five new species of listeria (l. floridensis sp. nov, l. aquatica sp. nov., l. cornellensis sp. nov....
TRANSCRIPT

Listeria floridensis sp. nov., Listeria aquatica sp.nov., Listeria cornellensis sp. nov., Listeria ripariasp. nov. and Listeria grandensis sp. nov.,from agricultural and natural environments
Henk C. den Bakker,1 Steven Warchocki,1 Emily M. Wright,1
Adam F. Allred,2 Christina Ahlstrom,33 Clyde S. Manuel,34Matthew J. Stasiewicz,1 Angela Burrell,2 Sherry Roof,1 Laura K. Strawn,1
Esther Fortes,11 Kendra K. Nightingale,4 Daniel Kephart2
and Martin Wiedmann1
Correspondence
Henk C. den Bakker
1Department of Food Science, Cornell University, Ithaca, NY 14853, USA
2Life Technologies, 2130 Woodward Street, Austin, TX 78744, USA
3Department of Animal Sciences, Colorado State University, Fort Collins, CO 80523, USA
4Department of Animal and Food Sciences, Texas Tech University, Lubbock, TX 79409, USA
Sampling of agricultural and natural environments in two US states (Colorado and Florida) yielded
18 Listeria-like isolates that could not be assigned to previously described species using
traditional methods. Using whole-genome sequencing and traditional phenotypic methods, we
identified five novel species, each with a genome-wide average BLAST nucleotide identity (ANIb) of
less than 85 % to currently described species. Phylogenetic analysis based on 16S rRNA gene
sequences and amino acid sequences of 31 conserved loci showed the existence of four
well-supported clades within the genus Listeria; (i) a clade representing Listeria monocytogenes,
L. marthii, L. innocua, L. welshimeri, L. seeligeri and L. ivanovii, which we refer to as Listeria
sensu stricto, (ii) a clade consisting of Listeria fleischmannii and two newly described species,
Listeria aquatica sp. nov. (type strain FSL S10-1188T5DSM 26686T5LMG 28120T5BEI
NR-42633T) and Listeria floridensis sp. nov. (type strain FSL S10-1187T5DSM 26687T5LMG
28121T5BEI NR-42632T), (iii) a clade consisting of Listeria rocourtiae, L. weihenstephanensis
and three novel species, Listeria cornellensis sp. nov. (type strain TTU A1-0210T5FSL
F6-0969T5DSM 26689T5LMG 28123T5BEI NR-42630T), Listeria grandensis sp. nov.
(type strain TTU A1-0212T5FSL F6-0971T5DSM 26688T5LMG 28122T5BEI NR-42631T)
and Listeria riparia sp. nov. (type strain FSL S10-1204T5DSM 26685T5LMG 28119T5BEI
NR- 42634T) and (iv) a clade containing Listeria grayi. Genomic and phenotypic data suggest
that the novel species are non-pathogenic.
3Present address: Production Animal Health, University of Calgary, Alberta, Canada.
4Present address: Department of Food, Bioprocessing and Nutrition Sciences, North Carolina State University, Raleigh, NC 27695-7624, USA.
1Present address: Hinton State Laboratory Institute, Massachusetts Department of Public Health, Jamaica Plain, MA, USA.
Abbreviations: AAI, average amino acid identity; ANIb, BLAST average nucleotide identity; MR, methyl red; PI-PLC, phosphoinositide phospholipase C; VP,Voges–Proskauer.
The GenBank/EMBL/DDBJ accession numbers for the 16S rRNA gene sequences of strains TTU A1-0210T, TTU A1-0212T, FSL S10-1187T, FSLS10-1188T and FSL S10-1204T are JX961634, JX961635, JX961636, JX961637 and JX961638, respectively. The accession numbers for thegenome sequences of strains TTU A1-0210T, TTU A1-0212T, FSL S10-1187T, FSL S10-1188T and FSL S10-1204T are AODE00000000,AODD00000000, AODF00000000, AOCG00000000 and AODL00000000, respectively; the versions described in this paper are the first versions,AODE01000000, AODD01000000, AODF01000000, AOCG01000000 and AODL01000000.
Five supplementary figures and five supplementary tables are available with the online version of this paper.
International Journal of Systematic and Evolutionary Microbiology (2014), 64, 1882–1889 DOI 10.1099/ijs.0.052720-0
1882 052720 G 2014 IUMS Printed in Great Britain

The genus Listeria was described by Pirie (1940), and atpresent comprises 10 recognized species, Listeria monocy-togenes (Pirie, 1940), Listeria grayi (Errebo Larsen &Seeliger, 1966), Listeria innocua (Seeliger, 1981), Listeriawelshimeri (Rocourt & Grimont, 1983), Listeria seeligeri(Rocourt & Grimont, 1983), Listeria ivanovii (Seeliger et al.,1984), Listeria marthii (Graves et al., 2010), Listeria rocourtiae(Leclercq et al., 2010), Listeria fleischmannii (Bertsch et al.,2013) and Listeria weihenstephanensis (Lang Halter et al.,2013). Additionally, two subspecies have been recognizedwithin L. ivanovii (subsp. ivanovii and subsp. londoniensis;Boerlin et al., 1992), L. grayi [subsp. grayi and subsp. murrayi(Stuart & Welshimer, 1973), though these names are notvalidly published] and L. fleischmannii (subsp. fleischmanniiand subsp. coloradonensis; den Bakker et al., 2013).
During sampling projects of agricultural and naturalenvironments in the US states of Florida and Colorado,we isolated 70 isolates, predominantly from water, with acolony morphology on Listeria monocytogenes platingmedium (LMPM) reminiscent of strains of Listeria. Theseisolates could be placed in the family Listeriaceae by phy-logenetic analysis of partial sequences of the 16S rRNAgene, and 52 isolates could be assigned to L. fleischmannii(32 isolates from Florida and 20 isolates from Colorado);however, 18 isolates did not cluster phylogenetically withinpreviously described species and therefore could not beclassified as members of any of the previously describedspecies. Based on further genotypic characterization of theseisolates, including by sigB sequencing (Sauders et al., 2012),one or two isolates per putative novel taxon were selectedfor further phenotypic and genomic characterization (seeFig. S1, available in the online Supplementary Material, formore information).
Draft genome sequences were assembled as detailed by denBakker et al. (2013) from Ion PGM instrument readsobtained for (i) Brochothrix thermosphacta ATCC 11509T
and B. campestris ATCC 43754T, (ii) the type strains L.monocytogenes ATCC 15313T, L. weihenstephanensis DSM24698T and L. rocourtiae CIP 109804T and L. grayi ATCC25401 (the type strain of Listeria murrayi), (iii) an additionalstrain of L. fleischmannii (FSL S10-1203) and (iv) five isolatesthat showed Listeria-like characteristics, but could not beclassified to species by traditional approaches. Briefly, denovo assembly was performed using the MIRA assembler(Chevreux, 1999), and homopolymers were corrected usinga homopolymer correction script (available at http://tinyurl.com/oke6xaa); this script uses output from VarScan(Koboldt et al., 2012) and BWA (Li & Durbin, 2009) toobtain a consensus about the homopolymer length, which isthen used to correct the homopolymers. De novo assemblies(draft genomes) of Ion Torrent reads ranged in total sizefrom 2.37 (B. campestris) to 3.37 (L. weihenstephanensis) Mbp(see Table S1 for total draft genome lengths and additionalgenome assembly statistics). Comparison of the draft genomeobtained here for L. monocytogenes ATCC 15313T and arecently finished genome of L. monocytogenes SLCC5850(Kuenne et al., 2013), a strain closely related to the type strain
(McLauchlin & Rees, 2009), showed that the draft genomecovers 97.3 % of the finished genome. We therefore concludethat the draft genomes obtained in this study are ofhigh quality and suitable for comparative analysis. Thesedraft genomes will also provide a starting point for futurein-depth genomic and evolutionary analyses of the genusListeria.
BLAST average nucleotide identities (ANIb) were calculatedbased on draft genome sequences and publicly availablegenome sequences using JSpecies (Richter & Rossello-Mora,2009). Additionally, BLAST average amino acid identities(AAI) (Konstantinidis & Tiedje, 2005) were calculated usingthe AAI.rb script from the Enve-omics package (http://enve-omics.ce.gatech.edu/). Pairwise comparisons between cur-rently recognized species in Listeria sensu stricto yieldedANIb and AAI values ,95 % for comparisons betweendifferent species (Fig. 1; Table S2); an ANIb value of 95–96 %generally corresponds to 70 % DNA–DNA hybridizationand is typically used as a cut-off value for species delineationin bacterial taxonomy (Richter & Rossello-Mora, 2009). Ourdata also support conclusions from previous studies thatANIb values are robust even with fragmented unfinisheddraft assemblies. This robustness is demonstrated by anANIb value of 99.94 % for the comparison between the draftgenome for the L. monocytogenes type strain, ATCC 15313T
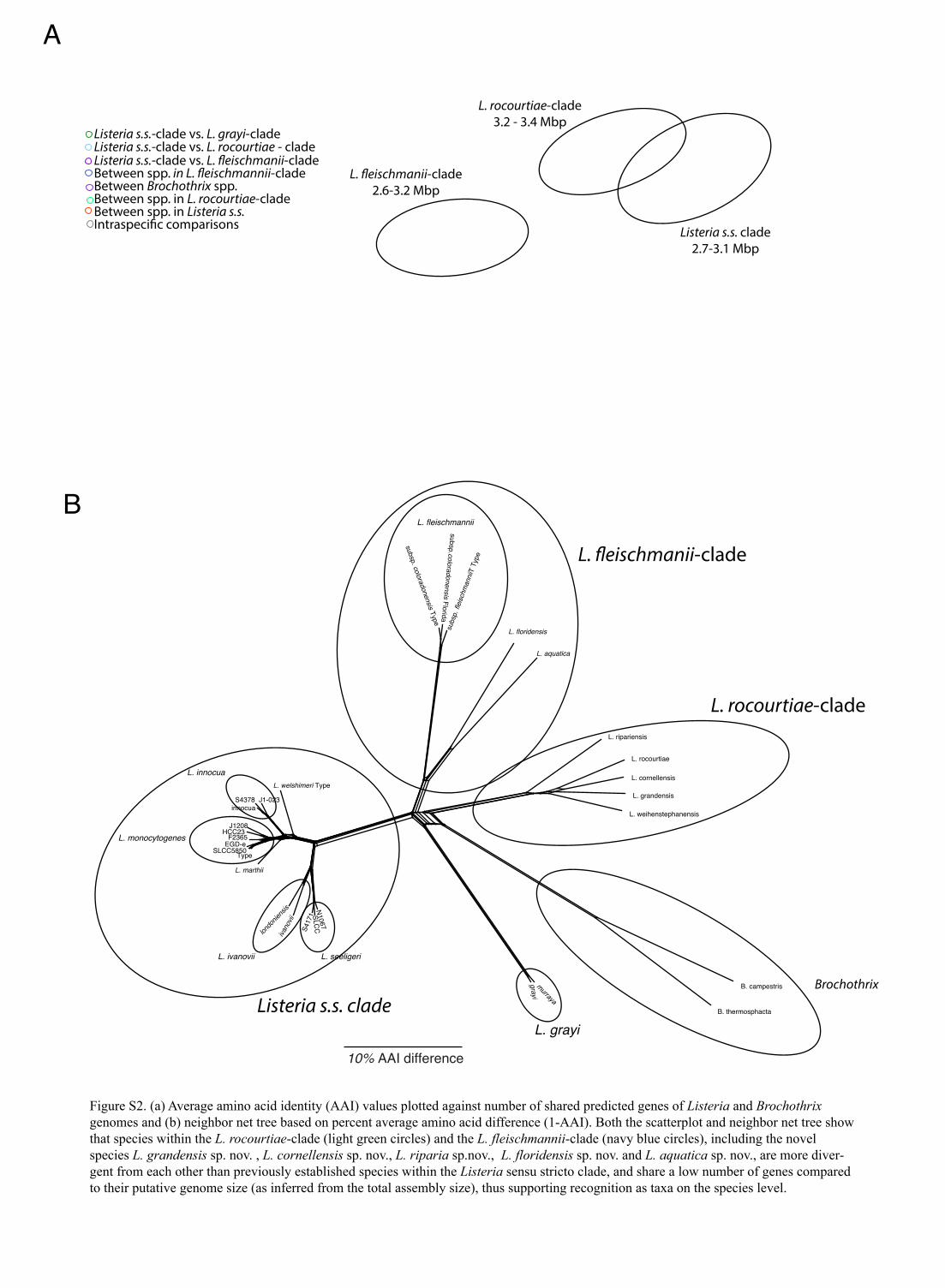
(a draft genome obtained in this study), and the finishedgenome for L. monocytogenes SLCC5850. For all genomesrepresenting previously reported species, ANIb valuessupported prior species classification, despite the mixeduse of finished and draft genomes (Fig. 1). Most impor-tantly, our analyses showed that all isolates representingnovel species proposed in this study show ANIb values,84 % and AAI values ,87.9 % (Table S2) when theirgenomes are compared to each other and to phylogeneticallyrelated, previously described species (L. fleischmannii,L. rocourtiae and L. weihenstephanensis). While the high16S rRNA gene sequence similarity (.99 %) between L.rocourtiae CIP 109804T, L. weihenstephanensis DSM 24998T,L. cornellensis sp. nov. TTU A1-0210T, L. grandensis sp. nov.TTU A1-0212T and L. riparia sp. nov. FSL S10-1204T couldbe interpreted as suggesting that these taxa representecotypes of a single species (Cohan, 2001; Konstantinidiset al., 2006), ANIb and AAI values and shared gene contentdo not support this hypothesis. AAI and ANIb valuesbetween these species range from 83.2 to 88.6 % andfrom 77.7 to 83.2 %, respectively, and the proportionof shared gene content between each species pair rangesfrom 63 to 75 %, justifying recognition as individual species(Fig. S2).
Phylogenetic analysis of (i) 16S rRNA gene sequences(Fig. S2) and (ii) concatenated sequences of 31 amino acidsequences (Wu & Eisen, 2008) (Fig. 2) was performed asdescribed previously (den Bakker et al., 2013), with theexception that Gblocks version 0.91b (Castresana, 2000)was used to remove ambiguous sites from the aminoacid sequence alignment. Maximum-parsimony analyseswere performed in PAUP* version 4.0b10 (Wilgenbusch &
Novel species of Listeria from environmental samples
http://ijs.sgmjournals.org 1883

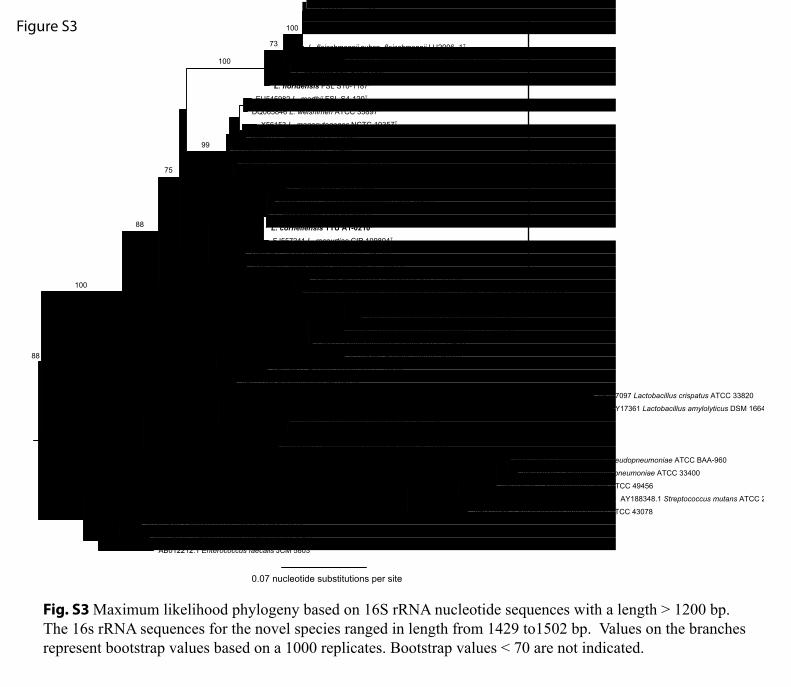
Swofford, 2003). Phylogenies obtained from the max-imum-parsimony analysis (data not shown) did not differfrom those found by maximum-likelihood-based analyses,and hence the results of the maximum-likelihood analysesare discussed here. Both 16S rRNA gene and amino acidsequence phylogenies confirmed the placement of the noveltaxa in the genus Listeria, and revealed four well-supportedclades [100 % bootstrap support (BS)] within the genusListeria: (i) a clade containing L. monocytogenes and relatedspecies (Listeria sensu stricto: L. marthii, L. innocua, L.welshimeri, L. seeligeri and L. ivanovii), (ii) a clade consistingof L. rocourtiae, L. weihenstephanensis, L. cornellensis sp.nov., L. grandensis sp. nov. and L. riparia sp. nov., (iii) aclade consisting of L. fleischmannii, L. floridensis sp. nov.and L. aquatica sp. nov., and (iv) a clade consisting ofL. grayi. The 16S rRNA gene sequence analysis (Fig. S3)also indicates that a previously unidentified Listeria-likebacterium (referred to as ‘unidentified bacterium isolateMB405’ in Fig. S3) from Belgium (Rijpens et al., 1998)represents a putative sister species of L. aquatica sp. nov.Phylogenetic analysis of individual genes, such as iap (Fig.S4), also reveal the subdivision of Listeria into four distinctclades. The phylogenetic relationships between the fourclades are unresolved in the 31 amino acid loci tree (Fig. 1;BS values ,60 %); however, the 16S rRNA gene sequenceanalysis suggests that the L. rocourtiae clade and the L. grayiclade are sister groups (Fig. S3; 85 % BS). AAI valuesbetween members of each phylogenetically distinct claderange from 67 to 70 % (Fig. S2). While Konstantinidis &Tiedje (2005, 2007) show that AAI between 65 and 72 %coincides with traditional genus delimitations in bac-terial taxonomy, we do not propose classification of theseclades into separate genera at this point, in part because
reclassification of L. grayi into a new genus has previouslybeen unsuccessful and controversial (McLauchlin & Rees,2009).
For methyl red (MR) and Voges–Proskauer (VP) tests,all isolates were cultured aerobically on BHI agar (BectonDickinson) for 18 h at 30 uC. A single colony was suspendedin 5 ml MR-VP broth (pH 6.9±0.2; Becton Dickinson);inoculated tubes were incubated aerobically for 5 days at35 uC. MR and VP tests were carried out according to themanufacturer’s guidelines (Becton Dickinson; procedureL007474). Results are listed in the species descriptions andTable 1.
Testing for catalase activity was performed according toBacteriological Analytical Manual (BAM) protocol R12(US Food and Drug Administration, 2013). An isolatedcolony from BHI agar was emulsified in a drop of 3 %hydrogen peroxide on a glass slide. Bubbling within 15 swas interpreted as a positive result for the catalase test.Results were scored independently by two individuals andare listed in the species descriptions and Table 1.
To test for nitrate reduction, isolates were grown innitrate broth (BAM medium M108; US Food and DrugAdministration, 2013) with Durham tubes for 48 h at30 uC without shaking. Durham tubes allowed detectionof gas production at 24 and 48 h, which is indicative ofN2 production. Reduction of nitrate to nitrite was tested byadding 50 ml bacterial culture grown in nitrate broth to0.9 ml phosphate buffer (50 mM, pH 7.4), followed by theaddition of 1 ml sulfanilic acid/HCl (5.0 g sulfanilic acid,100 ml concentrated HCl, 400 ml water; final pH 20.38)and 1 ml 0.58 % N-(1-naphthyl) ethylenediamine. An
L. aquatica FSL S10-1188T
L. floridensis FSL S10-1187T
L. cornellensis TTU A1-0210T
L. grandensis TTU A1-0212T
L. riparia FDL S10-1204T
L. fleischmannii subsp. coloradensis FSL S10 -1203L. fleischmannii subsp. coloradensis TTU M1-001T
L. fleischmannii subsp. fleischmannii DSM 24998T
L. monocytogenes ATCC 15313T
L. monocytogenes EGD-eL. monocytogenes F2365L. monocytogenes HCC23L. monocytogenes FSL J1-208L. marthii FSL S4-120T
L. innocua Clip 11262L. innocua FSL S4-378L. innocua FSL J1-023L. welshimeri SLCC5334T
L. ivanovii subsp. londoniensis ATCC 49954T
L. ivanovii subsp. ivanovii PAM 55L. seeligeri FSL N1-067L. seeligeri FSL S4-171L. seeligeri SLCC3954T
L. rocourtiae CIP 109804T
L. weihenstephanensis DSM 24699T
L. grayi DSM 20601T
‘L. grayi subsp. murrayi ’ ATCC 25401B. campestris ATCC 43754T
B. thermosphacta ATCC 11509T Brochothrix
Listeria
65 8575 95
Fig. 1. UPGMA cluster diagram based onpercentage average nucleotide difference(5100 % - ANIb; see Chan et al., 2012). Thescale indicates ANIb (%). The grey verticalbar marks 5 % average nucleotide difference,or 95 % ANIb, a value that has been shownto correlate with the traditional DNA–DNAhybridization value of 70 %. Novel species areshown in bold.
H. C. den Bakker and others
1884 International Journal of Systematic and Evolutionary Microbiology 64

immediate colour change to deep pink to purple indicatedthe production of nitrite. For all isolates, nitrite reductionwas determined using a protocol comparable to the nitratereduction protocol described above, except that KNO2 wasused instead of KNO3 in BAM medium M108. Nitritereduction was negative if the reaction solution producedmagenta to purple coloration. If an isolate reduces nitrateto an oxidation state lower than nitrite, the nitrate reductiontest will produce a false negative. Therefore, separatelytesting nitrite reduction and the generation of N2 (Durhamtubes) measures the reduction of nitrate to nitrite. All non-motile species were found to be able to reduce nitrate, withthe exception of L. floridensis sp. nov. None of the novelspecies reported here was able to reduce nitrite.
Haemolysis for each isolate was determined using theCAMP test as described in BAM (US Food and DrugAdministration, 2013) and performed in duplicate. To testfor phosphatidylinositol-specific phospholipase C (PI-PLC) activity, single colonies from BHI agar plates weresubcultured to LMPM (R&F Laboratories). PI-PLC activ-ity-positive strains colour blue on LMPM medium. None ofthe novel species described here showed haemolytic activityor PI-PLC activity, suggesting that these species lack thegenes (i.e. hly and plcA) associated with virulence in L.
monocytogenes and L. ivanovii. Virulence genes (i.e. genesfound in the prfA cluster; Schmid et al., 2005) or LIPI-2(Domınguez-Bernal et al., 2006) were absent from the draftgenomes, further supporting the conclusion that none ofthe novel species described here are pathogenic.
API Listeria and API 50 CH (with API 50 CHB/E medium)kits (bioMerieux) were utilized for further phenotypiccharacterization, using bacterial colonies taken from BHIagar plates. API Listeria tests were incubated for 24 h at34 uC for all isolates (within the manufacturer’s recom-mended temperature range of 36±2 uC). API 50 CH testswere incubated for 48 h at 37 uC for all isolates. A mineraloil cover was not used, since all species studied hereare facultative anaerobes. Results were recorded at 48 h,unless there were positive or variable results at 24 h with acorresponding negative result at 48 h, in which case 24 hresults were reported (as instructed in the API 50 CHB/Emanual). Results are summarized in the species descrip-tions and Table 1. Results for individual isolates arereported in Table S3.
To test growth characteristics, isolates were grown in BHIbroth at 4, 7, 22.5, 30, 37 and 41 uC. A Synergy H1 hybridmulti-mode microplate reader (BioTek Instruments) wasemployed to test for growth at 22.5±0.5, 30±0.1, 37±0.1
L. monocytogenes EGD-e
L. monocytogenes F2365
L. marthii FSL S4-120T
L. innocua Clip11262
L. welshimeri serovar 6b SLCC5334T
L. seeligeri serovar 1/2b SLCC3954T
L. ivanovii subsp. ivanovii PAM55L. weihenstephanensis DSM 24699T
L. fleischmannii FSL S10 -1203
L. rocourtiae CIP 109804T
L. fleischmannii subsp. coloradensis TTU M1-001T
L. fleischmannii subsp. fleischmannii DSM 24998T
‘L. grayi subsp. murrayi ’ ATCC 25401
L. grayi DSM 20601T
B. campestris ATCC 43754TBrochothrix
Listeria
B. thermosphacta ATCC 11509T
Staphylococcus epidermidis ATCC 12228
Staphylococcus haemolyticus JCSC1435
Staphylococcus aureus USA300 TCH1516
Staphylococcus saprophyticus ATCC 15305T
Enterococcus faecalis V583
0.05
100
100
100
100
100
100100
100
100
100
100
84
85
100
96
96
L. cornellensis TTU A1-0210T
L. grandensis TTU A1-0212T
L. riparia FSL S10-1204T
L. aquatica FSL S10-1188T
L. floridensis FSL S10-1187T
Fig. 2. Maximum-likelihood phylogeny based on concatenated amino acid sequences of 31 core genes. Values on branchesrepresent bootstrap values based on 1000 replicates. Bootstrap values ,70 % are not indicated. Novel species are shownin bold.
Novel species of Listeria from environmental samples
http://ijs.sgmjournals.org 1885
and 41±0.1 uC (temperature tolerances as provided bymanufacturer). Colony-inoculated cultures were grown for24 h in BHI broth; an aliquot of these cultures (20 ml) was
used to inoculate Costar 96-well flat-bottom plates (clearpolystyrene) prefilled with 180 ml BHI. Inoculated plateswere incubated at 22.5 uC for 48 h, 30 uC for 41 h, 37 uC
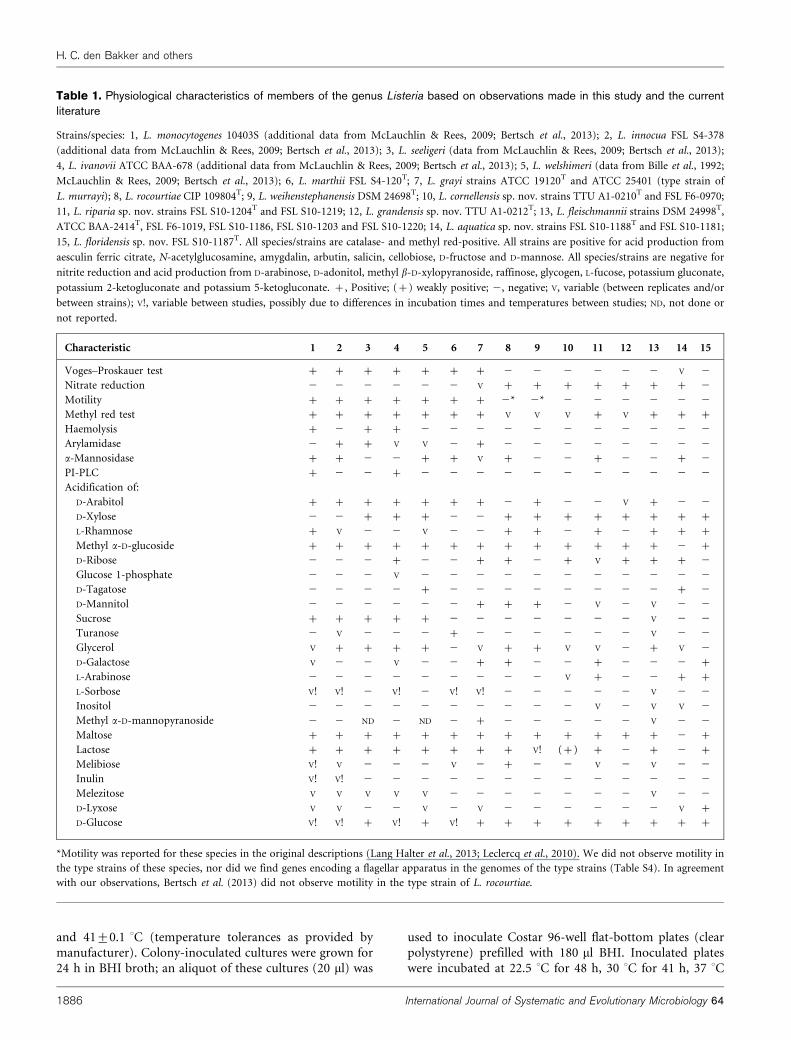
Table 1. Physiological characteristics of members of the genus Listeria based on observations made in this study and the currentliterature
Strains/species: 1, L. monocytogenes 10403S (additional data from McLauchlin & Rees, 2009; Bertsch et al., 2013); 2, L. innocua FSL S4-378
(additional data from McLauchlin & Rees, 2009; Bertsch et al., 2013); 3, L. seeligeri (data from McLauchlin & Rees, 2009; Bertsch et al., 2013);
4, L. ivanovii ATCC BAA-678 (additional data from McLauchlin & Rees, 2009; Bertsch et al., 2013); 5, L. welshimeri (data from Bille et al., 1992;
McLauchlin & Rees, 2009; Bertsch et al., 2013); 6, L. marthii FSL S4-120T; 7, L. grayi strains ATCC 19120T and ATCC 25401 (type strain of
L. murrayi); 8, L. rocourtiae CIP 109804T; 9, L. weihenstephanensis DSM 24698T; 10, L. cornellensis sp. nov. strains TTU A1-0210T and FSL F6-0970;
11, L. riparia sp. nov. strains FSL S10-1204T and FSL S10-1219; 12, L. grandensis sp. nov. TTU A1-0212T; 13, L. fleischmannii strains DSM 24998T,
ATCC BAA-2414T, FSL F6-1019, FSL S10-1186, FSL S10-1203 and FSL S10-1220; 14, L. aquatica sp. nov. strains FSL S10-1188T and FSL S10-1181;
15, L. floridensis sp. nov. FSL S10-1187T. All species/strains are catalase- and methyl red-positive. All strains are positive for acid production from
aesculin ferric citrate, N-acetylglucosamine, amygdalin, arbutin, salicin, cellobiose, D-fructose and D-mannose. All species/strains are negative for
nitrite reduction and acid production from D-arabinose, D-adonitol, methyl b-D-xylopyranoside, raffinose, glycogen, L-fucose, potassium gluconate,
potassium 2-ketogluconate and potassium 5-ketogluconate. +, Positive; (+) weakly positive; 2, negative; V, variable (between replicates and/or
between strains); V!, variable between studies, possibly due to differences in incubation times and temperatures between studies; ND, not done or
not reported.
Characteristic 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Voges–Proskauer test + + + + + + + 2 2 2 2 2 2 V 2
Nitrate reduction 2 2 2 2 2 2 V + + + + + + + 2
Motility + + + + + + + 2* 2* 2 2 2 2 2 2
Methyl red test + + + + + + + V V V + V + + +
Haemolysis + 2 + + 2 2 2 2 2 2 2 2 2 2 2
Arylamidase 2 + + V V 2 + 2 2 2 2 2 2 2 2
a-Mannosidase + + 2 2 + + V + 2 2 + 2 2 + 2
PI-PLC + 2 2 + 2 2 2 2 2 2 2 2 2 2 2
Acidification of:
D-Arabitol + + + + + + + 2 + 2 2 V + 2 2
D-Xylose 2 2 + + + 2 2 + + + + + + + +
L-Rhamnose + V 2 2 V 2 2 + + 2 + 2 + + +
Methyl a-D-glucoside + + + + + + + + + + + + + 2 +
D-Ribose 2 2 2 + 2 2 + + 2 + V + + + 2
Glucose 1-phosphate 2 2 2 V 2 2 2 2 2 2 2 2 2 2 2
D-Tagatose 2 2 2 2 + 2 2 2 2 2 2 2 2 + 2
D-Mannitol 2 2 2 2 2 2 + + + 2 V 2 V 2 2
Sucrose + + + + + 2 2 2 2 2 2 2 V 2 2
Turanose 2 V 2 2 2 + 2 2 2 2 2 2 V 2 2
Glycerol V + + + + 2 V + + V V 2 + V 2
D-Galactose V 2 2 V 2 2 + + 2 2 + 2 2 2 +
L-Arabinose 2 2 2 2 2 2 2 2 2 V + 2 2 + +
L-Sorbose V! V! 2 V! 2 V! V! 2 2 2 2 2 V 2 2
Inositol 2 2 2 2 2 2 2 2 2 2 V 2 V V 2
Methyl a-D-mannopyranoside 2 2 ND 2 ND 2 + 2 2 2 2 2 V 2 2
Maltose + + + + + + + + + + + + + 2 +
Lactose + + + + + + + + V! (+) + 2 + 2 +
Melibiose V! V 2 2 2 V 2 + 2 2 V 2 V 2 2
Inulin V! V! 2 2 2 2 2 2 2 2 2 2 2 2 2
Melezitose V V V V V 2 2 2 2 2 2 2 V 2 2
D-Lyxose V V 2 2 V 2 V 2 2 2 2 2 2 V +
D-Glucose V! V! + V! + V! + + + + + + + + +
*Motility was reported for these species in the original descriptions (Lang Halter et al., 2013; Leclercq et al., 2010). We did not observe motility in
the type strains of these species, nor did we find genes encoding a flagellar apparatus in the genomes of the type strains (Table S4). In agreement
with our observations, Bertsch et al. (2013) did not observe motility in the type strain of L. rocourtiae.
H. C. den Bakker and others
1886 International Journal of Systematic and Evolutionary Microbiology 64

for 26 h and 41 uC for 41 h in the Synergy H1 reader.The OD600 was measured after 10 s of orbital shaking asfollows: every 2 min for the first 20 min, every 5 min forthe next 40 min and every 10 min for the remainder ofthe experiment. Growth was quantified by finding the firsttime the cell number had increased by 0.6 log (c.f.u. ml21),i.e. a 4-fold increase in cell number. Measured increases inlog(OD600) were converted to increases in log(c.f.u. ml21)using the slope of the strain-specific linear function relatinglog(OD600) to log(c.f.u. ml21). The linear function wascreated with a series of eight 2-fold serial dilutions of a24 h culture of each strain, performed in a microtitre plate,and the resulting OD600 values for each dilution. The meanslope of the resulting best-fit lines was 0.92 log(OD600)/log(c.f.u. ml21), with a mean r2 of 0.99. For the measure-ment of growth characteristics at temperatures belowambient temperature, BHI culture tubes were inoculatedat 103 c.f.u. ml21 (±0.3 log) and incubated at 4 uC (low,3 uC; high, 9 uC) and 7 uC (6.8±0.2 uC) in a refrigeratedincubator. Cultures were spiral-plated on BHI agar (todetermine c.f.u. ml21) every 24 h for 12 days. Growthwas quantified by calculating the time for the numberof cells to increase by 1.0 log(c.f.u. ml21), i.e. a 10-foldincrease in cell number, by linear interpolation of themeasured points immediately before and after the observed1.0 log increase. Growth characteristics are reported inthe species descriptions and additional data on growthcharacteristics of the individual isolates can be found inTable S4.
Motility was determined in motility test medium (MTM)(prepared according to BAM medium M103; US Food andDrug Administration, 2013). For each isolate, 13 mm tubescontaining MTM were inoculated, 1 cm under the surface,with a stab from a colony on BHI agar. Isolates wereincubated aerobically for 7 days at 22, 30 or 37 uC or for10 days at 4 uC. An isolate was considered motile if cloudygrowth beyond the stab was observed in addition tothe characteristic mushroom- or umbrella-shaped growth.Results are reported in Table 1 and the species descriptions.L. monocytogenes 10403S was included as a positive control,while L. fleischmannii DSM 24998T was included as anegative control. In addition to the motility test, draftgenomes of the type strains sequenced were queried usingBLAST (Altschul et al., 1990) for genes associated with motilityin L. monocytogenes (Table S5). Consistent with observationsof the motility test, no flagellar motility-associated geneswere found in the novel species described here.
Light microscopic observations were performed on isolatesthat were cultured on BHI agar aerobically for 18 h at 30 uC.Stationary-phase bacteria were examined on agar-coatedslides and viewed by phase-contrast microscopy usingan Olympus BX61 microscope. Digital photographs werecaptured using the SlideBook version 4.0.2.2 software(Intelligent Imaging Innovations) and length/width ratioswere based on the mean length and width of five bacteria.Results are reported in Table 1 and the species descriptions.Photographs of individual isolates can be found in Fig. S5.
Description of Listeria floridensis sp. nov.
Listeria floridensis (flo.ri.den9sis. N.L. fem. adj. floridensis ofor belonging to Florida, the state in the United States fromwhich the type strain was isolated).
Cells are 0.661.3–1.9 mm; length/width ratio 2.7. Gram-stain-positive, straight rods with rounded ends. No growthat 7 uC or below. Optimum growth temperature 37–41 uC.No motility at 4, 22, 30 or 37 uC. Voges–Proskauer-negativeand catalase-positive. CAMP-negative. Negative for reduc-tion of nitrate and nitrite. Other characteristics are reportedin Table 1. Currently the only member of the genus Listeriathat both lacks motility and is unable to reduce nitrate.A unique API Listeria numerical profile (2 710) is observedfor the type strain.
The type strain, FSL S10-1187T (5DSM 26687T5LMG28121T5BEI NR-42632T), was isolated from running waterin Florida, USA. The DNA G+C content of the type strainis 41.8 mol% (determined by genome sequencing).
Description of Listeria aquatica sp. nov.
Listeria aquatica (a.qua9ti.ca. L. fem. adj. aquatica found inwater, aquatic).
Cells are 0.6–0.761.5–2.4 mm; mean length/width ratio3.2. Gram-stain-positive, straight rods with rounded ends.No growth at 7 uC or below. Optimum growth temper-ature 37–41 uC. No motility at 4, 22, 30 or 37 uC. Voges–Proskauer-negative for type strain; positive for strainFSL S10-1181. Catalase-positive. CAMP-negative. Nitratereduction-positive; nitrite reduction-negative. Other char-acteristics are reported in Table 1. Currently, the onlymember of the genus Listeria that is unable to fermentmaltose. Among non-motile species of the genus Listeria,it is currently unique in its ability to ferment D-tagatose. Aunique API Listeria numerical profile (6 731) is observedfor the type strain and FSL S10-1181.
The type strain, FSL S10-1188T (5DSM 26686T5LMG28120T5BEI NR-42633T), was isolated from running waterin Florida, USA. The DNA G+C content of the type strainis 40.9 mol% (determined by genome sequencing).
Description of Listeria cornellensis sp. nov.
Listeria cornellensis (cor.nel.len9sis. N.L. fem. adj. cornel-lensis named after Cornell, the university where most of theresearch was performed that led to the discovery of thespecies described in this study).
Cells are 0.4–0.762.4–3.8 mm; mean length/width ratio5.3. Gram-stain-positive, straight rods with rounded ends.Optimal growth at 30–37 uC. No motility at 4, 22, 30 or37 uC. Voges–Proskauer-negative, catalase-positive. CAMP-negative. Nitrate reduction-positive, nitrite reduction-negative. Other characteristics are indicated in Table 1.While phylogenetically and genomically distinct, this speciesresembles L. grandensis for the phenotypic characteristics
Novel species of Listeria from environmental samples
http://ijs.sgmjournals.org 1887

recorded here. Among the non-motile species of the genusListeria, L. cornellensis and L. grandensis are the only onesthat are L-rhamnose-negative in the API 50 CH test(performed at 37 uC); however, the API Listeria test strip(incubated at 34 uC) shows acidification of L-rhamnose forL. grandensis and for one of the two L. cornellensis strainstested. An operon involved in rhamnose utilization could befound in the draft genome of L. grandensis TTU A1-0212T,but is absent from the draft genome of the type strain ofL. cornellensis, suggesting the absence of rhamnose utiliza-tion for the type strain of L. cornellensis and temperature-dependent ability of L. grandensis to acidify L-rhamnose. L.cornellensis can be further distinguished from L. grandensisby weak acidification of lactose. API Listeria numericalprofile 2 330 is observed for the two known isolates (TTUA1-0210T and FSL F6-0970) of this species; this numericalprofile is associated with L. ivanovii according to the APIListeria manual (bioMerieux; version 04/2007).
The type strain, TTU A1-0210T (5DSM 26689T5LMG28123T5BEI NR-42630T), was isolated from water inColorado, USA. The DNA G+C content of the type strainis 42.5 mol% (determined by genome sequencing).
Description of Listeria grandensis sp. nov.
Listeria grandensis (gran.den9sis. N.L. fem. adj. grandensisof or belonging to Grand, the county where the type strainwas isolated).
Cells are 0.6–0.762.0–3.1 mm; mean length/width ratio 4.0.Gram-stain-positive, straight rods with rounded ends.Optimal growth temperature 30–37 uC. No motility at 4,22, 30 or 37 uC. Voges–Proskauer-negative, catalase-pos-itive. CAMP-negative. Nitrite reduction-negative and nitratereduction-positive. Other characteristics are reported inTable 1. See the description of L. cornellensis for differenti-ation of L. grandensis from this species. A unique APIListeria numerical profile (2 730) is observed for thetype strain.
The type strain, TTU A1-0212T (5DSM 26688T5LMG28122T5BEI NR-42631T), was isolated from water inColorado, USA. The DNA G+C content of the type strainis 43.0 mol% (determined by genome sequencing).
Description of Listeria riparia sp. nov.
Listeria riparia (ri.pa9ri.a. L. fem. adj. riparia of the bank ofa river or stream).
Cells are 0.5–0.762.3–3.7 mm; mean length/width ratio4.8. Gram-stain-positive, straight rods with rounded ends.Optimum growth temperature 37–41 uC. No motility at 4,22, 30 or 37 uC. Voges–Proskauer-negative and catalase-positive. CAMP-negative. Nitrite reduction-negative andnitrate reduction-positive. Other characteristics are reportedin Table 1. Can be differentiated from other non-motilespecies of the genus Listeria by a combination of a-mannosidase activity and the ability to acidify L-rhamnose,
D-galactose and L-arabinose. A unique API Listeria numer-ical profile (6 710) is observed for the isolates tested (FSLS10-1204T and FSL S10-1219).
The type strain, FSL S10-1204T (5DSM 26685T5LMG28119T5BEI NR-42634T), was isolated from running waterin Florida, USA. The DNA G+C content of the type strainis 41.9 mol% (determined by genome sequencing).
Acknowledgements
This project was supported by USDA NIFA Special Research Grant2010-34459-20756 as well as USDA NIFA National Integrated FoodSafety Initiative Grants 2008-51110-04333 and 2008-51110-04688.We thank Dr Esther Angert (Cornell University, Department ofMicrobiology) for help with microscopy. We would like to thank DrCatherine Donnelly (University of Vermont) and Drs Arnout deBruin and Wilma Jacobs (RIVM, Netherlands) for their help with theVP test. We would further like to thank Barbara Bowen for her helpwith some experiments.
Reference
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J.(1990). Basic local alignment search tool. J Mol Biol 215, 403–410.
Bertsch, D., Rau, J., Eugster, M. R., Haug, M. C., Lawson, P. A.,Lacroix, C. & Meile, L. (2013). Listeria fleischmannii sp. nov., isolatedfrom cheese. Int J Syst Evol Microbiol 63, 526–532.
Bille, J., Catimel, B., Bannerman, E., Jacquet, C., Yersin, M. N.,Caniaux, I., Monget, D. & Rocourt, J. (1992). API Listeria, a new andpromising one-day system to identify Listeria isolates. Appl EnvironMicrobiol 58, 1857–1860.
Boerlin, P., Rocourt, J., Grimont, F., Grimont, P. A. D., Jacquet, C. &Piffaretti, J.-C. (1992). Listeria ivanovii subsp. londoniensis subsp. nov.Int J Syst Bacteriol 42, 69–73.
Castresana, J. (2000). Selection of conserved blocks from multiplealignments for their use in phylogenetic analysis. Mol Biol Evol 17,540–552.
Chan, J. Z.-M., Halachev, M. R., Loman, N. J., Constantinidou, C. &Pallen, M. J. (2012). Defining bacterial species in the genomic era:insights from the genus Acinetobacter. BMC Microbiol 12, 302.
Chevreux, B., Wetter, T. & Suhai, S. (1999). Genome sequenceassembly using trace signals and additional sequence information. InGerman Conference on Bioinformatics, October 4–6 1999, Hannover,Germany, pp. 45–56.
Cohan, F. M. (2001). Bacterial species and speciation. Syst Biol 50,513–524.
den Bakker, H. C., Manuel, C. S., Fortes, E. D., Wiedmann, M. &Nightingale, K. K. (2013). Genome sequencing identifies Listeriafleischmannii subsp. coloradonensis subsp. nov., isolated from a ranch.Int J Syst Evol Microbiol 63, 3257–3268.
Domınguez-Bernal, G., Muller-Altrock, S., Gonzalez-Zorn, B.,Scortti, M., Herrmann, P., Monzo, H. J., Lacharme, L., Kreft, J. &Vazquez-Boland, J. A. (2006). A spontaneous genomic deletion inListeria ivanovii identifies LIPI-2, a species-specific pathogenicityisland encoding sphingomyelinase and numerous internalins. MolMicrobiol 59, 415–432.
Errebo Larsen, H. & Seeliger, H. P. R. (1966). A mannitol fermentingListeria, Listeria grayi sp. n. In Proceedings of the Third InternationalSymposium on Listeriosis, 13–16 July 1996, Bilthoven, Netherlands, p. 35.
H. C. den Bakker and others
1888 International Journal of Systematic and Evolutionary Microbiology 64

Graves, L. M., Helsel, L. O., Steigerwalt, A. G., Morey, R. E.,
Daneshvar, M. I., Roof, S. E., Orsi, R. H., Fortes, E. D., Milillo, S. R. &other authors (2010). Listeria marthii sp. nov., isolated from the
natural environment, Finger Lakes National Forest. Int J Syst Evol
Microbiol 60, 1280–1288.
Koboldt, D. C., Zhang, Q., Larson, D. E., Shen, D., McLellan, M. D.,Lin, L., Miller, C. A., Mardis, E. R., Ding, L. & Wilson, R. K. (2012).VarScan 2: somatic mutation and copy number alteration discovery
in cancer by exome sequencing. Genome Res 22, 568–576.
Konstantinidis, K. T. & Tiedje, J. M. (2005). Towards a genome-based
taxonomy for prokaryotes. J Bacteriol 187, 6258–6264.
Konstantinidis, K. T. & Tiedje, J. M. (2007). Prokaryotic taxonomy
and phylogeny in the genomic era: advancements and challenges
ahead. Curr Opin Microbiol 10, 504–509.
Konstantinidis, K. T., Ramette, A. & Tiedje, J. M. (2006). The bacterial
species definition in the genomic era. Philos Trans R Soc Lond B Biol
Sci 361, 1929–1940.
Kuenne, C., Billion, A., Mraheil, M. A., Strittmatter, A., Daniel, R.,Goesmann, A., Barbuddhe, S., Hain, T. & Chakraborty, T. (2013).Reassessment of the Listeria monocytogenes pan-genome reveals
dynamic integration hotspots and mobile genetic elements as major
components of the accessory genome. BMC Genomics 14, 47.
Lang Halter, E., Neuhaus, K. & Scherer, S. (2013). Listeria weihen-
stephanensis sp. nov., isolated from the water plant Lemna trisulca taken
from a freshwater pond. Int J Syst Evol Microbiol 63, 641–647.
Leclercq, A., Clermont, D., Bizet, C., Grimont, P. A. D., Le Fleche-Mateos, A., Roche, S. M., Buchrieser, C., Cadet-Daniel, V., Le Monnier,
A. & other authors (2010). Listeria rocourtiae sp. nov. Int J Syst Evol
Microbiol 60, 2210–2214.
Li, H. & Durbin, R. (2009). Fast and accurate short read alignment with
Burrows-Wheeler transform. Bioinformatics 25, 1754–1760.
McLauchlin, J. & Rees, C. (2009). Genus I. Listeria Pirie 1940a 383AL.
In Bergey’s Manual of Systematic Bacteriology, 2nd edn, vol. 3, pp.
244–257. Edited by P. De Vos, G. Garrity, D. Jones, N. R. Krieg,
W. Ludwig, F. A. Rainey, K.-H. Schleifer & W. B. Whitman.New York: Springer.
Pirie, J. H. (1940). The genus Listerella Pirie. Science 91, 383.
Richter, M. & Rossello-Mora, R. (2009). Shifting the genomic goldstandard for the prokaryotic species definition. Proc Natl Acad SciU S A 106, 19126–19131.
Rijpens, N., Vlaemynck, G., Rossau, R., Herman, L. & Jannes, G.(1998). Unidentified Listeria-like bacteria isolated from cheese. LettAppl Microbiol 27, 198–202.
Rocourt, J. & Grimont, P. A. D. (1983). Listeria welshimeri sp. nov. andListeria seeligeri sp. nov. Int J Syst Bacteriol 33, 866–869.
Sauders, B. D., Overdevest, J., Fortes, E., Windham, K., Schukken, Y.,Lembo, A. & Wiedmann, M. (2012). Diversity of Listeria species in urbanand natural environments. Appl Environ Microbiol 78, 4420–4433.
Schmid, M. W., Ng, E. Y. W., Lampidis, R., Emmerth, M., Walcher,M., Kreft, J., Goebel, W., Wagner, M. & Schleifer, K.-H. (2005).Evolutionary history of the genus Listeria and its virulence genes. SystAppl Microbiol 28, 1–18.
Seeliger, H. P. R. (1981). Apathogene Listerien: L. innocua sp.n.(Seeliger et Schoofs, 1977). Zentralbl Bakteriol Parasitenkd InfektionskrHyg Abt 1 Orig A 249, 487–493 (in German).
Seeliger, H. P. R., Rocourt, J., Schrettenbrunner, A., Grimont, P. A. D.& Jones, D. (1984). Listeria ivanovii sp. nov. Int J Syst Bacteriol 34,336–337.
Stuart, S. E. & Welshimer, H. J. (1973). Intrageneric relatedness ofListeria Pirie. Int J Syst Bacteriol 23, 8–14.
US Food and Drug Administration (2013). BacteriologicalAnalytical Manual. http://www.fda.gov/food/foodscienceresearch/laboratorymethods/ucm2006949.htm
Wilgenbusch, J. C. & Swofford, D. (2003). Inferring evolutionary treeswith PAUP*. In Current Protocols in Bioinformatics, unit 6.4. New York:Wiley.
Wu, M. & Eisen, J. A. (2008). A simple, fast, and accurate method ofphylogenomic inference. Genome Biol 9, R151.
Novel species of Listeria from environmental samples
http://ijs.sgmjournals.org 1889

Supplementary material
Five new species of Listeria (L. floridensis sp. nov, L. aquatica sp. nov., L. cornellensis
sp. nov. L. riparia sp. nov., and L. grandensis sp. nov.) from agricultural and natural
environments in the United States
Henk C. den Bakker,1* Steven Warchocki,1 Emily M. Wright,1 Adam F. Allred,2
Christina Ahlstrom,3† Clyde S. Manuel,3‡ Matthew J. Stasiewicz,1 Angela Burrell,2
Sherry Roof,1 Laura Strawn,1 Esther Fortes,1§ Kendra K. Nightingale,4 Daniel Kephart,2
and Martin Wiedmann1
1Department of Food Science, Cornell University, Ithaca, NY 14853, USA 2 Life Technologies, 2130 Woodward Street, Austin, TX 78744, USA 3 Department of Animal Sciences, Colorado State University, Fort Collins, CO 80523,
USA 4 Department of Animal and Food sciences, Texas Tech University, Lubbock, TX 79409,
USA
† Present address: Production Animal Health, University of Calgary, Alberta, Canada
‡ Present address: Department of Food, Bioprocessing and Nutrition Sciences, North
Carolina State University, Raleigh, NC 27695-7624, USA
§ Present address: Hinton State Laboratory Institute, Massachusetts Department of Public
Health, Jamaica Plain, Massachusetts, USA
Correspondence: Henk C. den Bakker, [email protected], Phone: (+1) 607 255-1266,
Fax: (+1) 607 254-4868
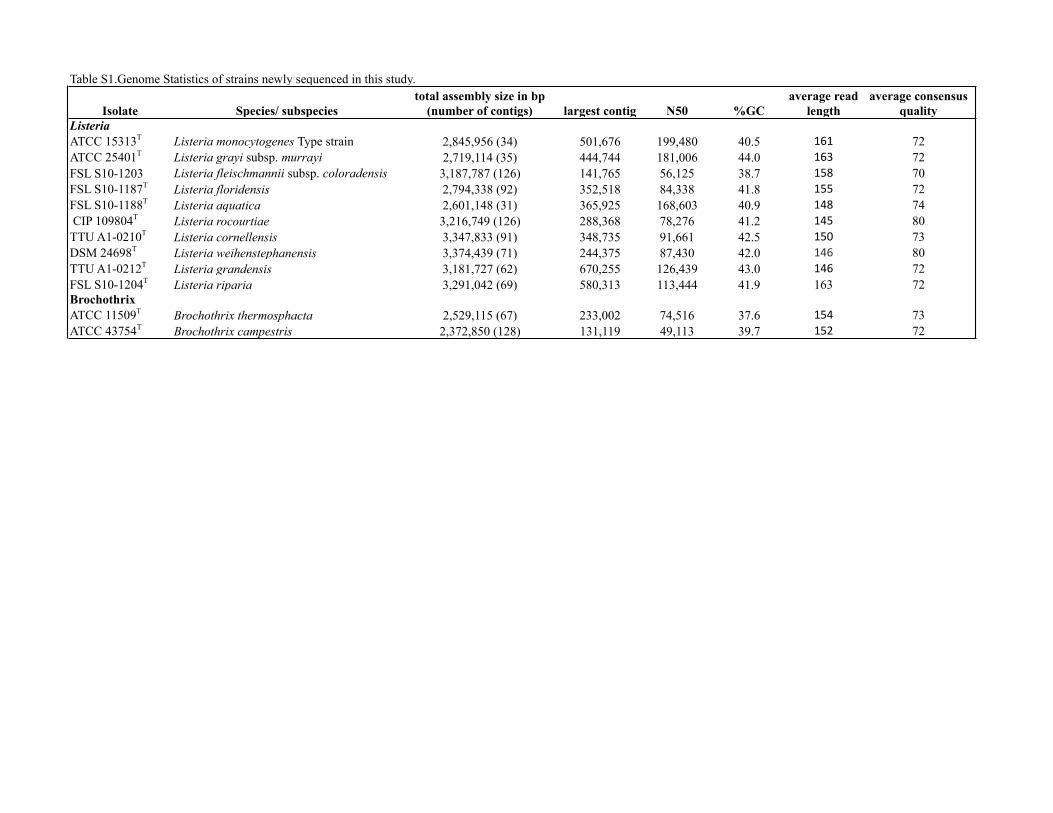
Table S1.Genome Statistics of strains newly sequenced in this study.
Isolate Species/ subspeciestotal assembly size in bp
(number of contigs) largest contig N50 %GCaverage read
lengthaverage consensus
qualityListeriaATCC 15313T Listeria monocytogenes Type strain 2,845,956 (34) 501,676 199,480 40.5 161 72ATCC 25401T Listeria grayi subsp. murrayi 2,719,114 (35) 444,744 181,006 44.0 163 72FSL S10-1203 Listeria fleischmannii subsp. coloradensis 3,187,787 (126) 141,765 56,125 38.7 158 70FSL S10-1187T Listeria floridensis 2,794,338 (92) 352,518 84,338 41.8 155 72FSL S10-1188T Listeria aquatica 2,601,148 (31) 365,925 168,603 40.9 148 74 CIP 109804T Listeria rocourtiae 3,216,749 (126) 288,368 78,276 41.2 145 80TTU A1-0210T Listeria cornellensis 3,347,833 (91) 348,735 91,661 42.5 150 73DSM 24698T Listeria weihenstephanensis 3,374,439 (71) 244,375 87,430 42.0 146 80TTU A1-0212T Listeria grandensis 3,181,727 (62) 670,255 126,439 43.0 146 72FSL S10-1204T Listeria riparia 3,291,042 (69) 580,313 113,444 41.9 163 72BrochothrixATCC 11509T Brochothrix thermosphacta 2,529,115 (67) 233,002 74,516 37.6 154 73ATCC 43754T Brochothrix campestris 2,372,850 (128) 131,119 49,113 39.7 152 72
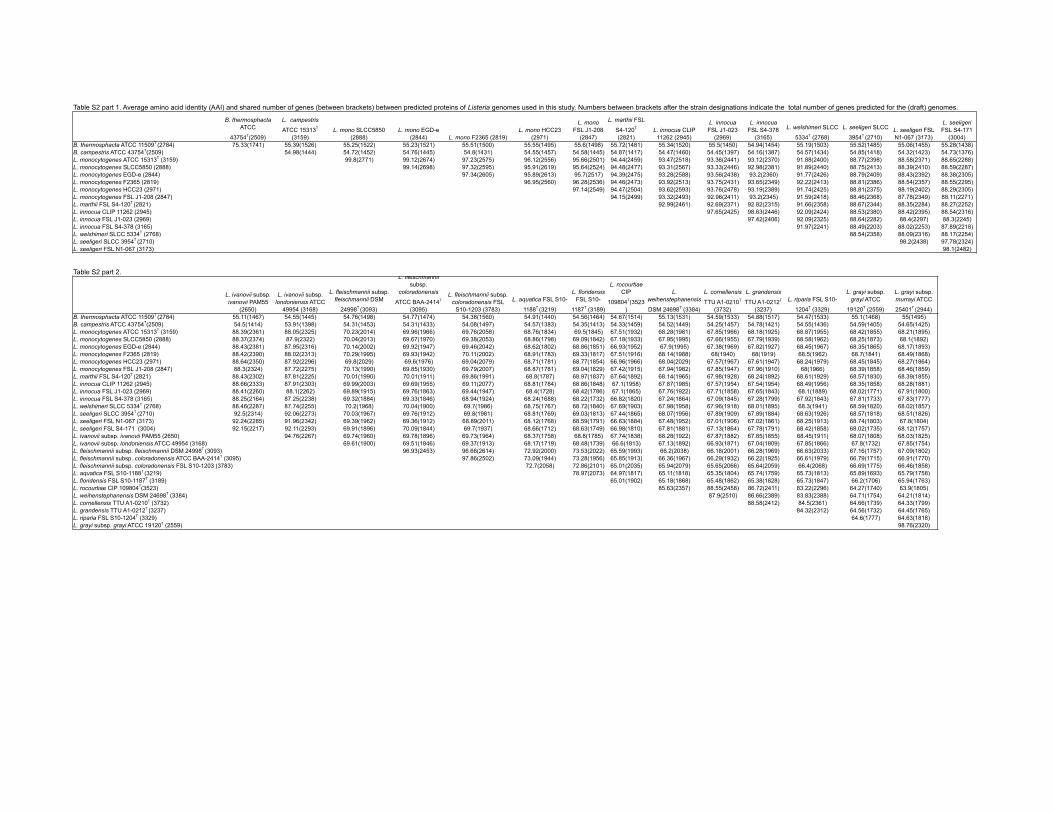
Table S2 part 1. Average amino acid identity (AAI) and shared number of genes (between brackets) between predicted proteins of Listeria genomes used in this study. Numbers between brackets after the strain designations indicate the total number of genes predicted for the (draft) genomes.
B. thermosphacta ATCC
43754T(2509)
L. campestris
ATCC 15313T
(3159)L. mono SLCC5850
(2888)L. mono EGD-e
(2844) L. mono F2365 (2819)L. mono HCC23
(2971)
L. mono FSL J1-208
(2847)
L. marthii FSL
S4-120T
(2821)L. innocua CLIP
11262 (2945)
L. innocua FSL J1-023
(2969)
L. innocua FSL S4-378
(3165)
L. welshimeri SLCC
5334T (2768)
L. seeligeri SLCC
3954T (2710)L. seeligeri FSL N1-067 (3173)
L. seeligeri FSL S4-171
(3004)B. thermosphacta ATCC 11509T (2784) 75.33(1741) 55.39(1526) 55.25(1522) 55.23(1521) 55.51(1500) 55.55(1495) 55.6(1498) 55.72(1481) 55.34(1520) 55.5(1450) 54.94(1454) 55.19(1503) 55.52(1485) 55.06(1455) 55.28(1438)B. campestris ATCC 43754T(2509) 54.98(1444) 54.72(1452) 54.76(1445) 54.8(1431) 54.55(1457) 54.58(1445) 54.87(1417) 54.47(1460) 54.45(1397) 54.16(1387) 54.57(1434) 54.85(1418) 54.32(1423) 54.73(1376)L. monocytogenes ATCC 15313T (3159) 99.8(2771) 99.12(2674) 97.23(2575) 96.12(2556) 95.66(2501) 94.44(2459) 93.47(2518) 93.36(2441) 93.12(2370) 91.88(2400) 88.77(2398) 88.58(2371) 88.65(2288)L. monocytogenes SLCC5850 (2888) 99.14(2698) 97.32(2595) 95.91(2619) 95.64(2524) 94.48(2477) 93.31(2567) 93.33(2446) 92.98(2381) 91.89(2440) 88.75(2413) 88.39(2410) 88.59(2287)L. monocytogenes EGD-e (2844) 97.34(2605) 95.89(2613) 95.7(2517) 94.39(2475) 93.28(2588) 93.56(2438) 93.2(2360) 91.77(2426) 88.79(2409) 88.43(2392) 88.38(2305)L. monocytogenes F2365 (2819) 96.95(2560) 96.28(2536) 94.46(2473) 93.92(2513) 93.75(2431) 93.65(2349) 92.22(2413) 88.81(2386) 88.54(2357) 88.55(2295)L. monocytogenes HCC23 (2971) 97.14(2549) 94.47(2504) 93.62(2593) 93.76(2478) 93.19(2389) 91.74(2425) 88.81(2375) 88.19(2402) 88.29(2305)L. monocytogenes FSL J1-208 (2847) 94.15(2499) 93.32(2493) 92.96(2411) 93.2(2345) 91.59(2418) 88.46(2368) 87.78(2349) 88.11(2271)L. marthii FSL S4-120T (2821) 92.99(2461) 92.69(2371) 92.82(2315) 91.66(2358) 88.67(2344) 88.35(2284) 88.27(2252)L. innocua CLIP 11262 (2945) 97.65(2425) 98.63(2446) 92.09(2424) 88.53(2380) 88.42(2395) 88.54(2316)L. innocua FSL J1-023 (2969) 97.42(2406) 92.09(2325) 88.64(2282) 88.4(2297) 88.3(2245)L. innocua FSL S4-378 (3165) 91.97(2241) 88.49(2203) 88.02(2253) 87.89(2218)L. welshimeri SLCC 5334T (2768) 88.54(2358) 88.09(2316) 88.17(2254)L. seeligeri SLCC 3954T (2710) 98.2(2438) 97.78(2324)L. seeligeri FSL N1-067 (3173) 98.1(2482)
Table S2 part 2.
L. ivanovii subsp. ivanovii PAM55
(2650)
L. ivanovii subsp. londoniensis ATCC
49954 (3168)
L. fleischmannii subsp. fleischmannii DSM
24998T (3093)
L. fleischmannii subsp.
coloradonensis
ATCC BAA-2414T
(3095)
L. fleischmannii subsp. coloradonensis FSL
S10-1203 (3783)
L. aquatica FSL S10-
1188T (3219)
L. floridensis FSL S10-
1187T (3189)
L. rocourtiae CIP
109804T(3523)
L. weihenstephanensis
DSM 24698T (3384)
L. cornellensis
TTU A1-0210T
(3732)
L. grandensis
TTU A1-0212T
(3237)
L. riparia FSL S10-
1204T (3329)
L. grayi subsp. grayi ATCC
19120T (2559)
L. grayi subsp. murrayi ATCC
25401T (2944)B. thermosphacta ATCC 11509T (2784) 55.11(1467) 54.55(1445) 54.76(1498) 54.77(1474) 54.38(1560) 54.91(1440) 54.56(1464) 54.67(1514) 55.13(1531) 54.59(1533) 54.88(1517) 54.47(1533) 55.1(1468) 55(1495)B. campestris ATCC 43754T(2509) 54.5(1414) 53.91(1398) 54.31(1453) 54.31(1433) 54.08(1497) 54.57(1383) 54.35(1413) 54.33(1459) 54.52(1449) 54.25(1457) 54.78(1421) 54.55(1436) 54.59(1405) 54.65(1425)L. monocytogenes ATCC 15313T (3159) 88.39(2361) 88.05(2325) 70.23(2014) 69.96(1966) 69.76(2058) 68.76(1834) 69.5(1845) 67.51(1932) 68.28(1981) 67.85(1966) 68.18(1925) 68.87(1955) 68.42(1855) 68.21(1895)L. monocytogenes SLCC5850 (2888) 88.37(2374) 87.9(2322) 70.04(2013) 69.67(1970) 69.38(2053) 68.86(1798) 69.09(1842) 67.18(1933) 67.95(1995) 67.66(1955) 67.79(1939) 68.58(1962) 68.25(1873) 68.1(1892)L. monocytogenes EGD-e (2844) 88.43(2381) 87.95(2316) 70.14(2002) 69.92(1947) 69.46(2042) 68.62(1802) 68.86(1851) 66.93(1952) 67.9(1995) 67.38(1969) 67.82(1927) 68.45(1967) 68.35(1865) 68.17(1893)L. monocytogenes F2365 (2819) 88.42(2390) 88.02(2313) 70.29(1995) 69.93(1942) 70.11(2002) 68.91(1783) 69.33(1817) 67.51(1916) 68.14(1988) 68(1940) 68(1919) 68.5(1962) 68.7(1841) 68.49(1868)L. monocytogenes HCC23 (2971) 88.64(2350) 87.92(2296) 69.8(2029) 69.6(1976) 69.04(2079) 68.71(1781) 68.77(1854) 66.96(1966) 68.04(2029) 67.57(1967) 67.61(1947) 68.24(1979) 68.45(1845) 68.27(1864)L. monocytogenes FSL J1-208 (2847) 88.3(2324) 87.72(2275) 70.13(1990) 69.85(1930) 69.79(2007) 68.87(1781) 69.04(1829) 67.42(1915) 67.94(1982) 67.85(1947) 67.96(1910) 68(1966) 68.39(1858) 68.46(1859)L. marthii FSL S4-120T (2821) 88.43(2302) 87.81(2225) 70.01(1990) 70.01(1911) 69.86(1991) 68.8(1787) 68.97(1837) 67.64(1892) 68.14(1965) 67.98(1928) 68.24(1892) 68.61(1929) 68.57(1830) 68.39(1855)L. innocua CLIP 11262 (2945) 88.66(2333) 87.91(2303) 69.99(2003) 69.69(1955) 69.11(2077) 68.81(1784) 68.86(1848) 67.1(1958) 67.87(1985) 67.57(1954) 67.54(1954) 68.49(1956) 68.35(1858) 68.28(1881)L. innocua FSL J1-023 (2969) 88.41(2260) 88.1(2262) 69.89(1915) 69.76(1863) 69.44(1947) 68.4(1728) 68.42(1786) 67.1(1865) 67.76(1922) 67.71(1858) 67.65(1843) 68.1(1889) 68.02(1771) 67.91(1800)L. innocua FSL S4-378 (3165) 88.25(2164) 87.25(2238) 69.32(1884) 69.33(1846) 68.94(1924) 68.24(1688) 68.22(1732) 66.82(1820) 67.24(1864) 67.09(1845) 67.28(1799) 67.92(1843) 67.81(1733) 67.83(1777)L. welshimeri SLCC 5334T (2768) 88.46(2287) 87.74(2255) 70.2(1968) 70.04(1900) 69.7(1986) 68.75(1767) 68.72(1840) 67.69(1903) 67.98(1958) 67.96(1918) 68.01(1895) 68.3(1941) 68.59(1820) 68.02(1857)L. seeligeri SLCC 3954T (2710) 92.5(2314) 92.06(2273) 70.03(1967) 69.76(1912) 69.8(1981) 68.81(1769) 69.03(1813) 67.44(1865) 68.07(1956) 67.89(1909) 67.89(1884) 68.63(1926) 68.57(1818) 68.51(1826)L. seeligeri FSL N1-067 (3173) 92.24(2285) 91.96(2342) 69.39(1962) 69.36(1912) 68.89(2011) 68.12(1768) 68.59(1791) 66.63(1884) 67.48(1952) 67.01(1906) 67.02(1861) 68.25(1913) 68.74(1803) 67.8(1804)L. seeligeri FSL S4-171 (3004) 92.15(2217) 92.11(2293) 69.91(1896) 70.09(1844) 69.7(1937) 68.66(1712) 68.63(1749) 66.98(1810) 67.81(1881) 67.13(1864) 67.78(1791) 68.42(1858) 68.02(1735) 68.12(1757)L. ivanovii subsp. ivanovii PAM55 (2650) 94.76(2267) 69.74(1960) 69.78(1896) 69.73(1964) 68.37(1758) 68.8(1785) 67.74(1838) 68.28(1922) 67.87(1882) 67.85(1855) 68.45(1911) 68.07(1808) 68.03(1825)L. ivanovii subsp. londoniensis ATCC 49954 (3168) 69.61(1900) 69.51(1846) 69.37(1913) 68.17(1719) 68.48(1739) 66.6(1813) 67.13(1892) 66.93(1871) 67.04(1809) 67.85(1866) 67.8(1732) 67.85(1754)L. fleischmannii subsp. fleischmannii DSM 24998T (3093) 96.93(2453) 96.66(2614) 72.92(2000) 73.53(2022) 65.59(1993) 66.2(2038) 66.18(2001) 66.28(1969) 66.63(2033) 67.16(1757) 67.09(1802)L. fleischmannii subsp. coloradonensis ATCC BAA-2414T (3095) 97.86(2502) 73.09(1944) 73.28(1956) 65.85(1913) 66.36(1967) 66.29(1932) 66.22(1925) 66.61(1979) 66.79(1715) 66.91(1770)L. fleischmannii subsp. coloradonensis FSL S10-1203 (3783) 72.7(2058) 72.86(2101) 65.01(2035) 65.94(2079) 65.65(2066) 65.64(2059) 66.4(2068) 66.69(1775) 66.46(1858)L. aquatica FSL S10-1188T (3219) 78.97(2073) 64.97(1817) 65.11(1818) 65.35(1804) 65.74(1759) 65.73(1813) 65.89(1693) 65.79(1758)L. floridensis FSL S10-1187T (3189) 65.01(1902) 65.18(1868) 65.48(1862) 65.38(1828) 65.73(1847) 66.2(1706) 65.94(1763)L. rocourtiae CIP 109804T(3523) 85.63(2357) 88.55(2458) 86.72(2411) 83.22(2296) 64.27(1740) 63.9(1805)L. weihenstephanensis DSM 24698T (3384) 87.9(2510) 86.66(2389) 83.83(2388) 64.71(1754) 64.21(1814)L. cornellensis TTU A1-0210T (3732) 88.58(2412) 84.5(2361) 64.66(1739) 64.33(1799)L. grandensis TTU A1-0212T (3237) 84.32(2312) 64.56(1732) 64.45(1765)L. riparia FSL S10-1204T (3329) 64.6(1777) 64.63(1818)L. grayi subsp. grayi ATCC 19120T (2559) 98.76(2320)

Table S3. Phenotypic characteristics of Listeriaceae strains
strain Species/subspecies
GLY
ERY
LAR
AR
IBD
XY
LLX
YL
GA
LG
LUFR
UM
NE
SBE
RH
AD
UL
INO
MA
NSO
RM
DM
MD
GN
AG
AM
YA
RB
ESC
SAL
CEL
MA
LLA
CM
ELSA
CTR
EIN
UM
LZA
MD
XLT
GEN
TUR
LYX
TAG
DFU
CD
AR
LLA
RL
DIM
ESC
αMan
Dar
lX
ylR
haM
dgR
ibG
1PTa
g
API Listeria numerical
profile Met
hyl R
edVo
ges-
Pros
kaue
rC
atal
ase
Nitr
ite R
educ
tion
Nitr
ate
Red
uctio
nC
AM
P +
Stap
hC
AM
P +
Rho
doco
ccus
Mot
ility
at 4
CM
otili
ty a
t 22C
Mot
ility
at 3
0CM
otili
ty a
t 37C
Listeria10403S L. monocytogenes - +/- - - - - - - + + + - + - - - - + + + + + + + + + + - - + - - - + + - - - - + - + + + - + + - - - 6 510 + + + - - + + - + + -
FSL S4-120T L. marthii - + - - - - - - + + + - - - - - - + + + + + + + + + +/- - - + - - - - + - - - - + - + + + - - + - - - 6 110 + + + - - - - - - - -FSL S4-378 L. innocua - +/- - - - - - - + + + - + - - - - + + + + + + + + + +/- - - + - - - + + - - - - + + + + + - + + - - - 7 510 + + + - - - - - + + +
ATCC BAA-678 L. ivanovii subsp. ivanovii - - - - + + - - + + + - - - - - - - + + + + + + + + - - - + - - - - + - - - - + + + - + + - + + + - 3 310 + + + - - - + - - - -ATCC 19120T L. grayii subsp. grayii +/- - - + - - + + + + - - - - + - + - + + + + + + + + - - + - - - + + - + - - + - + + + + - - + + - - 7 130 + + + - - - - - + + +ATCC 25401T L. grayii subsp. murrayi +/- - - + - - + + + + - + - - + - + - + + + + + + + + - - + - - + + + - + - - + - + + + + - + + + - - 7 530 + + + - + - - - + + +
ATCC BAA-2414T L. fleischmannii subsp. coloradensis + - - + + - - + + + - + - + + - + + + + + + + + + + - - + - - - +/- + - - - - +/- - - + - + + + + + - - 2 730 + - + - + - - - - - -FSL F6-1019 L. fleischmannii subsp. coloradensis + - - + + - - + + + - + - + +/- - - + + + + + + + + + - - + - - - + + - - - - - - - + - + + + + + - - 2 730 + - + - + - - - - - -
FSL S10-1186 L. fleischmannii subsp. coloradensis + - - + + - - + + + - + - + +/- - + + + + + + + + + + + + + - - - +/- + - - - - +/- - - + - + + + + + - - 2 730 + - + - + - - - - - -FSL S10-1203 L. fleischmannii subsp. coloradensis + - - + + - - + + + - + - + + - + + + + + + + + + + - - + - - - +/- + - - - - + - - + - + + + + + - - 2 730 + - + - + - - - - - -FSL S10-1220 L. fleischmannii + - - + + - - + + + - + - - +/- - - + + + + + + + + + - - + - - - +/- + - - - - + - - + - + + + + + - - 2 730 + - + - + - - - - - -
DSM 24998T L. fleischmannii subsp. fleischmannii + - - + + - - + + + - + - - + - +/- + + + + + + + + + - + + - + - + + +/- - - - + - - + - + + + + + - - 2 730 + - + - + - - - - - -FSL S10-1187T L. floridensis - - + - + - + + + + - + - - - - - + + + + + + + + + - - + - - - - + - + - - - - - + - + + + + - - - 2 710 + - + - - - - - - - -FSL S10-1188T L. aquatica + - + + + - - + + + - + - + - - - - + + + + + + - - - - + - - - - + - + + - - - - + + + + + + + - + 6 731 + - + - + - - - - - -FSL S10-1181 L. aquatica - - + + + - - + + + - + - - - - - - + + + + + + - - - - + - - - - + - +/- + - - - - + + + + + + + - + 6 731 + + + - + - - - - - -
CIP 109804T L. rocourtiae +/- - - + + - + + + + - + - - - - - +/- + + + + + + + + + - + - - - - + - - - - - - - + + + + + + + - - 6 730 + - + - + - - - - - -TTU A1-0210T L. cornellensis +/- - - + + - - + + + - - - - - - - + + + + + + + + +/- - - + - - - - + - - - - - - - + - + + - + + - - 2 330 + - + - + - - - - - -
FSL F6-0970 L. cornellensis - - + + + - - + + + - - - - - - - + + + + + + + + +/- - - + - - - - + - - - - - - - + - + + - + + - - 2 330 + - + - + - - - - - -DSM 24698T L. weihenstephanensis - - - + + - - + + + - +/- - - +/- - - + + + + + + + + - - - + - - - - + - - - - - - - + - + + + + - - - 2 710 + - +‡ -‡ +‡ - - - - - -
TTU A1-0212T L. grandensis - - - + + - - + + + - - - - - - - + + + + + + + + - - - + - - - - + - - - - - - - + - + + + + + - - 2 730 + - + - + - - - - - -FSL S10-1204T L. riparia - - + - + - + + + + - + - - +/- - - + + + + + + + + + + - + - - - + + - - - - + - - + + + + + + - - - 6 710 + - + - + - - - - - -FSL S10-1219 L. riparia +/- - + +/- + - + + + + - + - + +/- - - + + + + + + + + + +/- - + - - - - + - - - - - - - + + + + + + - - - 6 710 + - + - + - - - - - -
BrochothrixATCC 11509T B.thermosphacta - + - - + - - - + + + - - - + + - - - + + + + + + + - - + + - - - - + - - + - - - + - + - - + + - + 2 131 + + + - - - - - - - -ATCC 43754T B. campestris - + - - + - - - + + + - + - - + - - - + + + + + + + - - - + - - - - + - - - - - - + - + + + + + + + 2 771 + + + - - - - - - - -
* API 50 CH carried out at 37 °C and read at 48 hours, except Brochothrix thermosphacta ATCC 11509 and Brochothrix campestris ATCC 43754, which both were incubated at 30 °C for 48 h. Reactions positive at either timepoint are reported as positive. Variable reactions are designated as +/-.† API Listeria carried out at 34C and read at 24 hours. Variable reactions are designated as +/-.‡ Data from Lang Halter et al.( 2012). § GLY (glycerol), ERY (erythritol), DARA (D-arabinose), LARA (L-arabinose), RIB (D-ribose), DXYL (D-xylose), LXYL (L-xylose), ADO (D-adonitol), MDX (methyl-βD-xylopyranoside), GAL (D-galactose), GLU (D-glucose), FRU (D-fructose), MNE (D-mannose), SBE (L-sorbose), RHA (L-rhamnose), DUL (dulcitol), INO (inositol), MAN (D-mannitol), SOR (D-sorbitol), MDM (methyl-αD-mannopyranoside), MDG (methyl-αD-glucopyranoside), NAG (N-acetylglucosamine), AMY (amygdalin), ARB (arbutin), ESC (esculin), SAL (salicin), CEL (D-cellobiose), MAL (D-maltose), LAC (D-lactose), MEL (D-melibiose), SAC (D-saccharose), TRE (D-trehalose), INU (inulin), MLZ (D-melezitose), RAF (D-raffinose), AMD (amidon), GLYG (glycogen), XLT (xylitol), GEN (gentiobiose), TUR (D-turanose), LYX (D-lyxose), TAG (D-tagatose), DFUC (D-fuctose), LFUC (L-fucose), DARL (D-arabitol), LARL (L-arabitol), GNT (potassium gluconate), 2KG (potassium 2-ketogluconate), 5KG (potasium-5-ketogluconate), αMan (α-mannosidase), G1P (glucose-1-phosphate)|| All isolates in study negative for D-arabinose, D-adonitol, methyl-ßD-xylopyranoside, D-RAFfinose, glycogen, L-fucose, potassium gluconate, potassium 2-ketogluconate and potassium 5-ketogluconate
API Listeria† §API 50 CH* § Other
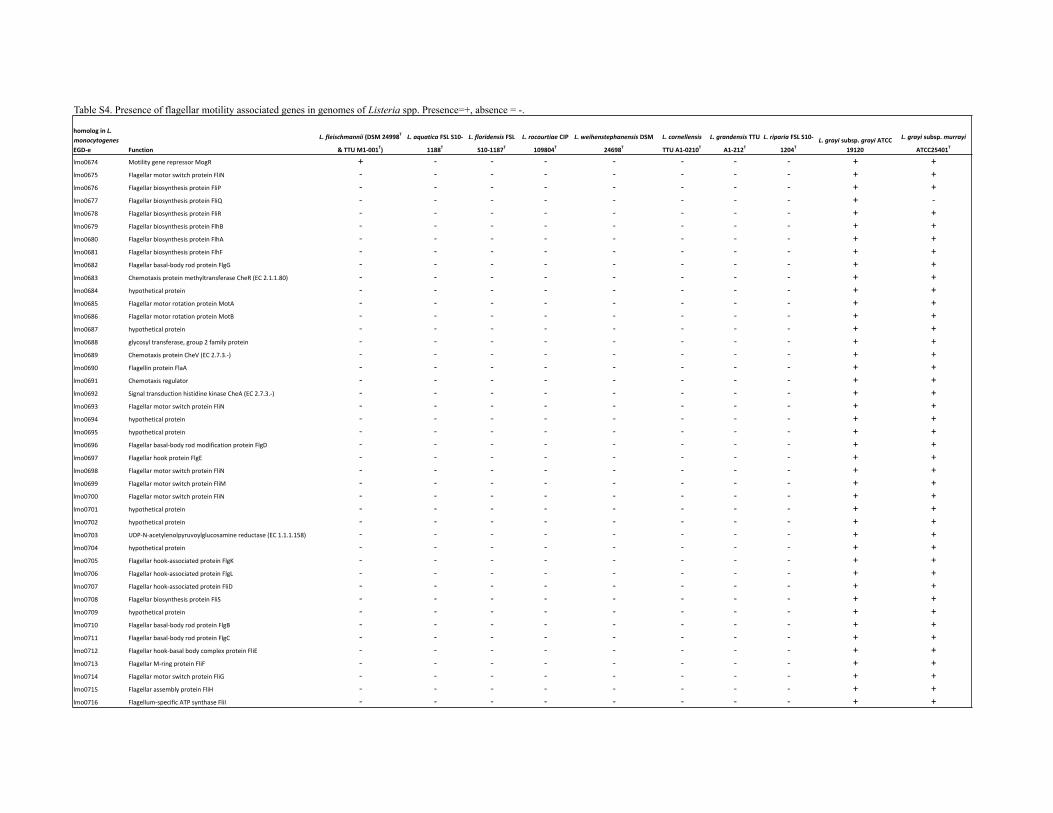
Table S4. Presence of flagellar motility associated genes in genomes of Listeria spp. Presence=+, absence = -.
homolog in L. monocytogenes EGD-‐e Function
L. fleischmannii (DSM 24998T
& TTU M1-‐001T)
L. aquatica FSL S10-‐
1188TL. floridensis FSL
S10-‐1187TL. rocourtiae CIP
109804TL. weihenstephanensis DSM
24698TL. cornellensis
TTU A1-‐0210TL. grandensis TTU
A1-‐212TL. riparia FSL S10-‐
1204TL. grayi subsp. grayi ATCC
19120
L. grayi subsp. murrayi
ATCC25401T
lmo0674 Motility gene repressor MogR + -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0675 Flagellar motor switch protein FliN -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0676 Flagellar biosynthesis protein FliP -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0677 Flagellar biosynthesis protein FliQ -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + -‐lmo0678 Flagellar biosynthesis protein FliR -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0679 Flagellar biosynthesis protein FlhB -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0680 Flagellar biosynthesis protein FlhA -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0681 Flagellar biosynthesis protein FlhF -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0682 Flagellar basal-‐body rod protein FlgG -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0683 Chemotaxis protein methyltransferase CheR (EC 2.1.1.80) -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0684 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0685 Flagellar motor rotation protein MotA -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0686 Flagellar motor rotation protein MotB -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0687 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0688 glycosyl transferase, group 2 family protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0689 Chemotaxis protein CheV (EC 2.7.3.-‐) -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0690 Flagellin protein FlaA -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0691 Chemotaxis regulator -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0692 Signal transduction histidine kinase CheA (EC 2.7.3.-‐) -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0693 Flagellar motor switch protein FliN -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0694 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0695 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0696 Flagellar basal-‐body rod modification protein FlgD -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0697 Flagellar hook protein FlgE -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0698 Flagellar motor switch protein FliN -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0699 Flagellar motor switch protein FliM -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0700 Flagellar motor switch protein FliN -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0701 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0702 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0703 UDP-‐N-‐acetylenolpyruvoylglucosamine reductase (EC 1.1.1.158) -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0704 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0705 Flagellar hook-‐associated protein FlgK -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0706 Flagellar hook-‐associated protein FlgL -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0707 Flagellar hook-‐associated protein FliD -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0708 Flagellar biosynthesis protein FliS -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0709 hypothetical protein -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0710 Flagellar basal-‐body rod protein FlgB -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0711 Flagellar basal-‐body rod protein FlgC -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0712 Flagellar hook-‐basal body complex protein FliE -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0713 Flagellar M-‐ring protein FliF -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0714 Flagellar motor switch protein FliG -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0715 Flagellar assembly protein FliH -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +lmo0716 Flagellum-‐specific ATP synthase FliI -‐ -‐ -‐ -‐ -‐ -‐ -‐ -‐ + +
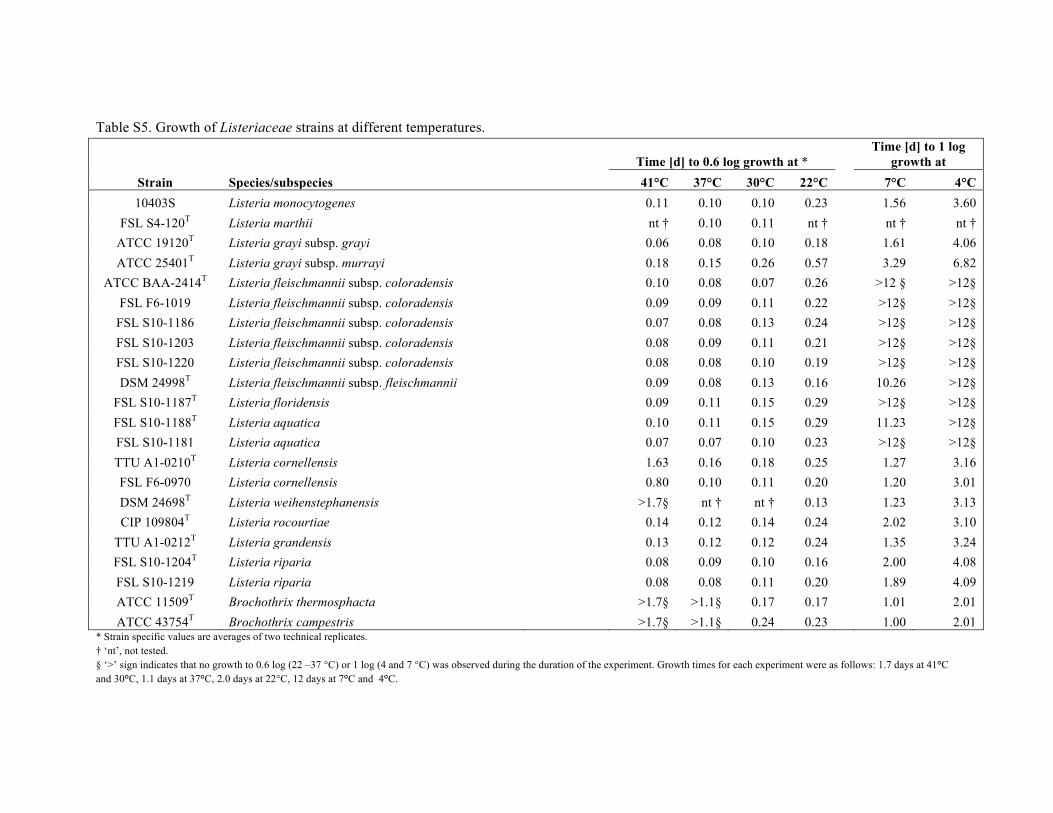
Table S5. Growth of Listeriaceae strains at different temperatures.
Strain Species/subspecies
Time [d] to 0.6 log growth at * Time [d] to 1 log
growth at 41°C 37°C 30°C 22°C 7°C 4°C
10403S Listeria monocytogenes
0.11 0.10 0.10 0.23 1.56 3.60 FSL S4-120T Listeria marthii
nt † 0.10 0.11 nt † nt † nt †
ATCC 19120T Listeria grayi subsp. grayi
0.06 0.08 0.10 0.18 1.61 4.06 ATCC 25401T Listeria grayi subsp. murrayi
0.18 0.15 0.26 0.57 3.29 6.82
ATCC BAA-2414T Listeria fleischmannii subsp. coloradensis
0.10 0.08 0.07 0.26 >12 § >12§ FSL F6-1019 Listeria fleischmannii subsp. coloradensis
0.09 0.09 0.11 0.22 >12§ >12§
FSL S10-1186 Listeria fleischmannii subsp. coloradensis
0.07 0.08 0.13 0.24 >12§ >12§ FSL S10-1203 Listeria fleischmannii subsp. coloradensis
0.08 0.09 0.11 0.21 >12§ >12§
FSL S10-1220 Listeria fleischmannii subsp. coloradensis
0.08 0.08 0.10 0.19 >12§ >12§ DSM 24998T Listeria fleischmannii subsp. fleischmannii
0.09 0.08 0.13 0.16 10.26 >12§
FSL S10-1187T Listeria floridensis
0.09 0.11 0.15 0.29 >12§ >12§ FSL S10-1188T Listeria aquatica
0.10 0.11 0.15 0.29 11.23 >12§
FSL S10-1181 Listeria aquatica
0.07 0.07 0.10 0.23 >12§ >12§ TTU A1-0210T Listeria cornellensis
1.63 0.16 0.18 0.25 1.27 3.16
FSL F6-0970 Listeria cornellensis
0.80 0.10 0.11 0.20 1.20 3.01 DSM 24698T Listeria weihenstephanensis
>1.7§ nt † nt † 0.13 1.23 3.13
CIP 109804T Listeria rocourtiae
0.14 0.12 0.14 0.24 2.02 3.10 TTU A1-0212T Listeria grandensis
0.13 0.12 0.12 0.24 1.35 3.24
FSL S10-1204T Listeria riparia
0.08 0.09 0.10 0.16 2.00 4.08 FSL S10-1219 Listeria riparia
0.08 0.08 0.11 0.20 1.89 4.09
ATCC 11509T Brochothrix thermosphacta
>1.7§ >1.1§ 0.17 0.17 1.01 2.01 ATCC 43754T Brochothrix campestris
>1.7§ >1.1§ 0.24 0.23 1.00 2.01
* Strain specific values are averages of two technical replicates. † ‘nt’, not tested. § ‘>’ sign indicates that no growth to 0.6 log (22 –37 °C) or 1 log (4 and 7 °C) was observed during the duration of the experiment. Growth times for each experiment were as follows: 1.7 days at 41°C and 30°C, 1.1 days at 37°C, 2.0 days at 22°C, 12 days at 7°C and 4°C.

Listeria_sp102
FSL_S10-1192
FSL_S10-1189
FSL_S10-1221
Listeria_innocua_ATCC33090
subsp. murrayi
FSL_S10-1204T
Listeria_marthii
FSL_S10-1199
Listeria_ivanovii_CLIP12510
TTU A1-0221
Bacillus_flexus
FSL_S10-1212
Listeria_monocytogenesNCTC10357
FSL_S10-1202
L. fleischmannii subsp. coloradensis TTU M1-001
FSL_S10-1206
TTU A1-0223
FSL_S10-1190
FSL_S10-1213
FSL_S10-1220
MB405
FSL_S10-1215
FSL_S10-1201
FSL_S10-1205
FSL_S10-1217
FSL_S10-1194
FSL_S10-1182
FSL_S10-1184
Listeria_welshimeri_ATCC_35897
TTU A1-0211
FSL_S10-1193
FSL_S10-1214
FSL_S10-1200
FSL_S10-1219
Bacillus_subtilis
FSL_S10-1183
FSL_S10-1195
FSL_S10-1207
FSL S10-1187T
FSL S10-1188
TTU_A1-0213
P. rocourtiae CIP 109804T
Brochothrix_thermosphacta
FSL_S10-1216
TTU A1-0224
FSL_S10-1208
L. fleischmannii_LU2006_1
Bacillus_cereus
FSL_S10-1186
Listeria_ivanovii_CLIP12229
FSL_S10-1210
Listeria_seeligeri_ATCC_35967
FSL_S10-1222
subsp. grayi
FSL_S10-1191
P. weihenstephanensis
FSL_S10-1203
FSL_S10-1209
Brochothrix_campestris
FSL_S10-1211
FSL_S10-1197
TTU_A1-0214
FSL_S10-1196
FSL_S10-1218
TTU_A1-0212T
FSL_S10-1185
FSL_S10-1198
FSL_S10-1181
FSL F6-969T; TTU A1-0210T
82
10099
99
100
100
91
100
93
88
98
92
80
100
72
100
99
100
9995
Streptococcus
Enterococcus
Lactobacillus
Staphylococcus
Brochothrix
L. grayi
L. aquatica
L. riparia
L. grandensis
L. cornellensis 2
L. cornellensis 1
0.06 nucleotide substitutions/site
Fig. S1.A Maximum likelihood tree based on partial (approximately 650 bp: see Ivy et al. (Appl Environ Microbiol. 2012, 78(6):1853-64), and complete or near complete (>1200 bp) 16S rDNA sequences. Values on the branches are bootstrap values based on 1000 replicates, values below 70 are not indicated. FSL and TTU isolates were obtained from environmental sampling projects in Florida and Colorado, respectively. L. fleischmannii isolates from Colorado (n=19) have not been included in this tree, with the exception of the type strain (TTU M1-001) of L. fleischmannii subsp. coloradensis. Black dots indicate isolates included in phenotypic and genomic characterizations described here (see Table 1 for details).

0.3 nucleotide substitutions/site
FSL S10-1216
FSL S10-1207
FSL S10-1187T
FSL S10-1213
FSL S10-1214
FSL S10-1218
L. ivanovii subsp. ivanovii ATCC BAA-678
FSL S10-1194
FSL S10-1181
FSL S10-1191
FSL S10-1196
FSL S10-1200
FSL S10-1222
TTU_A1-0223
TTU_A1-0212T
TTU_A1-0224
FSL S10-1202
subsp. grayi ATCC 19120T
FSL S10-1195
FSL S10-1189
FSL S10-1199
TTU_A1-0210T
TTU M1-001FSL S10-1203
L. rocourtiae CIP 109804T
TTU A1-222
FSL S10-1183
TTU_A1-0214
FSL S10-1221
L. welshimeri SLCC5334T
L. monocytogenes FSL F6-684
L. seeligeri SLCC3954T
FSL S10-1211
FSL S10-1210
FSL S10-1215
FSL S10-1190
FSL S10-1198
TTU_A1-0215
TTU A1-221
FSL S10-1184
FSL S10-1217
FSL S10-1204T
LU 2006_1
FSL S10-1188T
L. marthii FSL S4-120T
FSL S10-1185
TTU_A1-0213
FSL S10-1193
L. innocua FSL S4-378
FSL S10-1192
FSL S10-1208FSL S10-1209
FSLFSL S10-1186
subsp. murrayi ATCC 25401T
FSL S10-1197
TTU_A1-0211
L. weihenstephanensis DSM 24699T
FSL S10-1205
FSL S10-1212
FSL S10-1219FSL S10-1206
FSL S10-1182FSL S10-1201
99
95
71
87
94
99
100
98
99
94
70100
99
89
100
82
100
100
95
100
L. grandensis
L. cornellensis
L. riparia
L. aquatica
L. fleischmannii
Fig. S1.B Maximum likelihood tree based on partial ( approximately 600 bp) sigB sequences. Values on the branches are bootstrap values based on 1000 replicates, values below 70 are not indicated. FSL and TTU isolates were obtained from environmental sampling projects in Florida and Colorado, respectively. L. fleischmannii isolates from Colorado (n=19) have not been included in this tree, with the exception of the type strain (TTU M1-001) of M. fleischmannii subsp. colo-radensis. Black dots indicate isolates included in phenotypic and genomic characterizations described here (see Table 1 for details). L. riparia FSL S10-1204T and FSL S10-1219 were found to be divergent for a partial polC sequence ( polC is part of the MLST scheme described by den Bakker et al. [Appl Environ Microbiol. 2010, 76(18), 6085–6100], primers for amplification of this locus are given in den Bakker et al. [IJSEM 2013, in press, DOI 10.1099/ijs.0.048587-0]).
0.6 0.7 0.8 0.9 1.0
1400
1600
1800
2000
2200
2400
2600
2800
AAI
shar
ed g
enes
Brochothrix - ListeriaL. grayi -clade vs. L. rocourtiae - cladeL. rocourtiae-clade vs. L. �eischmanii-cladeL. grayi-clade vs. L. �eischmanii-cladeListeria s.s.-clade vs. L. grayi-cladeListeria s.s.-clade vs. L. rocourtiae - cladeListeria s.s.-clade vs. L. �eischmanii-cladeBetween spp. in L. �eischmannii-cladeBetween Brochothrix spp.Between spp. in L. rocourtiae-cladeBetween spp. in Listeria s.s.Intraspeci�c comparisons
B. campestris
B. thermosphacta
murraya
grayi
SLCCN1067
S417
1
londo
niens
isiva
novii
L. marthii
TypeSLCC5850EGD-eF2365
HCC23J1208
innocuaS4378 J1-023
L. welshimeri Type
subsp. coloradonensis Type
subsp.coloradonensis Florida
subs
p. fl
eisc
hman
niiT
Typ
e
L. floridensis
L. aquatica
L. ripariensis
L. rocourtiae
L. cornellensis
L. grandensis
L. weihenstephanensis
L. fleischmannii
L. monocytogenes
L. innocua
L. ivanovii L. seeligeri
L. grayi
L. �eischmanii-clade
Listeria s.s. clade
L. rocourtiae-clade
Brochothrix
10% AAI difference
A
B
Figure S2. (a) Average amino acid identity (AAI) values plotted against number of shared predicted genes of Listeria and Brochothrix genomes and (b) neighbor net tree based on percent average amino acid difference (1-AAI). Both the scatterplot and neighbor net tree show that species within the L. rocourtiae-clade (light green circles) and the L. fleischmannii-clade (navy blue circles), including the novel species L. grandensis sp. nov. , L. cornellensis sp. nov., L. riparia sp.nov., L. floridensis sp. nov. and L. aquatica sp. nov., are more diver-gent from each other than previously established species within the Listeria sensu stricto clade, and share a low number of genes compared to their putative genome size (as inferred from the total assembly size), thus supporting recognition as taxa on the species level.
L. �eischmanii-clade2.6-3.2 Mbp
L. rocourtiae-clade3.2 - 3.4 Mbp
Listeria s.s. clade2.7-3.1 Mbp
0.07 nucleotide substitutions per site
L. riparia FSL S10-1204T
Y12757 unidentified Bacterium isolate MB405
FR850019 L. weihenstephanensis DSM 24699T
AF003930.1 Streptococcus pneumoniae ATCC 33400
AF257097 Lactobacillus crispatus ATCC 33820
L36472 Staphylococcus aureus ATCC 12600
X56154 L. grayi subsp. murrayi NCTC 10812T
EU545982 L. marthii FSL S4-120T
X98527 L. innocua ATCC 33090T
AE016877 Bacillus cereus ATCC 14579
AF469172 Lactobacillus casei ATCC 393
X98528 L. ivanovii subsp. ivanovii CLIP12510T
X98526 L. grayi subsp. grayi ATCC 19120T
AB233327 Staphylococcus saprophyticus GTC 843
Y17361 Lactobacillus amylolyticus DSM 1664T
AJ965482 Lactobacillus plantarum NRRL B-14768
AJ276351 Bacillus subtilis DSM10
L. aquatica FSL S10-1188T
L. floridensis FSL S10-1187T
AY946290 L._sp102
AY612844.1 Streptococcus pseudopneumoniae ATCC BAA-960
DQ065845 L. seeligeri ATCC 35967T
AB012212.1 Enterococcus faecalis JCM 5803
AB021185 Bacillus flexus IFO15715
L. cornellensis TTU A1-0210T
L. fleischmannii subsp.coloradenisis TTU M1-001T
DQ065846 L. welshimeri ATCC 35897T
X56153 L. monocytogenes NCTC 10357T
L. fleischmannii subsp. fleischmannii LU2006_1T
AB002485.1 Streptococcus dysgalactiae ATCC 43078
X56156 Brochothrix campestris ATCC 43754T
AY543023 Brochothrix thermosphacta ATCC 11509T
D83363 Staphylococcus epidermidis ATCC 14990
X98529 L. ivanovii subsp. londoniensis CLIP12229T
FJ557241 L. rocourtiae CIP 109804T
AF003929.1 Streptococcus mitis ATCC 49456
L. grandensis TTU A1-0212T
AJ301830.1 Enterococcus faecium LMG 11423
AY188348.1 Streptococcus mutans ATCC 25175
X66100 Staphylococcus haemolyticus CCM2737
D16552 Lactobacillus rhamnosus JCM 1136
AY028437.2 Enterococcus phoeniculicola JLB-1
88
100
88
77
75
90
74
100
100
93
100
100
100
98
100
100
100
90
99
100
97
73
100
100
88
75
100
90
100
Listeria
Figure S3
Fig. S3 Maximum likelihood phylogeny based on 16S rRNA nucleotide sequences with a length > 1200 bp. The 16s rRNA sequences for the novel species ranged in length from 1429 to1502 bp. Values on the branches represent bootstrap values based on a 1000 replicates. Bootstrap values < 70 are not indicated.
Brochothrix

0.2
L. riparia FSL S10-1204T
L. rocourtiae CIP 109804T
L. floridensis FSL S10-1187T
L. ivanovii subsp. ivanovii PAM55
L. cornellensis_TTU_A1-0210T
L. monocytogenes EGD-e
Brochothrix thermosphacta ATCC 11509T
L. seeligeri SLCC 3954T
L. grayi subsp. grayi ATCC 19120T
L. grayi subsp. murrayi ATCC 25401T
L. weihenstephanensis DSM 24698T
L. innocua Clip11262
L. grandensis TTU A1-0212T
Brochothrix campestris ATCC 43754T
L. monocytogenes ATCC 15313T
L. ivanovii subsp. londoniensis ATCC 49954T
L._fleischmannii_subsp._coloradensis TTU M1-001T
L. marthii FSL S4-120T
L._fleischmannii_subsp. fleischmannii DSM 24998T
L. welshimeri SLCC 5334T
L. monocytogenes HCC23
L. aquatica FSL S10-1188T
L. monocytogenes F2365
97
100
100
100
82
100
100
100
93
100
100
95
96
91
100
100
100
Figure S4. Maximum likelihood phylogeny based on iap (invasion associated secreted endopeptidase)nucleotide sequences (homologs of L. monocytogenes EGD-e lmo0582). Values on the branchesrepresent bootstrap values based on 100 bootstrap replicates. Nucleotide sequences were aligned using Muscle 3.6 (R. C. Edgar, MUSCLE: multiple sequence alignment with high accuracy and highthroughput, Nucleic Acids Res 32, 1792–1797 (2004), and a Maximum likelihood tree with100 boot-strap replicates was generated using RaxML version 7.4.2.

Fig. S5 Phase contrast microscopy of strains of novel species and a representative selection of previously described species (L. monocytogenes strain 10403S; L. marthii strain FSL S4-120T; L. innocua strain FSL S4-378; L. ivanovii strain ATCC BAA-678; L. grayi subsp. grayi strain ATCC 19120Tand subsp. murrayi strain ATCC 25401T; L. rocourtiae strain CIP 109804T; L. weihenstephanensis strain DSM 24698T; L. cornellensis strains TTU A1-0210T and FSL F6-0970; L. riparia strains FSL S10-1204T, FSL S10-1219; L. grandensis strain TTU A1-0212T; L. fleischmannii strains DSM 24998T, ATCC BAA-2414T, FSL F6-1019, FSL S10-1186, FSL S10-1203, FSL S10-1220; L. aquatica strains FSL S10-1188T and FSL S10-1181; L. floridensis strain FSL S10-1187T; Brochothrix thermosphacta strain ATCC 11509T; Brochothrix campestris ATCC 43754T). Slides are labeled with strain designation and length- width ratios (average of five cells).
ATCC BAA-2414T 2.58
FSL S10-1203 3.22
FSL S10-1187T 2.70
FSL F6-1019 3.21
FSL S10-1186 2.50
ATCC 19120T 2.88
FSL S10-1181 2.87 FSL S10-1188T 3.45 CIP109804T 3.14
TTU A1-0212T 4.00 FSL F6-0970 4.84 TTU A1-0210T 5.72
FSL S10-1219 4.78 ATCC 11509T 6.82
DSM 24698T 4.75
ATCC 43754T 3.10 FSL S10-1204T 4.73
DSM 24698T 2.43 FSL S10-1220 3.47
ATCC 25401T 2.39
10403S 2.73
2
FSL S4-378 3.36 ATCC BAA-678 2.06
1
FSL S4-120T 3.50