hormone-like activity of humic substances in fagus sylvaticae forests
TRANSCRIPT

©
New Phytologist
(2001)
151
: 647–657
www.newphytologist.com
647
Research
Blackwell Science Ltd
Hormone-like activity of humic substances in
Fagus sylvaticae
forests
Diego Pizzeghello
1
, Gianni Nicolini
2
and Serenella Nardi
1
1
Dipartimento di Biotecnologie Agrarie, Università di Padova, Facoltà di Agraria – Agripolis, Strada Romea 16, 35020 Legnaro (Padova), Italy;
2
Centro di
Ecologia Alpina, 38040 Viote del Monte Bondone, Trento, Italy
Summary
• Soil A horizons from a large area typical for beech (
Fagus sylvatica
) in northernItaly were investigated to understand the phyto-hormone-like activity of humicsubstances (HS) with respect to the vegetation. These soils had developed underthermophilous (
Cephalonthero–Fagion
with
Carici albae–Fagetum
), mesophilous(
Fagion sylvaticae
with
Dentario pentaphylli–Fagetum
and
Galio-odourati–Fagetum
)or acidophilous (
Luzulo–Fagion
with
Luzulo niveae–Fagetum
) beech forests.• The development of organic and humic matter in the three forest types wasassessed by quantifying chemical parameters. Phyto-hormone-like activity of HSwas evaluated by measuring auxin-like (IAA
like
) and gibberellin-like (GA
like
) activityas well as IAA concentration, invertase and peroxidase activities.• Soil pH differed between the forest types. Soil C: N distinguished the thermophilousand the mesophilous from the acidophilous types but did not distinguish thermophilousfrom the mesophilous types. Phyto-hormone-like activity of HS varied significantlyacross the beech forests. Acid conditions were essential for the release of IAA
like
activity whereas neutral conditions promoted GA
like
activity. Plant isoenzymatic poly
-
morphism confirmed the different auxin activities of HS.• The phyto-hormone-like activity of HS is highly suitable, and better than soilchemical parameters, for differentiating between these ecosystems.
Key words:
humic substances, hormone-like activity, soil fertility, beech forests,
in vitro
and
in vivo
activity.
©
New Phytologist
(2001)
151
: 647–657
Author for correspondence:
S. Nardi
Tel:
+
39 0498272911
Fax:
+
39 0498272929
Email: [email protected]
Received:
6 November 2000
Accepted:
10 May 2001
Introduction
Soil organic matter (SOM) is one of the more useful indicatorsof soil fertility because it interacts with other numerous soilcomponents (Seybold
et al.
, 1998). Soil organic matter is akey component of terrestrial ecosystems and is functionallyand structurally integrated into basic ecosystem processes.Differences in SOM and its turnover are related to changes inclimate, parent rock, vegetation and to numerous complexbiological, chemical and physical soil processes.
Humic substances (HS), the largest constituent of SOM,can be considered the top rung in the stabilization of SOMand a key intermediate in the mineralization of SOM to CO
2
(Ertel
et al.
, 1988). The conservation and enhancement of SOMand HS in soils exerts beneficial effects on plant nutrientsupply (Stevenson, 1994), soil structure, compactability (Soane,1990) and water-holding capacity (DeJong
et al.
, 1983).
The term soil humus refers to a mixture of organic com-pounds produced by decomposition of plant tissue. Humus isdefined by Stevenson (1994) as the total organic fraction insoils exclusive of nondecomposed plant and animal material,their partial decomposition products, and the soil biomass.Humic substances (e.g. humic acids and fulvic acids) make upthe bulk of humus (Tan, 1998).
The biological activity of HS encompasses all the activitiesof HS in regulating plant biochemical and physiological pro-cesses, irrespective of their stimulatory or inhibitory roles.Most studies of HS influences on biochemical mechanismshave shown the influence of humic matter on membranepermeability, protein carriers of ions, activation of respirationand the Krebs cycle (Vaughan & Malcolm, 1985). There arealso effects on photosynthesis, formation of ATP, amino acids,carbohydrates and proteins, nucleic acids synthesis, and selectiveeffects on enzyme activities (Vaughan & Malcolm, 1985). This
NPH223.fm Page 647 Wednesday, August 1, 2001 9:27 AM

www.newphytologist.com
©
New Phytologist
(2001)
151
: 647–657
Research648
list is not comprehensive and does not include the possibilityof HS behaving as plant growth regulators (Bottomley, 1917;Hillitzer, 1932; Petrovic
et al.
, 1982; Cacco & Dell’Agnola,1984; Visser, 1986; Nardi
et al.
, 1996).Over the last 10 years we have investigated the phyto-
hormone-like activity of HS derived from earthworm faecesand agrarian soils (Dell’Agnola & Nardi, 1987; Muscolo &Nardi, 1997; Muscolo
et al.
, 1999), compost (Dell’Agnola
et al.
,1991; Calliari
et al.
, 1993), and forest soils (Concheri
et al.
,1996; Nardi
et al.
, 1998; Nardi
et al.
, 1999). Humic matterfrom forest soils has a very complex biological activity anddepending on its origin, molecular size, and concentrationexhibits high or low stimulations of plant metabolic parameters(Maggioni
et al.
, 1987; Albuzio
et al.
, 1993). In forest soils, littercomposition does not influence the chemical characteristicsof humic fractions greatly, as revealed by nuclear magneticresonance spectra (Nardi
et al.
, 2000a), but it does influencebiological activity and as a result forest species are affected indifferent ways by their humic substances (Nardi
et al.
, 2000a).Muscolo
et al
. (1998) revealed the presence of indoleaceticacid (IAA) in HS with high specificity monoclonal antibodiesand with different anti-indoleacetic acid antibodies. Neverthe-less, the low quantity of IAA found in HS explains the auxineffect in plant metabolism only in part and justifies comparativestudies relative to the auxin-like activity of HS. In a previouswork (Nardi
et al.
, 1988) we showed,
in vitro
, that the deliveryof the hormone-like activities in HS was influenced by theirparticular arrangement in the humic aggregate which is influ-enced by the pH of the humic extract. Acidification of thehumic extract elicited a high auxin-like activity, while basic orneutral conditions showed high gibberellin-like activity.
The aims of this study were to investigate if organic matterchemical parameters such as pH and C : N, or humic matterphyto-hormone-like activities are better able to differentiatebetween soils from different forest ecosystems. For this purposea large area typical for beech (
Fagus sylvaticae
L.) in northernItaly (Hofmann, 1991), was considered and the phyto-hormone-like activity of HS with respect to the vegetation was evaluated.The studies by Calabrese
et al
. (1996, 1998) in the Trentinoregion, which described three big groups of beech forests, thethermophilous (THE), mesophilous (MESO) and acidophilous(ACI), were used to reach our goal.
The phyto-hormone-like activity of HS was evaluated onthe basis of its effects on invertase and peroxidase activitiesin Norway spruce (
Picea abies
L. Karst.) seedlings. Invertaseactivity, which hydrolyses phloem-translocated sucrose intohexose substrate available to growing cells, is positivelycorrelated with plant growth (Kim & Suzuki, 1989). A closerelationship betwen gibberellic acid and invertase activitywas found by Kaufman
et al
. (1968) and by Kaur
et al
. (2000).Peroxidases are ubiquitous in plants and are involved, togetherwith others enzymes, in many processes such as lignification,differentiation, and growth; they have also been implicatedin organogenesis and their activities are related to IAA
(Krsnik-Rasol
et al.
, 1982). Soluble peroxidases also might beinvolved in the breakdown of hydrogen peroxide and in theoxidation of IAA (Sanchez
et al.
, 1996). Moreover, peroxidaseactivity occurs in different isoforms and may be an early indicatorof somatic embryogenesis (Krsnik-Rasol
et al.
, 1982).
Materials and Methods
Study area
The study area vegetation was investigated by Calabrese
et al
.(1996, 1998) and Zanella
et al
. (1997). Sites were chosen withthe help of the Forest Service of Autonomous Province ofTrento (north-east Italy) and by analysing 1 : 50 000 vegetationmap sheets (Pedrotti, 1981; Pedrotti, 1982; Pedrotti, 1987).To locate representative sites with respect to topography,morphology and management, information obtained fromthe Forest Planning Office was also considered. The fieldstudy allowed the most typical soils and soil cover conditionsof the beech population to be recognized. Forest populationswere only sampled if considered to be homogeneous with respectto morphological, pedological, vegetational and silviculturalcharacteristics. The statistical processing of the floristic listdata was carried out using ‘Anaphyto’, a program implementedby the Laboratoire de Biologie Végétale of Paris University.This program combines correspondence factorial analysis withcluster analysis. In the output, floristic lists are plotted both infactorial planes and on the branches of the dendrogram. Inthese diagrams, the distance between the sites is proportionalto the degree of floristic diversity and consequently, groups ofsimilar sites with respect to the flora are identified.
From the statistical processing of the floristic data, three largegroups, corresponding to key alliances of the order
Fagetaliasylvaticae
Pawl. were identified. The alliance
Cephalanthero-Fagion
with the association
Carici albae-Fagetum
, correspondingto thermophilous beech forests; the alliance
Fagion sylvaticae
with the associations
Dentario pentaphylli-Fagetum
and
Galio-odourati-Fagetum
, both corresponding to mesophilous beechforests and the alliance
Luzulo-Fagion
with the association
Luzulo niveae-Fagetum
corresponding to acidophilous beechforests (Pedrotti, 1981, 1982, 1994). The thermophilous com-munities are differentiated by
Juniperus communis
,
Cornussanguinea
,
Clematis vitalba
,
Coronilla emerus
,
Amelanchier ovalis
,
Viola mirabilis
,
Viola hirta
,
Carex humilis
,
Epipactis artrorubens
and
Polygala chamaebuxus
. The mesophilous communities aredifferentiated by
Cardamine enneaphyllos
,
Cardamine impatiens
,
Epipactis microphylla
,
Galium odouratum
,
Galium aristatum
,
Anemone trifolia
,
Paeonia officinalis
,
Lamiastrum galeobdolon
,
Polygonatum verticillatum
. The acidophilous communities aredifferentiated by
Luzula nivea
,
Luzula albida
,
Vacciniummyrtillus
,
Vaccinium vitis-idaea
.A number of soil profiles were sampled in each of the three
main types using a soil auger. The area of the sampling siteswas at least 400 square meters, as suggested by Lowe
et al
.
NPH223.fm Page 648 Wednesday, August 1, 2001 9:27 AM

©
New Phytologist
(2001)
151
: 647–657
www.newphytologist.com
Research 649
(1987). Twenty-seven typical soils were identified andsampled by excavating soil pits
c
. 1 m
2
in surface area. Thedepth of the soil pits never exceeded 1.50 m. Each profile wasdescribed morphologically during sampling and triplicatesamples for chemical and physical characterization were takenfrom each horizon. The thickness of the A horizons rangedfrom 8.5 to 65 cm for the THE sites, from 3 to 8 cm for theMESO sites and from 2.5 to 6 cm for the ACI sites. Afterscreening for large rock fragments, the soil samples wereplaced in plastic bags and transported to the laboratory. Thesoils were classified following the FAO-Unesco (1990) systemcriteria (Table 1).
To study the evolution of organic and humic matter inthe three types of beech forests only the soil A horizons werechosen, as the composition of the organic matter and thehumification process reach the last step of evolution in thishorizon (FitzPatrick, 1986). We considered as chemical andbiochemical parameters: organic carbon (OC) as an indicatorof soil fertility, the C : N ratio which indicates the proportionbetween soil mineralization and the humification processes,and the humic carbon (HC) which indicates the SOM frac-tion that reached a higher stabilization in the soil. Moreover,the molecular-weight-distribution in the humic extract wasconsidered because it was a result of the humification process(Dell’Agnola & Ferrari, 1971).
Preparation of humic substances
The humic substances were extracted from the air-dried sampleswith 0.1 M KOH (1 : 20 w/v) at room temperature for 16 hunder an N
2
atmosphere and were freed from the suspendedmaterial by centrifugation at 7000
×
g
for 20 min. Here, theterm HS is the fraction soluble in bases and comprensive ofhumic and fulvic acids. Humic extracts (50 ml) were transferredinto 18 000 mol. wt cut-off dialysis Visking tubing (MedicellLTD, London, UK) and dialysed against double-distilledwater. The water was changed daily until the liquid outsidethe dialysis tube was colourless. Subsequently the retainedsolution was desalted by ion exchange on Amberlite IR 120H
+
(Dell’Agnola & Ferrari, 1971).
Molecular-weight-distribution of humic substances
Molecular-weight-distribution and gel-permeation chromato-graphy of each humic extract was carried out on a SephadexG-100 gel packed in a 70
×
1.6 cm Pharmacia column (Pharmacia,Uppsala, Sweden). The gel packing solution and the eluentwere 0.02 M Na
2
B
4
O
7
. The apparent molecular weight of thefractions was: > 100 kDa, 100–10 kDa and < 10 kDa. Thecolumn calibration was based on previously assessed standardproteins (Kit MS-II, Serva, Heidelberg, Germany) (Dell’Agnola& Ferrari, 1971).
Phyto-hormone-like activities and indoleacetic acid content determinations
The auxin-like ( IAA
like
) and gibberellin-like (GA
like
) activitiesof the humic substances were assessed by checking thereduction in the growth of watercress (
Lepidium sativum
L.)roots and the increase in the length of lettuce (
Lactuca sativa
L.)epicotyls (Audus, 1972). Watercress and lettuce seeds weresurface-sterilized by immersion in 8% hydrogen peroxidefor 15-min. After rinsing five times with sterile distilled water,10 seeds were placed on a sterile filter paper in a sterile Petridish. For the watercress, the filter paper was wetted with1.2 ml of 1 mM CaSO
4
(control), or 1.2 ml of 20, 10, 1,0.1 mg indoleacetic acid l
−
1
(Sigma, St. Louis, MO, USA) forthe calibration curve or 1.2 ml of 10, 5, 2.5, 1, 0.5, 0.2 and0.1 mg C HS l
−
1
. For the lettuce, the experimental designwas the same as for the watercress except that the sterile filterpaper was wetted with 1.4 ml and not with 1.2 ml and thecalibration curve was a progression of 100, 10, 1, 0.1, mggibberellic acid l
−
1
(Sigma). The seeds were germinated in thedark at 25
°
C in a germination room. After 48-h for the watercressand 72-h for the lettuce, the seedlings were removed and theroot or epicotyl lengths measured (Audus, 1972).
The quantitative determination of IAA in HS was made by anenzyme linked immuno-sorbent assay (ELISA) (Phytodetek-IAA, Sigma). Tracer and Standard solutions were preparedfollowing the manufacturer’s instructions. To each well 100 µl
Table 1 Soil classification and vegetation cover of the 27 sites studied
Site Beech forest FAO classification
49 Thermophilous Skeletic-Calcaric Cambisol14 Thermophilous Skeletic-Calcaric Phaeozem217 Thermophilous Skeletic-Calcaric Phaeozem218 Thermophilous Skeletic-Calcaric Phaeozem106 Thermophilous Eutric Cambisol105 Thermophilous Calcaric Cambisol129 Thermophilous Haplic Phaeozem101 Thermophilous Calcaric Cambisol107 Mesophilous Eutric Cambisol08 Mesophilous Calcaric Cambisol109 Mesophilous Chromic Luvisol30 Mesophilous Skeletic-Calcaric Cambisol6 Mesophilous Haplic Luvisol110 Mesophilous Chromic Luvisol130 Mesophilous Luvic Phaeozem15 Mesophilous Haplic Luvisol34 Mesophilous Eutric Cambisol35 Mesophilous Skeletic-Calcaric Phaeozem4 Mesophilous Haplic Luvisol7 Mesophilous Calcaric Cambisol13 Mesophilous Albic Luvisol16 Mesophilous Haplic Luvisol112 Acidophilous Spodo-Dystric Cambisol26 Acidophilous Cambic Podzol103 Acidophilous Haplic Podzol25 Acidophilous Spodo-Dystric Cambisol12 Acidophilous Spodo-Dystric Cambisol
NPH223.fm Page 649 Wednesday, August 1, 2001 9:27 AM

www.newphytologist.com
©
New Phytologist
(2001)
151
: 647–657
Research650
of standard or humic extract and 100 µl of diluted Tracer wereadded. After an incubation at 4
°
C for 3-h, the wells weredecanted and washed by adding 200 µl of Wash solution andthen 200 µl of Substrate solution. After 60-mins at 37
°
C,50 µl of Stop reagent was added to each well and the colourabsorbance was than read at 405 nm using a 450 Biorad micro-plater reader (Biorad, Hercules, CA, USA). For each soil, a waterextract was made as a control and tested for IAA, IAA
like
, andGA
like
activity following the methods of Nardi
et al
. (2000b);no activities were detected.
Plant material
To avoid any complications of contamination by microorganismsthe plants were grown under sterile conditions. Norway spruce(
Picea abies
L. Karst.) seeds were surface-sterilized by 30-mincontact in 8% hydrogen peroxide solution. After rinsing five–six times with sterile distilled water, seeds were placed on a stain-less steel screen, kept in a steel cylinder (6.5 cm in diameter)and were covered daily with a 1-mM
-
CaSO
4
-saturated Whatmanno. 3 filter paper (Whatman, Maidstone, UK). To maintainsterility and optimal humidity, the supporting structure wasinserted into a Petri plate (9 cm in diameter). The seeds weregerminated in the dark at 25
°
C. Ten-day-old seedlings weregrown for 2 d in Hoagland’s nutrient solution (Hoagland &Arnon, 1950) added to 1.0 mg C l
−
1
of the extracted humicsubstances or with 2.0 mg l
−
1
of indoleacetic acid or 2.0 mg l−1
of gibberellic acid. Hormonal concentrations were based on theresults of previous experiments (Concheri et al., 1994), whichindicated that the concentration of all the hormones that providedmaximum stimulation for seedling growth was within therange 0.1–10 mg l−1. Seedlings were kept in a growth-chamberunder lamps giving off 100 µmol photon m−2 s−2 within a14–10 h, 24–18°C, 65–80% rh day : night cycle. A pumpwas used to bubble sterile air through the solution.
Invertase and peroxidase activities
At the end of 12th day, the Norway spruce seedlings (1 g) wereharvested, immersed in liquid N2 and homogenized (1 : 10 w/v)in 0.1 M potassium acetate buffer (pH 4.0) containing 0.1 Msucrose to determine invertase activity or in 0.1 M phosphatebuffer (pH 7.0) to test for peroxidase activity. The extracts werecentrifuged at 15 000 × g for 15-min at 2°C and the super-natants used as the enzyme source. Invertase activity wasevaluated according to Arnold (1965) and peroxidase activity asdescribed by Putter (1974). Peroxidase and invertase activitiesare expressed as a percentage of the control, where the controlswere 0.0045 optical density (OD) min−1 f. wt mg−1 and103.5 mg sucrose g f. wt−1 h−1, respectively.
Gel electrophoresis
Peroxidase isozymes were resolved on polyacrylamide gel,
according to Davis (1964), using 5% acrylamide as the stackinggel and 10% acrylamide as the separating gel in a verticalBiorad apparatus (Biorad, Hercules, CA, USA). The electrodebuffer contained 25 mM Tris and 192 mM glycine (pH 8.7).Gels were run for 2.5-h at 100 V, then peroxidase isozymes werevisualized following the Shaw & Prasad (1970) procedures.
Statistical analyses
Soil pH, organic carbon (OC), C : N ratio, humic carbon(HC), humic carbon : organic carbon ratio (HC : OC),> 100 kDa apparent molecular weight humic fraction, 100–10 kDa humic fraction, < 10 kDa humic fraction, auxin-likeactivity (IAAlike), gibberellin-like activity (GAlike), indoleaceticacid content (IAA), peroxidase activity (PEROX) and invertaseactivity (INV) data did not show the normal distributionor variance assumptions required for parametric tests. Con-sequently, the Kruskal–Wallis H-test was used and its multiplecomparison extension, a median test for ranked data, was usedin determining significant differences among pairs of foresttypes (Gibbons, 1976). Even though the assumption ofnormality was not satisfied, violation is usually acceptable,because the F-test was strong against nonnormality (Lindman,1992). For this reason, differences among forest groups(THE, MESO and ACI) were also determined using ANOVAunivariate and the Student-Newman-Keuls post hoc pairwisecomparison of means test. All statistical analyses were doneusing SPSS 8.0 for Windows (SPSS, Bologne, Italy).
Results
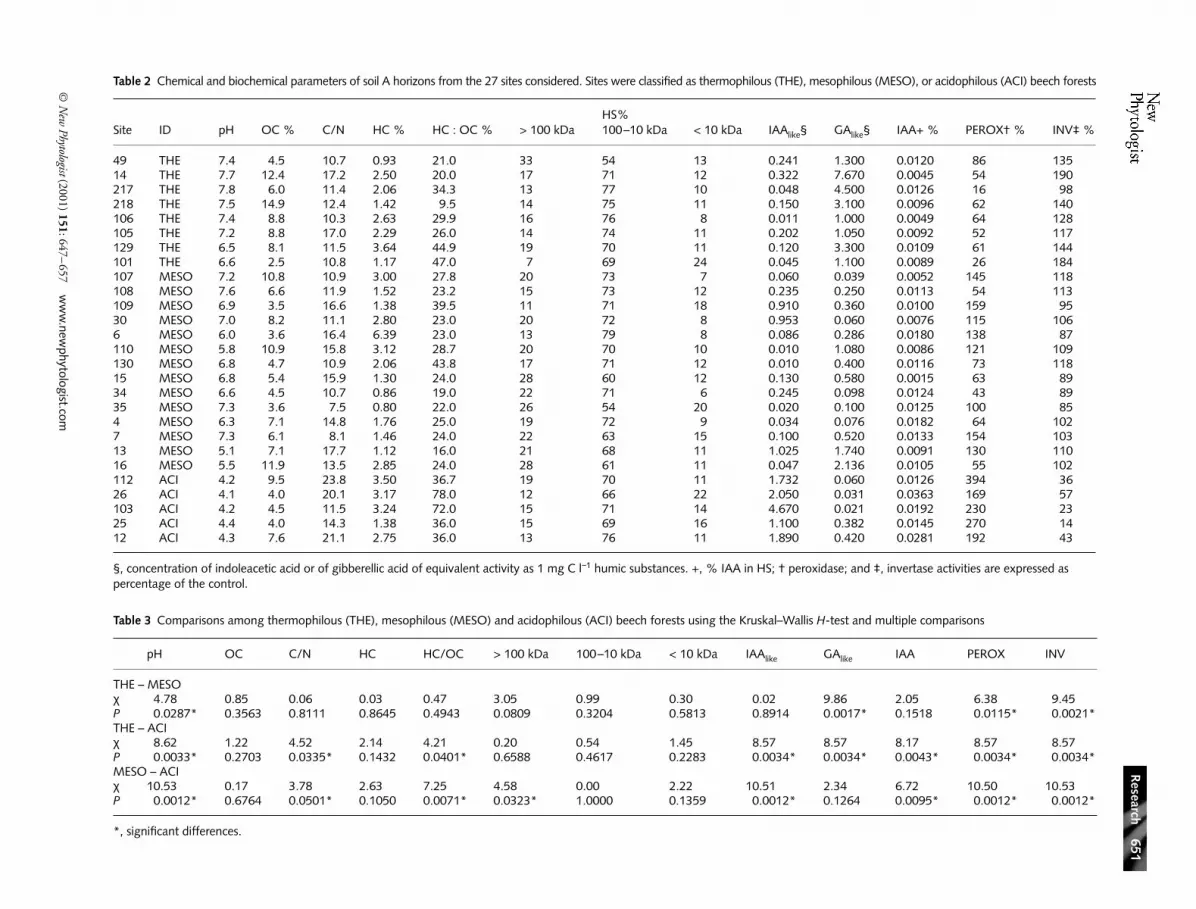
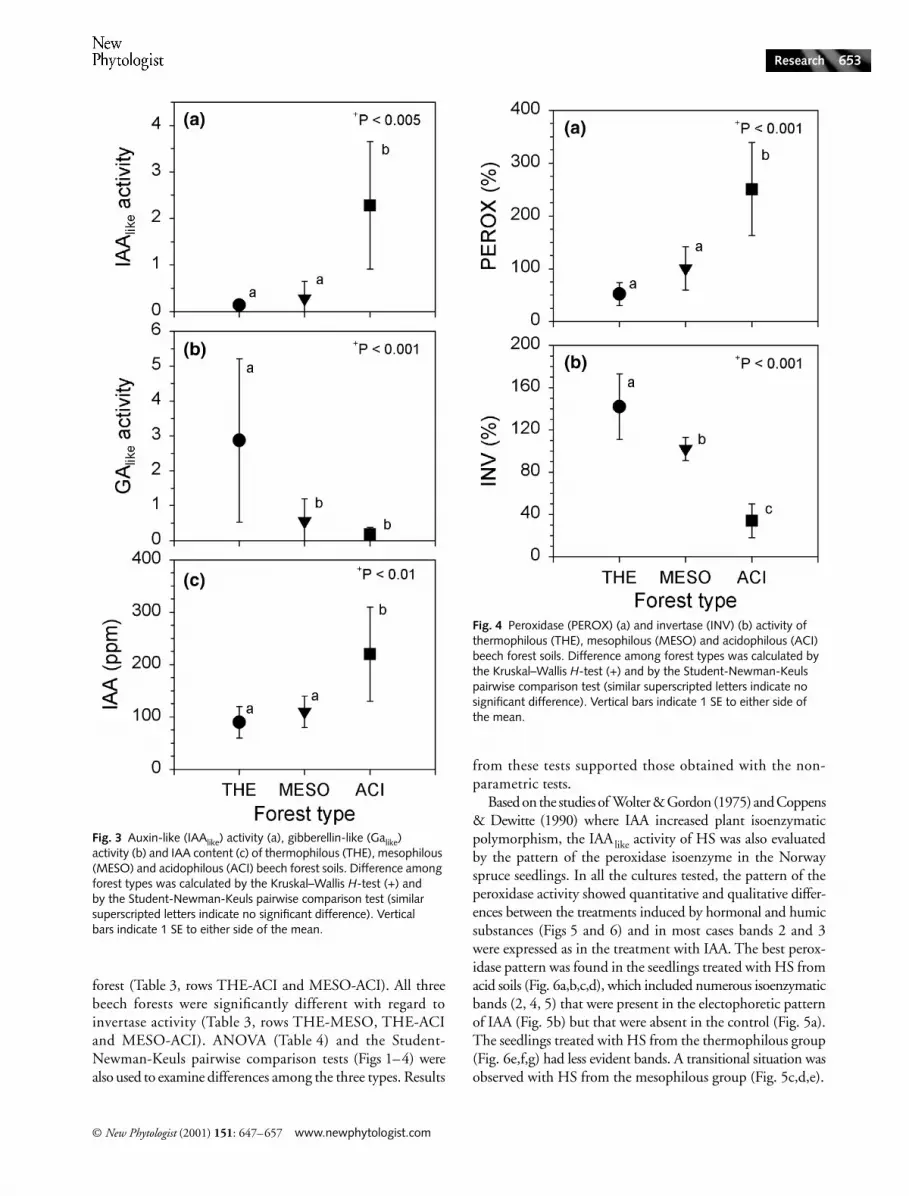
The chemical and biochemical parameters for each of the 27soils are shown in Table 2. Using the Kruskal–Wallis H-test toexamine overall differences among the three types of beechforests, only pH (Fig. 1a) and humic carbon : organic carbonratio (HC : OC) (Fig. 2b) among the chemical parametersshowed significant variation. However, the biological para-meters auxin-like activity ( IAAlike) (Fig. 3a), gibberellin-likeactivity (GAlike) (Fig. 3b), and indoleacetic acid content ( IAA)(Fig. 3C) all varied significantly across the beech forests.Peroxidase activity (Fig. 4a) and invertase activity (Fig. 4b)also showed significant variation among the forest types.
We also used the Kruskal–Wallis H-test for multiple com-parisons to examine differences among pairs of forest types.The three forest types each had different pH (Table 3, rowsTHE-MESO, THE-ACI and MESO-ACI). The C : N andHC :OC ratios did not distinguish the thermophilous fromthe mesophilous types (Table 3, row THE-MESO), while theydid distinguish the thermophilous and mesophilous from theacidophilous types (Table 3, rows THE-ACI and MESO-ACI).Organic carbon and HC did not vary among the three beechforests. The high molecular weight HS (> 100 kDa) signific-antly distinguished the mesophilous from the acidophiloustypes (Table 3, row MESO-ACI).
NPH223.fm Page 650 Wednesday, August 1, 2001 9:27 AM
© N
ew Phytologist (2001) 151: 647–
657w
ww
.newphytologist.com
Research
651
Table 2 Chemical and biochemical parameters of soil A horizons from the 27 sites considered. Sites were classified as thermophilous (THE), mesophilous (MESO), or acidophilous (ACI) beech forests
Table 3 Comparisons among thermophilous (THE), mesophilous (MESO) and acidophilous (ACI) beech forests using the Kruskal–Wallis H-test and multiple comparisons
Site ID pH OC % C/N HC % HC : OC % > 100 kDaHS% 100–10 kDa < 10 kDa IAAlike§ GAlike§ IAA+ % PEROX† % INV‡ %
49 THE 7.4 4.5 10.7 0.93 21.0 33 54 13 0.241 1.300 0.0120 86 13514 THE 7.7 12.4 17.2 2.50 20.0 17 71 12 0.322 7.670 0.0045 54 190217 THE 7.8 6.0 11.4 2.06 34.3 13 77 10 0.048 4.500 0.0126 16 98218 THE 7.5 14.9 12.4 1.42 9.5 14 75 11 0.150 3.100 0.0096 62 140106 THE 7.4 8.8 10.3 2.63 29.9 16 76 8 0.011 1.000 0.0049 64 128105 THE 7.2 8.8 17.0 2.29 26.0 14 74 11 0.202 1.050 0.0092 52 117129 THE 6.5 8.1 11.5 3.64 44.9 19 70 11 0.120 3.300 0.0109 61 144101 THE 6.6 2.5 10.8 1.17 47.0 7 69 24 0.045 1.100 0.0089 26 184107 MESO 7.2 10.8 10.9 3.00 27.8 20 73 7 0.060 0.039 0.0052 145 118108 MESO 7.6 6.6 11.9 1.52 23.2 15 73 12 0.235 0.250 0.0113 54 113109 MESO 6.9 3.5 16.6 1.38 39.5 11 71 18 0.910 0.360 0.0100 159 9530 MESO 7.0 8.2 11.1 2.80 23.0 20 72 8 0.953 0.060 0.0076 115 1066 MESO 6.0 3.6 16.4 6.39 23.0 13 79 8 0.086 0.286 0.0180 138 87110 MESO 5.8 10.9 15.8 3.12 28.7 20 70 10 0.010 1.080 0.0086 121 109130 MESO 6.8 4.7 10.9 2.06 43.8 17 71 12 0.010 0.400 0.0116 73 11815 MESO 6.8 5.4 15.9 1.30 24.0 28 60 12 0.130 0.580 0.0015 63 8934 MESO 6.6 4.5 10.7 0.86 19.0 22 71 6 0.245 0.098 0.0124 43 8935 MESO 7.3 3.6 7.5 0.80 22.0 26 54 20 0.020 0.100 0.0125 100 854 MESO 6.3 7.1 14.8 1.76 25.0 19 72 9 0.034 0.076 0.0182 64 1027 MESO 7.3 6.1 8.1 1.46 24.0 22 63 15 0.100 0.520 0.0133 154 10313 MESO 5.1 7.1 17.7 1.12 16.0 21 68 11 1.025 1.740 0.0091 130 11016 MESO 5.5 11.9 13.5 2.85 24.0 28 61 11 0.047 2.136 0.0105 55 102112 ACI 4.2 9.5 23.8 3.50 36.7 19 70 11 1.732 0.060 0.0126 394 3626 ACI 4.1 4.0 20.1 3.17 78.0 12 66 22 2.050 0.031 0.0363 169 57103 ACI 4.2 4.5 11.5 3.24 72.0 15 71 14 4.670 0.021 0.0192 230 2325 ACI 4.4 4.0 14.3 1.38 36.0 15 69 16 1.100 0.382 0.0145 270 1412 ACI 4.3 7.6 21.1 2.75 36.0 13 76 11 1.890 0.420 0.0281 192 43
§, concentration of indoleacetic acid or of gibberellic acid of equivalent activity as 1 mg C l−1 humic substances. +, % IAA in HS; † peroxidase; and ‡, invertase activities are expressed as percentage of the control.
pH OC C/N HC HC/OC > 100 kDa 100–10 kDa < 10 kDa IAAlike GAlike IAA PEROX INV
THE – MESOχ 4.78 0.85 0.06 0.03 0.47 3.05 0.99 0.30 0.02 9.86 2.05 6.38 9.45P 0.0287* 0.3563 0.8111 0.8645 0.4943 0.0809 0.3204 0.5813 0.8914 0.0017* 0.1518 0.0115* 0.0021*THE – ACIχ 8.62 1.22 4.52 2.14 4.21 0.20 0.54 1.45 8.57 8.57 8.17 8.57 8.57P 0.0033* 0.2703 0.0335* 0.1432 0.0401* 0.6588 0.4617 0.2283 0.0034* 0.0034* 0.0043* 0.0034* 0.0034*MESO – ACIχ 10.53 0.17 3.78 2.63 7.25 4.58 0.00 2.22 10.51 2.34 6.72 10.50 10.53P 0.0012* 0.6764 0.0501* 0.1050 0.0071* 0.0323* 1.0000 0.1359 0.0012* 0.1264 0.0095* 0.0012* 0.0012*
*, significant differences.
NP
H223.fm
Page 651 W
ednesday, August 1, 2001 9:27 A
M

www.newphytologist.com © New Phytologist (2001) 151: 647–657
Research652
Among the biological variables, the IAAlike activity, IAA content,and GAlike activity were complementary in distinguishing thethree types of beech forests. The IAAlike activity and IAA contentwere significantly different when comparing the thermophilousand mesophilous to the acidophilous beech forests (Table 3,
rows THE-ACI and MESO-ACI), while the GAlike activitydistinguished the thermophilous from the mesophilous andacidophilous types (Table 3, rows THE-MESO and THE-ACI).
Peroxidase activity significantly distinguished the thermo-philous and mesophilous forests from the acidophilous
Fig. 1 pH (a), C : N (b) and Humic carbon (HC) (c) of thermophilous (THE), mesophilous (MESO) and acidophilous (ACI) beech forest soils. Difference among forest types was calculated by the Kruskal–Wallis H-test (+) and by the Student-Newman-Keuls pairwise comparison test (similar superscripted letters indicate no significant difference). Vertical bars indicate 1 SE to either side of the mean.
Fig. 2 Organic C (a), HC : OC (b) and HS > 100 kDa (c) of thermophilous (THE), mesophilous (MESO) and acidophilous (ACI) beech forest soils. Difference among forest types was calculated by the Kruskal–Wallis H-test (+) and by the Student-Newman-Keuls pairwise comparison test (similar superscripted letters indicate no significant difference). Vertical bars indicate i SE to either side of the mean.
NPH223.fm Page 652 Wednesday, August 1, 2001 9:27 AM
© New Phytologist (2001) 151: 647–657 www.newphytologist.com
Research 653
forest (Table 3, rows THE-ACI and MESO-ACI). All threebeech forests were significantly different with regard toinvertase activity (Table 3, rows THE-MESO, THE-ACIand MESO-ACI). ANOVA (Table 4) and the Student-Newman-Keuls pairwise comparison tests (Figs 1–4) werealso used to examine differences among the three types. Results
from these tests supported those obtained with the non-parametric tests.
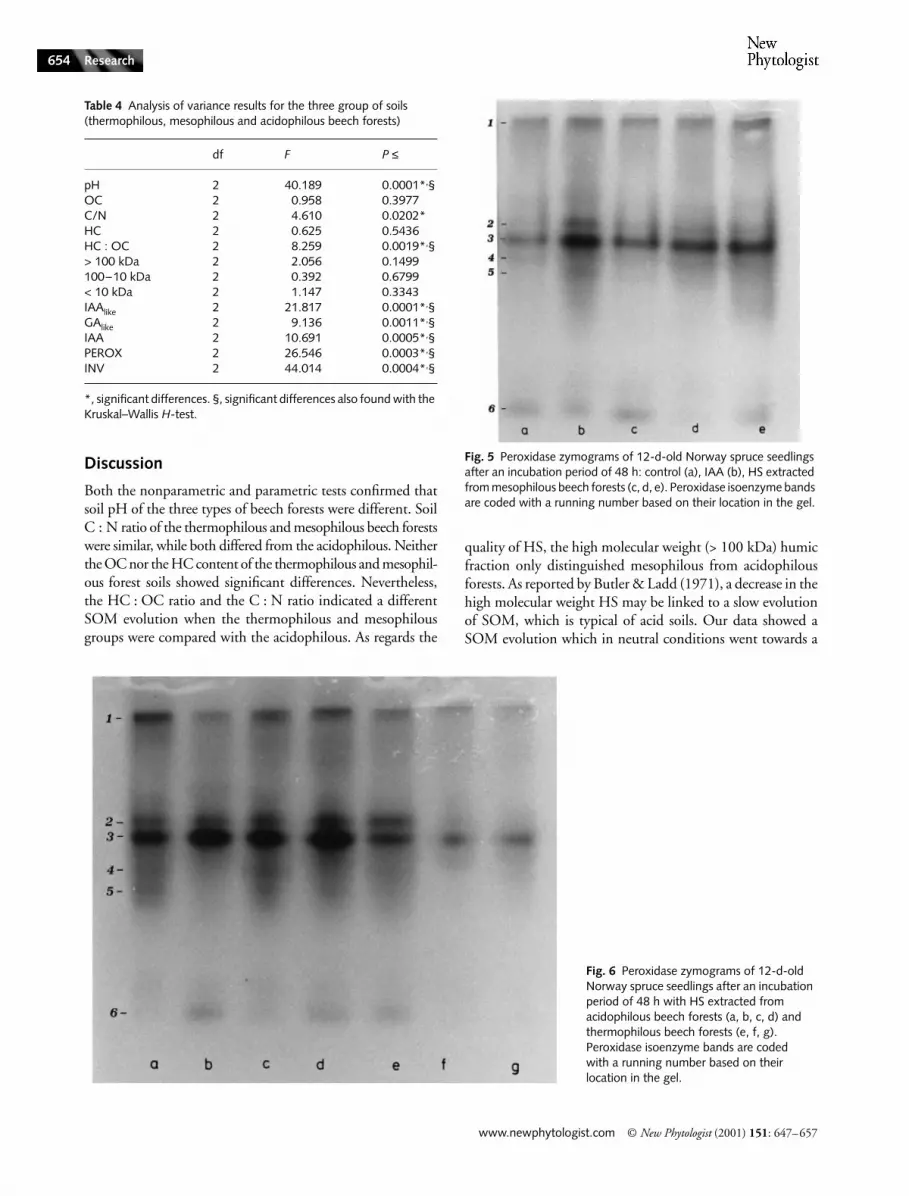
Based on the studies of Wolter & Gordon (1975) and Coppens& Dewitte (1990) where IAA increased plant isoenzymaticpolymorphism, the IAAlike activity of HS was also evaluatedby the pattern of the peroxidase isoenzyme in the Norwayspruce seedlings. In all the cultures tested, the pattern of theperoxidase activity showed quantitative and qualitative differ-ences between the treatments induced by hormonal and humicsubstances (Figs 5 and 6) and in most cases bands 2 and 3were expressed as in the treatment with IAA. The best perox-idase pattern was found in the seedlings treated with HS fromacid soils (Fig. 6a,b,c,d), which included numerous isoenzymaticbands (2, 4, 5) that were present in the electophoretic patternof IAA (Fig. 5b) but that were absent in the control (Fig. 5a).The seedlings treated with HS from the thermophilous group(Fig. 6e,f,g) had less evident bands. A transitional situation wasobserved with HS from the mesophilous group (Fig. 5c,d,e).
Fig. 3 Auxin-like (IAAlike) activity (a), gibberellin-like (Galike) activity (b) and IAA content (c) of thermophilous (THE), mesophilous (MESO) and acidophilous (ACI) beech forest soils. Difference among forest types was calculated by the Kruskal–Wallis H-test (+) and by the Student-Newman-Keuls pairwise comparison test (similar superscripted letters indicate no significant difference). Vertical bars indicate 1 SE to either side of the mean.
Fig. 4 Peroxidase (PEROX) (a) and invertase (INV) (b) activity of thermophilous (THE), mesophilous (MESO) and acidophilous (ACI) beech forest soils. Difference among forest types was calculated by the Kruskal–Wallis H-test (+) and by the Student-Newman-Keuls pairwise comparison test (similar superscripted letters indicate no significant difference). Vertical bars indicate 1 SE to either side of the mean.
NPH223.fm Page 653 Wednesday, August 1, 2001 9:27 AM
www.newphytologist.com © New Phytologist (2001) 151: 647–657
Research654
Discussion
Both the nonparametric and parametric tests confirmed thatsoil pH of the three types of beech forests were different. SoilC : N ratio of the thermophilous and mesophilous beech forestswere similar, while both differed from the acidophilous. Neitherthe OC nor the HC content of the thermophilous and mesophil-ous forest soils showed significant differences. Nevertheless,the HC : OC ratio and the C : N ratio indicated a differentSOM evolution when the thermophilous and mesophilousgroups were compared with the acidophilous. As regards the
quality of HS, the high molecular weight (> 100 kDa) humicfraction only distinguished mesophilous from acidophilousforests. As reported by Butler & Ladd (1971), a decrease in thehigh molecular weight HS may be linked to a slow evolutionof SOM, which is typical of acid soils. Our data showed aSOM evolution which in neutral conditions went towards a
Table 4 Analysis of variance results for the three group of soils (thermophilous, mesophilous and acidophilous beech forests)
df F P ≤
pH 2 40.189 0.0001*,§OC 2 0.958 0.3977C/N 2 4.610 0.0202*HC 2 0.625 0.5436HC : OC 2 8.259 0.0019*,§> 100 kDa 2 2.056 0.1499100–10 kDa 2 0.392 0.6799< 10 kDa 2 1.147 0.3343IAAlike 2 21.817 0.0001*,§GAlike 2 9.136 0.0011*,§IAA 2 10.691 0.0005*,§PEROX 2 26.546 0.0003*,§INV 2 44.014 0.0004*,§
*, significant differences. §, significant differences also found with the Kruskal–Wallis H-test.
Fig. 6 Peroxidase zymograms of 12-d-old Norway spruce seedlings after an incubation period of 48 h with HS extracted from acidophilous beech forests (a, b, c, d) and thermophilous beech forests (e, f, g). Peroxidase isoenzyme bands are coded with a running number based on their location in the gel.
Fig. 5 Peroxidase zymograms of 12-d-old Norway spruce seedlings after an incubation period of 48 h: control (a), IAA (b), HS extracted from mesophilous beech forests (c, d, e). Peroxidase isoenzyme bands are coded with a running number based on their location in the gel.
NPH223.fm Page 654 Wednesday, August 1, 2001 9:27 AM

© New Phytologist (2001) 151: 647–657 www.newphytologist.com
Research 655
humification process as shown by the C : N ratio whichreached a value c. 10, while in acid conditions it went towardsa humification-immobilization process as evidenced by aC : N ratio c. 20.
When the biological activity of HS was considered, however,the IAAlike activity and the IAA content showed significantdifferences between the thermophilous and acidophilousforests and between the mesophilous and acidophilous forests.The GAlike activity showed differences between the acidophil-ous and mesophilous compared with the thermophilous forests.The in vitro and in vivo enzyme activities confirmed the sig-nificant differences when comparing the three groups. Thesedata suggest that acid conditions are essential for the release ofthe auxin-like activity and agree with previous results obtainedin vitro (Nardi et al., 1988). Peroxidase activity, a plant enzymepositively influenced by IAA (Seitz & Lang, 1968; Lee, 1972),followed the same trend as did HS auxin content. In fact, theHS induced a lower peroxidase activity in the neutral soils,while it showed a higher one in the acid soils. When GAlikeactivity was compared with the IAAlike activity, it increasedmore in neutral conditions than in acid ones which was inaccordance with the release of gibberellin-like activity in neutralconditions (Nardi et al., 1988). The results of plant isoenzymaticpolymorphism agreed with the in vitro peroxidase activity andconfirmed that HS from the three groups could be considereddifferent in their auxin activity. The GAlike activity of HS wasconfirmed by the stimulation of invertase in plant metabolismwhich has already been described by Concheri et al. (1994).
These data indicate that vegetation cover is important inSOM evolution. In a previous study we found that litter com-position influences the biological activity of humic matter(Nardi et al., 2000a). Moreover, the characteristics of individualplants have also been found to regulate SOM decompositionand the nutrient cycle in mesic ecosystems (Chen & Stark,2000).
It is known that different soils vary in their native auxincontent (Hamence, 1946) and fertile soils contain greateramounts of auxin than less fertile ones (Stewart & Anderson,1942; Dahm et al., 1977). Auxin and gibberellin levels areusually higher in the rhizosphere than in the bulk soil, prob-ably as a consequence of increased microbial populations orof an accelerated metabolism owing to the presence of rootexudates. Although numerous soil and rhizosphere micro-organisms as well as the root systems of higher plants have beenreported to produce auxins (Lebuhn & Hartmann, 1993) andgibberellins (Rademacher, 1992), there is little informationabout their stability and only indirect conclusions have beenmade about their presence in amounts high enough to be bio-logically active (Frankenberger & Arshad, 1995).
Could HS be considered as a sort of memory of microbialpopulation and plant cover? Frankenberger & Arshad (1995)found that the active ingredients in humus were not mineralnutrients, but were organic substances and biologically activemetabolites of various microbes. Mineral substances applied
in equal amounts to soil had little effects on plant growth. Thefavourable effects of organic substances were observed primarilyafter decomposition and processing of humus, compost, andpeat. Biological and biochemical transformations are mostlikely to occur upon degradation of these materials, whichimplies that the biologically active substances of humus or otherorganic materials are not the original parent compounds, butare products of microbial metabolism. These parental organicmaterials may comprise compounds that serve as precursorsor substrates for the synthesis of biologically active substancesby the heterotrophic activity of the soil microbiota. The plantgrowth regulators kept within HS are of ecological import-ance because they do not leach, and at the same time they arebio-available for plant metabolism (Nardi et al., 2000a).
To conclude, we found that variables related to SOM evolu-tion distinguished two types of soils, the thermophilous andmesophilous from the acidophilous, reflecting the neutral andthe acid conditions, respectively. However, the phyto-hormone-like activity of HS significantly distinguished all three typesof beech forests and showed striking differences within thiswidespread forest ecosystem. Our results showed that vegetationcover was well-reflected in the phyto-hormone-like activityof HS.
Acknowledgements
We thank the Forest Service of Autonomous Province ofTrento for helping us choose the sites studied; M. S. Calabrese,G. Sartori, A. Mancabelli, M. Tomasi and C. De Siena for thefield sampling and the soil classification and A. Zanella for thestatistical processing of the floristic lists. Special thanks toMiss L. Tomat for her help concerning the statistical elaborationand to Ms. Adriana Tuz for kindly improving the English ofthe manuscript. Funds from the Autonomous Province ofTrento have supported this work.
References
Albuzio A, Dell’Agnola G, Dibona D, Concheri G, Nardi S. 1993. Humic constituents of forest soils as plant growth regulating substances. In: Paoletti MG, Foissner W, Coleman D, eds. Soil biota, nutrient cycling and farming systems. Boca Raton, FL, USA: Lewis Publishers, 15–25.
Arnold WN. 1965. β-Fructofuranosidase from grape berries. Biochimica Biophysica Acta 110: 134–147.
Audus LJ. 1972. Plant Growth Substances, vol. 1. Chemistry and Physiology. London, UK: Leonard Hill.
Bottomley WB. 1917. Some effects of organic growth-promotion substances (auximones) on the growth of Lemma minor in mineral cultural solutions. Proceedings of the Royal Society of London (Biological) 89: 481–505.
Butler JHA, Ladd JN. 1971. Importance of the molecular weight of humic and fulvic acids in determining their effects on protease activity. Soil Biology and Biochemistry 3: 249–257.
Cacco G, Dell’Agnola G. 1984. Plant growth regulator activity of soluble humic complexes. Canadian Journal of Soil Science 64: 225–228.
Calabrese MS, Mancabelli A, Nicolini G, Sartori G, Zanella A. 1996. Forest Humus in Trentino (Italy). Report 9. Trento, Italy: Centro di Ecologia Alpina.
NPH223.fm Page 655 Wednesday, August 1, 2001 9:27 AM

www.newphytologist.com © New Phytologist (2001) 151: 647–657
Research656
Calabrese MS, Nardi S, Sartori G, Pizzeghello D, Zanella A, Nicolini G. 1998. Forest humus in the province of Trento (Italian Alps). In: World Congress of Soil Science, 20–26 August, 1998, Montpellier, France. Montpellier, France: Je Megne H.
Calliari I, Concheri G, Pegoraro A, Nardi S. 1993. An application of EDXRF on the study of barley seedlings growth on sewage sludge. Biological Trace Element Research 36: 209–218.
Chen J, Stark JM. 2000. Plant species effects and carbon and nitrogen cycling in a sagebrush-crested wheatgrass soil. Soil Biology and Biochemistry 32: 47–57.
Concheri G, Nardi S, Dell’Agnola G. 1996. Productivity and biological humus activities in forest soils, pp. 131–138. In: Clapp CE, Hayes MHB, Senesi N, Griffith SM, eds. Humic substances and organic matter in soil and water environments: characterization, transformations and interactions. St. Paul, MN, USA: International Humic Substances Society Inc.
Concheri G, Nardi S, Piccolo A, Rascio N, Dell’Agnola G. 1994. Effects of humic fractions on morphological changes related to invertase and peroxidase activities in wheat seedlings. In: Senesi N, Miano TM, eds. Humic substances in the global environment and implications on human health. Amsterdam, The Netherlands: Elsevier Science, 257–262.
Coppens L, Dewitte B. 1990. Esterase and peroxidase zymograms from barley (Hordeum vulgare L.) callus as a biochemical marker system of embryogenesis and organogenesis. Plant Science 67: 97–105.
Dahm H, Sitek JM, Strzelczyk E. 1977. Synthesis of auxins by bacterial isolated from the roots of pine seedlings inoculated with rusty forest soil. Polish Journal of Soil Science 10: 131–137.
Davis BJ. 1964. Disc electrophoresis II. Method and application to human serum proteins. Annals of the New York Academy of Sciences 121: 404–427.
DeJong R, Campbell CA, Nicholaichuk W. 1983. Water retention and their relationship to soil organic matter and their particle size distributions for disturbed samples. Canadian Journal of Soil Science 63: 291–302.
Dell’Agnola G, Ferrari G. 1971. Molecular sizes and functional groups of humic substances extracted by 0.1 M pyrophosphate from soil. Journal of Soil Science 22: 342–349.
Dell’Agnola G, Nardi S. 1987. On overview of earthworm activity in the soil. In: Bonvicini Pagliai AM, Omodeo P, eds. On earthworms selected symposia and monographs 2. Modena, Italy: Mucchi Editore, 103–112.
Dell’Agnola G, Nardi S, Concheri G. 1991. Ruolo dei materiali umici derivanti dalla bioconversione dei residui organici sulla nutrizione e crescita delle piante. In: Senesi N, Miano TM, eds. Riciclo di biomasse di rifiuto e di scarto e fertilizzazione organica del suolo. Bologna, Italy: Patron Editore, 31–38.
Ertel JR, Behmel P, Christman RF, Flaig WJA, Haider KM, Hatcher PG, Hedges JI, Martin JP, Pfaender FK, Schulten HR. 1988. Genesis group report. In: Frimmel FH, Christman RF, eds. Humic substances and their role in the environment. New York, USA: Wiley and Sons, 104–112.
FAO-Unesco. 1990. Soil map of the world. Revised Legend. Roma, Italy: FAO.FitzPatrick EA. 1986. An introduction to soil science, 2nd edn. Essex, UK:
Longman.Frankenberger WT, Arshad M. 1995. Phytormones in soils. New York, USA:
Marcel Dekker Inc.Gibbons JD. 1976. Non-parametric methods for quantitative analysis. New
York, USA: Holt, Rinehart and Winston.Hamence JH. 1946. The determination of auxins in soils, including a note
on synthetic growth substances. Analyst 71: 111–116.Hillitzer A. 1932. Uber den einfluss der humusstoffe auf das wurzelwachstum.
Beih Botany Zbl 49: 467–480.Hoagland DR, Arnon DI. 1950. The water-culture method for growing plants
without soil. California, Agriculture Experimental Station Circular 347. Berkeley, CA, USA: University of California.
Hofmann A. 1991. Il Faggio E le Faggete in Italia Collana verde. Roma, Italy: Ministero dell’Agricoltura e delle Foreste.
Kaufman P, Ghosheh N, Ikuma H. 1968. Promotion of growth and invertase activit by gibberellic aacid in developing avena internodes. Plant Physiology 43: 29–34.
Kaur S, Gupta AK, Kaur N. 2000. Effect of GA3, kinetin and indole acetic acid on carbohydrate metabolism in chickpea seedlings germinating under water stress. Plant Growth Regulation 30: 61–70.
Kim HY, Suzuki Y. 1989. Changes in assimilated 13C distribution and soluble acid invertase activity of Zinnia elegans induced by Uniconazol an inhibitor of gibberellin biosynthesis. Plant Physiology 90: 316–321.
Krsnik-Rasol M, Jelaska S, Sermann D. 1982. Isoperoxydases: early indicators of somatic embryoid differentiation in pumpkin tissue. Acta Botany Croat 41: 33–39.
Lebuhn M, Hartmann A. 1993. Method for determination of indole-3-acetic acid and related compounds of L-tryptophan catabolism in soils. Journal of Chromatography 629: 255–266.
Lee TT. 1972. Interaction of cytokinin, auxin, and gibberellin on peroxidase isozyme in tobacco tissues cultured in vitro. Canadian Journal of Botany 50: 2471–2477.
Lindman HR. 1992. Analysis of variance in experimental design. New York, USA: Springer Verlag.
Lowe LE, Scagel AM, Klinka K. 1987. Chemical properties and classification of organic horizons from selected soils in British Columbia. Canadian Journal of Soil Science 67: 383–394.
Maggioni A, Varanini Z, Nardi S, Pinton R. 1987. Action of soil humic matter on plant roots: stimulation of ion uptake and effects on (Mg2+ and K+) ATPase activity. Science of the Total Environment 62: 355–363.
Muscolo A, Bovalo F, Gionfriddo F, Nardi S. 1999. Earthworm humic matter produces auxin-like effects on Daucus carota cell growth and nitrate metabolism. Soil Biology and Biochemistry 31: 1303–1311.
Muscolo A, Cutrupi S, Nardi S. 1998. IAA detection in humic substances. Soil Biology and Biochemistry 30: 1199–1201.
Muscolo A, Nardi S. 1997. Auxin or auxin-like activity of humic matter. In: Drozd J, Gonet SS, Senesi N, Weber J, eds. The role of humic substances in the ecosystems and environmental protection. Wroclaw, Poland: Polish Society of Humic Substances, 987–992.
Nardi S, Arnoldi G, Dell’Agnola G. 1988. Release of the hormone-like activities from Allolobophora rosea (Sav.) and Allolobophora caliginosa (Sav.) faeces. Canadian Journal of Soil Science 68: 563–567.
Nardi S, Concheri G, Dell’Agnola G. 1996. Biological activity of humus. In: Piccolo A, ed. Humic substances in terrestrial ecosystems. New York, USA: Elsevier, 361–406.
Nardi S, Concheri G, Pizzeghello D, Sturaro A, Rella R, Parvoli G. 2000b. Soil organic matter mobilization by root exudates. Chemosphere 41: 653–658.
Nardi S, Pizzeghello D, Muscolo A, Dalla Vecchia F, Concheri G. 1998. Effects of forest humus on biological activity in roots of Pinus sylvestris related to chemical humus fraction characteristics. Fresenius Environmental Bulletin 7: 203–208.
Nardi S, Pizzeghello D, Reniero F, Muscolo A. 1999. Biological activity of humic substances extracted from soils under different vegetation cover. Communications in Soil Science and Plant Analysis 30: 621–634.
Nardi S, Pizzeghello D, Reniero F, Rascio N. 2000a. Chemical and biochemical properties of humic substances isolated from forest soils and plant growth. Soil Science Society of America Journ 64: 639–645.
Pedrotti F. 1981. Carta Della Vegetazione Del Foglio Di Trento. Roma, Italy: CNR.
Pedrotti F. 1982. Carta Della Vegetazione Del Foglio Di Mezzolombardo. Roma, Italy: CNR.
Pedrotti F. 1987. Carta Della Vegetazione Del Foglio Di Borgo Valsugana. Roma, Italy: CNR.
Pedrotti F. 1994. Guida All’escursione Della Società Italiana Di Fitosociologia in Trentino. 1–5 luglio 1994. Camerino, Italy: CNR.
Petrovich P, Vitorovic D, Jablanovic M. 1982. Investigations of biological effects of humic acids. Acta Biological Medicine Experiment 7: 21–25.
Putter J. 1974. Peroxidases. In: Bergmeyer HV, ed. Methods of enzymatic analysis II. New York, USA: Academic Press, 685–690.
Rademacher W. 1992. Occurrence of gibberellins in different species of the fungal genera Sphaceloma and Elsinoe. Phytochemistry 31: 4155–4157.
NPH223.fm Page 656 Wednesday, August 1, 2001 9:27 AM

© New Phytologist (2001) 151: 647–657 www.newphytologist.com
Research 657
Sanchez M, Pena MJ, Revilla G, Zarra I. 1996. Changes in dehydrodiferulic acids and peroxidase activity against ferulic acid associated with cell walls during growth of Pinus pinaster hypocotyls. Plant Physiology 111: 941–946.
Seitz K, Lang A. 1968. Invertase activity and cell growth in lentil epycotyls. Plant Physiology 43: 1075–1082.
Seybold CA, Mausbach MJ, Karlen DL, Rogers HH. 1998. Quantification of soil quality. In: Lal R, Kimble JM, Follett RF, Stewart BA, eds. Soil processes and the carbon cycle. London, UK: CRC Press, 387–404.
Shaw CR, Prasad R. 1970. Starch gel electrophoresis of enzymes: a compilation of recipes. Biochemical Genetics 4: 297–320.
Soane BD. 1990. The role of organic matter in soil compactability: a review of some practical aspects. Soil Tillage Researches 16: 179–201.
Stevenson FJ. 1994. Humus chemistry: genesis, composition, reactions, 2nd edn. New York, USA: Wiley.
Stewart WS, Anderson MS. 1942. Auxins in some American soils. Botany Gazette 103: 570–575.
Tan KW. 1998. Principles of soil chemistry, 3rd edn. New York, USA: Marcel Dekker Inc.
Vaughan D, Malcolm RE. 1985. Influence of humic substances on growth and physiological processes. In: Vaughan D, Malcolm RE, eds. Soil organic matter and biological activity. Boston, USA: Martinus-Nijhoff, 37–75.
Visser SA. 1986. Effetto delle sostanze umiche sulla crescita delle piante. In: Burns RG, Dell’Agnola G, Miele S, Savoini G, Schnitzer M, Sequi P, Vaughan D, Visser SA, eds. Sostanze umiche effetti sul terreno e sulle piante. Milano, Italy: Reda, 96–114.
Wolter KE, Gordon JC. 1975. Peroxidases as indicator of growth and differ-entiation in aspen callus cultures. Physiologia Plantarum 33: 219–223.
Zanella A, Sartori G, Calabrese MS, Nicolini G, Mancabelli A. 1997. Verso una tipologia delle faggete ed abieti-faggete del Trentino. Monti e Boschi 3: 14–19.
NPH223.fm Page 657 Wednesday, August 1, 2001 9:27 AM