the circadian system of crustaceans
TRANSCRIPT

Ctvwwbiology ln/ernational Vol. IO, No. I, pp. l-19 Z, 1993 International Society of Chronobiology
Review
The Circadian System of Crustaceans
Hugo Arkchiga, Fernando Fernhndez-Quiroz, Francisco Fernandez de Miguel, and Leonardo Rodriguez-Sosa
Llepartamento de Fisiologiu, Biojfsica y Newociencias, Centro de Investigucicin y de Estlldios Avunzados de1 IPN. Colimu, MLisico
Summary: Crustaceans exhibit a variety of overt circadian rhythms. Observations on intact animals suggest the existence of more than one circadian pacemaker in the nervous system. Ablation experiments so far have been inconclusive in pin- pointing the location of putative pacemakers. However, various structures, most notably the optic peduncle. have been shown to sustain circadian rhythmicity in vitro. Retinal sensitivity and neurosecretory activity display circadian rhythms in the isolated optic peduncle, but they are also responsive to synchronizing influ- ences from other regions of the central nervous system, most notably the supra- esophageal ganglion. A model based on a number of circadian pacemakers distrib- uted in the central nervous system best fits the experimental results at present. Coupling of rhythmicity between independent circadian pacemakers is likely to occur. and a neuroendocrine stage of integration has been proposed for several rhythms. Various entraining agents have been identified, and more than one may play a part in the synchronization of a given rhythm. Key Words: Crustacean biological rhythms-Circadian pacemaker-Entraining agents.
Crustaceans display a variety of circadian, ultradian, and infradian rhythms in their overt behavior and physiological functions ( 1,2). A great number of crustaceans are nocturnal, performing their social activities (exploration, foraging, mating) dur- ing twilight hours in the natural environment. They are spontaneously active during darkness under laboratory conditions. Circadian rhythmicity has been noted in activi- ties as diverse as locomotion, egg-hatching, sensory responsiveness, position of tegu- mentary and retinal shielding pigments, heart rate, and some metabolic functions (3,4). As in other organisms, each function has a definite acrophase, and the acro-
Received September 20, I99 I: accepted with revisions January 27. 1992. Address correspondence and reprint requests to Dr. H. Arkhiga, Departamento de Fisiologia. Biofisica,
y Neurociencias, Centro de Investigacibn y de Estudios Avanzados del IPN. Apartado Postal 14-740, 07000 Mkxico D.F.
Fernando Fernhnder-Quinbz’s present address is Facultad de Medicina. Universidad Autbnoma de Colima, Colima. Mtxico.

2 II. ARl?CHIGA ET AL
phases of various rhythms bear fixed phase relations to one another. Circadian rhythms interact with those of other frequencies in the modulation of a given func- tion. For instance, the level of locomotor activity exhibits a tidal as well as a circadian rhythm, with similar features in common, and is also influenced by the molt cycle, which has a much longer period. Ultradian components have also been described (5).
Some of the basic features of circadian rhythms have been established for crusta- ceans, as indicated by the following examples.
CIRCADIAN RHYTHMS
Persistence Under Constant Conditions of Illumination
For many years it has been known that rhythms persist in constant darkness (DD) or under continuous illumination (LL). The rate of damping under constant condi- tions is a feature of a rhythm. For instance, the locomotory rhythm is quite labile, disappearing within 2 weeks under DD. It continues for even less time under LL. There are, however, reports that it goes on for several weeks (3). More persistent are the rhythms of migration of tegumentary and retinal shielding pigments, electroreti- nogram amplitude, and responsiveness of visual interneurons (4,6). The rhythmicity conforms to Aschoffs empirical rule. It was initially established for the rhythm of responsiveness of visual interneurons of the crayfish Procamharzw clarki (7) and subsequently for the rhythm of locomotion and electroretinogram amplitude of Pro- cumharzu houvieri, that the period (7) was shorter and the activity/rest (a/p) quotient greater under DD than under LL (8). Recently 7 has been analyzed further for the locomotory rhythm of Procambarus clarki (9). The length of r shortens under DD to -22 h and increases under LL to -26 h. The a/p quotient is much greater under DD (1.2 1) than LL (0.34).
Entrainment
Entrainment has been studied mainly for light stimuli in the locomotory rhythm. In P. clarki the onset of darkness appears to be the entraining cue ( 10). Other entrain- ing agents are salinity and hydrostatic pressure, as in the case of the tidal rhythm of Carcinus (11). In the same preparation, chilling brings about a tidal rhythm lost after housing under constant conditions (12). Recently, food has been proposed as an effective entraining agent (9). Little information is available regarding phase-re- sponse curves (PRC) for crustacean circadian rhythms. After the initial reports of Webb ( 13) and Stevens ( 14) for the chromatophore rhythm in C%a, a PRC was determined for the circatidal rhythm of swimming in Excirolana (1.5) and more recently for the circatidal locomotor rhythm in Curcinus ( 16).
Temperature Compensation
The Qlo was first explored for the chromatophore rhythm in LTca ( 17,18) and later for the locomotory rhythm in several crustacean species (19). In all cases a Qlo in the range of 0.9- 1 .O was found. For the circadian rhythm of electrical activity in visual neurons, between 14.0 and 24.O”C the Qlo was estimated to be 1 .O (7).
Chronoh/ol Inl, Vol. 10, No. 1. 19Y.i

THE CIRCADIAN SYSTEM OF CRUSTACEANS
Internal Desynchronization
As in other animal groups, crustaceans under constant conditions not only exhibit a damping of their rhythms but they also undergo changes in their time structure. Some ultradian components appear, while circadian rhythms show differences in their periods and phases as well as in their rates of damping. These observations have been the basis for various interpretations about the nature of rhythm-generating mechanisms.
In Curcinw kept in the laboratory without tidal exposure but with maintenance of the LD cycle, a selective damping out of the circatidal component of the locomotory rhythm results, while the circadian component persists (27). In the crayfish Pwcum- hurz~s howieri simultaneous recording of locomotion and electrical activity in the retina shows that the two circadian rhythms fade away at different rates (20). This finding suggests that the rhythms affecting different physiological functions are driven by separate mechanisms. But even for a single rhythm, more than one compo- nent has been identified. For instance, two independent peaks have been described in the circatidal rhythmicity of various marine crustaceans (2 1,22), and Fernandez de Miguel and Arechiga (9) have shown that a food entrainment peak of locomotor activity in crayfish free-runs with a different periodicity from that of the LD entrain- ment peak. Two phase-control mechanisms also have been postulated for the tegu- mentary pigment position rhythm in UCU pzgilutor (23). Findings suggest that more than one central pacemaker may control circadian rhythmicity in crustaceans. The question of whether a single pacemaker controls the various expressions of rhythmi- city, through different phase-setting mechanisms. or whether different independent but coupled pacemakers generate the variety of overt rhythms remains unanswered.
THE PHYSIOLOGICAL MECHANISMS UNDERLYING OVERT RHYTHMS
Analysis of the physiological mechanisms underlying the various phenomenologi- cal properties of circadian rhythms in crustaceans is quite limited. Three aspects of the question are reviewed here: identification of the circadian pacemakers, location of the entrainment pathways, and analysis of internal coupling mechanisms for se- lected rhythms.
Circadian Pacemakers in Crustaceans
The mechanisms regulating circadian rhythms in crustaceans have been explored for several decades. As for other zoological groups, it was initially hypothesized that a single specialized organ generated the circadian timing and experiments using le- sions, ablations, or implantations could locate it. Two main organs have been identi- fied as essential for the maintenance of circadian rhythms in decapod crustaceans: the optic peduncle in the eyestalk and the supraesophageal ganglion.
The Optic Peduncle (OP) in the Eyestalk
The optic peduncle was the first candidate suggested as a site of generation of circadian rhythmicity in crustaceans. Its ablation was first proposed by Kalmus (24,25), and later confirmed by others, to abolish the circadian rhythm of locomotion

4 H. ARl?HIGA ET AL.
TABLE 1. Eyestalk hormones participating in circa&m rhythms
Circadian rhythm Hormone(s) Acrophase Reference(s)
Blood glucose level Distal retinal
pigment position Proximal retinal
pigment position Electroretinogram
amplitude
Locomotor activity
Tegumentary chromatophores
Egg-hatching
Hyperglycemic hormone Night 44,45,77 Pigment-dispersing hormone Day 4,6,29,75,76
SHydroxytryptamine Night 6,28,42,16
Pigment-dispersing hormone 5-Hydroxytryptamine
Day 4.3 1,38,39,42, 75,76,77
Neurodepressing hormone, 5-Hydroxytryptamine
Night
Day 1,4,6,64,73,77
Red pigment-concentrating hormone
Pigment-dispersing hormone Gonad-inhibiting hormone
Night Night 6,15,76,77
Day Night 2.17
(4). One more reason to consider the OP as the site of the circadian pacemaker is that the first rhythms identified in crustaceans were those of chromatophore position, metabolic activity, and locomotion. All these functions are influenced by hormones produced in the OP. The OP contains the X organ-sinus gland system, a conglomer- ate of 1 OO- 150 neurons that synthesize, store, and release neurohormones responsi- ble for the control of circadian rhythmic physiological functions (Table I). Thus, it was a plausible hypothesis that the same organ performing the synthesis and release of the neurosecretory substances could generate the time signal for their circadian control. This view was strengthened by the finding in Carcimts that the tidal rhythm of locomotion, after fading out, could be reinstated by local cooling of the eye- stalks ( 12).
Some experiments, however, failed to support the theory of the loss of rhythmicity after eyestalk ablation. Webb et al. (26) showed that in the crab Uca the rhythm of tegumentary pigment position continued after eyestalk ablation. Later, other au- thors, most notably Page and Larimer (lo), confirmed the persistence of locomotor rhythmicity after eyestalk ablation in the crayfish (4). From ablation experiments. therefore, the role of the OP as a circadian pacemaker has never been completely established.
One useful criterion to identify circadian clocks is the transfer of a rhythm by implantation of the putative pacemaker. In the early literature, there were some reports of rhythm restoration in animals without eyestalks (4). However, implant experiments require a clear-cut demonstration of a loss of rhythmicity after ablation, and since this is not the case for the eyestalk, restoration experiments are not conclu- sive. The failure to suppress locomotor rhythmicity by eyestalk ablation is one argu- ment in support of the notion of a multioscillatory system controlling circadian rhythmicity in crustaceans.
The other criterion used to establish the identity of a circadian clock at the organ level is the presence of an endogenous rhythmicity in isolation. Circadian rhythmi- city has been reported in the isolated OP, primarily in two of its functions: response of

THE CIRCADIAN SYSTEM OF CRUSTACEANS 5
FIG. I. Schematic representation of a longitudinal section of the crayfish retina. Under lightadapta- tion (left), the proximal pigment and the distal pigment are dis- persed. covering most of the ret- ina. Under dark adaptation (right), both sets of pigments are retracted. The position of both pigments can be quantitatively ex- pressed as an index, calculated in the manner shown at left. DPI, distal pigment index: PPI, proxi- mal pigment index.
MEMBRANE
retinal photoreceptors to light and neurosecretion. Since the rhythmicity in the iso- lated organ differs from that described in intact animals, both will be reviewed and compared here.
Circadian rhythmicity in the retina of intact animals. The circadian rhythm of visual sensitivity was first found in retinal photoreceptors and visual interneurons by Arechiga and Wiersma (7) using whole specimens of P. clurki and, later, P. houvieri (8) and C. n?uenas (27). The rhythm persists in P. bouvieri in DD, with a period of 22.5 h, and in LL, with a period of -26 h, thus conforming to Aschoffs law. The rhythm dampens after several weeks and is phase-shifted by light pulses. The length of the circadian period does not change within a range of temperature from 14.0 to 24.O”C.
When considering the cellular substrate of the rhythmicity at the retinal level, three possibilities have been contemplated. The amplitude of the photoreceptor response to a light pulse of a given intensity depends on the functional activities of three sets of cells. The first is the gain at the photoreceptors themselves. The second is the position of a set of intracellular dark pigment granules within the photoreceptor cells, the so-called proximal pigment (PP). In darkness these granules are retracted in the axonal end of the photoreceptors (Fig. l), thus leaving the photoreceptor surface exposed to light. Under illumination the granules are dispersed. thereby shielding the photoreceptor cells from stray light. The third is the position of another type of shielding pigment granule, the so-called distal pigment (DP), which is located inside a set of long, slender cells parallel to the photoreceptors and covering the whole length of the retina, from the cornea to the basal lamina. These pigments also are retracted under darkness or at night, and they disperse under illumination. Some years ago the proximal pigment was shown to maintain its circadian rhythm of changes in position under DD (28) and the distal pigment was shown to maintain its rhythmicity under both DD and LL (29). In some crustacean species, other pigments participate in the modulation of retinal output (75).
The physiologies of the PP and DP are quite different (30). The response of the proximal pigment to light has a much lower threshold than that of the distal pigment, the difference being large enough for there to be almost no overlap in the range of
Chronohd Int, Vol IO, No 1, 1993
6 H. ARk’HIGA ET AL.
I- z % h 05 2 h 05
2 5 2 :: E
E 0 0 24 12 24 12 24 12
f 24 12 24 12 24 Ii
TIME (HOUR OF DAY) TIME (HOUR OF DAY I
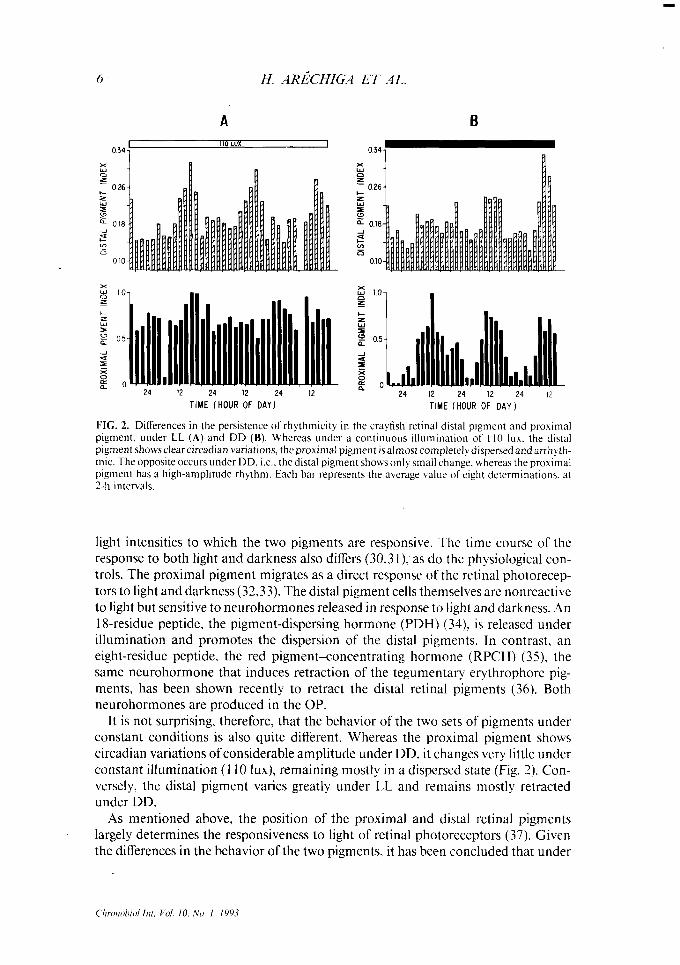
FIG. 2. Ditferences in the persistence of rhythmicity in the crayfish retinal distal pigment and proximal pigment. under LL (A) and DD (B). Whereas under a continuous illumination of I IO lux. the distal pigment showsclearcircadian variations, the proximal pigment isalmost completelydispersed and arrhyth- mic. The opposite occurs under DD. i.e.. the distal pigment shows only small change. whereas the proximal pigment has a high-amplitude rhythm. Each bar represents the average value of eight determinations. at 2-h intervals.
light intensities to which the two pigments are responsive. The time course of the response to both light and darkness also differs (30,3 l), as do the physiological con- trols. The proximal pigment migrates as a direct response of the retinal photorecep- tors to light and darkness (32,33). The distal pigment cells themselves are nonreactive to light but sensitive to neurohormones released in response to light and darkness. An 1 &residue peptide, the pigment-dispersing hormone (PDH) (34). is released under illumination and promotes the dispersion of the distal pigments. In contrast, an eight-residue peptide, the red pigment-concentrating hormone (RPCH) (35) the same neurohormone that induces retraction of the tegumentary erythrophore pig- ments, has been shown recently to retract the distal retinal pigments (36). Both neurohormones are produced in the OP.
It is not surprising, therefore, that the behavior of the two sets of pigments under constant conditions is also quite different. Whereas the proximal pigment shows circadian variations of considerable amplitude under DD, it changes very little under constant illumination (110 lux), remaining mostly in a dispersed state (Fig. 2). Con- versely, the distal pigment varies greatly under LL and remains mostly retracted under DD.
As mentioned above, the position of the proximal and distal retinal pigments largely determines the responsiveness to light of retinal photoreceptors (37). Given the differences in the behavior of the two pigments, it has been concluded that under
THE CIRCADIAN SYSTEM OF CRUSTACEANS
0 NORMALLY PIGMENTED LA)
A OPTIC NERVE CUT
0 “SILVER EYED” (Cl
I
a
, Lo 9 21 9
TIME (HOUR OF DAY)
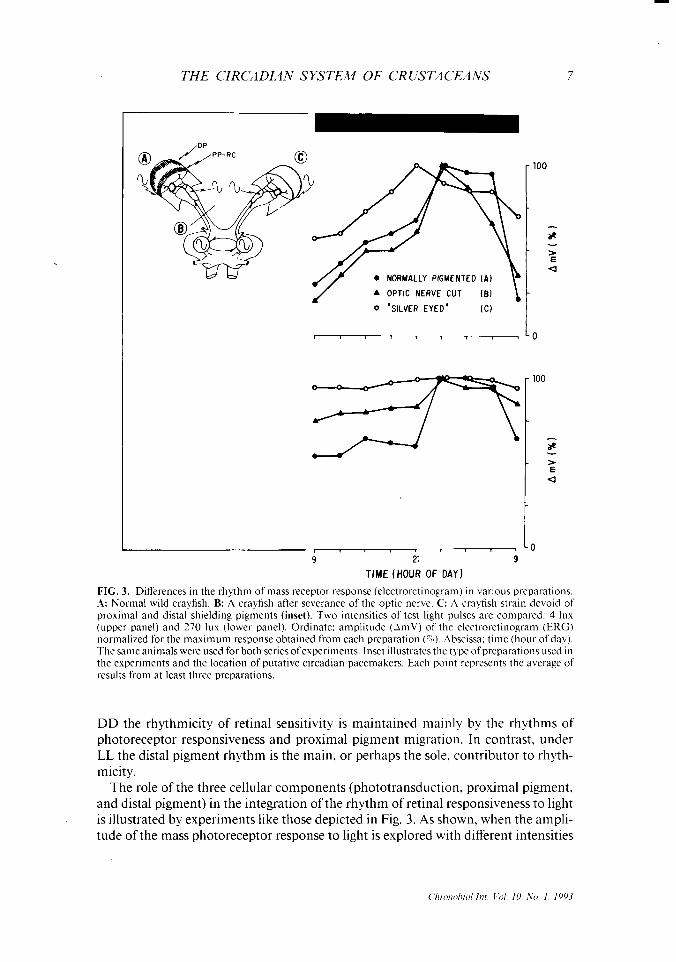
FIG. 3. Differences in the rhythm of mass receptor response (electroretinogram) in various preparations. A: Normal wild crayfish. B: A crayfish after severance of the optic nerve. C: A crayfish strain devoid of proximal and distal shielding pigments (inset). Two intensities of test light pulses are compared: 4 lux (upper panel) and 270 lux (lower panel). Ordinate: amplitude (AmV) of the electroretinogram (ERG) normalized for the maximum response obtained from each preparation (%). Abscissa: time (hour ofday). The same animals were used for both series ofexperiments. Inset illustrates the type of preparations used in the experiments and the location of putative circadian pacemakers. Each point represents the average of results from at least three preparations.
DD the rhythmicity of retinal sensitivity is maintained mainly by the rhythms of photoreceptor responsiveness and proximal pigment migration. In contrast, under LL the distal pigment rhythm is the main, or perhaps the sole, contributor to rhyth- micity.
The role of the three cellular components (phototransduction, proximal pigment. and distal pigment) in the integration of the rhythm of retinal responsiveness to light is illustrated by experiments like those depicted in Fig. 3. As shown, when the ampli- tude of the mass photoreceptor response to light is explored with different intensities

H. ARECHIGA ET AL.
of test light pulses, the rhythmicity is quite different. Three experimental conditions are compared: wild control crayfish; a crayfish in which the optic nerve has been severed; and a mutant strain (silver-eyed) of crayfish, devoid of both proximal and distal retinal pigments (40). With test light pulses of low intensity (4 lux, upper panel) there is a clear-cut rhythm of responsiveness. At a higher intensity of test light pulses (270 lux, lower panel), the rhythm in the wild control animals remains quite clear. while those.in the mutant strain and in the one with a severed optic nerve are of negligible amplitude. Presumably, the responses are near saturation at the higher intensity of the test light pulses; only the distal pigment mechanism is acting, while
_ both the photoreceptor and the proximal pigment are saturated. The fact that the rhythm persists after severance of the optic nerve partly supports the notion of an independent rhythm-generating mechanism, as illustrated in the inset.
In summary, it seems clear that under DD the main contributor to the circadian rhythmicity in the retina is the mechanism controlling photoreceptor activity and proximal pigment migration, whereas under LL the distal pigment mechanism is the chief contributor to rhythmicity.
Circadian rhythmicity in isolated eyestalk and retina. The circadian rhythm of light responsiveness has been confirmed in isolated eyestalks kept in organ culture for 15 days (38) and even in isolated retinas (39). Under DD, the rhythm in the photorecep- tor of the light response persists for several cycles, but its amplitude is less than that in the intact animal. Of the three cellular components, only the rhythms inherent in the retinal photoreceptors (i.e., the phototransduction and the proximal pigment posi- tion) are maintained, thus explaining why the rhythm of light responsiveness persists only in DD and with a much smaller amplitude than in the intact animal. The lack of rhythmicity of the distal pigment in the isolated retina isnot surprising given the fact that these pigment cells are only targets for hormones whose cells of origin are outside the retina. The lack of distal pigment rhythmicity in isolated eyestalks, in turn, may be artifactual, since the secretion of neurohormones driving this set of pigments continues in the isolated eyestalk, as we will show later. Presumably, what is lacking after isolation is the diffusion pathway from the neurosecretory cells to the retina.
Another limitation of the studies in the isolated organ is related to the survival of the preparation, which so far has been limited to 1 week. Consequently, experiments showing in vitro entrainment are still inconclusive.
Given the persistence of the rhythms of electroretinogram amplitude and proximal pigment migration in isolated eyestalks and retinas, as mentioned above, various possibilities arise with regard to the cellular origins of such rhythms. It is possible that the photoreceptors themselves are capable of generating the rhythm: however, no direct evidence is yet available. Moreover, the isolated retina contains, besides the photoreceptors, the various cells in the lumina gunglionaris (41), where efferent axons also have been noted in close apposition to photoreceptors (42). Further work is clearly needed to locate the cells responsible for the generation of retinal rhyth- micity.
Circadian rhythm of neurosecretion. The origin of the neurohormones participat- ing in the control of circadian rhythms has been studied for several decades. In the early literature, the optic peduncle was found to contain most of these substances (43). As mentioned above, several crustacean circadian rhythms are known to exist in
Chmnohd Int. Vol. IO, No. 1. 1993
THE CIRCADIAN SYSTEM OF CRUSTACEANS 9
40
35 1 FIG. 4. Changes of neurodepress- 30-
ing hormone (NDH) content in isolated eyestalks of crayfish, kept ; 25.
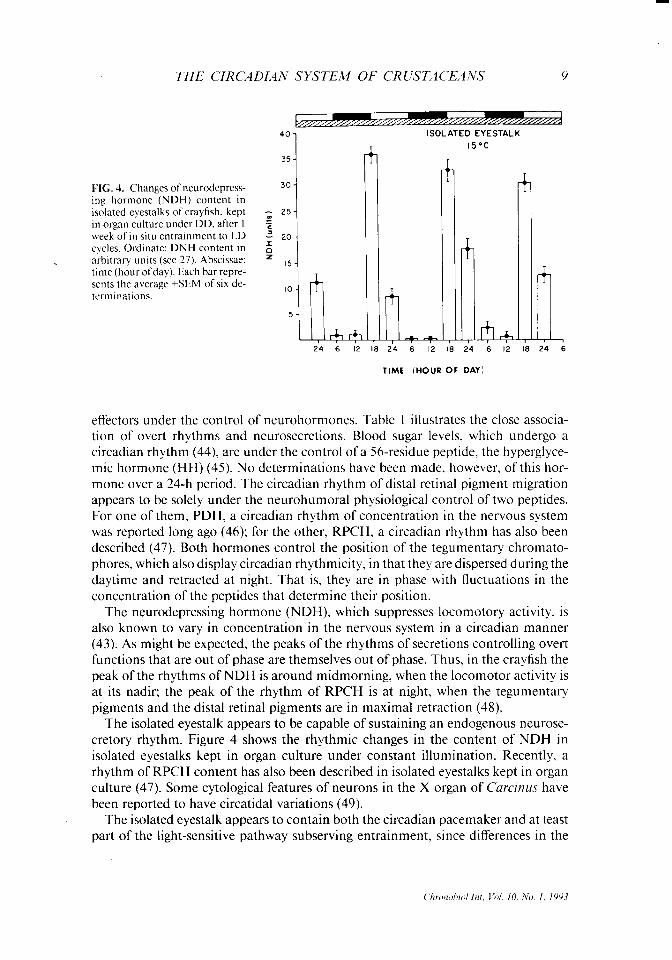
in organ culture under DD. after I “, week of in situ entrainment to LD = 20. cycles. Ordinate: DNH content in z arbitrary units (see 27). Abscissae: z ,s _ time (hour ofday). Each bar repre- sents the average GEM of six de- terminations.
Al+ 24 6 12
-4
l-
I ISOLATED EYESTALK
15oc
I 18 24 6 I2 1 I8 24 6 12 18 24 6
TIME (HOUR OF DAY)
effecters under the control of neurohormones. Table 1 illustrates the close associa- tion of overt rhythms and neurosecretions. Blood sugar levels. which undergo a circadian rhythm (44), are under the control of a 56-residue peptide. the hyperglyce- mic hormone (HH) (45). No determinations have been made, however, of this hor- mone over a 24-h period. The circadian rhythm of distal retinal pigment migration appears to be solely under the neurohumoral physiological control of two peptides. For one of them, PDH, a circadian rhythm of concentration in the nervous system was reported long ago (46); for the other, RPCH, a circadian rhythm has also been described (47). Both hormones control the position of the tegumentary chromato- phores, which also display circadian rhythmicity, in that they are dispersed during the daytime and retracted at night. That is, they are in phase with fluctuations in the concentration of the peptides that determine their position.
The neurodepressing hormone (NDH), which suppresses locomotory activity, is also known to vary in concentration in the nervous system in a circadian manner (43). As might be expected, the peaks of the rhythms of secretions controlling overt functions that are out of phase are themselves out of phase. Thus, in the crayfish the peak of the rhythms of NDH is around midmorning, when the locomotor activity is at its nadir: the peak of the rhythm of RPCH is at night, when the tegumentary pigments and the distal retinal pigments are in maximal retraction (48).
The isolated eyestalk appears to be capable of sustaining an endogenous neurose- cretory rhythm. Figure 4 shows the rhythmic changes in the content of NDH in isolated eyestalks kept in organ culture under constant illumination. Recently, a rhythm of RPCH content has also been described in isolated eyestalks kept in organ culture (47). Some cytological features of neurons in the X organ of C’urcinus have been reported to have circatidal variations (49).
The isolated eyestalk appears to contain both the circadian pacemaker and at least part of the light-sensitive pathway subserving entrainment, since differences in the
10 H. ARkHIGA ET AL.
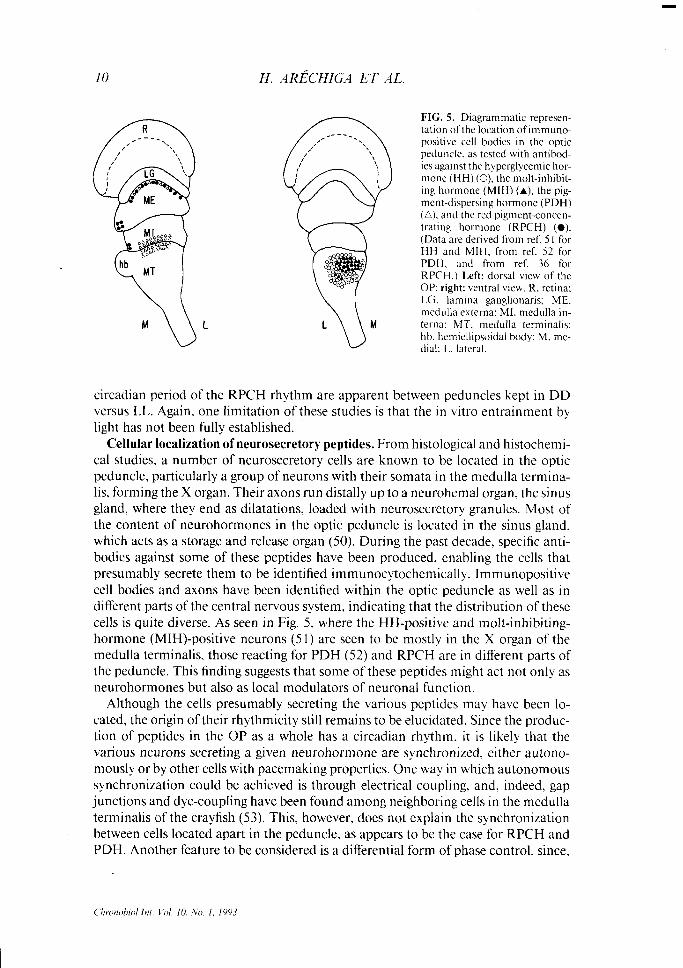
FIG. 5. Diagrammatic represen- tation ofthe location of immuno- positive cell bodies in the optic peduncle. as tested with antibod- ies against the hyperglycemic hor- mone (HH) (0), the molt-inhibit- ing hormone (MIH) (A), the pig- ment-dispersing hormone (PDH) (A). and the red pigment-concen- trating hormone (RPCH) (0). (Data are derived from ref. 5 1 for HH and MIH, from ref. 52 for PDH. and from ref. 36 for RPCH.) Left: dorsal view of the OP: right: ventral view. R. retina: LG. lamina ganglionaris; ME. medulla externa: MI. medulla in- terna: MT. medulla terminalis: hb. hemiellipsoidal body: M. me- dial: 1.. lateral.
circadian period of the RPCH rhythm are apparent between peduncles kept in DD versus LL. Again, one limitation of these studies is that the in vitro entrainment by light has not been fully established.
Cellular localization of neurosecretory peptides. From histological and histochemi- cal studies, a number of neurosecretory cells are known to be located in the optic peduncle, particularly a group of neurons with their somata in the medulla termina- lis, forming the X organ. Their axons run distally up to a neurohemal organ, the sinus gland, where they end as dilatations, loaded with neurosecretory granules. Most of the content of neurohormones in the optic peduncle is located in the sinus gland, which acts as a storage and release organ (50). During the past decade, specific anti- bodies against some of these peptides have been produced, enabling the cells that presumably secrete them to be identified immunocytochemically. Immunopositive cell bodies and axons have been identified within the optic peduncle as well as in different parts of the central nervous system, indicating that the distribution of these cells is quite diverse. As seen in Fig. 5, where the HH-positive and molt-inhibiting- hormone (MIH)-positive neurons (5 1) are seen to be mostly in the X organ of the medulla terminalis, those reacting for PDH (52) and RPCH are in different parts of the peduncle. This finding suggests that some of these peptides might act not only as neurohormones but also as local modulators of neuronal function.
Although the cells presumably secreting the various peptides may have been lo- cated, the origin of their rhythmicity still remains to be elucidated. Since the produc- tion of peptides in the OP as a whole has a circadian rhythm, it is likely that the various neurons secreting a given neurohormone are synchronized, either autono- mously or by other cells with pacemaking properties. One way in which autonomous synchronization could be achieved is through electrical coupling. and, indeed, gap junctions and dye-coupling have been found among neighboring cells in the medulla terminalis of the crayfish (53). This, however, does not explain the synchronization between cells located apart in the peduncle, as appears to be the case for RPCH and PDH. Another feature to be considered is a differential form of phase control, since,

THE CIRCADIAN SYSTEM OF CRUSTACEANS II
as mentioned above, some peptides are secreted with a diurnal acrophase and others with a nocturnal one.
The intracellular mechanisms underlying the rhythm of secretion are still to be analyzed. There is the possibility of a rhythm of biosynthesis as well as a rhythm of axonal transport and release-for example, in the crayfish X organ-sinus gland sys- tem (54). The electrical activity in neurosecretory cells in the optic peduncle has been studied, and light responses have been reported. Some neurons become excited and some inhibited by illumination of the retina (55). Various patterns of electrical activ- ity have been described in X organ cells of crabs (56) and crayfish (57). However, no systematic analysis of the rhythm of electrical activity or release from neurosecretory cells has been performed. The biosynthesis of some peptides has been shown to vary during the 24 h. Such is the case for RPCH (44) and NDH (43). For both peptides, the rhythm has been documented in intact animals and isolated eyestalks.
The same neuropeptides found in the optic peduncle have also been identified in other regions of the central nervous system (CNS). This has been established both by direct measurements ofcontent and by immunocytochemical localization ofneurose- cretory cells. The presence of neurosecretory cells in the various central ganglia of several crustacean species poses interesting questions about their physiological role. One possibility is that a given neuropeptide may act as a local neuromodulator. besides functioning as a neurohormone. There are indications, however, of peptide release into the circulation even after bilateral eyestalk ablation (58). These findings help explain the failure of eyestalk ablation to suppress the overt rhythm in the target organs of neurosecretion (26). They also add one more dimension to the problem of determining the synchronization of secretion in cells producing the same peptide in the intact animal.
The Supraesophageal Ganglion (SOG) as a Circadian Pacemnker
One obvious property of overt circadian rhythms is synchrony in the rhythmicity ofthe effecters displaying a given rhythm. In some instances, the final stage ofphysio- logical control involves the release of a neurohormone into the bloodstream, as is the case with metabolic rhythms or with rhythms affecting pigmentary effecters. There- fore, a neuroendocrine integration ofthe circadian rhythm is a necessary assumption. On the other hand, assuming the optic peduncle could function as a circadian pace- maker, it still remains to be explained how the rhythms in the two optic peduncles are synchronized. Both peduncles are connected to the SOG via the optic nerves. The SOG is a likely contributor to the integration of circadian rhythmicity.
Indeed, by severing the descending pathways from the SOG to the thoracic ganglia (where the motoneurons to the legs are located), Page and Larimer (59) were able to suppress the locomotor circadian rhythm; severing the optic nerve resulted in a unilat- eral suppression of the rhythm of retinal responsiveness to light (60). Arechiga et al. (8) showed that ablation of the SOG nullified the rhythm of retinal responsiveness to light. Soon after, however, Barrera-Mera found that the rhythm of retinal receptor responsiveness was still present after ablation of the SOG, provided that it was ex- plored with dim test light pulses (61). This observation was confirmed by Larimer and Smith (62) and it has led to the hypothesis (Fig. 3) that ablation of the ganglion

12 H. ARECHIGA ET AL
destroys the component of the rhythm dependent on the distal pigment while pre- serving the components dependent on the proximal pigment and the retinal photore- ceptors that appear capable of generating their own circadian signals. However, even for these components, a mechanism for the synchronization of the rhythm in the two retinas has to be postulated. Indeed, the rhythm in the proximal pigment in the isolated retina is of much smaller amplitude than that in the intact animal (39) particularly the retraction at night, which under certain conditions may not even appear in isolated retinas (Frixione, unpublished observations).
Two possible pathways for the bilateral synchronization of the retinal rhythms have been suggested. A hormonal pathway quite likely exists in the intact animal, since both shielding pigments are responsive to such blood-borne agents as PDH, RPCH, and 5-hydroxytryptamine. Axonal coupling is a second possibility suggested by several lines of evidence, for example, the presence of axons running from one side of the SOG to the other and of efferent fibers in the optic nerve and of efferent fibers reaching the retina and making appositional contacts with the axons of the photore- ceptors. Some of these axons appear to contain serotonin (42).
There is also physiological evidence supporting an axonal coupling between the rhythms in the two retinas. The longitudinal bisection of the ganglion results in the loss of phase-coupling between these rhythms (6 I). Since the rhythm in each retina persists and is quite robust, these observations imply a mutually entrained bilateral system of circadian oscillators (63) as represented in the inset of Fig. 3. This differs from the possible desynchronization between circadian cellular pacemakers in a sin- gle eyestalk, which presumably underlies the gradual damping out of the rhythmicity under constant conditions (4).
The cellular elements responsible for rhythm generation in the SOG are still to be located. Different sites have been proposed. Whereas Page and Larimer (59,60) pos- tulated the olfactory lobe in the tritocerebral part of the crayfish SOG as the site of the elements generating the rhythm, Barrera-Mera (6 1) has proposed a more rostra1 site, in the protocerebral part of the ganglion. An attempt to identify the axons conveying the circadian time signal from the SOG to the optic peduncle was made by Larimer and Smith (62) who placed selective lesions in the optic nerve. The central part of the nerve was found to contain these axons, but no individual fibers could be identified. In some cases there was also a lesion to the main blood vessel that runs in the central part of the optic nerve. No attempts to explore possible circadian rhythms in isolated SOG in organ culture have been reported.
Other Possible Sites of Origin gf Circadian Rh~Yhmicity
From experiments in isolated ganglia of the crayfish, the spontaneous activity in some identified motoneurons has been shown to vary during the 24 h, with a noctur- nal peak (64). However, no further characterization has been made of the origin of this rhythmicity. Figure 6 shows a multichannel recording of activity in one identi- fied motoneuron, one of the superficial abdominal flexor neurons (F5) in the second abdominal ganglion (65). The recording was done during the first 24-h period after

THE CIRCADIAN SYSTEM OF CRUSTACEANS 13
6 I6
FIG. 6. Twenty-four-hour changes in the spontaneous firing rate of an identified motoneuron (F-5) in the second abdominal ganglion ofthe isolated abdominal chain ofcrayfish. A: Diagrammatic representation of the location of cell bodies of the six superficial flexor motoneurons in the ganglion (from 65): their axons run together in the superficial branch (SB) ofthe third root. Their activity can be simultaneously recorded and separated, as shown next to the diagram, with a 2.min sample display ofthe separate activity in each of the six motoneurons, from the one generating the smallest spikes (Fl, left) to that firing the largest spikes (F6. right). The height of the records indicates the total number of spikes fired by each neuron. As seen F5 and F4 are the most active. The amplitude of each peak represents the number of action potentials generated at hourly intervals. B: Samples of I-h duration taken at different clock times (indicated in each panel); hence. the spread of spike heights for a given neuron over the 24 h. Center: the waveform of the spike. Cal. 1 mV and 1 ms for spike shown at center of figure.
isolating the ganglion. No long-term recordings are available on identified neurons to establish their possible role as circadian pacemakers.
Direct responses to illumination have been reported in motoneurons of abdominal ganglia (66). As mentioned above, neurons have been found, by immunocytological techniques, in the various ganglia, which contain some of the neuropeptides whose concentrations in the eyestalk and hemolymph are known to vary in a circadian manner. It will be interesting to explore whether the ganglia containing these ele- ments display rhythmicity in isolation.
Identification of the cells generating the various overt rhythms remains to be done. From the evidence mentioned, the anatomical segregation of cells with circadian pacemaker properties seems clear. The specific elements participating in the control of the various rhythms remain to be elucidated, as do the mechanisms by which the output of the various pacemakers is integrated to generate the overt rhythms. This issue will be discussed herein.

14 H. ARECHIGA ET AL.
The Entrainment Pathways
There is scant information about the receptors and entrainment pathways for circadian rhythmicity in crustaceans. Several sources of entrainment have been noted. As might be expected, light is the most prominent entraining agent. For the locomotory rhythm of P. cfurki, Page and Larimer ( 10) showed that the photorecep- tors participating in the entrainment are not located in the retina but in the central nervous system, particularly in the SOG (67). More recently, Fuentes-Pardo and Inclan-Rubio (68) ascertained in P. bouvieri that local illumination of the sixth ab-
c dominal ganglion is capable of phase-shifting the rhythm of electroretinogram ampli- tude. It is interesting to note in this regard that the release of PDH from the crayfish eyestalk is triggered by a neuroendocrine reflex. The reflex is initiated by light acting on extraretinal photoreceptors that are widely distributed in the CNS and that have a different spectral sensitivity from that of retinal rhodopsin (48).
Other sources of entrainment also have been described, as in the example of hydro- static pressure for the entrainment of the circatidal rhythm of locomotion in the crab Curcinus maenas (11). The receptors and pathways for this entrainment have not been characterized. Entrainment by food also has been observed recently for the locomotory rhythm of P. clarki (9). The origin of the entrainment appears to be the surface chemoreceptor neurons. No anatomical or physiological studies have been made of the elements subserving the entrainment. Since the peaks of locomotor activity entrained by mechanical or chemical inputs have phases and periodicities different from those of the LD-entrained rhythms, it is likely that a physiological segregation of mechanisms exists in the CNS such that different entrainment path- ways provide inputs to different circadian pacemakers. .
Internal Coupling Mechanisms
Analysis of the mechanisms by which the internal coupling of rhythmic influences is achieved is one of the least-known aspects of the physiology of circadian rhythms. For crustaceans, evidence suggests that the physiological mechanisms by which the internal coupling of the various circadian rhythms is organized are multiple and indirect. Little is known about their nature and interplay; however, some informa- tion is available on possible coupling pathways. Most behavioral activities of noctur- nal crustaceans in their normal habitat take place during the twilight hours. This has been related to a circadian rhythm of visual responsiveness to light, coupled to a rhythm of locomotion (69). In Nephrops norvegicus under field observation, two peaks of activity have been detected, one after dawn and another near dusk. It has been hypothesized that a dual response to light is the underlying factor (69). At the low intensities of illumination prevailing at twilight, the response to light serves as an attraction. At the high intensities around midday, light inhibits locomotion. This has been shown recently to be the case in P. clurki. Under controlled laboratory condi- tions, locomotor activity is enhanced by dim light, and the animals walk toward the light source. It is inhibited at higher intensities (above 10 lux for dark-adapted ani- mals) (74). The latency of both attraction and withdrawal responses is a function of the intensity of illumination. It is also shorter during the night than in the daytime. As
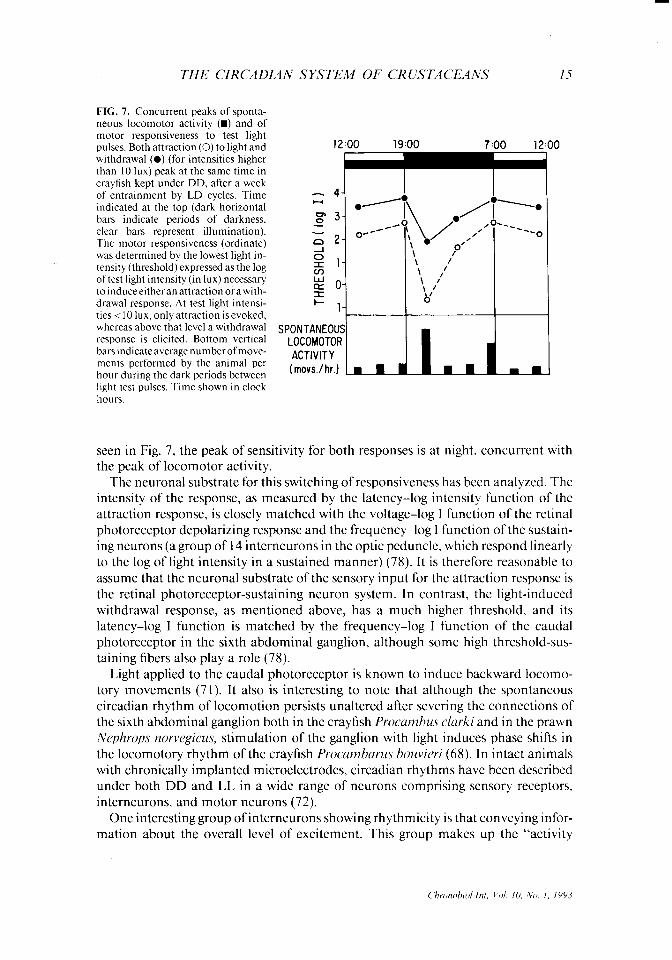
THE CIRCADIAN SYSTEM OF CRUSTACEANS 15
FIG. 7. Concurrent peaks of sponta- neous locomotor activity (m) and of motor responsiveness to test light pulses. Both attraction (0) to light and withdrawal (0) (for intensities higher than IO lux) peak at the same time in crayfish kept under DD, after a week of entrainment by LD cycles. Time indicated at the top (dark horizontal bars indicate periods of darkness, clear bars represent illumination). The motor responsiveness (ordinate) was determined by the lowest light in- tensity (threshold) expressed as the log of test light intensity (in lux) necessary toinduceeitheranattractionorawith- drawal response. At test light intensi- ties i 10 lux, only attraction is evoked, whereas above that level a withdrawal response is elicited. Bottom vertical bars indicate average number of move- ments performed by the animal per hour during the dark periods between light test pulses. Time shown in clock hours.
SPONTANEOU! LOCOMOTOR ACTIVITY (movs./hr.) .B fl
seen in Fig. 7, the peak of sensitivity for both responses is at night. concurrent with the peak of locomotor activity.
The neuronal substrate for this switching of responsiveness has been analyzed. The intensity of the response, as measured by the latency-log intensity function of the attraction response, is closely matched with the voltage-log I function of the retinal photoreceptor depolarizing response and the frequency-log I function of the sustain- ing neurons (a group of 14 interneurons in the optic peduncle, which respond linearly to the log of light intensity in a sustained manner) (78). It is therefore reasonable to assume that the neuronal substrate of the sensory input for the attraction response is the retinal photoreceptor-sustaining neuron system. In contrast, the light-induced withdrawal response, as mentioned above, has a much higher threshold, and its latency-log I function is matched by the frequency-log I function of the caudal photoreceptor in the sixth abdominal ganglion, although some high threshold-sus- taining fibers also play a role (78).
Light applied to the caudal photoreceptor is known to induce backward locomo- tory movements (71). It also is interesting to note that although the spontaneous circadian rhythm of locomotion persists unaltered after severing the connections of the sixth abdominal ganglion both in the crayfish Procamhs clurki and in the prawn Nephrops norvegicus, stimulation of the ganglion with light induces phase shifts in the locomotory rhythm of the crayfish Procamharus houvieri (68). In intact animals with chronically implanted microelectrodes, circadian rhythms have been described under both DD and LL in a wide range of neurons comprising sensory receptors, interneurons, and motor neurons (72).
One interesting group of interneurons showing rhythmicity is that conveying infor- mation about the overall level of excitement. This group makes up the “activity

I6 H. ARECHIGA ET AL.
neurons” of Wiersma and Yamaguchi (cf. 30). Their spontaneous firing rate is much higher during the night than in the day (7). Their output influences both sensory and motor neurons, and it is of a facilitory nature, i.e., these neurons enhance the sponta- neous activity and the responsiveness of both sensory and motor neurons (7). They are plausible candidates for the role of exerting a modulatory phasic control on sensory input and reflex activity. Some command interneurons also have been specifi- cally related to the control of circadian rhythmicity by Larimer and Smith (62). The excitability of the medial giant axon, a command interneuron for withdrawal reflexes in the crayfish, is higher at night than in the day (R. Aceves unpublished data).
As for the neuronal elements subserving locomotion in crustaceans, a great num- ber of identified neurons have been established and their dynamic properties have been studied. However, the mechanisms controlling their activity on a circadian basis are still unknown. The motoneurons involved in locomotion are under the modula- tory influence of neurohumoral agents. Amines and peptides are known to enhance or suppress their activity (50,73). Of particular interest in this regard is the neurode- pressing hormone produced mainly in the eyestalk. This hormone suppresses the nocturnal peak of locomotion under DD as well as the spontaneous activity in moto- neurons, both in intact animals and in isolated ganglia (43,50).
In brief, although some valuable information is now available about the neuronal substrate of internal coupling, we are still far from a comprehensive model of the interplay among the different elements.
REFERENCES
I. Page TL. Neural and endocrine control of circadian rhythmicity in invertebrates. In: Aschoff J. ed. Hundhook of hc~haviorul nezrrohro/o,qy New York: Plenum Press.’ 198 1: 145-8 I
2. De Coursey P. Biological timing. In: Vernberg J. Vernberg WB. eds. Bio/o~~~ of’c~r~stac~~u. vol. 7. New York: Academic Press, 1983: 107-6 1.
3. Palmer JD. Contributions made to chronobiology by studies of fiddler crab rhythms. C’hronohiol Int 1991:x:1 10-30.
4. Arechiga H, Naylor E. Endogenous factors in the control of rhythmicity in decapod crustaceans. In: de Coursey PJ, ed. Biological rhylhms in the marine rnl’ironmc’nf. Columbia, South Carolina: University of South Carolina Press, 1976: l- 16.
5. Dowse HB. Ringo JM. The search for hidden periodicities in biological time series revisited. J IVwro- hid 1989; 139:487-5 15.
6. Brown FA. Physiological rhythms, In: Waterman TH. ed. The physiologic of'cmstucru New York: Academic Press, 196 1:401-30.
7. Arechiga H, Wiersma CAG. Circadian rhythm of activity in the crayfish visual units. .I Nwrohid 1969;1:71-85.
8. Arechiga H, Fuentes B, Barrera B. Circadian rhythm of responsiveness in the visual system of the crayfish. In: J Salanki, ed. Nenrohiology of invertchrurcs. Budapest: Publishing House of the Hungar- ian Academy of Sciences, 1973:403-26.
9. Fernandez de Miguel F, Arechiga H. Entrainment by food of circadian locomotor activity rhythm in the crayfish. Sot Ncwosci Abstr I988;14: 1299.
IO. Page TL, Larimer JL. Entrainment ofthe circadian locomotor activity rhythm in the crayfish. JC‘omp Physiol 1972;78:107-20.
I I. Naylor E, Atkinson RJA. Entrainment of a tidal rhythm by hydrostatic pressure. In: Sleigh M. Alister Ci. eds. The effecrs ~fpressure on living orgunisms. New York: Academic Press. 1972:395-4 15.
12. Naylor E, Williams BG. Effects ofeyestalk removal on rhythmic locomotor activity in Curcinus. Jhp Bid 1968;49:107-16.
13. Webb HM. Diurnal variations in response to light in the fiddler crab, Uea. Ph~:sio/ Zoo1 1950;23:3 IO- 37.
14. Stevens CC. Influence of temperature fluctuations on the diurnal melanophore rhythm of the fiddler crab, Ucu. Physiol Zoo1 1957;30:55-69.
C‘hronohd Int. Vol. IO, No. I, 1993

THE CIRCADIAN SYSTEM OF CRUSTACEANS
15. Enright JT. Resetting a tidal clock: a phase-response curve for E.wirolana. In: De Coursey PJ, ed. Bioloeicul rhvthm.s in the rnurinr environment. Columbia, South Carolina: Universitv of South Caro- lina Press, 1976:103-14.
16. Naylor E, Williams BG. Phase-responsiveness of the circatidal locomotor activity rhythm of Ilemr- grupws edwrd.si (Hilgendorg) to simulated high tide. J ,Warine Biol A woe VK 1984:64:8 I - 150.
17. Brown FA, Webb HM. Temperature relations of an endogenous daily rhythmicity in the fiddler crab. C;cu. Ph.t,siol Zoo1 I948;2 I:27 I-8 1.
18. Stevens CC. Responses ofthe diurnal melanophore rhythm of Cc,u pu,qu.\- to changes in temperature. BIO/ Bull 1955;109:352.
19. Palmer JD. Comparative studies of circadian locomotory rhythms in four species of terrestrial crabs. .Im jMiddl Nut 1971;85:97-107.
20. Fuentes-Pardo B, In&n-Rubio V. Correlation between motor and electroretinographic circadian systems in the crayfish Procurnbutxs bozwieri (Ortmann). (‘o,np Biochcm Ph),siol 198 1:68A:477-85.
2 I. Palmer JD, Williams BG. Comparative studies oftidal rhythms. III. Spontaneous splitting ofthc peaks of crab locomotory rhythms. Marine Behuv P/IJl.sio/ 1987:13:63-75.
22. Reid DC, Naylor E. Entrainment of bimodal circatidal rhythms in the shore crab Curcinus muenu.s. J Biol Rhythms 1990:5:333-47.
23. Brown FA Jr. Webb M. Studies ofthe daily rhythmicity of the fiddler crab Uca. Modifications by light. PI~~:siol Zoo/ 1949;22: 136-48.
24. Kalmus H. Uber einen latenten physiologischen Farbwechel beim Flusskrebses. Potamobius astacus. sowie seine hormonale Beeinflussung. Z vergleich P/I),.siol 1938:25:784-97.
25. Kalmus H. Das Aktogram des Flusskrebses und seine Beeinflussung durch organextrakte. Z vcrgleich Phl:sio/ 1938:25:798-802.
26. Webb HM, Bennett MF, Brown FA. Persistence ofan endogenous diurnal rhythmicity in eyestalkless C’cu pugilufor. Biol Bull 1954:106:37 l-7.
27. Arechiga H. Huberman A. Naylor E. Hormonal modulation of circadian neural activity in the crab Curci&s muenu.s (L.). Proc RNv Sot Lond B 1974; 1871299-3 13.
28. Bennitt R. Diurnal rhvthm in the uroximal cells of the cravfish retina. Phlxiol Zoo/ 1932:5:65-9 29. Welsh J. Diurnal rhythm of the distal pigment cells in the eyes of certain crustaceans. Proc Nut .-lad
Sci USA 1930:16:386-95. 30. Arechiga H. Modulation of visual input in the craytish. In: Hoyle G. ed. Identified neuron.\ and
behuvior in urthropod.s. New York: Plenum Press 1977:387-403. 3 I, Brycesson KP. Diurnal changes in photoreceptor sensitivity in a reflecting superposition eye. J C’or?z[l
Ph.wiolA 1986;158:573-82. 32. Olive R, Larsen ME. Brief exposure to light initiates screening pigment migration in the retinula cells
of the crayfish Procamburu.s. .I C’omp PhJ>.siol 1978; 125A:9 I-6. 33. Frixione E, Arechiga H, Tsutsumi V. Photomechanical migrations of pigment granules along the
retinula cells of the crayfish. .I Nenrohiol 1979: 10:573-90. 34. Fernlund P. Structure of a light-adapting hormone from the shrimp Pundulns horeulis Biochem
Biophys ACIU 1976:439: 17-25. 35. Fernlund P, Josefsson L. Crustacean color change hormone: aminoacid sequence and chemical synthe-
sis. Science 1972:177:173-5. 36. Rodriguez-Sosa L. Ritmo circadico de neurosecrecibn en el tallo ocular de1 crustaceo decapodo.
[Dissertation]. CINVESTAV-IPN. Mexico, 1989. 37. Rodriguez-Sosa L. Arechiga H. Range of modulation of light sensitivity by accessory pigments in the
crayfish compound eye. Vision Res 1982;22: 15 15-24. 38. Sanchez J, Fuentes-Pardo B. Circadian rhythm in the amplitude of the electroretinogram in the
isolated eyestalk of the crayfish. Camp Biochem PhJ’sio/ 1977:56A:60 l-9. 39. Noguerbn I, Arechiga H. Ritmo circadico de sensibilidad a la luz en la retina aislada del acocil. Bol
E&d Med Biol Me; 1987:35:165. 40. Black JB, Huner JZ. A silver eyed mutant in the crayfish ~roc~umharw clurkii (Girard). .*l.s.soc SE Biol
Bull 1976;23:44. 4 I. Wiersma CAG, Roach J, Glantz RM. Neural integration in the optic system. In: Bliss DE, Sandeman
DC. Atwood HL. eds. The bio/oFr ofcru.stuceu. vol. 4. New York: Academic Press. 1982: l-3 1. 42. Arechiga H, Baiiuelos E, Frixione E, Picones A, Rodriguez-Sosa L. Modulation of crayfish retinal
sensitivity by 5-hydroxytryptamine. J Exp Biol 1990; 150: 123-43. 43. ArCchiga H, Huberman A. Hormonal control of circadian rhythmicity in crustaceans. In: Arechiga H.
Valverde C, eds. Frontiers in hormone rrseurch, vol. 6. Basel: Karger, 1980: 16-34. 44. Hamman A. Die neuroendokrine steverung tagesrhythmischer blutzuckerschwankungen durch die
sinusdruse beim flusskrebs. J Camp Physiol 1974;89: 197-2 14. 45. Keller R. Purification and aminoacid composition of the hyperglycaemic neurohormone from the

18 H. ARkXIGA ET AL.
46.
41.
sinus gland of OrcdM(‘(.(L’Y limo.rzls and comparison with the hormone from C’urcinlrv I~N(‘~I~.Y J C’ofnp Phr.sio/ 1981;141:445-50.
Arkchiga H. Mena F. Circadian variations of hormonal content in the nervous system ofthe crayfish. Camp Biochem Phjxiol 1975:52A:58 1-4. Rodriguez-Sosa L, Calderbn J. Hernander J. ArCchiga H. The isolated eyestalk of the crayfish main- tains a circadian rhythm of neurosecretion content in the crayfish eyestalk. Sot ,Ncwosci Ahsrr 1990:16:909.
48.
49.
-. 50.
Arechiga H, Cartes JL. Garcia U, Rodriguez Sosa L. Neuroendocrine correlates ofcircadian rhythmic- ity in crustaceans. .4/ncr Zoo/ 1985;25:265-74. Williams JA. Pullin RSV. Naylor E. Smith G, Williams BG. The role of Hanstroms organ in clock control in Curcin~rs ~r?~lerr(l.s. In: Naylor E, Hartnoll RG. eds. Cyclic phcwomcwa in murlne plants und animal.s. Oxford: Pergamon Press, 1979:459-66.
Arechiga H, Garcia U, Rodriguez-Sosa L. Neurosecretory role of crustacean eyestalk in the control of neuronal activity. In: Selverston AL ed. Model nczrrul nct~~wli.c und hclrurior New York: Plenum Press. 1985:361-79.
51.
52.
Dircksen H. Webster SC. Keller R. Immunocytochemical demonstration of the neurosecretory sys- tems containing putative moult-inhibiting and hyperglycemic hormone in the eyestalk of brachyuran crustaceans. Ccl1 Ti.s.slre Ret I988;25 I :3- 12. Mangerich S, Keller R, Dircksen H, Rao RK, Riehm JP. Immunocytochcmical localization of pig- ment-dispersing hormone (PDH) and its coexistence with FMRF-amtde immunoreactive material in the eyestalks of the decapod crustaceans Curcrnxs mue?!u.v and Orconwtrc Irmosrrs. <‘c/l Ti.ss~~ Rcc 1986:245:377-86.
53.
54.
Arechiga H. Chavez B, Glantz RM. Dye-coupling and gap-junctions between crustacean neurosecre- tory cells. Bruin Re.r 1985;326: 183-7.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
Arechiga H. Flores Loper J. Garcia U. Control of biosynthesis and release of the crustacean neurode- pressing hormone. In: Lofts B, Helms WM. eds. Clown/ trcnd.s in compurcrf~~~c cndocrinolog~~. Hong Kong: University Press. 1985:787-9 1. Glantz RM. Kirk MD. Arechiga H. Light input to crustacean neurosecretory cells. Brain Rcc 1983:265:307-II.
Nagano M, Cooke IM. Comparison of electrical responses of terminals, axons and somata of a pepti- dergic neurosecretory system. J IVc~lrosci 1988:7:634-B.
Onetti C. Garcia U. Valdiosera RF. Arechiga H. Ionic currents in crustacean neurosecretory cells. J Ne~rro~ph~~.sro/ 1990;64: I5 14-26. Arechiga H. Cabrera-Peralta C. Huberman A. Functional characterization of the neuro-depressing hormone in the crayfish. J Newohio/ 1979; 10:409-22.
Page TL. Larimer J. Neural control ofcircadian rhythmicity in the crayfish. I. The locomotor rhythm. .I Camp PhJ,.siol 1975:97:59-80.
Page TL. Larimer J. Neural control of circadian rhythmicity in the crayfish. II. The ERG amplitude rhythm. J Comp PI~wiol 1975;97:8 l-96. Barrera-Mera B. Effect ofcerebroid ganglion lesions on ERG circadian rhythm in the crayfish. Ph~~col Brha~~ 1976:11:59-64. Larimer JL, Smith JT. Circadian rhythm of retinal sensitivity in crayfish: modulation by the cerebral and optic ganglia. J Camp Pl~ysrol 1980; 136:3 13-26. Barrera-Mera B. Neural coupling between left and right electroretinographic circadian oscillations in the crayfish P Bouvieri. Camp Biochem Physisiol 1978;6 I A:427-32. Arechiga H. Circadian rhythmicity in the nervous system of crustaceans. p& Proc, 1977:36:2036-4 1.
Wine JJ, Mittenthal J. Kennedy DM. The structure oftonic flexor motoneurons in crayfish abdominal ganglia. J Camp Ph~~siol 1974;93:3 15-35.
Edwards DH. Crayfish extraretinal photoreception. I. Behavioural and motoneuron responses to abdominal illumination. J Ex~ Biol 1984; 109:29 l-306.
Page TL, Larimer J. Extraretinal photoreception in entrainment of crustacean circadian rhythms, Photochern Photohiol 19761231245-5 I.
69.
Fuentes-Pardo B, Inclin-Rubio V. Caudal photoreceptors synchronize circadian rhythms in crayfish. I. Synchronization of ERG and locomotor circadian rhythm. Camp Brochr~n Pl~!:siol 1987;86A: 523-l.
Arechiga H. Atkinson RJA. The eye and some effect of light on the locomotor activity in Nqhrop.s norvegicu.s. .Vtarine Biol 1975:32:63-76.
Chronohiol Int. Vol. 10, No 1, 1993

THE CIRCADIAN SYSTEM OF CRUSTACEANS 19
70. Glantz RM. Peripheral versus central adaptation in the crustacean visual system. J Neurophysiol 197 I ;34:485-92.
7 I. Simon TW, Edwards DH. Light-evoked walking in the crayfish: behavioral and neuronal responses triggered by the caudal photoreceptor. J Comp physiol [A] i990;166:745-55.
72. ArCchiga H. Circadian modulation of behavior in crustaceans. Neurosci Res Proa Bull 1979: 17:672-9. 73. Kraviti EA. Hormonal control of behavior: amines as gain-setting elementsthat bias behavioral
output in lobsters. Amer Zoo1 1990;30:595-608. 74. Fernandez de Miguel F, Cohen J, Zamora L, Arechiga H. An automated system for detection and
analysis of locomotor behavior in crustaceans. Bol Estud Med Biol Mex 1989;37:7 1-6. 75. Rao KR. Pigmentary effecters. In: Bliss DE, Mantel LH, eds. The biology ofcrustaceu, vol. 9. New
York: Academic Press, 1985:362-95. 76. Thurman C. Rhythmic physiological color change in crustacea: a review. Comp Biochem Ph~siol
1988;91C:171-85. 77. Beltz SB. Crustacean neurohormones. In: Laufer H, Downer RGH, eds. Invertebrate endocrinology
(endocrinology of selected invertebrate types), vol. 2. New York: Alan R. Liss, 1988:235-58.
(‘hronobrolln!, Vol. IO, No. 1, I993