gaba receptor subtype antagonists in the nucleus accumbens shell and ventral tegmental area...
TRANSCRIPT

B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res
Research Report
GABA receptor subtype antagonists in the nucleus accumbensshell and ventral tegmental area differentially alter feedingresponses induced by deprivation, glucoprivation andlipoprivation in rats
Y. Kandova, Y. Israela, A. Kesta, I. Dostovaa, J. Verasammya, S.Y. Bernala,b,L. Kasselmana,b, R.J. Bodnara,b,⁎aDepartment of Psychology, Doctoral Sub-Program, Queens College, CUNY, Flushing, NY 11367, USAbNeuropsychology Doctoral Sub-Program, Queens College, CUNY, Flushing, NY 11367, USA
A R T I C L E I N F O
⁎ Corresponding author. Fax: +1 718 997 3257.E-mail address: richard_bodna [email protected] (R
0006-8993/$ – see front matter © 2006 Elsevidoi:10.1016/j.brainres.2006.01.112
A B S T R A C T
Article history:Accepted 28 January 2006Available online 6 March 2006
GABAA and GABAB receptor agonists stimulate feeding following microinjection into thenucleus accumbens shell and ventral tegmental area, effects blocked selectively andrespectively by GABAA and GABAB receptor antagonists. GABA antagonists also differentiallyalter opioid-induced feeding responses elicited from these sites. Although GABA agonists andantagonists have been shown to modulate feeding elicited by deprivation or glucoprivation,there has been no systematic examination of feeding elicited by homeostatic challengesfollowing GABA antagonists in these sites. Therefore, the present study examined the dose-dependent ability of GABAA (bicuculline, 75–150 ng) and GABAB (saclofen, 1.5–3 μg) antagonistsadministered into the nucleus accumbens shell or ventral tegmental area upon feedingresponses elicited by food deprivation (24 h), 2-deoxy-D-glucose-induced glucoprivation (500mg/kg) or mercaptoacetate-induced lipoprivation (70 mg/kg). A site-specific effect of GABA receptorantagonismwas observed for deprivation-induced feeding in that both bicuculline and saclofenadministered into the nucleus accumbens shell, but not the ventral tegmental area, producedshort-term (1–4 h), but not long-term (24–48 h) effects upon deprivation-induced intake withoutmeaningfully altering bodyweight recovery. In contrast to the relative inability of GABA receptorantagonism in both sites to alter 2-deoxy-D-glucose-induced intake, mercaptoacetate-inducedintake was eliminated by saclofen and significantly reduced by bicuculline in the nucleusaccumbens shell and eliminated by both bicuculline and saclofen in the ventral tegmental area.These data reinforce the findings that GABAA and GABAB receptors in the nucleus accumbensshell and ventral tegmental area are not only important in themodulation of pharmacologicallyinduced feeding responses, but also participate in differentially mediating the short-termfeeding response to food deprivation in the nucleus accumbens shell as well stronglymodulating lipoprivic, but not glucoprivic feeding responses in both sites.
© 2006 Elsevier B.V. All rights reserved.
Keywords:BicucullineSaclofenGABAA receptorGABAB receptor2-deoxy-D-glucoseMercaptoacetate
.J. Bod nar).
er B.V. All rights reserved.

87B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
1. Introduction
Site-specific feeding responses can be elicited by directadministration of GABA or its agonists into the ventraltegmental area (VTA: e.g., Arnt and Scheel-Kruger, 1979;Echo et al., 2002; Khaimova et al., 2004; Klitenick andWirtshafter, 1988), nucleus accumbens (NAC) shell (e.g.,Soderpalm and Berridge, 2000; Stratford and Kelley, 1997;Ward et al., 2000; Znamensky et al., 2001), substantia nigra(SN: Redgrave et al., 1984; but see Kelly et al., 1977), median(Klitenick and Wirtshafter, 1989; Wirtshafter et al., 1993) anddorsal (Bendotti et al., 1986; Borsini et al., 1983) raphe nucleiand ventro-medial hypothalamus (VMH: Grandison andGuidotti, 1977; Kelly et al., 1977; Tsujii and Bray, 1991) infree-feeding rats. In contrast, GABA agonists suppress foodintake following microinjections into the central nucleus ofthe amygdala and lateral hypothalamus (LH) (Kelly et al.,1977; Minano et al., 1992). Among these sites, the NAC shelland tegmental area (VTA) have been implicated in themediation of reward and reinforcement related to appetitive,ingestive, stress-related, aversive and drug-seeking beha-viors (e.g., see reviews: Kelley, 1999, 2004) traditionallythrough dopamine mechanisms (e.g., see reviews: Berridgeand Robinson, 1998; Salamone, 1994). Although dopamineplays an important role in these processes, other modulatorytransmitters, including GABA connections between thesenuclei (Van Bockstaele and Pickel, 1995), have been impli-cated in mediating ingestive responses. Thus, GABA receptorsubtype antagonists selectively and specifically reducefeeding induced by GABA receptor subtype agonists in theVTA (Echo et al., 2002) and NAC shell (Stratford and Kelley,1997; Znamensky et al., 2001). Thus, feeding elicited by theselective GABAA agonist muscimol is blocked by pretreat-ment with the GABAA antagonist bicuculline but not theGABAB antagonist saclofen in either the NAC shell or theVTA. Correspondingly, feeding elicited by the selectiveGABAB agonist baclofen is selectively blocked by pretreat-ment with the GABAB antagonist saclofen but not bicucullinein either the NAC shell or the VTA. Region-specific effects inthe NAC shell were observed such that muscimol increasesfeeding, visits to food, place preferences and positivehedonic reactions after administration into the rostral NACshell and defensive paw-treading and burial behaviors afteradministration into the caudal NAC shell (Reynolds andBerridge, 2001, 2002). Muscimol in the NAC shell activatesFos-like immunoreactivity in the lateral hypothalamus,lateral septum, paraventricular hypothalamus, ventral teg-mental area, substantia nigra and nucleus of the solitarytract (Stratford and Kelley, 1999), stimulates intake of bothcarbohydrates and fats regardless of whether the macronu-trients are presented singly or together and increases intakeof sucrose, but not water, saccharin or saline (Basso andKelley, 1999). However, increases in spontaneous intake, butnot operant lever-pressing for food, are observed followingmuscimol, DAMGO or amphetamine administered into theNAC shell (Hanlon et al., 2004). Finally, muscimol adminis-tration into the central amygdala reduces GABAA-agonist-induced feeding elicited from the NAC shell (Baldo et al.,2005).
GABA receptor agonists and antagonists interact withother transmitter systems in modulating feeding responses.Within the hypothalamic paraventricular nucleus (PVN),combined muscimol and NPY microinjections synergize toincrease food intake, an effect not observed in senescentrats (Coppola et al., 2005; Pu et al., 1999). NPY Y1 or Y5receptor antagonists block muscimol-induced feeding eli-cited from the NAC shell (Stratford and Wirtshafter, 2004).Muscimol infusion into the NAC shell increases c-Fos inorexin-containing neurons in the perifornical area and NPYneurons in the arcuate nucleus, while decreasing c-Fos incocaine–amphetamine-related transcript- and proopiomela-nocortin-containing neurons in the arcuate nucleus (Zhenget al., 2003). Glutamate–GABA interactions in feeding areevident by the facts that feeding elicited by AMPA/kainiteantagonists in the NAC shell is blocked by muscimolinfusions into the LH (Maldonado-Irizarry et al., 1995) andthat muscimol-induced feeding in the NAC shell is blockedby LH administration of D-AP5 (Stratford and Kelley, 1999).GABA receptor subtype antagonists in the NAC shell andVTA also locally modulate opioid-agonist-induced feedingresponses. Feeding elicited by the μ opioid agonist DAMGOin the NAC shell was respectively enhanced and reduced byNAC shell pretreatment with GABAA (bicuculline) andGABAB (saclofen) antagonists (Znamensky et al., 2001).DAMGO-induced feeding elicited from the VTA was signif-icantly reduced by VTA pretreatment with saclofen in theabsence of conditioned aversions and pretreatment withbicuculline in the presence of seizure activity (Echo et al.,2002). In contrast, if either GABA antagonist was adminis-tered in one site (e.g., NAC shell) and the opioid agonist wasadministered into the second site (e.g., VTA), the resultantopioid-mediated orexigenic responses were unaffected,indicating a lack of regional GABA–opioid interactions(Ackerman et al., 2003). Moreover, DAMGO-induced feedingelicited from the NAC shell is blocked by temporaryinactivation with high doses of muscimol of the dorsome-dial hypothalamus, LH, VTA, nucleus tractus solitarius oramygdala (Will et al., 2003, 2004). In turn, opioid receptorsubtype antagonists locally modulate feeding responseselicited by GABA receptor agonists within the NAC shelland VTA. Muscimol-induced feeding elicited from the VTAwas significantly enhanced by μ or δ antagonists and wassignificantly reduced by κ antagonists. Baclofen-inducedfeeding elicited from the VTA was significantly reduced byμ or κ, but not δ antagonists. Muscimol-induced feedingelicited from the NAC was significantly reduced by μ, κ or δantagonists. Baclofen-induced feeding elicited from the NACwas significantly reduced by κ or δ, but not μ antagonists(Khaimova et al., 2004).
Whereas the role of GABA in modulating pharmacologi-cally induced feeding responses elicited from the NAC shelland VTA has been studied extensively, relatively little isknown about GABAergic modulation in the NAC shell or VTAof homeostatically controlled feeding responses. Previousstudies demonstrated that systemic pretreatment with eitherthe GABA-transaminase inhibitor ethanolamine-O-sulfate(Olgiati et al., 1980) or the GABA agonist THIP (Blavet et al.,1982) decreased intake in both freely fed and food-deprivedrats, whereas systemic picrotoxin or bicuculline failed to alter

88 B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
chlordiazepoxide-induced increases in deprivation-inducedintake (Sanger, 1984). Muscimol, but not baclofen, microinjec-tions into the central nucleus of the amygdala decreaseddeprivation-induced intake that was blocked by bicucullinepretreatment (Minano et al., 1992). Moreover, memories oftasks related to food reward in food-restricted rats wererespectively attenuated and enhanced by muscimol andbicuculline treatment in the amygdala (Salinas and McGaugh,1996). Muscimol administration into the VMH stimulatedspontaneous intake in lean, but not genetically obese Zuckerrats (Tsujii and Bray, 1991). Whereas GABAA receptor anta-gonism with bicuculline in the anterior piriform cortexstimulated intake of an amino acid imbalanced, but not abasal diet, GABAB receptor antagonism with phaclofen in theanterior piriform cortex stimulated intake of the basal, but notthe amino acid imbalanced diet (Truong et al., 2002). Acuteglucoprivic feeding responses elicited by either 2-deoxy-D-glucose (2DG) or insulin were abolished by intracisternaladministration of ethanolamine-O-sulfate (Nobrega and Cos-cina, 1982). Furthermore, glucoprivic feeding responses in-duced by 2DG, 5-thioglucose or insulin were blocked by eithersystemic or VMH, but not LH microinjections of eitherpicrotoxin or bicuculline (Lenin Kamatchi et al., 1984, 1986).
Fig. 1 – Representation of cannula sites aimed unilaterally at ththe stereotaxic atlas of Paxinos and Watson (1998). Multiple aconsiderable overlap of placements is depicted on the panelsfood deprivation (F) and mercaptoacetate (M) paradigms.
In contrast, microinjections of bicuculline into the LH en-hanced sweetened milk intake (Kelly et al., 1977).
The present study examined the dose-dependent and time-dependent ability of theGABAA receptor antagonist bicucullineand the GABAB receptor antagonist saclofen administered intoeither the NAC shell or the VTA to alter feeding responsesinduced by 24 h of food deprivation, 2DG or mercaptoacetate(MA). Both the anti-metabolic glucose analogue 2DG (e.g.,Wicket al., 1957) and the free fatty acid oxidation inhibitor MA (e.g.,Bauche et al., 1981) elicit glucoprivic (e.g., Smith and Epstein,1969; Smith and Root, 1969) and lipoprivic (e.g., Langhans andScharrer, 1987; Scharrer and Langhans, 1986) feedingresponses respectively.
2. Results
2.1. Histological verification
The mesencephalic cannula placements of all animals partic-ipating in the protocols were found to be within the region ofthe VTA, bordered as far rostral as the level of the fasciculusretroflexus and commissure of the superior colliculus,
e VTA region (A) and bilaterally at the NAC shell (B) usingnimals had highly similar cannula placements, and theamong rats participating in the 2-deoxy-D-glucose (D),
89B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
bordered as far caudally as the level of the interpeduncularnucleus, bordered as far dorsally as the rostral linear nucleusof the raphe and bordered as far laterally as the substantianigra pars compacta (Fig. 1, left panel). Bilateral forebraincannulae placements were all localized to the rostral shellregion of the NAC and immediately adjacent ventral diagonalband area; these placements were all medial to the core regionof the NAC (Fig. 1, right panel). The cannula placements werefound to be intermingled within the NAC shell and VTArespectively across the three protocols. The cannula place-ments were highly similar to those reported previously by ourlaboratory for the VTA and NAC shell regions (e.g., Ackermanet al., 2003; Bodnar et al., 1995; Echo et al., 2002; Khaimova etal., 2004; Ragnauth et al., 1997; Znamensky et al., 2001) andsimilar to rostral NAC shell sites specifically stimulatingfeeding and not defensive responses (Reynolds and Berridge,2001, 2002).
2.2. GABA antagonists in the NAC shell anddeprivation-induced intake
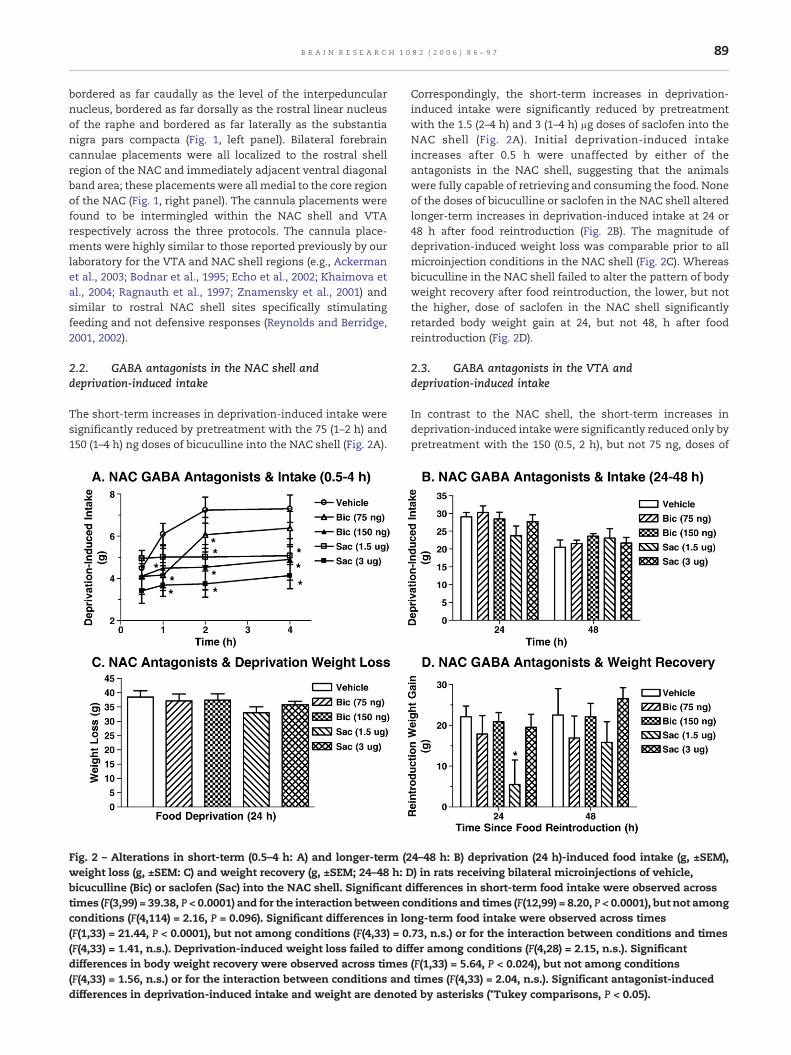
The short-term increases in deprivation-induced intake weresignificantly reduced by pretreatment with the 75 (1–2 h) and150 (1–4 h) ng doses of bicuculline into the NAC shell (Fig. 2A).
Fig. 2 – Alterations in short-term (0.5–4 h: A) and longer-term (2weight loss (g, ±SEM: C) and weight recovery (g, ±SEM; 24–48 h: Dbicuculline (Bic) or saclofen (Sac) into the NAC shell. Significant dtimes (F(3,99) = 39.38, P < 0.0001) and for the interaction between cconditions (F(4,114) = 2.16, P = 0.096). Significant differences in lo(F(1,33) = 21.44, P < 0.0001), but not among conditions (F(4,33) = 0(F(4,33) = 1.41, n.s.). Deprivation-induced weight loss failed to difdifferences in body weight recovery were observed across times(F(4,33) = 1.56, n.s.) or for the interaction between conditions anddifferences in deprivation-induced intake and weight are denote
Correspondingly, the short-term increases in deprivation-induced intake were significantly reduced by pretreatmentwith the 1.5 (2–4 h) and 3 (1–4 h) μg doses of saclofen into theNAC shell (Fig. 2A). Initial deprivation-induced intakeincreases after 0.5 h were unaffected by either of theantagonists in the NAC shell, suggesting that the animalswere fully capable of retrieving and consuming the food. Noneof the doses of bicuculline or saclofen in the NAC shell alteredlonger-term increases in deprivation-induced intake at 24 or48 h after food reintroduction (Fig. 2B). The magnitude ofdeprivation-induced weight loss was comparable prior to allmicroinjection conditions in the NAC shell (Fig. 2C). Whereasbicuculline in the NAC shell failed to alter the pattern of bodyweight recovery after food reintroduction, the lower, but notthe higher, dose of saclofen in the NAC shell significantlyretarded body weight gain at 24, but not 48, h after foodreintroduction (Fig. 2D).
2.3. GABA antagonists in the VTA anddeprivation-induced intake
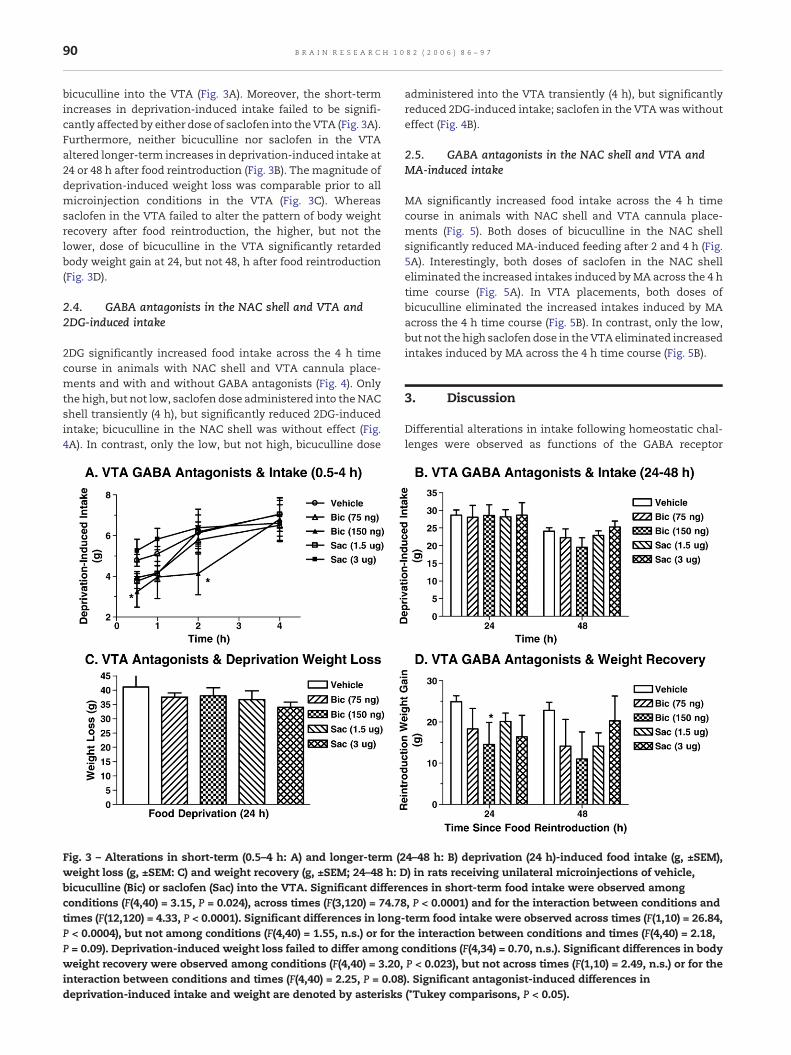
In contrast to the NAC shell, the short-term increases indeprivation-induced intake were significantly reduced only bypretreatment with the 150 (0.5, 2 h), but not 75 ng, doses of
4–48 h: B) deprivation (24 h)-induced food intake (g, ±SEM),) in rats receiving bilateral microinjections of vehicle,ifferences in short-term food intake were observed acrossonditions and times (F(12,99) = 8.20, P < 0.0001), but not amongng-term food intake were observed across times.73, n.s.) or for the interaction between conditions and timesfer among conditions (F(4,28) = 2.15, n.s.). Significant(F(1,33) = 5.64, P < 0.024), but not among conditionstimes (F(4,33) = 2.04, n.s.). Significant antagonist-inducedd by asterisks (*Tukey comparisons, P < 0.05).
90 B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
bicuculline into the VTA (Fig. 3A). Moreover, the short-termincreases in deprivation-induced intake failed to be signifi-cantly affected by either dose of saclofen into the VTA (Fig. 3A).Furthermore, neither bicuculline nor saclofen in the VTAaltered longer-term increases in deprivation-induced intake at24 or 48 h after food reintroduction (Fig. 3B). The magnitude ofdeprivation-induced weight loss was comparable prior to allmicroinjection conditions in the VTA (Fig. 3C). Whereassaclofen in the VTA failed to alter the pattern of body weightrecovery after food reintroduction, the higher, but not thelower, dose of bicuculline in the VTA significantly retardedbody weight gain at 24, but not 48, h after food reintroduction(Fig. 3D).
2.4. GABA antagonists in the NAC shell and VTA and2DG-induced intake
2DG significantly increased food intake across the 4 h timecourse in animals with NAC shell and VTA cannula place-ments and with and without GABA antagonists (Fig. 4). Onlythe high, but not low, saclofen dose administered into the NACshell transiently (4 h), but significantly reduced 2DG-inducedintake; bicuculline in the NAC shell was without effect (Fig.4A). In contrast, only the low, but not high, bicuculline dose
Fig. 3 – Alterations in short-term (0.5–4 h: A) and longer-term (2weight loss (g, ±SEM: C) and weight recovery (g, ±SEM; 24–48 h: Dbicuculline (Bic) or saclofen (Sac) into the VTA. Significant differeconditions (F(4,40) = 3.15, P = 0.024), across times (F(3,120) = 74.7times (F(12,120) = 4.33, P < 0.0001). Significant differences in longP < 0.0004), but not among conditions (F(4,40) = 1.55, n.s.) or for tP = 0.09). Deprivation-induced weight loss failed to differ amongweight recovery were observed among conditions (F(4,40) = 3.20,interaction between conditions and times (F(4,40) = 2.25, P = 0.08deprivation-induced intake and weight are denoted by asterisks
administered into the VTA transiently (4 h), but significantlyreduced 2DG-induced intake; saclofen in the VTAwas withouteffect (Fig. 4B).
2.5. GABA antagonists in the NAC shell and VTA andMA-induced intake
MA significantly increased food intake across the 4 h timecourse in animals with NAC shell and VTA cannula place-ments (Fig. 5). Both doses of bicuculline in the NAC shellsignificantly reduced MA-induced feeding after 2 and 4 h (Fig.5A). Interestingly, both doses of saclofen in the NAC shelleliminated the increased intakes induced by MA across the 4 htime course (Fig. 5A). In VTA placements, both doses ofbicuculline eliminated the increased intakes induced by MAacross the 4 h time course (Fig. 5B). In contrast, only the low,but not the high saclofen dose in theVTA eliminated increasedintakes induced by MA across the 4 h time course (Fig. 5B).
3. Discussion
Differential alterations in intake following homeostatic chal-lenges were observed as functions of the GABA receptor
4–48 h: B) deprivation (24 h)-induced food intake (g, ±SEM),) in rats receiving unilateral microinjections of vehicle,nces in short-term food intake were observed among8, P < 0.0001) and for the interaction between conditions and-term food intake were observed across times (F(1,10) = 26.84,he interaction between conditions and times (F(4,40) = 2.18,conditions (F(4,34) = 0.70, n.s.). Significant differences in bodyP < 0.023), but not across times (F(1,10) = 2.49, n.s.) or for the). Significant antagonist-induced differences in(*Tukey comparisons, P < 0.05).
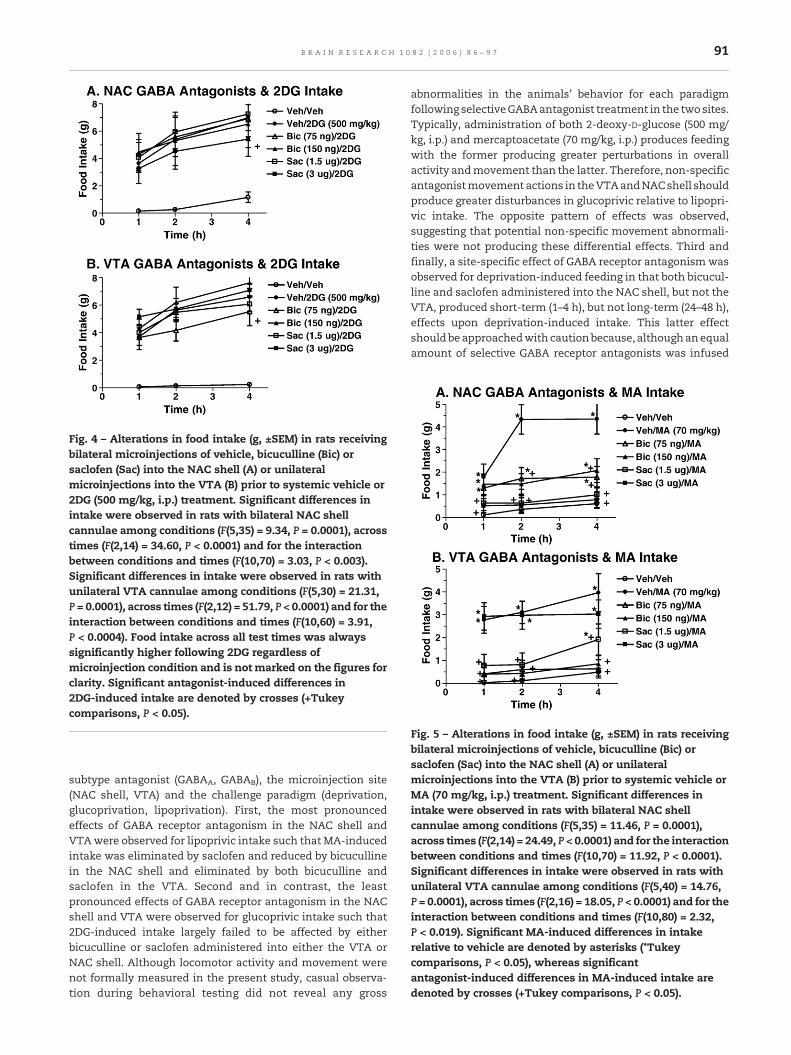
Fig. 4 – Alterations in food intake (g, ±SEM) in rats receivingbilateral microinjections of vehicle, bicuculline (Bic) orsaclofen (Sac) into the NAC shell (A) or unilateralmicroinjections into the VTA (B) prior to systemic vehicle or2DG (500 mg/kg, i.p.) treatment. Significant differences inintake were observed in rats with bilateral NAC shellcannulae among conditions (F(5,35) = 9.34, P = 0.0001), acrosstimes (F(2,14) = 34.60, P < 0.0001) and for the interactionbetween conditions and times (F(10,70) = 3.03, P < 0.003).Significant differences in intake were observed in rats withunilateral VTA cannulae among conditions (F(5,30) = 21.31,P = 0.0001), across times (F(2,12) = 51.79, P < 0.0001) and for theinteraction between conditions and times (F(10,60) = 3.91,P < 0.0004). Food intake across all test times was alwayssignificantly higher following 2DG regardless ofmicroinjection condition and is not marked on the figures forclarity. Significant antagonist-induced differences in2DG-induced intake are denoted by crosses (+Tukeycomparisons, P < 0.05).
Fig. 5 – Alterations in food intake (g, ±SEM) in rats receivingbilateral microinjections of vehicle, bicuculline (Bic) orsaclofen (Sac) into the NAC shell (A) or unilateralmicroinjections into the VTA (B) prior to systemic vehicle orMA (70 mg/kg, i.p.) treatment. Significant differences inintake were observed in rats with bilateral NAC shellcannulae among conditions (F(5,35) = 11.46, P = 0.0001),across times (F(2,14) = 24.49, P < 0.0001) and for the interactionbetween conditions and times (F(10,70) = 11.92, P < 0.0001).Significant differences in intake were observed in rats withunilateral VTA cannulae among conditions (F(5,40) = 14.76,P = 0.0001), across times (F(2,16) = 18.05, P < 0.0001) and for theinteraction between conditions and times (F(10,80) = 2.32,P < 0.019). Significant MA-induced differences in intakerelative to vehicle are denoted by asterisks (*Tukeycomparisons, P < 0.05), whereas significantantagonist-induced differences in MA-induced intake aredenoted by crosses (+Tukey comparisons, P < 0.05).
91B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
subtype antagonist (GABAA, GABAB), the microinjection site(NAC shell, VTA) and the challenge paradigm (deprivation,glucoprivation, lipoprivation). First, the most pronouncedeffects of GABA receptor antagonism in the NAC shell andVTAwere observed for lipoprivic intake such that MA-inducedintake was eliminated by saclofen and reduced by bicucullinein the NAC shell and eliminated by both bicuculline andsaclofen in the VTA. Second and in contrast, the leastpronounced effects of GABA receptor antagonism in the NACshell and VTA were observed for glucoprivic intake such that2DG-induced intake largely failed to be affected by eitherbicuculline or saclofen administered into either the VTA orNAC shell. Although locomotor activity and movement werenot formally measured in the present study, casual observa-tion during behavioral testing did not reveal any gross
abnormalities in the animals' behavior for each paradigmfollowing selectiveGABAantagonist treatment in the two sites.Typically, administration of both 2-deoxy-D-glucose (500 mg/kg, i.p.) and mercaptoacetate (70 mg/kg, i.p.) produces feedingwith the former producing greater perturbations in overallactivity andmovement than the latter. Therefore, non-specificantagonistmovement actions in theVTAandNACshell shouldproduce greater disturbances in glucoprivic relative to lipopri-vic intake. The opposite pattern of effects was observed,suggesting that potential non-specific movement abnormali-ties were not producing these differential effects. Third andfinally, a site-specific effect of GABA receptor antagonismwasobserved for deprivation-induced feeding in that both bicucul-line and saclofen administered into the NAC shell, but not theVTA, produced short-term (1–4 h), but not long-term (24–48 h),effects upon deprivation-induced intake. This latter effectshouldbe approachedwith cautionbecause, althoughanequalamount of selective GABA receptor antagonists was infused

92 B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
into the two sites, administration into the NAC shell wasaccomplished through bilateral cannulae, whereas adminis-tration into the VTA regionwas accomplished through a singlecannula aimedat themidline.Although the bilateral cannulaewere localized to the NAC shell, the unilateral cannulaeaimed at the VTA were sometimes found to be off themidline, raising the possibility that only unilateral infusionsinto the VTA region were administered. Therefore, whereasthere is assurance that a substantial area of the NAC shellreceived GABA antagonists in these studies, it is possiblethat the VTA region did not. However, there was substantialoverlap of VTA cannula placements across the threeparadigms, and the effects of VTA GABA antagonists uponlipoprivic intake were equal to (saclofen) or even greater(bicuculline) than NAC shell effects.
3.1. GABA antagonism and deprivation-induced intake
As previously indicated, manipulations of GABAergic sys-tems modulate deprivation-induced feeding. Thus, systemicpretreatment with either ethanolamine-O-sulfate (Olgiati etal., 1980) or THIP (Blavet et al., 1982) decreased intake inboth freely fed and food-deprived rats, whereas systemicpicrotoxin or bicuculline failed to alter chlordiazepoxide-induced increases in deprivation-induced intake (Sanger,1984). Muscimol, but not baclofen, microinjections into thecentral nucleus of the amygdala decreased deprivation-induced intake that was blocked by bicuculline pretreat-ment (Minano et al., 1992). Moreover, memories of tasksrelated to food reward in food-restricted rats were respec-tively attenuated and enhanced by muscimol and bicucul-line treatment in the amygdala (Salinas and McGaugh,1996). In this study, we find that selective GABAA andGABAB antagonists administered into the NAC shell, but notinto the VTA, decreased short-term increases in depriva-tion-induced intake, thereby indicating that perturbations ofGABA actions (through exogenous administration of GABAantagonists, GABA agonists (muscimol) or GABA modulators(ethanolamine-O-sulfate)) can all act to decrease thishomeostatic challenge.
These antagonist actions are specific in several respects.First, the decreases in deprivation-induced intake by bicucul-line and saclofen in the NAC shell were temporally delimitedin that the decreases were noted from 1 to 4 h after foodreintroduction. Interestingly, both doses of bicuculline andsaclofen in the NAC shell failed to affect the initial largeincrease in intake noted 0.5 h following food reintroduction,suggesting that these antagonists are acting on the mainte-nance, but not the initiation of deprivation-induced intake.This lack of effect serves as an important control because itminimizes the possibility that these antagonists are actingnon-specifically on intake through interference with motoractivity. Thus, the animals ate normally in the first 30 minfollowing food reintroduction, and the GABAA and GABAB
antagonist effects acted subsequently. However, the long timecourse of antagonist effects raises the possibility that drugdiffusion to remote sites other than the NAC shell may be afactor in producing these effects. Although there were nomisplaced cannulae that could serve as anatomical controls inthis study, previous studies using selective GABA agonists
failed to display feeding effects when cannulae were placedoutside of the NAC shell (e.g., Khaimova et al., 2004;Znamensky et al., 2001). Second, neither GABA antagonistproduced any consistent longer-term ingestive effects withnormal intake noted 24 and 48 h following food reintroduc-tion. Therefore, compensatory increases in deprivation-in-duced intake occurred in the ensuing 20 h following the initial4 h of antagonist-induced decreases. Third, neither GABAantagonist interfered with body weight recovery noted in the48 h following food reintroduction, suggesting that GABAantagonist actions on short-term intake did not extend topronounced metabolic effects. Fourth, the GABA antagonisteffects upon deprivation-induced feeding were site-specificwith both antagonists active in the NAC shell, but not in theVTA. Importantly, the receptor subtype selective effects ofGABAA and GABAB agonists and antagonists are present inboth the NAC shell and VTA (Echo et al., 2002; Stratford andKelley, 1997; Znamensky et al., 2001). Hence, muscimol-induced feeding is selectively blocked by bicuculline, but notsaclofen pretreatment in both the NAC shell and the VTA,whereas baclofen-induced feeding is selectively blocked bysaclofen, but not bicuculline, pretreatment in both the NACshell and the VTA. Thus, the same dose ranges of bothantagonists are active in both sites for pharmacologicallyinduced feeding, but selective in the NAC shell, but not theVTA, for deprivation-induced feeding. A parallel pattern ofeffects has been observed for selective opioid antagonists inthese sites in that deprivation-induced feeding is decreased byμ and κ opioid antagonists administered into the NAC shell,but not the VTA (Bodnar et al., 1995; Kelley et al., 1996;Ragnauth et al., 1997). Yet, μ and κ opioid antagonists decreasefeeding induced by μ opioid agonists in both the NAC shell andVTA (Lamonte et al., 2002; Ragnauth et al., 2000).
3.2. GABA antagonism and lipoprivic andglucoprivic-induced intake
As previously indicated, glucoprivic feeding responseselicited by either 2-deoxy-D-glucose (2DG) or insulin wereabolished by intracisternal administration of ethanolamine-O-sulfate (Nobrega and Coscina, 1982) and by systemic orVMH administration of either picrotoxin or bicuculline(Lenin Kamatchi et al., 1984, 1986). The present studyindicated that 2DG-induced intake was largely unaffectedby either bicuculline or saclofen administration into eitherthe VTA or NAC shell, suggesting a relative lack of GABAantagonist action upon glucoprivic feeding in these sites. Incontrast, lipoprivic feeding induced by MA was eliminatedby saclofen and reduced by bicuculline in the NAC shell andeliminated by both bicuculline and saclofen in the VTA. Anample literature exists indicating dissociations betweenlipoprivic and glucoprivic feeding. Although MA and 2DGadministration each elicit c-fos responses in the nucleus ofthe solitary tract, lateral parabrachial nucleus, centralnucleus of the amygdala and the dorsal motor nucleus ofthe vagus, the former, but not latter, response is abolishedby vagotomy (Ritter and Dinh, 1994). Indeed, lesions placedin the lateral parabrachial nucleus abolish MA-inducedfeeding but fail to affect 2DG-induced feeding (Calingasanand Ritter, 1993), whereas lesions placed in the central

93B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
nucleus of the amygdala produce more marked reductionsof MA-induced feeding relative to 2DG-induced feeding(Ritter and Hutton, 1995). Correspondingly, MA-induced,but not 2DG-induced, feeding is reduced by either destruc-tion of visceral sensory neurons with capsaicin (Ritter andTaylor, 1989) or subdiaphragmatic vagotomy (Ritter andTaylor, 1990). Indeed, whereas 2DG increased sympathoa-drenal plasma epinephrine levels, MA increased sym-pathoadrenal plasma norepinephrine levels (Scheurink andRitter, 1993). Moreover, the beta-2-adrenoceptor agonistsalbutamol antagonizes MA-induced feeding but fails toaffect 2DG-induced feeding (Nisoli et al., 1996). However,2DG, but not MA, increases c-fos responses in the adrenalmedulla and sympathetic preganglionic spinal cord neurons(Ritter et al., 1995). The general ability of the present studyand these previous lesion studies in observing disruptionsof lipoprivic relative to glucoprivic intake may reflect agreater survival importance of glucoprivic signals in ani-mals whose customary diets are high in carbohydrate andlow in fat and thereby reflect a greater regional anatomicaland chemical coding of glucoprivic relative to lipoprivicenergy regulation.
Given that the present study employed chow which ishigher in carbohydrate and less in fat, would the pattern ofresults reverse if the animals were maintained on analternative high fat and low carbohydrate diet? Thiswould not appear to be the case given that brain GABAincreases with fat intake especially in animals resistant toobesity (Fisler et al., 1989) and that baclofen increasesthermogenic activity of brown adipose tissue (Addae et al.,1986). Furthermore, muscimol administered into the medialNAC shell increases intakes of both high fat and highcarbohydrate diets presented separately or together (Bassoand Kelley, 1999), suggesting that diet per se is not a criticalfactor. Moreover, these differential challenge responsesextend to different effects upon macronutrient intake.Whereas 2DG stimulates intake of all three macronutrientsacross a wide range of dietary conditions, MA reliablystimulates protein intake, stimulates carbohydrate intakeonly when carbohydrate palatability is enhanced and failsto affect or decreases fat intake (Ritter et al., 1999; Singer etal., 1998). 2DG, but not MA, elicits interoceptive sensorysignals similar to that of food deprivation (Benoit andDavidson, 1996). Moreover, whereas intravenous and centralglucose selectively and significantly reduces 2DG-inducedfeeding, MA-induced feeding is significantly reduced byintravenous glucose, lipid or fructose pretreatment but isunaffected by central glucose (Singer and Ritter, 1994, 1996).Whereas rat strains that are differentially susceptible todietary obesity show similar responses to 2DG-inducedfeeding, MA is more effective in eliciting feeding in thediet-susceptible strain relative to the diet-resistant strain(Singer et al., 1997).
Food intake in the three homeostatically driven para-digms inducing deprivation, glucoprivation and lipoprivationrespectively was measured as total consumption only anddid not examine any potential GABA-induced changes inhedonic responses. The NAC shell and VTA have beenconsidered to be reward-related sites and thereby implicatedin the appetitive rather than consummatory aspects of
ingestive behavior (e.g., see reviews: Kelley, 1999, 2004;Salamone, 1994), or “wanting” rather than “liking”, usingtaste-reactivity paradigms (see review: Berridge, 1996, 2004;Berridge and Robinson, 1998). GABA has been implicated inthese latter hedonic issues such that intra-VTA GABAdecreased food approach behavior at doses that failed toaffect spontaneous consumption (Ikemoto and Panksepp,1996), whereas systemic administration of the benzodiaze-pine diazepam facilitated positive palatability reactions(Treit and Berridge, 1990). Specific regions within the NACshell mediate GABAA-agonist induced behaviors with therostral region increasing feeding, visits to food, placepreferences and positive hedonic reactions and the caudalregion increasing defensive and burial behaviors (Reynoldsand Berridge, 2001, 2002). Taste reactivity paradigms dif-ferentiating “wanting” and “liking” are usually employed inshort-term tests immediately after a given manipulation. YetGABAA and GABAB receptor antagonists administered intothe NAC shell, but not the VTA, significantly reduced short-term (1–4 h) deprivation-induced feeding without affectingthe initiation of intake after 0.5 h, suggesting that theseantagonists were ineffective during the peak period whenone would ordinarily observe taste reactivity results. Theeffectiveness of both bicuculline and saclofen in the NACshell and VTA in immediately reducing lipoprivic feedingelicited by MA, but not glucoprivic, feeding elicited by 2DGindicates the participation of GABA in these sites in thedissociative responses elicited by these two challenges andopens the possibility that changes in hedonic and appetitivefactors may also play a role in this lipoprivic response.Interestingly, additional recent evidence suggests that GABAneurons in the NAC shell influence hypothalamic effectormechanisms for feeding motor patterns (e.g., consummatorycomponent: Baldo et al., 2004; Kelley et al., 2005) but fail toalter either lever pressing or the break point for foodacquisition (e.g., appetitive component: Hanlon et al., 2004;Zhang et al., 2003). Taken together, these data reinforce thefindings that GABAA and GABAB receptors in the NAC shelland VTA are not only important in the modulation ofpharmacologically induced feeding responses, but alsodifferentially participate in mediating feeding responses toimportant homeostatic challenges.
4. Experimental procedures
4.1. Subjects, surgery and histology
All of the following experiments and procedures wereapproved by the Queens College Institutional Animal Careand Use Committee. Fifty-one adult male albino Sprague–Dawley rats (Charles River Laboratories, Kingston, NY, 80–120days of age) were housed individually in wire mesh cages andmaintained on a 12 h light/12 h dark cycle with Purina ratchow and water available ad libitum. Each rat was pretreatedwith chlorpromazine (3 mg/kg, i.p.) and anesthetized withKetamine HCl (120 mg/kg, i.m.). Stainless steel guide cannulae(26-gauge, Plastics One) were aimed stereotaxically (KopfInstruments) at bilateral placements in the NAC shell in 24rats using the following coordinates: incisor bar (+5 mm),

94 B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
3.1 mm anterior to the bregma suture, 1.7 mm and angled 10°towards each side of the sagittal suture and 6.8 mm from thetop of the skull; and unilaterally in the VTA in 27 rats using thefollowing coordinates: incisor bar (+5mm), 5.6mmposterior tothe bregma suture, 2.5 mm lateral to and angled 10° towardsthe sagittal suture and 8.4 mm from the top of the skull. Thecannulae were secured to the skull by four anchor screws withdental acrylic. To allow full drug clearance, all animals wereallowed at least 2 weeks to recover from stereotaxic surgerybefore behavioral testing began. At the completion of testing,all rats were overdosedwith an anesthetic (Euthasol) andwereinjected transcardially with potassium chloride (15 mg/ml,0.9% saline). Transcardiac perfusions were performed with0.9% normal saline followed by 10% buffered formalin. Coronal40-μm sections, stained with Cresyl violet, were examined bylight microscopy by an observer unfamiliar with the behav-ioral data; only animals with confirmed cannula placementswere included in the data analysis.
4.2. Drugs
Bicuculline and saclofen were purchased from Aldrich-SigmaChemical Company (St. Louis, MO) and were dissolved indistilled water or in 0.1 M NaOH respectively. All microinjec-tions were administered in 0.5 μl volumes over 30 s through astainless internal cannula (33-gauge, Plastics One) thatextended 0.5 mm past the guide cannula and was connectedby polyethylene tubing to a Hamilton microsyringe. Thisrelatively high injection volume was necessary because of thelimited solubility of some of the pharmacological agents. Thedoses of bicuculline (75 and 150 ng) and saclofen (1.5 and 3 μg)and antagonist pretreatment interval (20 min) were chosen onthe basis of their effectiveness in previous studies in the NACshell and VTA in altering GABA and opioid-agonist-inducedfeeding (Ackerman et al., 2003; Echo et al., 2002; Khaimova etal., 2004; Stratford and Kelley, 1997; Znamensky et al., 2001).Particular microinjection conditions were presented in coun-terbalanced order at specified (at least weekly) intervals tominimize potential order and carry-over effects in all exper-imental protocols. 2DG and MA were purchased from Aldrich-Sigma Chemical Company and were dissolved in 0.9% saline.The doses of 2DG (500 mg/kg) and MA (70 mg/kg) were chosenon the basis of their effectiveness in inducing feeding that wasaltered by central pharmacological treatments (e.g., Bodnar etal., 1995; Ragnauth et al., 1997; Stein et al., 2000).
4.3. Deprivation-induced intake protocol
To assess the effects of food deprivation upon intake and bodyweight, all rats with NAC shell (n = 8) and VTA (n = 11) cannulaeunderwent an initial 4-day protocol beginning at approxi-mately 3 h into the light cycle. At the beginning of Day 1, allanimals were weighed (Pre-Deprivation Weight) and thenfood-deprived (water continuously available) for the next 24 h.On Day 2, all animals were re-weighed (Post-DeprivationWeight) to assess deprivation-induced weight loss andreceived a vehicle (0.9% saline, 0.5 μl) microinjection. Twentyminutes thereafter, food was reintroduced by placing pre-weighed pellets on the floor of the wire mesh cages, andcumulative deprivation-induced intakes, adjusted for spillage
collected beneath each cage by paper towels, were assessed0.5, 1, 2 and 4 h thereafter. Pre-weighed food bins and waterwere thenmade available for the next 44 h, allowing for overall24 h intake to be measured at the beginning of Days 3 (24 hIntake) and 4 (48 h Intake). Rats were re-weighed again onDays 3 and 4 to assess recovery of body weight 24 and 48 hfollowing reintroduction of food. Rats in each site werematched for the magnitude of deprivation-induced intakefollowing vehicle treatment for testing under the differentdrug conditions. A 2-week interval elapsed between subse-quent deprivation conditions in which all rats were observedto completely recover normal body weights. The eight ratswith NAC shell cannulae received the following bilateralmicroinjection conditions following 24 h of food deprivation:bicuculline at total doses of 75 (n = 7) and 150 (n = 7) ng (37.5and 75 ng/0.5 μl each side respectively) and saclofen at totaldoses of 1.5 (n = 8) and 3 (n = 8) μg (0.75 and 1.5 μg/0.5 μl eachside respectively). The eleven rats with a VTA cannulareceived the following microinjection conditions following24 h of food deprivation: bicuculline at total doses of 75 (n = 7)and 150 (n = 7) ng and saclofen at total doses of 1.5 (n = 7)and 3 (n = 7) μg. Food was reintroduced 20 min after themicroinjections.
4.4. 2DG intake protocol
Rats with bilateral NAC shell cannulae (n = 8) and unilateralVTA cannulae (n = 7) received the following conditions atweekly intervals: vehicle (0.5 μl bilaterally in NAC)/vehicle(1 ml/kg 0.9% normal saline, i.p.) and vehicle/2DG (500 mg/kg) with 20 min elapsing between central and systemicinjections. Pre-weighed food pellets were placed on the floorof the wire mesh cages, and cumulative intakes, adjusted forspillage, were assessed 1, 2 and 4 h after systemic drugtreatment. Rats in each site were matched for the magnitudeof vehicle/2DG-induced intake for testing under the differentdrug conditions. The eight rats with NAC shell cannulaereceived the following bilateral microinjection conditions incounterbalanced order at weekly intervals 20 min prior to2DG (500 mg/kg): bicuculline at total doses of 75 (n = 6) and150 (n = 6) ng and saclofen at total doses of 1.5 (n = 6) and 3(n = 6) μg. The seven rats with a VTA cannula received thefollowing microinjection conditions in counterbalanced orderat weekly intervals 20 min prior to 2DG (500 mg/kg): bicucul-line at total doses of 75 (n = 6) and 150 (n = 6) ng and saclofenat total doses of 1.5 (n = 6) and 3 (n = 6) μg.
4.5. MA intake protocol
Rats with bilateral NAC shell cannulae (n = 8) and unilateralVTA cannulae (n = 9) received the following conditions atweekly intervals: vehicle (0.5 μl bilaterally in NAC)/vehicle(1 ml/kg 0.9% normal saline, i.p.) and vehicle/MA (70 mg/kg)with 20 min elapsing between central and systemic injec-tions. Pre-weighed food pellets were placed on the floor ofthe wire mesh cages, and cumulative intakes, adjusted forspillage, were assessed 1, 2 and 4 h after systemic drugtreatment. Rats in each site were matched for the magnitudeof vehicle/MA-induced intake for testing under the differentdrug conditions. The 8 rats with NAC shell cannulae received

95B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
the following bilateral microinjection conditions in counter-balanced order at weekly intervals 20 min prior to MA(70 mg/kg): bicuculline at total doses of 75 (n = 6) and 150(n = 6) ng and saclofen at total doses of 1.5 (n = 6) and 3(n = 6) μg. The 9 rats with a VTA cannula received thefollowing microinjection conditions in counterbalancedorder at weekly intervals 20 min prior to MA (70 mg/kg):bicuculline at total doses of 75 (n = 6) and 150 (n = 6) ng andsaclofen at total doses of 1.5 (n = 6) and 3 (n = 6) μg.
4.6. Statistical analyses
Two-way repeated-measures analyses of variance were per-formed to assess short-term (0.5–4 h) and longer-term (24–48 h) alterations in deprivation-induced intake as well as bodyweight recovery (24–48 h) for each site with experimentalcondition as one variable and time as the second variable. Aone-way repeated-measures analysis of variance was per-formed to assess deprivation-induced weight loss amongexperimental conditions for each site. In the 2DG and MAprotocols, two-way randomized-block analyses of variancewere performed to assess alterations in intake for each sitewith experimental condition as the between-subject variableand time as the within-subject variable. Tukey comparisons(P < 0.05) were used to assess significant alterations followingantagonist treatment relative to corresponding deprivation,glucoprivic or lipoprivic treatment.
Acknowledgments
This research was supported in part by PSC/CUNY Grants65285 and 66325 to RJB; YI was a student in the CUNY HonorsCollege at Queens College.
R E F E R E N C E S
Ackerman, T.F., Lamonte, N., Bodnar, R.J., 2003. Lack of intersiteGABA receptor subtype antagonist effects upon mu opioidreceptor agonist-induced feeding elicited from either theventral tegmental area or nucleus accumbens shell in rats.Physiol. Behav. 79, 191–198.
Addae, J.I., Rothwell, N.J., Stock, M.J., Stone, T.W., 1986. Activationof thermogenesis of brown fat in rats by baclofen.Neuropharmacology 25, 627–631.
Arnt, J., Scheel-Kruger, J., 1979. GABA in the ventral tegmentalarea: differential regional effects on locomotion, aggressionand food intake after microinjection of GABA agonists andantagonists. Life Sci. 25, 1351–1360.
Baldo, B.A., Gual-Bonilla, L., Sijapati, K., Daniel, R.A., Landry, C.F.,Kelley, A.E., 2004. Activation of a subpopulation of orexin/hypocretin-containing hypothalamic neurons by GABAAreceptor-mediated inhibition of the nucleus accumbens shell,but not by exposure to a novel environment. Eur. J. Neurosci.19, 376–386.
Baldo, B.A., Alsene, K.M., Negron, A., Kelley, A.E., 2005.Hyperphagia induced by GABAA receptor-mediated inhibitionof the nucleus accumbens shell: dependence on intact neuraloutput from the central amygdaloid region. Behav. Neurosci.119, 1195–1206.
Basso, A.M., Kelley, A.E., 1999. Feeding induced by GABA(A)receptor stimulation within the nucleus accumbens shell:
regional mapping and characterization of macronutrient andtaste preference. Behav. Neurosci. 113, 324–336.
Bauche, F., Sabourault, D., Giudicelli, Y., Nordmann, J., Nordmann,R., 1981. 2-Mercaptoacetate administration depresses thebeta-oxidation pathway through an inhibition of long-chainacyl-CoA dehydrogenase activity. Biochem. J. 196,803–809.
Bendotti, C., Berettera, C., Invernizzi, R., Samanin, R., 1986.Selective involvement of dopamine in the nucleus accumbensin the feeding response elicited by muscimol injection in thenucleus raphe dorsalis of sated rats. Pharmacol. Biochem.Behav. 24, 1189–1193.
Benoit, S.C., Davidson, T.L., 1996. Interoceptive sensory signalsproduced by 24-hr food deprivation, pharmacologicalglucoprivation, and lipoprivation. Behav. Neurosci. 110,168–181.
Berridge, K.C., 1996. Food reward: substrates of wanting and liking.Neurosci. Biobehav. Rev. 20, 1–25.
Berridge, K.C., 2004. Motivation concepts in behavioralneuroscience. Physiol. Behav. 81, 179–209.
Berridge, K.C., Robinson, T.E., 1998. What is the role of dopaminein reward: hedonic impact, reward learning, or incentivesalience. Brain Res. Rev. 28, 309–369.
Blavet, N., DeFeudis, F.V., Clostre, F., 1982. THIP inhibits feeding infasted rats. Psychopharmacology 76, 75–78.
Bodnar, R.J., Glass, M.J., Ragnauth, A., Cooper, M.L., 1995. General,mu and kappa opioid antagonists in the nucleus accumbensalter food intake under deprivation, glucoprivic and palatableconditions. Brain Res. 700, 205–212.
Borsini, F., Bendotti, C., Przewlocka, B., Samanin, R., 1983.Monoamine involvement in the overeating caused bymuscimol injection in the rat nucleus raphe dorsalis and theeffects of D-fenfluramine and D-amphetamine. Eur. J.Pharmacol. 94, 109–115.
Calingasan, N.Y., Ritter, S., 1993. Lateral parabrachialsubnucelus lesions abolish feeding induced bymercaptoacetate but not by 2-deoxy-D-glucose. Am. J.Physiol. 265, R1168–R1178.
Coppola, J.D., Horowitz, B.A., Hamilton, J., Blevins, J.E., McDonald,R.B., 2005. Reduced feeding response to muscimol andneuropeptide Y in senescent F344 rats. Am. J. Physiol. 288,R1492–R1498.
Echo, J.A., Lamonte, N., Ackerman, T.F., Bodnar, R.J., 2002.Alterations of food intake elicited by GABA and opioid agonistsand antagonists administered into the ventral tegmental area.Physiol. Behav. 76, 107–116.
Fisler, J.S., Shimizu, H., Bray, G.A., 1989. Brain 3-hydroxybutyrate,glutamate, and GABA in a rat model of dietary obesity. Physiol.Behav. 45, 571–577.
Grandison, L., Guidotti, A., 1977. Stimulation of food intake bymuscimol and beta-endorphin. Neuropharmacology 16,533–536.
Hanlon, E.C., Baldo, B.A., Sadeghian, K., Kelley, A.E., 2004.Increases in food intake or food-seeking behavior induced byGABAergic, opioid or dopaminergic stimulation of thenucleus accumbens: is it hunger? Psychopharmacology 172,241–247.
Ikemoto, S., Panksepp, J., 1996. Dissociations between appetitiveand consummatory responses by pharmacologicalmanipulations of reward-relevant brain regions. Behav.Neurosci. 110, 331–345.
Kelley, A.E., 1999. Neural integrative activities of nucleusaccumbens subregions in relation to learning and motivation.Psychobiology 27, 198–213.
Kelley, A.E., 2004. Ventral striatal control of appetitive motivation:role in ingestive behavior and reward-related learning.Neurosci. Biobehav. Rev. 27, 765–776.
Kelly, J., Alheid, G.F., Newberg, A., Grossman, S.P., 1977. GABAstimulation and blockade in the hypothalamus and midbrain:

96 B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
effects of feeding and locomotor activity. Pharmacol. Biochem.Behav. 7, 537–541.
Kelley, A.E., Bless, E.P., Swanson, C.J., 1996. Investigation of theeffects of opiate antagonists infused into the nucleusaccumbens on feeding and sucrose drinking in rats.J. Pharmacol. Exp. Ther. 278, 1499–1507.
Kelley, A.E., Baldo, B.A., Pratt, W.E., Will, M.J., 2005.Corticostriatal–hypothalamic circuitry and food motivation:integration of energy, action and reward. Physiol. Behav. 86,773–795.
Khaimova, E., Kandov, Y., Israel, Y., Cataldo, G., Hadjimarkou, M.M., Bodnar, R.J., 2004. Opioid receptor subtype antagonistsdifferentially alter GABA agonist-induced feeding elicited fromeither the nucleus accumbens shell or ventral tegmental arearegions in rats. Brain Res. 1026, 284–294.
Klitenick, M.A., Wirtshafter, D., 1988. Comparative studies of theingestive behaviors produced by microinjections ofmuscimol into the midbrain raphe nuclei of the ventraltegmental area of the rat. Life Sci. 42, 775–782.
Klitenick, M.A., Wirtshafter, D., 1989. Elicitation of feeding,drinking and gnawing following microinjections of muscimolinto the median raphe nucleus of rats. Behav. Neural Biol. 51,436–441.
Lamonte, N., Echo, J.A., Ackerman, T.F., Christian, G., Bodnar, R.J.,2002. Analysis of opioid receptor subtype antagonist effectsupon mu opioid agonist-induced feeding elicited from theventral tegmental area of rats. Brain Res. 929, 96–100.
Langhans, W., Scharrer, E., 1987. Evidence for a vagallymediated satiety signal derived from hepatic fatty acidoxidation. J. Auton. Nerv. Syst. 18, 13–18.
Lenin Kamatchi, G., Bhakthavatsalam, P., Chandra, D., Bapna, J.S.,1984. Inhibition of insulin hyperphagia by gamma aminobutyric acid antagonists in rats. Life Sci. 34, 2297–2301.
Lenin Kamatchi, G., Veeraragavan, K., Chandra, D., Bapna, J.S.,1986. Antagonism of acute feeding response to 2-deoxyglucoseand 5-thioglucose by GABA antagonists: the relative role ofventromedial and lateral hypothalamus. Pharmacol. Biochem.Behav. 25, 59–62.
Maldonado-Irizarry, C.S., Swanson, C.J., Kelley, A.E., 1995.Glutamate receptors in the nucleus accumbens shell controlfeeding behavior via the lateral hypothalamus. J. Neurosci. 15,6779–6788.
Minano, F.J., Meneres Sancho, M.S., Sancibrian, M., Salinas, P.,Myers, R.D., 1992. GABA-A receptors in the amygdala:role in feeding in fasted and satiated rats. Brain Res. 586,104–110.
Nisoli, E., Garosi, V., Blundell, J.E., Carruba, M.O., 1996. Salbutamolantagonizes insulin- and sodium mercaptoacetate-inducedbut not 2-deoxy-D-glucose-induced hyperphagia. Pharmacol.Biochem. Behav. 54, 409–413.
Nobrega, J.N., Coscina, D.V., 1982. Inhibition of acute feedingresponses to systemic 2-deoxy-D-glucose and insulin in ratspretreated with the GABA-transaminase blockerethanolamine-O-sulfate (EOS). Pharmacol. Biochem. Behav. 17,1145–1148.
Olgiati, V.R., Netti, C., Guidobono, F., Pecile, A., 1980. Thecentral GABAergic system and control of food intake underdifferent experimental conditions. Psychopharmacology 68,163–167.
Paxinos, G., Watson, C., 1998. The Rat Brain in StereotaxicCoordinates, Fourth ed. NeuroScience Associates,Knoxville, TN.
Pu, S., Jain, M.R., Horvath, T.L., Diano, S., Kalra, P.S., Kalra, S.P.,1999. Interactions between neuropeptide Y andgamma-aminobutyric acid in stimulation of feeding: amorphological and pharmacological analysis. Endocrinology140, 933–940.
Ragnauth, A., Ruegg, H., Bodnar, R.J., 1997. Evaluation of opioidreceptor subtype antagonist effects in the ventral tegmental
area upon food intake under deprivation, glucoprivic andpalatable conditions. Brain Res. 767, 8–16.
Ragnauth, A., Moroz, M., Bodnar, R.J., 2000. Multiple opioidreceptors mediate feeding elicited by mu and delta opioidagonists in the shell region of the nucleus accumbens. BrainRes. 876, 65–72.
Redgrave, P., Dean, P., Taha, E.B., 1984. Feeding induced byinjections of muscimol into the substantia nigra of rats:unaffected by haloperidol but abolished by large lesions of thesuperior colliculus. Neuroscience 13, 77–85.
Reynolds, S.M., Berridge, K.C., 2001. Fear and feeding in thenucleus accumbens shell: rostrocaudal segregation ofGABA-elicited defensive behavior versus eating behavior.J. Neurosci. 21, 3261–3270.
Reynolds, S.M., Berridge, K.C., 2002. Positive and negativemotivation in nucleus accumbens shell: bivalent rostrocaudalgradients for GABA-elicited eating, taste “liking”/“disliking”reactions, place preference/avoidance, and fear. J. Neurosci. 22,7308–7320.
Ritter, S., Dinh, T.T., 1994. 2-Mercaptoacetate and 2-deoxy-D-glucoseinduce fos-like immunoreactivity in rat brain. Brain Res. 641,111–120.
Ritter, S., Hutton, B., 1995. Mercaptoacetate-induced feeding isimpaired by central nucleus of the amygdala lesions. Physiol.Behav. 58, 1215–1220.
Ritter, S., Taylor, J.S., 1989. Capsaicin abolishes lipoprivic but notglucoprivic feeding in rats. Am. J. Physiol. 256, R1232–R1239.
Ritter, S., Taylor, J.S., 1990. Vagal sensory neurons are required forlipoprivic but not glucoprivic feeding in rats. Am. J. Physiol. 258,R1395–R1401.
Ritter, S., Scheurink, A., Singer, L.K., 1995. 2-Deoxy-D-glucose butnot 2-mercaptoacetate increases Fos-like immunoreactivity inadrenal medulla and sympathetic preganglionic neurons.Obes. Res. 3, 729S–734S.
Ritter, S., Ritter, J.B., Cromer, L., 1999. 2-Deoxy-D-glucose andmercaptoacetate induce different patterns of macronutrientingestion. Physiol. Behav. 66, 709–715.
Salamone, J.D., 1994. The involvement of nucleus accumbensdopamine in appetitive and aversive motivation. Behav. BrainRes. 61, 117–133.
Salinas, J.A., McGaugh, J.L., 1996. The amygdala modulatesmemory for changes in reward magnitude: involvement of theamygdaloid GABAergic system. Behav. Brain Res. 80, 87–98.
Sanger, D.J., 1984. Chlordiazepoxide-induced hyperphagia in rats:lack of effect of GABA agonists or antagonists.Psychopharmacology 84, 388–392.
Scharrer, E., Langhans, W., 1986. Control of food intake by fattyacid oxidation. Am. J. Physiol. 250, R1003–R1006.
Scheurink, A., Ritter, S., 1993. Sympathoadrenal responses toglucoprivation and lipoprivation in rats. Physiol. Behav. 53,995–1000.
Singer, L.K., Ritter, S., 1994. Differential effects of infused nutrientson 2-deoxy-D-glucose- and 2-mercaptoacetate-inducedfeeding. Physiol. Behav. 56, 193–196.
Singer, L.K., Ritter, S., 1996. Intraventricular glucose blocks feedinginduced by 2-deoxy-D-glucose but not mercaptoacetate.Physiol. Behav. 59, 921–923.
Singer, L.K., York, D.A., Bray, G.A., 1997. Feeding responses tomercaptoacetate in Osborne-Mendel and S5B/PL rats. Obes.Res. 5, 587–594.
Singer, L.K., York, D.A., Bray, G.A., 1998. Macronutrient selectionfollowing 2-deoxy-D-glucose and mercaptoacetateadministration in rats. Physiol. Behav. 65, 115–121.
Smith, G.P., Epstein, A.N., 1969. Increased feeding response todecreased glucose utilization in the rat and the monkey. Am. J.Physiol. 217, 1083–1087.
Smith, G.P., Root, A.W., 1969. Effects on feeding on hormonalresponses to 2-deoxy-D-glucose in conscious monkeys.Endocrinology 85, 963–966.

97B R A I N R E S E A R C H 1 0 8 2 ( 2 0 0 6 ) 8 6 – 9 7
Soderpalm, A.H., Berridge, K.C., 2000. Food intake afterdiazepam, morphine or muscimol: microinjections in thenucleus accumbens shell. Pharmacol. Biochem. Behav. 66,429–434.
Stein, J.A., Znamensky, V., Baumer, F., Rossi, G.C., Pasternak, G.W.,Bodnar, R.J., 2000. Mercaptoacetate induces feeding throughcentral opioid-mediated mechanisms in rats. Brain Res. 864,240–251.
Stratford, T.R., Kelley, A.E., 1997. GABA in the nucleus accumbensshell participates in the central regulation of feeding behavior.J. Neurosci. 17, 4434–4440.
Stratford, T.R., Kelley, A.E., 1999. Evidence of a functionalrelationship between the nucleus accumbens shell and lateralhypothalamus subserving the control of feeding behavior.J. Neurosci. 19, 11040–11048.
Stratford, T.R., Wirtshafter, D., 2004. NPY mediates the feedingelicited by muscimol injections into the nucleus accumbensshell. NeuroReport 15, 2673–2676.
Treit, D., Berridge, K.C., 1990. A comparison of benzodiazepine,serotonin, and dopamine agents in the taste-reactivityparadigm. Pharmacol. Biochem. Behav. 37, 451–456.
Truong, B.G., Magrum, L.J., Gietzen, D.W., 2002. GABA(A) and GABA(B) receptors in the anterior piriform cortex modulate feedingin rats. Brain Res. 924, 1–9.
Tsujii, S., Bray, G.A., 1991. GABA-related feeding control ingenetically obese rats. Brain Res. 540, 48–54.
Van Bockstaele, E.J., Pickel, V.M., 1995. GABA-containing neuronsin the ventral tegmental area project to the nucleusaccumbens in rat brain. Brain Res. 682, 215–221.
Ward, B.O., Somerville, E.M., Clifton, P.G., 2000. Intraccumbensbaclofen selectively enhances feeding behavior in the rat.Physiol. Behav. 68, 463–468.
Wick, A.N., Drury, R., Nakada, H.I., Wolfe, J.B., 1957. Localization ofthe primary metabolic block produced by 2-deoxy-D-glucose.J. Biol. Chem. 224, 963–979.
Will, M.J., Franzblau, E.B., Kelley, A.E., 2003. Nucleus accumbensmu-opioids regulate intake of a high-fat diet via activation of adistributed brain network. J. Neurosci. 23, 2882–2888.
Will, M.J., Franzblau, E.B., Kelley, A.E., 2004. The amygdala iscritical for opioid-mediated binge eating of fat. NeuroReport 15,1857–1860.
Wirtshafter, D., Stratford, T.R., Pitzer, M.R., 1993. Studies on thebehavioral activation produced by stimulation of GABA-Breceptors in the median raphe nucleus. Behav. Brain Res. 59,83–93.
Zhang, M., Balmadrid, C., Kelley, A.E., 2003. Nucleus accumbensopioid, GABAergic, and dopaminergic modulation of palatablefood motivation: contrasting effects revealed by a progressiveratio study in the rat. Behav. Neurosci. 117, 202–211.
Zheng, H., Corkern, M., Stoyanova, I., Patterson, L.M., Tian, R.,Berthoud, H.R., 2003. Peptides that regulate food intake:appetite-inducing accumbens manipulation activateshypothalamic orexin neurons and inhibits POMC neurons. Am.J. Physiol. 284, R1436–R1444.
Znamensky, V., Echo, J.A., Lamonte, N., Christian, G., Ragnauth, A.,Bodnar, R.J., 2001. GABA receptor subtype antagonistsdifferentially alter opioid-induced feeding in the shell region ofthe nucleus accumbens. Brain Res. 906, 84–91.