expression of nuclear hormone receptors in the rat supraoptic nucleus
TRANSCRIPT

0013.7227/95/$03.00/O Endocrinology Copyright 0 1995 by The Endocrine Society
Vol. 136, No. 5 Prmted III U.S.A.
Expression of Nuclear Hormone Receptors in the Rat Supraoptic Nucleus
SOFIA LOPES DA SILVA, A. MARTIN VAN HORSSEN, CHAWNSHANG WANG, AND
J. PETER H. BURBACH
Rudolf Magnus Institute for Neurosciences (S.L.d.S., M.v.H., J.P.H.B.), Department of Medical Pharmacology, Utrecht University, 3584 CG Utrecht, The Netherlands; and Department of Human Oncology and Program in Endocrinology and Reproductive Physiology (C.C.), University of Wisconsin, Madison, Wisconsin 53792
ABSTRACT To investigate the role of nuclear hormone receptors on neuropep-
tide gene expression in the hypothalamo-neurohypophyseal system (HNS) of the rat, a survey was made of members of the nuclear hormone receptor superfamily that are expressed in the supraoptic nucleus (SON). A polymerase chain reaction cloning strategy based on homologies in the DNA-binding domain of AGGTCA-binding factors was devised for the identification of receptors in microdissected SON tissue. Cloning of the amplified products led to the identification of five true receptors, thyroid hormone receptor-a (THRo), retinoic acid receptor-o, retinoic acid receptor-y, retinoidxreceptor-cu, and retinoid X receptor-y, as well as four orphan receptors, apolipoprotein AI regulatory protein (ARP-11, chicken ovalbumin upstream promoter transcription factor I (COUP-TF I), estrogen-related receptor 2, and testis receptor 4 (TR4). Dot-blot screening of amplified gene fragment
analysis showed that THRa, ARP-1, TR4, and COUP-TF I were the most abundant factors expressed in the SON region, in the order THRa > ARP-1 > TR4 = COUP-TF I. THRa has previously been localized to HNS neurons. In situ hybridization analysis showed that ARP-1, COUP-TF I, and TR4 were not expressed in magnocellular neurons at appreciable levels, but rather in surrounding structures. Furthermore, in lactating female rats there were no significant dif- ferences in the composition of the nine identified nuclear hormone receptors in the SON region compared with control animals. From these experiments, it is concluded that there is a multitude of hypo- thalamically expressed nuclear hormone receptors, but that only THRa is expressed at relatively high abundance in HNS neurons. This indicates that the peptide-producing magnocellular neurons of the SON express a specific set of transcription factors of the nuclear hormone receptor family. (Endocrinology 136: 2276-2283, 1995)
T HE hypothalamo-neurohypophyseal system (HNS) pro- vides a conduit for influences of the nervous system on
the endocrine system and as such plays a critical homeostatic role. The HNS consists of magnocellular neurons in the hy- pothalamus that send their axons to the posterior lobe of the pituitary gland, where they release vasopressin (VP) and oxytocin (OT) as major products into the general circulation (1). Adequate functioning of the HNS requires critical reg- ulation of the synthesis and release of its peptide products. Recent research aims to understand the transcriptional events that control the synthesis of neuropeptides. One ap- proach is to identify transcription factors in the HNS. At present, a number of transcription factors belonging to dis- tinct families have been identified in this system (2). Tran- scription factors of the basic leucine zipper proteins family (3-5), such as c-fos and isoforms of the CAMP response ele- ment-binding protein modulators (6), have been localized in the supraoptic nucleus (SON), as well as transcription factors belonging to class III of the family of POU domain proteins (7). Their functions are unknown.
From the viewpoint of endocrine regulation of the VP and OT genes in the HNS, particular attention has been paid to steroid hormone receptors. OT and VP gene expression are stimulated during pregnancy (B), suggesting a regulatory
Received July 12, 1994. Address all correspondence and requests for reprints to: Dr. J. Peter
H. Burbach, Rudolf Magnus Institute for Neurosciences, Department of Medical Pharmacology, Utrecht University, Universiteitsweg 100, 3584 CG Utrecht, The Netherlands.
role of steroid hormones. A composite hormone response element was identified in the rat OT (rOT) gene that inte- grates responses to estrogen, thyroid hormone, and retinoic acid (RA) (9, 10). The rVP gene also contains a potential hormone response element, although actions of nuclear hor- mone receptors on this element have not been found as yet (2, 11). The nuclear hormone receptor family includes not only true receptors for ligands, such as steroid and thyroid hormones, but also transcription factors for which no ligands are known (12). A large amount of these so-called orphan receptors have been cloned recently (13), and their role in neuropeptide gene regulation remains to be elucidated.
The focus of this study was on the presence of a category of nuclear hormone receptors that bind &-elements com- posed of AGGTCA motifs. These motifs are present in the composite hormone response element of the OT gene (9,10, 14) as well as in the VP gene (2,ll). Most receptors, including all orphans, bind to &-elements composed of repeats of AGGTCA motifs (15). They share common amino acid res- idues in the DNA-binding domain (DBD). Only a limited subfamily that includes the progesterone receptor, the glu- cocorticoid receptor, the mineralocorticoid receptor, and the androgen receptor recognizes a different &element, com- posed of a palindrome of the TGTTCT motif. To identify hypothalamic nuclear hormone receptors, a polymerase chain reaction (PCR) cloning strategy based on homologies in the DBD of AGGTCA-binding factors was devised. The most abundant clones were analyzed by means of in situ hybridization for their presence in magnocellular neurons.
2276

EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON 2277
Furthermore, it was investigated whether the composition of nuclear hormone receptors differed in lactating rats com- pared with controls.
Materials and Methods
Animals
Wistar rats (TN0 Zeist, The Netherlands) from our own breeding stock (250-300 g) were decapitated, and their brains were rapidly dis- sected. The SONS were microdissected from 2-mm-thick fresh tissue slices using needles with a diameter of 1 mm, according to Palkovits (16). In several experiments, female rats lactating litters for 4-6 days were used.
RNA isolation and first-strand complementary DNA (cDNA) synthesis
Total RNA was isolated from SON tissue by means of extraction with RNAzol according to the manufacturer’s intructions (Cinna/Biotecx, Friendswood, TX). First-strand cDNA was synthesized using Super- Script, a Moloney murine leukemia virus reverse transcriptase (BRL, Gaithersburg, MD). Briefly, 2.5 kg total RNA were annealed to 1 pg random hexanucleotide primers and reverse transcribed in 20 ~1 first- strand buffer containing 50 rnM Tris-HCl (pH 8.31, 75 rnM KCl, 3 mM MgCl,, 10 mM dithiothreitol (DTT), and 1.25 mM deoxynucleoside triphosphate each at 37 C for 1.5 h using 400 U Superscript reverse transcriptase. Subsequently, Superscript was heat inactivated at 90 C for 5 min. Finally, 30 U RNaseH (Amersham, Little Chalfont, UK) were added, and the mixture was incubated at 37 C for 20 min.
PCR amplification
One tenth of the cDNA was supplemented with 50 pmol of the appropriate 5’.primer and 3’-primer and amplified in a 50-~1 reaction mixture containing 1.0 U Replitherm polymerase (Biozym, Madison, WI), 50 mM KCI, 10 rnM Tris-HCl (pH 8.3), 1.5 rnM MgCl,, 0.1% (wt/vol) gelatin, and 200 PM deoxynucleoside triphosphate each. The degenerate primers were directed to the DBDs of all nuclear hormone receptors except those that contain a phosphorylation box with the amino acid sequence GSCKV (glucocorticoid, mineralocorticoid, progesterone, and androgen receptors). All other receptors, including the recently identi- fied members, were included in this study. Nucleotide sequences of the highly conserved DBDs of these nuclear hormone receptors were ob- tained from the GenBank/EMBL database, release 69/28. Alignments were performed with the Genetics Computer Group Sequence Analysis Software Package (Genetics Computer Group, Madison, WI) (17). The nuclear hormone receptors used in the alignment are (the GenBank/ EMBL accession numbers are given in parentheses): human apolipopro- tein AI regulatory protein (hARP-1; M64497) (18), human chicken ovalbumin upstream promoter transcription factor I (COUP-TF I; X16155) (19), human ear2 (X127994) (20), mouse embryonal long termi- nal repeat-binding protein (D90530) (21), Mouse estrogen receptor (mER; M38651) (22), rER (YOO102) (23), hER (X03635) (24), human es- trogen-related receptor 1 (hERR-1; X51416) (25), hERR-2 (X51417) (25), rat hepatic nuclear factor 4 (X57133) (26), rat nerve growth factor 1B (JO41131 (271, human RA receptor-a (hRARol; X06538) (28), mRARa (M60909) (29), hRARy (M38258) (30), mRARy (X15848) (31), human retinoid X receptor-a (hRXRa; X52773) (32), mRXRlr (M84817) (33), mouse H-2 region II binding protein (M26804) (34), mRXRy (M84819) (331, Rat thyroid hormone receptor-a (rTHRu; M18028) (351, human c-e&A (X55005) (36), human testis receptor 2 (hTR-2; M21985) (37), human vitamin D receptor (JO3258) (38), and rat vitamin D receptor (JO4147) (39).
Based on this alignment, the following 5’-primer and 3’-primer were designed: (DBD66-100, 5’.primer, 5’.GGA GTC GGT ACC TG[T/CJ GA[G/A] GGC TGC AAG GG[T/C] TTC TT-3’; and DBD210-238,3’- primer, 5’-TC CTT [G/ClNG CAT GCC CAC TTC GA[G/A/TJ GCA CTT3’). To facilitate cloning, the 5’.primer contained an internal K,nnI restriction site, and the 3’-primer contained a SphI site (underlined). PCR amplifications were carried out by 30 cycles under the following con-
ditions: 1 min of denaturation at 94 C, 1 min of primer annealing at 37 C during the first 15 cycles and 45 C during the following cycles, and 2 min of extension at 75 C. During the final cycle, PCR mixtures were incubated at 75 C for 10 min to obtain full extension of all PCR products.
Cloning and PCR library screening
PCR-amplified material was digested with K@ and SyhI restriction enzymes, purified on a 1% low melting point (LMP) agarose gel (Sigma, St. Louis, MO), and cloned into the pGEM7 Zf(+) vector (Promega, Madison, WI) using standard procedures (40). After transformation of competent DH5cu cells, 300 independent colonies were plated and screened. Screening was performed by subsequent steps of sequencing and screening, using the sequenced PCR products as probe. DNA se- quence analysis was performed on double-stranded DNA templates using T7 DNA polymerase and T7 and SP6 sequencing primers accord- ing to the manufacturer’s instructions (Pharmacia, Uppsala, Sweden; T7 Sequencing Kit). Sequenced clones were used as a probe to screen the PCR library that was transferred to Colony/Plaque Screen membranes (DuPont New England Nuclear, Boston, MA). To avoid cross-hybrid- ization, the 5’.DBD66-100 primer of sequenced PCR products was re- moved with an appropriate restriction enzyme. Single-stranded fss) probes were subsequently made by means of linear PCR with an internal 3’-primer in the presence of [ol-32P]deoxycytidine triphosphate (dCTP1. Linear PCR was performed on an LMP agarose gel-purified template DNA (5 ng) supplemented with 50 pmol of the appropriate 3’.primer in a 50-~1 reaction mixture containing 50 mM KCI, 10 mM Tris-HCI (pH 8.3), 1.5 mM MgCl, 0.1% (wt/vol) gelatin, 200 mM d[G/A/TJtriphosphate each, 50 &i [cu-“P]dCTP (Amersham), and 1.0 U Replitherm poly- merase (Biozym). Linear PCR consisted of 50 cycles, which were carried out under the following conditions: 1 min denaturation at 94 C, 1 min primer annealing at 50 C, and 2 min extension at 75 C. The “I’ ssDNA probe was precipitated with 0.3 M sodium acetate and ethanol and 10 pg carrier DNA and finally dissolved in water. Filters were hybridized in 3X SSC (1 X SSC = 0.15 M NaCl, 0.015 M Na citrate, pH 7.0),10% (wt/vol) dextran sulfate, 0.5% sodium dodecyl sulfate, 5X Denhardt’s solution, 50 Kg/ml denatured salmon sperm DNA, and 2 X 10’ cpm/ml [“ZP]ssDNA probe at 70 C overnight. After hybridization, the filters were washed three times in 0.1 X SSC, 0.1% sodium dodecyl sulfate at 70 C for 20 min and exposed to autoradiographic film (X-OMAT AR, Kodak, Rochester, NY) at -80 C for 2 h using an intensifying screen. Negative clones were sequenced and screened in consecutive rounds.
In situ hybridization
Mouse and rat brains were quickly dissected and frozen in isopen- tane, which was cooled to approximately ~25 C in an ethanol-dry ice mixture. Cryostate sections of 16 km were mounted on poly-r-lysine- coated slides and stored at -80 C. Slides were placed at room temper- ature for 20 min and fixed in freshly prepared 4% paraformaldehyde in PBS for 5 min. The slides were washed twice with PBS for 5 min and transferred to 0.25% acetic anhydride in 0.1 M triethanolamine fpH 8.0) for 10 min. This was followed by washing in PBS for 5 min and in 0.83% NaCl for 5 min. The slides were dehydrated slowly by immersing them in solutions with increasing concentrations of ethanol (30%, 50%, 7OYc, 85%, 96%, and 100%). The dehydrated slides were delipidated in chlo- roform for 1 min and washed in 100% ethanol again. The slides were air dried for 30 min and used for hybridization on the same day.
Mouse ARP-1 and COUP-TF I cRNA probes were derived from 3’-noncoding cDNA fragments, which were cloned in pBluescript SK-II (SmaI; Stratagene, San Diego, CA). The mouse ARP-1 fragment is a 322-base pair (bp) SspI-MscI fragment (1588-1910). The mouse COUP-TF I fragment is a 146.bp RsnI-RsaI fragment (1797-1943) (41). [n-“Sluridine triphosphate-labeled antisense transcripts were generated on linearized plasmids using T3 RNA polymerase (Promega). As a control for non- specific binding, sense RNA probes were made using T7 RNA poly- merase (Promega). An rTR4 cRNA probe was made using nucleotides 66-693, which were cloned in pT7-blue [a-‘“Sluridine triphosphate- labeled antisense transcripts made on linearized plasmid with T7 RNA polymerase. As a control for nonspecific binding, a 50.fold excess of unlabeled transcript was used.
Hybridization was performed in 50% formamide, 2X SSC, 10% dex-

2278 EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON Endo. 1995 Vol 136. No 5
tran sulfate, 1X Denhardt’s solution, 5 mM EDTA, 10 mM phosphate buffer (DH = 8.0). 100 rnM DTT, and 0.5 mcr/ml veast transfer RNA containing 10h cpm ‘“S-labeled cRNA probe. T?,e solution was heated at 80 C for 2 min and cooled on ice before being applied on slides (75 PI/slide). The slides were covered with coverslips, placed in a plastic slide box, and incubated overnight at 50 C. Slides were washed in 200 ml 5~ SSC, 10 rnM DTT (fresh) at 65 C for 30 min for coverslips to fall off. High stringency washing was performed in 50% formamide, 2X SSC, and 10 mM DTT (fresh) at 65 C for 30 min. This was followed by washing three times for 10 min each in NTE buffer (0.5 M NaCl, 10 mM Tris-HCl, and 5 mM EDTA, pH = 8.0) at 37 C. The slides were treated with 20 pg/ml RNAse A in NTE buffer at 37 C for 30 min and washed again with NTE buffer at 37 C for 15 min. The high stringency wash in 50% for- mamide, 2X SSC, and 10 mM DTT at 65 C was repeated. The slides were washed at room temperature in 2~ SSC and in 0.1 X SSC for 15 min each and then dehydrated quickly by putting them through 30%, 60%, SO%, and 95% ethanol, containing 0.3 M ammonium acetate, followed by 100% ethanol, twice. Slides were air dried and exposed to Kodak X-OMAT AR autoradiography film for 3 days. For analysis at the microscopic level, the slides were dipped in Kodak NBT2 autoradiography liquid emulsion and exposed for 6 weeks or longer.
Dot-blot screening of amplified gene fragments
Dot-blot screening of amplified gene fragments (DOSAGE) was used to determine the relative amounts of receptors in microdissected SONS and to determine their composition in different brain regions and pe- ripheral organs of the rat. Therefore, PCR reactions were carried out on cDNA derived from different tissues using DBD66-100 and DBD210- 238. The PCR products were separated from the primers on a 1% LMP agarose gel and random-prime labeled using Klenow polymerase (Boehringer, Mannheim, Germany) and 10 FCi [~-32PldCTP. The labeled PCR products were used as probes on dot blots (Hybond nylon mem- branes, Amersham, Den Bosch, The Netherlands) containing the fol- lowing nuclear hormone receptor cDNAs: rARP-1, rCOIJP-TF I, hER, rERR2, rRARa, rRARy, rRXRol, rRXRy, rTHRcr, and rTR4. In addition, the DOSAGE technique was used to survey for differences in amounts of nuclear hormone receptors between SONS of lactating female rats and those of adult male rats. Therefore, SON tissue of three lactating or adult male rats was pooled and used for the synthesis of cDNA. Nine inde- pendent cDNA samples derived from lactating rats were compared to nine independent samples from adult male rats. The intensities of the various hybridization signals were quantitated with a phosphoimage analyzer (Fujix BAS 1000 Bio-Imaging Analyzer System, Fuji, Strauben- hardt, Germany). The presence of the cloned receptors was also verified in SON tissue derived from neonatal rats.
Results
Cloning and PCR library screening
Reverse transcriptase PCR on RNA of microdissected SON tissue, using degenerated primers directed to the DBD of nuclear hormone receptors (DBD66-100 and DBD210-238), resulted in a single band in the size range of 160-180 bp. This is the size expected for amplification of messenger RNAs (mRNAs) of the nuclear hormone receptor family. Since for most receptors, the zinc fingers Cl and C2 are encoded by separate exons (36,42), contamination of PCR-amplified hy- pothalamic cDNA with genomic DNA is unlikely. PCR-am- plified material was cloned in a plasmid vector, and a library consisting of 300 independent clones was screened by sub- sequent steps of sequencing followed by hybridization using the sequenced PCR products as probes. An individual nu- clear hormone receptor that consisted of more than 1% of the mixture of PCR products was likely (P > 0.95) to be included in this library. Of these clones, 263 contained inserts char-
acteristic for the DBD of the nuclear hormone receptor fam- ily, whereas 37 clones consisted of unrelated sequences.
Nine distinct receptor sequences (clones l-9) were identified. Comparison with DNA and amino acid sequences in the GenBank/EMBL database showed that eight of these receptors were rat homologs of known nuclear hormone receptors. One clone represented a novel nuclear hormone receptor of which the full-length cDNA cloning has been reported recently (43). The nucleotide and amino acid se- quences of the nine clones, aligned to the most similar se- quences in the GenBank/EMBL database, are given in Fig. 1.
The amino acid sequence predicted by the nucleotide se- quence of clone 1 showed 97.1% similarity with hARP-1 (nucleotide homology, 97.1%) (18). Clone 2 was identical to hCOUP-TF I (nucleotide homology, 100%) (19), and clone 3 was identical to hERR2 at the amino acid level (nucleotide homology, 85.3%) (25). The amino acid sequences of clones 4 and 5 were identical to mRARa (nucleotide homology, 97.1%) (29) and hRARy (nucleotide homology, 91.2%) (30), respectively. Clone 6 was 97.1% similar both at the nucleotide and at the amino acid level to mRXRa (33). Clone 7 was identical to mRXRy (nucleotide homology, 91.2%) (33), and clone 8 was identical to rTHRa (nucleotide homology, 100%) (35). Comparison of nucleotide sequences of clone 9 (estab- lished by sequence analysis of six independent clones) to known nucleotide sequences revealed a similarity to chicken RXRa (78.2%) (44) and mRXRa (70.5%) (331, the rTR2 (76.5%) (37) and hARP-1 (70.3%) (18). However, comparison of the predicted amino acid sequence of clone 9 with protein se- quences in the GenBank/EMBL database revealed the high- est similarity to both hTR2 and rTR2 (82.4%). These data showed that clone 9 represented a novel member of the nuclear hormone receptor superfamily. Clone 9 has been used as a probe to isolate and characterize the full-length cDNA of a novel orphan receptor, termed TR4 (43).
In situ hybridization
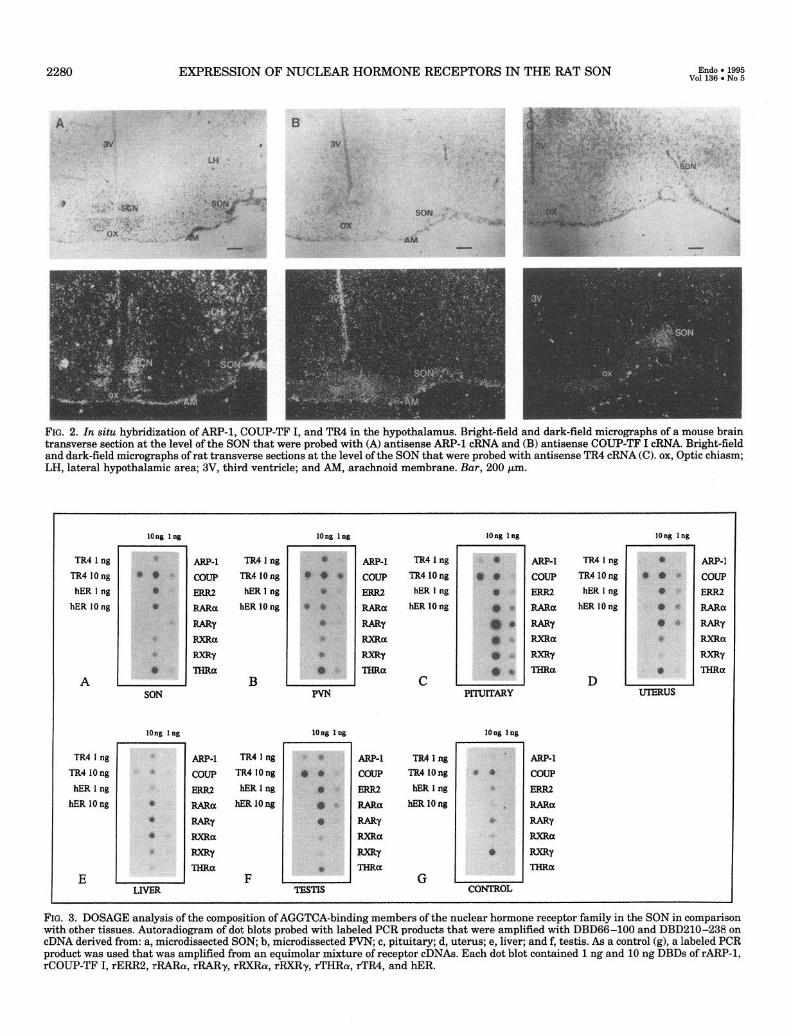
From the cloning experiment and DOSAGE studies (see below) it seemed that THRcw, ARP-1, COUP-TF I, and TR4 were the most abundant factors expressed in microdissected SON tissue. Previous studies had already demonstrated THRa expression in the SON, in both magnocellular and parvocellular cells of the paraventricular nucleus (PVN), in the preoptic nucleus, and in the suprachiasmatic nucleus (SCN) in the rat hypothalamus (45,46). In situ hybridization for ARP-1, COUP-TF I, and TR4 was performed to determine their sites of hypothalamic expression. For ARP-1 and COUP-TF I, cRNA probes were available that were derived from the 3’-untranslated region of the mouse mRNA (41). These probes were therefore used on mouse hypothalamus sections at the level of the SON (Fig. 2, A and B). ARP-1 mRNA was detected in the lateral hypothalamic area, in the SCN, in the wall of the third ventricle, and in the arachnoid membrane (Fig. 2A). COUP-TF I expression was not ob- served in hypothalamic nuclei but was only detected in the wall of the third ventricle and in the arachnoid membrane (Fig. 2B). Sense cRNA probes did not give a hybridization signal. In situ hybridization of the orphan receptor TR4 was performed on hypothalamic sections of the rat brain using a

EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON 2279
FIG. 1. Comparison of the nucleotide and deduced amino acid se- quences of the PCR-amplified cDNAs cloned from the microdissected SON of the rat (clones l-9) to the most similar nucleotide sequences
rat-specific cDNA probe (43). A low level of specific labeling was observed throughout the hypothalamus (Fig. 2C). This hybridization signal was not found when a 50-fold molar excess of unlabeled transcript was added to the TR4 probe. Notably, none of these factors seemed to be expressed in magnocellular neurons of the HNS at appreciable levels.
DOSAGE
DOSAGE (47) was used to determine the relative amounts of receptors in microdissected SON and to compare this composition with that in the microdissected PVN and with that in other tissues. Furthermore, the DOSAGE technique was used to screen for possible pronounced changes in the expression of the identified nuclear hormone receptors in lactating and neonatal rats.
Figure 3 shows autoradiograms of dot blots probed with labeled PCR products, which were amplified with DBD66- 100 and DBD210-238 from cDNA derived from microdis- sected SON, PVN, rat pituitary gland, uterus, liver, and testis. As a comparison, a labeled PCR product was used that was amplified from an equimolar mixture of receptor cDNAs. Each dot blot contained fixed amounts of the DBDs of rARP-1, rCOUP-TF I, rERR2, rRARLu, rRARy, rRXRa, rRXRy, rTHRa, rTR4, and hER. Hybridization with labeled PCR product amplified from the equimolar mixture of all cloned cDNAs showed a preferential amplification of COUP-TF I, less abundant amplification of ARP-1, ERR2, RARy, and THRa, and almost no amplification of ER, indicating a large difference in amplification efficiency among receptors. The DOSAGE technique is therefore not suitable for the deter- mination of the precise amount of receptors that are present in a certain tissue. However, the technique provides an ad- equate tool to compare the composition of receptors in a certain tissue under different conditions or with another tissue. DOSAGE of independent SON samples (n = 24) showed that the composition of receptors was very repro- ducible. When corrected for the bias in amplification effi- ciency, the following order of abundancy is obtained for the identified factors in microdissected SON: THRo > ARP-1 > TR4 = COUP-TF I > other factors (Fig. 3).
THRo was also the most abundant factor in the microdis- sected PVN. Low proportions of THRa in the total receptor population in liver and testis were found in comparison to the amounts of RARs in these tissues. In the microdissected SON and PVN, the RARs were not predominant, in contrast to the liver, in which a large proportion of the receptor population consisted of RXRs. In the receptor population of the SON region, RARa and ERR2 were represented more than in the PVN population. The proportion of COUP-TF I was higher in the PVN region than in the SON region. A notable difference was the detection of ER in PVN samples, but not in SON samples. The ER was amplified with low
in the GenBanWEMBL database and to the predicted amino acid sequences. A, Clone 1 compared with hARP-1; B, clone 2 compared with hCOUP-TF I; C, clone 3 compared with hERR2; D, clone 4 compared with mRARu; E, clone 5 compared with hRARy; F, clone 6 compared with mRXRa; G, clone 7 compared with mRXR-y; H, se- quence of clone 8, which is identical to rTHRa; and I, sequence ofclone 9, which has been termed rTR4 (43).
2280 EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON Endo . 1995 Voll36. No 5
FIG. 2. In situ hybridization of ARF’-1, COUP-TF I, and TR4 in the hypothalamus. Bright-field and dark-field micrographs of a mouse brain transverse section at the level of the SON that were probed with (A) antisense ARP-1 cRNA and (B) antisense COUP-TF I cRNA. Bright-field and dark-field micromanhs of rat transverse sections at the level of the SON that were probed with antisense TR4 cRNA (C). ox, Optic chiasm; LH, lateral hypothalamic area; 3V, third ventricle; and AM, arachnoid membrane. Bar, 200 pm.
TR41ng
TR4 10 ng hER I ng
hER 10 ng
A
long lng long lag
long lng
LIVER
TR41ng
TR41ong
hER 1 ng
hER 10 ng
B
m-1 TR4 I ng
COUP TR4 10 ng
ERR2 hERing
RARa hERlOng
RARY
FtXRa
mr
THRa F
long log
\ ”
. \ \ _ ..:< : ;:~.: . . \
. ; 1,; ‘,: 1.. _, *‘..’ \
TESTIS
Am-1 TR4lng
COUP TR4 10 ng
ERR2 hER 1 ng
RARa hER 10 ng
MY
RXRa
Rxxy
THRa C
long 1ng long 1ng
\\ PITUITARY
long 1ng
ARP-1
COUP
Em2
RARa
mY
RXRa F-w THRa
TR4lng
TR4 long
hER 1 ng
hER 10 ng
ARP-1 TR4 I ng
COUP TR4 long
ERR2 hER I ng
RARa hER 10 ng
RARY RXRa
=Y TXRa
D
ARP-1
COUP
ERR2
RARa
RARY RXRa
=w
lW.Ra
UmRus
FIG. 3. DOSAGE analysis of the composition of AGGTCA-binding members of the nuclear hormone receptor family in the SON in comparison with other tissues. Autoradiogram of dot blots probed with labeled PCR products that were amplified with DBD66-100 and DBD210-238 on cDNA derived from: a, microdissected SON; b, microdissected PVN; c, pituitary; d, uterus; e, liver; and f, testis. As a control (g), a labeled PCR product was used that was amplified from an equimolar mixture of receptor cDNAs. Each dot blot contained 1 ng and 10 ng DBDs of rARP-1, rCOUP-TF I, rERR2, rRARcy, rR.ARy, rRXRo, rI?XRy, rTHRa, rTR4, and hER.
EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON 2281
5
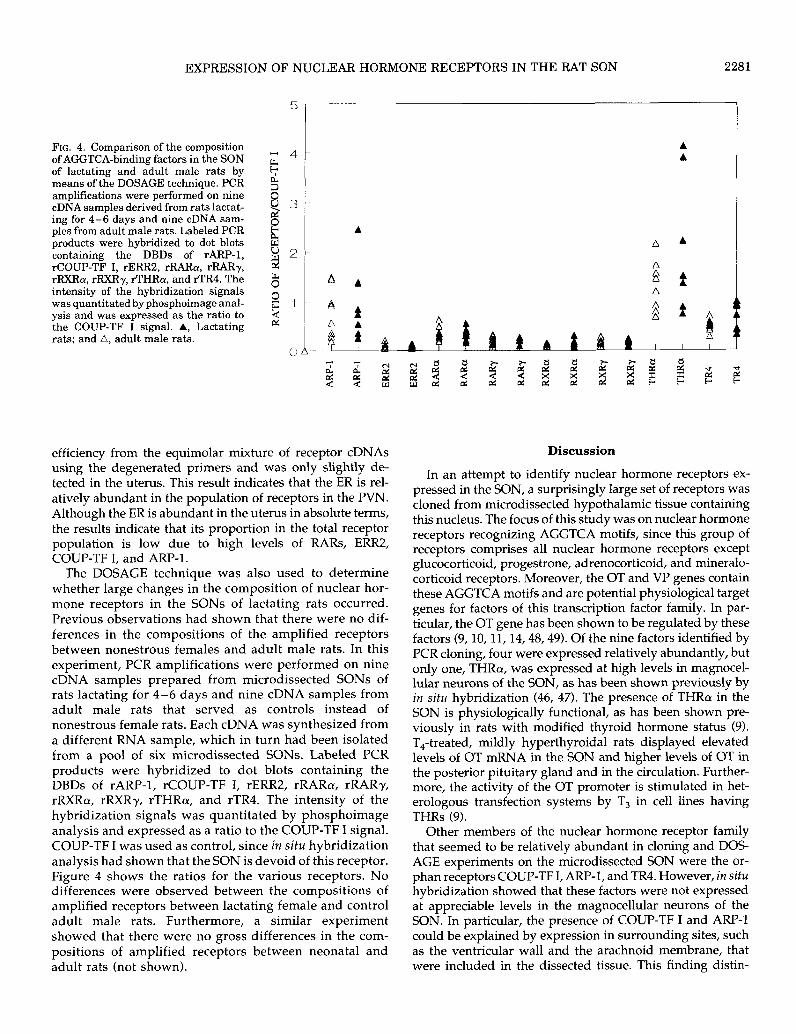
FIG. 4. Comparison of the composition of AGGTCA-binding factors in the SON M 4 of lactating and adult male rats by 6 means of the DOSAGE technique. PCR amplifications were performed on nine
3
cDNA samples derived from rats lactat- 83 ing for 4-6 days and nine cDNA sam- ples from adult male rats. Labeled PCR
8
products were hybridized to dot blots e
containing the DBDs of rARP-1, 82 rCOUP-TF I, rERR2, rRARe, rRARy, 2 rRXRcr, rRXRy, rTHRe, and rTR4. The intensity of the hybridization signals
8
was quantitated byphosphoimage anal- ysis and was expressed as the ratio to
$1
the COUP-TF I signal. A, Lactating !s
rats; and A, adult male rats. 0 h
A A
A A A
g A
efficiency from the equimolar mixture of receptor cDNAs using the degenerated primers and was only slightly de- tected in the uterus. This result indicates that the ER is rel- atively abundant in the population of receptors in the PVN. Although the ER is abundant in the uterus in absolute terms, the results indicate that its proportion in the total receptor population is low due to high levels of RARs, ERR& COUP-TF I, and ARP-1.
The DOSAGE technique was also used to determine whether large changes in the composition of nuclear hor- mone receptors in the SONS of lactating rats occurred. Previous observations had shown that there were no dif- ferences in the compositions of the amplified receptors between nonestrous females and adult male rats. In this experiment, PCR amplifications were performed on nine cDNA samples prepared from microdissected SONS of rats lactating for 4-6 days and nine cDNA samples from adult male rats that served as controls instead of nonestrous female rats. Each cDNA was synthesized from a different RNA sample, which in turn had been isolated from a pool of six microdissected SONS. Labeled PCR products were hybridized to dot blots containing the DBDs of rARP-1, rCOUP-TF I, rERR2, rRARa, rRARy, rRXRa, rRXRy, rTHRa, and rTR4. The intensity of the hybridization signals was quantitated by phosphoimage analysis and expressed as a ratio to the COUP-TF I signal. COUP-TF I was used as control, since in situ hybridization analysis had shown that the SON is devoid of this receptor. Figure 4 shows the ratios for the various receptors. No differences were observed between the compositions of amplified receptors between lactating female and control adult male rats. Furthermore, a similar experiment showed that there were no gross differences in the com- positions of amplified receptors between neonatal and adult rats (not shown).
Discussion
In an attempt to identify nuclear hormone receptors ex- pressed in the SON, a surprisingly large set of receptors was cloned from microdissected hypothalamic tissue containing this nucleus. The focus of this study was on nuclear hormone receptors recognizing AGGTCA motifs, since this group of receptors comprises all nuclear hormone receptors except glucocorticoid, progestrone, adrenocorticoid, and mineralo- corticoid receptors. Moreover, the OT and VP genes contain these AGGTCA motifs and are potential physiological target genes for factors of this transcription factor family. In par- ticular, the OT gene has been shown to be regulated by these factors (9,10,11,14,48,49). Of the nine factors identified by PCR cloning, four were expressed relatively abundantly, but only one, THRa, was expressed at high levels in magnocel- lular neurons of the SON, as has been shown previously by in situ hybridization (46, 47). The presence of THRa in the SON is physiologically functional, as has been shown pre- viously in rats with modified thyroid hormone status (9). T,-treated, mildly hyperthyroidal rats displayed elevated levels of OT mRNA in the SON and higher levels of OT in the posterior pituitary gland and in the circulation. Further- more, the activity of the OT promoter is stimulated in het- erologous transfection systems by T, in cell lines having THRs (9).
Other members of the nuclear hormone receptor family that seemed to be relatively abundant in cloning and DOS- AGE experiments on the microdissected SON were the or- phan receptors COUP-TF I, ARP-1, and TR4. However, in situ hybridization showed that these factors were not expressed at appreciable levels in the magnocellular neurons of the SON. In particular, the presence of COUP-TF I and ARE-1 could be explained by expression in surrounding sites, such as the ventricular wall and the arachnoid membrane, that were included in the dissected tissue. This finding distin-

2282 EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON Endo. 1995 Vol 136. No 5
guishes magnocellular OT neurons from OT-expressing granulosa cells of the bovine corpus luteum, which express COUP-TF I and ARP-1 (50). In these cells, the OT gene and the ARP-1 and COUP-TF I genes are expressed in an opposite fashion (50). This expression pattern has been interpreted as a functional repressive activity of COUP-TF I and ARP-1 on the OT gene (50). Indeed, these factors can strongly repress the positive action of the hormone-activated THR, ER, and RAR in transfected cell lines through competition for the composite hormone response element in the 5’-flanking re- gion of the OT gene (49). It is possible that the composite hormone response element in the rOT gene is only used for the regulation by thyroid hormone in the HNS but is a more complex site of regulation in other OT-expressing tissues. On the other hand, still other factors may exist in the HNS that use this element.
Comparison of the composition of nuclear hormone re- ceptors in the SON region of lactating rats and control rats did not reveal a significant difference in the receptor popu- lation, taking as internal standard the non-SON-expressed factor COUP-TF I. These data suggest that no gross changes in the composition of these nuclear hormone receptors occur during lactation, a condition that results in two to four times higher OT mRNA levels (8). However, the semiquantitative nature of the DOSAGE technique does not rule out that more subtle changes occur. These may be determined for each factor separately by in situ hybridization, which was beyond the scope of this study.
The receptors identified at relatively low abundance were RAR~Y, RARy, RXR~K, RXRy, and the orphan receptor ERR2 In view of their low representation in the microdissected SON and the lack of regulation in lactating animals, no at- tempts were made to localize sites of expression. RARs and RXRs are expressed in many tissues. With regard to the RARs, RARP was found to be the most abundantly expressed RAR in the mouse adult brain (31). Our data show that in the SON area, both RARa and RARy are expressed without evidence for the expression of RARP. RARcY~ and RARa2 are present in the mouse brain (511, but our data do not distin- guish between the RARa isoforms. In addition to the RARs, two RA-related receptors were identified, RXRa and RXRy. RXRa! was shown by Northern analysis of poly(A)+ RNA to be present in low amounts in the adult rat brain (33). RXRp, although ubiquitous and abundant in most tissues, was not encountered in our study. On the other hand, we found RXRy, which is known to be less abundant and more re- stricted in its distribution. Our findings indicate that the composition of RAR and RXR members in the microdissected SON differs from that seen overall in the brain.
Little is known about the orphan receptor ERR2 In con- trast to ERRl, a closely related factor, it has not been found to occur in the brain, and functions of it are unknown. ERR1 and ERR2 bear a distant structural relationship to ER but do not bind estrogens. We did not find the rER in the PCR library nor in DOSAGE analysis of the SON. This is in agreement with previous immunohistochemical studies that failed to detect the ER in the SON (52). Axelson and Van Leeuwen (53) had shown previously that the PVN displayed ER immu- noreactivity in a restricted subset of neurons (53).
The findings in this study indicate that the magnocellular
neurons of the SON are restricted in the expression of nuclear hormone receptors and differ from peripheral OT-producing cells. This suggests that specific regulatory mechanisms exist in the HNS.
Acknowledgments
We thank Dr. W. Hendriks for his protocols of the DOSAGE tech- nique. Drs. G. Jonk and W. Kruijer provided the probes for mCOUP-TF I and mARP-1, respectively.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
References
Swanson LW 1993 Patterns of transcriptional regulation in the neu- roendocrine system. In: Burbach JPH, de Wied D feds) Brain Func- tions of Neuropeptides. Parthenon Publishing Group, New York, pp 41-64 Burbach JPH, Lopes da Silva S, Van Schaick HSA, Cox JJ, Adan RAH 1993 Transcription factors of the supraoptic nucleus: activation and repression of the oxytocin gene. In: Gross P, Richter D, Robertson GL feds) Vasopressin. John Libbey Eurotext, pp 95-109 Habener JF 1990 Cyclic AMP response element binding proteins: a cornucopia of transcription factors. Mol Endocrinol 4:1087-1094 Sharp FR, Sagar SM, Hicks K, Lowenstein D, Hisanaga K 1991 c&s mRNA, fos and @-related antigen induction by hypertonic saline and stress. J Neurosci 11:2321-2331 Carter DA, Murphy D 1990 Regulation of c-fos and c-jun expression in the rat supraoptic nucleus. Mol Cell Neurobiol 10:435-446 MellstrSm B, Naranjo JR, Foulkes NS, Lafarga M, Sassone-Corsi P 1993 Transcriptional response to CAMP in brain: specific distri- bution and induction of CREM antagonists. Neuron 10:655-665 Ruvkun G, Finney M 1991 Regulation of transcription and cell identity by POU domain proteins. Cell 64475-478 Van To1 HHM, Bolwerk ELM, Liu B, Burbach JPH 1988 Oxytocin and vasopressin gene expression in the hypothalamo-neurohypo- physeal system of the rat during the estrous cycle, pregnancy, and lactation. Endocrinology 122:945-951 Adan RAH, Cox JJ, Van Kats JP, Burbach JPH 1992 Thyroid hor- mone regulates the oxytocin gene. J Biol Chem 2673771-3777 Adan RAH, Cox JJ, Beischlag TV, Burbach JPH 1993 A composite hormone response element mediates the truns-activation of the rat oxytocin gene by different classes of nuclear hormone receptors. Mol Endocrinol 7:47-57 Burbach JPH, Adan RAH, Cox JJ, Lopes da Silva S 1993 Transac- tivation of the oxytocin and vasopressin promoters by nuclear hormone receptors. Regul Pept 45:31-35 Laudet V, Hanni C, Co11 J, Catzeflis F, Stehelin D 1992 Evolution of the nuclear receptor gene superfamily. EMBO J 11:1003-1013 O’Malley BW, Conneely OM 1992 Orphan receptors: in search of a unifying hypothesis for activation. Mol Endocrinol 6:1359-1361 Richard S, Zingg HH 1990 The human oxytocin gene promoter is regulated by estrogens. J Biol Chem 265:6098-6103 Parker MG 1993 Steroid and related receptors. Curr Op Cell Biol 5:499-504 Palkovits M 1973 Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Res 59:449-450 Devereux J, Haeberli P, Smithies 0 1984 A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res 12: 387-395 Ladias JAA, Karanthanasis SK 1991 Regulation of the apolipopro- tein AI gene by ARP-1, a novel member of the steroid receptor superfamily. Science 251:561-565 Wang LH, Tsai SY, Cook RG, Beattie WG, Tsai M-J, O’Malley BW 1989 COUP transcription factor is a member of the steroid receptor superfamily. Nature 340:163-166 Miyajima N, Kadowaki Y, Fukusjoge SI, Shimizu SI, Semba K, Yamanashi Y, Matsubara KI, Toyoshima K, Yamamoto T 1988 Identification of two novel members of erbA superfamily by mo- lecular cloning. Nucleic Acids Res 16:11057-11066 Tsukiyama T, Ueda H, Hirose S, Niwa 0 1992 Embryonal long

EXPRESSION OF NUCLEAR HORMONE RECEPTORS IN THE RAT SON
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
terminal repeat-binding protein is a murine homolog of FTZ-Fl, a member of the steroid receptor superfamily. Mol Cell Biol 12:1286- 1291 White R, Lees JA, Needham Ham J, Parker M 1987 Structural organization and expression of the mouse estrogen receptor. Mol Endocrinol 1:735 Koike S, Sakai M, Muramatsu M 1987 Molecular cloning and char- acterization of rat estrogen receptor cDNA. Nucleic Acids Res 15: 2459-2513 Green S, Walter P, Kumar V, Krust A, Bomert JM, Argos P, Cham- bon P 1986 Human oestrogen receptor cDNA: sequence expression and homology to v-erbA. Nature 320:134-139 G&u&e V, Yang N, Segui P, Evans R 1988 Identification of a new class of steroid hormone receptors. Nature 331:91-94 Sladek FM, Zhong W, Lai E, Dame11 JE 1990 Liver-enriched tran- scription factor HNF-4 is a novel member of the steroid hormone receptor superfamily. Genes Dev 4:2353-2365 Milbrandt J 1988 Nerve growth factor induces a gene homologous to the glucocorticoid receptor gene. Neuron 1:183-188 Petkovitch M, Brand NJ, Krust A, Chambon P 1987 A human retinoic acid receptor which belongs to the family of nuclear recep- tors. Nature 330&P-450 Pratt MA, Kralova J, McBumey MW 1990 A dominant negative mutation of the cr-retinoic acid receptor gene in a retinoic acid- nonresponsive embryonal carcinoma cell. Mol Cell Biol 10:6445- 6453 Lehmann JM, Hoffmann 8, Pfahl M 1991 Genomic organization of the retinoic acid receptor y gene. Nucleic Acids Res 19:573-578 Zelent A, Krust A, Petkovich M, Kastner P, Chambon P 1989 Cloning of murine (Y and p retinoic acid receptors and a novel receptor y predominantly expressed in skin. Nature 339:714-717 Mangelsdorf DJ, Ong ES, Dyck JA, Evans RM 1990 Nuclear re- ceptor that identifies a novel retinoic acid response pathway. Nature 345:224-229 Mangelsdorf DJ, Borgmeyer U, Heyman RA, Zhou JY, Ong ES, Oro AE, Kakizuka A, Evans R 1992 Characterization of three RXR genes that mediate the action of 9-cis retinoic acid. Genes Dev 6:329-344 Hamada K, Gleason SL, Levi BZ, Hirschfeld S, Appella E, Ozato K 1989 H-2RllBP, a member of the nuclear hormone receptor su- perfamily that binds to both the regulatory element of major his- tocompatibility class 1 genes and the estrogen response element. Proc Nat1 Acad Sci USA 86:8289-8293 Thompson CC, Weinberger C, Lebo R, Evans RM 1987 ldentifi- cation of a novel thyroid hormone receptor expressed in the mam- malian nervous system. Science 237:1610-1614 Laudet V, Btigue A, Henry-Duthoit C, Joubel A, Martin P, StChelin D, Saule S 1991 Genomic organization of the human thyroid hor- mone receptor (Y (c-erbA-1) gene. Nucleic Acids Res 19:1105-l 113 Chang C, Kokontis J 1988 Identification of a new member of the steroid receptor superfamily by cloning and sequence analysis. Bio- them Biophys Res Commun 155:971-977 Baker AR, McDonnell DP, Hughes M, Crisp TM, Mangelsdorf DJ, Haussler MR, Pike JW, Shine J, O’Malley BW 1988 Cloning and
39
40
41
42. 43.
44
45.
46.
47.
48.
49.
50.
51.
52.
53.
expression of full-length cDNA encoding human vitamin D recep- tor. Proc Nat1 Acad Sci USA 85:3294-3298 Burmester JK, Maeda N, DeLuca HF 1988 Isolation and expression of the rat 1,25 dihydroxyvitamin D, receptor cDNA. Proc Nat1 Acad Sci USA 85:9499-9502 Maniatis T, Fritsch EF, Sambrook J 1989 Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY Jonk LJC, De Jonge MEJ, Pals CEGM, Wissink S, Vervaart JMA, Schoorlemmer J, Kruijer W 1993 Cloning and expression during development of three members of the COUP family of nuclear orphan receptors. Mech Dev 47:81-97 Beato M 1989 Gene regulation by steroid hormones. Cell 56:335-344 Chang C, Lopes da Silva S, Ideta R, Lee Y, Yeh S, Burbach JPH 1994 Human and rat TR4 orphan receptors specify a subclass of the steroid receptor superfamily. Proc Nat1 Acad Sci USA 91:6040-6044 Rowe A, Eager NSC, Brickell PM 1991 A member of the RXR nuclear receptor family is expressed in neural-crest-derived cells of the developing chick peripheral nervous system. Development 111: 771-778 Bradley DJ, Young WS, Weinberger C 1989 Differential expression of LY and p thyroid hormone receptor genes in rat brain and pituitary. Proc Nat1 Acad Sci USA 86:7250-7254 Bradley DJ, Towle HC, Young WS 1992 Spatial and temporal ex- pression of (Y- and P-thyroid hormone receptor mRNAs, including the &-subtype, in the developing mammalian nervous system. J Neuroendocrinol 12:2288-2303 Cox JJ, Lopes da Silva S, Hendriks W, Burbach JPH 1994 Regulation of neuropeptide genes: determination of responsiveness to steroids and identification of receptors in brain nuclei. In: De ER Kloet, Sutanto W (eds) Methods in Neurosciences. Vo122. Neurobiology of Steroids. Academic Press, New York, pp 277-295 Burbach JPH, Adan RAH, Van To1 HHM, Verbeeck MAE, Axelson JF, Van Leeuwen FW, Beekman H, ABG 1990 Regulation of the rat bxytocin gene by estradiol. J Neuroendocrinol 2:633-639 Burbach IPH, Loues da Silva S, Cox II, Adan RAH, Coonev Al, Tsai M-J, Tsai S i99dRepression of estrogen-dependent stim;lacion of the oxytocin gene by COUP transcription factor 1. J Biol Chem 269:15046-15053 Wehrenberg U, Ivell R, Walther N 1992 The COUP transcription factor (COUP-TF) is directly involved in the regulation of oxytocin gene expression in luteinizing bovine granulosa cells. Biochem Biophys Res Commun 189:496-503 Leroy P, Krust A, Zelent A, Mendelsohn Gamier J-M, Kastner P, Dierich A, Chambon P 1991 Multiple isoforms of the mouse retinoic acid receptor (Y are generated by alternative splicing and differential induction by retinoic acid. EMBO J 10:59-69 Burbach JPH, Adan RAH, Van To1 HHM, Verbeeck MAE, Axelson JF, Van Leeuwen FW, Beekman H, Ab G 1990 Regulation of the rat oxytocin gene by estradiol. J Neuroendocrinol 2:633-639 Axelson JF, Van Leeuwen FW 1990 Differential localization of es- trogen receptors in various vasopressin synthesizing nuclei of the rat brain. J Neuroendocrinol 2:209-216