functional modeling and phylogenetic distribution of putative cylindrospermopsin biosynthesis...
TRANSCRIPT

Functional Modeling and Phylogenetic Distribution of Putative
Cylindrospermopsin Biosynthesis Enzymes
Ralf Kellmann, Toby Mills, Brett A. Neilan
School of Biotechnology and Biomolecular Sciences, Section Microbiology, The University of New South Wales, Biotechnology, Sydney,
N.S.W. 2052, Australia
Received: 2 February 2005 / Accepted: 14 June 2005 [Reviewing Editor: Dr. Martin Kreitman]
Abstract. The alkaloid cylindrospermopsin is themost recently discovered cyanotoxin and has causedepidemic outbreaks of human poisoning. Cylindro-spermopsin producing cyanobacteria have in recenttimes appeared in countries all over the world wherethey had not been observed previously and, thus,represent a global public health concern. Threeputative cylindrospermopsin biosynthesis genes,encoding an amidinotransferase (aoaA), a nonribos-omal peptide synthetase (aoaB), and a polyketidesynthase (aoaC), have been described. Most cyano-toxins are the product of nonribosomal peptide andpolyketide synthesis, but the involvement of an ami-dinotransferase is novel. In the present study, func-tional modeling was carried out to gain insight intothe mechanism of precursor recruitment in cylindro-spermopsin biosynthesis. In addition, the molecularphylogenies of putative cylindrospermopsin biosyn-thesis genes and producer organisms were determined.The model indicated that AoaA may catalyze theformation of guanidino acetate from glycine andarginine. The catalytic site of the AoaB adenylationdomain provided two aspartate residues, instead ofthe usual one, which may be involved in the binding ofthe guanidino moiety of guanidino acetate. Molecularphylogenetic analysis grouped cylindrospermopsinproducing cyanobacteria into two divergent groups.Although the phylogeny of the cylindrospermopsinbiosynthesis genes followed that of the producerorganisms, they were less divergent, which may indi-cate the recent horizontal transfer of these genes.
Key words: Cylindrospermopsin — Biosynthesisgenes — Protein modeling — Phylogeny — Ami-dinotransferase nonribosomal peptide — Polyketide
Introduction
Cylindrospermopsin (CYLN) is an alkaloid that isproduced by various cyanobacteria, includingCylindrospermopsis raciborskii, Aphanizomenonovalisporum, Anabaena bergii, Umezakia natans, andRaphidiopsis curvata (Banker et al. 1997; Haradaet al. 1994; Li et al. 2001; Schembri et al. 2001). Itinterferes with multiple metabolic pathways, causinghepatotoxicity (Hawkins et al. 1985), general cyto-toxicity (Runnegar et al. 1994), and neurotoxicity(Kiss et al. 2002), while it may also induce carcino-genesis (Humpage et al. 2000). Human poisoning byCYLN has been reported from Australia after theingestion of contaminated drinking water and iscolloquially known as the ‘‘Palm Island MysteryDisease’’ (Bourke et al. 1983), ‘‘Barcoo Disease,’’ or‘‘Belyando Fever’’ (Hayman 1992). Although inci-dents of CYLN poisoning have been limited toAustralia so far (Saker et al. 1999b), the producerorganisms are recent and highly invasive neophytes intemperate and tropical regions all over the world(Neilan et al. 2003). CYLN thus represents a publichealth concern on a global scale.
Themolecular structure, biosynthetic pathway, andputative biosynthesis enzymes of CYLN have beenrecently described (Burgoyne et al. 2000; Ohtani et al.1992; Schembri et al. 2001; Shalev-Alon et al. 2002). AsCorrespondence to: Brett Neilan; email: [email protected]
J Mol Evol (2006) 62:267–280DOI: 10.1007/s00239-005-0030-6
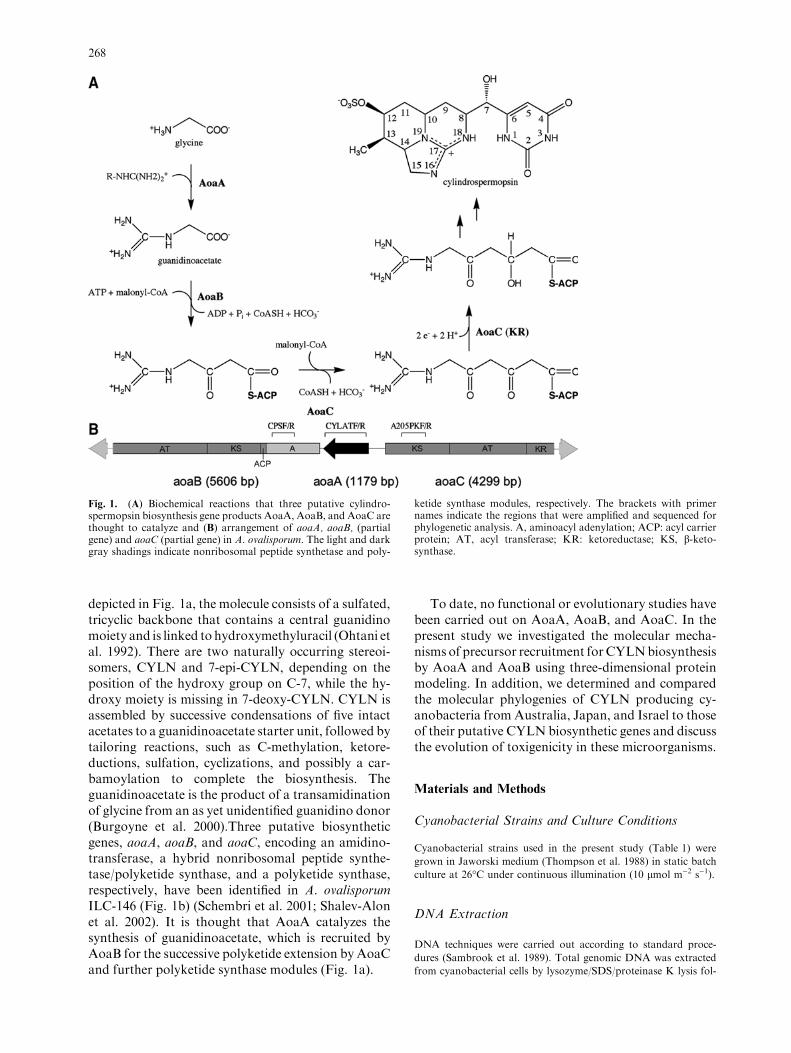
depicted in Fig. 1a, the molecule consists of a sulfated,tricyclic backbone that contains a central guanidinomoiety and is linked to hydroxymethyluracil (Ohtani etal. 1992). There are two naturally occurring stereoi-somers, CYLN and 7-epi-CYLN, depending on theposition of the hydroxy group on C-7, while the hy-droxy moiety is missing in 7-deoxy-CYLN. CYLN isassembled by successive condensations of five intactacetates to a guanidinoacetate starter unit, followed bytailoring reactions, such as C-methylation, ketore-ductions, sulfation, cyclizations, and possibly a car-bamoylation to complete the biosynthesis. Theguanidinoacetate is the product of a transamidinationof glycine from an as yet unidentified guanidino donor(Burgoyne et al. 2000).Three putative biosyntheticgenes, aoaA, aoaB, and aoaC, encoding an amidino-transferase, a hybrid nonribosomal peptide synthe-tase/polyketide synthase, and a polyketide synthase,respectively, have been identified in A. ovalisporumILC-146 (Fig. 1b) (Schembri et al. 2001; Shalev-Alonet al. 2002). It is thought that AoaA catalyzes thesynthesis of guanidinoacetate, which is recruited byAoaB for the successive polyketide extension by AoaCand further polyketide synthase modules (Fig. 1a).
To date, no functional or evolutionary studies havebeen carried out on AoaA, AoaB, and AoaC. In thepresent study we investigated the molecular mecha-nisms of precursor recruitment forCYLNbiosynthesisby AoaA and AoaB using three-dimensional proteinmodeling. In addition, we determined and comparedthe molecular phylogenies of CYLN producing cy-anobacteria from Australia, Japan, and Israel to thoseof their putative CYLN biosynthetic genes and discussthe evolution of toxigenicity in these microorganisms.
Materials and Methods
Cyanobacterial Strains and Culture Conditions
Cyanobacterial strains used in the present study (Table 1) were
grown in Jaworski medium (Thompson et al. 1988) in static batch
culture at 26�C under continuous illumination (10 lmol m)2 s)1).
DNA Extraction
DNA techniques were carried out according to standard proce-
dures (Sambrook et al. 1989). Total genomic DNA was extracted
from cyanobacterial cells by lysozyme/SDS/proteinase K lysis fol-
Fig. 1. (A) Biochemical reactions that three putative cylindro-spermopsin biosynthesis gene products AoaA, AoaB, and AoaC arethought to catalyze and (B) arrangement of aoaA, aoaB, (partialgene) and aoaC (partial gene) in A. ovalisporum. The light and darkgray shadings indicate nonribosomal peptide synthetase and poly-
ketide synthase modules, respectively. The brackets with primernames indicate the regions that were amplified and sequenced forphylogenetic analysis. A, aminoacyl adenylation; ACP: acyl carrierprotein; AT, acyl transferase; KR: ketoreductase; KS, b-keto-synthase.
268

lowing phenol–chloroform extraction as described previously
(Neilan 1995). DNA in the supernatant was precipitated with 0.6
volumes isopropanol, washed with 70% ethanol, dissolved in TE
buffer (10:1), and stored at –20�C.
PCR Amplification and Sequencing
Fragments of the putative CYLN biosynthesis genes, aoaA, aoaB,
and aoaC (Fig. 1), and 16S rRNA gene were amplified by the
polymerase chain reaction (PCR) using the primer pairs listed in
Table 2. PCR was performed in 20-ll reaction volumes containing
1· Taq polymerase buffer, 2.5 mM MgCl2, 0.2 mM deoxynucleo-
tide triphosphates, 10 pmol forward and reverse primers, between
10 and 100 ng genomic DNA, and 0.2 units Taq polymerase
(Fischer Biotech, Perth, Australia). Thermal cycling was performed
in a GeneAmp PCR System 2400 Thermocycler (Perkin Elmer
Corp., Norwalk, CT, USA). Cycling began with a denaturing step
at 94�C for 3 min followed by 30 cycles of denaturing at 94�C for
10 s, primer annealing at 50�C or 55�C for 20 s, and a DNA strand
extension at 72�C for 1 min. Amplification was completed by a final
extension step at 72�C for 7 min. DNA was separated by agarose
gel electrophoresis in TAE buffer (40 mM Tris–acetate, 1 mM
EDTA, pH 7.8) and visualized by UV translumination after
staining in ethidium bromide (0.5 lg/ml).
Automated DNA sequencing was performed using the PRISM
Big Dye cycle sequencing system and a model 373 sequencer (Ap-
plied Biosystems Inc., Foster City, CA, USA). Sequence data were
Table 1. Cyanobacteria strains used in the present study
Strain Origin Toxicity Reference
C. raciborskii AWT205 NSW, AU + Hawkins et al. (1997)
C. raciborskii 05E NSW, AU + Schembri et al. (2001)
C. raciborskii 23B QSL, AU + Wilson et al. (2000)
C. raciborskii 23D QSL, AU + Wilson et al. (2000)
C. raciborskii 24C QSL, AU + Schembri et al. (2001)
C. raciborskii GOON QSL, AU + Saker and Griffiths (2000)
C. raciborskii SDC QSL, AU + Saker et al. (1999a)
C. raciborskii T3 Sao Paulo,BRZ ) Lagos et al. (1999)
C. raciborskii GERM1 Melangsee, D ) Neilan et al. (2003)
C. raciborskii GERM2 Melangsee, D ) Neilan et al. (2003)
C. raciborskii VOLL1 Melangsee, D ) Neilan et al. (2003)
C. raciborskii VOLL2 Melangsee, D ) NA
C. raciborskii HUNG1 Lake Bulton, H ) NA
C. raciborskii HUNG2 Lake Bulton, H ) NA
C. raciborskii MARAU1 Maranhao Res., P ) Neilan et al. (2003)
C. raciborskii 4799 Odivelas Res, P ) Neilan et al. (2003)
C. raciborskii CAIA Caia Res, P ) Neilan et al. (2003)
C. raciborskii FL-D Florida, USA ? Neilan et al. (2003)
C. raciborskii FL-F Florida, USA ? Neilan et al. (2003)
C. raciborskii FL-G Florida, USA ? Neilan et al. (2003)
C. raciborskii FL-I Florida, USA ? Neilan et al. (2003)
A. ovalisporum APH028A QSL, AU + Shaw et al. (1999)
U. natans TAC101 Lake Mikata, J + Harada et al. (1994)
Note. AU, Australia; BRZ, Brazil; D, Germany; H, Hungary; J, Japan; P, Portugal; USA, United States of America; NSW, New South
Wales; Queensland; Res, Reservoir; NA, not available; +, CYLN producer; –, non-producer of CYLN; ?, not tested.
Table 2. PCR primers used in the amplification and sequencing of 16S rDNA, putative amidinotransferase (AMT), peptide synthetase A-domain (NRPS), and polyketide synthetase b-ketosynthase domain (PKS) genes (the length of PCR products and the 5¢-binding sites aregiven in parentheses)
Primer Sequence Reference
16S rDNA (1367 bp)
27F1 (5¢-27) 5¢-AGAGTTTGATCCTGGCTCAG Neilan et al. (1997)
1494Rc (5¢-1494) 5¢-TACGCGCTACCTTGTTACGAC
NRPS (478 bp)
CPSF (5’-338) 5¢-AGTATATGTTGCGGGACTCG This study
CPSR (5’-816) 5¢-CCCGCCAAGACAGAGGGTAG
PKS (514 bp)
A205PKF (5¢-966) 5¢-AATGACAGAGACTTGTGCGGGG This study
A205PKR(5¢-452) 5¢-TTATCGGTATTGGTGGTAGCAACT
AMT (1105 bp)
CYLATF (5¢-13) 5¢-ATTGTAAATAGCTGGAATGAGTGG This study
CYLATR (5¢-1118) 5¢-TTAGGGAAGTAATCTTCACAG
269

analyzed using ABI Prism-Autoassembler software, and percentage
similarity and identity to other translated sequences determined
using BLAST in conjunction with the National Center for Bio-
technology Information (NIH, MD). Accession numbers are pre-
sented in the legends to Figs. 6a to 6d.
Phylogenetic Analysis
DNA sequences were aligned using the multiple alignment tool
from Clustal X (Thompson et al. 1997). Sequence alignments were
manually confirmed. Phylogenetic trees were reconstructed from a
pairwise distance matrix (Jukes and Cantor 1969) using neighbor-
joining (Saitou and Nei 1987). All alignments were bootstrapped
with 1000 resampling events. Alignments were also applied to
parsimony and maximum likelihood programs of the Phylip
package version 3.6 (Felsenstein 1989). Phylogenetic trees and se-
quence alignments were reproduced using the software NJPlot
(Perriere and Gouy 1996) and TeXshade (Beitz 2000), respectively.
Three-Dimensional Protein Modeling
A suitable template for the three-dimensional modeling of AaoA
was determined by a PSI-BLAST search of the nonredundant
PDB database using BLOSUM62 with 10 iterations (http://
bioinformatics.ljcrf.edu/pdb_blast/). A structural alignment was
obtained by sequence-structure comparison (FUGUE) of AoaA
against the HOMESTRAD database (Shi et al. 2001). The protein
was modeled using the interactive three-dimensional graphics
programs, O (Jones et al. 1991) and LSQMAN (Kleywegt 1996).
The AoaA model was prepared, using the human arginine:glycine
amidinotransferase (GATM) as a template structure (PDB acces-
sion no. 1JDW), by manually replacing each residue of GATM
with the corresponding amino acid of AoaA, as determined by the
structural alignment described above. Rotamer libraries were used
to replace side chains. Equivalent rotamers were used when
replacing a side chain unless it resulted in a steric clash. The final
model, consisting of residues 4 to 372, was verified using Verify3D
(Luthy et al. 1992). Substrate arginine was placed into the catalytic
site of the AoaA model by using the atom coordinates of substrate
arginine in GATM (PDB accession no. JDW4). Graphic repre-
sentations of protein models were prepared using the software
Pymol (Molecular Graphics System, deLano Scientific LLC).
Extensive modeling of nonribosomal peptide synthetase ade-
nylation domains (A-domains) has been carried out (Challis et al.
2000; Stachelhaus et al. 1999). Eight topologically conserved amino
acid residues determine the substrate specificity and were identified
for the A-domain of AoaB as described elsewhere (Challis et al.
2000; Stachelhaus et al. 1999). The substrate determining residues
were compared to an assigned database of residues, where the
substrate for the corresponding A-domain has been determined
experimentally, and an unassigned database, where the substrate is
putative or unknown (Challis et al. 2000).
Results and Discussion
Functional Modeling of AoaA
Burgoyne et al. (2000) have demonstrated that gua-nidino acetate is the starter unit in cylindrospermop-sin biosynthesis, however, they failed to demonstratehow guanidino acetate is produced. While the authorsdemonstrated that an intact molecule of glycine isincorporated into guanidino acetate, no incorpora-tion of label was achieved in feeding experiments with
ubiquitously 13C- and 15N-labeled arginine. Guani-dino acetate is found in a wide variety of organismsand is the product of amidinotransferases, a mono-phyletic enzyme family that is highly conserved inboth prokaryotic and eukaryotic organisms (Bedekaret al. 1998). While the natural guanidino acceptorvaries in amidinotransferases from different organ-isms, the preferred guanidino donor is arginine in allcases investigated (Bedekar et al. 1998; Lee et al. 2002;Srivenugopal and Adiga 1980). Considering the highlevel of conservation in the structure and function ofamidinotransferases, it is surprising that arginineshould not be the guanidino donor in the biosynthesisof guanidino acetate in C. raciborskii. On the otherhand, the failure to achieve incorporation of labelfrom arginine into cylindrospermopsin in the study byBurgoyne et al. (2000) is not conclusive evidence thatarginine is not the natural substrate in cylindrosper-mopsin biosynthesis. Reasons for this experimentalfailure can be manifold. For example, not all cyano-bacteria provide basic amino acid transporter systems(Montesinos et al. 1997), and evidence for the uptakeof arginine by C. raciborskii was not presented.
AoaA provided the highest amino acid sequencesimilarity (PSI-BLAST of PDB: 40% identities/56%similarities) to the human arginine:glycine amidino-transferase, GATM (PDB accession no. 1JDW). AFUGUE search revealed high structural similarityamong AoaA, GATM, and the Streptomyces griseusarginine:inosamine phosphate amidinotransferase,StrB1 (PDB accession no. 1BWD) (FUGUE Z-score = 42.29). As shown in the structural alignmentin Figure 2, residues forming hydrogen bonds or saltbridges with the substrate arginine (D170, M302,R305, R322, S354, S355) and those forming the cat-alytic triad (D254, H303, C407) were completelyconserved in GATM, AoaA, and StrB1, with theexception that M302 was substituted by serine inAoaA and deleted in StrB1. In contrast, substrateglycine binding residues were only conserved inGATM and AoaA, and not in StrB1, which utilizesinosamine phosphate as the natural guanidineacceptor (Fritsche et al. 1998).
Two secondary structures, helix H9 and 300-flap,are involved in conformational changes of GATMduring catalysis (Fritsche et al. 1999). Upon enteringthe catalytic channel, substrate arginine stericallyclashes with N300, displacing the 300-flap towardhelix H9, which in turn is displaced to avoid thesterical collision of P299, H236, and N300 with R235.The regions corresponding to H9 and the 300-flapwere structurally conserved in AoaA, but their aminoacid sequences differed from GATM. In particular,P299 has been replaced by isoleucine, and N300 byphenylalanine.
The three-dimensional structure of AoaA wasmodeled using the crystal structure of GATM
270

(1JDW) as a template. AoaA contained nine inser-tions (C105, C148, P149, V238, H239, Q270, E271,D295, E296), which were not modeled. The quality ofthe model was verified by plotting the mean 3DVerifyscore of both 1JDW and the AoaA model versus theresidue number (Fig. 3). The mean score of all resi-dues in the AoaA model (0.3778) was only slightlylower than that of the template (0.5051), indicatingthat the majority of residues were folded correctly.Two regions of the model (A42 to K52 and G226 toP238) provided residues with negative scores andwere probably folded incorrectly. Whereas the firstregion was located on the surface of the protein andprobably has only structural function, the secondregion included the 300-flap. Another misfold notevident from the 3DVerify score occurred near helixH9. The last four residues of the loop leading up toH9, PIHS in GATM, were replaced in AoaA byfunctionally different amino acids, WEWP. In themodel, the two hydrophobic tryptophans were ex-posed to the solvent at the surface of the enzyme,
while the charged glutamate was buried and theproline was twisted into an unlikely rotamer. Thismisfold had flow-on effects up to residue 250, whichincluded the entire helix H9.
In GATM, substrate arginine forms 11 hydrogenbonds to amino acid residues M302, H303, R322,S354, S355, and C407. All residues were 100%conserved in sequence and topology in the AoaAmodel, apart from M302, which was replaced byserine (S247). This replacement is a significantchange, as a large hydrophobic amino acid wasexchanged with a small polar amino acid, and itseffect on substrate specificity can only be speculatedon. The topolgies of substrate binding and catalyticresidues of AoaB are shown in Figure 4. InGATM, substrate arginine forms a hydrogen bondbetween its a-amino group and the main chaincarboxylate of M302. The residue corresponding toM302 has been deleted in StrB1, and substratearginine forms a hydrogen bond with the mainchain carboxylate of the adjacent T226 (Fritsche et
Fig. 2. Structural alignment of amidinotransferases. AoaA,A. ovalisporum amidinotransferase; AT, human amidinotransfer-ase; StrB1, S. griseus amidinotransferase. (h), nonconserved; (n),similar; (n), conserved; (n), all match. Bracket with arrow indicatesb-sheet; bracket indicates a-helix. H9, helix H9; flp, 300-flap. Let-
ters above alignment indicate substrate binding residues of AT;letters below alignment indicate substrate binding residues ofStrB1. G, glycine binding residue; I, inosamine phosphate bindingresidue; R, arginine binding residue. Amino acids in italics arecatalytic residues.
271

al. 1998). It cannot be excluded that S247 in AoaAperforms a similar function as M302 and T226 inGATM and StrB1, respectively.
Substrate glycine is held in the catalytic site ofGATM by hydrogen bonds from its carboxylateto the side chains of H303 and via a water molecule toY164, while the amino group is hydrogen-bonded tothe side chain of S354. These residues were 100%
conserved in both sequence and topology in theAoaA model.
Due to regional errors involving helix H9 and the300-flap, it was not possible to determine whetherAoaA would undergo the same substrate-inducedconformational changes as GATM. This is, however,not essential to the function of the enzyme but isthought to increase the catalytic rate by aiding in the
Fig. 3. Verify 3D mean score plottedversus residue number. Abbreviations asexplained in the text. (—) Mean score of1JDW; (—) mean score of AoaA model.
Fig. 4. Topology of substrate binding and catalyticresidues with arginine in the catalytic site of AoaA.
272

release of ornithine from the catalytic site (Fritscheet al. 1999).
The high structural similarity of the AoaA modelto GATM, in addition to the high conservation ofsubstrate binding residues, suggested that AoaA maycatalyze the transamidination of glycine from argi-nine. Biochemical studies on AoaA will be requiredto verify this claim.
Upon synthesis, guanidino acetate is thought to berecruited by the hybrid NRPS/PKS, AoaB, for poly-ketide extension (Burgoyne et al. 2000). So far, noother NRPS has been described that utilizes guanidinoacetate as a substrate. The eight substrate specificityconferring residues of the AoaB A-domain (DFH-FITHD), determined as described by Challis et al.(2000), did not closelymatch any otherA-domain fromeither the assigned or the unassigned databases (http://raynam.chm.jhu.edu/�nrps/). The most similarA-domains activated proline, methyl-proline, andpipecolic acid (Fig. 5), all of which provide modifieda-amino groups. Unfortunately, the overall sequencesimilarity of the AoaB A-domain to other A-domainswith a known crystal structurewas too low (<40%) forreliable modeling of its three-dimensional structure,and it could not be determined how the eight substratebinding residues described abovemay interact with theguanidino acetate substrate. One interesting feature ofAoaB is that the eighth substrate binding residues,which in other A-domains may be one of the hydro-
phobic amino acids leucine, isoleucine, or valine,consisted of an aspartate (D362). Like other A-do-mains, AoaB provided a conserved lysine (K558),which in GrsA forms a hydrogen bond to the a-car-boxylate of the substrate amino acid (Conti et al. 1997).AoaB also provided a conserved aspartate (D258), theside chain of which forms a second hydrogen bond tothe a-amino group of the substrate in GrsA. Using theatom coordinates of the substrate and substrate bind-ing residues of GrsA, and replacing these residues withthe corresponding residues of AoaB, a simple model ofthe catalytic site was prepared (data not shown). In thismodel, guanidino acetate was held in the catalytic siteby hydrogen bonds between its a-carboxylate and theK558 side chain, between its imino nitrogen and theD258 side chain, aswell as between its guanidino amineand the side chain of D362. Other residues, such asT330 and H262, may also play a role in substratebinding. A crystal structure is required to verify thisproposal.
Molecular Phylogeny of CYLN ProducingCyanobacteria
The molecular phylogeny of CYLN producing cy-anobacteria was investigated using partial 16S rDNAgene sequences (954 bp). A phylogenetic tree of the16S rDNA from CYLN producing strains and ref-erence strains is given in Fig. 6a. As already reportedelsewhere (Neilan et al. 2003), C. raciborskii strainsgroup closely together and form a separate branchwithin the order Nostocales. In contrast to theirmorphological classification, a close relationshipamong U. natans, A. bergii, and A. ovalisporum be-came apparent, and positioned these strains withinthe same group as Anabaenopsis and Cyanospira(Fig. 6a). Neilan et al. (2003) reported that U. natanshas a distant phylogenetic position to other cyano-bacteria, however, on closer inspection, the 16SrDNA determined in that study may have been aPCR hybrid. In the present study, the sequence sim-ilarity among U. natans, the Australian and Israeliisolates of A. ovalisporum, Anabaenopsis, andCyanospira was at least 98%, while that of A. bergiiwas slightly lower (97%). The Israeli isolateA. ovalisporum ILC-146 is not included in Figure 6a,since only a short fragment (479 bp; AF044269) wasavailable from the database. This fragment was,however, 99% identical to the corresponding regionof the A. ovalisporum APH028 16S rDNA. Othermembers belonging to the genera Aphanizomenon andAnabaena were located on a branch well separatedfrom the above stains, and provided a sequencesimilarity of 93% or less to the above strains.Unfortunately, no data or culture was available forthe CYLN-producer strain Raphidiopsis curvata, andtherefore its phylogenetic relationship to other
Fig. 5. Alignment of substrate binding residues of NRPS adeny-lation domains. (h), Nonconserved; (n), similar; (n), conserved;(n), all match. Access codes are given, followed by the proteinname, adenylation module number, and three-letter amino acidcode of the substrate amino acid. 3h4mPhe, 3-hydroxy-4-meth-oxyphenylalanine; mePro, methylproline; DAB, diaminobutyricacid; DhbF, 2,3-dihydroxybenzoylglycine synthetase; NosA/C,nostopeptolide synthetase; SafA, saframycin synthetase; Cda1/2,calcium-dependent antibiotic; GrsA, gramicidin synthetase; PvdD,pyoverdin synthetase; McyC, microcystin synthetase; PstB, peptidesynthetase; TyrB, tyocidin synthetase; SnbD, pristinamycin/vir-giniamycin synthetase; Pps4, fengycin synthetase; RapP, rapamycinsynthetase; FkbP, FK506 synthetase; PvsA, pyoverdin synthetaseA; AoaB, cylindrospermopsin synthetase.
273

CYLN producers could not be established. Phyloge-netic trees reconstructed by parsimony and maximumlikelihood methods did not differ in their topologyand geometry from those produced by the neighbor-joining method.
Filamentous heterocyst-forming cyanobacteriacomprise a monophyletic group, but the generic andspecific assignments of its members are based onclassical morphological definitions, and often do notreflect their true phylogeny (Iteman et al. 2002).Apart from Cylindrospermopsis (Neilan et al. 2003),the phylogeny of CYLN producers has not beenpreviously examined by molecular methods. As evi-dent from the present study, the CYLN-producerstrains A. ovalisporum and Anabaena bergii may have
been taxonomically misclassified. Iteman et al. (2002)divided filamentous heterocyst-forming cyanobacte-ria into six subgroups. Two of these groups, groupIV, containing Anabaenopsis/Cyanospira, and groupV (Nodularia), provide a high tolerance to elevatedlevels of salinity, pH, and temperature (Iteman et al.2002). This trait is also shared by the Australianisolate Aphanizomenon ovalisporum APH028A, abloom of which was attributed to the high salinityand alkalinity of the water (Shaw et al. 1999). Nophysiological data were available for A. ovalisporumILC-146, U. natans, and A. bergii 283A. Based onshared physiological features and the high similarityin rDNA and ITS sequences, Iteman et al. (2002)suggested that Cyanospira and Anabaenopsis should
Fig. 6a. Phylogenetic affiliations ofselected filamentous, heterocyst formingcyanobacteria derived from partial 16SrDNA gene sequences. The phenogram wasreconstructed from a pairwise distancematrix (Jukes and Cantor 1969) using theneighbor-joining method (Saitou and Nei1987). The scale represents the number ofsubstitutions per 100 nucleotides. Bootstrapvalues (1000 resampling cycles) above 500are shown and represent the statisticalsignificance at each node.Cylindrospermopsin producing strains arein boldface. Accession numbers: AY897614,AF516724, AF516729 to AF516736,AF516738, AF516743 to AF516747,AF247577, AJ133154, AF092504,AF067819, AJ781131, AF160256,AY038033, AF044270, AY494701,AB039003, AY513505, AJ309733,AY038036, AF027655.
274

be assigned to a single genus. The close relationshipof U. natans, A. ovalisporum, and Anabaena bergii283A to Anabaenopsis/Cyanospira suggests that theymay have to be included in such a revised genus.
Phylogeny of CYLN Biosynthesis Genes
Twenty-one cyanobacterial strains, nine of whichproduced CYLN (Table 1), were screened by PCRfor the presence of putative CYLN biosynthesisgenes, using the primer pairs listed in Table 2. Posi-tive PCR products were sequenced using the sameprimers. As depicted in Figure 1, primer set CY-LATF/R amplified almost the entire aoaA gene (1105of 1179 bp), while CPSF/R amplified a 478-bp frag-
ment of the aoaB A-domain, which included the eightsubstrate specificity conferring residues. Finally, theprimer set A205PKF/R was used to amplify part (514bp) of the b-ketosynthase domain of aoaC.
All three putative CYLN biosynthesis genes,aoaA, aoaB, and aoaC, were exclusively present inCYLN producers, with one exception. The Hungar-ian strain, C. raciborskii Hung1, a nonproducer ofCYLN, possessed an aoaB homologue but no ho-mologues of aoaA or aoaC. Corresponding homo-logues of aoaA, aoaB, and aoaC in the differentstrains and species examined in the present studyshared at least 95% nucleotide sequence identity.Multiple sequence alignment divided the corre-sponding aoaA, aoaB, and aoaC homologues into
Fig. 6b. Phylogenetic tree ofamidinotransferase gene sequences fromCYLN producing cyanobacteria. Thephenogram was reconstructed as describedin the legend to Fig. 6a.Cylindrospermopsin producing strains arein boldface. Accesssion numbers:AY897605 to AY897613, AF395828, andAAM48710.
275

two distinct groups (Figs. 6b to 6d), one containingsequences exclusively from Cylindrospermopsis andthe other containing sequences from Aphanizomenon,Umezakia, and Anabaena, with one exception. TheaoaB homologue of C. raciborskii Hung1 groupedwith those of Aphanizomenon/Umezakia/Anabaena(Fig. 6b). No DNA of A. bergii 183A was available atthe time of the present study, however, a fragment ofa NRPS sequence (AF170843) outside the regioninvestigated in the present study showed greatersimilarity (99%) to that of Aphanizomenon than toCylindrospermopsis (96%).
The phylogenetic grouping of aoaA, aoaB, andaoaC homologues followed that of the 16S rRNAgenes of the producer organisms, however, the phy-logenetic distance of corresponding aoaA, aoaB, andaoaC homologues between the Cylindrospermopsisand the Aphanizomenon/Umezakia/Anabaena group,as well as within the latter group, was smaller thanthat of their 16S rRNA genes. The sequence similaritybetween aoaA, aoaB, and aoaC homologues withinCylindrospermopsis strains and within the Aphani-zomenon/Umezakia/Anabaena group ranged from 98to 100% at the nucleotide level, while the similarity
Fig. 6c. Phylogenetic tree ofnonribosomal peptide synthetase genesequences from CYLN producingcyanobacteria. The phenogram wasreconstructed as described in the legend toFig. 6a. Cylindrospermopsin producingstrains are in boldface. Accession numbers:AY89790 to AY89797, AAF15891,AAM33468, AAF76934, AF204805.
276

between the two groups was still 95 to 98%. The 16SrDNA sequence similarity within the Cylindrosperm-opsis group was 99%, but only 95 to 97% within theAphanizomenon/Umezakia/Anabaena group and 91 to92% between the two groups. Considering that 16SrDNA is one of the most conserved genes (Honda etal. 1999), the discrepancy in the divergence betweenrespective 16S rDNA genes and the putative CYLNbiosynthesis genes becomes even more apparent.There is mounting evidence that horizontal genetransfer is an important factor in shaping bacterialgenomes (e.g., Palenik et al. 2003; Sawada et al. 2002).The present data suggest that a gene transfer event
may have taken place between the Cylindrospermopsisand Aphanizomenon/Umezakia/Anabaena groups, aswell as among the Aphanizomenon/Umezakia/Anaba-ena group. The aoaB homologue in the Hungarianisolate C. raciborskii Hung1 could represent either aremnant or an otherwise ancestral intermediate of afunctional CYLN gene cluster and an evolutionarylink between theAphanizomenon/Umezakia/Anabaenaand Cylindrospermopsis groups.
There is indirect evidence that CYLN originated inAustralia and spread to other countries in the world.CYLN poisoning has been reported in Australia sincetimes of early European settlement, then referred to
Fig. 6d. Phylogenetic tree of polyketidesynthase genes from CYLN producingcyanobacteria. The phenogram wasreconstructed as described in the legend toFig. 6a. Cylindrospermopsin producingstrains are in boldface. Accession numbers:AY897598 to AY897604, AAF76933,AAG38956, AAM33470, CAD19091,AAG38958.
277

as ‘‘Belyando Fever’’ or ‘‘Barcoo Disease’’ (Hayman1992), and the largest number of reported CYLNproducing species occurs in Australia. Up until earlylast century, C. raciborskii was limited to tropicalregions, with a primary evolutionary center in thedeep lakes of tropical Africa and a secondary centerin Northern Australia, from where it is believed tohave recently spread to other parts of the world,including Europe (Padisak 1997). The invasiveness ofC. raciborskii has been described (Neilan et al. 2003),while A. ovalisporum may share similar physiologicalfeatures (Shaw et al. 1999). The close phylogeneticrelatedness between Israeli and Australian A. ovali-sporum isolates indicates that they may have origi-nated from a common source and are capable ofbeing dispersed over vast distances.
Conclusion
The AoaA model suggested that the putative functionof AoaA is the production of guanidino acetate fromglycine with arginine as the guanidino donor. Asopposed to Strb1, secondary structures involved insubstrate induced conformational changes in GATMwere present in AoaA, however, their amino acidsequence was poorly conserved, and the topology ofthese structures could not be modeled accurately. Noprediction was made about whether AoaA wouldundergo substrate induced conformational changesor whether it would be static as in the case of Strb1.
Previously established models were used to at-tempt the prediction of the substrate of the AoaB A-domain. Secondary structural alignment revealed thepresence of a universally conserved lysine (K558) andaspartate residue (D258) in AoaB, which in other A-domains forms hydrogen bonds to the a-carboxylateand a-amino group of the amino acid substrate,respectively. In addition, AoaB provided a secondaspartate (D362) residue, which in other A-domainsmay be one of the hydrophobic amino acids, leucine,isoleucine, or valine. Assuming that the catalyticresidues of AoaB provide the same topology as inGrsA guanidino acetate may be held in the catalyticsite by hydrogen bonds between its a-carboxylate andK558, between its imino-nitrogen of the guanidinogroup and D258, as well as between its positivelycharged guanidino amine and D362.
CYLN producing strains have been assigned toseveral genera of filamentous heterocyst-forming cy-anobacteria based on morphological characteristics.The molecular phylogeny of CYLN producing strainsdetermined in the present study clearly showed thatthe morphological classification is not precise. TheCYLN producing strains examined belonged to twophylotypes. One consisted of strains belonging to themonophyletic and monospecific C. raciborskii and theother, containing Aphanizomenon/Umezakia/Anabae-
na, should be combined into a revised genus togetherwith Anabaenopsis/Cyanospira.
There was no evident correlation between phy-logeny and the capacity for toxin production. Thephylogeny of CYLN biosynthesis genes followed, ingeneral, that of the producer strains, however,the divergence between Cylindrospermopsis andAphanizomenon/Umezakia/Anabaena phylotypes, aswell as between strains within the latter phylotype,was found to be greater than that between their aoahomologues. This may be the result of horizontalgene transfer of aoa homologues.
CYLN producing cyanobacteria have the capacityfor global distribution and represent a public healthconcern worldwide. The presence of an amidino-transferase in a cyanotoxin gene cluster is unique toCYLN producing cyanobacteria and thus representsan ideal target as a gene probe for water qualitymonitoring and, also, for the metabolic engineeringof complex biosyntheses.
Acknowledgments. The authors wish to acknowledge the finan-
cial support of the Australian Research Council and the assistance
given by Paul Curmi (UNSW) in the protein modeling. Strains used
in this study were supplied by Peter Baker (AWQC) and Martin
Saker (University of Porto).
References
Banker R, Carmeli S, Hadas O, Teltsch B, Porat R, Sukenik A
(1997) Identification of cylindrospermopsin in Aphanizomenon
ovalisporum (Cyanophyceae) isolated from lake Kinneret, Is-
rael. J Phycol 33:613–616
Bedekar A, Zink RM, Sherman DH, Line TV, Vanpilsum JF
(1998) The comparative amino acid sequences, substrate speci-
ficities and gene or cDNA nucleotide sequences of some pro-
karyote and eukaryote amidinotransferases—implications for
evolution. Comp Biochem Physiol B Biochem Mol Biol
119:677–690
Beitz E (2000) TeXshade: shading and labeling of multiple se-
quence alignments using LaTeX2e. Bioinformatics 16:135–139
Bourke ATC, Hawes RB, Neilson A, Stallman ND (1983) An
outbreak of hepato-enteritis (the Palm Island mystery disease)
possibly caused by algal intoxication. Toxicon 21:45–48
Burgoyne DL, Hemscheidt TK, Moore RE, Runnegar MTC (2000)
Biosynthesis of cylindrospermopsin. J Org Chem 65:152–156
Challis GL, Ravel J, Townsend CA (2000) Predictive, structure-
based model of amino acid recognition by nonribosomal pep-
tide synthetase adenylation domains. Chem Biol 7:211–224
Conti E, Stachelhaus T, Marahiel MA, Brick P (1997) Structural
basis for the activation of phenylalanine in the nonribosomal
biosynthesis of gramicidin S. EMBO J 16:4174–83
Felsenstein J (1989) PHYLIP. Phylogeny inference package. Cla-
distics 5:164–166
Fritsche E, Bergner A, Humm A, Piepersberg W, Huber R (1998)
Crystal structure of L-arginine:inosamine-phosphate amidino-
transferase StrB1 from Streptomyces griseus: An enzyme
involved in streptomycin biosynthesis. Biochemistry 37:17664–
17672
Fritsche E, Humm A, Huber R (1999) The ligand-induced struc-
tural changes of human L-arginine:glycine amidinotransferase.
278

A mutational and crystallographic study. J Biol Chem
274:3026–3032
Harada K, Ohtani I, Iwamoto K, Suzuki M, Watanabe MF, Wa-
tanabe M, Terao K (1994) Isolation of cylindrospermopsin
from a cyanobacterium Umezakia natans and its screening
method. Toxicon 32:73–84
Hawkins PR, Runnegar MT, Jackson AR, Falconer IR (1985)
Severe hepatotoxicity caused by the tropical cyanobacterium
(blue–green alga) Cylindrospermopsis raciborskii (Woloszynska)
Seenaya and Subba Raju isolated from a domestic water supply
reservoir. Appl Environ Microbiol 50:1292–5
Hawkins PR, Chandrasena NR, Jones GJ, Humpage AR, Falconer
IR (1997) Isolation and toxicity of Cylindrospermopsis racibor-
skii from an ornamental lake. Toxicon 35:341–346
Hayman J (1992) Beyond the Barcoo––probable human tropical
cyanobacterial poisoning in outback Australia. Med J Aust
157:794–796
Honda D, Yokota A, Sugiyama J (1999) Detection of seven major
evolutionary lineages in cyanobacteria based on the 16S rRNA
gene sequence analysis with new sequences of five marine
Synechococcus strains. J Mol Evol 48:723–739
Humpage AR, Fenech M, Thomas P, Falconer IR (2000) Micro-
nucleus induction and chromosome loss in transformed human
white cells indicate clastogenic and aneugenic action of the cy-
anobacterial toxin, cylindrospermopsin. Mutat Res Genet
Toxicol Environ Mutat 472:155–161
Iteman I, Rippka R, de Tandeau Marsac N, Herdman M (2002)
rDNA analyses of planktonic heterocystous cyanobacteria,
including members of the genera Anabaenopsis and Cyanospira.
Microbiology 148:481–496
Jones TA, Zou JY, Cowan SW, Kjeldgaard (1991) Improved
methods for building protein models in electron density maps
and the location of errors in these models. Acta Crystallograf A
47(2):110–119
Jukes TH, Cantor CR (1969) Evolution of protein molecules. In:
Munro HN (ed) Mammalian protein metabolism. Academic
Press, New York, pp 121–132
Kiss T, Vehovszky A, Hiripi L, Kovacs A, Voros L (2002) Mem-
brane effects of toxins isolated from a cyanobacterium, Cylin-
drospermopsis raciborskii, on identified molluscan neurones.
Comp Biochem Physiol C Toxicol Pharmacol 131:167–176
Kleywegt GJ (1996) Use of non-crystallographic symmetry in
protein structure refinement. Acta Crystallograf D 52:842–857
Lagos N, Onodera H, Zagatto PA, Andrinolo D, Azevedo S,
Oshima Y (1999) The first evidence of paralytic shellfish toxins
in the freshwater cyanobacterium Cylindrospermopsis racibor-
skii, isolated from Brazil. Toxicon 37:1359–1373
Lee GT, Kim WJ, Cho YD (2002) Polyamine synthesis in plants.
Purification and properties of amidinotransferase from soybean
(Glycine max) axes. Phytochemistry 61:781–789
Li RH, Carmichael WW, Brittain S, Eaglesham GK, Shaw GR,
Liu YD, Watanabe MM (2001) First report of the cyanotoxins
cylindrospermopsin and deoxycylindrospermopsin from
Raphidiopsis curvata (Cyanobacteria). J Phycol 37:1121–1126
Luthy R, Bowie JU, Eisenberg D (1992) Assessment of protein
models with three-dimensional profiles. Nature 356:83–85
Montesinos ML, Herrero A, Flores E (1997) Amino acid transport
in taxonomically diverse cyanobacteria and identification of
two genes encoding elements of a neutral amino acid permease
putatively involved in recapture of leaked hydrophobic amino
acids. J Bacteriol 179:853–862
Neilan BA (1995) Identification and phylogenetic analysis of toxi-
genic cyanobacteria by multiplex randomly amplified polymor-
phic DNA PCR. Appl Environ Microbiol 61:2286–2291
Neilan BA, Jacobs D, DelDot T, Blackall LL, Hawkins PR, Cox
PT, Goodman AE (1997) rRNA sequences and evolutionary
relationships among toxic and nontoxic cyanobacteria of the
genus Microcystis. Int J Syst Bacteriol 47:693–697
Neilan BA, Saker ML, Fastner J, Torokne A, Burns BP
(2003) Phylogeography of the invasive cyanobacterium
Cylindrospermopsis raciborskii. Mol Ecol 12:133–140
Ohtani I, Moore RE, Runnegar MTC (1992) Cylindrospermop-
sin—a potent hepatotoxin from the blue-green-alga
Cylindrospermopsis raciborskii. J Am Chem Soc 114:7941–7942
Padisak J (1997) Cylindrospermopsis raciborskii (Wolonszynska)
Seenayva et Subba Raju, an expanding, highly adaptive cya-
nobacterium: worldwide distribution and review of its ecology.
Arch Hydrobiol Suppl 107:563–593
Palenik B, Brahamsha B, Larimer FW, Land M, Hauser L, Chain
P, Lamerdin J, Regala W, Allen EE, McCarren J, Paulsen I,
Dufresne A, Partensky F, Webb EA, Waterbury J (2003) The
genome of a motile marine Synechococcus. Nature 424:1037–
1042
Perriere G, Gouy M (1996) WWW-Query: An on-line retrieval
system for biological sequence banks. Biochemie 78:364–369
Runnegar MT, Kong SM, Zhong YZ, Ge JL, Lu SC (1994) The
role of glutathione in the toxicity of a novel cyanobacterial
alkaloid cylindrospermopsin in cultured rat hepatocytes. Bio-
chem Biophys Res Commun 201:235–241
Saitou N, Nei M (1987) The neighbour-joining method: a new
method for reconstructing phylogenetic trees. Mol Biol Evol
4:406–425
Saker ML, Griffiths DJ (2000) The effect of temperature on
growth and cylindrospermopsin content of seven isolates of
Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae)
from water bodies in northern Australia. Phycologia 39:349–
354
Saker ML, Neilan BA, Griffiths DJ (1999a) Two morphological
forms of Cylindrospermopsis raciborskii (Cyanobacteria) iso-
lated from Solomon Dam, Palm Island, Queensland. J Phycol
35:599–606
Saker ML, Thomas AD, Norton JH (1999b) Cattle mortality
attributed to the toxic cyanobacterium Cylindrospermopsis
raciborskii in an outback region of North Queensland. Environ
Toxicol 14:179–182
Sambrook J, Fritsch E, Maniatis T (1989) Molecular cloning: A
laboratory manual. Cold Spring Harbor Laboratory, Cold
Spring Harbor, NY
Sawada H, Kanaya S, Tsuda M, Suzuki F, Azegami K, Saitou N
(2002) A phylogenomic study of the OCTase genes in Pseudo-
monas syringae pathovars: the horizontal transfer of the argK-
tox cluster and the evolutionary history of OCTase genes on
their genomes. J Mol Evol 54:437–457
Schembri MA, Neilan BA, Saint CP (2001) Identification of genes
implicated in toxin production in the cyanobacterium
Cylindrospermopsis raciborskii. Environ Toxicol 16:413–421
Shalev-Alon G, Sukenik A, Livnah O, Schwarz R, Kaplan A (2002)
A novel gene encoding amidinotransferase in the cylindrosper-
mopsin producing cyanobacterium Aphanizomenon
ovalisporum. FEMS Microbiol Lett 209:83–87
Shaw GR, Sukenik A, Livne A, Chiswell RK, Smith MJ, Seawright
AA, Norris RL, Eaglesham GK, Moore MR (1999) Blooms of
the cylindrospermopsin containing cyanobacterium, Aphani-
zomenon ovalisporum (Forti), in newly constructed lakes,
Queensland, Australia. Environ Toxicol 14:167–177
Shi J, Blundell TL, Mizuguchi K (2001) FUGUE: sequence-
structure homology recognition using environment-specific
substitution tables and structure–dependent gap penalties. J
Mol Biol 310:243–257
Srivenugopal KS, Adiga PR (1980) Partial purification and prop-
erties of a transamidinase from Lathyrus sativus seedlings.
Involvement in homoarginine metabolism and amine intercon-
versions. Biochem J 189:553–560
Stachelhaus T, Mootz HD, Marahiel MA (1999) The specificity-
conferring code of adenylation domains in nonribosomal pep-
tide synthetases. Chem Biol 6:493–505
279

Thompson AS, Rhodes JC, Pettman I (1988) Catalogue of strains.
Natural Environment Research Council Culture Collection of
Algae and Protozoa, p 22
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F,
Higgins DG (1997) The CLUSTAL_X windows inter-
face: flexible strategies for multiple sequence alignment
aided by quality analysis tools. Nucleic Acids Res 25:
4876–4882
Wilson KM, Schembri MA, Baker PD, Saint CP (2000) Molecular
characterization of the toxic cyanobacterium Cylindrospermop-
sis raciborskii and design of a species-specific PCR. Appl
Environ Microbiol 66:332–338
280