6 the anterior and posterior margins are producedinto bosses, for articulation with the next segment. Theanterior bosses are more prominent than the posterior, andeach is accompanied posteroventrally by a pit. The ante-rior boss on segment 7 is in a similar position to that onsegments 5 and 6, but the posterior boss is situated at theventral limit of the segment. The cuticle is ornamented withclosely spaced narrow ridges which encircle the segmentsimmediately adjacent to the anterior margin, but elsewhereare anterodorsally to posteroventrally inclined, curving intoa cusp anteriorly (Figs 16, 17). These ridges extend intoshort spines along the posterior margin of the segments
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
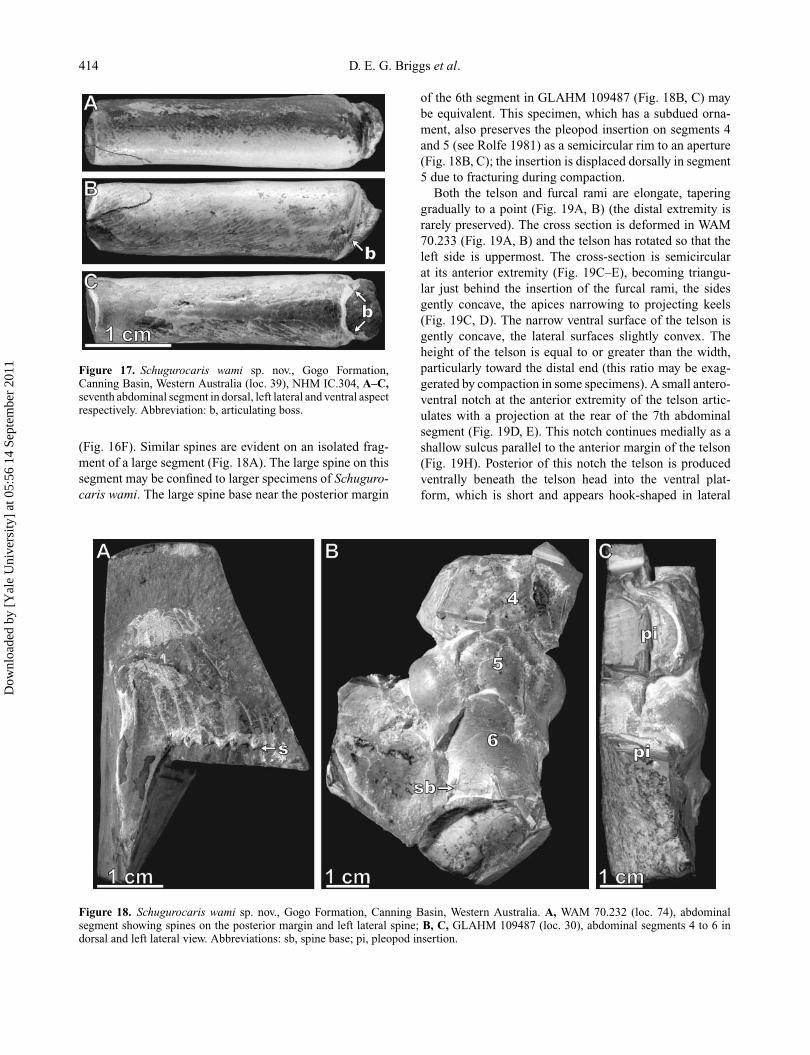
414 D. E. G. Briggs et al.
Figure 17. Schugurocaris wami sp. nov., Gogo Formation,Canning Basin, Western Australia (loc. 39), NHM IC.304, A–C,seventh abdominal segment in dorsal, left lateral and ventral aspectrespectively. Abbreviation: b, articulating boss.
(Fig. 16F). Similar spines are evident on an isolated frag-ment of a large segment (Fig. 18A). The large spine on thissegment may be confined to larger specimens of Schuguro-caris wami. The large spine base near the posterior margin
of the 6th segment in GLAHM 109487 (Fig. 18B, C) maybe equivalent. This specimen, which has a subdued orna-ment, also preserves the pleopod insertion on segments 4and 5 (see Rolfe 1981) as a semicircular rim to an aperture(Fig. 18B, C); the insertion is displaced dorsally in segment5 due to fracturing during compaction.
Both the telson and furcal rami are elongate, taperinggradually to a point (Fig. 19A, B) (the distal extremity israrely preserved). The cross section is deformed in WAM70.233 (Fig. 19A, B) and the telson has rotated so that theleft side is uppermost. The cross-section is semicircularat its anterior extremity (Fig. 19C–E), becoming triangu-lar just behind the insertion of the furcal rami, the sidesgently concave, the apices narrowing to projecting keels(Fig. 19C, D). The narrow ventral surface of the telson isgently concave, the lateral surfaces slightly convex. Theheight of the telson is equal to or greater than the width,particularly toward the distal end (this ratio may be exag-gerated by compaction in some specimens). A small antero-ventral notch at the anterior extremity of the telson artic-ulates with a projection at the rear of the 7th abdominalsegment (Fig. 19D, E). This notch continues medially as ashallow sulcus parallel to the anterior margin of the telson(Fig. 19H). Posterior of this notch the telson is producedventrally beneath the telson head into the ventral plat-form, which is short and appears hook-shaped in lateral
Figure 18. Schugurocaris wami sp. nov., Gogo Formation, Canning Basin, Western Australia. A, WAM 70.232 (loc. 74), abdominalsegment showing spines on the posterior margin and left lateral spine; B, C, GLAHM 109487 (loc. 30), abdominal segments 4 to 6 indorsal and left lateral view. Abbreviations: sb, spine base; pi, pleopod insertion.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
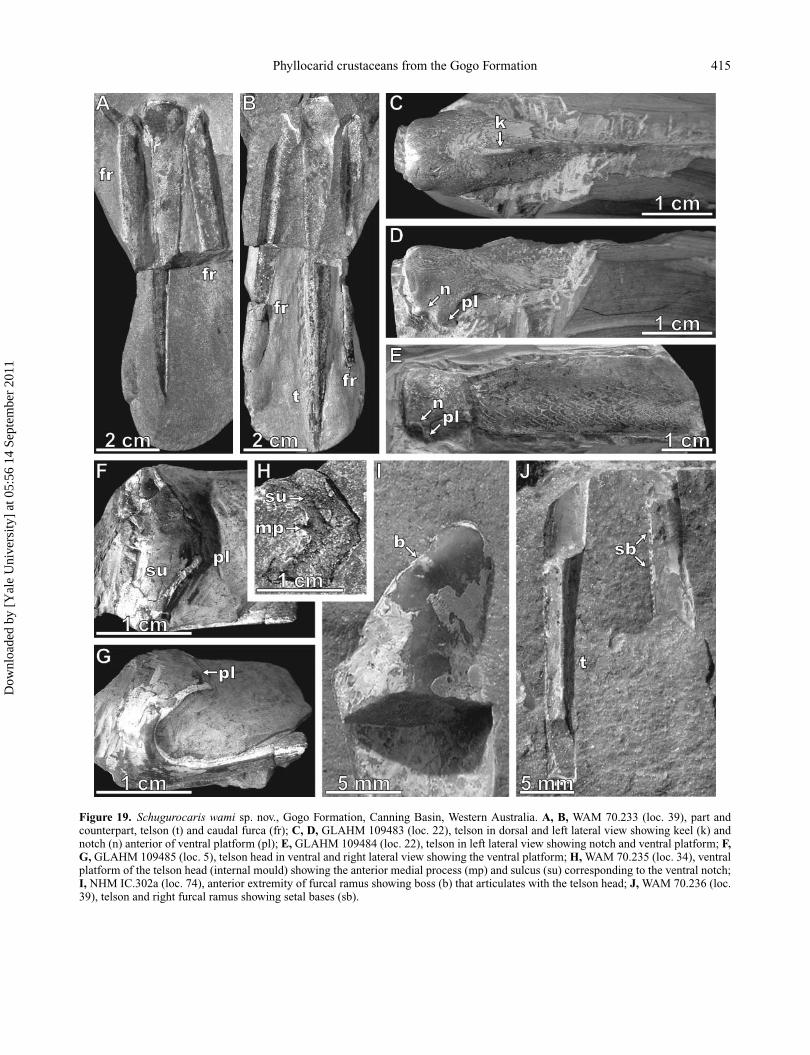
Phyllocarid crustaceans from the Gogo Formation 415
Figure 19. Schugurocaris wami sp. nov., Gogo Formation, Canning Basin, Western Australia. A, B, WAM 70.233 (loc. 39), part andcounterpart, telson (t) and caudal furca (fr); C, D, GLAHM 109483 (loc. 22), telson in dorsal and left lateral view showing keel (k) andnotch (n) anterior of ventral platform (pl); E, GLAHM 109484 (loc. 22), telson in left lateral view showing notch and ventral platform; F,G, GLAHM 109485 (loc. 5), telson head in ventral and right lateral view showing the ventral platform; H, WAM 70.235 (loc. 34), ventralplatform of the telson head (internal mould) showing the anterior medial process (mp) and sulcus (su) corresponding to the ventral notch;I, NHM IC.302a (loc. 74), anterior extremity of furcal ramus showing boss (b) that articulates with the telson head; J, WAM 70.236 (loc.39), telson and right furcal ramus showing setal bases (sb).
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

416 D. E. G. Briggs et al.
aspect (Fig. 19F, G). The ventral platform is gently convexventrally and V-shaped in outline, the sides converging toa short transverse, slightly concave, posterior margin (Fig.19F, G). The anterior margin forms a broad V-shaped inden-tation, with a short median process (Fig. 19H). The furcalrami are triangular in cross section but much flatter than thetelson, the ventral surface wider than the lateral. A smallanterior boss (Fig. 19I) forms part of the articulation withthe telson head. The relative lengths of telson and rami areunknown, but extrapolation suggests that the telson was alittle longer. The posterior part of the telson bears a seriesof closely spaced setae (∼2 per mm) on both ventral keels,and setae are also present on the inner margin of the furcalrami, except anteriorly (Fig. 19J). The setae themselves arenot preserved. The cuticle of both the furcal rami and telsondisplays a ridged sculpture of anteriorly closing cusps (Fig.19E) similar to that on the abdomen, except along the crestof the telson which is smooth. On the telson head thesecusps are replaced by lines radiating out from the anteriormargin (Fig. 19C–E).
Dimensions. No complete specimens of Schugurocariswami are known. The distal end of the telson is rarelyevident, and the maximum telson length observed is120 mm (GLAHM 109488). The cross-section of othertelsons, such as GLAHM 109484 (Fig. 19E), indicates thatthey represent larger specimens. Omitting short, brokenfragments, the mean length of 38 incomplete telsons is55.7 mm, ranging from 23 to 120 mm. The ratio of incom-plete telson length:width ranges from 4.4 to 11.0, the meanratio of 35 specimens being 6.7. The longest furcal ramusknown, which is incomplete, occurs in association with thelongest telson, and is 98 mm long (GLAHM 109488).
Discussion. Only one specimen (WAM 70.237b) preservesa fragment of carapace with a volute ornament associatedwith a furcal ramus of Schugurocaris wami type. Of 24isolated carapace fragments occurring at localities that alsoyield Schugurocaris wami tails but no examples of Dithyro-caris, 18 bear the volute ornament, four show slots and theremaining two lack ornament. This provides further supportfor the association of these carapaces with the abdomen andfurcal ramus of Schugurocaris wami rather than Dithyro-caris (see below). A similar ornament was illustrated byStumm & Chilman (1969) in Hebertocaris wideneri andDithyrocaris sp. cf. D. neptuni, and by Racheboeuf (1998)in D. oculeus.
Krestovnikov (1961) erected a number of species ofSchugurocaris based mainly on small numbers of tail spec-imens. The potential intraspecific variability in the dimen-sions and sculpture on the tail in his material remain to beinvestigated, and such partial information does not providea secure basis for the erection of species. The Gogo specieswas assigned to Schugurocaris by Rolfe in his original(1966) report of the discovery of the fauna. The distinc-
tive sculpture of the abdomen and the tail, combined withthe morphology and dimensions of the telson and furcalrami, confirm the generic assignment.
Rolfe (1963) observed that the triangular telson of C.cornwallisensis Copeland, 1960 “is probably sufficient forgeneric distinction from Ceratiocaris . . . and this specimenmay be referable to Schugurocaris”. Following discoveryof Schugurocaris wami described here, Rolfe (1966) againsuggested that the genus may occur in the Ludlovian ofBritish Columbia and in the Gedinnian and Emsian of theCzech Republic, i.e. represented by other species formerlyassigned to Ceratiocaris: S. coherbaria (Chlupac, 1963),S. cornwallisensis (Copeland, 1960) and S. cornwallisensisdamesi (Chlupac, 1963). These identifications are main-tained here and S. wami is compared with all the speciesthat can be assigned to the genus. Rolfe (1963) tentativelysynonymized Neurocaris with Schugurocaris; this identifi-cation is also accepted here. Racheboeuf (1994) suggestedthat the triangular section of the telson alone may be aninsufficient character to discriminate Schugurocaris fromCeratiocaris.
All Krestovnikov’s (1961) species (apart from S. sinuosa)display a distinct median keel running along the telson, likethat in S. wami. The type species S. magnifica (as wellas the two subspecies, which are of doubtful value, beingbased on the proportions of the telson of 1 and 2 specimensrespectively) is diagnosed on the basis of incomplete speci-mens of the telson and furcal rami. Schugurocaris magnificadisplays a pattern of cusps, with anterior facing apices, onboth the telson and furcal rami (Rolfe 1969, fig. 152,1). In S.magnifica the telson cusps are hook-like and concentrateddorsally alongside the telson keel, the flanks of the telsonmainly showing only the long oblique limbs of the cusps,extending posteroventrally (Krestovnikov 1961, p. 19; pl.1, fig. 4; pl. 3, fig. 1). Schugurocaris wami differs in havingthe cusps evenly spread over the flanks of the telson, withno extensive prolongation of the cusp limbs (Fig. 19E). Incontrast to S. wami, S. magnifica also lacks setae on eitherthe telson or furca.
Schugurocaris ignota is known from just one telson frag-ment (Krestovnikov 1961, pp. 17–18, pl. 1, fig. 8). Thesculpture is preserved on only one side of what was inter-preted as a keel, and although similar to that of S. wami,the lack of other morphological features precludes furtherdiscussion. This fragment may only be a ventral telsonsurface of S. magnifica, making S. ignota truly ignota, i.e.unknown. The ornament on the telson and furcal rami of S.libelluliformis is characteristically different, consisting ofwavy obliquely transverse ridges. Schugurocaris sinuosa,which is known only from a fragment of the carapace orcuticle of an abdominal segment, displays a discontinuousornament quite different from that on S. wami.
Schugurocaris cornwallisensis from the Silurian ofCanada is the only Schugurocaris species known to preservethe complete carapace. The outline varies with orientation
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

Phyllocarid crustaceans from the Gogo Formation 417
to bedding, as it is preserved in a flattened state, butCopeland (1971) did not record a median ridge. Distinc-tive external ornamentation is lacking on the carapace, theabdominal segments and elements of the tail of S. cornwal-lisensis (Copeland 1960, 1962), in contrast to those of theGogo S. wami.
The morphology of the tail of S. c. damesi and S.coherbaria, from the Upper Silurian of the Czech Repub-lic, is markedly different from that of S. wami. The telsonof small S. c. damesi bears five longitudinal ridges proxi-mally, becoming diamond-shaped in cross section distally(Chlupac 1963, fig. 4). Chlupac (1994) therefore main-tained this species within Ceratiocaris and referred it tohis nominate subgenus Ceratiocaris (Ceratiocaris). Theventral articulation socket of the telson head of S. c. damesiis a circular pit (Rolfe 1963, l969, fig. 126), which contrastswith the laterally elongate sulcus in the Gogo species. Thefurcal rami, in contrast to those of S. wami, are smooth. Apair of short curved lateral spines projects half-way alongthe telson of S. coherbaria. The rami have a longitudinalkeel on the internal margin and may bear setae. The orna-ment on the telson forms chevrons on either side of a centralkeel and that on the rami runs diagonally across the surface(Chlupac 1963, pl. 12, fig. 6).
The subdued nature of the cuspate ornament and the pairof dorsal spines on the posterior dorsal margin of segment 6in Gogo specimen GLAHM 109487 (Fig. 18B) are unusual,and may indicate the presence of another Gogo phyllocaridtaxon. These paired spines are like those in Echinocarisarmata (Hall, 1863), a taxon which Hall subsequently foundwas only a large specimen of E. punctata (Hall, 1863) (see
Hall & Clarke 1888, p. 170, pl. 28, figs 6, 7; Rolfe 1969, fig.142,3a, b). Lacking further material, we prefer to be parsi-monious and follow Hall’s example by regarding GLAHM109487 as an older and larger instar of S. wami.
A reconstruction of the Gogo Schugurocaris describedhere was published by Racheboeuf (1994, fig. 3C), cour-tesy of W. D. I. Rolfe. In the absence of evidence for thecarapace, its outline was based on that of Ceratiocaris; theslot ornament shown was subsequently found to be that ofthe Dithyrocaris sp indet. described below. The carapace ofS. wami reconstructed here is very different (Fig. 20).
The material from the Gogo Formation is clearly differentfrom all other described species of the genus, justifying itsassignment to a new species, S. wami, here.
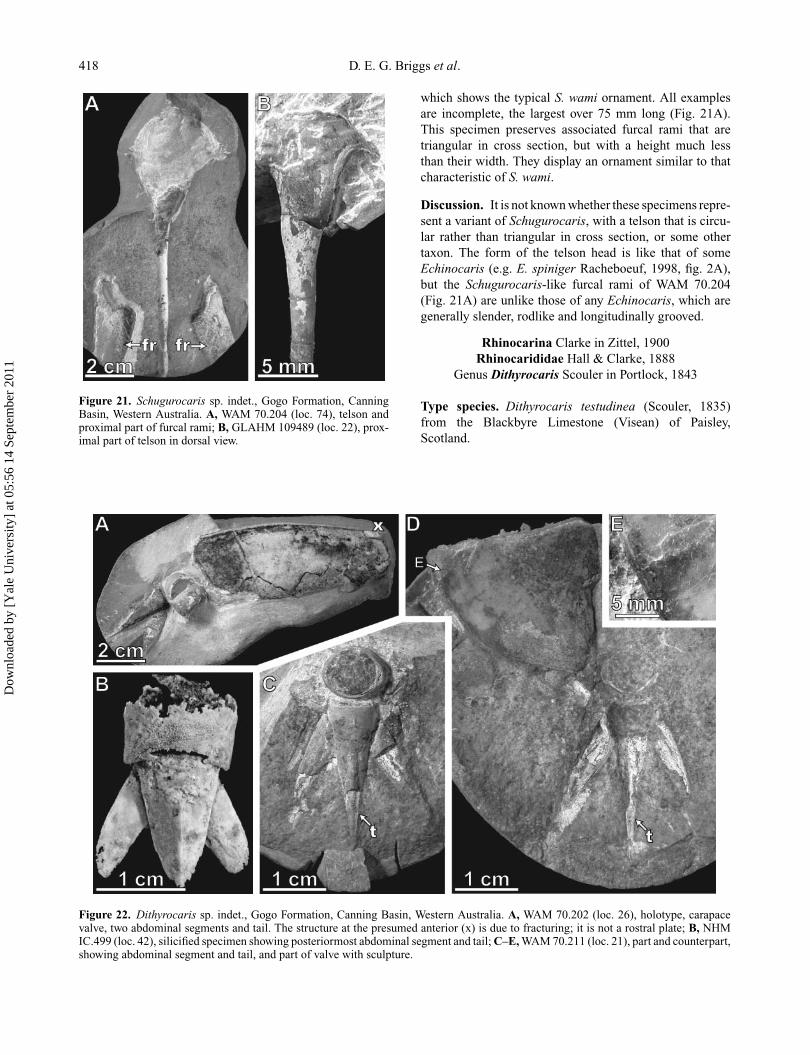
Schugurocaris sp. indet.(Fig. 21)
Material. GLAHM 109489, WAM 70.204, WAM 70.239,NHM IC.298.
Occurrence. Virgin Bore area, Gogo Station, 125◦ 54′
E, 18◦ 35′ S. Schugurocaris sp. indet. occurs at threeof the localities collected by the 1967 Gogo expedition(Appendix). It is present in the Stromatoporoid Camp andPaddy’s Spring areas.
Description. Four elongate telsons, two associated withfurcal rami morphologically identical to those of Schuguro-caris wami, are circular in cross section (Fig. 21A, B). Theynarrow distally from a normal S. wami telson head to a slen-der rod. These telsons are smooth except for WAM 70.239
Figure 20. Schugurocaris wami sp. nov., Gogo Formation, Canning Basin, Western Australia, reconstruction (drawn by J. K. Ingham).
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
418 D. E. G. Briggs et al.
Figure 21. Schugurocaris sp. indet., Gogo Formation, CanningBasin, Western Australia. A, WAM 70.204 (loc. 74), telson andproximal part of furcal rami; B, GLAHM 109489 (loc. 22), prox-imal part of telson in dorsal view.
which shows the typical S. wami ornament. All examplesare incomplete, the largest over 75 mm long (Fig. 21A).This specimen preserves associated furcal rami that aretriangular in cross section, but with a height much lessthan their width. They display an ornament similar to thatcharacteristic of S. wami.
Discussion. It is not known whether these specimens repre-sent a variant of Schugurocaris, with a telson that is circu-lar rather than triangular in cross section, or some othertaxon. The form of the telson head is like that of someEchinocaris (e.g. E. spiniger Racheboeuf, 1998, fig. 2A),but the Schugurocaris-like furcal rami of WAM 70.204(Fig. 21A) are unlike those of any Echinocaris, which aregenerally slender, rodlike and longitudinally grooved.
Rhinocarina Clarke in Zittel, 1900Rhinocarididae Hall & Clarke, 1888
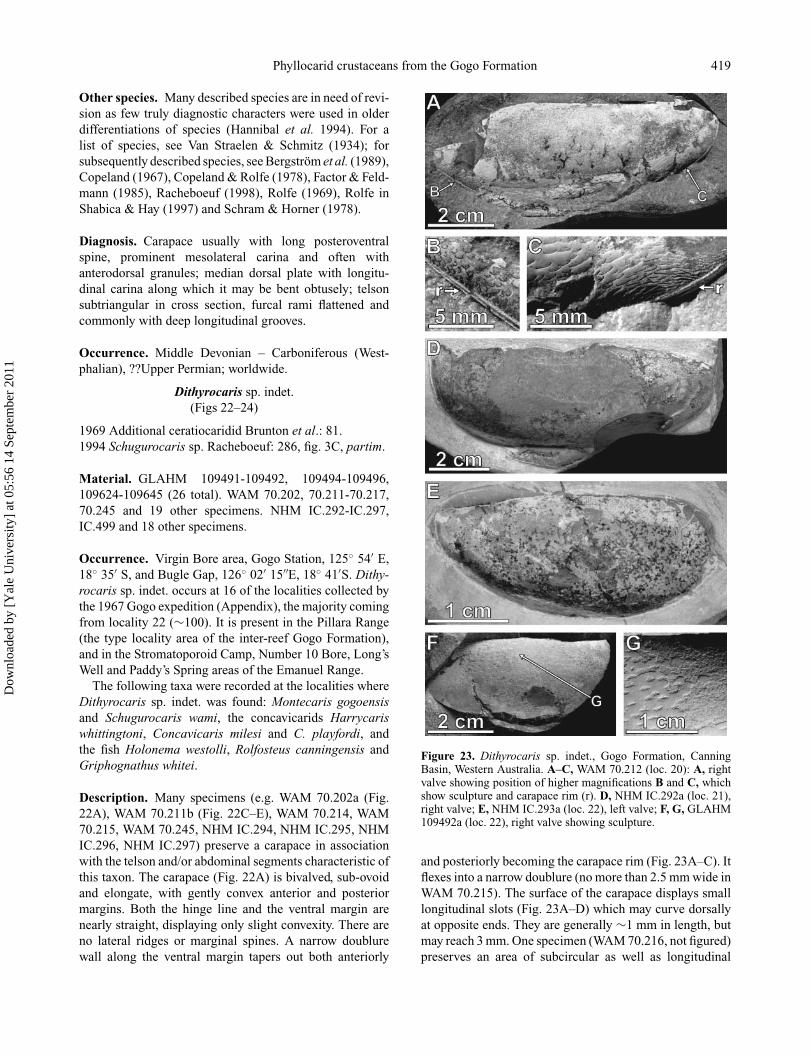
Genus Dithyrocaris Scouler in Portlock, 1843
Type species. Dithyrocaris testudinea (Scouler, 1835)from the Blackbyre Limestone (Visean) of Paisley,Scotland.
Figure 22. Dithyrocaris sp. indet., Gogo Formation, Canning Basin, Western Australia. A, WAM 70.202 (loc. 26), holotype, carapacevalve, two abdominal segments and tail. The structure at the presumed anterior (x) is due to fracturing; it is not a rostral plate; B, NHMIC.499 (loc. 42), silicified specimen showing posteriormost abdominal segment and tail; C–E, WAM 70.211 (loc. 21), part and counterpart,showing abdominal segment and tail, and part of valve with sculpture.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
Phyllocarid crustaceans from the Gogo Formation 419
Other species. Many described species are in need of revi-sion as few truly diagnostic characters were used in olderdifferentiations of species (Hannibal et al. 1994). For alist of species, see Van Straelen & Schmitz (1934); forsubsequently described species, see Bergstrom et al. (1989),Copeland (1967), Copeland & Rolfe (1978), Factor & Feld-mann (1985), Racheboeuf (1998), Rolfe (1969), Rolfe inShabica & Hay (1997) and Schram & Horner (1978).
Diagnosis. Carapace usually with long posteroventralspine, prominent mesolateral carina and often withanterodorsal granules; median dorsal plate with longitu-dinal carina along which it may be bent obtusely; telsonsubtriangular in cross section, furcal rami flattened andcommonly with deep longitudinal grooves.
Occurrence. Middle Devonian – Carboniferous (West-phalian), ??Upper Permian; worldwide.
Dithyrocaris sp. indet.(Figs 22–24)
1969 Additional ceratiocaridid Brunton et al.: 81.1994 Schugurocaris sp. Racheboeuf: 286, fig. 3C, partim.
Material. GLAHM 109491-109492, 109494-109496,109624-109645 (26 total). WAM 70.202, 70.211-70.217,70.245 and 19 other specimens. NHM IC.292-IC.297,IC.499 and 18 other specimens.
Occurrence. Virgin Bore area, Gogo Station, 125◦ 54′ E,18◦ 35′ S, and Bugle Gap, 126◦ 02′ 15′′E, 18◦ 41′S. Dithy-rocaris sp. indet. occurs at 16 of the localities collected bythe 1967 Gogo expedition (Appendix), the majority comingfrom locality 22 (∼100). It is present in the Pillara Range(the type locality area of the inter-reef Gogo Formation),and in the Stromatoporoid Camp, Number 10 Bore, Long’sWell and Paddy’s Spring areas of the Emanuel Range.
The following taxa were recorded at the localities whereDithyrocaris sp. indet. was found: Montecaris gogoensisand Schugurocaris wami, the concavicarids Harrycariswhittingtoni, Concavicaris milesi and C. playfordi, andthe fish Holonema westolli, Rolfosteus canningensis andGriphognathus whitei.
Description. Many specimens (e.g. WAM 70.202a (Fig.22A), WAM 70.211b (Fig. 22C–E), WAM 70.214, WAM70.215, WAM 70.245, NHM IC.294, NHM IC.295, NHMIC.296, NHM IC.297) preserve a carapace in associationwith the telson and/or abdominal segments characteristic ofthis taxon. The carapace (Fig. 22A) is bivalved, sub-ovoidand elongate, with gently convex anterior and posteriormargins. Both the hinge line and the ventral margin arenearly straight, displaying only slight convexity. There areno lateral ridges or marginal spines. A narrow doublurewall along the ventral margin tapers out both anteriorly
Figure 23. Dithyrocaris sp. indet., Gogo Formation, CanningBasin, Western Australia. A–C, WAM 70.212 (loc. 20): A, rightvalve showing position of higher magnifications B and C, whichshow sculpture and carapace rim (r). D, NHM IC.292a (loc. 21),right valve; E, NHM IC.293a (loc. 22), left valve; F, G, GLAHM109492a (loc. 22), right valve showing sculpture.
and posteriorly becoming the carapace rim (Fig. 23A–C). Itflexes into a narrow doublure (no more than 2.5 mm wide inWAM 70.215). The surface of the carapace displays smalllongitudinal slots (Fig. 23A–D) which may curve dorsallyat opposite ends. They are generally ∼1 mm in length, butmay reach 3 mm. One specimen (WAM 70.216, not figured)preserves an area of subcircular as well as longitudinal
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011
420 D. E. G. Briggs et al.
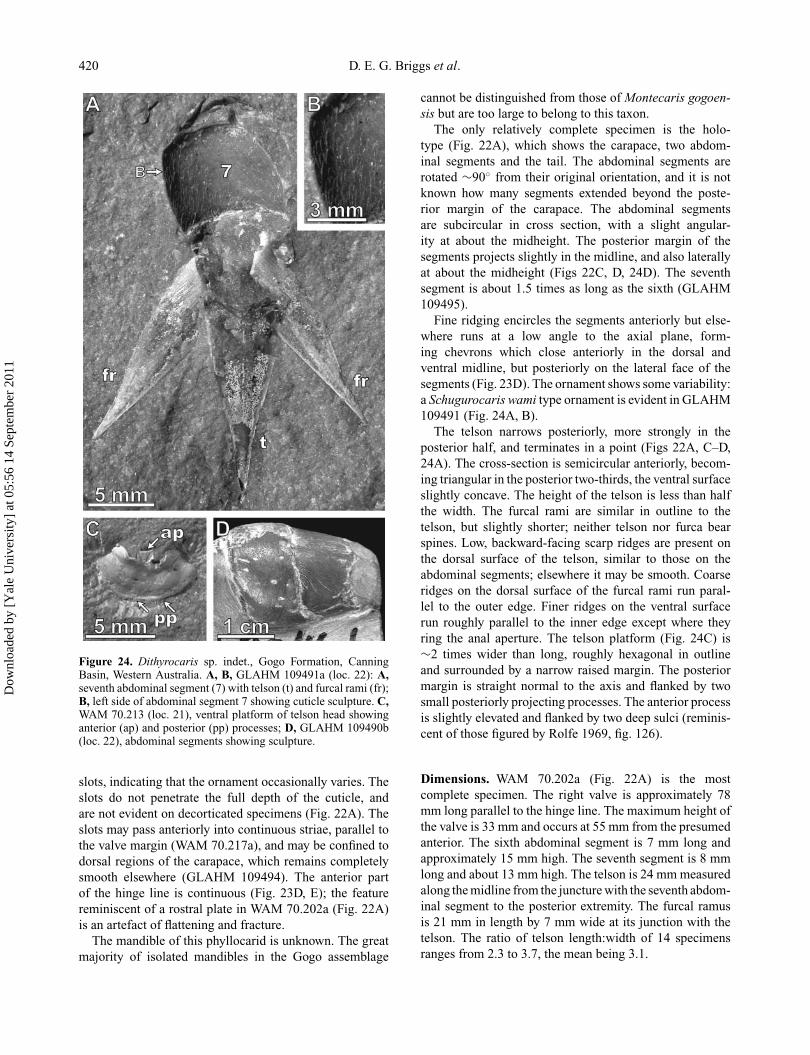
Figure 24. Dithyrocaris sp. indet., Gogo Formation, CanningBasin, Western Australia. A, B, GLAHM 109491a (loc. 22): A,seventh abdominal segment (7) with telson (t) and furcal rami (fr);B, left side of abdominal segment 7 showing cuticle sculpture. C,WAM 70.213 (loc. 21), ventral platform of telson head showinganterior (ap) and posterior (pp) processes; D, GLAHM 109490b(loc. 22), abdominal segments showing sculpture.
slots, indicating that the ornament occasionally varies. Theslots do not penetrate the full depth of the cuticle, andare not evident on decorticated specimens (Fig. 22A). Theslots may pass anteriorly into continuous striae, parallel tothe valve margin (WAM 70.217a), and may be confined todorsal regions of the carapace, which remains completelysmooth elsewhere (GLAHM 109494). The anterior partof the hinge line is continuous (Fig. 23D, E); the featurereminiscent of a rostral plate in WAM 70.202a (Fig. 22A)is an artefact of flattening and fracture.
The mandible of this phyllocarid is unknown. The greatmajority of isolated mandibles in the Gogo assemblage
cannot be distinguished from those of Montecaris gogoen-sis but are too large to belong to this taxon.
The only relatively complete specimen is the holo-type (Fig. 22A), which shows the carapace, two abdom-inal segments and the tail. The abdominal segments arerotated ∼90◦ from their original orientation, and it is notknown how many segments extended beyond the poste-rior margin of the carapace. The abdominal segmentsare subcircular in cross section, with a slight angular-ity at about the midheight. The posterior margin of thesegments projects slightly in the midline, and also laterallyat about the midheight (Figs 22C, D, 24D). The seventhsegment is about 1.5 times as long as the sixth (GLAHM109495).
Fine ridging encircles the segments anteriorly but else-where runs at a low angle to the axial plane, form-ing chevrons which close anteriorly in the dorsal andventral midline, but posteriorly on the lateral face of thesegments (Fig. 23D). The ornament shows some variability:a Schugurocaris wami type ornament is evident in GLAHM109491 (Fig. 24A, B).
The telson narrows posteriorly, more strongly in theposterior half, and terminates in a point (Figs 22A, C–D,24A). The cross-section is semicircular anteriorly, becom-ing triangular in the posterior two-thirds, the ventral surfaceslightly concave. The height of the telson is less than halfthe width. The furcal rami are similar in outline to thetelson, but slightly shorter; neither telson nor furca bearspines. Low, backward-facing scarp ridges are present onthe dorsal surface of the telson, similar to those on theabdominal segments; elsewhere it may be smooth. Coarseridges on the dorsal surface of the furcal rami run paral-lel to the outer edge. Finer ridges on the ventral surfacerun roughly parallel to the inner edge except where theyring the anal aperture. The telson platform (Fig. 24C) is∼2 times wider than long, roughly hexagonal in outlineand surrounded by a narrow raised margin. The posteriormargin is straight normal to the axis and flanked by twosmall posteriorly projecting processes. The anterior processis slightly elevated and flanked by two deep sulci (reminis-cent of those figured by Rolfe 1969, fig. 126).
Dimensions. WAM 70.202a (Fig. 22A) is the mostcomplete specimen. The right valve is approximately 78mm long parallel to the hinge line. The maximum height ofthe valve is 33 mm and occurs at 55 mm from the presumedanterior. The sixth abdominal segment is 7 mm long andapproximately 15 mm high. The seventh segment is 8 mmlong and about 13 mm high. The telson is 24 mm measuredalong the midline from the juncture with the seventh abdom-inal segment to the posterior extremity. The furcal ramusis 21 mm in length by 7 mm wide at its junction with thetelson. The ratio of telson length:width of 14 specimensranges from 2.3 to 3.7, the mean being 3.1.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

Phyllocarid crustaceans from the Gogo Formation 421
Discussion. The abdomen/telson morphology and orna-ment are characteristic of Dithyrocaris, but the absenceof a prominent mesolateral carina and anterodorsal tuber-culation on the carapace is unusual in this genus. Analmost smooth carapace does occur, however, in Dithyro-caris glabra Woodward & Etheridge, 1873 (? = Colpocarischesterensis Worthen, 1884). Dithyrocaris glabra hasnuchal and faint cephalic ridges, and anterior tubercula-tion, while Dithyrocaris? sp. has distinctive slots.
Tropidocaris bicarinata? Beecher, 1884 from the Middleto Upper Frasnian of the Willesden Borehole number 1(BGS WN 367: Stubblefield 1947; Butler 1981) shows asimilar ornament dorsally on the carapace. These slots aresmaller and less deeply incised than in Dithyrocaris? sp. butthe specimens too are much smaller. On small specimensof the type material of T. bicarinata this ornament wasdescribed as striae or lines (Beecher 1884, p. 17, 1902, p.446). The pattern of slots in Dithyrocaris? sp. superficiallyresembles that of the posteriorly directed ‘papillae’ coveringthe carapace of D. rolfei Schram & Horner, 1978 (Factor& Feldman 1985, p. 324). These features of D. rolfei are∼0.5 mm long ridges or elongate tubercles terminating inbroken spine bases? in Carnegie Museum specimen LundUMV 3824. Similar short ridges occur in D. insignis Jones& Woodward, 1898 (pl. 27, fig. 1b, c; pl. 30, fig. 3c, d).
The distinctive carapace ornament of Dithyrocaris sp.indet. is similar to some of the straight lines on D. oculeusRacheboeuf, 1998, D. sp. cf. D. neptuni (Hall, 1863) Stumm& Chilman, 1969 and Hebertocaris wideneri Stumm &Chilman, 1969, but it is often more rectilinear and deeplyincised (Fig. 23F, G). Dithyrocaris sp. indet. differs fromthese in lacking a mesolateral carina.
The raised ridge ornament on the abdominal segmentsof many species of Dithyrocaris encircles the segments,forming anteriorly closing chevrons on the dorsum, whilstposteriorly closing ventrally. This is clearly evident in thetype species D. testudinea (Jones & Woodward, 1899, pl.29 fig. 14), D. sp. nov. Rolfe & Richardson in Shabica &Hay, 1997, Nahecaris stuertzi Jaekel, 1921 (Rolfe 1969;Bergstrom et al. 1987; suggested to be a junior synonym ofDilophaspis by Hahn 1990, but see Briggs & Bartels 2003),and also in Rhinocaris ehlersi Stewart, 1933 (Stumm &Chilman 1969). Dithyrocaris sp. indet. is unusual in show-ing anterior chevron closure on both dorsal and ventralsurfaces, although this is also evident on the sixth abdom-inal segment of D. oculeus Racheboeuf, 1998, but not onthe seventh segment.
Racheboeuf’s (1994) reconstruction is included in theabove synonymy of Dithyrocaris sp. indet. as some speci-mens here assigned to this taxon were formerly included inour concept of the Gogo Schugurocaris species, explainedabove in the Discussion of S. wami.
While this Gogo phyllocarid can be assigned to Dithy-rocaris, the incompleteness of the specimens and lack ofother key characters precludes its identification to species.
Palaeoecology
The Gogo fauna is dominated by pelagic organisms,comprising abundant radiolarians, ammonoids and fish.Rolfe (1966) noted the presence of benthic life (bryozoans,gastropods and ostracods), while Campbell & Barwick(1999, p. 107) recorded “multitudes of small Devonianmolluscs . . . indicating the fauna was living close to thebottom of the sea and that it was overcome by a cloud ofsediment that was deposited down the front of the reef”.The present investigation has revealed brachiopods andrare sponge spicules in concretions. Briggs & Rolfe (1983)believed that the concavicarids may have been benthic, butdismissed a burrowing paradigm. It is clear that the bottomfauna at Gogo is restricted.
The streamlined shape of the carapace of Montecarisgogoensis, and the substantial tail fan made up of telson,furca and furcal spines, suggest that this phyllocarid (Fig.13) was epibenthic or nektonic. The spinose abdomenwould not have been conducive to a burrowing habit. Manyisolated valves of M. gogoensis consist of anterodorsal frag-ments that may be the result of predator attacks (Fig. 12A,B), which have also been recorded on concavicarids and fish(Briggs & Rolfe 1983; Long 1991). The robust mandiblewith a well developed incisor process, strong molar denti-tion and powerful musculature suggests a carnivorous orscavenging mode of life, picking up large detritus directlywith the mouthparts (Rolfe 1969, p. 309). The mode of lifeof the other two phyllocarids is unknown.
Chlupac (1960) noted that the formations in the CzechRepublic that yield Montecaris are all of a similar lithologyand preserve a low diversity benthos. He attributed thisto low oxygen levels resulting from poor circulation. TheMontecaris from western Canada (Pratt 1987) occurs indeeper water sediments. The Bergisch Gladbach lithologiesyield a limited benthos of brachiopods and bivalves thatJux (1960) interpreted as a reflection of low nutrient levelsrather than lack of oxygen. Montecaris typically occurs indeeper water sediments offshore of carbonate banks (Pratt1987).
Schugurocaris, in contrast, is more commonly found inrocks dominated by a nektonic fauna, principally grapto-lites and rare arthropods (Copeland 1960, 1962, 1971). Theco-occurrence of Schugurocaris and Montecaris is rare,and otherwise recorded only in the Schugurov layers of theCentral Volga region (Krestovnikov 1961; Chlupac 1963;Rolfe 1966) where the type and other species of Schuguro-caris co-occur with Montecaris tatarica.
Montecaris gogoensis dominates at the Gogo localitieswhere it occurs in highest numbers (localities 30, 37) asdoes Schugurocaris wami (locality 22). Such differencesin relative abundance were noted in the field (Bruntonet al. l969, p. 81) and are borne out by this analysis.Montecaris gogoensis tended to predominate in collectionsmade closest to the reef, whereas S. wami predominated at
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

422 D. E. G. Briggs et al.
locality 22, 150 yards out from the reef edge. Dithyrocarissp. indet. is an order of magnitude rarer than the other twobut is likewise most abundant at locality 22. Although someadmixing of concretions by surface transport must haveoccurred, an indication of original relative abundance maybe retained in the samples. Such variation was also recordedin the distribution of Gogo concavicarids, but its interpreta-tion is likewise problematic (Briggs & Rolfe 1983, p. 265).Nevertheless, M. gogoensis and S. wami may have occu-pied separate habitats within the reef complex, reflectingdifferent modes of life.
Acknowledgements
The material was prepared by a succession of prepara-tors employed at the Hunterian Museum under ManpowerServices Commission auspices, supervised by ChristinaHeywood, Valerie Boa and Stan Wood. Camilla Nicholdesigned the database for Gogo material used in this study.Kim Bryan assisted with locality data. Neil D. L. Clark(Glasgow) arranged the loan of material from the Hunte-rian Museum; Claire Mellish facilitated recent access tothe material at the Natural History Museum, London,with which Andrew Ross and Julia Rolfe assisted. SimonPowell (Bristol) assisted with photography and SEM, EricaChampion prepared the photographic figures and assistedwith editing, and Jean Vannier (Lyon) contributed valuableinsights on recent leptostracan biology. Professor UlrichJux (Cologne) generously assisted W.D.I.R. in field collect-ing and discussing comparative German material in the1960s; Fig. 4D is his photograph. Professor Harry Whit-tington obtained funding from the National Science Foun-dation which enabled W.D.I.R. to join the 1967 GogoExpedition. P.D.B. was funded by a Natural EnvironmentResearch Council studentship. D.E.G.B. completed partof the research for this paper while a Visiting Professorat the Department of Geophysical Sciences, University ofChicago.
References
Attenborough, D. 1980. Life on Earth. 2nd edition. Reader’sDigest, London, 368 pp.
Attenborough, D. 1981. Discovering life on Earth. Collins,London, 223 pp.
Beecher, C. E. 1884. Ceratiocaridae from the Chemung andWaverly Groups at Warren, Pennsylvania. Report of Progress,Second Geological Survey of Pennsylvania, pp. 1–22.
Beecher, C. E. 1902. Revision of the Phyllocarida from theChemung and Waverly Groups of Pennsylvania. QuarterlyJournal of the Geological Society, 58, 441–449.
Bergstrom, J., Briggs, D. E. G., Dahl, E., Rolfe, W. D. I.& Sturmer, W. 1987. Nahecaris stuertzi, a phyllocaridcrustacean from the Lower Devonian Hunsruck Slate.Palaontologische Zeitschrift, 61, 273–298.
Bergstrom, J., Briggs, D. E. G., Dahl, E., Rolfe, W. D. I. &Sturmer, W. 1989. Rare phyllocarid crustaceans from the
Devonian Hunsruck Slate. Palaontologische Zeitschrift, 63,319–333.
Briggs, D. E. G. & Bartels, C. 2003. Case 3281: NahecarisJaekel, 1921 (Malacostraca, Phyllocarida, Archaeostraca):proposed precedence over Dilophaspis Traquair in Walther,1903. Bulletin of Zoological Nomenclature, 60, 269–271.
Briggs, D. E. G. & Rolfe, W. D. I. 1983. New Concavicarida(New Order: ?Crustacea) from the Upper Devonian of Gogo,Western Australia, and the palaeoecology and affinities of thegroup. Special Papers in Palaeontology, 30, 249–276.
Briggs, D. E. G., Sutton, M. D., Siveter, David J. & Siveter,Derek J. 2003. A new phyllocarid (Crustacea: Malacostraca)from the Silurian Fossil-Lagerstatte of Herefordshire, UK.Proceedings of the Royal Society of London B, 271, 131–138.
Brunton, C. H. C., Miles, R. S. & Rolfe W. D. I. 1969. Gogoexpedition 1967. Proceedings of the Geological Society ofLondon, 1655, 79–83.
Butler, D. E. 1981. Marine faunas from concealed Devonian rocksof southern England and their reflection of the Frasnian trans-gression. Geological Magazine, 118, 679–697.
Campbell, K. S. W. & Barwick, R. E. 1988. Geological andpalaeontological information and phylogenetic hypotheses.Geological Magazine, 125, 207–227.
Campbell, K. S. W. & Barwick, R. E. 1999. Dipnoan fishesfrom the Late Devonian Gogo Formation of Western Australia.Records of the Western Australian Museum, Supplement, 57,107–138.
Campbell, K. S. W. & Barwick, R. E. 2002. The axial postcra-nial structure of Griphognathus whitei from the Upper Devo-nian Gogo Formation of Western Australia: comparisons withother Devonian Dipnoans. Records of the Western AustralianMuseum, 21, 167–201.
Chlupac, I. 1960. Die Gattung Montecaris Jux (Crustacea,Phyllocarida) im alteren Palaozoikum der Tschechoslowakei.Geologie, 9, 638–649.
Chlupac, I. 1963. Phyllocarid crustaceans from the Silurian andDevonian of Czechoslovakia. Palaeontology, 6, 97–118.
Chlupac, I. 1994. Assemblages of phyllocarid crustaceans inthe Silurian and Devonian of Bohemia and their analogues.Geologica et Palaeontologica, 28, 1–25.
Choo, B., Long, J. A. & Trinajstic, K. 2009. A new genus andspecies of basal actinopterygian fish from the Upper DevonianGogo Formation of Western Australia. Acta Zoologica, 90,194–210.
Clarke, J. M. 1898. Notes on some crustaceans from the ChemungGroup of New York. 1. A singularly ornamented phyllocaridgenus, Pephricaris. New York State Museum, 49th AnnualReport of the Regents, 1895, 2, 731–733.
Claus, C. 1888. Uber den Organismus der Nebaliden und diesystematische Stellung der Leptostraken. Arbeiten aus demZoologischen Institute der Universitat Wien und der zoologis-chen Station in Triest, 8, 1–148.
Copeland, M. J. 1960. New occurrences of Ceratiocaris andPtychocaris (Phyllocarida) from the Canadian Arctic. Bulletinof the Geological Survey of Canada, 60, 49–54.
Copeland, M. J. 1962. Additional Silurian arthropods from theCanadian Arctic. Geological Survey of Canada Bulletin, 91,52–54.
Copeland, M. J. 1967. A new species of Dithyrocaris (Phyl-locarida) from the Imo Formation, Upper Mississippian, ofArkansas. Journal of Paleontology, 41, 1195–1196.
Copeland, M. J. 1971. Additional Silurian Arthropoda fromArctic and Eastern Canada. Geological Survey of CanadaBulletin, 200, 20–26.
Copeland, M. J. & Rolfe, W. D. I. 1978. Occurrence ofa large phyllocarid crustacean of Late Devonian – Early
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

Phyllocarid crustaceans from the Gogo Formation 423
Carboniferous age from Yukon Territory. Current Research,Part B, Geological Survey of Canada, 78–1B, 1–5.
Dahl, E. 1992. Aspects of malacostracan evolution. Acta Zoolog-ica, 73, 339–346.
Druce, E. C. 1976. Conodont biostratigraphy of the Upper Devo-nian reef complexes of the Canning Basin, Western Australia.Bureau of Mineral Resources Australia Bulletin, 158, 2 vols.,303 pp. + 97 pls.
Dzik, J. 1980. Isolated mandibles of early Palaeozoic phyllocaridCrustacea. Neues Jahrbuch fur Geologie und Palaontologie.Monatshefte, 1980, 87–106.
Factor, D. F. & Feldman, R. M. 1985. Systematics and paleoecol-ogy of malacostracan arthropods in the Bear Gulch Limestone(Namurian) of Central Montana. Annals of Carnegie Museum,54, 319–356.
Feldmann, R. M., Garassino, A. A. & Schweitzer, C. A. 2004.The presumed decapod, Palaeopemphix Gemmellaro, 1890,is a unique member of the Phyllocarida (Palaeopemphicida:Palaeopemphidae). Journal of Paleontology, 78, 340–348.
Gardiner, B. G. & Miles, R. S. 1975. Devonian fishes of theGogo Formation, Western Australia. Problemes actuels depaleontologie-evolution des vertebres. Colloque InternationalCNRS, 218, 73–79.
Glenister, B. F. 1958. Upper Devonian ammonoids from theManticoceras zone, Fitzroy Basin, Western Australia. Journalof Paleontology, 32, 58–96.
Glenister, B. F. & Klapper, G. J. 1966. Upper Devonianconodonts from the Canning Basin. Journal of Paleontology,40, 777–842.
Hahn, G. 1990. Revision von Dilophaspis lata (Crustacea, Unter-Devon). Geologica et Palaeontologica, 24, 11–15.
Hall, J. 1863. On the occurrence of crustacean remains of thegenera Ceratiocaris and Dithyrocaris, with a notice on somenew species from the Hamilton Group and Genesee Slate.Sixteenth Annual Report of the Condition of the New YorkState Cabinet of Natural History, Appendix D, pp. 71–75.
Hall, J. & Clarke, J. M. 1888. Trilobites and other Crus-tacea of the Oriskany, Upper Helderberg, Hamilton, Portage,Chemung and Catskill groups. New York State GeologicalSurvey, Natural History of New York, Palaeontology, 7, 64 +236 pp.
Hannibal, J. T. 2009. Inside-out Arthropoda: new taxa andthe reinterpretation of the enigmatic Devonian phyllocaridOhiocaris. 9th North American Paleontological Conven-tion, Abstracts, Cincinnati Museum Center, Cincinnati,212–213.
Hannibal, J. T., Feldmann, R. M. & Rolfe, W. D. I. 1994. Phyl-locarid crustaceans from the Devonian of Bolivia. Pp. 59–69in R. Suarez-Soruco (ed.) Fosiles y facies de Bolivia, vol. IIInvertebrados y Paleobotanica. Revista Tecnica de Yacimien-tos Petroliferos Fiscales Bolivianos, 13–14.
Jaekel, O. 1921. Uber einen neuen Phyllocariden aus demUnterdevon der Bundenbacher Dachschiefer. Zeitschrift derDeutschen Geologischen Gesellschaft, 72, 290–292.
Jones, T. R. & Woodward, H. 1898–1899. A monograph ofthe British Palaeozoic Phyllopoda (Phyllocarida, Packard).Palaeontographical Society Monograph, pp. 125–211.
Jux, U. 1959. Phyllocariden-Reste aus dem oberen Mitteldevonder Bergisch Gladbach-Paffrather Mulde. PalaontologischeZeitschrift, 33, 166–171.
Jux, U. 1960. Montecaris lehmanni, a new crustacean from theRhenish Devonian and the problem of its systematic position.Journal of Paleontology, 34, 1129–1152.
Krestovnikov, V. N. 1961. Novyye rakoobraznye fillokaridyPaleozoya Russkoi platformy, Urala, Timana i Donbassa
[New phyllocarid crustaceans from Palaeozoic of the Russianplateau, the Urals, Timan and the Donetz Basin]. TrudyGeologicheskogo Instituta, 52, 1–67.
Latreille, P. A. 1806. Histoire naturelle, generale et particuliere,des Crustaces et des Insectes. In Histoire naturelle, 80. Dufart,Paris, 408 pp.
Long, J. A. 1988a. The extraordinary fishes of Gogo. New Scien-tist, 120, 40–44.
Long, J. A. 1988b. Late Devonian fishes from Gogo, WesternAustralia. National Geographic Research, 4, 436–449.
Long, J. A. 1991. Arthrodire predation by Onychodus (Pisces,Crossopterygii) from the Late Devonian Gogo Forma-tion, Western Australia. Records of the Western AustralianMuseum, 15, 479–481.
Long, J. A. 1995a. A new plourdosteid arthrodire from the UpperDevonian Gogo Formation of Western Australia. Palaeontol-ogy, 38, 39–62.
Long, J. A. 1995b. The rise of fishes. University of New SouthWales Press, Sydney, 223 pp.
Long, J. A. 2006. Swimming in stone. The amazing Gogo fossilsof the Kimberley. Arts Centre Press, Fremantle, 320 pp.
Long, J. A. & Trinajstic, K. 2010. The Late Devonian GogoFormation Lagerstatten of Western Australia: Exceptionalearly vertebrate preservation and diversity. Annual Reviewof Earth and Planetary Sciences, 38, 255–279.
Miles, R. S. 1971. The Holonematidae (placoderm fishes), areview based on new specimens of Holonema from the UpperDevonian of Western Australia. Philosophical Transactionsof the Royal Society, B263, 101–234.
Nazarov, B. B. & Ormiston, A. R. 1983. Upper Devonian (Fras-nian) radiolarian fauna from the Gogo Formation, WesternAustralia. Micropaleontology, 28, 454–466.
Nazarov, B. B., Cockbain, A. E. & Playford, P. E. 1982. LateDevonian Radiolaria from the Gogo Formation, CanningBasin, Western Australia. Alcheringa, 6, 161–173.
Nicoll, R. S. 1977. Conodont apparatuses in an Upper Devonianpalaeoniscid fish from the Canning Basin, Western Australia.BMR Journal of Australian Geology and Geophysics, 2,217–228.
Novak, O. 1885. Nouveau Crustace Phyllocaride de l’etage F-f2, en Boheme. Sitzungsberichte der Konigl. BohmischenGesellschaft der Wissenschaften, 1886, 343–347.
Packard, A. S. Jr 1879. The nebaliad Crustacea as types of a neworder. American Naturalist, 13, 128.
Pennant, T. 1777. British Zoology, Volume 4. Crustacea,Mollusca, Testacea, 4th Edition. White, London, i-viii + 136pp.
Playford, P. E. 1980. Devonian “Great Barrier Reef” of CanningBasin, Western Australia. Bulletin American Association ofPetroleum Geologists, 64, 814–840.
Playford, P. E. & Lowry, D. C. 1966. Devonian reef complexes ofthe Canning Basin, Western Australia. Bulletin of the Geolog-ical Survey of Western Australia, 118, 150 pp.
Playford, P. E., Hocking, R. M. & Cockbain, A. E., eds.2009. Devonian reef complexes of the Canning Basin, West-ern Australia. Bulletin of the Geological Survey of WesternAustralia, 145, 1–444.
Playford, P. E., Hurley, N. F., Kerans, C. & Middleton, M. F.1989. Reefal platform development, Devonian of the CanningBasin, Western Australia. Pp. 187–202 in P. D. Crevello, J. L.Wilson, J. F. Sarg, & J. F. Read (eds) Controls on carbonateplatform and basin development. Society of Economic Pale-ontologists and Mineralogists, Tulsa, Special Publication, 44.
Portlock, J. E. 1843. Report of the geology of the countyof Londonderry, and of parts of Tyrone and Fermanagh.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011

424 D. E. G. Briggs et al.
Longman, Brown, Green and Longmans, London, 784 pp.+37 pls.
Pratt, B. R. 1987. The phyllocarid crustacean Montecaris from theDevonian of Northwest Canada. Canadian Journal of EarthSciences, 24, 1267–1269.
Racheboeuf, P. 1994. Silurian and Devonian phyllocarid crus-taceans from the Massif Armoricain, NW France. Revue dePaleobiologie, 13, 281–305.
Racheboeuf, P. 1998. Mid Devonian phyllocarid Crustacea fromBolivia. Palaeontology, 41, 103–124.
Rich, P. V., Van Tets, G. F. & Knight, F. 1985. Kadimakara,extinct vertebrates of Australia. Pioneer Design Studio, Lily-dale, 384 pp.
Rode, A. L. & Lieberman, B. S. 2002. Phylogenetic and biogeo-graphic analysis of Devonian phyllocarid crustaceans. Journalof Paleontology, 76, 271–286.
Rolfe, W. D. I. 1962. A new phyllocarid crustacean from theUpper Devonian of Ohio. Breviora, 151, 7 pp.
Rolfe, W. D. I. 1963. Morphology of the telson in Ceratiocaris?cornwallisensis (Crustacea: Phyllocarida) from Czechoslo-vakia. Journal of Paleontology, 37, 486–488.
Rolfe, W. D. I. 1966. Phyllocarid crustacean fauna of Europeanaspect from the Devonian of Western Australia. Nature, 209,192.
Rolfe, W. D. I. 1969. Phyllocarida. Pp. 296–331 in R. C. Moore(ed.) Treatise on Invertebrate Paleontology. Part R, Arthro-poda 4. Geological Society of America and University ofKansas Press, Lawrence, Kansas.
Rolfe, W. D. I. 1981. Phyllocarida and the origin of the Malacos-traca. Geobios, 14, 17–27.
Rolfe, W. D. I. & Edwards, V. A. 1979. Devonian Arthropoda(Trilobita and Ostracoda excluded). Pp. 325–329 in M. R.House, C. T. Scrutton & M. G. Bassett (eds). The DevonianSystem. Special Papers in Palaeontology, PalaeontologicalAssociation, Aberystwyth, 23.
Salter, J. W. 1860. On new fossil Crustacea from the SilurianRocks. The Annals and Magazine of Natural History, Series3, 5, 153–162.
Schram, F. R. 1986. Crustacea. Oxford University Press, Oxford,606 pp.
Schram, F. R. & Horner, J. 1978. Crustacea of the Mississip-pian Bear Gulch Limestone of Central Montana. Journal ofPaleontology, 52, 394–406.
Scouler, J. 1835. Account of some fossil Crustacea which occurin the Coal Formation. Records of General Science, 1, 136–141.
Seddon, G. 1970. Frasnian conodonts from the Sadler Ridge-Bugle Gap area, Canning Basin, Western Australia. Journalof the Geological Society of Australia, 16, 723–753.
Sepkoski, J. J. 2000. Crustacean biodiversity through the marinefossil record. Contributions to Zoology, 69, 213–222.
Shabica, C. W. & Hay, A. A. (eds) 1997. Richardson’s Guide tothe fossil fauna of Mazon Creek. Northeastern Illinois Univer-sity, Chicago, 18 + 308 pp.
Shuster, C. N. Jr. 1982. A pictorial review of the natural historyand ecology of the horseshoe crab Limulus polyphemus, withreference to other Limulidae. Pp. 1–52 in Physiology andbiology of horseshoe crabs: studies on normal and environ-mentally stressed animals. Alan R. Liss Inc., New York.
Stewart, G.A. 1933. A phyllocarid crustacean from the Devo-nian rocks of Ohio. American Midland Naturalist, 14, 363–366.
Stubblefield, C. J. 1947. Comments on cores from D’Arcy Explo-ration Company’s Borehole at Willesden. Proceedings of theGeological Society of London, 1437, 20.
Stumm, E. C. & Chilman, R. B. 1969. Phyllocarid crustaceansfrom the Middle Devonian Silica Shale of northwesternOhio and southeastern Michigan. Contributions from theMuseum of Paleontology, University of Michigan, 23, 53–71.
Tetlie, O. E., Braddy, S. J., Butler, P. D. & Briggs, D. E.G. 2004. A new eurypterid (Chelicerata: Eurypterida) fromthe Upper Devonian Gogo Formation of Western Australia,with a review of the Rhenopteridae. Palaeontology, 47, 801–809.
Trinajstic, K., Marshall, C., Long, J. A. & Bifield, K. 2007.Exceptional preservation of nerve and muscle tissues in LateDevonian placoderm fish and their evolutionary implications.Biology Letters, 3, 197–200.
Van Straelen, V. 1933. Note sur les Phyllocarides paleozoıques ouArchaeostraca de la Belgique et remarques sur la systematiquede ces Crustaces. Bulletin du Musee Royal d’HistoireNaturelle de Belgique, 9(16), 1–12.
Van Straelen, V. & Schmitz, G. 1934. Crustacea Phyllocarida(=Archaeostraca). Fossilium Catalogus I: Animalia, 64.W.Junk, Berlin, 246 pp.
Woodward, H. & Etheridge, R. 1873. Explanation ofSheet 23. Memoirs of the Geological Survey, Scotland,p. 99.
Won, M. Z. 1997a. Review of the family Entactiniidae (Radi-olaria), and taxonomy and morphology of Entactiniidae inthe Late Devonian (Frasnian) Gogo Formation, Australia.Micropalaeontology, 43, 333–369.
Won, M. Z. 1997b. The proposed new radiolarian subfam-ily Retentactiniinae (Entactiniidae) from the Late Devonian(Frasnian) Gogo Formation, Australia. Micropaleontology,43, 371–418.
Worthen, A. H. 1884. Descriptions of two new species of Crus-tacea, fifty-one species of Mollusca, and three species ofcrinoids, from the Carboniferous formations of Illinois andadjacent states. Illinois State Museum of Natural History,Bulletin, 2, 1–27.
Young, G. C., Burrow, C. J., Long, J. A., Turner, S., & Choo, B.2010. Devonian macrovertebrate assemblages and biogeog-raphy of East Gondwana (Australasia, Antarctica). Palaeo-world, 19, 55–74.
Zittel, K. A. 1900. Text-book of palaeontology. Macmillan,London and New York, 706 pp.
Dow
nloa
ded
by [
Yal
e U
nive
rsity
] at
05:
56 1
4 Se
ptem
ber
2011