extracellular and intracellular mechanisms that mediate the metastatic activity of exogenous...
TRANSCRIPT

Extracellular and Intracellular Mechanisms Mediating MetastaticActivity of Exogenous Osteopontin
Jami Mandelin, PhD1, Emme C. K. Lin, PhD2, Dana D. Hu, PhD1, Susan K. Knowles, PhD2,Kim-Anh Do, PhD1, Xuemei Wang, MS1, E. Helene Sage, PhD3, Jeffrey W. Smith, PhD2,Wadih Arap, MD, PhD1, and Renata Pasqualini, PhD1
1 The University of Texas M. D. Anderson Cancer Center, 1515 Holcombe Boulevard, Houston,TX2 The Program of Cell Adhesion, Cancer Research Center, The Burnham Institute, 10901 NorthTorrey Pines Road, La Jolla, CA3 Hope Heart Program, Benaroya Research Institute at Virginia Mason, 1201 Ninth Avenue,Seattle, WA
AbstractBACKGROUND—Osteopontin affects several steps of the metastatic cascade. Despite directcorrelation with metastasis in experimental systems and in patient studies, the extracellular andintracellular basis for these observations remains unsolved. We used human melanoma andsarcoma cell lines to evaluate the effects of soluble osteopontin on metastasis.
METHODS—Exogenous osteopontin or negative controls, including a site-directed mutantosteopontin, were used in functional assays in vitro, ex vivo, and in vivo designed to testextracellular and intracellular mechanisms involved in experimental metastasis.
RESULTS—In the extracellular environment, we confirm that soluble osteopontin is required forits pro-metastatic effects; this phenomenon is specific, RGD-dependent, and evident inexperimental models of metastasis. In the intracellular environment, osteopontin initially inducesrapid Tyr-418 dephosphorylation of c-Src, with decreases in actin stress fibers and increasedbinding to the vascular endothelium. This heretofore undescribed Tyr dephosphorylation isfollowed by a tandem c-Src phosphorylation after tumor cell attachment to the metastatic site.
CONCLUSION—Our results reveal a complex molecular interaction as well as a dual role forosteopontin in metastasis that is dependent on whether tumor cells are in circulation or attached.Such context-dependent functional insights may contribute to anti-metastasis strategies.
Keywordsc-Src; endothelium; metastasis; osteopontin
INTRODUCTIONOsteopontin (reviewed in 1–4) plays a central role in the metastatic potential of both humanand experimental tumors.5–7 Clinically, elevated levels of circulating osteopontin in cancerpatients correlate with increased metastasis and poor prognosis for many solid tumors, most
Requests for reprints: Wadih Arap, MD, PhD The University of Texas M. D. Anderson Cancer Center, 1515 Holcombe Boulevard,Houston, TX 77030 (e-mail: [email protected]), Jeffrey W. Smith The Burnham Institute, 10901 North Torrey Pines Road, LaJolla, CA 92037 (e-mail: [email protected]), Renata Pasqualini, PhD The University of Texas M. D. Anderson Cancer Center,1515 Holcombe Boulevard, Houston, TX 77030 (e-mail: [email protected]; phone: (713) 792-3873, fax: (713) 745-2999).
NIH Public AccessAuthor ManuscriptCancer. Author manuscript; available in PMC 2010 April 15.
Published in final edited form as:Cancer. 2009 April 15; 115(8): 1753–1764. doi:10.1002/cncr.24170.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

notably documented in malignant tumors of breast6, 8, prostate9, 10, liver11, and head andneck.12 Since the original discovery of this matricellular protein by Oldberg et al.13, theprecise extracellular and intracellular mechanisms through which osteopontin promotesmetastasis and correlates with poor prognosis in cancer patients remain unresolved, althoughthere is a recognized dependence on the arginine-glycine-aspartic acid (RGD) motif.5
Osteopontin (also termed secreted phosphoprotein-1, urinary stone protein, and early Tlymphocyte activation-1) is an acidic, secreted, non-collagenous matricellular protein withcytokine- and chemokine-like functions.1, 14 It was first isolated as a major bonesialoprotein containing an RGD motif.13 Early work established that osteopontin binds toαvβ3 integrin on osteoclasts; later, it became clear that osteopontin also recognizes severalother members of the αv family and β1 family of integrins (reviewed in 1–4). Such a broadintegrin-binding profile indicates that osteopontin mediates cell adhesion, proliferation,migration, and survival. Therefore, it is likely to be relevant in the context of tumorprogression, angiogenesis, and metastasis.
Herein, we have developed functional assays in vitro, in tumor cells, in mouse models, andex vivo with patient-derived samples to gain mechanistic insight into the role of solubleosteopontin in metastasis. Together, the results indicate the existence of an RGD-dependent,osteopontin-triggered activation cascade involving the Src oncogene pathway in non-adherent tumor cells. Because we have also shown that human cancer cells expressfunctional osteopontin cell-surface receptors regardless of tumor type, these findings havepotential implications for therapeutic intervention.
MATERIALS AND METHODSCloning and production of wild-type osteopontin and mutant osteopontin-RGE
Total RNA was isolated from cultured human KS1767 Kaposi sarcoma cells with Trizol®(Invitrogen, Carlsbad, CA). After DNase (Promega, Madison, WI) treatment, 1 μg of totalRNA was transcribed with oligo dT primers and SuperScript™ III Reverse Transcriptase(Invitrogen). One μl cDNA was amplified with forward primer 5′-ACTCGGATCCATGAGAATTGCAGTGATTTGCTTT-3′ and reverse primer 5′-TTTGCGGCCGCTTAATTGACCTCAGAAGATGCACTATCT-3′ containing BamHI andNotI restriction sites (nucleotides shown in italics). After digestion with BamHI and NotI(Roche, Basel, Switzerland), PCR products were purified by electrophoresis and cloned intopGEX-6P-1 expression vector (GE Healthcare, Piscataway, NJ). GST-osteopontin-RGE wasgenerated by amplification of the cloned osteopontin with the following primers: 5-primeend forward primer 5′-CTCGGATCCATGAGAATTGCAGTGATTTGCTTT-3′ and reverseprimer 5′-AAACCACACTTTCACCTCGGCCATCATATGTGTCT-3′, and 3-prime endforward primer 5′-ATGGCCGAGGTGAAAGTGTGGTTTATGGACTGAGGT-3′ andreverse primer 5′-TTTGCGGCCGCTTAATTGACCTCAGAAGATGCACTATCT-3′ thatgenerate 5′- and 3′-ends containing the Asp→Glu mutation. The corresponding nucleotidefor this single point mutation is shown in bold. The ends were purified, mixed in 1:1 molarratio, and used to generate a full-length mutant osteopontin-RGE. The amplification primersused were the same as those for wild-type osteopontin. The integrity of constructs wasverified by DNA sequencing and by restriction enzyme mapping.
Purified or recombinant proteins and synthetic peptidesThe GST-osteopontin (wild-type or mutant) constructs were used to transform E. coli strainBL21 (Novagen, Madison, WI). Recombinant proteins were produced as described.15Briefly, cells were grown to O.D. approximately 1.0, harvested, resuspended in phosphate-buffered saline (PBS) containing 1% Triton® X-100 and protease inhibitor cocktail (Roche),
Mandelin et al. Page 2
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and lysed by sonication. The lysate was cleared by centrifugation, and the GST proteinswere bound to glutathione-Sepharose (GE Healthcare). After three washing steps, theproteins were eluted with reduced glutathione (Sigma, St Louis, MO).
Fibronectin was purified by affinity chromatography on gelatin-Sepharose.16 A polymericform of fibronectin (superfibronectin, sFN) was generated as described.17, 18 Fibrinogenwas purchased from Enzyme Research Laboratories (South Bend, IN). Soluble peptideswere synthesized and cyclized (if necessary) at AnaSpec (San Jose, CA) to ourspecifications.
Cell cultureCell culture reagents were purchased from Invitrogen unless otherwise indicated. Humantumor cell lines C8161 (melanoma) and KRIB (osteosarcoma) were maintained in DMEMsupplemented with 10% FCS, vitamins, non-essential amino acids, and antibiotics. Cellswere grown to approximately 80% confluence at the time of use. After detachment withphosphate-buffered saline (PBS) containing 2.5 mM EDTA, cells were washed three timeswith DMEM, counted, and resuspended in DMEM prior to subsequent treatments. Cellviability was monitored before and after treatments by counting of Trypan blue-excludingcells.
In vivo assayAll animal experimentation was reviewed and approved by each respective InstitutionalAnimal Care and Use Committee (IACUC) at the Burnham Institute and at the University ofTexas M. D. Anderson Cancer Center (UTMDACC). Two-month-old nude mice (HarlanSprague Dawley) were anesthetized with Avertin® (0.015 ml/g) and were injectedintravenously (tail vein) with 106 tumor cells, which were previously treated with 200 μlDMEM containing either 1 μg/ml or 10 μg/ml of GST-osteopontin for 10 min at roomtemperature (RT). The RGE mutant of GST-osteopontin was used in the same manner and atthe same concentrations. GRGDSP and RGD-4C peptides were used at 1 mg/ml (n = 10 foreach group). These experiments were repeated three times.
Metastases were monitored during the study by the killing of sentinel tumor-bearing micefrom different cohorts at different time points (12–16 weeks) and by determination ofmetastatic tumor loads.18 In selected cohorts, the number of metastatic foci was countedunder a dissecting microscope, and a histological examination was performed after tissuefixation. Sentinel tumor-bearing mice served to estimate tumor burden prior to terminationof each cohort. Experiments were terminated when the animals displayed signs ofdiscomfort or weight loss. Actuarial survival is demonstrated in the Kaplan-Meier plot.19
Actin, integrin, and CD44 visualizationC8161 and KRIB cells were cultured on CultureWell™ cover glasses (Invitrogen). Afterseveral washes, the cells were stimulated with GST-osteopontin or GST-osteopontin-RGEfor 10 min. Cells were washed three times with PBS, fixed with PBS containing 2%paraformaldehyde, rendered permeable with 0.2% Triton® X-100 for 5 min, and stainedwith AlexaFluor488 phalloidin (Invitrogen), anti-integrin αv (Chemicon, Temecula, CA),and anti-CD44 (clone IM7; AbCam, Cambridge, MA) antibodies. These experiments wererepeated three times in duplicate.
Cell signalingThe cells were stimulated for 1, 10, 20, 30, and 60 min in suspension, after which they werecentrifuged and lysed in RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 0.25%sodium deoxycholate; pH 7.4) containing Complete™ protease inhibitors (Roche) and
Mandelin et al. Page 3
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

phosphatase inhibitors (1 mM Na3VO4 and 1 mM NaF; Sigma). Lysates (30 μg per lane)were resolved on SDS-polyacrylamide gels, transferred onto nitrocellulose membranes, andprobed with mouse anti-phosphotyrosine (clone 4G10) and rabbit anti-phospho-Src(Tyr-418) antibodies (Upstate, Temecula, CA). Total Src was detected with rabbit anti-SrcIgG (Cell Signaling Technology, Danvers, MA). These experiments were repeated threetimes.
Ex vivo cell binding assayFor analysis of the binding tumor cells to vascular endothelium, aortas were dissected frommice and cut open longitudinally, after which they were attached on 4% agarose withstaples. 105 tumor cells were treated in suspension for 10 minutes with osteopontin, withosteopontin-RGE, with osteopontin and phosphatase inhibitors (1 mM Na3VO4 and 1 mMNaF; inhibitors were administered 2 minutes before osteopontin), or DMEM. The cells wereallowed to attach to the aortas (n = 3 for each group) for 10 minutes, after which they weregently washed three times with PBS and fixed with 2% paraformaldehyde. The number ofcells bound to the vascular endothelium was determined under the light microscope. Thisexperiment was repeated twice.
Statistical analysisGiven that there were no substantially skewed data points and metastatic foci weredistributed evenly throughout the lung, the statistical significance of the differences in theextent of metastases between tumor-bearing mice cohorts was initially determined by a t-test. However, to expand statistical data to support the significance of our findings, weperformed additional non-parametric tests (i.e., Kruskal-Wallis and Wilcoxon). In addition,summary statistics, such as mean, median and standard deviation, were calculated for lungweight and metastatic foci in the lung. Scatter plots were generated based on the results,with a bar denoting the mean of each group. Due to the number of samples in each cohort,non-parametric tests were carried out based on the ranks of each variable. Dunnett’smethodology was applied to adjust for the inflated type I error rate generated from multiplecomparisons. This method provides a better power in the cases of comparing between asingle control and each of the treatment groups, while holding the maximum experiment-wise error rate to a level not exceeding the stated type I error rate. All of the statisticalmethods used supported our conclusions.
RESULTSHuman melanoma and sarcoma cells express osteopontin receptors
We selected well-established cell lines that are of human origin, from different solid tumorpathological types (i.e., melanomas and sarcomas), and with wide metastatic capability.C8161 (human melanoma) and KRIB (human osteosarcoma) cells contain the principalknown osteopontin receptors that include αvβ3 integrin, αvβ5 integrin, and CD44. The CD44receptor mediates attachment, homing, and aggregation of immune system cells.4Immunofluorescence revealed that all human tumor cell types studied expressed αv integrinsand CD44 (Fig. 1). Flow cytometric analysis confirmed high expression of these osteopontinreceptors (data not shown).
Exogenous soluble osteopontin increases experimental metastasis in mouse models in anRGD-dependent manner
We reasoned that osteopontin likely functions at several sites in the multi-step metastaticcascade (e.g., the primary tumor, the circulation, and/or the metastasis site). Relatively littleis known about the role of circulating osteopontin. We therefore asked whether exogenous
Mandelin et al. Page 4
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

administration of osteopontin--which would presumably mimic what cancer cells encounterin the bloodstream--might have an effect on tumor cell adhesion, spreading, and metastasis.After confirming that tumor cells exhibit osteopontin surface receptors, we performedexperimental metastasis assays in vivo. Tumor cell burden in the lungs was determined bymeasuring lung mass, a methodology we have previously validated as a suitable surrogate.18 Treatment of cancer cells with recombinant osteopontin enhanced colony formation inthe lungs (Fig. 2A). Treatment of C8161 melanoma cells with recombinant osteopontin wasassociated with a significant increase in lung weight (p<0.01; Fig. 2B) relative to negativecontrols that included mice treated with vehicle (DMEM) and control proteins. To excludethe possibility that the effect of exogenous osteopontin was restricted to a specific melanomacell line, we treated the KRIB human osteosarcoma cell line in a manner identical to that ofthe C8161 cells. The metastatic enhancement effect of osteopontin on sarcoma cells wassimilar to that observed with melanoma cells. Consistently, osteopontin increasedexperimental lung metastasis of osteosarcoma cells, in comparison to negative controls(p<0.01; Fig. 2C).
After establishing that exogenous osteopontin increases experimental metastasis formation,we asked whether this action is integrin-mediated. It is well established that the tripeptideRGD is an essential integrin recognition site. Thus, to address this question, we replacedaspartic acid with glutamic acid (Asp→Glu) in the RGD motif of recombinant osteopontin(GST-osteopontin-RGE mutant protein) and treated human melanoma and osteosarcomacells with it as was done with wild-type GST-osteopontin. Mutant GST-osteopontin-RGEtreatment did not increase tumor metastasis to the lungs of either cell line (p<0.001 in eachcase), relative to tumor cells treated with vehicle only (Fig. 2D and E and data not shown).This effect was reflected in the actuarial survival of the tumor-bearing mice. Indeed, micereceiving osteosarcoma cells that were pre-incubated with wild-type osteopontin had agreatly reduced actuarial survival relative to mice receiving cells that were pre-incubatedwith mutant osteopontin-RGE (Fig. 2F).
Enhancement of experimental metastasis is specific for osteopontinBecause RGD is present in other adhesive extracellular proteins that bind to αv integrins, itis possible that the pro-metastatic effect of osteopontin might merely represent a non-specific effect of any RGD motif in circulating proteins. To exclude this possibility, wecompared the effects of two RGD-containing circulating control proteins (fibronectin andfibrinogen) with that of osteopontin on experimental metastasis. Neither of these RGD-containing circulating proteins had detectable effects on lung colonization of C8161melanoma cells (Fig. 3A). Finally, to analyze the role of soluble RGD-containing syntheticpeptides in experimental metastasis formation, we also treated melanoma cells with the“classic” linear RGD peptide (sequence GRGDSP20, 21) or with the double-cyclic RGD-4Cpeptide (sequence ACDCRGDCFCG) that has binds selectively to αv integrins.22–26 Incontrast to osteopontin, treatment with RGD synthetic peptides was associated only with amild inhibitory effect on lung colonization (Fig. 3B). These data indicate that theosteopontin-mediated enhancement of experimental metastasis is relatively broad todifferent tumor cells but specific to osteopontin.
Soluble osteopontin modulates the actin cytoskeleton of tumor cellsEffective antigen-presenting cell (APC) interaction requires decreased cell rigidity, which isachieved by disengagement of the cortical actin cytoskeleton from the membrane.27 Toevaluate whether osteopontin would similarly induce relaxation of the actin cytoskeleton tofacilitate binding of tumor cells to the endothelium, we stained adherent human cancer cellswith Alexa Fluor 488-conjugated phalloidin to reveal cytoskeletal changes after osteopontinstimulation. Consistently, treatment with osteopontin for only ten minutes markedly altered
Mandelin et al. Page 5
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the actin fiber network and reduced the density of transcytoplasmic actin cables, relative tothose in cells treated with osteopontin-RGE (Fig. 4). These results indicate that cytoskeletalconfiguration could be relevant to tumor/endothelial cell functional interactions, as observedin other cell types such as certain specialized T-lymphocytes.27, 28
Osteopontin activates tumor cells through a Src-mediated signal transduction pathwayOsteopontin activates several intracellular signaling molecules after ligand-receptor bindingto cell surface integrins.3 One such molecule is c-Src, a member of the non-receptor proteintyrosine kinase family that plays a central role in signaling downstream from integrins thatregulate actin dynamics.29 We therefore investigated the activation of c-Src in osteosarcomacells after osteopontin stimulation.30 Surprisingly, we found that rapid dephosphorylation ofc-Src at Tyr-418 occurred when cells in suspension were stimulated with osteopontin foronly one minute. When cells were treated with several protein or peptide controls (e.g.,mutant osteopontin-RGE, GST, or RGD-4C peptide) or left unstimulated, suchdephosphorylation did not occur (Fig. 5A). Moreover, this result was also observed whenosteosarcoma lysates were analyzed for general tyrosine phosphorylation. Several proteins(predominantly between 50–98 kDa) were found to be rapidly dephosphorylated afterexposure of cells to osteopontin, in comparison to proteins after exposure of cells toosteopontin-RGE or to those from unstimulated cells (Fig. 5B).
Osteopontin enhances tumor cell binding to vascular endotheliumTo ask whether the specific and rapid dephosphorylation of proteins such as we observedhas biological relevance, we analyzed the binding of sarcoma cells to vascular endotheliumafter exposure to osteopontin but in the presence of phosphatase inhibitors (1 mM Na3VO4and 1 mM NaF). Indeed, osteopontin enhanced tumor cell binding to vascular endothelium,and phosphatase inhibitors completely abolished this binding (Fig. 6A and B), dataindicating that the interaction of osteopontin with disseminated tumor cells leading to a rapiddephosphorylation of proteins, including c-Src, has a significant impact on circulating tumorcell behavior. This effect was also seen in vivo: the hearts of the mice contained moretumors after they were injected with cells that were treated with osteopontin relative to thosetreated with DMEM (Fig. 6C).
DISCUSSIONOsteopontin has pleotropic roles in tumor cell biology. Although it is involved inangiogenesis, apoptosis, anchorage-independent cell growth, and metastasis, themechanisms accounting for these functions have not been entirely explained.4 Many studieshave demonstrated the effect of osteopontin in tumor growth and metastasis, with differenttypes of solid tumors and sites of metastasis.5–7, 31–34 However, these reports havefocused on the effect of endogenous osteopontin. Because osteopontin presumably functionsat several sites in the multi-step metastatic cascade, altering the level of endogenousexpression of osteopontin does not necessarily elucidate the role of circulating osteopontinin metastasis. We reasoned that direct stimulation of tumor cells with exogenous osteopontinwould minimize systemic effects and gain insight on whether circulating osteopontin affectsthe metastasis of disseminated tumor cells. We decided to use recombinant osteopontinbecause cells adhere to recombinant and native osteopontin by similar mechanisms.15, 35Our findings show that osteopontin has marked and reproducible pro-metastatic effects onhuman melanoma and sarcoma cells. Osteopontin often induced a doubling in experimentallung metastasis that was ultimately reflected in a severely-reduced actuarial survival rate oftumor-bearing mice. This result is particularly relevant because cancer patients withincreased serum osteopontin also have more metastases and poor overall survival.2–4, 6, 8–12 Although the lung colony assay is indeed not a bone fide metastasis model and the
Mandelin et al. Page 6
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

findings might not apply generally to the metastasis of tumors in vivo, our functional resultssuggest that circulating osteopontin is not only a correlative biomarker for metastaticpotential but also enhances the capacity of circulating tumor cells to establish metastases.Our functional results indicate that circulating osteopontin is not only a correlativebiomarker for metastatic potential but also enhances the capacity of circulating tumor cellsto establish metastases. Thus, the studies presented here provide a previously unrecognizedfunction for circulating osteopontin.
The pro-metastatic effects of osteopontin require an intact RGD motif, an observationindicating that a molecular interaction with integrins is critical for enhancement of lungcolonization. Although this possibility could perhaps have been anticipated because theRGD domain in osteopontin contributes to lymphatic metastases of breast cancer5, our workprovides novel insights for this finding and for the role of osteopontin in metastatic activity.First, soluble exogenous osteopontin appears to function in a manner similar to that ofendogenous osteopontin. Second, cells do not need to be adherent (i.e., they can be incirculation) to respond to osteopontin. Third, our results demonstrate that osteopontinpromotes RGD-dependent endothelial adherence of disseminated tumor cells and thusexplain the early advantage of osteopontin to metastatic spread. Fourth, a previous report onthe functional role of RGD used an osteopontin construct in which the entire RGD-sequencewas deleted5; in contrast, in this work we used the classic negative control (RGE) in thissetting: a single point mutation (Asp→Glu) is less likely to generate unpredictable structuraleffects (i.e., steric hindrance) within the osteopontin molecule.
Finally, the effects of osteopontin on lung colonization contrast with the effects of other(control) RGD-containing proteins and peptides; serum proteins such as soluble fibrinogenand fibronectin have little or no discernible effects. As an additional positive control, apolymeric form of fibronectin blocked metastasis (data not shown), an effect that has beenpreviously reported.18 Small RGD-containing peptides also exhibited a mild inhibitoryeffect on experimental metastasis; this result is consistent with several earlier reportsshowing that soluble RGD-peptides and RGD-containing snake venom proteins can reduceexperimental metastasis.36–38 Therefore, osteopontin behaves quite differently from otherRGD-containing molecules. Although the reason for this functional distinction is unclear,one possibility is that CD44 also functions as a receptor for osteopontin in our model.31,39–41 Both CD44 and αv integrins are expressed on the tumor cells used in our studies, andwe have confirmed that the isoforms v6/7 and v9 of CD44 that have been shown to bindosteopontin35, 42 are expressed in osteosarcoma cells (unpublished observations). However,our findings in in vitro, in vivo, and ex vivo assays consistently indicate that the RGD motifis required for the pro-metastatic function of osteopontin. Whether or not other receptors areinvolved and how they cooperate with αv (and perhaps other) integrins in the interactionbetween cancer cells and osteopontin remain to be determined.
It is possible, although unlikely, that the pro-metastatic effects of osteopontin are due to itsdirect effects on the host nude mice. First, mixture of tumor cells and osteopontin results inan approximately 10-fold dilution of osteopontin (with a total blood volume in mice ofapproximately 2 ml). This dilution reduces the concentration of osteopontin below integrin-binding levels. Second, attached tumor cells responded to exogenous soluble osteopontinwith a reduction in cytoskeletal actin fibers and, in suspension, with a rapid inactivation ofc-Src that led to enhanced binding to vascular endothelium. These in vitro events are host-independent and were osteopontin-specific. Thus, despite evidence that host-related factorscan also influence metastasis43, the in vitro data presented here clearly indicate thatosteopontin directly affects tumor cells (the “seed”) rather than the lung microenvironment(the “soil”). However, our results do not exclude the non-mutually exclusive possibility thatlocally produced osteopontin (by host or tumor cell) also affects either tumor or host cells
Mandelin et al. Page 7
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

after the tumor cells exit the bloodstream and establish metastases at distant sites; suchpossibility can be addressed in further studies.
The oncogene Src is important in the regulation of cytoskeletal structure and actin dynamics.Osteopontin affects osteoclast migration and function; notably, osteoclasts are extremelymotile, bone-resorbing cells with high rates of cytoskeletal turnover29 through c-Src.44Thus, we evaluated c-Src activation in cancer cells after exposure to osteopontin bydetermination of the state of phosphorylation of the conserved residue Tyr-418 within theactivation loop. Phosphorylation of c-Src at that site enhances kinase activity.30Unexpectedly, osteopontin induced rapid dephosphorylation of Tyr-418, data indicatinginactivation of c-Src after osteopontin exposure. This new observation is in striking contrastto earlier reports describing the effects of osteopontin in the context of c-Src activation.However, all the previous studies were carried out with immobilized osteopontin andadherent cells45, or in different experimental settings.44, 46 Indeed, we were able toconfirm similar c-Src activation through Tyr-418 phosphorylation when cells were allowedto adhere to plastic dishes. Arguably, the analysis of c-Src phosphorylation in a cellsuspension is more relevant, because circulating tumor cells encounter osteopontin inplasma. Our experiments show that rapid changes in signaling take place after tumor cellsinteract with osteopontin. This result is further supported by the fact that addition of solubleosteopontin to adherent cells causes reduction of actin stress fibers. In addition, exposure toosteopontin induced tumor cell binding to vascular endothelium, which was abolished byphosphatase inhibitors. Aortic metastasis rarely (if ever) occurs in patients but one mightspeculate that the strong blood flow accompanied by shear forces within the aorticcirculation could retard, or even prevent, metastasis. Thus, we believe that this“reductionist” aortic assay in vitro--while limited and restricted in scope relative to in vivosettings--might simulate at least some of the early general events that may occur when tumorcells encounter an intact vascular endothelium. Based on all our in vitro results, one mightsurmise that osteopontin induces similar cytoskeletal arrangements in tumor cells that occurduring T-cell activation28, when interaction with APCs involves the formation of large areasof intimate cell-membrane contact. Faure et al. has reported that disanchoring of the corticalactin cytoskeleton from the plasma membrane decreases cellular rigidity and leads to moreefficient T cell-APC complex formation; such disanchoring was achieved through rapidinactivation of ezrin-radixin-moesin proteins via a Vav1-Rac1 pathway.27 This type of cellrelaxation could allow clustering of adhesion molecules on the cell membrane and thusfacilitate tumor cell binding and penetration of the vascular endothelium. Although ourresults support this hypothesis, extensive further experimentation will be required to confirmand expand this interpretation.
In summary, we show that osteopontin has marked pro-metastatic effects in experimentalmodels of metastasis. We also show that the RGD motif in osteopontin is functional andrequired for the effects. Finally, in non-adherent cells we observed Src dephosphorylation,and enhanced tumor cell binding to the vascular endothelium that was completely blockedby phosphatase inhibitors. This osteopontin-dependent biological phenomenon might be amolecular mechanism for tumor cells in suspension. If so, osteopontin could affect Srcregulation and the release of adhesion molecules from the cytoskeleton, a reaction allowingtheir rearrangement on the cell membrane that facilitates enhanced cell binding to vascularendothelium. Presumably, tumor cells undergo two types of osteopontin-mediated sequentialeffects: (i) an initial rapid Src dephosphorylation accompanied by an increase in thecompliance of the cell membrane while in circulation, and (ii) a later Src phosphorylationafter cells attach to a metastasis site. If confirmed, these mechanistic data might lead tonovel anti-metastatic strategies in human cancer.
Mandelin et al. Page 8
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgmentsWe thank Dr. Emmanuel Dias-Neto for comments on the manuscript and Dr. Amin Hajitou for technical assistance.
This work was supported by grants CA69306 (JWS), CA30199 (RP and JWS), and CA90270 (RP and WA) fromthe National Institutes of Health, grant DAMD 17-98-1-8041 from the Department of Defense (RP) and grants fromthe California Breast Cancer Research Program (JWS and ECKL). The Army Breast Cancer Research Program(DDH) provided additional support. WA and RP received support from the Gillson-Longenbaugh Foundation. JMreceived support from the Helsingin Sanomat Centennial Foundation, the Emil Aaltonen Foundation, the Researchand Science Foundation of Farmos, and the Maud Kuistila Memorial Foundation.
References1. Denhardt DT, Noda M, O'Regan AW, Pavlin D, Berman JS. Osteopontin as a means to cope with
environmental insults: regulation of inflammation, tissue remodeling, and cell survival. J ClinInvest. 2001; 107:1055–1061. [PubMed: 11342566]
2. Furger KA, Menon RK, Tuck AB, Bramwell VH, Chambers AF. The functional and clinical roles ofosteopontin in cancer and metastasis. Curr Mol Med. 2001; 1:621–632. [PubMed: 11899236]
3. Rangaswami H, Bulbule A, Kundu GC. Osteopontin: role in cell signaling and cancer progression.Trends Cell Biol. 2006; 16:79–87. [PubMed: 16406521]
4. Weber GF. The metastasis gene osteopontin: a candidate target for cancer therapy. Biochim BiophysActa. 2001; 1552:61–85. [PubMed: 11825687]
5. Allan AL, George R, Vantyghem SA, et al. Role of the integrin-binding protein osteopontin inlymphatic metastasis of breast cancer. Am J Pathol. 2006; 169:233–246. [PubMed: 16816376]
6. Bramwell VH, Doig GS, Tuck AB, et al. Serial plasma osteopontin levels have prognostic value inmetastatic breast cancer. Clin Cancer Res. 2006; 12:3337–3343. [PubMed: 16740755]
7. Kang Y, Siegel PM, Shu W, et al. A multigenic program mediating breast cancer metastasis to bone.Cancer Cell. 2003; 3:537–549. [PubMed: 12842083]
8. Singhal H, Bautista DS, Tonkin KS, et al. Elevated plasma osteopontin in metastatic breast cancerassociated with increased tumor burden and decreased survival. Clin Cancer Res. 1997; 3:605–611.[PubMed: 9815727]
9. Hotte SJ, Winquist EW, Stitt L, Wilson SM, Chambers AF. Plasma osteopontin: associations withsurvival and metastasis to bone in men with hormone-refractory prostate carcinoma. Cancer. 2002;95:506–512. [PubMed: 12209742]
10. Ramankulov A, Lein M, Kristiansen G, Loening SA, Jung K. Plasma osteopontin in comparisonwith bone markers as indicator of bone metastasis and survival outcome in patients with prostatecancer. Prostate. 2006; 67:330–340. [PubMed: 17192877]
11. Ye QH, Qin LX, Forgues M, et al. Predicting hepatitis B virus-positive metastatic hepatocellularcarcinomas using gene expression profiling and supervised machine learning. Nat Med. 2003;9:416–423. [PubMed: 12640447]
12. Le QT, Sutphin PD, Raychaudhuri S, et al. Identification of osteopontin as a prognostic plasmamarker for head and neck squamous cell carcinomas. Clin Cancer Res. 2003; 9:59–67. [PubMed:12538452]
13. Oldberg A, Franzen A, Heinegard D. Cloning and sequence analysis of rat bone sialoprotein(osteopontin) cDNA reveals an Arg-Gly-Asp cell-binding sequence. Proc Natl Acad Sci U S A.1986; 83:8819–8823. [PubMed: 3024151]
14. Bornstein P, Sage EH. Matricellular proteins: extracellular modulators of cell function. Curr OpinCell Biol. 2002; 14:608–616. [PubMed: 12231357]
15. Xuan JW, Hota C, Chambers AF. Recombinant GST-human osteopontin fusion protein isfunctional in RGD-dependent cell adhesion. J Cell Biochem. 1994; 54:247–255. [PubMed:8175899]
16. Engvall E, Ruoslahti E. Binding of soluble form of fibroblast surface protein, fibronectin, tocollagen. Int J Cancer. 1977; 20:1–5. [PubMed: 903179]
17. Morla A, Zhang Z, Ruoslahti E. Superfibronectin is a functionally distinct form of fibronectin.Nature. 1994; 367:193–196. [PubMed: 8114919]
Mandelin et al. Page 9
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

18. Pasqualini R, Bourdoulous S, Koivunen E, Woods VL Jr, Ruoslahti E. A polymeric form offibronectin has antimetastatic effects against multiple tumor types. Nat Med. 1996; 2:1197–1203.[PubMed: 8898745]
19. Kaplan E, Meier P. Nonparametric estimation from incomplete observations. J Am Stat Assoc.1958; 53:157–481.
20. Pierschbacher MD, Ruoslahti E. Variants of the cell recognition site of fibronectin that retainattachment-promoting activity. Proc Natl Acad Sci U S A. 1984; 81:5985–5988. [PubMed:6237366]
21. Ruoslahti E. RGD and other recognition sequences for integrins. Annu Rev Cell Dev Biol. 1996;12:697–715. [PubMed: 8970741]
22. Arap W, Pasqualini R, Ruoslahti E. Cancer treatment by targeted drug delivery to tumorvasculature in a mouse model. Science. 1998; 279:377–380. [PubMed: 9430587]
23. Assa-Munt N, Jia X, Laakkonen P, Ruoslahti E. Solution structures and integrin binding activitiesof an RGD peptide with two isomers. Biochemistry. 2001; 40:2373–2378. [PubMed: 11327857]
24. Hajitou A, Trepel M, Lilley CE, et al. A hybrid vector for ligand-directed tumor targeting andmolecular imaging. Cell. 2006; 125:385–398. [PubMed: 16630824]
25. Koivunen E, Wang B, Ruoslahti E. Phage libraries displaying cyclic peptides with different ringsizes: ligand specificities of the RGD-directed integrins. Biotechnology (N Y). 1995; 13:265–270.[PubMed: 9634769]
26. Pasqualini R, Koivunen E, Ruoslahti E. Alpha v integrins as receptors for tumor targeting bycirculating ligands. Nat Biotechnol. 1997; 15:542–546. [PubMed: 9181576]
27. Faure S, Salazar-Fontana LI, Semichon M, et al. ERM proteins regulate cytoskeleton relaxationpromoting T cell-APC conjugation. Nat Immunol. 2004; 5:272–279. [PubMed: 14758359]
28. Dustin ML, Cooper JA. The immunological synapse and the actin cytoskeleton: molecularhardware for T cell signaling. Nat Immunol. 2000; 1:23–29. [PubMed: 10881170]
29. Linder S, Aepfelbacher M. Podosomes: adhesion hot-spots of invasive cells. Trends Cell Biol.2003; 13:376–385. [PubMed: 12837608]
30. Tang H, Hao Q, Rutherford SA, Low B, Zhao ZJ. Inactivation of SRC family tyrosine kinases byreactive oxygen species in vivo. J Biol Chem. 2005; 280:23918–23925. [PubMed: 15851473]
31. Ito T, Hashimoto Y, Tanaka E, et al. An inducible short-hairpin RNA vector against osteopontinreduces metastatic potential of human esophageal squamous cell carcinoma in vitro and in vivo.Clin Cancer Res. 2006; 15(12):1308–1316. [PubMed: 16489088]
32. Shevde LA, Samant RS, Paik JC, et al. Osteopontin knockdown suppresses tumorigenicity ofhuman metastatic breast carcinoma, MDA-MB-435. Clin Exp Metastasis. 2006; 23:123–133.[PubMed: 16830223]
33. Suzuki M, Mose E, Galloy C, Tarin D. Osteopontin gene expression determines spontaneousmetastatic performance of orthotopic human breast cancer xenografts. Am J Pathol. 2007;171:682–692. [PubMed: 17620367]
34. McAllister SS, Gifford AM, Greiner AL, et al. Systemic endocrine instigation of indolent tumorgrowth requires osteopontin. Cell. 2008; 133:994–1005. [PubMed: 18555776]
35. Katagiri YU, Sleeman J, Fujii H, et al. CD44 variants but not CD44s cooperate with beta1-containing integrins to permit cells to bind to osteopontin independently of arginine-glycine-aspartic acid, thereby stimulating cell motility and chemotaxis. Cancer Res. 1999; 59:219–226.[PubMed: 9892210]
36. Humphries MJ, Olden K, Yamada KM. A synthetic peptide from fibronectin inhibits experimentalmetastasis of murine melanoma cells. Science. 1986; 233:467–470. [PubMed: 3726541]
37. Morris VL, Schmidt EE, Koop S, et al. Effects of the disintegrin eristostatin on individual steps ofhematogenous metastasis. Exp Cell Res. 1995; 219:571–578. [PubMed: 7641809]
38. Trikha M, De Clerck YA, Markland FS. Contortrostatin, a snake venom disintegrin, inhibits beta 1integrin-mediated human metastatic melanoma cell adhesion and blocks experimental metastasis.Cancer Res. 1994; 54:4993–4998. [PubMed: 7520832]
39. Lin YH, Yang-Yen HF. The osteopontin-CD44 survival signal involves activation of thephosphatidylinositol 3-kinase/Akt signaling pathway. J Biol Chem. 2001; 276:46024–46030.[PubMed: 11590166]
Mandelin et al. Page 10
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

40. Teramoto H, Castellone MD, Malek RL, et al. Autocrine activation of an osteopontin-CD44-Racpathway enhances invasion and transformation by H-RasV12. Oncogene. 2005; 24:489–501.[PubMed: 15516973]
41. Weber GF, Ashkar S, Glimcher MJ, Cantor H. Receptor-ligand interaction between CD44 andosteopontin (Eta-1). Science. 1996; 271:509–512. [PubMed: 8560266]
42. Khan SA, Cook AC, Kappil M, et al. Enhanced cell surface CD44 variant (v6, v9) expression byosteopontin in breast cancer epithelial cells facilitates tumor cell migration: novel post-transcriptional, post-translational regulation. Clin Exp Metastasis. 2005; 22:663–673. [PubMed:16691370]
43. Crawford HC, Matrisian LM, Liaw L. Distinct roles of osteopontin in host defense activity andtumor survival during squamous cell carcinoma progression in vivo. Cancer Res. 1998; 58:5206–5215. [PubMed: 9823334]
44. Hruska KA, Rolnick F, Huskey M, Alvarez U, Cheresh D. Engagement of the osteoclast integrinalpha v beta 3 by osteopontin stimulates phosphatidylinositol 3-hydroxyl kinase activity.Endocrinology. 1995; 136:2984–2992. [PubMed: 7540546]
45. Das R, Mahabeleshwar GH, Kundu GC. Osteopontin induces AP-1-mediated secretion ofurokinase-type plasminogen activator through c-Src-dependent epidermal growth factor receptortransactivation in breast cancer cells. J Biol Chem. 2004; 279:11051–11064. [PubMed: 14704150]
46. Chellaiah M, Fitzgerald C, Alvarez U, Hruska K. c-Src is required for stimulation of gelsolin-associated phosphatidylinositol 3-kinase. J Biol Chem. 1998; 273:11908–11916. [PubMed:9565618]
Mandelin et al. Page 11
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIGURE 1.Osteopontin receptors are expressed on human cancer cells. Cultured human C8161melanoma and KRIB osteosarcoma cells were immunostained for two of the majorosteopontin receptors: αv integrins and CD44. Abundant punctuate αv integrinimmunoreactivity was detected in all cell types. The same cells displayed also CD44immunoreactivity. Negative staining controls with irrelevant isotype control plus secondaryantibodies displayed no immunoreactivity. Scale bar, 20 μm.
Mandelin et al. Page 12
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIGURE 2.Tumor cell exposure to exogenous osteopontin increases experimental metastasis that isRGD-dependent. A, Typical gross appearance of lungs after intravenous administration oftumor cells. Arrows indicate tumor foci in the lungs. Scale bar, 0.5 cm. B, C8161 melanomacells were pre-incubated for 10 min with DMEM, GST or GST-osteopontin and weresubsequently intravenously injected into the tail vein of nude mice (106 cells/mouse). After12–16 weeks, animals were killed and the lung weights were measured (18). The weight wassignificantly increased (p<0.01; asterisk) in nude mice receiving human melanoma cellspreviously incubated with GST-osteopontin relative to DMEM- or GST-treated cells. C,Lung weight of mice receiving human KRIB osteosarcoma cells, which were pretreated for10 min with GST or GST-osteopontin. Average normal lung weight (0.175 g) is indicated bythe line. A significant increase in lung metastasis was observed when tumor cells weretreated with GST-osteopontin relative to GST (p<0.01). D and E, Tumor cells were pre-incubated for 10 min with GST-osteopontin-RGE or GST-osteopontin. The Asp→Glu(termed RGE) mutation, which disrupts the integrin-binding RGD-domain in osteopontin,abolished its effects on tumor cells as seen in a statistically significant difference in lungweights of mice receiving C8161 melanoma cells (p<0.001) or KRIB osteosarcoma cells(p<0.001). Average normal lung weight (0.175 g) is indicated by the line. F, In a relatedexperiment, mice received KRIB osteosarcoma cells treated with either GST-osteopontin orGST-osteopontin-RGE prior to administration. Survival of tumor-bearing mice wasmonitored, and the experiment was terminated at day 80 as indicated.
Mandelin et al. Page 13
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
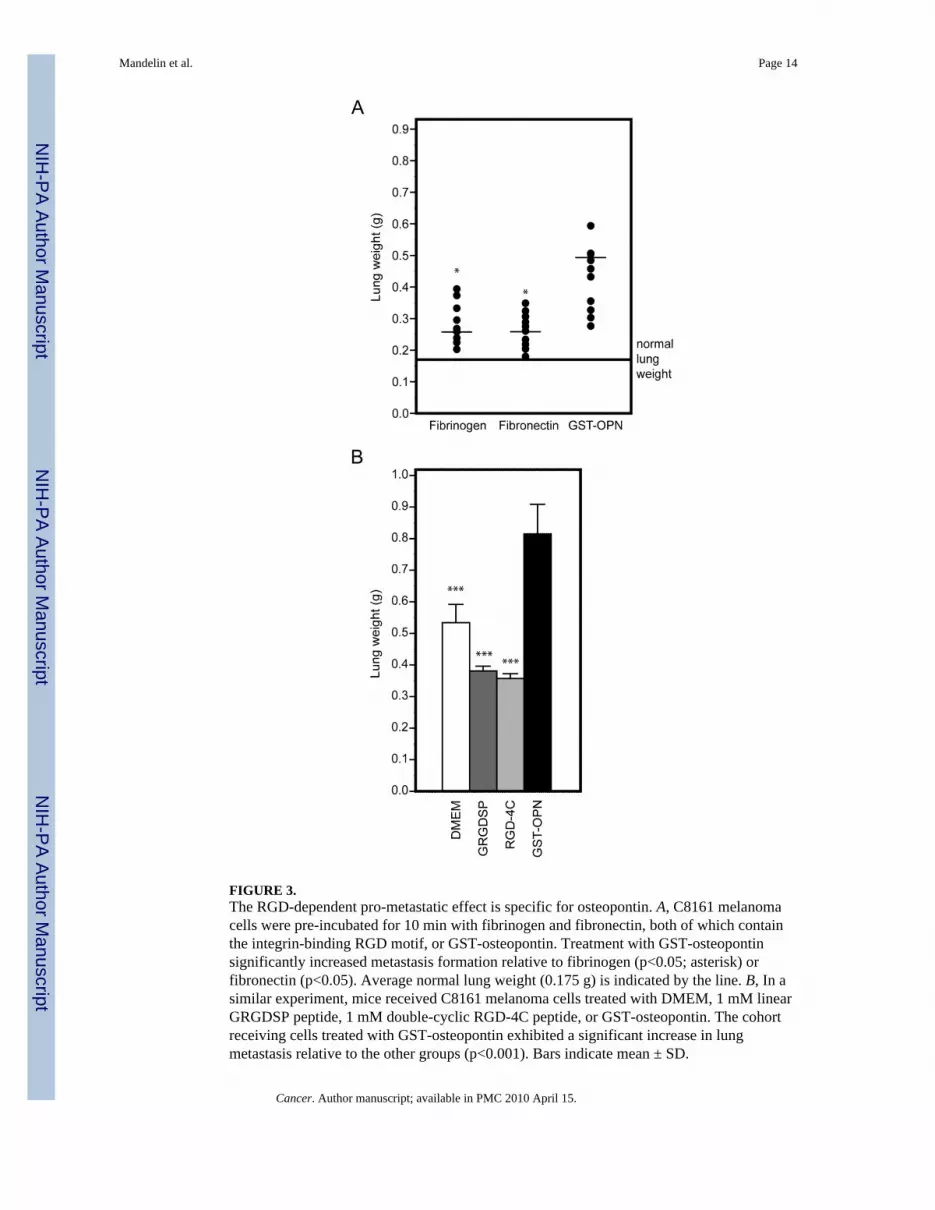
FIGURE 3.The RGD-dependent pro-metastatic effect is specific for osteopontin. A, C8161 melanomacells were pre-incubated for 10 min with fibrinogen and fibronectin, both of which containthe integrin-binding RGD motif, or GST-osteopontin. Treatment with GST-osteopontinsignificantly increased metastasis formation relative to fibrinogen (p<0.05; asterisk) orfibronectin (p<0.05). Average normal lung weight (0.175 g) is indicated by the line. B, In asimilar experiment, mice received C8161 melanoma cells treated with DMEM, 1 mM linearGRGDSP peptide, 1 mM double-cyclic RGD-4C peptide, or GST-osteopontin. The cohortreceiving cells treated with GST-osteopontin exhibited a significant increase in lungmetastasis relative to the other groups (p<0.001). Bars indicate mean ± SD.
Mandelin et al. Page 14
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIGURE 4.Osteopontin reduces actin fibers in adherent tumor cells. Adherent KRIB osteosarcoma cellswere treated with GST-osteopontin or GST-osteopontin-RGE for 10 min, after which thecells were fixed and stained for cytoskeletal actin. Tumor cells treated with GST-osteopontindisplayed a reduced actin network, with a decrease in transcytoplasmic actin cables, incomparison with cells treated with GST-osteopontin-RGE (arrows). Scale bar, 20 μm.
Mandelin et al. Page 15
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIGURE 5.Osteopontin treatment is associated with Src dephosphorylation of detached tumor cells. A,KRIB osteosarcoma cells were detached from tissue culture substrates. Some of the cellswere allowed to adhere to the culture dish, while others were treated for 1 min with GST-osteopontin, GST-osteopontin-RGE, GST, RGD-4C peptide, or DMEM only. The cells weresubsequently lysed, and 30 μg of the lysate was analyzed for the phosphorylation of Tyr-418c-Src. Adhesion of the cells to plastic induced transient activation of c-Src. Treatment of thecells with GST-osteopontin induced dephosphorylation of c-Src. Analysis of total Srcconfirmed equal loading of the proteins. B, Analysis of the tyrosine phosphorylation of thesame lysates (30 μg) revealed general dephosphorylation of tyrosine in proteins (note range50–98 kDa proteins). Arrow indicates c-Src; cellular actin (40kDa) was detected at the sametime to ensure equal loading of the proteins.
Mandelin et al. Page 16
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIGURE 6.Promotion of tumor cell binding to vascular endothelium by osteopontin. A and B, KRIBosteosarcoma cells were treated with DMEM, GST-osteopontin, GST-osteopontin-RGE, orphosphatase inhibitors prior to GST-osteopontin and were subsequently incubated with theendothelium of the dorsal aorta. Relative to the GST-osteopontin-RGE and phosphataseinhibition groups, GST-osteopontin significantly increased cell binding (arrows) to theaortas (p<0.05; asterisk). Bars indicate mean ± SD. Scale bar, 20 μm. C, The number oftumor foci was increased in nude mice receiving human sarcoma cells previously incubatedwith GST-osteopontin relative to DMEM-treated cells.
Mandelin et al. Page 17
Cancer. Author manuscript; available in PMC 2010 April 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript